 Frederieke A. J. Gigase1,2*
Frederieke A. J. Gigase1,2* Anna Suleri1,2Elena Isaevska1,2
Anna Suleri1,2Elena Isaevska1,2 Anna-Sophie Rommel3
Anna-Sophie Rommel3 Myrthe G. B. M. Boekhorst4
Myrthe G. B. M. Boekhorst4 Olga Dmitrichenko3
Olga Dmitrichenko3 Hanan El Marroun1,5
Hanan El Marroun1,5 Eric A. P. Steegers6
Eric A. P. Steegers6 Manon H. J. Hillegers1
Manon H. J. Hillegers1 Ryan L. Muetzel1,7Whitney Lieb8Charlotte A. M. Cecil1,9,10Victor J. M. Pop4Michael Breen3Veerle Bergink3,8,11†Lot D. de Witte3,12,13†
Ryan L. Muetzel1,7Whitney Lieb8Charlotte A. M. Cecil1,9,10Victor J. M. Pop4Michael Breen3Veerle Bergink3,8,11†Lot D. de Witte3,12,13†- 1Department of Child and Adolescent Psychiatry, Erasmus University Medical Center, Rotterdam, Netherlands
- 2The Generation R Study Group, Erasmus University Medical Center, Rotterdam, Netherlands
- 3Department of Psychiatry, Icahn School of Medicine at Mount Sinai, New York, NY, United States
- 4Department of Medical and Clinical Psychology, Tilburg University, Tilburg, Netherlands
- 5Department of Psychology, Education and Child Studies, Erasmus School of Social and Behavioral Sciences, Erasmus University, Rotterdam, Netherlands
- 6Department of Obstetrics and Gynecology, Erasmus University Medical Center, Rotterdam, Netherlands
- 7Department of Radiology and Nuclear Medicine, Erasmus University Medical Center, Rotterdam, Netherlands
- 8Department of Obstetrics, Gynecology and Reproductive Science, Icahn School of Medicine at Mount Sinai, New York, NY, United States
- 9Department of Epidemiology, Erasmus University Medical Center, Rotterdam, Netherlands
- 10Department of Biomedical Data Sciences, Molecular Epidemiology, Leiden University Medical Center, Leiden, Netherlands
- 11Department of Psychiatry, Erasmus University Medical Center, Rotterdam, Netherlands
- 12Department of Human Genetics, Radboud University Medical Center, Nijmegen, Netherlands
- 13Department of Psychiatry, Radboud University Medical Center, Nijmegen, Netherlands
Introduction: Adaptations of the immune system throughout gestation have been proposed as important mechanisms regulating successful pregnancy. Dysregulation of the maternal immune system has been associated with adverse maternal and fetal outcomes. The design and interpretation of human biomarker studies require additional insights in the trajectories and drivers of peripheral immune markers.
Methods: The current study mapped maternal inflammatory markers (C-reactive protein (CRP), interleukin (IL)-1β, IL-6, IL-17A, IL-23, interferon-γ) during pregnancy and investigated the impact of demographic, environmental and genetic drivers on maternal inflammatory marker levels in four multi-ethnic and socio-economically diverse population-based cohorts with more than 12,000 pregnant participants. Additionally, pregnancy inflammatory markers were compared to pre-pregnancy levels.
Results: Cytokines showed a high correlation with each other, but not with CRP. Inflammatory marker levels showed high variability between individuals, yet high concordance within an individual over time during and pre-pregnancy. Pre-pregnancy body mass index (BMI) explained ~ 9.6% of the variance in CRP, but less than 1% of the variance in cytokines. The polygenic score of CRP was the best predictor of variance in CRP (14.1%). Gestational age and previously identified inflammation drivers, including tobacco use and parity, explained less than 1% of variance in both cytokines and CRP.
Discussion: Our findings corroborate differential underlying regulatory mechanisms of CRP and cytokines and are suggestive of an individual inflammatory marker baseline which is, in part, genetically driven.
