 Veronika Szentes
Veronika Szentes Mária Gazdag
Mária Gazdag István Szokodi
István Szokodi Csaba A. Dézsi
Csaba A. Dézsi- 1Department of Cardiology, Petz Aladár County Teaching Hospital, Győr, Hungary
- 2Gedeon Richter Plc., Budapest, Hungary
- 3Heart Institute, Medical School, and Szentágothai Research Centre, University of Pécs, Pécs, Hungary
The chemokine receptor CXCR3 and associated CXC chemokines have been extensively investigated in several inflammatory and autoimmune diseases as well as in tumor development. Recent studies have indicated the role of these chemokines also in cardiovascular diseases. We aimed to present current knowledge regarding the role of CXCR3-binding chemokines in the pathogenesis of atherosclerosis and during acute myocardial infarction.
Introduction
Atherosclerosis is a chronic inflammatory disease, with immune cells and their effector molecules initiating and maintaining the progression of atherosclerotic lesion formation, accompanying and also precipitating acute coronary events and the following reparatory processes (1, 2). Chemotactic cytokines, or so-called chemokines have been shown to facilitate leukocyte migration during inflammatory responses to various stimuli, including their recruitment to the sites of atherosclerotic lesions (3).
Several chemokines have been associated with cardiovascular inflammatory changes. Chemokines CCL2, CCL5, CCL20, CXCL1, MIF (migration inhibitory factor), and CX3CL1 play a role in monocyte mobilization and recruitment (4). Monocyte binding to endothelial cells and their diapedesis into the subendothelial space is promoted by chemokine heterodimers CXCL4-CCL5. CXCL4 also affects monocyte differentiation into M4 macrophages, predominantly present in the adventitia and intima (5). Recruitment and survival of neutrophils is facilitated by CCL2, CCL3, CCL5, and CXCL1; (4) they also interact with CXCL4 (6) and CXCL12 (7).
Activated T lymphocytes (primarily Th1 cells) accumulate early and abundantly in the atherosclerotic lesions and are present in the plaques at all stages (3, 8). The Th1 cells recruited to the lesion recognize LDL as antigen and produce proinflammatory mediators such as interferon-gamma (IFN-γ) and tumor necrosis factor (TNF) (3, 8, 9). IFN-γ is the major proatherogenic cytokine, promoting local expression of adhesion molecules, cytokines and chemokines such as CXCL9, CXCL10, and CXCL11 and their main receptor CXCR3 by macrophages and endothelial cells (10). Chemokine signaling through CXCR3 facilitates recruitment and selective homing of active Th1 cells to the site of plaque development or rupture (Figure 1) (10–12).
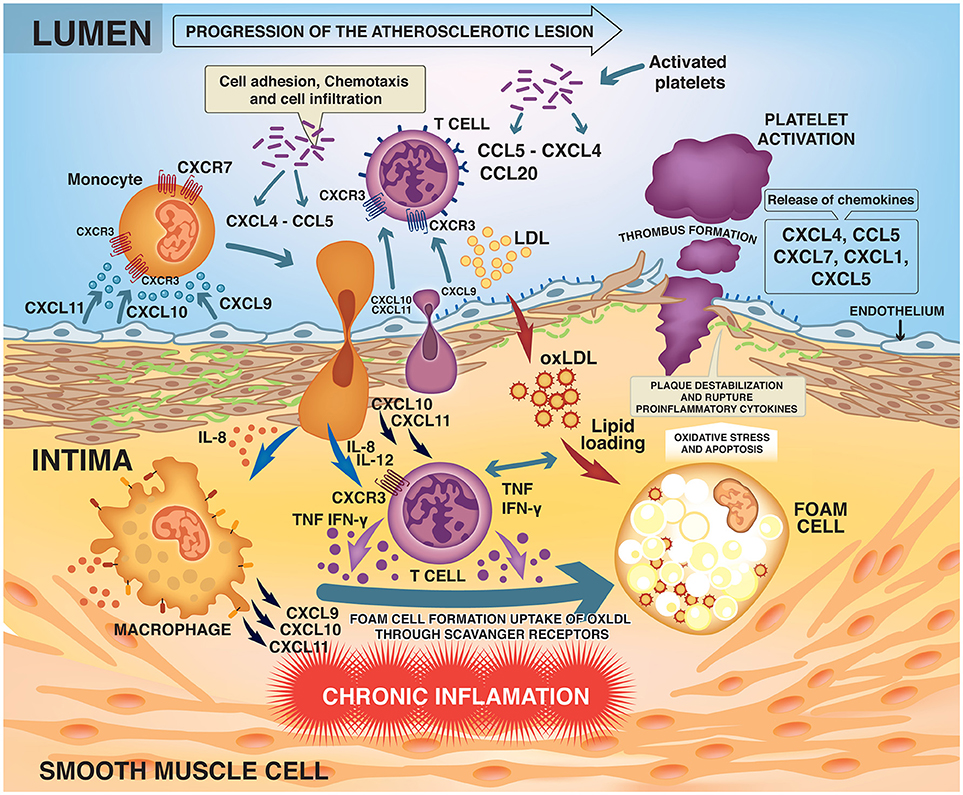
Figure 1. Development and progression of the atherosclerotic lesion.
The present review focuses on the role of the IFN-γ inducible chemokines and their receptor CXCR3 in the development of atherosclerosis and consequent coronary artery disease. Possible clinical implications of the presented findings are not entirely clear, but the currently available clinical studies suggest that this might be a promising area of intervention in the future of cardiovascular therapy and prevention (13).
