 Zara Raza1,2,3,†
Zara Raza1,2,3,† Syeda F. Hussain1,2,†
Syeda F. Hussain1,2,† Victoria S. Foster1,2,4,†Joseph Wall3,5Peter J. Coffey6John F. Martin7
Victoria S. Foster1,2,4,†Joseph Wall3,5Peter J. Coffey6John F. Martin7 Renata S. M. Gomes1,2,8*
Renata S. M. Gomes1,2,8*
- 1Research & Innovation, Blind Veterans UK, London, United Kingdom
- 2BRAVO VICTOR, Research & Innovation, London, United Kingdom
- 3Hull York Medical School, University of York, York, United Kingdom
- 4St George’s Hospital Medical School, London, United Kingdom
- 5Haxby Group Hull, General Practice Surgery, Hull, United Kingdom
- 6Development, Ageing and Disease, UCL Institute of Ophthalmology, University College London, London, United Kingdom
- 7Centre for Cardiovascular Biology and Medicine, University College London, London, United Kingdom
- 8Northern Hub for Veterans and Military Families Research, Department of Nursing, Midwifery and Health, Faculty of Health and Life Sciences, Northumbria University, Newcastle upon Tyne, United Kingdom
War and conflict are global phenomena, identified as stress-inducing triggers for epigenetic modifications. In this state-of-the-science narrative review based on systematic principles, we summarise existing data to explore the outcomes of these exposures especially in veterans and show that they may result in an increased likelihood of developing gastrointestinal, auditory, metabolic and circadian issues, as well as post-traumatic stress disorder (PTSD). We also note that, despite a potential “healthy soldier effect”, both veterans and civilians with PTSD exhibit the altered DNA methylation status in hypothalamic–pituitary–adrenal (HPA) axis regulatory genes such as NR3C1. Genes associated with sleep (PAX8; LHX1) are seen to be differentially methylated in veterans. A limited number of studies also revealed hereditary effects of war exposure across groups: decreased cortisol levels and a heightened (sex-linked) mortality risk in offspring. Future large-scale studies further identifying the heritable risks of war, as well as any potential differences between military and civilian populations, would be valuable to inform future healthcare directives.
Background
The epigenome, a collaborative effort of DNA and its modifications, can be readily modified in response to internal and external changes (1–9). Epigenetics may impact the individual, for example, work-related stress and depression result in altered DNA methylation of the glucocorticoid receptor (GR) gene (NR3C1; nuclear receptor subfamily 3 group C member 1), an area well reviewed by Bakusic et al. (10). Epigenetic modifications may be inherited by progeny, and even in the offspring of progeny. Such inheritance is termed “intergenerational”, except in instances where the generation could never have been exposed to stress stimuli (e.g., the grandparent was not pregnant at the time), in which case it is “transgenerational” (11). Non-genetic influence includes the impact of experiences from previous generations, passed down through shared stories or the spoken word, an effect referred to as “intergenerational trauma,” which may contribute to PTSD (12–14).
Our review discusses epigenetic impacts and phenotypic outcomes on individuals and families affected by conflict and war, summarised in Figure 1. Exploring whether military populations differ epigenetically in response to conflict when compared to civilians is important. First, as the rigorous military selection process generates a population of highly fit and healthy individuals compared to general society. Second, both military and civilian populations may witness and experience the long-term physical, psychological and economic impacts of war, but it is, in most instances, only military personnel who are expected to actively participate in planned, targeted assault. Therefore, these differential impacts may result in different health outcomes where a one-size-fits-all treatment approach will be ineffective. Current research demonstrates the long-reaching impacts of stressors, such as famine on metabolic, cardiovascular and cerebral health (15). The current global issues, including the war in Ukraine, civil unrest in Iran, upheaval in Afghanistan, conflict in Ethiopia, South Sudan, and Syria (and undoubtably numerous more yet less widely publicised unrest), result in a drive to take in refugees. One review has noted primary healthcare as the prevalent support type used by refugees (98.7%) and found to be the most useful (27.5%) (16). This highlights the necessity to understand epigenetic impacts that may further contribute to, or drive, psychological and/or physiological complications to improve support for vulnerable populations.
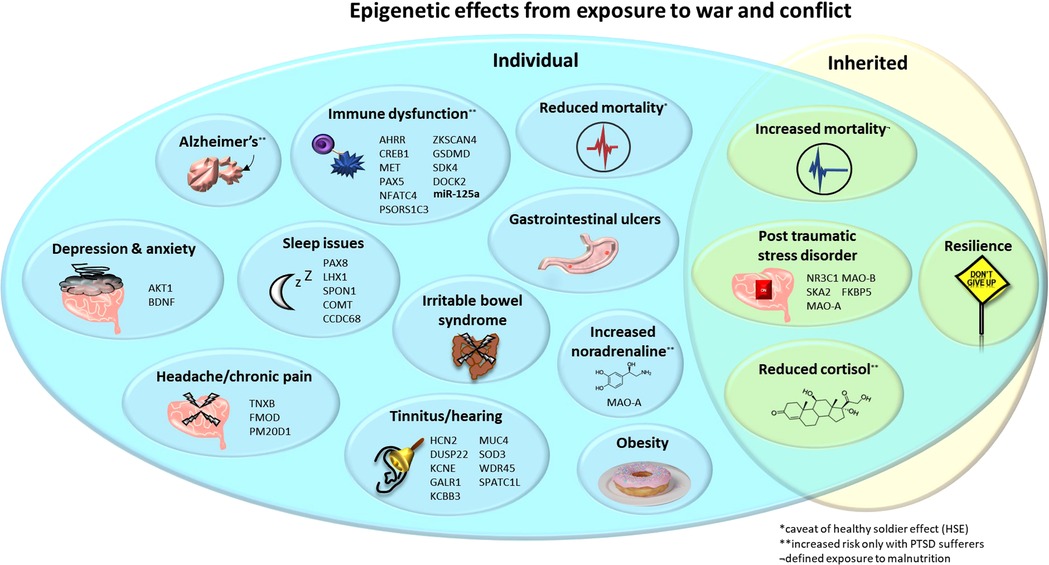
Figure 1. Epigenetic effects from exposure to war and conflict. Upon the individual: decreased cortisol, mortality* and sleep quality/quantity; increased mortality, PTSD, noradrenaline**, tinnitus, immune dysfunction, Alzheimer's disease**, obesity, anxiety, depression, gastrointestinal ulcers and IBS in children exposed to war. Inherited: decreased cortisol and mortality*, and increased risk of PTSD but also resilience. Noted are genes (and the microRNA miR-125a where expression is altered) that are associated with these pathologies and may occur differentially methylated and/or expressed. *Decreased mortality, potentially as a confounding “healthy solider effect”. **Increased risk only in individuals with PTSD. IBS, irritable bowel syndrome; PTSD, post-traumatic stress disorder.
Post-Traumatic Stress Disorder
Hypothalamic–pituitary–adrenal (HPA) axis dysfunction is one of the aetiological factors linked with PTSD (17, 18). The GR, responding to cortisol, is a key player in this axis (19). Increases in cortisol are often associated with higher levels of stress. Paradoxically, patients with chronic PTSD may exhibit lower cortisol, and lowered urinary cortisol after a traumatic event may be predictive of PTSD development (18, 20). It is important to note that cortisol levels are not pathologically low, and still follow a circadian rhythm, but are significantly lower compared to individuals without PTSD (18, 21). In PTSD, a spiking of cortisol levels may also be seen in acute therapy stages, and the administration of a low dose of cortisol may hold promise for alleviation (18, 22). Indeed, administration of dexamethasone (to suppress cortisol) in veterans resulted in a positive correlation between suppression and the severity of PTSD symptoms (18, 23). Yehuda et al. summarise from their research that reduced cortisol may occur due to enhanced negative feedback sensitivity of the HPA axis, i.e., cortisol suppression (18). As an increased number of GRs are seen in individuals with PTSD, the physiological response may still be high despite these lowered levels of cortisol (24).
Altered methylation may result in reduced (or increased) gene expression and, in genes related to the HPA axis, influence the suppression of the sympathetic nervous system. Individuals who have experienced emotional and/or sexual trauma have been reported to show both increased methylation at the promoter region of the NR3C1 (the GR gene) and reduced messenger RNA (mRNA) levels of the receptor (25, 26). Conversely, another study reported that individuals with lifetime PTSD showed reduced methylation of NR3C1 at promoter regions and an inverse correlation with overall mRNA expression, which, in turn, inversely correlated with cortisol levels (27). These data reveal the complexity of the HPA axis effects from epigenetic changes; given there are different promoter regions of the gene, this may play a role in differential results. FK506 binding protein 5 (FKBP5) is a negative regulator of GR via the signalling and translocation of glucocorticoids to modulate sensitivity (28) and is also associated with both genetic and environmental modifications (29, 30). Decreased methylation of FKBP5 (predicating increased expression) was found in individuals exposed to early trauma and associated with the GR-induced transcription of FKBP5, resulting in chronic dysregulation of stress hormones (30). Epigenetic marks, i.e., methylation of FKBP5 and/or NR3C1, have been proposed as a possible predictor for the severity of PTSD as well as to define treatment outcome (31–33).
Other genes have also been linked to PTSD symptoms (26). A sex-linked effect for higher levels of circulating peptide [pituitary adenylate cyclase-activating polypeptide (PACAP)], which binds to the pituitary adenylate cyclase-activating polypeptide type I receptor (ADCYAP1R1; PAC-1), was seen for the female symptoms/diagnosis of PTSD (34). Increased methylation of a CpG island of PAC-1 was predictive of PTSD, in a non-sex-specific manner (34) and found in male veterans with PTSD (35). This suggests functionally relevant epigenetic effects may occur as a result of PTSD exposure. Multiple microRNAs (miRNAs) have been shown to play a significant role in the regulation of fear, with differential expression observed in individuals with PTSD (36).
Understanding the role of epigenetics in the pathogenesis of PTSD has allowed for research to be conducted into the therapeutic uses of these epigenetic modifications. A common focus of PTSD treatment is the extinction of memories that cause fear (37). Studies on mice have revealed histone acetylation, methylation, and DNA methylation all contribute to memory extinction (38). Valproic acid (VPA), a histone deacetylate (HDAC) inhibitor, enhances extinction memory for fear associated with audio (39). Murphy and Singewald suggest increasing the occurrence of acetylation promotes de novo transcription, translation and gene expression to allow for the consolidation of long-term extinction memory (36). The use of VPA, an established mood stabiliser for bipolar disorders, has been suggested for the treatment of PTSD alongside psychotherapy (40, 41). Potential treatments, underpinned by epigenetic changes, indicate the importance of understanding these underlying modifications to better aid patients in future.
