 David Sanchis-Pascual1,2*
David Sanchis-Pascual1,2* Rosa Cámara Gómez1,2
Rosa Cámara Gómez1,2 Pilar Morillas-Amat3Darío Lara-Gálvez1,2Víctor Pérez-Cervantes1,2Pilar Masdeu-López-Cerón1,2
Pilar Morillas-Amat3Darío Lara-Gálvez1,2Víctor Pérez-Cervantes1,2Pilar Masdeu-López-Cerón1,2 Juan Francisco Merino-Torres1,2,4
Juan Francisco Merino-Torres1,2,4- 1Endocrinology and Nutrition Department, University and Polytechnic Hospital La Fe, Valencia, Spain
- 2Joint Research Unit on Endocrinology, Nutrition and Clinical Dietetics, Health Research Institute La Fe, Valencia, Spain
- 3Pediatrics Department, Primary Health Care Center of Silla, Silla, Spain
- 4Medicine Department, University of Valencia, Valencia, Spain
The pituitary gland, as a central regulator of endocrine function, may be affected by a wide range of biologically derived harmful substances present in nature. While most available literature focuses on neoplasms, trauma or autoimmune disorders, the potential impact of natural toxins and poisons on pituitary function remains underexplored. This narrative review addresses the effects of acute or chronic exposure to harmful agents originating from the five biological kingdoms—Animalia, Plantae, Fungi, Monera, and Protista—on the hypothalamic–pituitary axis. Drawing on clinical reports, experimental data, and physiological insights, we describe how various biological substances may alter hormonal regulation, leading to temporary or persistent dysfunction. Importantly, this review does not cover direct infectious involvement of the pituitary, such as pituitary abscess, but focuses instead on biologically active compounds produced by living organisms. The review highlights a dispersed body of knowledge with implications not only for endocrinologists and pituitary specialists, but also for clinicians in toxicology, emergency medicine, infectious diseases, and global health. Recognizing the pituitary as a vulnerable target in diverse environmental and ecological contexts may aid in the early diagnosis and management of otherwise unexpected endocrine disorders.
1 Introduction
The pituitary gland is a pivotal endocrine structure situated at the base of the brain, within the sella turcica, and connected to the hypothalamus via the pituitary stalk (1). Despite its small size, it plays a central role in regulating the entire endocrine system through its anterior and posterior lobes. The anterior pituitary (adenohypophysis) secretes trophic hormones and prolactin, that govern target organs such as the adrenal glands, thyroid, gonads, and liver (2). The posterior pituitary (neurohypophysis), derived from neural tissue, releases oxytocin and vasopressin in response to hypothalamic input (3). This bidirectional communication is tightly regulated by negative feedback loops, allowing for dynamic adaptation to internal and external stimuli such as stress, fasting, circadian rhythms, and physiological demands (4, 5). Pituitary integrity is therefore essential to homeostasis, and its disruption may result in substantial clinical consequences, including hormone deficiencies, metabolic dysregulation, and impaired stress responses (6–8).
Although the pituitary operates under robust physiological control, various insults may compromise its function or structure (9, 10). These insults may lead to partial or complete hypopituitarism, hormone hypersecretion, or mass effects due to glandular inflammation, hemorrhage, or expansion. Beyond endocrine dysregulation, hypopituitarism is associated with increased clinical risk. Mortality rates among patients with hypopituitarism are higher than those in the general population, with a meta-analysis reporting a standardized mortality ratio of 1.55 (95% CI 1.14–2.11), even after hormone replacement therapy (11, 12). Furthermore, they also show increased rates of ICU admission and longer hospital stays compared to individuals without pituitary dysfunction (13). These findings underscore the importance of early recognition and prevention of pituitary injury. It is therefore essential to understand how the pituitary gland can be damaged in various pathological contexts, as several causes such as mass effects, radiation, trauma, infections, autoimmune processes, ischemia, and infiltrative diseases have been described (14–20).
While these mechanisms are well established, the potential for naturally occurring biologically derived compounds—such as venoms, plant poisons, fungal toxins, and microbial exotoxins—to impair pituitary function remains underrecognized. This review addresses their impact across the five biological kingdoms, emphasizing an underexplored intersection between natural toxins and endocrine regulation, emphasizing its clinical relevance for recognizing pituitary dysfunction in toxic exposures where early hormonal screening may be crucial for diagnosis and management.
This work was conducted as a narrative review based on an exploratory search of the biomedical literature in PubMed and Google Scholar. Iterative combinations of keywords were used, including “pituitary” together with “toxin,” “venom,” “poison,” “endocrine,” and terms referring to biological sources such as animals, plants, fungi, bacteria, and protists. Additional references were identified from the bibliographies of selected papers. No formal time or language restrictions were applied, and relevant publications from the 1980s to June 2025 were considered.
2 Biological kingdoms and pituitary dysfunction
In this review, we adopt the classical five-kingdom model of biological classification proposed by Whittaker in 1969 as a conceptual framework to explore naturally occurring agents capable of injuring the pituitary gland (Figure 1) (21). Although more recent taxonomic systems such as the proposal by Cavalier-Smith, which incorporates Chromista and other eukaryotic groups, the five-kingdom scheme remains widely used in educational and medical literature and offers a practical structure for grouping biologically derived toxins (22). These kingdoms encompass a broad spectrum of organisms capable of producing venoms, poisons, or other biologically derived compounds that may affect pituitary function, either directly or indirectly.
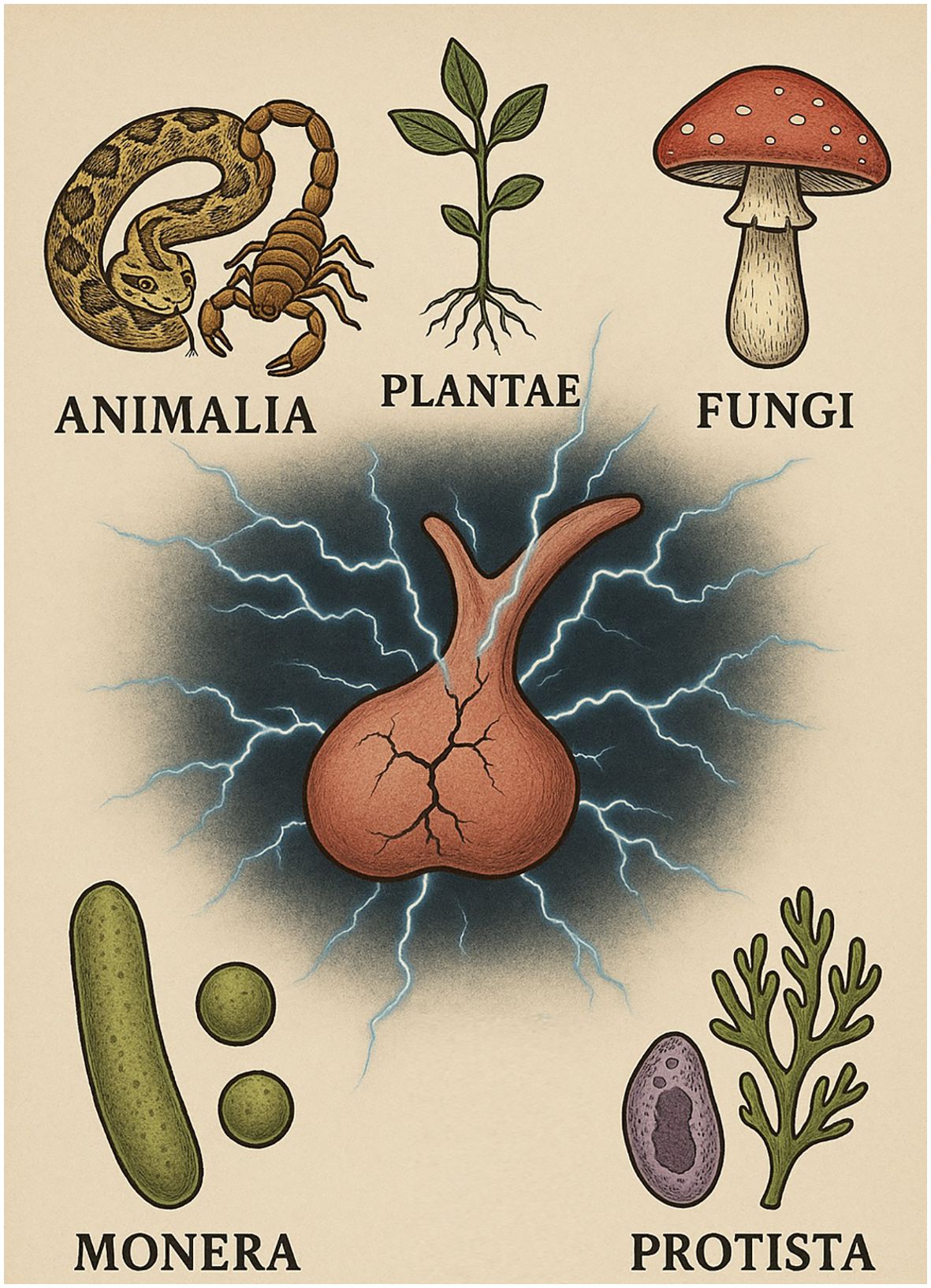
Figure 1. Schematic representation of the five biological kingdoms (Animalia, Plantae, Fungi, Monera and Protista) and their potential impact on pituitary function. AI-assisted visualization; verified by authors.
2.1 Kingdom Animalia
The kingdom Animalia comprises an immense variety of multicellular species, ranging from complex vertebrates to microscopic invertebrates, many of which have evolved specialized biochemical defenses or predatory tools in the form of venoms and toxins (23). These bioactive compounds often target critical physiological processes in prey or predators, such as the cardiovascular and nervous systems (24, 25). The endocrine system, and particularly the pituitary gland, can also be affected, most often in the context of snake envenomation, but also by venom components from other animal groups such as arachnids and insects, through mechanisms ranging from direct neuroendocrine interference to indirect vascular and inflammatory injury (26–28).
2.1.1 Snake envenomation
Snake envenomation is a major public health issue in tropical Asia, causing 1.8–2.7 million cases and 81,000–138,000 deaths annually, mostly in India (29, 30). It is a multisystemic condition involving renal failure, coagulopathy, myotoxicity, cardiac and neurological complications (31). Endocrine complications have also been described, most notably pituitary dysfunction, although adrenal and thyroid involvement have occasionally been reported (26, 32, 33).
The association between snakebite and hypopituitarism was first recognized in 1958, when Wolff reported a case following Bothrops jararacussu envenomation (34). However, the majority of clinical evidence originates from envenoming by Russell’s viper (Daboia russelii and D. siamensis), which has become the paradigm species linking snake venom to pituitary failure (35–39). Although most reports concern anterior pituitary dysfunction, involvement of the neurohypophysis has also been described, including cases of ADH deficiency (40–42).
Russell’s viper venom is a complex mixture of bioactive molecules, notably snake venom metalloproteinases, serine proteases, phospholipase A2, lectin-like proteins, disintegrins, and hyaluronidases (43, 44). These enzymes act synergistically on hemostasis, directly activating clotting factors V and X, driving rapid thrombin generation and fibrin formation, while thrombin-like enzymes accelerate fibrinogen consumption and fibrinolysis (Figure 2) (45). Concomitantly, metalloproteinases (“hemorrhagins”) damage vascular endothelium, and PLA2 and lectin-like proteins impair platelet function, further amplifying the coagulopathy (46, 47). The net effect is a state of profound endothelial injury with disseminated intravascular coagulation and spontaneous hemorrhages (48).