Biased Signaling Through CXCR3
CXCR3 is a 7-transmembrane spanning (7-TMS) G-protein-coupled cell surface receptor that allows functional selectivity on tissue, receptor as well as ligand levels (6). It binds three inflammatory chemokines CXCL9, CXCL10, and CXCL11 (14, 15). It was also shown to weakly bind CXCL4 (platelet factor 4), with questionable in vivo significance (16). CXCR3 has three alternative splice variants: CXCR3A, CXCR3B, and CXCR3Alt that activate different intracellular signaling pathways, depending also on the ligand they bind (14). For example, Gαi heterotrimeric G protein activation and β-arrestin 1 and 2 recruitment was shown after stimulation with CXCL10 and CXCL11 on CXCR3A, however on CXCR3B it was shown only after stimulation with CXCL11 in high doses and was not detectable on CXCR3Alt. ERK1/2 phosphorylation and receptor internalization occurred on all three variants after stimulation, its intensity and signal duration depending on the chemokine ligand and splice variant assessed (14).
Different chemokines binding to CXCR3 appear to have slightly different roles in T cell trafficking. CXCL10 is abundantly expressed by all atheroma-associated cells such as T cells and monocytes and is supposed to facilitate T cell retention within the lesion (15, 17). CXCL11 interacts with CXCR3 with higher affinity and is a stronger agonist, demonstrated by its ability to mobilize intracellular calcium and also chemotactic migration of CXCR3+ cells. It is not active on resting or naïve T cells suggesting that CXCL11 does not play a role under normal conditions only during IL-2 stimulated T cell response (17, 18). CXCL11 was shown to be the physiologic inducer of CXCR3 down-regulation on the cellular surface after T cell contact with IFN-activated endothelial cells (19). This might serve as an arrest signal for the activated T cells and lead to restraining inflammatory responses (8). Besides CXCR3, CXCL11 also binds to receptor CXCR7 (ACKR3), which may also be a possible regulation point for CXCR3-mediated responses (16, 20). CXCL11 also has an antagonistic effect on CCR5, counteracting its inflammatory activities in leukocyte activation (21).
Biased signaling on CXCR3 results in different effect of its ligands during inflammatory events. It seems that CXCL9 and CXCL10 promote inflammation through inducing T cell polarization into Th1/Th17 cells, while CXCL11 drives the development of regulatory T cells (Treg) cells which play a role in restraining inflammation (22). Based on the above, CXCR3 may be hypothesized to play a dual role by mediating both proinflammatory and anti-inflammatory pathways.
CXCR3 Binding Chemokines in Atheroma Development
Experimental data demonstrated that targeted deletion or pharmacological inhibition of CXCR3 results in reduced plaque formation, which is accompanied by reduced recruitment of Th1 cells and increased migration of regulatory T-lymphocytes to lesions in apoE–/– mice (23, 24). In line, Apoe–/– /Cxcl10–/– mice showed reduced atherogenesis with enhanced numbers and activity of Treg cells (25). Moreover, antibody-mediated CXCL10 inhibition resulted in a more stable plaque phenotype in a vulnerable plaque mouse model (26).
High levels of IFN-γ induced chemokines CXCL9, CXCL10, and CXCL11 can be detected in human atheromas throughout all stages of plaque development (7). Niki et al. found elevated CLXCL10 levels to be associated with coronary atherosclerosis (27), while Segers et al. revealed a close correlation between high local concentrations of CXCL10 and unstable plaque characteristics by analyzing human carotid plaque specimens (26). CXCL4 and CXCL12 were also detected within atherosclerotic lesions (7, 28). CXCL12 was suggested to mediate anti-inflammatory action through neutrophil cells (7). CXCL4 is produced by platelets and plays a role in T cell-platelet interactions (29). Its levels were found to be correlated with the histological and clinical severity of atherosclerosis (28).
CXCR3 Binding Chemokines in Angina Pectoris
There is an increased systemic inflammatory activity present in patients with coronary artery disease, characterized by an increased proportion of IFN-γ positive Th1 lymphocytes. In patients with stable angina pectoris, enhanced systemic expression of CXCL9, CXCL10, and CXCR3 can be observed. Interestingly, lower levels of these chemokines and CXCR3 were found in the peripheral cells of patients with acute coronary syndrome, which indicates a sequestering of circulating CXCR positive cells from blood to the site of infarction via an intense in situ release of these chemokines (10, 11). Plasma levels of CXCL12 are decreased in patients with stable and unstable angina compared with healthy controls. CXCL12 thus might have a protective effect in unstable angina through stabilizing the atherosclerotic plaque (7).
Other anti-inflammatory molecules known for their protective effect in cardiovascular diseases were found to influence T cell trafficking through the chemokine system. Adiponectin was shown to inhibit CXCR3 ligand production in macrophages, while heparin competes for binding with CXCL9, CXCL10, and CXCL11 on endothelial cells (30, 31).
CXCR3 Binding Chemokines in Myocardial Infarction
It has been reported that CXCL10 and CXCR3 mRNA levels are up-regulated in the infarcted murine myocardium, with a marked increase in the number of CXCR3+/CD45+ leukocytes, CXCR3+/CD3+ T lymphocytes and CXCR3+ myofibroblasts (32, 33). Although CXCR3 is well-known to activate pro-inflammatory Th1 lymphocyte responses, deficiency of CXCR3 did not affect post-infarction cardiac remodeling (34). In contrast, Cxcl10–/– mice subjected to myocardial infarction were characterized by enhanced adverse ventricular remodeling, early expansion of the fibrotic scar, and increased neutrophil infiltration with marked reduction of recruitment of CXCR3 expressing leukocytes and T cells (33). Notably, CXCR3-independent proteoglycan signaling may mediate the anti-fibrotic effects of CXCL10 in the infarcted heart (34). In contrast to CXCL10, the role of CXCL9 and CXCL11 in infarct healing is not known.