Introduction
Adaptations of the immune system during pregnancy have fascinated the immunological field for decades. Immunological changes have likely evolved as a way for placental mammals to tolerate the developing fetus, which expresses both paternal and maternal allo-antigens (1). Accordingly, aberrant adaptations of the maternal immune system during the prenatal period have been suggested to contribute to increased chance of adverse pregnancy and offspring outcomes (2–8). Identifying which factors contribute to atypical immune adaptation during pregnancy is therefore of clinical importance.
Complex immunological processes at the maternal-fetal interface have been mainly studied in rodent models. Preclinical studies have shown that the equilibrium of T-, B- and uterine natural killer (NK) cells (9) and the production of immunosuppressive cytokines by regulatory T cells (10) is important in a typical pregnancy. These models have also shown that a shift in T-helper (Th) cell balance toward increased Th1 cells, complement activation and the depletion of regulatory T cells or NK cells may all lead to adverse pregnancy outcomes, including fetal rejection, impaired fetal growth, and abnormal neurodevelopment (11–16). Mechanistic preclinical models allow for the investigation of the maternal immunological system in ways that cannot be done in humans, including knock-out models to assess the role of particular immune cells, and harvesting of immune system tissues at different timepoints during gestation (17). Together, animal studies have been particularly useful in revealing that a well-balanced immune system is crucial for a successful pregnancy.
In humans, assessment of peripheral blood biomarkers, including cytokines, chemokines and cell-type composition has been employed in an effort to translate preclinical findings to human pregnancy. Signaling molecules such as C-reactive protein (CRP) and cytokines tightly regulate immunological processes and are therefore frequently measured in clinical studies. CRP is an acute-phase protein produced by liver cells and its production is stimulated by proinflammatory cytokines including interleukin (IL)-6 and IL-1. Cytokines are produced by a variety of immune and non-immune cells. During pregnancy, the placenta is an important additional source. IL-1β, IL-6, IL-17A, IL-23 and interferon (IFN)-γ have been implicated in early pregnancy processes, such as implantation (18, 19) and spiral artery remodeling of the uterine vessels (20). Various studies (sample size range n=20-1274) have investigated inflammatory marker patterns over the course of pregnancy and have suggested shifts in the levels of inflammatory biomarkers over time (21–30). For example, the largest study to date suggested an increase of IL-6 and IFN-y between early (median 8.5 weeks) and mid-pregnancy (median 25 weeks) (22), while the second largest study showed decreased cytokine levels from early to mid-pregnancy, reflective of a pro-inflammatory environment in the first trimester followed by an anti-inflammatory state during the second trimester (27). For the majority of cytokines, a consistent trend throughout pregnancy was not observed across the literature (3). Existing studies were limited in terms of generalizability (i.e. no independent replication cohorts; exclusion of susceptible subgroups) and methodological variability between cohorts (21–30). Large studies with repeated measures, identical assays, and replication cohorts are needed to expand our knowledge of the drivers of immune markers in pregnancy. A robust characterization of inflammatory markers, including trajectories and driving forces, throughout normal gestation may add to our understanding of the role of atypical immune adaptation.
Well-known triggers such as viral or bacterial infection might be causal to immune activation. Yet, other inflammatory factors such as high maternal age, high body mass index (BMI), tobacco use and parity, as well as child sex, might also play a role, as suggested by studies of pregnant (21, 22, 27, 31–33) and non-pregnant (34) populations. Additionally, the genetic susceptibility to elevated inflammatory marker levels may play a role as indicated for example by a large genome-wide study (GWAS) reporting genetic loci associated with CRP levels (35). While there is evidence that immune dysregulation is associated with adverse maternal and fetal outcomes, it is currently unclear to what extent these factors contribute to dysregulation of the maternal immune system in the general pregnant population. Previous studies have typically employed a cross-sectional design (36), or included single or limited number of biomarkers (37). Identifying robust drivers of systemic immune adaptations will enhance our understanding of successful maintenance of pregnancy, as well as of adverse outcomes.