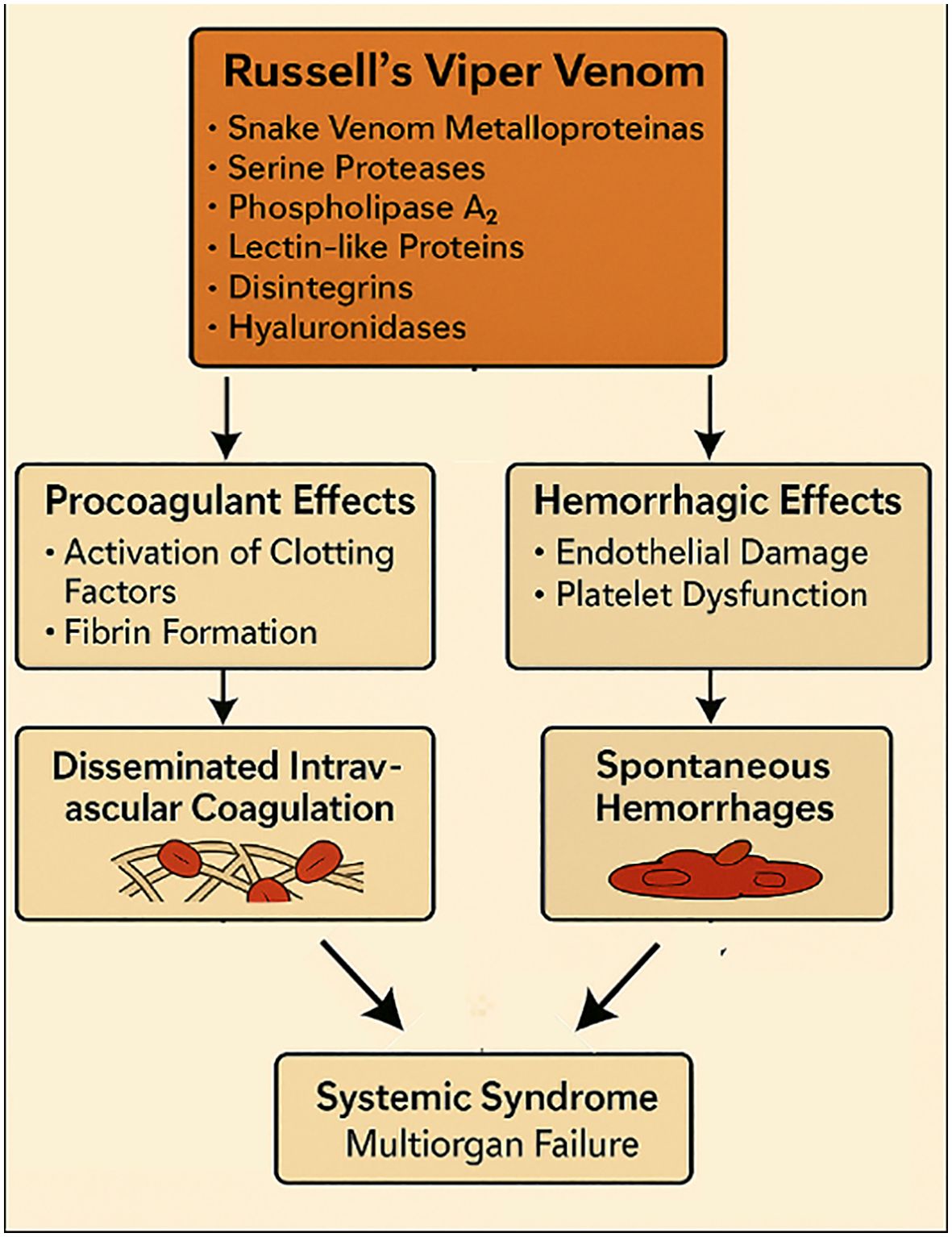
Figure 2. Mechanisms of Russell’s viper venom, including coagulopathy, endothelial injury, and disseminated intravascular coagulation leading to multiorgan failure. AI-assisted visualization; verified by authors.
Clinically, two forms are recognized: acute and chronic hypopituitarism (35). Diagnosis of the acute form is challenging because glucocorticoid therapy and non-thyroidal illness can mask hormonal deficits. Nevertheless, Tun-Pe et al. identified combined deficiencies of cortisol, prolactin, and growth hormone (GH) in nine patients with altered consciousness and hemodynamic instability after Russell’s viper envenomation (35). Similarly, Proby et al. described multiple hormonal deficits in patients with snakebite-associated acute kidney injury, with 10 of 15 exhibiting cortisol deficiency, 19 of 20 reduced thyroid stimulating hormone (TSH) and T4, and 12 of 17 men showing low testosterone with inappropriately normal gonadotropins (49). Although methodological limitations exist, the persistence of deficits in 11 of 12 patients argues against transient illness-related changes and supports true pituitary damage. Rajagopala et al. further reported that acute hypopituitarism typically emerges around 9 days after envenomation, with unexplained hypoglycemia (100%) and refractory hypotension (67%) as key clinical markers (36). Autopsy studies have revealed focal pituitary hemorrhages and fibrin thrombi consistent with ischemic and hemorrhagic injury, while other work has suggested a paradoxical early hypersecretion of TSH, GH, and adrenocorticotropic hormone (ACTH) within the first hours, probably reflecting toxin-induced stimulation rather than destruction (50, 51).
Chronic hypopituitarism is typically diagnosed years after the initial bite, with a mean delay of 8.1 ± 3.6 years (37). The nonspecific nature of symptoms—including fatigue, hypothyroidism, hypogonadism, and adrenal insufficiency—often accounts for diagnostic delay. Central hypothyroidism and gonadotropin deficiency are almost universal, with ACTH deficiency present in approximately 93% of cases (38). GH deficiency shows wider variability, ranging from 15% in the cohort studied by Bhat et al. to 100% in that of Naik et al. (39, 52) Radiological changes are frequent, as Ayan Roy et al. reported magnetic resonance imaging abnormalities in all patients studied (n=15), with partial (47%) or complete (53%) empty sella (38). Furthermore, stalk thinning and proximal T1 hyperintensity was identified in 23% and 15% respectively.
Predictors of pituitary dysfunction remain controversial. Naik et al. found no clear differentiating factors between patients with and without hypopituitarism (39). In contrast, Bhat et al. identified coagulopathy and the need for hemodialysis as predictors in patients with acute kidney injury, while Rajagopala et al. associated multiorgan failure, thrombocytopenia, and transfusion requirements with the later development of pituitary insufficiency (36, 39, 52). These endocrine alterations likely reflect a complex combination of direct ischemic–hemorrhagic injury to the gland and systemic inflammatory stress associated with coagulopathy, rather than a purely toxin-mediated pituitary effect.
Pituitary involvement has also been described beyond Russell’s viper. In fact, a case report of Gloydius blomhoffii envenomation documented transient secondary adrenal insufficiency presenting with severe hyponatremia, low cortisol, and inadequate responses to dynamic stimulation tests, in the context of a pre-existing empty sella (53).Furthermore, experimental work in rabbits showed that the neurotoxic fraction of Naja haje (Egyptian cobra) venom generates an hyperglycemic status, as evidenced by a paradoxical reduction in pituitary ACTH secretion alongside elevated circulating cortisol (54). Another noteworthy example is the green mamba (Dendroaspis angusticeps) venom, from which the peptide mambaquaretin (MQ1) has been isolated (55). Although it does not target the pituitary directly, this venom-derived peptide MQ1 is a highly selective vasopressin V2 receptor competitive antagonist, blocking β-arrestin and MAP kinase (MAPK) pathways, and in vivo producing a pure aquaretic effect, with potential therapeutic applications in disorders of vasopressin signaling such as polycystic kidney disease or inappropriate ADH secretion.
2.1.2 Other venomous species
Beyond reptiles, evidence from the phylum Arthropoda—particularly arachnids and insects—also points to the modulation of pituitary function (56). Daachi et al. reported that Androctonus australis hector scorpion venom increased ACTH and corticosterone levels and disrupted circadian rhythmicity. Histological analysis revealed enhanced ACTH immunoexpression in the anterior pituitary, together with increased oxidative markers and greater vascular permeability in the hypothalamus (57). In addition, venoms from species such as Leiurus quinquestriatus var. hebraeus and Buthus martensii have been used in more mechanistic studies of pituitary physiology, the former to investigate regulatory feedback in ACTH secretion and the latter to explore sodium current–dependent membrane potentials in anterior pituitary cells (27, 58). Moreover, α-latrotoxin, a component of black widow spider venom, has been shown to trigger vasopressin and oxytocin release through its interaction with calcium-independent receptors localized in the plasma membrane as well as calcium-dependent receptors for latrotoxin (59). Regarding insects, the available evidence is more limited but nonetheless illustrative. Sapeptin B, an antibacterial peptide from the flesh fly, has been shown to inhibit potassium currents in GH3 pituitary cells, while mastoparan, a wasp venom peptide, stimulates prolactin secretion in rats through increased intracellular calcium (28, 60). Finally, bee venom has been reported to deplete secretory granules in corticotrophs and somatotrophs, accompanied by elevated plasma ACTH and GH concentrations. These effects have led some authors to propose bee venom as a potential enhancer of growth performance and a modulator of pubertal timing in certain mammalian species (61, 62).
In these species, the observed hormonal changes are thought to result primarily from neuroendocrine stress responses rather than direct pituitary cytotoxicity, as no histological lesions or direct cellular injury have been demonstrated, supporting a predominantly neuroendocrine or systemic stress mechanism.
2.2 Kingdom Plantae
Kingdom Plantae is characterized by multicellular, autotrophic organisms with cell walls of cellulose and photosynthetic capacity (63). Although plants play a role in numerous human physiological functions and are widely used for their therapeutic potential, some plant-derived compounds can also exert harmful effects (64, 65). However, their influence on the hypothalamic–pituitary axis remains poorly understood. This section focuses on the impact of whole plants or natural extracts on pituitary function, excluding pharmacological agents or highly modified derivatives of plant origin —such as opioids (from Papaver somniferum) or aspirin (from Salix alba)— as well as synthetic endocrine disruptors.
The vast majority of scientific evidence regarding the influence of plants on pituitary function relates to their beneficial effects, particularly in the modulation of the stress response, reproductive health and obesity via the hypothalamic–pituitary–adrenal (HPA) axis (66–70). In contrast, evidence on potentially harmful or disruptive effects—whether functional or structural—is limited and often anecdotal, highlighting a gap in current research. This imbalance is further underscored by the fact that certain plant species, such as Ginkgo biloba and Garcinia kola, have even been investigated as protective agents against known toxicants like lead acetate or sodium arsenate, owing to their antioxidant and anti-inflammatory properties (71, 72).
Several plant species have been shown to modulate the hypothalamic–pituitary–thyroid (HPT) axis. Extracts of Lithospermum officinale, Lycopus virginicus, Melissa officinalis, and Thymus serpyllum have demonstrated the ability to suppress both serum and pituitary TSH levels in animal models, even under hypothyroid conditions (73). These effects appear to combine central hormone-blocking actions with peripheral thyroid hormone-like activity. Other species such as Chelidonium majus, Curcuma longa, Dorema aucheri, and Peganum harmala contain flavonoids or alkaloids capable of modulating thyrotropin–releasing hormone (TRH) and TSH secretion via dopaminergic, serotonergic, or second messenger pathways (74). Specifically, flavonoids from D. aucheri and Humulus lupulus inhibit thyroid peroxidase and type-1 deiodinase, limiting iodine organification and peripheral T3 formation, whereas alkaloids from C. majus increase TRH and TSH release by blocking catechol-O-methyltransferase and enhancing calcium–phosphatidylinositol signaling in pituitary thyrotrophs. In contrast, harmaline alkaloids from P. harmala suppress TRH secretion through monoamine oxidase inhibition and serotonin accumulation, effects further modulated by leptin-mediated inhibition of neuropeptide Y neurons.
Regarding the hypothalamic–pituitary–gonadal (HPG) axis, traditional extracts from Cola nitida, Afrormosia laxiflora, and Pterocarpus erinaceus have been reported to inhibit ovulation and disrupt the estrous cycle in experimental models (75). In vitro studies further support these observations, showing that these plants selectively inhibit luteinizing hormone (LH) release in cultured rat pituitary cells, without affecting follicle stimulating hormone (FSH) secretion or cell viability (76). This inhibition appears to result from the formation of heterodimers between plant-derived compounds and basic glycoproteins, suggesting a non-cytotoxic, extracellular mechanism of gonadotropin modulation. Furthermore, Li et al. found that exposure to swainsonine (an indolizidine alkaloid) during pregnancy in mice led to significant disruption of reproductive hormone secretion (77). This effect was linked to impaired glycoprotein function in the anterior pituitary, resulting from the alteration of the glycosylation pattern of gonadotropins. Interestingly, some saponins such as those from Quillaja saponaria and Gypsophila paniculata have shown a stimulatory effect on LH secretion in pituitary cell cultures, in contrast to the inhibitory effects reported for other plant compounds (78). These triterpenoid and steroidal glycosides interact with membrane cholesterol, modifying lipid microdomains and, in some cases, inducing pore formation that facilitates Ca2+ influx and exocytotic release of gonadotropins in cultured pituitary cells. Notably, G. paniculata exhibits marked hemolytic activity whereas soybean saponins appear less lytic, suggesting structural heterogeneity within their sapogenin cores.