Through receptor CXCR3, CXCL9, and CXCL10 promote T cell polarization into effector Th1/Th17 cells releasing pro-inflammatory mediators. Meanwhile, CXCL4 and CXCL11 promoted the differentiation of T cells into Treg1 cells, responsible for restraining the inflammatory response through IL-10, TGF-β and contact dependent pathways (22, 35, 36). Platelet surface expression of CXCR4 and CXCR7 receptors is elevated in acute coronary syndrome compared to stable angina. High CXCR7 levels are also associated with better improvement of left ventricular function after myocardial infarction. CXCR7 expression might contribute to regenerative function of platelets following acute coronary events (37).
Timely resolution of cardiac inflammatory responses plays a pivotal role in optimal tissue reparation (38, 39). Excessive, prolonged or inadequately contained inflammation can cause several complications such as cardiac rupture or dilatative ventricular remodeling and may lead to impaired cardiac function. Activation of pro-apoptotic pathways can cause unnecessary loss of cardiomyocytes and the extension of the inflammation to the non-infarcted area results in enhanced fibrosis and increased infarct size (38–40). Blockade of leukocyte related inflammatory mediators was shown to cause a marked reduction in infarct size and prevented the extension of ischemic cardiomyocyte injury following reperfusion in experimental studies (38).
During myocardial infarction, the dual role of CXCR3 in inflammatory processes might enable CXCR3+ cells to set off an appropriately rapid and robust inflammatory response in the beginning (1). Also, it might contribute to the timely resolution of symptoms by restraining the inflammation afterwards. It seems that the activation of the plaque rather than the degree of coronary stenosis precipitates ischemia and infarction. Endothelial erosion or plaque rupture was found to be responsible for the majority of coronary thrombotic events (1, 9) (Box 1).
Box 1. Chemokines, CXCR3, and CXCL9 (Mig), CXCL10 (IP-10), and CXCL11 (I-TAC)
Chemokines
Chemokines are a structurally related superfamily of more than 50 small signaling proteins (cytokines) that were originally named after their chemotactic effect on leukocytes. They all share a conserved cysteine motif in the mature sequence of the chemokines. Based on the number and arrangement of the N-terminal cysteine residues in this motif, chemokines can be divided into four families (CXC, CC, C, and CX3C) (41, 42). Besides regulating leukocyte migration and degranulation, chemokines take active part in a number of complex processes like angiogenesis or hematopoiesis and were found to participate in several diseases related to the immune system such as atherogenesis, multiple sclerosis, asthma, HIV-infection or cancer (7, 18, 41–43).
Chemokines bind to 7-TMS G-protein-coupled cell surface receptors. The activation of chemokine receptors can be followed by one of several signaling pathways, including inhibition of adenylate cyclase, activation of phosphoinositol 3-kinase, phospholipase C and D, protein kinase C and A, inositol triphosphate generation and transient calcium influx (44). More than 20 chemokine receptors have been discovered so far; their names mirror the nomenclature of chemokine family names (CXCR1-7, CCR1-10, etc.) (45).
CXCR3
CXCR3 is a chemokine receptor expressed by activated T lymphocytes, including CD4+ T helper 1 (Th1) cells, CD8+ cytotoxic T lymphocytes (CTL), and CD4+ and CD8+ memory T cells, as well as monocytes, M1 macrophages, natural killer (NK) cells, leukemic B-cells, eosinophils, mast cells, plasmocytoid dendritic cells, endothelial cells (ECs) and vascular smooth muscle cells (SMCs) (44, 46). Up-regulation of CXCR3 has been described in multiple sclerosis and transplant rejection (47). CXCR3 is also expressed by various tumor cells (48).
CXCL9 (Mig), CXCL10 (IP-10), and CXCL11 (I-TAC)
These three non-ELR chemokines are on the same branch of the phylogenetic tree and consequently share common characteristics. Their main receptor is CXCR3, but they can also act as antagonists for CCR3. They are constitutively expressed at low levels in normal tissues including thymus and spleen, where they are probably involved in activated (CXCR3+) T cell trafficking. Their expression is strongly induced by IFN-γ and they are produced in a wide variety of cell types, including atheroma-associated endothelial cells and macrophages (7, 17, 41, 44).
Clinical Experience With CXCR3 Binding Chemokines in Ischemic Heart Disease
The clinical relevance of CXCR3 binding chemokines in ischemic heart disease is not fully understood. As summarized in Table 1, clinical studies to date aimed to find an association between plasma levels of different cytokines and several aspects of coronary events. It seems that complex patterns rather than individual changes in plasma chemokine levels might be associated with cardiovascular risk (50, 53, 59). Ardigo et al. found that when using a combined multimarker chemokine model (including CXCL10), serum concentrations of the chemokines were differentially regulated in individuals with clinical coronary artery disease compared with subjects with no such history. Their findings suggest that chemokine profile models using multiple chemokines may represent a strong signal of coronary artery disease with even higher specificity than traditional risk factors (49).