To summarise, a number of players (e.g., FKBP5, NR3C1, PACAP, PAC-1, and HDAC) can be linked to the pathophysiology of PTSD, likely due to influences on the HPA axis. Next, we will discuss how the trauma of war may evoke these symptoms.
War and military personnel
Trauma, the exposure to a distressing or disturbing experience, can have long-term negative impacts on individuals that may lead to PTSD (a common diagnosis made in the aftermath of war and conflict) (17, 42). As PTSD-related epigenetic modifications may impact the longevity of individuals, we first evaluate the normal longevity seen in military personnel.
Hartal et al. compared the life expectancy of retired military personnel to the general population and found the mean age at death was greater in comparison to their sex- and birth-matched equivalents in the civilian population. The average life expectancy was found to be exceeded by 67.9% of retired service members (43). The authors attributed many of these results to the “healthy soldier effect” (HSE), which persists even 30 years after service (44), a variation of the “healthy worker effect” (45). This is a tendency for people who work in physically demanding environments—particularly the military—to exhibit lower mortality and morbidity rates relative to the general population, due to the exclusion of those with poor health from participating (46). Soldiers in the US who are wounded in battle receive a Purple Heart (PH) decoration. In veterans aged 65 years and over, PH recipients with or without PTSD showed halved rates of mortality in comparison to those without the award (47). This could be attributed to a “survivor effect,” where the less healthy in this cohort of injured veterans did not survive to this timepoint. Alternatively, veterans who have been injured during combat may receive priority medical care. The epigenetic impact of stressors that military personnel experience may be influenced by these factors.
Verhoeven et al. found the epigenetic age of veterans with PTSD, defined by the methylation of leukocyte DNA, was significantly lower than those without (48). This was partly explained through use of antidepressants, which was associated with reducing telomerase activity and, in turn, genetic aging. However, veterans with PTSD, versus those who have been exposed to trauma but did not have PTSD, have been shown to exhibit the following: (1) increased plasma lipids (49); (2) an exacerbated startle response upon exposure to yohimbine, a noradrenaline inhibitor antagonist (50); and (3) a heightened experience of odour intensity with decreased heart rate in response to burning rubber (a trauma cue) (51).
Therefore, despite a proposed increase in longevity among veterans, the physiological impacts of PTSD persist. We next consider how the epigenetic factors specifically may occur after exposure to conflict.
Epigenetic impacts of war
Exposure to war results in a plethora of biological impacts, described below and summarised in Figure 1. The physiological effects of war trauma seen in veterans with PTSD include co-morbid chronic pain (which may reduce alongside PTSD treatment) (52, 53). These findings were applicable to both male and female veterans (53), although the lack of data pertaining to female veterans often makes sex-specific effects difficult to explore. Other pathologies incurred from war include a higher incidence of peptic ulcers in veterans and increases in dyspepsia and heartburn (2). Australian combat prisoners of war (POWs) in World War II (WWII) Japanese camps exhibited an increased rate of gastrointestinal ulcers (1). Children exposed to war exhibit an increased likelihood of irritable bowel syndrome (IBS), a condition also more common in veteran populations (4). An increased risk of obesity was observed in male veterans but, when adjusted for, were no more likely to exhibit hypertension or diabetes (7).
One group found no change in POW medical mortality (54) but, later, found a clear increase at 5–14 years after WWII, which reduced after this point. This indicates an individual's age at the time of war ending may influence mortality (55). Costa et al. also highlighted an age-related effect; after 30, the age at which a POW became imprisoned resulted in a range of differing effects during ageing. Survivors with the worst conditions showed reduced mortality versus non-POWs (or POWs when conditions were not so poor) 35 years post-imprisonment (56). An established increased risk of mortality through ischemic heart disease was found to be even more prevalent after 75 years of age (57, 58). POWs aged younger than 30 years showed an increased risk of cardio- and cerebrovascular disease as well as morbidity from the former (56). As an increased risk of cardiovascular disease is also seen in those prenatally exposed to famine, the malnutrition experienced by POWs may therefore convolute specific impacts of war (e.g., combat exposure) (59, 60). During WWII, the genocide of six million European Jews and other ethnic and societal minorities occurred. Studies on individuals exposed to the Shoah provide information on those experiencing the effects of war through torture, and genocide, but also severe famine. Despite these impacts, small-scale Holocaust studies did not reveal an increased risk of mortality at two timepoints [20–41 (61) and 40–50 (62) years after WWII].
As with other civilian populations, veterans (as well as those exposed to the Holocaust 63), despite often showing increased resilience, are at risk of mental health conditions such as depression and anxiety, which negatively impact on quality of life (64–69). A reduced expression of brain-derived neurotrophic factor (BDNF) is exhibited in depression (70) and has been found to be dysregulated in PTSD/traumatic brain injury (TBI) in veterans (71). In addition, one study revealed the increased methylation of the BDNF promoter region is associated with a PTSD diagnosis in Vietnam War veterans (72). Furthermore, older veterans with PTSD exhibit a twofold increase in the risk for dementia (73, 74). Veterans showed increased levels of depression 40 years on compared to non-POWs; however, in this study, there were no differences in alcohol issues or anxiety (54).
As mentioned above regarding civilian groups with PTSD, lowered cortisol has also been shown in combat veterans and Holocaust survivors with PTSD (75–77). The hormone and neurotransmitter noradrenaline, the mediator of the HPA-stress system, was present at the baseline in higher levels in the cerebrospinal fluid (CSF) from male combat veterans with PTSD, an effect not replicated in blood plasma (78). Blood plasma again was reported by Yahyavi et al. to show normal levels of adrenaline and noradrenaline in the plasma of veterans with PTSD (and their offspring) but decreased cortisol in those with PTSD (79). A study using CSF samples from 52 veterans with trauma revealed no significant difference in noradrenaline levels between those with or without PTSD/trauma when taking the medication prazosin (which treats high blood pressure and heart failure). However, in those not taking the drug, a significantly positive correlation between behavioural symptoms (e.g., depression and insomnia) and higher noradrenaline levels was revealed (80). The authors suggest trauma exposure may influence responsiveness to noradrenaline, and these data reveal other medication may influence hormone levels.
Above, we have discussed the physiological ramifications of war and conflict exposure, such as gastrointestinal upset (included in Figure 1). We will further expand upon this in the next section while specifically discussing epigenetic alterations (summarised in Table 1), predominantly pertaining to gene methylation.
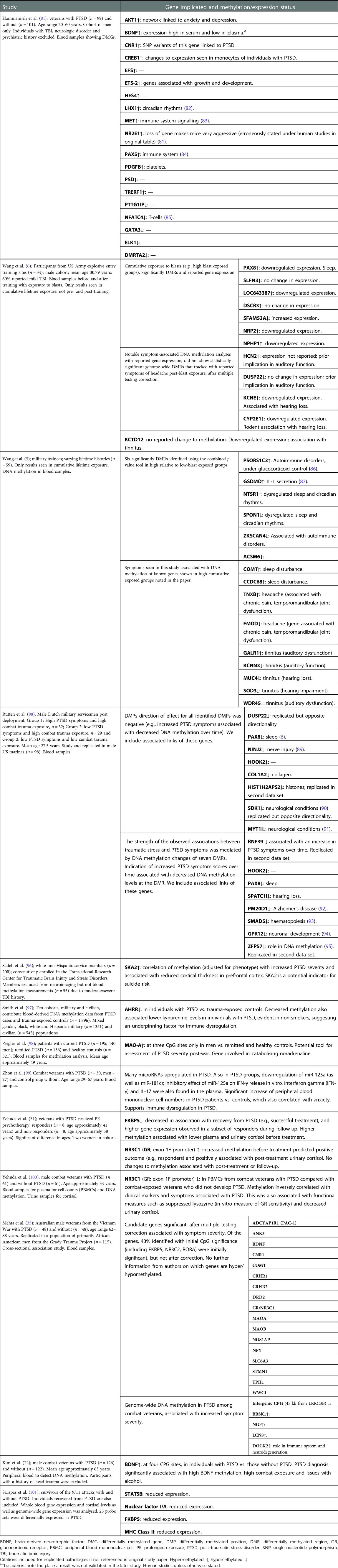
Table 1. Exposure to war and subsequent effects on epigenetics to individuals measured through gene methylation.
Monoamine oxidase A (MAO-A) is an enzyme critically involved in catalysing adrenaline, noradrenaline, dopamine and serotonin (102). Hypermethylation of the MAO-A gene is linked to PTSD (35) and was suggested as a tool to assess the severity of the disorder in male patients with PTSD due to war exposure; a sex-linked effect was seen, although the female sample size was comparatively small (see Table 1) (98). Gene hypermethylation often results in reduced expression, in this case potentially impeding catabolic activity on noradrenaline resulting in increased circulating levels upon stress exposure. Such dysregulation of stress hormones is a hallmark of PTSD.
In combat veteran studies, those with PTSD exhibited NR3C1 hypermethylation when compared to those without PTSD, an effect not seen in those exposed to childhood trauma (31, 35, 100). The impact of the Tutsi genocide in Rwanda was studied in 25 pregnant women and found higher CpG methylation in the nuclear receptor subfamily 3 group C member 2 (NR3C2; a mineralocorticoid receptor, also binds glucocorticoids) gene (103). The increased methylation of spindle and kinetochore-associated protein 2 (SKA2) plays a role in GR chaperoning, providing negative feedback to the HPA by removing the receptor from the cytoplasm and to the nucleus (104). Blood methylation of SKA2 and reduced expression, along with aberrant glucocorticoid signalling, was identified as a potential indicator for suicide risk (105, 106). In veterans, a positive correlation of SKA2 methylation in the blood was seen with severity of PTSD symptoms (96).