The effects of various plant-based formulations on prolactin secretion have been studied both in terms of increasing and suppressing its levels (79). Most of this research has focused on potentially beneficial outcomes—such as enhancing prolactin to support lactation, or reducing it in cases of hyperprolactinemia—rather than investigating possible dysregulation or adverse effects on pituitary control (80, 81). In line with this therapeutic perspective, two natural compounds derived from Glycyrrhiza glabra radix—18β-glycyrrhetinic acid and liquiritigenin—have shown antiproliferative and pro-apoptotic activity in prolactin-secreting pituitary adenoma cells (82, 83). These effects were mediated by distinct intracellular pathways (ROS–CaMKII–JNK/P38 activation and Ras/ERK inhibition, respectively) and were accompanied by a consistent reduction in prolactin synthesis both in vitro and in vivo, supporting their potential utility in the management of prolactin-related pituitary disorders.
Despite these observations, most data on plant-derived compounds come from experimental or preclinical settings, with considerable variability in extraction methods, active constituents, and dosing. The absence of standardized preparations and controlled human studies continues to limit the interpretation and translational relevance of these findings.
2.3 Kingdom Monera
Monera comprises prokaryotic organisms characterized by the absence of a true nucleus and membrane-bound organelles (84). Beyond their well-recognized roles in infectious disease, members of this kingdom can influence endocrine function through structural components and bioactive metabolites that interact with the hypothalamic–pituitary axes without directly invading the gland, instead acting via systemic mediators. (Figure 3) (85) This section focuses on pituitary modulation mediated by bacterial components or metabolites and, as discussed previously, direct pituitary infections—such as abscess formation— are not included in the present analysis.
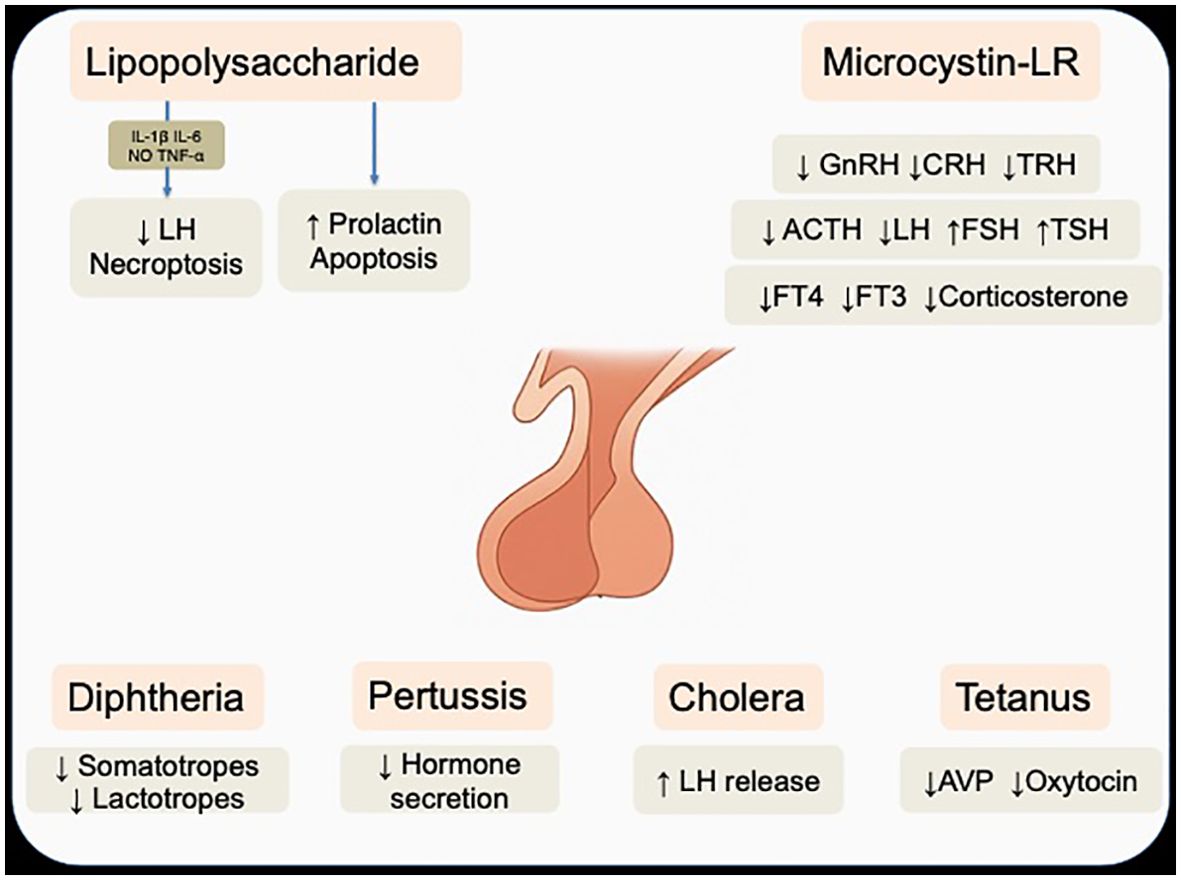
Figure 3. Effects of LPS, microcystin, and classical bacterial toxins on pituitary hormone regulation and cellular function. Note that lipopolysaccharide can act either directly on pituitary cells or indirectly through cytokine-mediated pathways. Acronyms: IL-1β, interleukin-1β; IL-6, interleukin-6, TNF-α, Tumor necrosis factor-α; NO, nitric oxide; GnRH, gonadotropin–releasing hormone; CRH, corticotropin–releasing hormone; TRH, thyrotropin–releasing hormone; ACTH, adrenocorticotropic hormone; LH, luteinizing hormone; FSH, follicle stimulating hormone; TSH, thyroid stimulating hormone; FT4, free T4; FT3, free T3; ADH, antidiuretic hormone. Note that the anatomical illustration of the pituitary gland was AI-generated using OpenAI DALL·E (2025) and verified by authors.
2.3.1 Effect of lipopolysaccharide on pituitary function
Lipopolysaccharide (LPS) is a major structural component of the outer membrane of Gram-negative bacteria, composed of a lipid A moiety, a core oligosaccharide, and an O-antigen polysaccharide chain (86). It acts as a potent endotoxin by binding to Toll-like receptor 4 (TLR4) and triggering a strong innate immune response (87).
Evidence indicates that LPS can modulate prolactin secretion through both indirect and direct pituitary mechanisms. Regarding indirect mechanisms, LPS binds to TLR4 via LPS-binding protein and CD14, triggering the release of proinflammatory cytokines such as interleukin (IL) -1β, IL-6, and tumor necrosis factor-α (TNF-α) (88). These mediators can suppress LH secretion from gonadotropes, stimulate prolactin release via folliculostellate cell-derived IL-6, remodel the pituitary microenvironment (e.g., increased aquaporin-4 expression), and activate necroptosis in the hypothalamus, pituitary, and adrenal glands (89–92). Other mediators such as nitric oxide (NO) also participate, influencing LPS-induced prolactin release in a manner dependent on thyroid status (93). Furthermore, developmental exposure, particularly during the prenatal period, disrupts gonadotropin–releasing hormone (GnRH) neuron maturation and leads to long-term reductions in GnRH and LH and altered sex steroid profiles, whereas chronic low-dose exposure may paradoxically increase gonadotropin output through pituitary transcriptomic reprogramming (94, 95). In addition, LPS can act directly on endocrine cells, as lactotrophs express functional TLR4; stimulation promotes their proliferation and prolactin synthesis via MAPK and nuclear factor κB (NF-κB) activation—effects abolished by TLR4 blockade—demonstrating that bacterial endotoxin can influence pituitary hormonal output and cell growth independently of hypothalamic input (96). However, these direct effects have been demonstrated exclusively in experimental models, and their physiological relevance in humans with sepsis remains speculative.
On the other hand, under conditions where NF-κB–dependent survival signaling is impaired, either pharmacologically or via estradiol, LPS exposure can also trigger apoptosis in anterior pituitary cells, particularly lactotrophs and somatotrophs, accompanied by reduced expression of the anti-apoptotic protein Bcl-xL (97). Notably, LPS has also been shown to suppress the growth of TLR4-positive pituitary adenomas, an effect closely linked to IL-6 production via p38αMAPK, as evidenced by its reversal with specific kinase inhibitors (98).
2.3.2 Bacterial toxins and pituitary modulation
Cyanobacteria, commonly known as “blue–green algae,” are photosynthetic prokaryotes that inhabit a wide range of aquatic environments and can form dense blooms under eutrophic conditions (99). Many cyanobacterial species produce potent secondary metabolites, including microcystins, which are cyclic heptapeptide hepatotoxins with recognized endocrine-disrupting potential (100).
In mammals, exposure to microcystin-LR (MC-LR) has been reported to reduce hypothalamic GnRH and Kiss1 transcript expression, accompanied by decreased GnRH concentrations but increased FSH levels, while effects on LH appear variable, with most studies reporting elevations, although Dos Santos et al. observed reduced levels (101–103).These endocrine alterations are associated with impaired gametogenesis and reduced ovarian follicle counts, indicating downstream reproductive toxicity (104). In addition to its effects on the reproductive axis, MC-LR disrupts the HPA axis by reducing hypothalamic corticotropin–releasing hormone (CRH) expression, circulating ACTH, and plasma corticosterone, and impairs the HPT axis by decreasing hypothalamic TRH expression and free T4 and T3 concentrations, while concomitantly increasing TSH levels in a compensatory manner (102, 103).
In fish and amphibians, waterborne MC-LR exposure disrupts the hypothalamic–pituitary–gonadal–liver axis, reducing GnRH, FSHβ, LHβ and LH receptor mRNA expression, impairing vitellogenin synthesis, and inducing gonadal dysplasia, often with sex-specific transcriptional profiles (105, 106).
Early life-stage exposures in zebrafish have demonstrated activation and dysregulation of the hypothalamic–pituitary–interrenal (HPI) axis, with an hypothalamic CRH and pituitary proopiomelanocortin (POMC) expression, elevated cortisol and altered glucocorticoid receptor expression, indicating feedback dysregulation and suggesting potential long-term programming effects (107). Combined exposures with other aquatic contaminants, such as nitrite, can exacerbate these endocrine disruptions through synergistic effects (108).
Some toxins from classical pathogenic bacteria also show pathogenic relevance for pituitary function. In rodents, the Staphylococcus aureus superantigen enterotoxin B and Clostridium difficile toxin A both increase plasma corticosterone and ACTH levels, the latter likely acting through the release of inflammatory mediators such as TNF-α and prostaglandin E2 (109–111). In humans, vaccination with tetanus toxoid has been associated with transient rises in cortisol and ACTH, consistent with acute HPA axis activation (112).
By contrast, although several other bacterial toxins have been investigated in relation to pituitary function, most evidence derives from animal models in which these molecules serve as experimental tools rather than as causes of clinically relevant pituitary dysfunction. For example, Diphtheria toxin has been employed in transgenic mouse models to conditionally ablate specific pituitary cell types, such as somatotropes or lactotropes, via inducible expression of the diphtheria toxin receptor (113–115). These models have revealed a remarkable regenerative capacity of the adult pituitary, involving Sox2+ stem/progenitor cell activation, expansion of the marginal-zone niche, proliferation of surviving endocrine cells, and, in some cases, transdifferentiation between lineages. Pertussis toxin, through ADP-ribosylation of Gi/o proteins, has been instrumental in demonstrating that somatostatin receptor signaling in pituitary cells is Gi/o-dependent, clarifying inhibitory pathways that regulate hormone secretion (116–118). Cholera toxin, in contrast, enhances GnRH-induced LH release by shifting hormone stores from a nonreleasable to a releasable pool via Gs protein activation and cyclic adenosine monophosphate (cAMP) production, a sensitization lost during desensitization but recoverable under specific stimulation patterns (119). Clostridial toxins have been used to probe vesicular trafficking and cytoskeletal regulation, for example showing that Clostridium tetani toxin blocks synaptobrevin-dependent exocytosis of vasopressin and oxytocin, and that other Clostridium toxins can induce actin depolymerization and aquaporin-2 translocation (120, 121). Collectively, these studies have provided valuable mechanistic insight into pituitary cell biology, although most represent experimental paradigms rather than evidence of clinically relevant pituitary pathology.