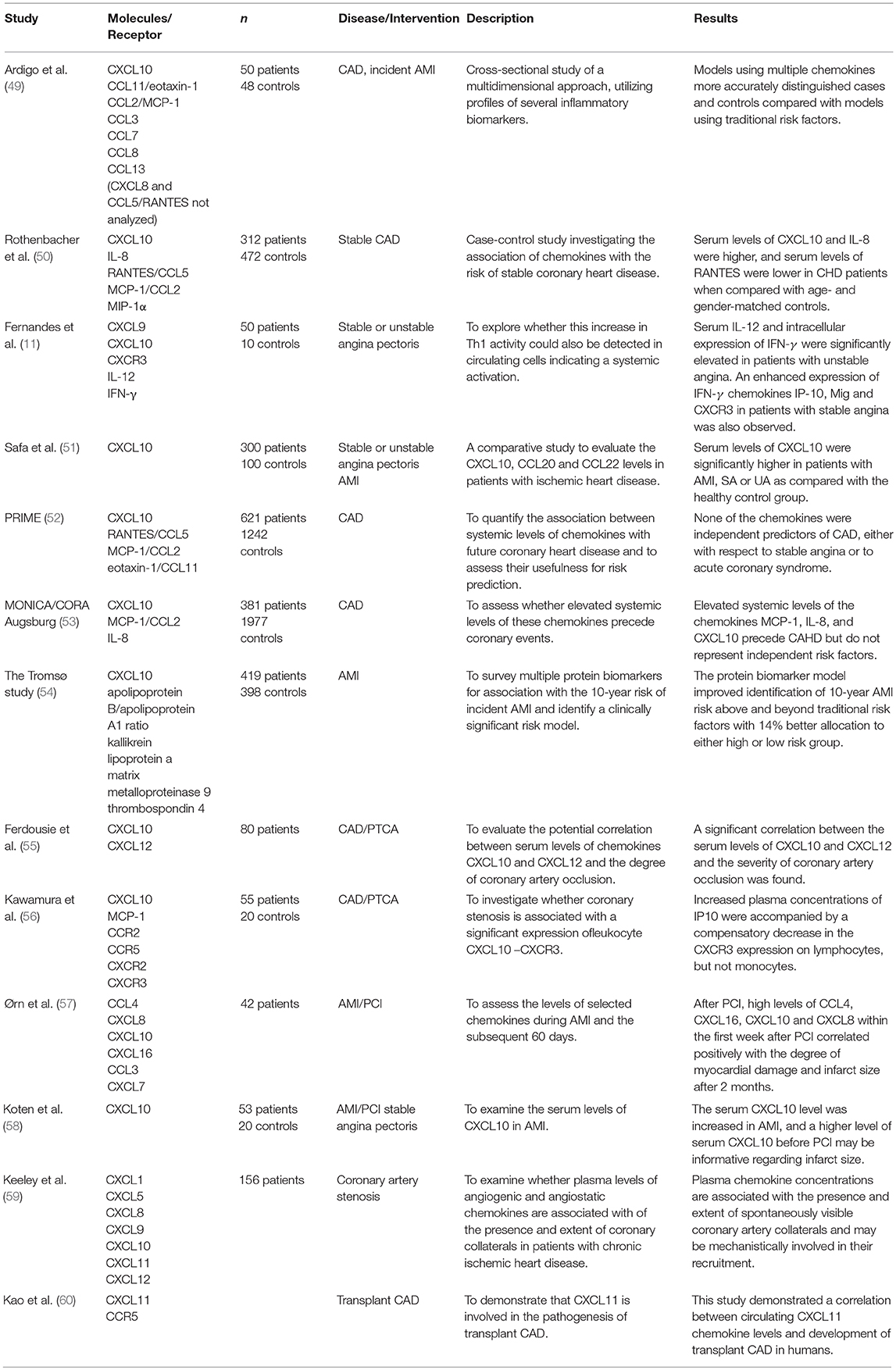
Table 1. Clinical studies with CXCR3 binding chemokines in coronary artery disease.
In a large case-control study of 312 patients with coronary heart disease and 472 controls, a significant association of increased serum CXCL10 was found with the risk of coronary heart disease. Higher CXCL10 levels were also found to be independently correlated with established laboratory risk markers of coronary heart disease such as acute-phase proteins and inflammatory cytokines (50).
In patients with stable angina pectoris, Fernandes et al. found significantly higher levels of CXCL9, CXCL10, and CXCR3 compared to healthy controls (11). In patients with unstable angina, increased inflammatory activity was confirmed compared to stable angina patients by elevated high sensitivity C-reactive protein and serum amyloid A protein levels. However, the levels of CXCL9, CXCL10, and CXCR3 remained low in patients with unstable angina, comparable to the control group and significantly lower than in patients with stable angina. The authors suggested local release and intense uptake of these molecules by circulating leukocytes migrating to the site of active inflammation, which would explain their lower levels in the peripheral blood. Blood samples were drawn within 48 h of the index consultation of the unstable patients, and it was hypothesized that samples taken in a different time frame might capture serum elevations in CXCR3 and related chemokines (11).
Safa et al. (51) in a larger study in 260 patients and 100 healthy controls managed to capture elevated CXCL10 levels in patients with unstable angina. In this study the serum levels of CXCL10 were measured at the time of admission and were found to be elevated both in patients with stable and unstable angina pectoris. CXCL10 was also elevated in acute myocardial infarction, measured 3–5 days after admission. The study also confirmed the correlation of tradition risk factor with CXCL10, as mean serum levels of CXCL10 in patients with hypertension, dyslipidemia, obesity, diabetes and smoking were significantly higher as compared to the control group (51).
While elevated serum CXCL10 was found to be significantly associated with increased risk of coronary heart disease, it was not an independent risk factor for future coronary events in population-based case-control studies (52, 53). CXCL10 modestly correlated with traditional cardiovascular risk factors in the PRIME study (49). Age was found to be the strongest positive confounder in the MONICA/CORA Augsburg cohort, with the levels of circulating immune mediators increasing with age (52). The investigators of the Tromsø Study found that higher CXCL10 levels were protective for women when assessing the 10-year risk of incident myocardial infarction. In the multivariable model, the composite risk of 6 biomarkers including CXCL10 improved the traditional risk factor model by 14% (54).