Sleep difficulties are often reported by veterans (8), including those with PTSD (107), making these epigenetic changes an area of interest. Indeed, an increase in methylation on the circadian clock gene, LHX1, has been seen in veterans with PTSD (81). Impacts on sleep-related genes were found in military personnel with chronic blast exposure, including differential methylation of NTSR1 and SPON1 (5), and differentially methylated regions (DMRs) for CCDC68 and COMT (also linked to sleep difficulties experienced by the individuals). An earlier study by the same group found, concurrent with impacts on sleep, chronic exposure to blasts resulted in the increased methylation of the paired box gene 8 (PAX8) antisense transcript, related to repressed gene expression (6). However, another study found decreased PAX8 methylation in veterans with PTSD (88). PAX8 is associated with thyroid function (108) as well as sleep duration (109). Sleep after exposure to emotional information was shown to consolidate memory up to 4 years later, so initial sleep disruption may be beneficial to reduce PTSD severity (110). However, this is a short-term benefit, given that persistently poor sleep impedes mental health (111). To alleviate PTSD, attempts to restore effective sleep should be undertaken; gene methylation may offer a useful marker of success.
Chronic blast exposure also revealed the differential methylation of auditory function genes (KCNN3, SOD3, MUC3, GALR1 and WDR45B) which were associated with tinnitus, and the DMRs within FMOD and TNXB were linked to headache and pain (5). Symptoms of tinnitus in low versus high blast exposure groups were also linked to DMRs in auditory genes KCNE1 and CYP2E1, showing an inverse correlation with methylation and expression (6). Finally, pre- versus post-blast exposure showed at least a 1.5-fold difference in expression in 67 genes, such as UFC1 and YOD1 (ubiquitin related proteins; others have been linked to TBI). The dysregulation of cytokine and chemokines was also exhibited in MCP-1, GCSF, HGF, MCSF, and RANTES after acute exposure to blasts (6). Blast exposure can result in neurological impacts, such as brain swelling and neurotrauma; these epigenetic markers are evidence of both the psychological and physical impacts of war (5, 6).
PTSD has been associated with increased immunological inflammation (112). A number of immunological changes were characterised in a study on combat veterans with PTSD (99). First, there was a significant increase in the numbers of (1) T-helper 1 (Th1; CD4+), (2) Th17, (3) cytotoxic T (CD8+), and (4) B cells. It was found that an increase in Th1 percentage was correlated with severity of PTSD score, as well as a reduction in the number of T-regulatory cells. These data suggest a heightened inflammatory response with reduced regulation. Reflecting this, upregulated levels of the proinflammatory cytokines interferon gamma (IFN-γ) and IL-17 were seen in the plasma. Zhou et al. then revealed that these changes were associated with the downregulation of the miRNA, miR-125a (as well as miR-181c), and went on to show the inhibitory effect of miR-125a on IFN-γ release in vitro (99). This signalling molecule has been linked to the upregulation of another proinflammatory cytokine, NF-κB, in human cancer cell lines (113). In rats, NF-κB is necessary for memory reconsolidation (e.g., to allow for alterations of negative associations) (114, 115). This suggests that the cytokine cascade in PTSD is dysregulated. Regular amounts of miR-125a would control the inflammatory response; by reducing IFN-γ and increasing NF-κB, a reduction of trauma responses may occur.
Further exploring the immunological impacts, the largest epigenetics study to date was performed on mixed veteran and civilian populations with and without PTSD (see Table 1) (97). The aryl-hydrocarbon receptor repressor (AHRR), a gene that regulates transcription, revealed four CpG sites in which DNA methylation was lower in individuals with PTSD versus the controls. This was found to be associated with the reduced kynurenine levels also seen in individuals with PTSD (97). The authors suggest that the reduced levels of kynurenine (a metabolite that promotes T-regulatory cell differentiation) may explain why increased inflammation is seen in individuals with PTSD. This form of gene methylation appears in both military and civilian populations with PTSD, which suggests that the HSE is not evident in this instance.
Differential expression of 25 genes associated with HPA, immune and/or cerebral function was seen in survivors of the 9/11 attack with PTSD (101). The altered expression of FKBP5 was revealed as an acute state marker for PTSD as well as major histocompatibility complex (MHC) Class II (vs. lifetime PTSD). In addition, the reduced expression of Stat five B inhibitor (STAT5B; a GR inhibitor 116) and nuclear factor I/A (key for astrocytic function 117) was seen in PTSD. These genes reveal that epigenetic impacts from PTSD impact on both the HPA axis and the immune system.
Rutten et al. compared the longitudinal changes of genome-wide blood DNA methylation profiles of two different subgroups of soldiers and identified the genes associated with stress in a military context (included in Table 1) (88). In the first cohort, consisting of Dutch military personnel, they discovered 17 differentially methylated positions (DMPs) and 12 DMRs in individuals after they had been deployed to a combat zone. The DMPs and DMRs were then used to explore replication in a different cohort of US marines. In this group, a decrease in DNA methylation at the genes ZFP57, RNF39, and HIST1H2APS2 was associated with an increase in PTSD symptoms over time. They found that the association between trauma exposure and PTSD symptoms was mediated by DNA methylation; those who did not exhibit PTSD symptoms showed increased methylation, but those who did develop PTSD symptoms exhibited decreased DNA methylation at the DMP and DMRs. Altered methylation on DUSP22 and PAX8 genes, earlier identified, revealed decreased methylation in both, with no change in expression to the former but increased expression in the latter (6, 88).
PTSD is often defined solely by symptom severity, despite the heterogenous nature of the symptoms themselves. To challenge this perception, Yang and colleagues defined the epigenetic biotypes of veterans/active-duty personnel and propensity for PTSD (118). They revealed two main subsets with opposing symptoms: G1 (faster recovery from PTSD, lower methylation vs. controls) and G2 (increased risk of PTSD, higher methylation vs. controls), enriched for individuals also exhibiting major depressive disorder (MDD). That study highlights the complex aetiology of PTSD and variation within military subsets. However, the authors did not discuss immunological components or pro-inflammatory processes, which also contribute to PTSD (99, 112, 119). The identification of if, and how, G1 and G2 express different immunological profiles would inform on whether immune function also differs with opposing symptoms.
Therefore, there is a large body of research exploring a plethora of epigenetic changes upon the individual exposed to war (listed with details in Table 1), which we also attempt to summarise here. As noted earlier, the FKBP5 and NR3C1 genes are epigenetically modified; in addition, research on war and conflict exposure has revealed epigenetic impacts to players in auditory, sleep and immune systems.
Intergenerational impacts of war
Paternal trauma experienced by the survivors of Confederate POW camps, during the US Civil War (1861–1865), was found to impact the life expectancy of their children (see Table 2) (120). During the first 2 years of conflict, there was an exchange period during which prisoners were swapped between the warring sides. However, a “no-exchange period” period occurred between July 1863 to 1864, during which increased camp populations resulted in worsened conditions and many resultant deaths of POWs on both sides. It was found that sons born post-war to POWs who experienced the no-exchange period were 9% and 11% more likely to die early in comparison to sons of exchange-period POWs and non-POWs, respectively. Within families, when comparing sons born pre-war to those born post-war from no-exchange POW fathers, the latter were 2.23 times more likely to die early. However, in this comparison, the authors note the caveat of small sample size. No significance was seen on the daughters of POWs. Post-war factors, such as paternal socioeconomic status, did not impact these results. The sex-specific effects seen in this study population are similar to an effect observed in the Överkalix population of Sweden, where grandparents experiencing an abundant harvest resulted in an increased risk of cardiovascular disease and reduced longevity (131, 132). As in the Överkalix studies, the authors propose that the epigenetic response is transmitted via the Y chromosome, which could explain why the effects are seen only in sons.
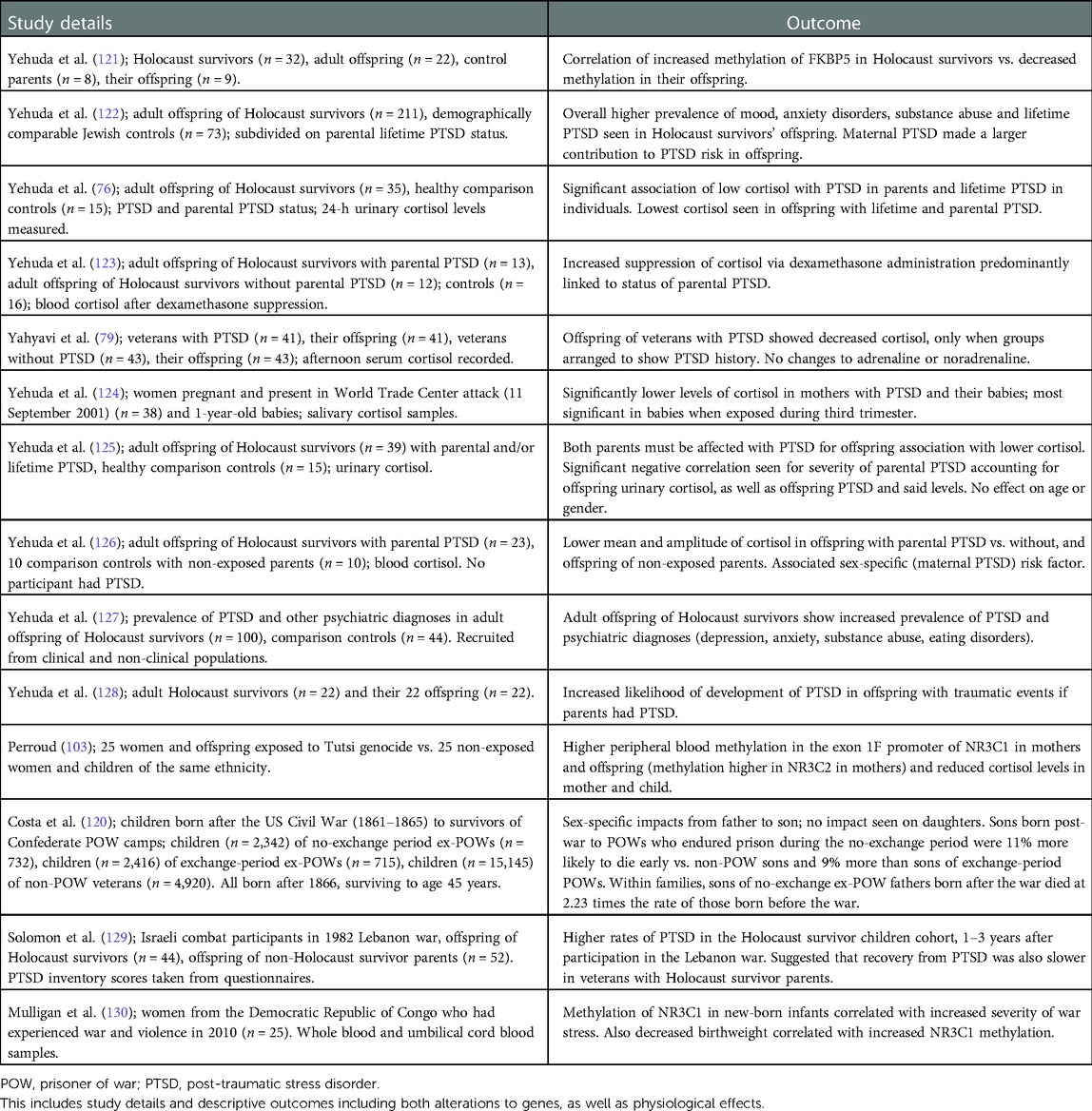
Table 2. Exposure to war and subsequent inherited epigenetics to offspring.