2.4 Kingdom Fungi
The kingdom Fungi comprises a diverse group of eukaryotic organisms that includes yeasts, molds, and mushrooms (122). Characterized by a chitinous cell wall and heterotrophic metabolism, fungi play essential ecological roles as decomposers and symbionts, but also produce a wide array of bioactive metabolites (123). Some of these secondary metabolites have been recognized as potent modulators of endocrine function, capable of affecting the hypothalamic–pituitary axes.
2.4.1 Mycotoxin T-2 and pituitary impairment
Most of the current scientific evidence on pituitary dysfunction related to fungi centers around T-2 toxin, a trichothecene mycotoxin produced by Fusarium species (124). This compound gained notoriety in the 1940s in the former Soviet Union, where contaminated grain led to outbreaks of alimentary toxic aleukia—a severe disease characterized primarily by hematological suppression, mucosal ulceration, and high mortality (125). T-2 toxin is known to widely contaminate barley, maize, oats, and even processed human food, raising ongoing concerns about its impact on both public health and endocrine function (126).
The effects of T-2 toxin on the HPG axis appear to be highly context-dependent, varying with dose, sex, and developmental stage. In adult male mice, chronic oral exposure to T-2 toxin led to a dose-dependent decline in fertility and disrupted spermatogenesis, which correlated with reduced serum levels of GnRH, LH, FSH, testosterone, and progesterone (127). These endocrine alterations were supported by suppressed expression of hypothalamic GnRH1 and pituitary LHβ mRNA. In contrast, prepubertal exposure in female rats resulted in the opposite pattern: accelerated pubertal onset, increased circulating LH, FSH, and estradiol, and upregulation of GnRH and GnRHR expression, along with histological signs of premature reproductive maturation (128). These divergent effects suggest that T-2 toxin may suppress or activate the HPG axis depending on the physiological window, possibly through differential modulation of upstream regulators such as kisspeptin signaling.
Regarding the somatotropic axis, T-2 toxin significantly reduces GH synthesis and secretion in GH3 pituitary cells, primarily through excessive NO production via inducible NO synthase (129). This leads to mitochondrial dysfunction, oxidative stress, and caspase-mediated apoptosis, further enhanced by proinflammatory cytokines such as IL-6, IL-1β, and IL-11. These findings align with the well-established cytotoxic role of NO in the pituitary, contributing to both cell death and hormonal deficiencies (130, 131).
Beyond its effects on hormone secretion, T-2 toxin has been shown to cause direct structural damage to the pituitary gland. In fact, Pu Guo et al. demonstrated that T-2 crosses the blood–brain barrier and induces distinct histopathological changes in the anterior pituitary, including early signs of vascular congestion and hemorrhage, followed by apoptotic degeneration (132). The pituitary responded primarily with apoptosis, unlike other brain regions where autophagy was more prominent, suggesting tissue-specific mechanisms of toxicity.
2.4.2 Other toxins and fungal molecules
Regarding other mycotoxins, zebrafish models have shown that both zearalenone (ZEA) and ochratoxin A (OTA) can disrupt pituitary-regulated endocrine function through distinct mechanisms. ZEA, a mycotoxin produced by Fusarium species, selectively altered LH-related pathways in adult females, as evidenced by upregulation of LHr and multiple genes involved in estrogen synthesis and steroidogenesis, while FSH receptor expression remained unchanged—suggesting a targeted effect on LH-mediated signaling (133). In contrast, OTA—produced mainly by Aspergillus and Penicillium species—did not affect prolactin release itself but interfered with its downstream action by upregulating prolactin receptor expression following miR-731 suppression in embryonic zebrafish, contributing to vascular instability (134). In addition, the mycotoxin deoxynivalenol, produced by Fusarium species and commonly found in contaminated cereals, has been shown to activate the HPA axis in a necroptosis-dependent manner, as evidenced by the attenuation of these effects with necrostatin-1, a well-recognized necroptosis inhibitor (135).
On the other hand, species of Amanita are among the most dangerous wild mushrooms due to their highly toxic compounds, including amatoxins and phallotoxins, and pose a risk of accidental ingestion (136). In relation to endocrine disregulation, Amanita mushroom poisoning resulted in suppressed thyroid function, with reduced thyroxine levels and undetectable triiodothyronine accompanied by inappropriately low or normal TSH concentrations in most cases, suggesting impaired hypothalamic–pituitary responsiveness or a euthyroid sick syndrome (137). Whether Amanita exerts a direct cytotoxic effect on pituitary cells remains unknown; this contrasts with other endocrine organs, such as the pancreas, where β-cell injury has been reported (138).
Beyond classical mycotoxins, other fungal-derived compounds may also modulate hypothalamic–pituitary function. For instance, β-glucan—a polysaccharide present in fungal cell walls—has been shown to stimulate prolactin secretion both in vivo, after intravenous administration, and ex vivo in incubated pituitary tissue (139).
Pectin derivatives may exert similar effects, suggesting that certain structural polysaccharides can influence anterior pituitary hormone release.
From a mechanistic standpoint, findings from different fungal species converge on a limited set of cellular processes—oxidative stress, inhibition of protein synthesis, and apoptotic signaling—suggesting a conserved pattern of endocrine cell injury related to redox imbalance and mitochondrial dysfunction. Such mechanisms parallel those observed in bacterial endotoxins and other biologically derived compounds, pointing to a broader susceptibility of hormone-secreting cells to metabolic and oxidative stress. Nevertheless, most available data originate from in vitro or rodent experiments, often employing supraphysiological concentrations, and human evidence remains largely inferential, based on dietary or environmental exposure rather than direct clinical observation.
2.5 Kingdom Protista
Protists are a diverse group of eukaryotic microorganisms traditionally defined by exclusion: they are not animals, plants, or fungi, yet share certain cellular features with each. Most are unicellular, though some form simple multicellular or colonial structures. They exhibit a wide range of nutritional strategies—including autotrophy, heterotrophy, and mixotrophy—and possess various forms of motility such as flagella, cilia, or pseudopodia (140). Despite their diversity, in practical terms the kingdom Protista is often represented by two major categories of medical and ecological interest: marine microalgae and pathogenic parasites.
2.5.1 Marine microalgae
Scientific evidence is limited in this group and derives almost exclusively from two species: Gambierdiscus toxicus and Prymnesium patelliferum. In more recent taxonomic systems these species are reclassified under the kingdom Chromista. Gambierdiscus is typically placed within the Alveolata group, while Prymnesium belongs to Haptophyta (141, 142).
Gambierdiscus toxicus is a marine dinoflagellate known as the primary source of ciguatoxin, a lipophilic neurotoxin responsible for ciguatera fish poisoning —a foodborne illness characterized by gastrointestinal, neurological, and cardiovascular disturbances following the ingestion of contaminated reef fish (143). Interestingly, another compound produced by this organism, maitotoxin, has been shown to interact directly with the pituitary gland, inducing a marked increase in intracellular calcium flux in pituitary cells, likely mediated by elevated levels of inositol trisphosphate (144). This calcium influx leads to enhanced secretion of several pituitary hormones, including GH, LH, and prolactin (145). However, the regulation of prolactin secretion appears to involve additional complexity, potentially mediated by an increase in leukotriene production (146). Notably, the secretagogue effect of maitotoxin seems to be specific to the pituitary gland, as it has been shown to inhibit parathyroid hormone secretion (147).
A similar mechanism has been observed with the ichthyotoxic flagellate Prymnesium patelliferum, whose toxin also enhanced calcium influx in GH4C1 pituitary cells by activating both T-type and L-type voltage-gated calcium channels, leading to elevated intracellular calcium levels and a dose-dependent increase in prolactin secretion (148).
2.5.2 Pathogenic parasites
Infection with Plasmodium spp. has been associated with impairment of the pituitary gland. Acute malaria typically activates the HPA axis, with elevated cortisol and DHEA levels, likely triggered by TNF-α and IL-1 release following erythrocyte rupture and antigen exposure during the parasitic replication phase, which acutely stimulate both the thermoregulatory center and corticotropic pathways (149). In contrast, prolonged disease may result in selective adrenal exhaustion, reflected in declining DHEA concentrations. Transient pituitary dysfunction has also been observed in the HPT axis, with low thyroxine levels and blunted TSH responses to TRH stimulation during severe malaria, which normalize upon recovery (150). However, these alterations are most likely driven by systemic inflammation due to TNF-α and interferon gamma (IFN-γ) oversecretion and metabolic adaptation to critical illness, rather than specific peptide-mediated pituitary toxicity (151).
Several protozoan infections have been associated with pituitary dysfunction. but current evidence suggests that such alterations are not mediated by selective distant pituitary modulators. In Trypanosoma spp. infections, extensive data from both animal models and human studies reveal central endocrine disturbances, such as ACTH exhaustion and corticotroph remodeling in T. brucei brucei, persistent hypogonadism in T. rhodesiense, and central hypothyroidism in T. evansi (152–154). Findings in Trypanosoma congolense infections are less consistent, with conflicting reports regarding ACTH dynamics, although the detection of parasites in both cerebrospinal fluid and pituitary microvasculature suggests direct involvement of the gland (155–157). Similarly, other protozoa such as Toxoplasma gondii and Leishmania spp. have been investigated for their effects on pituitary axes, particularly the HPG axis (158). Chronic T. gondii infection triggers sustained systemic inflammation, with IL-1 and TNF-α reaching the hypothalamus and stimulating corticotropin-releasing hormone (CRH) secretion, thereby activating the HPA axis and secondarily suppressing the HPG axis through inhibition of GnRH release. Similarly, elevated TNF-α and IFN-γ levels reported in visceral leishmaniasis may further impair hypothalamic and gonadal function, providing a plausible immuno-endocrine mechanism linking protozoan infection to reproductive axis dysfunction. Yet, no pituitary-specific toxins have been identified, and endocrine manifestations in these infections are most likely secondary to systemic cytokine-driven inflammation, though limited evidence suggests that direct pituitary involvement may occasionally occur (159–162).
Taken together, evidence from parasitic and non-parasitic protists points to two predominant processes of endocrine disruption: (1) cytokine-driven activation of hypothalamic–pituitary pathways during systemic infection, and (2) calcium-dependent exocytotic or cytopathic effects induced by marine metabolites. These phenomena illustrate how distinct protistan lineages can trigger comparable neuroendocrine alterations through immune signaling and intracellular calcium dynamics. However, most of these data derive from experimental systems rather than human studies, and the direct endocrine relevance of these mechanisms in clinical settings remains to be confirmed.
After reviewing the different biological kingdoms, the main biologically derived agents and their effects on pituitary function can be summarized comparatively. Table 1 provides an integrative overview of representative toxins, their principal mechanisms, and documented endocrine outcomes according to the available type of evidence.
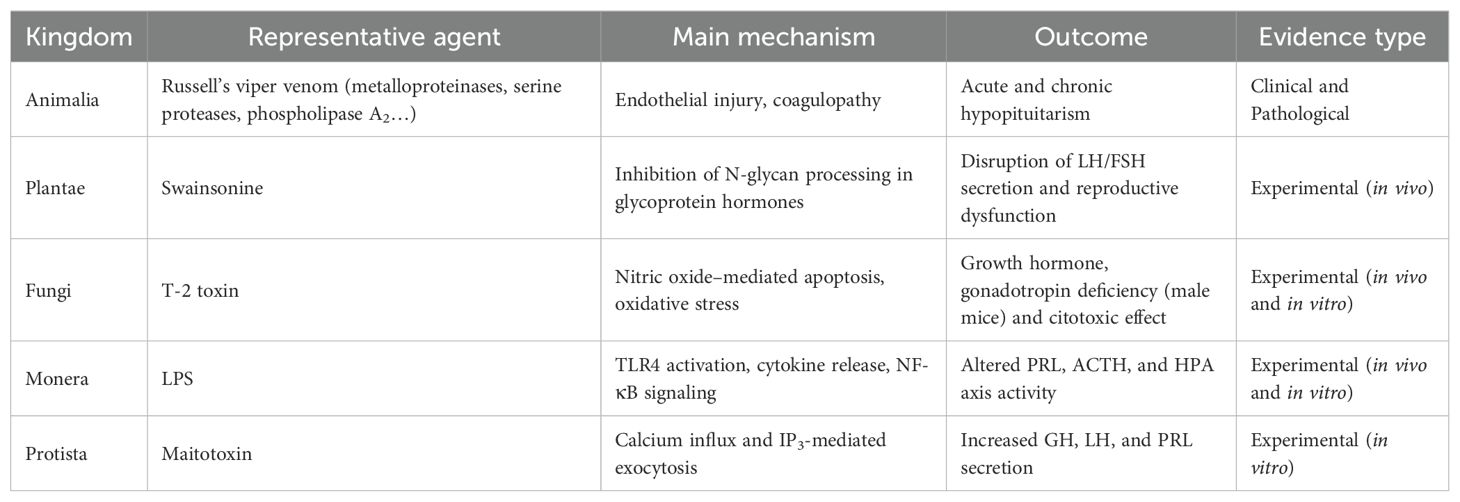
Table 1. Representative natural toxins affecting pituitary function, summarizing their main mechanisms, endocrine outcomes, and supporting evidence.