A significant correlation was found between elevated serum CXCL10 and CXCL12 levels and the severity of coronary artery occlusion in patients with coronary heart disease who underwent PTCA (55). In patients with restenosis after PTCA, decreased concentrations of CXCL10 were followed by the decrease of CXCR3 expression on lymphocytes but not monocytes, suggesting a possible role of CXCL10 signaling on monocytes in neointimal hyperplasia in patients with restenosis (56).
CXCL8, CXCL10, and CXCL16 were found to be correlated with maximum troponin T levels, infarct size and impaired myocardial function assessed by cardiac magnetic resonance in patients after successful PCI (57). Serum CXCL10 level before PCI also proved to be an independent predictor of cumulative CK release and was negatively correlated with infarct size, as indicated by peak CK and CK-MB enzymes (58).
Better clinical outcome was found to be associated with recruitment of coronary collaterals (61). This form of vascular remodeling was shown to be accompanied by alterations in chemokine levels (59). Higher levels of angiogenic ligands CXCL5, CXCL8, and CXCL12 indicate the presence of collaterals, while the concentration of the angiostatic CXCL11 was associated with their absence. The higher extent of collateralization was associated with increased CXCL1 and decreased CXCL9, CXCL10, and CXCL11 (59).
Several chemokines have been linked to the development of acute transplant rejection episodes and transplant coronary artery disease in animals and also in human studies (60). Following heart transplantation, elevated CXCL11 levels have shown an association with the development of severe transplant coronary artery disease (60). CXCR3 ligands have also been studied in patients with left ventricular dysfunction and heart failure (62–64). Circulating levels of CXCL9, CXCL10, and CXCL11 were increased in subclinical as well as symptomatic left ventricular dysfunction, reaching statistical significance only in symptomatic patients (62). Addition of these CXCR3 ligands to established risk factors significantly improved the risk prediction models for left ventricular dysfunction (63). In a pilot study by Altara et al. levels of CXCL10 positively correlated with the severity of heart failure, especially in patients with advanced heart failure (64). Also, higher systemic levels of CXCL10 have been demonstrated to be independent risk factors for ischemic stroke (52).
Conclusions
The chemokine network specifically directs the trafficking of immune cells in homeostasis and during inflammation. Excessive or inappropriate chemokine expression can lead to unnecessary leukocyte recruitment typical for autoimmune or allergic diseases. Chemokines have been extensively studied in diseases associated with T cell mediated inflammatory responses like multiple sclerosis, asthma bronchiale, AIDS and also in patients with transplant rejection (47, 60, 65).
Inflammatory processes in ischemic heart disease involve intense chemokine signaling from the forming of the atherosclerotic plaque and plaque destabilization to all phases of acute coronary events and infarct healing (36). IFN-γ inducible chemokines CXCL9, CXCL10, and CXCL11 attract activated T cells through CXCR3 receptor to the site of infarction. Modulation of their action might prevent the excessive recruitment of leukocytes to sites of inflammation and consequently influence the clinical outcome of the disease (47).
CXCR3 binding chemokines might be promising biomarkers for the risk assessment of coronary heart disease. Chemokine levels however have a short half-life and may have high intraindividual variability; (52) this results in difficulties in estimating the best sampling time and may generate conflicting clinical results.
CXCL10 is the most extensively studied of the three chemokines in the clinical setting of ischemic heart disease; less is known about the role of CXCL9 and CXCL11. New clinical studies are needed to fill in the gaps and properly map the role of alterations in chemokine levels in coronary artery disease and during acute coronary events.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Funding
This work was supported by a grant from the National Research, Development and Innovation Fund of Hungary (K120536).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
1. Hansson GK. Inflammation, atherosclerosis and coronary artery disease. N Engl J Med. (2005) 352:1685–95. doi: 10.1056/NEJMra043430
2. Santos-Gallego CG, Picatoste B, Badimón JJ. Pathophysiology of acute coronary syndrome. Curr Atheroscler Rep. (2014) 16:401. doi: 10.1007/s11883-014-0401-9
3. Li J, Ley K. Lymphocyte migration into atherosclerotic plaque. Arterioscler Thromb Vasc Biol. (2015) 35:40–49. doi: 10.1161/ATVBAHA.114.303227
4. Zernecke A, Weber C. Chemokines in atherosclerosis. proceedings resumed. Arterioscler Thromb Vasc Biol. (2014) 34:742–750. doi: 10.1161/ATVBAHA.113.301655
5. Domschke G, Gleissner CA. CXCL4-induced macrophages in human atherosclerosis. Cytokine (2017). doi: 10.1016/j.cyto.2017.08.021. [Epub ahead of print].