One recurrent effect noted in studies on the offspring of Holocaust survivors with PTSD was the impact of reduced cortisol levels and an increased risk of PTSD, with an association between the two (76, 123, 125–129). These data have been highlighted with the caveat of the family environment, which may influence behaviour and mindset. However, for Holocaust survivors, in keeping with the HPA axis effects, glucocorticoid sensitivity was increased in the offspring with maternal exposure to PTSD, yet decreased with paternal exposure, effects not influenced by parental care (122, 133). The children of Holocaust survivors exhibit either no prevalence towards psychiatric disorders (134), or in another study, higher instances of mood and anxiety disorders (122). A decreased cortisol effect was present in the offspring of veterans, but only those with a parental history of PTSD (79).
Individuals exposed to the World Trade Centre attacks produced both children and grandchildren who exhibit lower levels of salivary cortisol (124). The data are referred to as transgenerational, but, as grandmothers were pregnant at the time of the attacks, this, by definition, is an intergenerational effect.
Epigenetic effects via gene methylation on the offspring of those exposed to conflict have largely been shown in genes already associated with PTSD. The study discussed earlier by Perroud et al. on mothers exposed to the Tutsi genocide also explored impacts to offspring; both groups showed higher levels of PTSD and decreased cortisol in comparison to controls. Differential methylation was seen, with elevated methylation of NR3C1 (promoter region) seen only in offspring (vs. the increased level of NR3C2 methylation in mothers) (103). Prenatal exposure to maternal depression/anxiety has been associated with increased cortisol stress response at 3 months of age, and an associated increase of NR3C1 (predicted NGFI-A binding site) methylation (135). In the eastern Democratic Republic of Congo, mothers and their offspring in utero were exposed to the stressors of war. The offspring exhibited methylation on the NR3C1 (promoter region) gene, which differed from the mother, and correlated with prenatal exposure to stress (130). Finally, Yehuda et al. revealed, when compared to controls, enhanced methylation of FKBP5 (intron 7) in Holocaust survivors versus the lower levels of methylation in their offspring, outcomes that were significantly correlated (121). Differential methylation patterns between parent and offspring have been suggested as a form of compensation for the trauma experienced by the parents (136), further supported by a meta-analysis finding children of Holocaust survivors were generally well adapted, with little evidence to suggest secondary traumatisation (137).
To conclude this section, in comparison to the epigenetic effects impacting the individual, such as DNA methylation on a number of genes, data on inter- and transgenerational epigenetic effects of war and conflict are mainly limited to the well-established NR3C1 and FKBP5 genes, mortality, cortisol levels and risk of PTSD (summarised in Table 2). This highlights a large gap in the field; we do not know whether the progeny of those exposed to war may also exhibit higher risks of pathologies, such as sleep disruption, and metabolic and/or gastrointestinal effects as seen in their parent/s. Immunological dysfunction is linked to the HPA axis (138) and PTSD (112), so lowered cortisol levels in the progeny of those exposed to war could result in impacts on the immune system. To improve the health and well-being of those exposed to war, and their families, such effects should be defined to allow for appropriate and timely intervention.
Discussion
The present article shows that stressors associated with war and conflict have epigenetic impacts on health, at individual, inter- and/or transgenerational levels. Individual impacts uninfluenced by PTSD may present as gastrointestinal (1–4), auditory (5, 6), metabolic (7) and circadian (8) effects (see Figure 1). Shared effects between individuals and their progeny include an increased risk of PTSD (and co-occurring lowered cortisol), as well as changes to mortality; in some cases, these effects are sex-linked (120). Persistent epigenetic markers across generations exposed to war are seen in NR3C1, NR3C2, and FKBP5 genes, known players in the HPA axis. Future work may wish to consider the inter- and transgenerational inheritance of other genetic factors shown in Figure 1, such as whether AHRR methylation or reduced miR-125a expression in veterans (99) also occurs in the progeny of veterans and trauma-exposed civilians. We do not know what the long-term effects of the numerous conflicts mentioned in our introduction (e.g., the war in Ukraine or in Iran with the aggressive government measures against citizens during civil unrest) will be. However, we propose that there may be an increased risk of the pathologies described above. Thus, when providing effective international aid and support to individuals seeking refuge in the UK, being cognisant of such downstream epigenetic effects could inform mitigation strategies through short- and long-term health and social care. The British Medical Association (BMA) currently suggests a number of guidelines, named “Refugee and Asylum Seeker Health Toolkit,” which include issues refugees may have in general regarding the control of blood sugar and advises screening for communicable diseases. The GOV.UK “Migrant Health Guide” also provides specific guides for nutrition and mental health support, noting impacts of depression and anxiety. This highlights a focus on PTSD, recognising the necessity of addressing the pathology. We would propose additionally considering gastrointestinal dysfunction, impacts to hearing due to explosives and attempts to provide education on sleep.
We note potentially confounding (but unavoidable) factors from the sequalae of war. First, TBI from military trauma is known as a causative factor for PTSD and/or dementia (107). This therefore includes a psychological and physical influence upon PTSD. Another issue is concurrent malnutrition experienced in POW studies; famine results in cardiovascular impacts and the sole study revealing a cardiac effect (ischaemic heart disease) where POWs were reported as experiencing severe malnutrition (57, 58). Noxious auditory stimuli (e.g., blast exposure) may vary depending on the war environment, or job role within the military, and significant epigenetic effects occur with lifetime blast exposure. Age may also impact the prevalence of morbidity (55, 56). We mention the HSE influencing increased longevity in military personnel, but in more recent populations this effect appears eroded (139). As noted in Tables 1 and 2, most war-related studies have been conducted on Caucasian male veterans or African American individuals who have experienced trauma. More diverse studies, such as that of Smith and colleagues, are required to truly understand the possible contribution of factors such as race, sex, and cultural experiences (97). Despite the unique nature of military populations, many PTSD-linked alterations appear similar across veteran PTSD and PTSD linked to non-military adult trauma, e.g., AHRR methylation as a predicator for PTSD in both military and civilian populations (97). Thus far, when considering exposure to war, there is limited evidence to suggest, without PTSD or exposure to famine, that veterans specifically transmit substantive epigenetic impacts.
Author contributions
ZR performed the initial data search and established the original data collection. SFH expanded this research, added articles, original input and edited the manuscript. VSF structured and modified the manuscript, provided investigation to include further research and developed tables and figure. RSMG contributed intellectual and supervisory support as well as editing. PJC, JFM and JW also provided intellectual input. All authors contributed to the article and approved the submitted version.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ADCYAP1R1, pituitary adenylate cyclase-activating polypeptide type I receptor (PAC-1); AHRR, aryl-hydrocarbon receptor repressor; BDNF, brain-derived neurotrophic factor; CpG, cytosine and guanine appearing consecutively on the same strand of nucleic acid; gene methylation often occurring on the cytosine; DMP, differentially methylated position; DMR, differentially methylated region; FKBP5, FK506 binding protein 5, involved in assembly of glucocorticoid receptor complex; GC, glucocorticoid; GR, glucocorticoid receptor; IBS, irritable bowel syndrome; MAO-A, monoamine oxidase-A; MAO-B, monoamine oxidase-B; PACAP, pituitary adenylate cyclase-activating polypeptide; POW, prisoner of war; PTSD, post-traumatic stress disorder; SNP, single nucleotide polymorphism; TBI, traumatic brain injury; HDAC, histone deacetylate; HPA, hypothalamic-pituitary-adrenal; HSE, healthy soldier effect; MHC, major histocompatibility complex; NR3C1, nuclear receptor subfamily 3 group C member 1, glucocorticoid receptor gene; NR3C2, nuclear receptor subfamily 3 group C member 2, mineral glucocorticoid receptor gene; STAT5B, stat five B inhibitor, GR inhibitor; VPA, valproic acid, HDAC inhibitor.