The two-fold aim of the present study is to i) map inflammatory marker (high sensitivity (HS)-CRP, IL-1β, IL-6, IL-17A, IL-23, IFN-γ) patterns throughout pregnancy and ii) investigate the impact of demographic, environmental and genetic factors on maternal inflammatory marker levels. Blood samples were collected at two timepoints in the first half of pregnancy. Maternal serum inflammatory marker levels were measured by multiplex analysis. To account for immune marker inter-relationships, principal component analysis (PCA) was used to extract underlying inflammatory components and create a cytokine index, in line with prior approaches (38, 39). The Generation R Study, a multi-ethnic population-based prospective pregnancy cohort in the Netherlands (NL) (n=8,082) was used as a discovery cohort (40). Importantly, findings were replicated in three cohorts, namely the Generation C cohort (n=2,535, USA), the Brabant Study cohort (n=587, NL) and the Generation R Next Study (n=1,270, NL) with biomarkers assessed at multiple timepoints using identical assays.
Results
Characteristics of four cohorts
A total of 8,082 pregnant participants were included in the Discovery Cohort (Table 1). Of the included participants, 5,478 participants (67.8%) had repeated cytokine measurements and 5,938 participants (73.5%) had genotype data. In total, 13,467 samples were collected with complete cytokine data. Of these, 13,316 samples also had HS-CRP data. Results were replicated in the Generation C cohort (n=2,535) and the Brabant Study cohort (n=587), hereafter referred to as Replication Cohort I and Replication Cohort II, respectively (Table 1). In Replication Cohort I, 3,319 samples were collected from 2,535 participants and 541 of these participants (21.3%) had 2 to 6 repeated cytokine measurements. In Replication Cohort II, 1,170 samples were included from 587 participants and 387 of these participants (65.9%) had 2 to 3 repeated cytokine measurements. The Generation R Next, hereafter referred to as the pre-pregnancy cohort, collected samples preconception and during pregnancy. In the pre-pregnancy cohort, 1,779 samples were collected from 1,270 participants and 395 of these participants (31.3%) had up to 5 repeated cytokine measurements preconception and during pregnancy. Study design is visualized in Figure 1A. Demographics of the Discovery Cohort are visualized in Figures 1B–G.
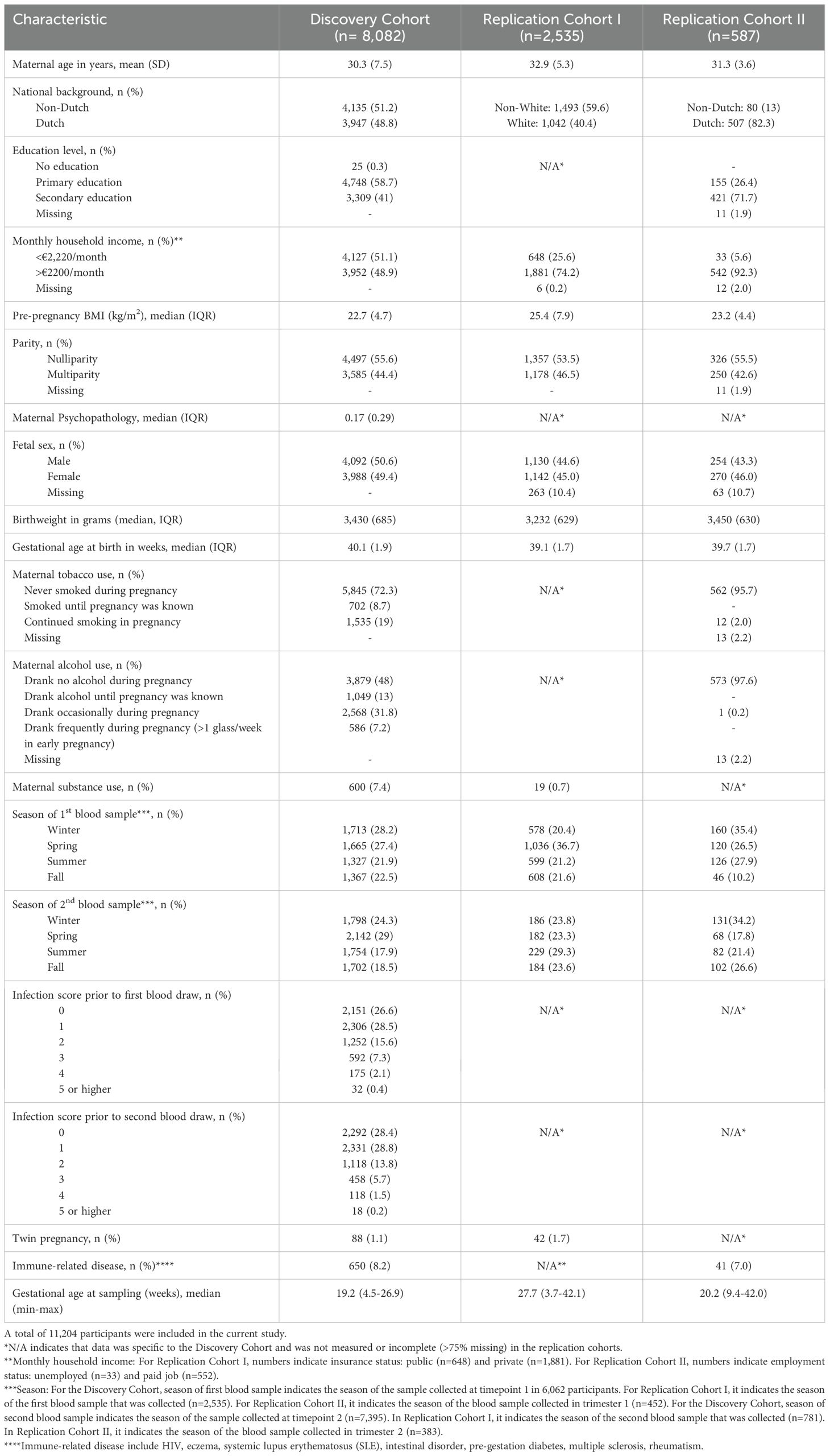
Table 1. Characteristics of the Discovery Cohort, Replication Cohort I, and Replication Cohort II.
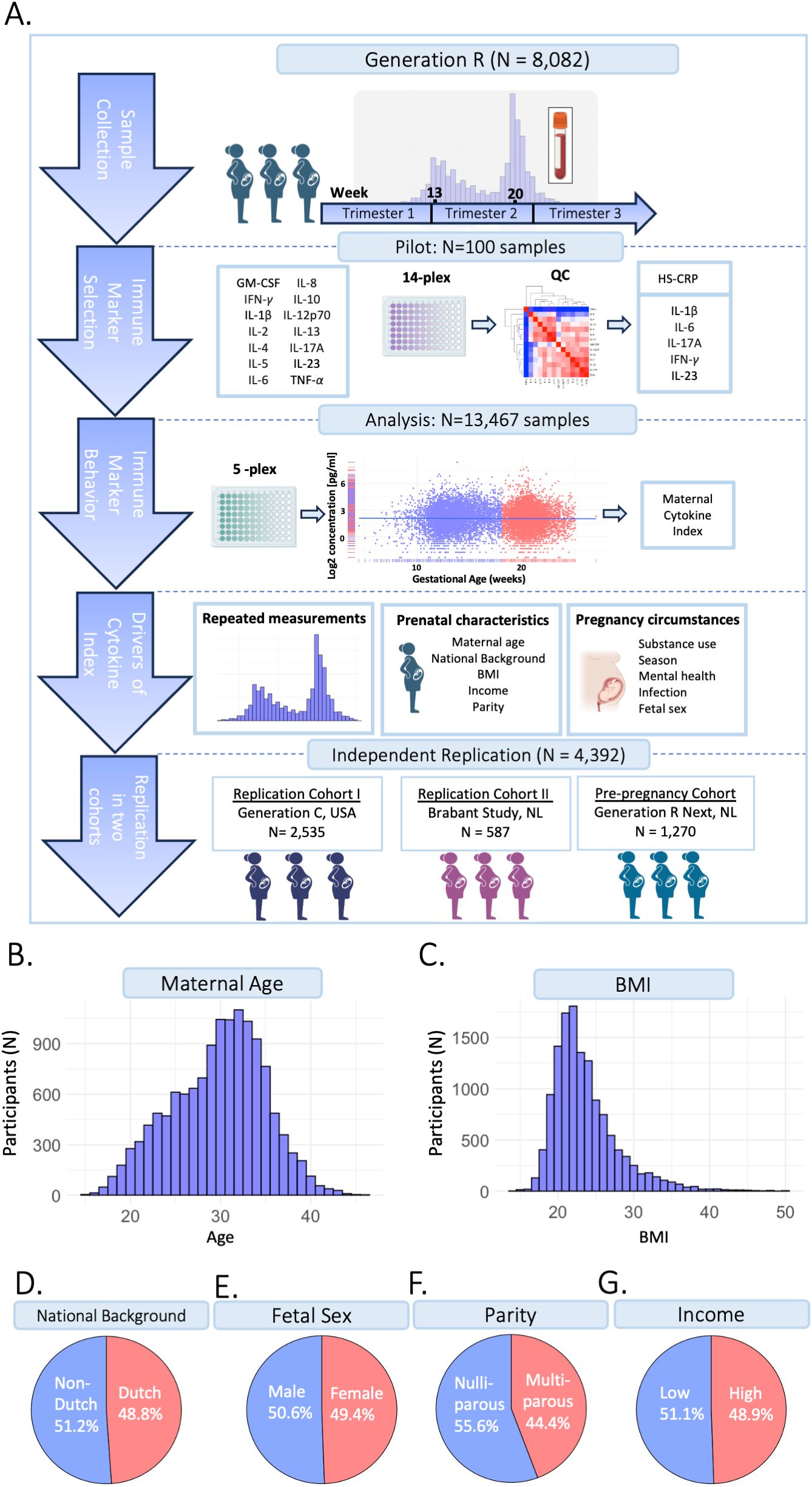