6. Pilatova K, Greplov K, Demlova R, Bencsikova B, Klement GL, Zdrazilova-Dubska L. Role of platelet chemokines, PF-4 and CTAP-III, in cancer biology. J Hematol Oncol. (2013) 6:42. doi: 10.1186/1756-8722-6-42
7. Zernecke A, Shagdarsuren E, Weber C. Chemokines in atherosclerosis. an update. Arterioscler Thromb Vasc Biol. (2008) 28:1897–908. doi: 10.1161/ATVBAHA.107.161174
8. Mach F, Sauty A, Iarossi AS, Sukhova GK, Neote K, Libby P, et al. Differential expression of three T lymphocyte-activating CXC chemokines by human atheroma-associated cells. J Clin Invest. (1999) 104:1041–50. doi: 10.1172/JCI6993
9. Gistera A, Hansson GK. The immunology of atherosclerosis. Nat Rev Nephrol. (2017) 13:368–80. doi: 10.1038/nrneph.2017.51
10. de Oliviera RTD, Mamoni RL, Souza JR, Fernandes JL, Rios FJ, Gidlund M, et al. Differential expression of cytokines, chemokines and chemokine receptors in patients with coronary artery disease. Int J Cardiol. (2009) 136:17–26. doi: 10.1016/j.ijcard.2008.04.009
11. Fernandes JL, Mamoni RL, Orford JL, Garcia C, Selwyn AP, Coelho OR, et al. Increased Th1 activity in patients with coronary artery disease. Cytokine (2004) 26:131–7. doi: 10.1016/j.cyto.2004.01.007
12. Qin S, Rottman JB, Myers P, Kassam N, Weinblatt M, Loetscher M, et al. The chemokine receptors CXCR3 and CCR5 mark subsets of T cells associated with certain inflammatory reactions. J Clin Invest. (1998) 101:746–54. doi: 10.1172/JCI1422
13. von Hundelshausen P, Schmitt MMN. Platelets and their chemokines in atherosclerosis—clinical applications. Front Physiol. (2014) 5:294. doi: 10.3389/fphys.2014.00294
14. Berchiche YA, Sakmar TP. CXC chemokine receptor 3 alternative splice variants selectively activate different signaling pathways. Mol Pharmacol. (2016) 90:483–95. doi: 10.1124/mol.116.105502
15. Metzemaekers M, Vanheule V, Janssens R, Struyf S, Proost P. Overview of the mechanisms that may contribute to the non-redundant activities of interferon-inducible cxc chemokine receptor 3 ligands. Front Immunol. (2018) 8:1970. doi: 10.3389/fimmu.2017.01970
16. Bachelerie F, Ben-Baruch A, Burkhardt AM, Combadiere C, Farber JM, Graham GJ, et al. International Union of Pharmacology. LXXXIX. update on the extended family of chemokine receptors and introducing a new nomenclature for atypical chemokine receptors. Pharmacol Rev. (2014) 66:1–79. doi: 10.1124/pr.113.007724
17. Booth V, Clark-Lewis I, Sykes BD. NMR structure of CXCR3 binding chemokine CXCL11 (ITAC). Protein Sci. (2004) 13:2022–8. doi: 10.1110/ps.04791404
18. Cole KE, Strick CA, Paradis TJ, Ogborne KT, Loetscher M, Gladue RP, et al. Interferon–inducible T cell alpha chemoattractant (I-TAC): a novel non-ELR CXC chemokine with potent activity on activated T cells through selective high affinity binding to CXCR3. J Exp Med. (1998) 187:2009–21. doi: 10.1084/jem.187.12.2009
19. Colvin RA, Campanella GS, Sun J, Luster AD. Intracellular domains of CXCR3 that mediate CXCL9, CXCL10, and CXCL11 function. J Biol Chem. (2004) 279:30219–27. doi: 10.1074/jbc.M403595200
20. Chatterjee M, Rath D, Gawaz M. Role of chemokine receptors CXCR4 and CXCR7 for platelet function. Biochem Soc Trans. (2015) 43:720–6. doi: 10.1042/BST20150113
21. Petkovic V, Moghini C, Paoletti S, Uguccioni M, Gerber B. I-TAC/CXCL11 is a natural antagonist for CCR5. J Leukoc Biol. (2004) 76:701–8. doi: 10.1189/jlb.1103570
22. Karin N, Wildbaum G, Thelen M. Biased signaling pathways via CXCR3 control the development and function of CD4+ T cell subsets. J Leukoc Biol. (2016) 99:857–62. doi: 10.1189/jlb.2MR0915-441R
23. Veillard NR, Steffens S, Pelli G, Lu B, Kwak BR, Gerard C, et al. Differential influence of chemokine receptors CCR2 and CXCR3 in development of atherosclerosis in vivo. Circulation (2005) 112:870–878. doi: 10.1161/CIRCULATIONAHA.104.520718
24. van Wanrooij EJ, de Jager SC, van Es T, de Vos P, Birch HL, Owen DA, et al. CXCR3 antagonist NBI-74330 attenuates atherosclerotic plaque formation in LDL receptor –deficient mice. Arterioscler Thromb Vasc Biol. (2008) 28:251–7. doi: 10.1161/ATVBAHA.107.147827
25. Heller EA, Liu E, Tager AM, Yuan Q, Lin AY, Ahluwalia N, et al. Chemokine CXCL10 promotes atherogenesis by modulating the local balance of effector and regulatory T cells. Circulation (2006) 113:2301–12. doi: 10.1161/CIRCULATIONAHA.105.605121
26. Segers D, Lipton JA, Leenen PJ, Cheng C, Tempel D, Pasterkamp G, et al. Atherosclerotic plaque stability is affected by the chemokine CXCL10 in both mice and humans. Int J Inflam. (2011) 2011:936109. doi: 10.4061/2011/936109
27. Niki T, Soeki T, Yamaguchi K, Taketani Y, Yagi S, Iwase T, et al. Elevated concentration of interferon-inducible protein of 10 kD (IP-10) is associated with coronary atherosclerosis. Int Heart J. (2015) 56:269–72. doi: 10.1536/ihj.14-300
28. Pitsilos S, Hunt J, Mohler ER, Prabhakar AM, Poncz M, Dawicki J, et al. Platelet factor 4 localization in carotid atherosclerotic plaques: correlation with clinical parameters. Thromb.Haemost. (2003) 90:1112–20. doi: 10.1160/TH03-02-0069