References
1. Goulston KJ, Dent OF, Chapuis PH, Chapman G, Smith CI, Tait AD, Tennant CC. Gastrointestinal morbidity among World War II prisoners of war: 40 years on. Med J Aust. (1985) 143(1):6–10. doi: 10.5694/j.1326-5377.1985.tb122757.x
2. Dominitz JA, Provenzale D. Prevalence of dyspepsia, heartburn, and peptic ulcer disease in veterans. Am J Gastroenterol. (1999) 94(8):2086–93. doi: 10.1111/j.1572-0241.1999.01282.x
3. Klooker TK, Braak B, Painter RC, de Rooij SR, van Elburg RM, van den Wijngaard RM, Roseboom TJ, Boeckxstaens GE. Exposure to severe wartime conditions in early life is associated with an increased risk of irritable bowel syndrome: a population-based cohort study. Am J Gastroenterol. (2009) 104(9):2250–6. doi: 10.1038/ajg.2009.282
4. Riddle MS, Welsh M, Porter CK, Nieh C, Boyko EJ, Gackstetter G, Hooper TI. The epidemiology of irritable bowel syndrome in the US military: findings from the Millennium Cohort Study. Am J Gastroenterol. (2016) 111(1):93–104. doi: 10.1038/ajg.2015.386
5. Wang Z, Wilson CM, Ge Y, Nemes J, LaValle C, Boutté A, Carr W, Kamimori G, Haghighi F. DNA Methylation patterns of chronic explosive breaching in U.S. military warfighters. Front Neurol (2020) 11:1010. doi: 10.3389/fneur.2020.01010
6. Wang Z, Wilson CM, Mendelev N, Ge Y, Galfalvy H, Elder G, Ahlers S, Yarnell AM, LoPresti ML, Kamimori GH, Carr W, Haghighi F. Acute and chronic molecular signatures and associated symptoms of blast exposure in military breachers. J Neurotrauma. (2020) 37(10):1221–32. doi: 10.1089/neu.2019.6742
7. Fryar CD, Herrick K, Afful J, Ogden CL. Cardiovascular disease risk factors among male veterans, U.S., 2009–2012. Am J Prev Med. (2016) 50(1):101–5. doi: 10.1016/j.amepre.2015.06.011
8. Alexander M, Ray MA, Hébert JR, Youngstedt SD, Zhang H, Steck SE, Bogan RK, Burch JB. The national veteran sleep disorder study: descriptive epidemiology and secular trends, 2000–2010. Sleep. (2016) 39(7):1399–410. doi: 10.5665/sleep.5972
9. Faulk C, Dolinoy DC. Timing is everything: the when and how of environmentally induced changes in the epigenome of animals. Epigenetics. (2014) 6(7):791–7. doi: 10.4161/epi.6.7.16209
10. Bakusic J, Schaufeli W, Claes S, Godderis L. Stress, burnout and depression: a systematic review on DNA methylation mechanisms. J Psychosom Res. (2017) 92:34–44. doi: 10.1016/j.jpsychores.2016.11.005
11. Nagy C, Turecki G. Transgenerational epigenetic inheritance: an open discussion. Epigenomics. (2015) 7(5):781–90. doi: 10.2217/epi.15.46
12. Dashorst P, Mooren TM, Kleber RJ, de Jong PJ, Huntjens RJC. Intergenerational consequences of the holocaust on offspring mental health: a systematic review of associated factors and mechanisms. Eur J Psychotraumatol. (2019) 10(1):1654065. –. doi: 10.1080/20008198.2019.1654065
13. Münyas B. Genocide in the minds of Cambodian youth: transmitting (hi)stories of genocide to second and third generations in Cambodia. J Genocide Res. (2008) 10(3):413–39. doi: 10.1080/14623520802305768
14. Raphael B, Swan P, Martinek N. Intergenerational aspects of trauma for Australian aboriginal people. In: Danieli Y, editors. International handbook of multigenerational legacies of trauma. Boston, MA: Springer (1998). p. 327–39.
15. Vaiserman A, Lushchak O. Prenatal famine exposure and adult health outcomes: an epigenetic link. Environ Epigenet (2021) 7(1): dvab013. doi: 10.1093/eep/dvab013
16. Zepinic V, Bogic M, Priebe S. Refugees’ views of the effectiveness of support provided by their host countries. Eur J Psychotraumatol. (2012) 3:8447. doi: 10.3402/ejpt.v3i0.8447
17. Bremner JD. Traumatic stress: effects on the brain. Dialogues Clin Neurosci. (2006) 8(4):445–61. doi: 10.31887/DCNS.2006.8.4/jbremner
18. Yehuda R, Giller EL, Mason JW. Psychoneuroendocrine assessment of posttraumatic stress disorder: current progress and new directions. Prog Neuropsychopharmacol Biol Psychiatry. (1993) 17(4):541–50. doi: 10.1016/0278-5846(93)90004-C
19. Laryea G, Schütz G, Muglia LJ. Disrupting hypothalamic glucocorticoid receptors causes HPA axis hyperactivity and excess adiposity. Mol Endocrinol. (2013) 27(10):1655. doi: 10.1210/me.2013-1187
20. Yehuda R. Post-traumatic stress disorder. N Engl J Med. (2002) 346(2):108–14. doi: 10.1056/NEJMra012941
21. Yehuda R, Golier JA, Kaufman S. Circadian rhythm of salivary cortisol in holocaust survivors with and without PTSD. Am J Psychiatry. (2005) 162(5):998–1000. doi: 10.1176/appi.ajp.162.5.998
22. Aerni A, Traber R, Hock C, Roozendaal B, Schelling G, Papassotiropoulos A, Nitsch RM, Schnyder U, de Quervain DJF. Low-dose cortisol for symptoms of posttraumatic stress disorder. Am J Psychiatry. (2004) 161(8):1488–90. doi: 10.1176/appi.ajp.161.8.1488
23. Yehuda R, Giller EL, Boisoneau D, Lowry MT, Southwick SM, Mason JW. Low dose DST in PTSD (abstract NR144). In: New research abstracts of the American Psychiatric Association 144th annual meeting. University of Connecticut Health Center, Farmington, CT; (1991).
24. Yehuda R, Boisoneau D, Mason JW, Giller EL. Glucocorticoid receptor number and cortisol excretion in mood, anxiety, and psychotic disorders. Biol Psychiatry. (1993) 34(1):18–25. doi: 10.1016/0006-3223(93)90252-9
25. Perroud N, Paoloni-Giacobino A, Prada P, Olié E, Salzmann A, Nicastro R, et al. Increased methylation of glucocorticoid receptor gene (NR3C1) in adults with a history of childhood maltreatment: a link with the severity and type of trauma. Transl Psychiatry. (2011) 1(12):e59. doi: 10.1038/tp.2011.60
26. McGowan PO, Sasaki A, D'Alessio AC, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci. (2009) 12(3):342–8. doi: 10.1038/nn.2270
27. Labonté B, Azoulay N, Yerko V, Turecki G, Brunet A. Epigenetic modulation of glucocorticoid receptors in posttraumatic stress disorder. Transl Psychiatry. (2014) 4(4):e368. doi: 10.1038/tp.2014.3
28. Young DA, Inslicht SS, Metzler TJ, Neylan TC, Ross JA. The effects of early trauma and the FKBP5 gene on PTSD and the HPA axis in a clinical sample of Gulf War veterans. Psychiatry Res. (2018) 270:961–6. doi: 10.1016/j.psychres.2018.03.037
29. Ising M, Depping A, Siebertz A, et al. Polymorphisms in the FKBP5 gene region modulate recovery from psychosocial stress in healthy controls. Eur J Neurosci. (2008) 28:389–98. doi: 10.1111/j.1460-9568.2008.06332.x
30. Klengel T, Mehta D, Anacker C, et al. Allele-specific FKBP5 DNA demethylation mediates gene–childhood trauma interactions. Nat Neurosci. (2013) 16(1):33–41. doi: 10.1038/nn.3275
31. Yehuda R, Daskalakis NP, Desarnaud F, et al. Epigenetic biomarkers as predictors and correlates of symptom improvement following psychotherapy in combat veterans with PTSD. Front Psychiatry. (2013) 4:118. doi: 10.3389/fpsyt.2013.00118
32. Zannas AS, Provençal N, Binder EB. Epigenetics of posttraumatic stress disorder: current evidence, challenges, and future directions. Biol Psychiatry. (2015) 78(5):327–35. doi: 10.1016/j.biopsych.2015.04.003
33. Pape JC, Carrillo-Roa T, Rothbaum BO, Nemeroff CB, Czamara D, Zannas AS, et al. DNA methylation levels are associated with CRF1 receptor antagonist treatment outcome in women with post-traumatic stress disorder. Clin Epigenetics. (2018) 10(1):136. doi: 10.1186/s13148-018-0569-x
34. Ressler KJ, Mercer KB, Bradley B, et al. Post-traumatic stress disorder is associated with PACAP and the PAC1 receptor. Nature. (2011) 470(7335):492. doi: 10.1038/nature09856
35. Mehta D, Bruenig D, Carrillo-Roa T, Lawford B, Harvey W, Morris CP, Smith AK, Binder EB, Young RM, Voisey J. Genomewide DNA methylation analysis in combat veterans reveals a novel locus for PTSD. Acta Psychiatr Scand. (2017) 136(5):493–505. doi: 10.1111/acps.12778
36. Murphy C, Singewald N. Potential of microRNAs as novel targets in the alleviation of pathological fear. Genes Brain Behav. (2018) 17(3):e12427. doi: 10.1111/gbb.12427
37. Foa E, Hembree E, Rothbaum BO. Prolonged exposure therapy for PTSD: Emotional processing of traumatic experiences therapist guide. New York: Oxford University Press (2007).
38. Kwapis JL, Wood MA. Epigenetic mechanisms in fear conditioning: implications for treating post-traumatic stress disorder. Trends Neurosci. (2014) 37(12):706–20. doi: 10.1016/j.tins.2014.08.005
39. Bredy TW, Barad M. The histone deacetylase inhibitor valproic acid enhances acquisition, extinction, and reconsolidation of conditioned fear. Learn Mem. (2008) 15(1):39–45. doi: 10.1101/lm.801108
40. Adamou M, Puchalska S, Plummer W, Hale AS. Valproate in the treatment of PTSD: systematic review and meta analysis. Curr Med Res Opin. (2007) 23(6):1285–91. doi: 10.1185/030079907X188116
41. Yoshiike T, Kuriyama K. Valproic acid in the treatment of post traumatic stress disorder. In: Martin CR, Preedy VR, Patel VB, editors. Comprehensive guide to post-traumatic stress disorder. Cham: Springer (2014). p. 1–11.