3 Conclusions
Across the five biological kingdoms, diverse natural agents demonstrate the pituitary’s vulnerability to toxic injury. Russell’s viper envenomation is a well-documented cause of both acute and chronic hypopituitarism, while other animals can also alter pituitary responses. Plants are mainly studied for their therapeutic potential, yet certain extracts interfere with gonadotropin secretion and reproductive regulation. Fungal metabolites, particularly the trichothecene T-2 toxin, exert direct cytotoxic damage on pituitary cells, raising concerns about dietary exposure. Among kingdom Monera, LPS can disrupt pituitary function both indirectly through systemic inflammation and directly at the cellular level, also highlighting the influence of bacterial toxins. Protists complete this spectrum, with marine microalgae producing potent secretagogues of pituitary hormones, while pathogenic protozoa impair endocrine axes largely through systemic inflammatory mechanisms. Together, these findings underscore the pituitary’s remarkable impairment to biologically derived agents and the need for greater clinical awareness of these impactful toxic insults. Clinicians should maintain a high index of suspicion for pituitary dysfunction in acute toxic exposures, as early assessment of ACTH, cortisol, and TSH levels may unmask evolving hypopituitarism and guide timely management.
Author contributions
DS: Data curation, Writing – review & editing, Investigation, Writing – original draft, Formal Analysis, Software. RC: Conceptualization, Investigation, Writing – review & editing, Writing – original draft, Supervision. PM: Investigation, Conceptualization, Writing – review & editing, Writing – original draft. DL: Writing – original draft, Investigation, Writing – review & editing, Conceptualization. VP: Writing – original draft, Methodology, Conceptualization, Writing – review & editing. PM: Writing – review & editing, Writing – original draft, Conceptualization, Data curation. JM: Writing – review & editing, Conceptualization, Investigation, Supervision, Methodology, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors acknowledge the use of artificial intelligence tools (OpenAI DALL·E and ChatGPT, 2025) for figure generation and minor language refinement during manuscript preparation. All authors reviewed and approved the use of these tools and remain fully responsible for the content, interpretation, and conclusions of the work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer PI declared a past co-authorship with the author RC to the handling editor.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Artificial intelligence was employed to create Figures 1 and 2, and partially Figure 3.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Sav A, Menna G, Serra C, Söztutar E, and Türe U. Anatomy of the pituitary gland. Best Pract Res Clin Endocrinol Metab. (2025) 39. doi: 10.1016/j.beem.2025.102013
2. Le Tissier P, Fiordelisio Coll T, and Mollard P. The processes of anterior pituitary hormone pulse generation. Endocrinology. (2018) 159:3524–35. doi: 10.1210/en.2018-00508
3. Brown CH. Magnocellular neurons and posterior pituitary function. Compr Physiol. (2016) 6:1701–41. doi: 10.1002/cphy.c150053
4. Le Tissier P, Campos P, Lafont C, Romanò N, Hodson DJ, and Mollard P. An updated view of hypothalamic-vascular-pituitary unit function and plasticity. Nat Rev Endocrinol. (2017) 13:257–67. doi: 10.1038/nrendo.2016.193
5. Sam S and Frohman LA. Normal physiology of hypothalamic pituitary regulation. Endocrinol Metab Clin North Am. (2008) 37:1–22. doi: 10.1016/j.ecl.2007.10.007
6. Garmes HM. Special features on insulin resistance, metabolic syndrome and vascular complications in hypopituitary patients. Rev Endocr Metab Disord. (2024) 25:489–504. doi: 10.1007/s11154-023-09872-8
7. Gogakos AI, Gogakos T, Kita M, and Efstathiadou ZA. Pituitary dysfunction as a cause of cardiovascular disease. Curr Pharm Des. (2020) 26:5573–83. doi: 10.2174/1381612824999201105165351
8. Herman JP, McKlveen JM, Ghosal S, Kopp B, Wulsin A, Makinson R, et al. Regulation of the hypothalamic-pituitary- adrenocortical stress response. Compr Physiol. (2016) 6:603–21. doi: 10.1002/cphy.c150015
9. Prodam F, Caputo M, Mele C, Marzullo P, and Aimaretti G. Insights into non-classic and emerging causes of hypopituitarism. Nat Rev Endocrinol. (2021) 17:114–29. doi: 10.1038/s41574-020-00437-2
10. Fleseriu M, Christ-Crain M, Langlois F, Gadelha M, and Melmed S. Hypopituitarism. Lancet. (2024) 403:2632–48. doi: 10.1016/S0140-6736(24)00342-8
11. Jasim S, Alahdab F, Ahmed AT, Tamhane S, Prokop LJ, Nippoldt TB, et al. Mortality in adults with hypopituitarism: a systematic review and meta-analysis. Endocrine. (2017) 56:33–42. doi: 10.1007/s12020-016-1159-3
12. Pappachan JM, Raskauskiene D, Kutty VR, and Clayton RN. Excess mortality associated with hypopituitarism in adults: A meta-analysis of observational studies. J Clin Endocrinol Metab. (2015) 100:1405–11. doi: 10.1210/jc.2014-3787
13. Ebrahimi F, Kutz A, Wagner U, Illigens B, Siepmann T, Schuetz P, et al. Excess mortality among hospitalized patients with hypopituitarism—A population-based, matched-cohort study. J Clin Endocrinol Metab. (2020) 105:1–9. doi: 10.1210/clinem/dgaa517
14. Tritos NA and Miller KK. Diagnosis and management of pituitary adenomas: A review. JAMA. (2023) 329:1386–98. doi: 10.1001/jama.2023.5444
15. Iglesias P. Pituitary apoplexy: an updated review. J Clin Med. (2024) 13:2508. doi: 10.3390/jcm13092508
16. Feldt-Rasmussen U and Klose MC. Pathophysiology and diagnosis of neuroendocrine abnormalities in patients with traumatic brain injury. Best Pract Res Clin Endocrinol Metab. (2025) 39:102020. doi: 10.1016/j.beem.2025.102020
17. Angelousi A, Alexandraki K, Tsoli M, Kaltsas G, and Kassi E. Hypophysitis (Including IgG4 and immunotherapy). Neuroendocrinology. (2020) 110:822–35. doi: 10.1159/000506903
18. Iplikcioglu AC, Bek S, Bikmaz K, Ceylan D, Gökduman CA, and Bayston R. Aspergillus pituitary abscess. Acta Neurochir (Wien). (2004) 146:521–4. doi: 10.1007/s00701-004-0256-x
19. Bouter J, Reznik Y, and Thariat J. Effects on the hypothalamo-pituitary axis in patients with CNS or head and neck tumors following radiotherapy. Cancers (Basel). (2023) 15:3820. doi: 10.3390/cancers15153820
20. Makras P, Erickson D, Davidge-Pitts CJ, Diamond EL, Allen CE, McClain KL, et al. Approach to the patient: from endocrinopathy to the diagnosis of a histiocytic disorder. J Clin Endocrinol Metab. (2025) 110:1756–66. doi: 10.1210/clinem/dgae827
21. Whittaker RH. New concepts of kingdoms of organisms. Evolutionary relations are better represented by new classifications than by the traditional two kingdoms. Science. (1969) 163:150–60. doi: 10.1126/science.163.3863.150
22. Cavalier-Smith T. A revised six-kingdom system of life. Biol Rev Camb Philos Soc. (1998) 73:203–66. doi: 10.1017/s0006323198005167
23. Herzig V, Cristofori-Armstrong B, Israel MR, Nixon SA, Vetter I, and King GF. Animal toxins — Nature’s evolutionary-refined toolkit for basic research and drug discovery. Biochem Pharmacol. (2020) 181:114096. doi: 10.1016/j.bcp.2020.114096
24. Avalo Z, Barrera MC, Agudelo-Delgado M, Tobón GJ, and Cañas CA. Biological effects of animal venoms on the human immune system. Toxins (Basel). (2022) 14:344. doi: 10.3390/toxins14050344
25. Mathis S, Carla L, Duval F, Nadal L, Solé G, and Le Masson G. Acute peripheral neuropathy following animal envenomation: A case report and systematic review. J Neurol Sci. (2022) 442:120448. doi: 10.1016/j.jns.2022.120448
26. Bhattacharya S, Krishnamurthy A, Gopalakrishnan M, Kalra S, Kantroo V, Aggarwal S, et al. Endocrine and metabolic manifestations of snakebite envenoming. Am J Trop Med Hyg. (2020) 103:1388. doi: 10.4269/ajtmh.20-0161
27. Bauer CK, Krylov B, Zhou PA, and Schwarz JR. Effects of Buthus martensii (Karsch) scorpion venom on sodium currents in rat anterior pituitary (GH3/B6) cells. Toxicon. (1992) 30:581–9. doi: 10.1016/0041-0101(92)90852-V
28. Mau SE, Witt MR, and Vilhardt H. Mastoparan, a wasp venom peptide, stimulates release of prolactin from cultured rat anterior pituitary cells. J Endocrinol. (1994) 142:9–18. doi: 10.1677/joe.0.1420009
29. Gutiérrez JM, Calvete JJ, Habib AG, Harrison RA, Williams DJ, and Warrell DA. Snakebite envenoming. Nat Rev Dis Primers. (2017) 3:1–21. doi: 10.1038/nrdp.2017.63
30. Mohapatra B, Warrell DA, Suraweera W, Bhatia P, Dhingra N, Jotkar RM, et al. Snakebite mortality in India: A nationally representative mortality survey. PLoS Negl Trop Dis. (2011) 5:e1018. doi: 10.1371/journal.pntd.0001018
31. Seifert SA, Armitage JO, and Sanchez EE. Snake envenomation. N Engl J Med. (2022) 386:68–78. doi: 10.1056/NEJMra2105228
32. Shetty S, Abi MS, Hickson R, David T, and Rajaratnam S. Hypopituitarism and autoimmune thyroiditis following snake bite: an unusual clinical scenario. J Assoc Physicians India. (2014) 11:55–7.
33. Senthilkumaran S, Almeida JR, Williams J, Williams HF, Thirumalaikolundusubramanian P, Patel K, et al. Rapid identification of bilateral adrenal and pituitary haemorrhages induced by Russell’s viper envenomation results in positive patient outcome. Toxicon. (2023) 15:225. doi: 10.1016/j.toxicon.2023.107068
34. Wolff H. Is it time to denominate hypopituitarism after snake bite? Q J Med. (2013) 106:390. doi: 10.1093/qjmed/hct007
35. Tun-Pe, Warrell DA, Tin-Nu-Swe, Phillips RE, Moore RA, Myint-Lwin, et al. Acute and chronic pituitary failure resembling sheehan’s syndrome following bites by Russell’s viper in Burma. Lancet. (1987) 330:763–7. doi: 10.1016/s0140-6736(87)92500-1
36. Rajagopala S, Thabah MM, Ariga KK, and Gopalakrishnan M. Acute hypopituitarism complicating Russell’s viper envenomation: Case series and systematic review. Q J Med. (2015) 108:719–28. doi: 10.1093/qjmed/hcv011
37. Shivaprasad C, Aiswarya Y, Sridevi A, Anupam B, Amit G, Rakesh B, et al. Delayed hypopituitarism following Russell’s viper envenomation: a case series and literature review. Pituitary. (2019) 22:4–12. doi: 10.1007/s11102-018-0915-1
38. Roy A, Suryadevara V, Nagarajan K, Sahoo J, Naik D, Perumal NL, et al. Pituitary dysfunction following snakebite envenomation: A clinico-radiological assessment of 15 cases and review of the literature. Neurol India. (2022) 70:2093–9. doi: 10.4103/0028-3886.359201
39. Naik B, Bhalla A, Sharma N, Mokta J, Singh S, Gupta P, et al. Pituitary dysfunction in survivors of Russell’s viper snake bite envenomation: A prospective study. Neurol India. (2018) 66:1351–8. doi: 10.4103/0028-3886.241378
40. Golay V, Roychowdhary A, Pandey R, Pasari A, Praveen M, Arora P, et al. Growth retardation due to panhypopituitarism and central diabetes insipidus following Russell’s viper bite. Southeast Asian J Trop Med Public Health. (2013) 44:697–702.