29. Li N. CD4+ T cells in atherosclerosis: regulation by platelets. Thromb Haemost. (2013) 109:980–90. doi: 10.1160/TH12-11-0819
30. Okamoto Y, Folco EJ, Minami M, Wara AK, Feinberg MW, Sukhova GK, et al. Adiponectin inhibits the production of CXC receptor 3 chemokine ligands in macrophages and reduces T-lymphocyte recruitment in atherogenesis. Circ Res. (2008) 102:218–25. doi: 10.1161/CIRCRESAHA.107.164988
31. Ranjbaran H, Wang Y, Manes TD, Yakimov AO, Akhtar S, Kluger MS, et al. Heparin displaces interferon-γ-inducible chemokines (IP-10, I-TAC, and Mig) sequestered in the vasculature and inhibits the transendothelial migration and arterial recruitment of T cells. Circulation (2006) 114:293–1300. doi: 10.1161/CIRCULATIONAHA.106.631457
32. Dewald O, Frangogiannis NG, Zoerlein M, Duerr GD, Klemm C, Knuefermann P, et al. Development of murine ischemic cardiomyopathy is associated with a transient inflammatory reaction and depends on reactive oxygen species. Proc Natl Acad Sci USA. (2003) 100:2700–5. doi: 10.1073/pnas.0438035100
33. Bujak M, Dobaczewski M, Gonzalez-Quesada C, Xia Y, Leucker T, Zymek P, et al. Induction of the CXC chemokine interferon-γ-inducible protein (IP)-10 regulates the reparative response following myocardial infarction. Circ Res. (2009) 105:973–83. doi: 10.1161/CIRCRESAHA.109.199471
34. Saxena A, Chen W, Su Y, Rai V, Uche OU, Li N, et al. Interleukin-1 induces pro-inflammatory leukocyte infiltration and regulates fibroblast phenotype in the infarcted myocardium. J Immunol. (2013) 191:4838–48. doi: 10.4049/jimmunol.1300725
35. Liu CY, Battaglia M, Lee SH, Sun QH, Aster RH, Visentin GP. Platelet factor 4 differentially modulates CD4+ CD25+ (regulatory) versus CD4+ CD25- (nonregulatory) T cells. J Immunol. (2005) 174:2680–6. doi: 10.4049/jimmunol.174.5.2680
36. Bonaventura A, Montecucco F, Dallegri F. Cellular recruitment in myocardial ischaemia/reperfusion injury. Eur J Clin Invest. (2016) 46:590–601. doi: 10.1111/eci.12633
37. Rath D, Chatterjee M, Borst O, Müller K, Stellos K, Mack AF, et al. Expression of stromal cell-derived factor-1 receptors CXCR4 and CXCR7 on circulating platelets of patients with acute coronary syndrome and association with left ventricular functional recovery. Eur. Heart J. (2014) 35:386–94. doi: 10.1093/eurheartj/eht448
38. Christia P, Frangogiannis NG. Targeting inflammatory pathways in myocardial infarction. Eur J Clin Invest. (2013) 43:986–95. doi: 10.1111/eci.12118
39. Frangogiannis NG. Regulation of the inflammatory response in cardiac repair. Circ Res. (2012) 110:159–73. doi: 10.1161/CIRCRESAHA.111.243162
40. Matsumori A, Furukawa Y, Hashimoto T, Yoshida A, Ono K, Shioi T, et al. Plasma levels of the monocyte chemotactic and activating factor/monocyte chemotactic protein-1 are elevated in patients with acute myocardial infarction. J Mol Cell Cardiol. (1997) 29:419–23. doi: 10.1006/jmcc.1996.0285
42. Singh AK, Arya RK, Trivedi AK, Sanyal S, Baral R, Dormond O, et al. Chemokine receptor trio: CXCR3, CXCR4 and CXCR7 crosstalk via CXCL11 and CXCL12. Cytokine Growth Factor Rev. (2013) 24:41–9. doi: 10.1016/j.cytogfr.2012.08.007
43. Mackay CR. Chemokines: immunology's high impact factors. Nat Immunol. (2001) 2:95–101. doi: 10.1038/84298
44. Sauty A, Colvin RA, Wagner L, Rochat S, Spertini F, Luster AD. CXCR3 internalization following T cell-endothelial cell contact: preferential role of IFN-inducible T cell α chemoattractant (CXCL11). J Immunol. (2001) 167:7084–93. doi: 10.4049/jimmunol.167.12.7084
45. Zlotnik A, Yoshie O. Chemokines: a new classification review system and their role in immunity. Immunity (2000) 12:121–7. doi: 10.1016/S1074-7613(00)80165-X
46. Altara R, Mallat Z, Booz GW, Zouein FA. The CXCL10/CXCR3 axis and cardiac inflammation: implications for immunotherapy to treat infectious and noninfectious diseases of the heart. J Immunol Res. (2016) 2016:4396368. doi: 10.1155/2016/4396368
47. Power CA, Proudfoot AE. The chemokine system: novel broad-spectrum therapeutic targets. Curr Opin Pharmacol. (2001) 1:417–24. doi: 10.1016/S1471-4892(01)00072-8
48. Miekus K, Jarocha D, Trzyna E, Majka M. Role of I-TAC-binding receptors CXCR3 and CXCR7 in proliferation, activation of intracellular signaling pathways and migration of various tumor cell lines. Folia Histochem Cytobiol. (2010) 48:104–111. doi: 10.2478/v10042-008-0091-7
49. Ardigo D, Assimes TL, Fortmann SP, Go AS, Hlatky M, Hytopoulos E, et al. Circulating chemokines accurately identify individuals with clinically significant atherosclerotic heart disease. Physiol Genomics (2007) 31:402–9. doi: 10.1152/physiolgenomics.00104.2007
50. Rothenbacher D, Müller-Scholze S, Herder C, Koenig W, Kolb H. Differential expression of chemokines, risk of stable coronary heart disease, and correlation with established cardiovascular risk markers. Arterioscler Thromb Vasc Biol. (2006) 26:194–9. doi: 10.1161/01.ATV.0000191633.52585.14