42. Kessler RC, Sonnega A, Bromet E, Hughes M, Nelson CB. Posttraumatic stress disorder in the National Comorbidity Survey. Arch Gen Psychiatry. (1995) 52(12):1048–60. doi: 10.1001/archpsyc.1995.03950240066012
43. Hartal M, Kreiss Y, Yavnai N. Relative longevity among retired military personnel: a historical-cohort study. Mil Med Res. (2015) 2:29. doi: 10.1186/s40779-015-0057-y
44. Waller M, McGuire ACL. Changes over time in the “healthy soldier effect.” Popul Health Metr. (2011) 9(1):7. doi: 10.1186/1478-7954-9-7
45. Shah D. Healthy worker effect phenomenon. Indian J Occup Environ Med. (2009) 13(2):77–9. doi: 10.4103/0019-5278.55123
46. Lundquist J, Elo I, Barfield W, Xu Z. Racial disparities in US infant birth outcomes: a protective effect of military affiliation? Race Soc Probl. (2013) 6:161. doi: 10.1007/s12552-013-9108-8
47. Kimbrell T, Pyne JM, Kunik ME, Magruder KM, Petersen NJ, Yu HJ, Hudson TJ, Schulz PE, Qureshi SU. The impact of purple heart commendation and PTSD on mortality rates in older veterans. Depress Anxiety. (2011) 28(12):1086–90. doi: 10.1002/da.20850
48. Verhoeven JE, Yang R, Wolkowitz OM, Bersani FS, Lindqvist D, Mellon SH, Yehuda R, Flory JD, Lin J, Abu-Amara D, Makotkine I, Marmar C, Jett M, Hammamieh R. Epigenetic age in male combat-exposed war veterans: associations with posttraumatic stress disorder status. Mol Neuropsychiatry. (2018) 4(2):90–9. doi: 10.1159/000491431
49. Dzubur Kulenović A, Kucukalić A, Malec D. Changes in plasma lipid concentrations and risk of coronary artery disease in army veterans suffering from chronic posttraumatic stress disorder. Croat Med J. (2008) 49(4):506–14. doi: 10.3325/cmj.2008.4.506
50. Morgan CA. 3rd, Grillon C, Southwick SM, Nagy LM, Davis M, Krystal JH, Charney DS. Yohimbine facilitated acoustic startle in combat veterans with post-traumatic stress disorder. Psychopharmacology (Berl) (1995) 117(4): 466–71. doi: 10.1007/BF02246220
51. Wilkerson AK, Uhde TW, Leslie K, Freeman WC, LaRowe SD, Schumann A, Cortese BM. Paradoxical olfactory function in combat veterans: the role of PTSD and odor factors. Mil Psychol. (2018) 30(2):120–30. doi: 10.1080/08995605.2018.1425063
52. Shipherd JC, Keyes M, Jovanovic T, Ready DJ, Baltzell D, Worley V, Gordon-Brown V, Hayslett C, Duncan E. Veterans seeking treatment for posttraumatic stress disorder: what about comorbid chronic pain? J Rehabil Res Dev. (2007) 44(2):153–66. doi: 10.1682/JRRD.2006.06.0065
53. Asmundson GJ, Wright KD, Stein MB. Pain and PTSD symptoms in female veterans. Eur J Pain. (2004) 8(4):345–50. doi: 10.1016/j.ejpain.2003.10.008
54. Tennant CC, Goulston KJ, Dent OF. The psychological effects of being a prisoner of war: forty years after release. Am J Psychiatry. (1986) 143(5):618–21. doi: 10.1176/ajp.143.5.618
55. Dent OF, Richardson B, Wilson S, Goulston KJ, Murdoch CW. Postwar mortality among Australian World War II prisoners of the Japanese. Med J Aust. (1989) 150(7):378–82. doi: 10.5694/j.1326-5377.1989.tb136529.x
56. Costa DL. Scarring and mortality selection among Civil War POWs: a long-term mortality, morbidity, and socioeconomic follow-up. Demography. (2012) 49(4):1185–206. doi: 10.1007/s13524-012-0125-9
57. Page WF, Ostfeld AM. Malnutrition and subsequent ischemic heart disease in former prisoners of war of World War II and the Korean conflict. J Clin Epidemiol. (1994) 47(12):1437–41. doi: 10.1016/0895-4356(94)90087-6
58. Page WF, Brass LM. Long-term heart disease and stroke mortality among former American prisoners of war of World War II and the Korean conflict: results of a 50-year follow-up. Mil Med Res. (2001) 166(9):803–8. doi: 10.1093/milmed/166.9.803
59. van Abeelen AF, Veenendaal MV, Painter RC, de Rooij SR, Dijkgraaf MG, Bossuyt PM, Elias SG, Grobbee DE, Uiterwaal CS, Roseboom TJ. Survival effects of prenatal famine exposure. Am J Clin Nutr. (2012) 95(1):179–83. doi: 10.3945/ajcn.111.022038
60. Grey K, Gonzales GB, Abera M, Lelijveld N, Thompson D, Berhane M, Abdissa A, Girma T, Kerac M. Severe malnutrition or famine exposure in childhood and cardiometabolic non-communicable disease later in life: a systematic review. BMJ Glob Health. (2021) 6(3):e003161. doi: 10.1136/bmjgh-2020-003161
61. Williams RL, Medalie JH, Zyzanski SJ, Flocke SA, Yaari S, Goldbourt U. Long-term mortality of Nazi concentration camp survivors. J Clin Epidemiol. (1993) 46(6):573–5. doi: 10.1016/0895-4356(93)90130-S
62. Collins C, Burazeri G, Gofin J, Kark JD. Health status and mortality in Holocaust survivors living in Jerusalem 40–50 years later. J Trauma Stress (2004) 17(5):403–11. doi: 10.1023/B:JOTS.0000048953.27980.18
63. Trappler B, Cohen CI, Tulloo R. Impact of early lifetime trauma in later life: depression among Holocaust survivors 60 years after the liberation of Auschwitz. Am J Geriatr Psychiatry. (2007) 15(1):79–83. doi: 10.1097/01.JGP.0000229768.21406.a7
64. Liu Y, Collins C, Wang K, Xie X, Bie R. The prevalence and trend of depression among veterans in the United States. J Affective Disorders. (2019) 245:724–7. doi: 10.1016/j.jad.2018.11.031
65. Rauch SAM, Favorite T, Giardino N, Porcari C, Defever E, Liberzon I. Relationship between anxiety, depression, and health satisfaction among veterans with PTSD. J Affect Disord. (2010) 121(1):165–8. doi: 10.1016/j.jad.2009.05.026
66. Murphy D, Iversen A, Greenberg N. The mental health of veterans. J R Army Med Corps. (2008) 154(2):136–9. doi: 10.1136/jramc-154-02-13
67. Kypraiou A, Sarafis P, Tsounis A, Bitsi G, Andreanides E, Constantinidis T, Kotrotsiou E, Malliarou M. Depression and anxiety in Greek male veterans after retirement. Mil Med. (2017) 182(3):e1639–44. doi: 10.7205/MILMED-D-16-00299
68. Fogle BM, Tsai J, Mota N, Harpaz-Rotem I, Krystal JH, Southwick SM, Pietrzak RH. The national health and resilience in veterans study: a narrative review and future directions. Front Psychiatry. (2020) 11:538218. doi: 10.3389/fpsyt.2020.538218
69. Milanak ME, Gros DF, Magruder KM, Brawman-Mintzer O, Frueh BC. Prevalence and features of generalized anxiety disorder in Department of Veteran Affairs primary care settings. Psychiatry Res. (2013) 209(2):173–9. doi: 10.1016/j.psychres.2013.03.031
70. Lee B-H, Kim Y-K. The roles of BDNF in the pathophysiology of major depression and in antidepressant treatment. Psychiatry Investig. (2010) 7(4):231–5. doi: 10.4306/pi.2010.7.4.231
71. Kaplan GB, Vasterling JJ, Vedak PC. Brain-derived neurotrophic factor in traumatic brain injury, post-traumatic stress disorder, and their comorbid conditions: role in pathogenesis and treatment. Behav Pharmacol. (2010) 21(5–6):427–37. doi: 10.1097/FBP.0b013e32833d8bc9
72. Kim TY, Kim SJ, Chung HG, Choi JH, Kim SH, Kang JI. Epigenetic alterations of the BDNF gene in combat-related post-traumatic stress disorder. Acta Psychiatr Scand. (2017) 135(2):170–9. doi: 10.1111/acps.12675
73. Yaffe K, Vittinghoff E, Lindquist K, Barnes D, Covinsky KE, Neylan T, Kluse M, Marmar C. Posttraumatic stress disorder and risk of dementia among US veterans. Arch Gen Psychiatry. (2010) 67(6):608–13. doi: 10.1001/archgenpsychiatry.2010.61
74. Qureshi SU, Kimbrell T, Pyne JM, Magruder KM, Hudson TJ, Petersen NJ, Yu HJ, Schulz PE, Kunik ME. Greater prevalence and incidence of dementia in older veterans with posttraumatic stress disorder. J Am Geriatr Soc. (2010) 58(9):1627–33. doi: 10.1111/j.1532-5415.2010.02977.x
75. Yehuda R, Kahana B, Binder-Brynes K, Southwick SM, Mason JW, Giller EL. Low urinary cortisol excretion in Holocaust survivors with posttraumatic stress disorder. Am J Psychiatry. (1995) 152(7):982–6. doi: 10.1176/ajp.152.7.982
76. Yehuda R, Bierer LM, Schmeidler J, Aferiat DH, Breslau I, Dolan S. Low cortisol and risk for PTSD in adult offspring of holocaust survivors. Am J Psychiatry. (2000) 157(8):1252–9. doi: 10.1176/appi.ajp.157.8.1252
77. Yehuda R, Golier JA, Tischler L, Harvey PD, Newmark R, Yang RK, Buchsbaum MS. Hippocampal volume in aging combat veterans with and without post-traumatic stress disorder: relation to risk and resilience factors. J Psychiatr Res. (2007) 41(5):435–45. doi: 10.1016/j.jpsychires.2005.12.002
78. Geracioti TD Jr., Baker DG, Ekhator NN, West SA, Hill KK, Bruce AB, Schmidt D, Rounds-Kugler B, Yehuda R, Keck PE Jr., Kasckow JW. CSF Norepinephrine concentrations in posttraumatic stress disorder. Am J Psychiatry (2001) 158(8):1227–30. doi: 10.1176/appi.ajp.158.8.1227
79. Yahyavi ST, Zarghami M, Naghshvar F, Danesh A. Relationship of cortisol, norepinephrine, and epinephrine levels with war-induced posttraumatic stress disorder in fathers and their offspring. Braz J Psychiatry. (2015) 37(2):93–8. doi: 10.1590/1516-4446-2014-1414