Figure 1. Study design. (A) The current study is part of the Generation R Study pregnancy cohort (n=8,082 participants). Sample collection occurred between 2001-2006. Blood samples were collected at two timepoints at median 13.2 weeks (95% range: 9.6-17.6 weeks) and 20.3 weeks (95% range: 18.5-23.3 weeks) gestation, processed and stored for further analyses. A pilot analysis of 14 cytokines was performed in 100 samples. IL-1β, IL-6, IL-17A, IFN-γ and IL-23 were selected for analysis in the full cohort, as well as HS-CRP. Participants were excluded based on several exclusion criteria. Data analysis was performed in n=13,467 samples of n=8,082 included participants, of which n=5,478 had a repeated measurement. Inflammatory markers were characterized throughout gestation. A maternal cytokine index was generated for each sample, reflecting inflammatory marker behavior. To identify potential drivers of inflammatory markers, the association between pre-pregnancy characteristics and pregnancy circumstances as predictors of the maternal cytokine index and HS-CRP was assessed. Findings were replicated in two replication cohorts: the Generation C study (n=2,535) and the Brabant Study (n=587) and in a unique pre-pregnancy cohort, the Generation R Next study (n=1,270). (B) Distribution of maternal age (years). (C) Distribution of BMI (kg/m2). (D) Distribution of national background (Dutch/non-Dutch). (E) Distribution of fetal sex (female/male). (F) Distribution of parity (nulliparous/multiparous). (G) Distribution of household income (low: <€2,220/month/high: >€2200/month).