41. Krishnan MN, Kumar S, and Ramamoorthy. Severe panhypopituitarism KP. and central diabetes insipidus following snake bite: unusual presentation as torsades de pointes. J Assoc Physicians India. (2001) 49:923–4.
42. Gupta UC, Garg OP, and Kataria ML. Cranial diabetes insipidus due to viper bite. J Assoc Physicians India. (1992) 40:686–7.
43. Castro-Amorim J, Oliveira A, Mukherjee AK, Ramos MJ, and Fernandes PA. Unraveling the reaction mechanism of Russell’s viper venom factor X activator: A paradigm for the reactivity of zinc metalloproteinases? J Chem Inf Model. (2023) 63:4056–69. doi: 10.1021/acs.jcim.2c01156
44. Kalita B, Mackessy SP, and Mukherjee AK. Proteomic analysis reveals geographic variation in venom composition of Russell’s Viper in the Indian subcontinent: implications for clinical manifestations post-envenomation and antivenom treatment. Expert Rev Proteomics. (2018) 15:837–49. doi: 10.1080/14789450.2018.1528150
45. Op Den Brouw B, Coimbra FCP, Casewell NR, Ali SA, Vonk FJ, and Fry BG. A genus-wide bioactivity analysis of daboia (Viperinae: Viperidae) viper venoms reveals widespread variation in haemotoxic properties. Int J Mol Sci. (2021) 22:13486. doi: 10.3390/ijms222413486
46. Srinivasan K, Nampoothiri M, Khandibharad S, Singh S, Nayak AG, and Hariharapura RC. Proteomic diversity of Russell’s viper venom: exploring PLA2 isoforms, pharmacological effects, and inhibitory approaches. Arch Toxicol. (2024) 98:3569–84. doi: 10.1007/s00204-024-03849-5
47. Alangode A, Reick M, and Reick M. Sodium oleate, arachidonate, and linoleate enhance fibrinogenolysis by Russell’s viper venom proteinases and inhibit FXIIIa; a role for phospholipase A2 in venom induced consumption coagulopathy. Toxicon. (2020) 186:83–93. doi: 10.1016/j.toxicon.2020.07.011
48. Jeevagan V, Katulanda P, Gnanathasan CA, and Warrell DA. Acute pituitary insufficiency and hypokalaemia following envenoming by Russell’s viper (Daboia russelii) in Sri Lanka: Exploring the pathophysiological mechanisms. Toxicon. (2013) 63:78–82. doi: 10.1016/j.toxicon.2012.11.012
49. Proby C, Tha-Aung, Thet-Win, Hla-Mon, Burrin JM, and Joplin GF. Immediate and long-term effects on hormone levels following bites by the Burmese Russell’s viper. Q J Med. (1990) 75:399–411.
50. Than-Than, Francis N, Tin-Nu-Swe, Myint-Lwin, Tun-Pe, Soe-Soe, et al. Contribution of focal haemorrhage and microvascular fibrin deposition to fatal envenoming by Russell’s viper (Vipera russelli siamensis) in Burma. Acta Trop. (1989) 46:23–38. doi: 10.1016/0001-706x(89)90013-2
51. Hart GR, Proby C, Dedhia G, Yeo TH, Joplin GF, and Burrin JM. Burmese Russell’s viper venom causes hormone release from rat pituitary cells in vitro. J Endocrinol. (1989) 122:489–94. doi: 10.1677/joe.0.1220489
52. Bhat S, Mukhopadhyay P, Raychaudhury A, Chowdhury S, and Ghosh S. Predictors of hypopituitarism due to vasculotoxic snake bite with acute kidney injury. Pituitary. (2019) 22:594–600. doi: 10.1007/s11102-019-00990-8
53. Sugimoto R, Kenzaka T, and Nishisaki H. Hyponatremia due to adrenal insufficiency after a Mamushi bite: a case report. BMC Endocr Disord. (2023) 23:219. doi: 10.1186/s12902-023-01466-4
54. el-Fiky MA. Hyperglycemic effect of a neurotoxic fraction (F3) from Naja haje venom: role of hypothalamo-pituitary adrenal axis (HPA). J Nat Toxins. (1999) 8:203–12.
55. Droctové L, Lancien M, Tran VL, Susset M, Jego B, Theodoro F, et al. A snake toxin as a theranostic agent for the type 2 vasopressin receptor. Theranostics. (2020) 10:11580. doi: 10.7150/thno.47485
56. Kobzina-Didukh DS. The influence of scorpion venom on the hypothalamo-pituitary-adrenal axis (review). Rep Vinnytsia Natl Med University. (2024) 28:524–9. doi: 10.31393/reports-vnmedical-2024-28(3)-24
57. Daachi F, Adi-Bessalem S, Megdad-Lamraoui A, and Laraba-Djebari F. Immune-toxicity effects of scorpion venom on the hypothalamic pituitary adrenal axis during rest and activity phases in a rodent model. Comp Biochem Physiol C Toxicol Pharmacol. (2020) 235:108787. doi: 10.1016/j.cbpc.2020.108787
58. Yamashita M, Oki Y, Iino K, Hayashi C, Matsushita F, Faje A, et al. The role of ether-a-go-go-related gene K+ channels in glucocorticoid inhibition of adrenocorticotropin release by rat pituitary cells. Regul Pept. (2009) 152:73–8. doi: 10.1016/j.regpep.2008.09.002
59. Hlubek M, Tian D, and Stuenkel EL. Mechanism of α-latrotoxin action at nerve endings of neurohypophysis. Brain Res. (2003) 992:30–42. doi: 10.1016/j.brainres.2003.08.025
60. Suzuki N, Hirono M, Kawahara K, and Yoshioka T. Sapecin B, a novel fly toxin, blocks macroscopic K+ currents in the GH3 rat pituitary cell line. Am J Physiol Cell Physiol. (1997) 273:289–96. doi: 10.1152/ajpcell.1997.273.1.C289
61. Florea A, El Hof FA, Hazi GM, and Oprea MC. Bee venom stimulates hormone secretion in rat somatotroph and corticotroph cells: digital image analysis of secretory granules. Microscopy Microanalysis. (2019) 25:1234–45. doi: 10.1017/S1431927619014910
62. Khalifa EI, Grawish SI, Gomaa AAAI, M.M GA, Metwally AAA, and Mahdy T. Influence of honey bee venom on growth performance of zaraibi kids from weaning to sexual puberty. EJAP. (2024) 61:83–92. doi: 10.21608/EJAP.2024.281834.1080
64. Monjotin N, Amiot MJ, Fleurentin J, Morel JM, and Raynal S. Clinical evidence of the benefits of phytonutrients in human healthcare. Nutrients. (2022) 14:1712. doi: 10.3390/nu14091712
65. Kristanc L and Kreft S. European medicinal and edible plants associated with subacute and chronic toxicity part I: Plants with carcinogenic, teratogenic and endocrine-disrupting effects. Food Chem Toxicol. (2016) 92:150–64. doi: 10.1016/j.fct.2016.04.007
66. Li H, Ge M, Lu B, Wang W, Fu Y, Jiao L, et al. Ginsenosides modulate hypothalamic–pituitary–adrenal function by inhibiting FKBP51 on glucocorticoid receptor to ameliorate depression in mice exposed to chronic unpredictable mild stress. Phytother Res. (2024) 38:5016–29. doi: 10.1002/ptr.8075
67. Mohan Gowda CM, Murugan SK, Bethapudi B, Purusothaman D, Mundkinajeddu D, and D’Souza P. Ocimum tenuiflorum extract (HOLIXERTM): Possible effects on hypothalamic-pituitary-adrenal (HPA) axis in modulating stress. PloS One. (2023) 18:e0285012. doi: 10.1371/journal.pone.0285012
68. Shams E, Zohrabi D, Omrani O, Sanati MH, Karimi-Dehkordi M, Yazdanpanahi N, et al. The effect of quercetin on obesity and reproduction through the expression of genes involved in the hypothalamus-pituitary-gonadal axis. J Bras Reprod Assist. (2025) 29:211–8. doi: 10.5935/1518-0557.20240097
69. Abedi H, Zarrin-Mehr A, Ebrahimi B, Haghshenas H, Parvin N, and Kargar Jahromi H. The effect of aqueous extract of orchid root on the structure of ovary and hypothalamic-pituitary-gonadal hormones in polycystic ovary syndrome rat model: An experimental study. Int J Reprod Biomed. (2024) 22:203–10. doi: 10.18502/ijrm.v22i3.16164
70. Oh DR, Yoo JS, Kim Y, Kang H, Lee H, Lm SJ, et al. Vaccinium bracteatum Leaf extract reverses chronic restraint stress-induced depression-like behavior in mice: Regulation of hypothalamic-pituitary-adrenal axis, serotonin turnover systems, and ERK/Akt phosphorylation. Front Pharmacol. (2018) 9:604. doi: 10.3389/fphar.2018.00604
71. Asiwe JN, Ekene EN, Agbugba LC, Moke EG, Akintade AV, Ben-Azu B, et al. Ginkgo biloba supplement abates lead-induced endothelial and testicular dysfunction in Wistar rats via up-regulation of Bcl-2 protein expression, pituitary-testicular hormones and down-regulation of oxido-inflammatory reactions. J Trace Elem Med Biol. (2023) 79:127216. doi: 10.1016/j.jtemb.2023.127216
72. Omotola MB, Adeosun IO, Obuotor EM, Akomolafe RO, and Ayannuga OA. Assessment of the effects of graded doses of polyphenolic-rich fraction of Garcinia kola seeds on pituitary–testicular axis of male Wistar rats. Dose-Response. (2017) 15:1559325817729260. doi: 10.1177/1559325817729260
73. Sourgens H, Winterhof H, Gumbinger HG, and Kemper FH. Antihormonal effects of plant extracts. TSH- and prolactin-suppressing properties of lithospermum officinale and other plants. Planta Med. (1982) 45:78–86. doi: 10.1055/s-2007-971251
74. Aleebrahim-Dehkordy E, Ansaripour S, Rafieian-Kopaei M, and Saberianpour S. Effects of substances on plants’ active compounds on changes in the hormone levels of the pituitary-thyroid axis in hyperthyroidism and hypothyroidism. Pharmacogn Rev. (2018) 12:1. doi: 10.4103/phrev.phrev_48_17
75. Benie T, Duval J, and Thieulant ML. Effects of some traditional plant extracts on rat oestrous cycle compared with clomid. Phytother Res. (2003) 17:748–55. doi: 10.1002/ptr.1206
76. Benie T and Thieulant ML. Mechanisms underlying antigonadotropic effects of some traditional plant extracts in pituitary cell culture. Phytomedicine. (2004) 11:157–64. doi: 10.1078/0944-7113-00326
77. Zhongyang L, Shengxiang W, Yuyun Q, Tian T, Baoyu Z, and Chenchen W. Swainsonine affects the secretion of reproductive hormones by inhibiting N-glycan processing. Acta Vet Zootec Sin. (2020) 51:3171–80.