80. Hendrickson RC, Raskind MA, Millard SP, Sikkema C, Terry GE, Pagulayan KF, Li G, Peskind ER. Evidence for altered brain reactivity to norepinephrine in veterans with a history of traumatic stress. Neurobiol Stress. (2018) 8:103–11. doi: 10.1016/j.ynstr.2018.03.001
81. Hammamieh R, Chakraborty N, Gautam A, Muhie S, Yang R, Donohue D, Kumar R, Daigle BJ Jr., Zhang Y, Amara DA, Miller SA, Srinivasan S, Flory J, Yehuda R, Petzold L, Wolkowitz OM, Mellon SH, Hood L, Doyle FJ, 3rd., Marmar C, Jett M. Whole-genome DNA methylation status associated with clinical PTSD measures of OIF/OEF veterans. Transl Psychiatry. (2017) 7(7):e1169. doi: 10.1038/tp.2017.129
82. Bedont JL, LeGates TA, Buhr E, Bathini A, Ling JP, Bell B, Wu MN, Wong PC, van Gelder RN, Mongrain V, Hattar S, Blackshaw S. An LHX1-regulated transcriptional network controls sleep/wake coupling and thermal resistance of the central circadian clockworks. Curr Biol. (2017) 27(1):128–36. doi: 10.1016/j.cub.2016.11.008
83. Papaccio F, Della Corte CM, Viscardi G, Di Liello R, Esposito G, Sparano F, Ciardiello F, Morgillo F. HGF/MET and the immune system: relevance for cancer immunotherapy. Int J Mol Sci. (2018) 19(11):3595. doi: 10.3390/ijms19113595
84. Calderón L, Schindler K, Malin SG, Schebesta A, Sun Q, Schwickert T, Alberti C, Fischer M, Jaritz M, Tagoh H, Ebert A, Minnich M, Liston A, Cochella L, Busslinger M. Pax5 regulates B cell immunity by promoting PI3K signaling via PTEN down-regulation. Sci Immunol. (2021) 6(61):eabg5003. doi: 10.1126/sciimmunol.abg5003
85. Macian F. NFAT Proteins: key regulators of T-cell development and function. Nat Rev Immunol. (2005) 5(6):472–84. doi: 10.1038/nri1632
86. Azad F M, Malakootian M, Mowla SJ. lncRNA PSORS1C3 is regulated by glucocorticoids and fine-tunes OCT4 expression in non-pluripotent cells. Sci Rep. (2019) 9(1):8370. doi: 10.1038/s41598-019-44827-7
87. Lieberman J, Wu H, Kagan Jonathan C. Gasdermin D activity in inflammation and host defense. Sci Immunol. (2019) 4(39):eaav1447. doi: 10.1126/sciimmunol.aav1447
88. Rutten BP VE, Vinkers CH, Ursini G, Daskalakis NP, Pishva E, de Nijs L, et al. Longitudinal analyses of the DNA methylome in deployed military servicemen identify susceptibility loci for post-traumatic stress disorder. Mol Psychiatry. (2018) 23:1145–56. doi: 10.1038/mp.2017.120
89. Guo HM, Zhang Y, Zhang Y, Jiao PF, Fan XC, Kong CL, Wang T, Li XX, Zhang HW, Zhang LR, Ma MY, Bu HL. Spinal Ninjurin2 contributes to the neuropathic pain via NF-κB-mediated neuroinflammation in the spared sciatic nerve injury rats. Int Immunopharmacol. (2021) 99:107918. doi: 10.1016/j.intimp.2021.107918
90. Yamagata M. Structure and functions of sidekicks. Front Mol Neurosci. (2020) 13:139. doi: 10.3389/fnmol.2020.00139
91. Mansfield P, Constantino JN, Baldridge D. MYT1L: a systematic review of genetic variation encompassing schizophrenia and autism. American Journal of Medical Genetics Part B: Neuropsychiatric Genetics. (2020) 183(4):227–33. doi: 10.1002/ajmg.b.32781
92. Sanchez-Mut JV, Heyn H, Silva BA, Dixsaut L, Garcia-Esparcia P, Vidal E, Sayols S, Glauser L, Monteagudo-Sánchez A, Perez-Tur J, Ferrer I, Monk D, Schneider B, Esteller M, Gräff J. PM20D1 is a quantitative trait locus associated with Alzheimer’s disease. Nat Med. (2018) 24(5):598–603. doi: 10.1038/s41591-018-0013-y
93. Liu B, Mao N. Smad5: signaling roles in hematopoiesis and osteogenesis. Int J Biochem Cell Biol. (2004) 36(5):766–70. doi: 10.1016/S1357-2725(03)00250-4
94. Allende G, Chávez-Reyes J, Guerrero-Alba R, Vázquez-León P, Marichal-Cancino BA. Advances in neurobiology and pharmacology of GPR12. Front Pharmacol. (2020) 11:628. doi: 10.3389/fphar.2020.00628
95. Acurzio B, Verma A, Polito A, Giaccari C, Cecere F, Fioriniello S, Della Ragione F, Fico A, Cerrato F, Angelini C, Feil R, Riccio A. Zfp57 inactivation illustrates the role of ICR methylation in imprinted gene expression during neural differentiation of mouse ESCs. Sci Rep. (2021) 11(1):13802. doi: 10.1038/s41598-021-93297-3
96. Sadeh N, Spielberg JM, Logue MW, et al. SKA2 methylation is associated with decreased prefrontal cortical thickness and greater PTSD severity among trauma-exposed veterans. Mol Psychiatry. (2016) 21(3):357. doi: 10.1038/mp.2015.134
97. Smith AK, Ratanatharathorn A, Maihofer AX, Naviaux RK, Aiello AE, Amstadter AB, Ashley-Koch AE, Baker DG, Beckham JC, Boks MP, Bromet E, Dennis M, Galea S, Garrett ME, Geuze E, Guffanti G, Hauser MA, Katrinli S, Kilaru V, Kessler RC, Kimbrel NA, Koenen KC, Kuan PF, Li K, Logue MW, Lori A, Luft BJ, Miller MW, Naviaux JC, Nugent NR, Qin X, Ressler KJ, Risbrough VB, Rutten BPF, Stein MB, Ursano RJ, Vermetten E, Vinkers CH, Wang L, Youssef NA, Uddin M, Nievergelt CM. Epigenome-wide meta-analysis of PTSD across 10 military and civilian cohorts identifies methylation changes in AHRR. Nat Commun. (2020) 11(1):5965. doi: 10.1038/s41467-020-19615-x
98. Ziegler C, Wolf C, Schiele MA, et al. Monoamine oxidase A gene methylation and its role in posttraumatic stress disorder: first evidence from the South Eastern Europe (SEE)-PTSD study. Int J Neuropsychopharmacol. (2017) 21(5):423–32. doi: 10.1093/ijnp/pyx111
99. Zhou J, Nagarkatti P, Zhong Y, Ginsberg JP, Singh NP, Zhang J, Nagarkatti M. Dysregulation in microRNA expression is associated with alterations in immune functions in combat veterans with post-traumatic stress disorder. PLoS One. (2014) 9(4):e94075. doi: 10.1371/journal.pone.0094075
100. Yehuda R, Flory JD, Bierer LM, et al. Lower methylation of glucocorticoid receptor gene promoter 1F in peripheral blood of veterans with posttraumatic stress disorder. Biol Psychiatry. (2015) 77(4):356–64. doi: 10.1016/j.biopsych.2014.02.006
101. Sarapas C, Cai G, Bierer L, Golier J, Galea S, Ising M, Rein T, Schmeidler J, Müller-Myhsok B, Uhr M, Holsboer F, Buxbaum J, Yehuda R. Genetic markers for PTSD risk and resilience among survivors of the World Trade Center attacks. Dis Markers. (2011) 30(2–3):101–10. doi: 10.1155/2011/328054
102. Shih JC. Molecular basis of human MAO A and B. Neuropsychopharmacology. (1991) 4(1):1–7. PMID: 2003865
103. Perroud N, Rutembesa E, Paoloni-Giacobino A., Mutabaruka J, Mutesa L, Stenz L, Malafosse A, Karege F. The Tutsi genocide and transgenerational transmission of maternal stress: epigenetics and biology of the HPA axis. World J Biol Psychiatry (2014) 15(4):334–45. doi: 10.3109/15622975.2013.866693
104. Rice L, Waters CE, Eccles J, Garside H, Sommer P, Kay P, Blackhall FH, Zeef L, Telfer B, Stratford I, Clarke R, Singh D, Stevens A, White A, Ray DW. Identification and functional analysis of SKA2 interaction with the glucocorticoid receptor. J Endocrinol. (2008) 198(3):499–509. doi: 10.1677/JOE-08-0019
105. Guintivano J, Brown T, Newcomer A, Jones M, Cox O, Maher BS, Eaton WW, Payne JL, Wilcox HC, Kaminsky ZA. Identification and replication of a combined epigenetic and genetic biomarker predicting suicide and suicidal behaviors. Am J Psychiatry. (2014) 171(12):1287–96. doi: 10.1176/appi.ajp.2014.14010008
106. Niculescu AB, Levey D, Le-Niculescu H, Niculescu E, Kurian SM, Salomon D. Psychiatric blood biomarkers: avoiding jumping to premature negative or positive conclusions. Mol Psychiatry. (2015) 20(3):286–8. doi: 10.1038/mp.2014.180
107. Raza Z, Hussain SF, Ftouni S, Spitz G, Caplin N, Foster RG, Gomes RSM. Dementia in military and veteran populations: a review of risk factors-traumatic brain injury, post-traumatic stress disorder, deployment, and sleep. Mil Med Res. (2021) 8(1):55. doi: 10.1186/s40779-021-00346-z
108. Mansouri A, Chowdhury K, Gruss P. Follicular cells of the thyroid gland require Pax8 gene function. Nat Genet. (1998) 19(1):87–90. doi: 10.1038/ng0598-87
109. Gottlieb DJ, Hek K, Chen TH, Watson NF, Eiriksdottir G, Byrne EM, Cornelis M, Warby SC, Bandinelli S, Cherkas L, Evans DS, Grabe HJ, Lahti J, Li M, Lehtimäki T, Lumley T, Marciante KD, Pérusse L, Psaty BM, Robbins J, Tranah GJ, Vink JM, Wilk JB, Stafford JM, Bellis C, Biffar R, Bouchard C, Cade B, Curhan GC, Eriksson JG, Ewert R, Ferrucci L, Fülöp T, Gehrman PR, Goodloe R, Harris TB, Heath AC, Hernandez D, Hofman A, Hottenga JJ, Hunter DJ, Jensen MK, Johnson AD, Kähönen M, Kao L, Kraft P, Larkin EK, Lauderdale DS, Luik AI, Medici M, Montgomery GW, Palotie A, Patel SR, Pistis G, Porcu E, Quaye L, Raitakari O, Redline S, Rimm EB, Rotter JI, Smith AV, Spector TD, Teumer A, Uitterlinden AG, Vohl MC, Widen E, Willemsen G, Young T, Zhang X, Liu Y, Blangero J, Boomsma DI, Gudnason V, Hu F, Mangino M, Martin NG, O'Connor GT, Stone KL, Tanaka T, Viikari J, Gharib SA, Punjabi NM, Räikkönen K, Völzke H, Mignot E, Tiemeier H. Novel loci associated with usual sleep duration: the CHARGE Consortium Genome-Wide Association Study. Mol Psychiatry. (2015) 20(10):1232–9. doi: 10.1038/mp.2014.133