Descriptive analysis of maternal inflammatory markers
Discovery cohort
In the Discovery Cohort, blood samples were collected at two timepoints during gestation. Median gestational age at sample collection of the first blood sample was 13.2 weeks (95% range: 9.6-17.6 weeks) in early pregnancy, and 20.3 weeks (95% range: 18.5-23.3 weeks) in mid-pregnancy (Figure 2A). Of the 13,467 samples, 6,072 (45%) were collected in early pregnancy and 7,395 (55%) in mid pregnancy. Median inflammatory marker levels were 3.95 pg/mL for IL-1β, 1.61 pg/mL for IL-6, 23.98 pg/mL for IL-17A, 1,107.4 pg/mL for IL-23, 14.75 pg/mL for IFN-γ, and 4.3 mg/L for HS-CRP (Table 2). The univariate analysis showed that group-level inflammatory marker levels were significantly lower at the second timepoint compared to the first (Figure 2B; Table 2). Repeated measures were median 50 days apart among participants with a repeated measurement (n=5,478 participants, n=10,956 samples). The intra-individual correlation of inflammatory markers between timepoints in these participants was strong (r=0.68-0.93, p<0.001) (Figures 2C, D).
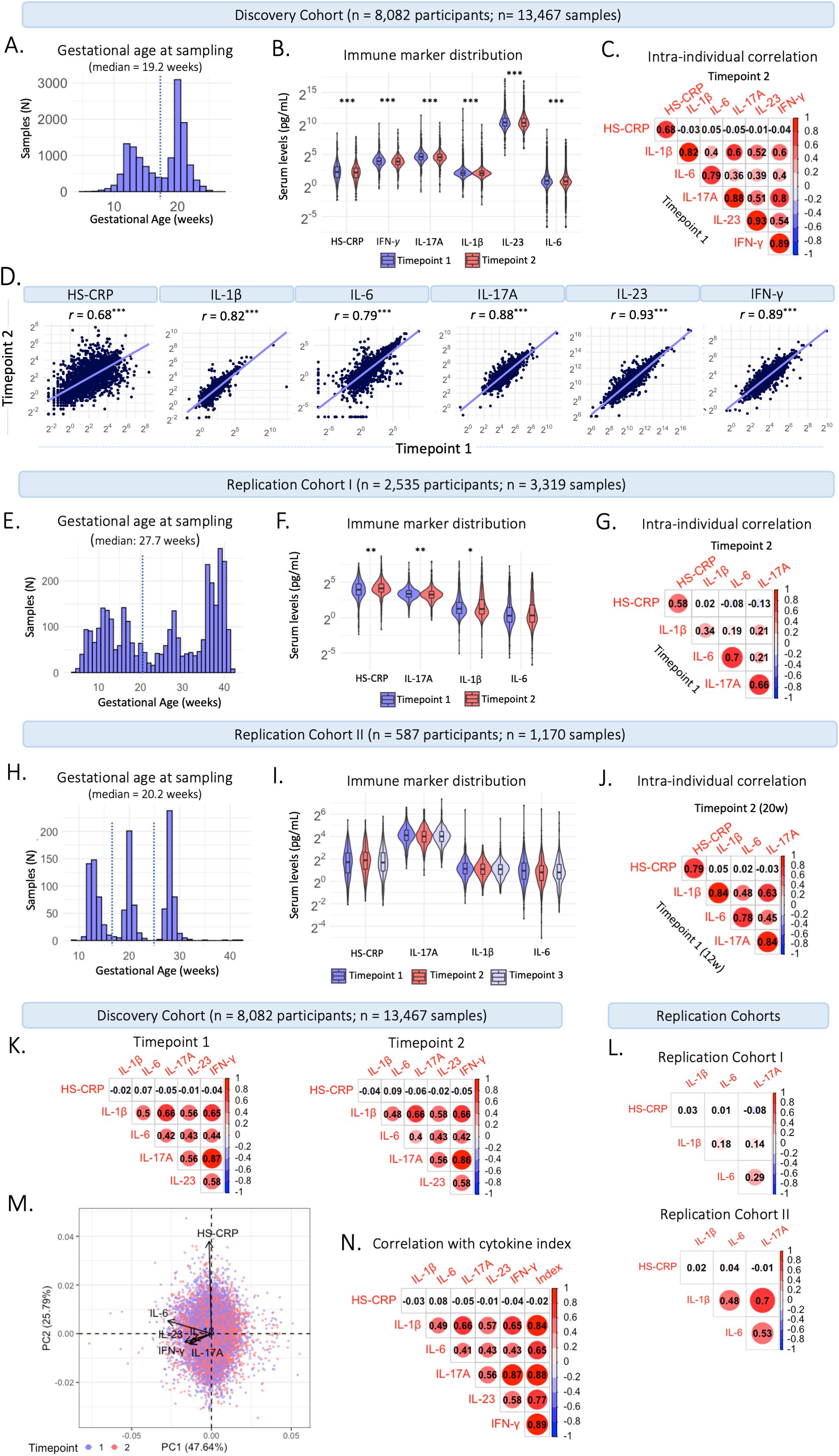
Figure 2. Characteristics of maternal inflammatory markers. (A) Distribution of gestational age (weeks) at sample collection in the Discovery Cohort. The dotted line indicates the division between timepoint 1 (trimester 2; 13.2 weeks) and timepoint 2 (trimester 2; 20.3 weeks). (B) Violin plots of inflammatory markers measured at timepoint 1 at median 13.2 weeks gestation (95% range: 9.6-17.6 weeks) and timepoint 2 at median 20.3 weeks gestation (95% range: 18.5-23.3 weeks). (C) Intra-individual correlation between samples collected at timepoint 1 and at timepoint 2 among participants