78. Levavi-Sivan B, Hedvat R, Kanias T, Francis G, Becker K, and Kerem Z. Exposure of tilapia pituitary cells to saponins: Insight into their mechanism of action. Comp Biochem Physiol C Toxicol Pharmacol. (2005) 140:79–86. doi: 10.1016/j.cca.2005.01.002
79. Kasiri K, Heidari-Soureshjani S, and Pocock L. Medicinal plants effect on prolactin: A systematic review. WFM/MEJFM. (2017) 15. doi: 10.5742/MEWFM.2017.93164
80. Ushiroyama T, Sakuma K, Souen H, Nakai G, Morishima S, Yamashita Y, et al. Xiong-gui-tiao-xue-yin (Kyuki-chouketsu-in), a traditional herbal medicine, stimulates lactation with increase in secretion of prolactin but not oxytocin in the postpartum period. Am J Chin Med. (2007) 35:195–202. doi: 10.1142/S0192415X07004734
81. Wang X, Chen YG, Ma L, Li ZH, Li JY, Liu XG, et al. Effect of Chinese medical Herbs-Huiru Yizeng Yihao on hyperprolactinemia and hyperplasia of mammary gland in mice. Afr J Tradit Complement Altern Med. (2013) 10:24. doi: 10.4314/ajtcam.v10i4.5
82. Wang D, Wong HK, Bin FY, and Zhang ZJ. 18beta-Glycyrrhetinic acid induces apoptosis in pituitary adenoma cells via ROS/MAPKs-mediated pathway. J Neurooncol. (2014) 116:221–30. doi: 10.1007/s11060-013-1292-2
83. Wang D, Wong HK, Feng YB, and Zhang ZJ. Liquiritigenin exhibits antitumour action in pituitary adenoma cells via Ras/ERKs and ROS-dependent mitochondrial signalling pathways. J Pharm Pharmacol. (2014) 66:408–17. doi: 10.1111/jphp.12170
84. Margulis L. Biodiversity: molecular biological domains, symbiosis and kingdom origins. BioSystems. (1992) 27:39–51. doi: 10.1016/0303-2647(92)90045-z
85. Webster JI and Sternberg EM. Role of the hypothalamic-pituitary-adrenal axis, glucocorticoids and glucocorticoid receptors in toxic sequelae of exposure to bacterial and viral products. J Endocrinol. (2004) 181:207–21. doi: 10.1677/joe.0.1810207
86. Di Lorenzo F, Duda KA, Lanzetta R, Silipo A, De Castro C, and Molinaro A. A journey from structure to function of bacterial lipopolysaccharides. Chem Rev. (2022) 122:15767–821. doi: 10.1021/acs.chemrev.0c01321
87. Wang X and Quinn PJ. Endotoxins: Lipopolysaccharides of gram-negative bacteria. Subcell Biochem. (2010) 53:3–25. doi: 10.1007/978-90-481-9078-2_1
88. Tsukamoto H, Takeuchi S, Kubota K, Kobayashi Y, Kozakai S, Ukai I, et al. Lipopolysaccharide (LPS)-binding protein stimulates CD14-dependent Toll-like receptor 4 internalization and LPS-induced TBK1-IKKα-IRF3 axis activation. J Biol Chem. (2018) 293:10186–201. doi: 10.1074/jbc.M117.796631
89. Herman AP, Krawczyńska A, Bochenek J, Dobek E, Herman A, and Tomaszewska-Zaremba D. LPS-induced inflammation potentiates the IL-1 β -mediated reduction of LH secretion from the anterior pituitary explants. Clin Dev Immunol. (2013) 2013:926937. doi: 10.1155/2013/926937
90. Tomaszewska-Zaremba D, Haziak K, Tomczyk M, and Herman AP. Inflammation and LPS-binding protein enable the stimulatory effect of endotoxin on prolactin secretion in the ovine anterior pituitary: ex vivo study. Mediators Inflamm. (2018) 2018:5427089. doi: 10.1155/2018/5427089
91. Kuwahara-Otani S, Maeda S, Tanaka K, Hayakawa T, and Seki M. Systemic administration of lipopolysaccharide increases the expression of aquaporin-4 in the rat anterior pituitary gland. J Vet Med Sci. (2013) 75:1081–4. doi: 10.1292/jvms.13-0083
92. Zhou B, Xu Q, Guo J, Chen Q, Lv Q, Xiao K, et al. Necroptosis contributes to LPS-induced activation of the hypothalamic-pituitary-adrenal axis in a piglet model. Int J Mol Sci. (2022) 23:11218. doi: 10.3390/ijms231911218
93. Rodriguez TT, Biscarde EFS, Muniz RFDG, Amoedo MK, and Ramalho MJ. Prolactin secretion in hypothyroid endotoxemic rats: Involvement of L-arginine and nitric oxide synthase. Shock. (2005) 5:448–52. doi: 10.1097/01.shk.0000160938.08385.b8
94. Izvolskaia MS, Tillet Y, Sharova VS, Voronova SN, and Zakharova LA. Disruptions in the hypothalamic-pituitary-gonadal axis in rat offspring following prenatal maternal exposure to lipopolysaccharide. Stress. (2016) 19:198–205. doi: 10.3109/10253890.2016.1149695
95. Garcia C, Velez LM, Ujagar N, Del Mundo Z, Nguyen T, Fox C, et al. Lipopolysaccharide-induced chronic inflammation increases female serum gonadotropins and shifts the pituitary transcriptomic landscape. Front Endocrinol (Lausanne). (2023) 14:1279878. doi: 10.3389/fendo.2023.1279878
96. Sabatino ME, Sosa LDV, Petiti JP, Mukdsi JH, Mascanfroni ID, Pellizas CG, et al. Functional Toll-like receptor 4 expressed in lactotrophs mediates LPS-induced proliferation in experimental pituitary hyperplasia. Exp Cell Res. (2013) 319:3020–34. doi: 10.1016/j.yexcr.2013.08.012
97. Eijo G, Zárate S, Jaita G, Ferraris J, Magri ML, Zaldivar V, et al. Inhibition of nuclear factor-kappa b sensitises anterior pituitary cells to tumour necrosis factor-α- and lipopolysaccharide-induced apoptosis. J Neuroendocrinol. (2011) 8:651–9. doi: 10.1111/j.1365-2826.2011.02157.x
98. Tichomirowa M, Theodoropoulou M, Lohrer P, Schaaf L, Losa M, Uhl E, et al. Bacterial endotoxin (Lipopolysaccharide) stimulates interleukin-6 production and inhibits growth of pituitary tumour cells expressing the Toll-like receptor 4. J Neuroendocrinol. (2005) 17:152–60. doi: 10.1111/j.1365-2826.2005.01286.x
99. Lindberg P, Kenkel A, and Bühler K. Introduction to cyanobacteria. Adv Biochem Eng Biotechnol. (2023) 183:1–24. doi: 10.1007/10_2023_217
100. Pearson L, Mihali T, Moffitt M, Kellmann R, and Neilan B. On the chemistry, toxicology and genetics of the cyanobacterial toxins, microcystin, nodularin, saxitoxin and cylindrospermopsin. Mar Drugs. (2010) 8:1650. doi: 10.3390/md8051650
101. Dos Santos FCF, Lima GFC, Merlo E, Januario C de F, Miranda-Alves L, Miranda RA, et al. Single microcystin exposure impairs the hypothalamic-pituitary-gonadal axis at different levels in female rats. Mol Cell Endocrinol. (2024) 586:112203. doi: 10.1016/j.mce.2024.112203
102. Chen L, Shi T, Wang YT, He J, Zhao X, Wang YK, et al. Effects of acute exposure to microcystins on hypothalamic-pituitary-adrenal (HPA), -gonad (HPG) and -thyroid (HPT) axes of female rats. Sci Total Environ. (2021) 778:145196. doi: 10.1016/j.scitotenv.2021.145196
103. Shi T, Xu LL, Chen L, He J, Wang YK, Chen F, et al. Acute exposure to microcystins affects hypothalamic-pituitary axes of male rats. Environ pollut. (2023) 318:120843. doi: 10.1016/j.envpol.2022.120843
104. Chen Y, Xu J, Li Y, and Han X. Decline of sperm quality and testicular function in male mice during chronic low-dose exposure to microcystin-LR. Reprod Toxicol. (2011) 31:551–7. doi: 10.1016/j.reprotox.2011.02.006
105. Liu W, Chen C, Chen L, Wang L, Li J, Chen Y, et al. Sex-dependent effects of microcystin-LR on hypothalamic-pituitary-gonad axis and gametogenesis of adult zebrafish. Sci Rep. (2016) 6:22819. doi: 10.1038/srep22819
106. Zhang H, Wu J, Fang N, Zhang S, Su X, Jiang H, et al. Waterborne exposure to microcystin-leucine arginine induces endocrine disruption and gonadal dysplasia of Pelophylax nigromaculatus tadpoles via the hypothalamic-pituitary-gonadal-liver axis. Sci Total Environ. (2024) 906:167644. doi: 10.1016/j.scitotenv.2023.167644
107. Chen L, Wang Y, Giesy JP, Chen F, Shi T, Chen J, et al. Microcystin-LR affects the hypothalamic-pituitary-inter-renal (HPI) axis in early life stages (embryos and larvae) of zebrafish. Environ pollut. (2018) 241:540–8. doi: 10.1016/j.envpol.2018.05.024
108. Lin W, Guo H, Li Y, Wang L, Zhang D, Hou J, et al. Single and combined exposure of microcystin-LR and nitrite results in reproductive endocrine disruption via hypothalamic-pituitary-gonadal-liver axis. Chemosphere. (2018) 211:1137–46. doi: 10.1016/j.chemosphere.2018.08.049
109. Shurin G, Shanks N, Nelson L, Hoffman G, Huang L, and Kusnecov AW. Hypothalamic-pituitary-adrenal activation by the bacterial superantigen staphylococcal enterotoxin B: Role of macrophages and T cells. Neuroendocrinology. (1997) 65:18–28. doi: 10.1159/000127161
110. Castagliuolo I, Karalis K, Valenick L, Pasha A, Nikulasson S, Wlk M, et al. Endogenous corticosteroids modulate Clostridium difficile toxin A-induced enteritis in rats. Am J Physiol Gastrointest Liver Physiol. (2001) 280:539–45. doi: 10.1152/ajpgi.2001.280.4.G539
111. Mykoniatis A, Anton PM, Wlk M, Wang CC, Ungsunan L, Blüher S, et al. Leptin mediates Clostridium difficile toxin A-induced enteritis in mice. Gastroenterology. (2003) 124:683–91. doi: 10.1053/gast.2003.50101
112. Oken E, Kasper DL, Gleason RE, and Adler GK. Tetanus toxoid stimulation of the hypothalamic-pituitary-adrenal axis correlates inversely with the increase in tetanus toxoid antibody titers. J Clin Endocrinol Metab. (1998) 83:1691–6. doi: 10.1210/jcem.83.5.4829
113. Fu Q and Vankelecom H. Regenerative capacity of the adult pituitary: Multiple mechanisms of lactotrope restoration after transgenic ablation. Stem Cells Dev. (2012) 21:3245–57. doi: 10.1089/scd.2012.0290
114. Fu Q, Gremeaux L, Luque RM, Liekens D, Chen J, Buch T, et al. The adult pituitary shows stem/progenitor cell activation in response to injury and is capable of regeneration. Endocrinology. (2012) 153:3224–35. doi: 10.1210/en.2012-1152
115. Luque RM, Lin Q, Córdoba-Chacón J, Subbaiah PV, Buch T, Waisman A, et al. Metabolic impact of adult-onset, isolated, growth hormone deficiency (AOiGHD) due to destruction of pituitary somatotropes. PloS One. (2011) 6:e15767. doi: 10.1371/journal.pone.0015767
116. Sosa L del V, Picech F, Perez P, Gutierrez S, Leal RB, De Paul A, et al. Regulation of FGF2-induced proliferation by inhibitory GPCR in normal pituitary cells. Front Endocrinol (Lausanne). (2023) 14:1183151. doi: 10.3389/fendo.2023.1183151
117. Di Muro G, Mangili F, Esposito E, Barbieri AM, Catalano R, Treppiedi D, et al. A β-arrestin 2-biased dopamine receptor type 2 (DRD2) agonist is more efficacious than cabergoline in reducing cell proliferation in PRL-secreting but not in non-functioning pituitary tumor cells. Cancers (Basel). (2023) 15:3218. doi: 10.3390/cancers15123218
118. Günther T, Culler M, and Schulz S. Research resource: Real-time analysis of somatostatin and dopamine receptor signaling in pituitary cells using a fluorescence-based membrane potential assay. Mol Endocrinol. (2016) 30:479–90. doi: 10.1210/me.2015-1241
119. Ann JO and Conn PM. A cholera toxin-sensitive guanyl nucleotide binding protein mediates the movement of pituitary luteinizing hormone into a releasable pool: loss of this event is associated with the onset of homologous desensitization to gonadotropin-releasing hormone. Endocrinology. (1993) 132:2131–5. doi: 10.1210/endo.132.5.8386609