110. Wagner U, Hallschmid M, Rasch B, Born J. Brief sleep after learning keeps emotional memories alive for years. Biol Psychiatry. (2006) 60(7):788–90. doi: 10.1016/j.biopsych.2006.03.061
111. Hertenstein E, Feige B, Gmeiner T, Kienzler C, Spiegelhalder K, Johann A, Jansson-Fröjmark M, Palagini L, Rücker G, Riemann D, Baglioni C. Insomnia as a predictor of mental disorders: a systematic review and meta-analysis. Sleep Med Rev. (2019) 43:96–105. doi: 10.1016/j.smrv.2018.10.006
112. Gill JM, Saligan L, Woods S, Page G. PTSD is associated with an excess of inflammatory immune activities. Perspect Psychiatr Care. (2009) 45(4):262–77. doi: 10.1111/j.1744-6163.2009.00229.x
113. Kim SW, Ramasamy K, Bouamar H, Lin AP, Jiang D, Aguiar RC. MicroRNAs miR-125a and miR-125b constitutively activate the NF-κB pathway by targeting the tumor necrosis factor alpha-induced protein 3 (TNFAIP3, A20). Proc Natl Acad Sci U S A. (2012) 109(20):7865–70. doi: 10.1073/pnas.1200081109
114. Si J, Yang J, Xue L, Yang C, Luo Y, Shi H, Lu L. Activation of NF-κB in basolateral amygdala is required for memory reconsolidation in auditory fear conditioning. PLoS One. (2012) 7(9):e43973. doi: 10.1371/journal.pone.0043973
115. Merlo E, Freudenthal R, Maldonado H, Romano A. Activation of the transcription factor NF-kappaB by retrieval is required for long-term memory reconsolidation. Learn Mem. (2005) 12(1):23–9. doi: 10.1101/lm.82705
116. Stöcklin E, Wissler M, Gouilleux F, Groner B. Functional interactions between Stat5 and the glucocorticoid receptor. Nature. (1996) 383(6602):726–8. doi: 10.1038/383726a0
117. Laug D, Huang TW, Huerta NAB, Huang AY, Sardar D, Ortiz-Guzman J, Carlson JC, Arenkiel BR, Kuo CT, Mohila CA, Glasgow SM, Lee HK, Deneen B. Nuclear factor I-A regulates diverse reactive astrocyte responses after CNS injury. J Clin Invest. (2019) 129(10):4408–18. doi: 10.1172/JCI127492
118. Yang R, Gautam A, Getnet D, Daigle BJ, Miller S, Misganaw B, Dean KR, Kumar R, Muhie S, Wang K, Lee I, Abu-Amara D, Flory JD, Consortium PSB, Hood L, Wolkowitz OM, Mellon SH, Doyle FJ, 3rd, Yehuda R, Marmar CR, Ressler KJ, Hammamieh R, Jett M Epigenetic biotypes of post-traumatic stress disorder in war-zone exposed veteran and active duty males. Mol Psychiatry. (2021) 26(8):4300–14. doi: 10.1038/s41380-020-00966-2
119. Somvanshi PR, Mellon SH, Yehuda R, Flory JD, Makotkine I, Bierer L, Marmar C, Jett M, Doyle FJ, 3rd. Role of enhanced glucocorticoid receptor sensitivity in inflammation in PTSD: insights from computational model for circadian-neuroendocrine-immune interactions. Am J Physiol: Endocrinol Metab. (2020) 319(1):E48–66. doi: 10.1152/ajpendo.00398.2019
120. Costa DL, Yetter N, DeSomer H. Intergenerational transmission of paternal trauma among US Civil War ex-POWs. Proc Natl Acad Sci U S A. (2018) 115(44):11215–20. doi: 10.1073/pnas.1803630115
121. Yehuda R, Daskalakis NP, Bierer LM, Bader HN, Klengel T, Holsboer F, et al. Holocaust exposure induced intergenerational effects on FKBP5 methylation. Biol Psychiatry. (2016) 80(5):372–80. doi: 10.1016/j.biopsych.2015.08.005
122. Yehuda R, Bell A, Bierer LM, Schmeidler J. Maternal, not paternal, PTSD is related to increased risk for PTSD in offspring of Holocaust survivors. J Psychiatr Res. (2008) 42(13):1104–11. doi: 10.1016/j.jpsychires.2008.01.002
123. Yehuda R, Blair W, Labinsky E, Bierer LM. Effects of parental PTSD on the cortisol response to dexamethasone administration in their adult offspring. Am J Psychiatry. (2007) 164(1):163–6. doi: 10.1176/ajp.2007.164.1.163
124. Yehuda R, Engel SM, Brand SR, Seckl J, Marcus SM, Berkowitz GS. Transgenerational effects of posttraumatic stress disorder in babies of mothers exposed to the World Trade Center attacks during pregnancy. J Clin Endocrinol Metab. (2005) 90(7):4115–8. doi: 10.1210/jc.2005-0550
125. Yehuda R, Halligan SL, Bierer LM. Cortisol levels in adult offspring of Holocaust survivors: relation to PTSD symptom severity in the parent and child. Psychoneuroendocrinology. (2002) 27(1):171–80. doi: 10.1016/S0306-4530(01)00043-9
126. Yehuda R, Teicher MH, Seckl JR, Grossman RA, Morris A, Bierer LM. Parental posttraumatic stress disorder as a vulnerability factor for low cortisol trait in offspring of holocaust survivors. Arch Gen Psychiatry. (2007) 64(9):1040–8. doi: 10.1001/archpsyc.64.9.1040
127. Yehuda R, Schmeidler J, Wainberg M, Binder-Brynes K, Duvdevani T. Vulnerability to posttraumatic stress disorder in adult offspring of holocaust survivors. Am J Psychiatry. (1998) 155(9):1163–71. doi: 10.1176/ajp.155.9.1163
128. Yehuda R, Schmeidler J, Giller EL, Siever LJ, Binder-Brynes K. Relationship between posttraumatic stress disorder characteristics of holocaust survivors and their adult offspring. Am J Psychiatry. (1998) 155(6):841–3. doi: 10.1176/ajp.155.6.841
129. Solomon Z, Kotler M, Mikulincer M. Combat-related posttraumatic stress disorder among second-generation Holocaust survivors: preliminary findings. Am J Psychiatry. (1988) 145(7):865–8. doi: 10.1176/ajp.145.7.865
130. Mulligan CJ, D’Errico NC, Stees J, Hughes DA. Methylation changes at NR3C1 in newborns associate with maternal prenatal stress exposure and newborn birth weight. Epigenetics. (2012) 7(8):853–7. doi: 10.4161/epi.21180
131. Kaati G, Bygren LO, Edvinsson S. Cardiovascular and diabetes mortality determined by nutrition during parents’ and grandparents’ slow growth period. Eur J Hum Genet. (2002) 10(11):682–8. doi: 10.1038/sj.ejhg.5200859
132. Bygren LO, Kaati G, Edvinsson S. Longevity determined by paternal ancestor’s nutrition during their slow growth period. Acta Biotheor. (2001) 49(1):53–9. doi: 10.1023/A:1010241825519
133. Lehrner A, Bierer LM, Passarelli V, Pratchett LC, Flory JD, Bader HN, Harris IR, Bedi A, Daskalakis NP, Makotkine I, Yehuda R. Maternal PTSD associates with greater glucocorticoid sensitivity in offspring of Holocaust survivors. Psychoneuroendocrinology. (2014) 40:213–20. doi: 10.1016/j.psyneuen.2013.11.019
134. Schwartz S, Dohrenwend BP, Levav I. Nongenetic familial transmission of psychiatric disorders? Evidence from children of Holocaust survivors. J Health Soc Behav. (1994) 35(4):385–402. doi: 10.2307/2137216
135. Oberlander TF, Weinberg J, Papsdorf M, Grunau R, Misri S, Devlin AM. Prenatal exposure to maternal depression, neonatal methylation of human glucocorticoid receptor gene (NR3C1) and infant cortisol stress responses. Epigenetics. (2008) 3(2):97–106. doi: 10.4161/epi.3.2.6034
136. Youssef NA, Lockwood L, Su S, Hao G, Rutten BPF. The effects of trauma, with or without PTSD, on the transgenerational DNA methylation alterations in human offsprings. Brain Sci. (2018) 8(5):83. doi: 10.3390/brainsci8050083
137. van IMH, Bakermans-Kranenburg MJ, Sagi-Schwartz A. Are children of Holocaust survivors less well-adapted? A meta-analytic investigation of secondary traumatization. J Trauma Stress. (2003) 16(5):459–69. doi: 10.1023/A:1025706427300
138. Vizi ES, Elenkov IJ. Nonsynaptic noradrenaline release in neuro-immune responses. Acta Biol Hung. (2002) 53(1-2):229–44. doi: 10.1556/ABiol.53.2002.1-2.21
Keywords: epigenetics, war, conflict, stress, PTSD, inherited epigenetics, trauma, healthy soldier effect
Citation: Raza Z, Hussain SF, Foster VS, Wall J, Coffey PJ, Martin JF and Gomes RSM (2023) Exposure to war and conflict: The individual and inherited epigenetic effects on health, with a focus on post-traumatic stress disorder. Front. Epidemiol. 3:1066158. doi: 10.3389/fepid.2023.1066158
Received: 10 October 2022; Accepted: 3 January 2023;
Published: 16 February 2023.
Edited by:
Eliana Mattos Lacerda, University of London, United KingdomReviewed by:
Marcela Tamayo-Ortiz, Instituto Mexicano del Seguro Social, MexicoLauren Godier-McBard, Anglia Ruskin University, United Kingdom
© 2023 Raza, Hussain, Foster, Wall, Coffey, Martin and Gomes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Renata S. M. Gomes cmVuYXRhLmdvbWVzQGJyYXZvdmljdG9yLm9yZw==
†These authors have contributed equally to this work and share first authorship
Specialty Section: This article was submitted to Occupational and Environmental Epidemiology, a section of the journal Frontiers in Epidemiology