51. Safa A, Rashidinejad HR, Khalili M, Dabiri S, Nemati M, Mohammadi MM, et al. Higher circulating levels of chemokines CXCL10, CCL20 and CCL22 in patients with ischemic heart disease. Cytokine (2016) 83:147–57. doi: 10.1016/j.cyto.2016.04.006
52. Canouï-Poitrine F, Luc G, Mallat Z, Machez E, Bingham A, Ferrieres J, et al. Systemic chemokine levels, coronary heart disease, and ischemic stroke events: the PRIME study. Neurology (2011) 77:1165–73. doi: 10.1212/WNL.0b013e31822dc7c8
53. Herder C, Baumert J, Thorand B, Martin S, Löwel H, Kolb H. et al. Chemokines and incident coronary heart disease results from the MONICA/KORA augsburg case-cohort study, 1984–2002. Arterioscler Thromb Vasc Biol. (2006) 26:2147–52. doi: 10.1161/01.ATV.0000235691.84430.86
54. Wilsgaard T, Mathiesen EB, Patwardhan A, Rowe MW, Schirmer H, Løchen ML, et al. Clinically significant novel biomarkers for prediction of first ever myocardial infarction the tromsø study. Circ Cardiovasc Genet. (2015) 8:363–71. doi: 10.1161/CIRCGENETICS.113.000630
55. Tavakolian Ferdousie V, Mohammadi M, Hassanshahi G, Khorramdelazad H, Khanamani Falahati-Pour S, Mirzaei M, et al. Serum CXCL10 and CXCL12 chemokine levels are associated with the severity of coronary artery disease and coronary artery occlusion. Int J Cardiol. (2017) 233:23–28. doi: 10.1016/j.ijcard.2017.02.011
56. Kawamura A, Miura S, Fujino M, Nishikawa H, Matsuo Y, Tanigawa H. et al. CXCR3 chemokine receptor–plasma IP10 interaction in patients with coronary artery disease. Circ J. (2003) 67:851–4. doi: 10.1253/circj.67.851
57. Orn S, Breland UM, Mollnes TE, Manhenke C, Dickstein K, Aukrust P, et al. The chemokine network in relation to infarct size and left ventricular remodeling following acute myocardial infarction. Am J Cardiol. (2009) 104:1179–83. doi: 10.1016/j.amjcard.2009.06.028
58. Koten K, Hirohata S, Miyoshi T, Ogawa H, Usui S, Shinohata R, et al. Serum interferon-gamma-inducible protein 10 level was increased in myocardial infarction patients, and negatively correlated with infarct size. Clin Biochem. (2008) 41:30–7. doi: 10.1016/j.clinbiochem.2007.10.001
59. Keeley EC, Moorman JR, Liu L, Gimple LW, Lipson LC, Ragosta M, et al. Plasma chemokine levels are associated with the presence and extent of angiographic coronary collaterals in chronic ischemic heart disease. PLoS ONE (2011) 6:e21174. doi: 10.1371/journal.pone.0021174
60. Kao J, Kobashigawa J, Fishbein MC, MacLellan WR, Burdick MD, Belperio JA, et al. Elevated serum levels of the CXCR3 chemokine ITAC are associated with the development of transplant coronary artery disease. Circulation (2003) 107:1958–61. doi: 10.1161/01.CIR.0000069270.16498.75
61. Regieli JJ, Jukema JW, Nathoe HM, Zwinderman AH, Ng S, Grobbee DE, et al. Coronary collaterals improve prognosis in patients with ischemic heart disease. Int J Cardiol. (2009) 132:257–262. doi: 10.1016/j.ijcard.2007.11.100
62. Altara R, Gu YM, Struijker-Boudier HA, Thijs L, Staessen JA, Blankesteijn WM. Left ventricular dysfunction and CXCR3 ligands in hypertension: from animal experiments to a population-based pilot study. PLoS ONE (2015) 10:e0141394. doi: 10.1371/journal.pone.0141394
63. Altara R, Gu YM, Struijker-Boudier HA, Staessen JA, Blankesteijn WM. Circulating CXCL-9,−10 and−11 levels improve the discrimination of risk prediction models for left ventricular dysfunction. FASEB J. (2015) 29:46.2.
64. Altara R, Manca M, Hessel MH, Gu Y, van Vark LC, Akkerhuis KM, et al. CXCL10 is a circulating inflammatory marker in patients with advanced heart failure: a pilot study. J Cardiovasc Trans Res. (2016) 9:302–14. doi: 10.1007/s12265-016-9703-3
65. Zhao DX, Hu Y, Miller GG, Luster AD, Mitchell RN, Libby P. Differential expression of the IFN-γ-inducible CXCR3-binding chemokines, IFN-inducible protein 10, monokine induced by IFN, and IFN-inducible T cell α chemoattractant in human cardiac allografts: association with cardiac allograft vasculopathy and acute rejection. J Immunol. (2002) 169:1556–60. doi: 10.4049/jimmunol.169.3.1556
Keywords: inflammation, chemokine, I-TAC, atherosclerosis, coronary artery disease, myocardial infarction
Citation: Szentes V, Gazdag M, Szokodi I and Dézsi CA (2018) The Role of CXCR3 and Associated Chemokines in the Development of Atherosclerosis and During Myocardial Infarction. Front. Immunol. 9:1932. doi: 10.3389/fimmu.2018.01932
Received: 11 March 2018; Accepted: 06 August 2018;
Published: 27 August 2018.
Edited by:
Pietro Enea Lazzerini, University of Siena, ItalyReviewed by:
Jose Miguel Rodriguez Frade, Consejo Superior de Investigaciones Científicas (CSIC), SpainRaffaele Altara, Oslo University Hospital, Norway
Copyright © 2018 Szentes, Gazdag, Szokodi and Dézsi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Csaba A. Dézsi, ZGNzYTYyQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work