120. Klussmann E, Tamma G, Lorenz D, Wiesner B, Maric K, Hofmann F, et al. An inhibitory role of rho in the vasopressin-mediated translocation of aquaporin-2 into cell membranes of renal principal cells*. J Biol Chem. (2001) 276:20451–7. doi: 10.1074/jbc.M010270200
121. Dayanithi G, Forostyak O, Forostyak S, Kayano T, Ueta Y, and Verkhratsky A. Vasopressin and oxytocin in sensory neurones: expression, exocytotic release and regulation by lactation. Sci Rep. (2018) 8:13084. doi: 10.1038/s41598-018-31361-1
122. Roberson RW. Subcellular structure and behaviour in fungal hyphae. J Microsc. (2020) 280:75–85. doi: 10.1111/jmi.12945
123. Naranjo-Ortiz MA and Gabaldón T. Fungal evolution: diversity, taxonomy and phylogeny of the Fungi. Biol Rev Camb Philos Soc. (2019) 94:2101–37. doi: 10.1111/brv.12550
124. Meneely J, Greer B, Kolawole O, and Elliott C. T-2 and HT-2 toxins: toxicity, occurrence and analysis: A review. Toxins (Basel). (2023) 15:481. doi: 10.3390/toxins15080481
125. Pitt JI and David Miller J. A concise history of mycotoxin research. J Agric Food Chem. (2017) 65:7021–33. doi: 10.1021/acs.jafc.6b04494
126. Carballo D, Moltó JC, Berrada H, and Ferrer E. Presence of mycotoxins in ready-to-eat food and subsequent risk assessment. Food Chem Toxicol. (2018) 121:558–65. doi: 10.1016/j.fct.2018.09.054
127. Yang X, Zhang X, Yao Q, Song M, Han Y, Shao B, et al. T-2 toxin impairs male fertility by disrupting hypothalamic-pituitary-testis axis and declining testicular function in mice. Chemosphere. (2019) 234:909–16. doi: 10.1016/j.chemosphere.2019.06.145
128. Yang R, Wang YM, Zhang L, Zhao ZM, Zhao J, and Peng SQ. Prepubertal exposure to T-2 toxin advances pubertal onset and development in female rats via promoting the onset of hypothalamic-pituitary-gonadal axis function. Hum Exp Toxicol. (2016) 12:1276–85. doi: 10.1177/0960327116629529
129. Liu X, Guo P, Liu A, Wu Q, Xue X, Dai M, et al. Nitric oxide (NO)-mediated mitochondrial damage plays a critical role in T-2 toxin-induced apoptosis and growth hormone deficiency in rat anterior pituitary GH3 cells. Food Chem Toxicol. (2017) 102:11–23. doi: 10.1016/j.fct.2017.01.017
130. Velardez MO, Poliandri AH, Cabilla JP, Bodo CCA, Machiavelli LI, and Duvilanski BH. Long-term treatment of anterior pituitary cells with nitric oxide induces programmed cell death. Endocrinology. (2004) 145:2064–70. doi: 10.1210/en.2003-1263
131. Caride A, Lafuente A, and Cabaleiro T. Endosulfan effects on pituitary hormone and both nitrosative and oxidative stress in pubertal male rats. Toxicol Lett. (2010) 197:106–12. doi: 10.1016/j.toxlet.2010.05.006
132. Guo P, Liu A, Huang D, Wu Q, Fatima Z, Tao Y, et al. Brain damage and neurological symptoms induced by T-2 toxin in rat brain. Toxicol Lett. (2018) 286:96–107. doi: 10.1016/j.toxlet.2018.01.012
133. Muthulakshmi S, Hamideh PF, Habibi HR, Maharajan K, Kadirvelu K, and Mudili V. Mycotoxin zearalenone induced gonadal impairment and altered gene expression in the hypothalamic–pituitary–gonadal axis of adult female zebrafish (Danio rerio). J Appl Toxicol. (2018) 38:1388–97. doi: 10.1002/jat.3652
134. Wu TS, Lin YT, Huang YT, Yu FY, and Liu BH. Ochratoxin A triggered intracerebral hemorrhage in embryonic zebrafish: Involvement of microRNA-731 and prolactin receptor. Chemosphere. (2020) 242:125143. doi: 10.1016/j.chemosphere.2019.125143
135. Qin X, Li X, Guo J, Zhou M, Xu Q, Lv Q, et al. Necroptosis contributes to deoxynivalenol-induced activation of the hypothalamic–pituitary–adrenal axis in a piglet model. Int Immunopharmacol. (2024) 143:113541. doi: 10.1016/j.intimp.2024.113541
136. Wu S, Wang Q, Duan H, Xiong J, Lai T, Wang C, et al. Mushroom poisoning deaths and prevention practices in Hunan, 2014-2023. PloS One. (2025) 20:e0326107. doi: 10.1371/journal.pone.0326107
137. Kelner MJ and Alexander NM. Endocrine hormone abnormalities in amanita poisoning. Clin Toxicol. (1987) 25:21–37. doi: 10.3109/15563658708992611
138. De Carlo E, Milanesi A, Martini C, Maffei P, Tamagno G, Parnigotto PP, et al. Effects of Amanita phalloides toxins on insulin release: In vivo and in vitro studies. Arch Toxicol. (2003) 77:441–5. doi: 10.1007/s00204-003-0475-6
139. Sepehri H, Renard C, and Houdebine LM. β-glucan and pectin derivatives stimulate prolactin secretion from hypophysis in vitro. Proc Soc Exp Biol Med. (1990) 94:193–7. doi: 10.3181/00379727-194-43077
140. Schoenle A, Francis O, Archibald JM, Burki F, de Vries J, Dumack K, et al. Protist genomics: key to understanding eukaryotic evolution. Trends Genet. (2025) 41(10):868–82. doi: 10.1016/j.tig.2025.05.004
141. Litaker RW, Vandersea MW, Faust MA, Kibler SR, Nau AW, Holland WC, et al. Global distribution of ciguatera causing dinoflagellates in the genus Gambierdiscus. Toxicon. (2010) 56:711–30. doi: 10.1016/j.toxicon.2010.05.017
142. Binzer SB, Svenssen DK, Daugbjerg N, Alves-de-Souza C, Pinto E, Hansen PJ, et al. A-, B- and C-type prymnesins are clade specific compounds and chemotaxonomic markers in Prymnesium parvum. Harmful Algae. (2019) 81:10–7. doi: 10.1016/j.hal.2018.11.010
143. Nishimura T, Sato S, Tawong W, Sakanari H, Uehara K, Shah MMR, et al. Genetic diversity and distribution of the ciguatera-causing dinoflagellate gambierdiscus spp. (Dinophyceae) in coastal areas of Japan. PloS One. (2013) 8:e60882. doi: 10.1371/journal.pone.0060882
144. Sortino MA, Delahunty TM, Yasumoto T, and Cronin MJ. Maitotoxin increases inositol phosphates in rat anterior pituitary cells. J Mol Endocrinol. (1991) 6:95–9. doi: 10.1677/jme.0.0060095
145. Schettini G, Koike K, and Login IS. Maitotoxin stimulates hormonal release and calcium flux in rat anterior pituitary cells in vitro. Am J Physiol Endocrinol Metab. (1984) 10:520–5. doi: 10.1152/ajpendo.1984
146. Koike K, Judd AM, Login IS, Yasumoto T, and MacLeod RM. Maitotoxin, a calcium channel activator, increases prolactin release from rat pituitary tumor 7315a cells by a mechanism that may involve leukotriene production. Neuroendocrinology. (1986) 43:283–90. doi: 10.1159/000124542
147. Fitzpatrick LA, Yasumoto T, and Aurbach GD. Inhibition of parathyroid hormone release by maitotoxin, a calcium channel activator. Endocrinology. (1989) 124:97–103. doi: 10.1210/endo-124-1-97
148. Kolderup J, Meldahl A -S, Eriksen S, Haug E, Fonnum F, and Sand O. Toxin of the marine alga Prymnesium patelliferum enhances voltage dependent Ca2+-currents, elevates the cytosolic Ca2+-concentration and facilitates hormone release in clonal rat pituitary cells. Acta Physiol Scand. (1995) 154:321–7. doi: 10.1111/j.1748-1716.1995.tb09916.x
149. Libonati RMF, de Mendonça BB, Maués JA, Quaresma JAS, and de Souza JM. Some aspects of the behavior of the hypothalamus-pituitary-adrenal axis in patients with uncomplicated Plasmodium falciparum malaria: Cortisol and dehydroepiandrosterone levels. Acta Trop. (2006) 98:270–6. doi: 10.1016/j.actatropica.2006.05.008
150. Davis TM, Supanaranond W, Pukrittayakamee S, Krishna S, Hart GR, Burrin JM, et al. The pituitary-thyroid axis in severe falciparum malaria: evidence for depressed thyrotroph and thyroid gland function. Trans R Soc Trop Med Hyg. (1990) 84:330–5. doi: 10.1016/0035-9203(90)90305-x
151. Kelly TR, Cannon AL, Stansberry KR, Kimball MG, and Lattin CR. Changes in hypothalamic–pituitary–adrenal axis function, immunity, and glucose during acute Plasmodium relictum infection in house sparrows (Passer domesticus). Gen Comp Endocrinol. (2024) 345:114388. doi: 10.1016/j.ygcen.2023.114388
152. Maina CI. Plasma ACTH concentration and pituitary gland histo-pathology in rats infected with Trypanosoma brucei brucei. Afr Health Sci. (2017) 17:1029. doi: 10.4314/ahs.v17i4.10
153. Petzke F, Heppner C, Mbulamberi D, Winkelmann W, Chrousos GP, Allolio B, et al. Hypogonadism in Rhodesian sleeping sickness: evidence for acute and chronic dysfunction of the hypothalamic-pituitary-gonadal axis. Fertil Steril. (1996) 65:68–75. doi: 10.1016/S0015-0282(16)58029-7
154. al-Qarawi AA, Abdel-Rahman H, and Elmougy SA. Impairment in the pituitary-thyroid axis of the Camelus dromedarius infected with Trypanosoma evansi. Dtsch Tierarztl Wochenschr. (2001) 108:172–4.
155. Mutayoba BM, Eckersall PD, Seely C, Gray CE, Cestnik V, Jeffcoate IA, et al. Effects of Trypanosoma congolense on pituitary and adrenocortical function in sheep: responses to exogenous corticotrophin-releasing hormone. Res Vet Sci. (1995) 58:180–5. doi: 10.1016/0034-5288(95)90074-8
156. Anyogu DC, Shoyinka SVO, and Ihedioha JI. Brain and pituitary-adrenal lesions of Trypanosoma brucei brucei and Trypanosoma congolense infections in the West African Dwarf rams: Is trypanotolerance overrated? Vet Pathol. (2022) 59:773–81. doi: 10.1177/03009858221100432
157. Abebe G, Shaw MK, and Eley RM. Trypanosoma congolense in the microvasculature of the pituitary gland of experimentally infected Boran cattle (Bos indicus). Vet Pathol. (1993) 30:401–9. doi: 10.1177/030098589303000501
158. Nourollahpour Shiadeh M, Niyyati M, Fallahi S, and Rostami A. Human parasitic protozoan infection to infertility: a systematic review. Parasitol Res. (2016) 115:469–77. doi: 10.1007/s00436-015-4827-y
159. Roberts CW, Walker W, and Alexander J. Sex-associated hormones and immunity to protozoan parasites. Clin Microbiol Rev. (2001) 14:476–88. doi: 10.1128/CMR.14.3.476-488.2001
160. Melo GD, Goyard S, Fiette L, Boissonnas A, Combadiere C, MaChado GF, et al. Unveiling Cerebral Leishmaniasis: parasites and brain inflammation in Leishmania donovani infected mice. Sci Rep. (2017) 7:8454. doi: 10.1038/s41598-017-09085-5
161. Schlüter D and Barragan A. Advances and challenges in understanding cerebral toxoplasmosis. Front Immunol. (2019) 10:242. doi: 10.3389/fimmu.2019.00242
Keywords: pituitary gland, poison, toxin, venom, nature
Citation: Sanchis-Pascual D, Cámara Gómez R, Morillas-Amat P, Lara-Gálvez D, Pérez-Cervantes V, Masdeu-López-Cerón P and Merino-Torres JF (2025) Biological poisons targeting the pituitary gland: insights across the five kingdoms. Front. Endocrinol. 16:1708792. doi: 10.3389/fendo.2025.1708792
Received: 19 September 2025; Accepted: 29 October 2025;
Published: 13 November 2025.
Edited by:
Masaaki Yamamoto, Cedars-Sinai, United StatesReviewed by:
Pedro Iglesias, Puerta de Hierro University Hospital Majadahonda, SpainŞeyma Erdem Torun, Yuksek Ihtisas Training and Research Hospital, Türkiye
Copyright © 2025 Sanchis-Pascual, Cámara Gómez, Morillas-Amat, Lara-Gálvez, Pérez-Cervantes, Masdeu-López-Cerón and Merino-Torres. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Sanchis-Pascual, ZGF2aWQuc2FuY2hpcy5wYXNjdWFsQGhvdG1haWwuY29t