with repeated measurements (n=5,478 participants; 10,956 samples). (D) Intra-individual correlation between timepoint 1 (median 13.2 weeks) and timepoint 2 (median 20.3 weeks) in participants with repeated measures in the Discovery Cohort. (E) Distribution of gestational age (weeks) at sample collection in Replication Cohort (I) The dotted line indicates the division between timepoint 1 (early gestation; 20 weeks) and timepoint 2 (late gestation; >20 weeks). (F) Violin plots of inflammatory markers measured in early gestation (20 weeks) and late gestation (>20 weeks). (G) Intra-individual correlation between the first and second sample collected among participants with repeated measurements (n=541). (H) Distribution of gestational age (weeks) at sample collection in Replication Cohort II. The dotted line indicates the division between samples collected at timepoint 1 (trimester 1; 12 weeks), timepoint 2 (trimester 2; 20 weeks), and timepoint 3 (trimester 3; 28 weeks). (I) Violin plots of inflammatory markers measured at timepoint 1-3. (J) Intra-individual correlation between timepoint 1 and 2 among participants with repeated measurements (n=387). (K) Correlation between cytokines and HS-CRP at timepoint 1 (left) and timepoint 2 (right) in the Discovery Cohort. (L) Correlation between inflammatory markers in Replication Cohort I (top) and Replication Cohort II (bottom). (M) Principal component analysis of cytokines and HS-CRP in the Discovery Cohort, indicating high loadings of IL-1β, IL-6, IL-17A, IFN-γ and IL-23 on Principal Component (PC) 1 and of HS-CRP on PC 2. Samples are color coded by timepoint. (N) Correlation of cytokines and HS-CRP with the maternal cytokine index across all samples in the Discovery Cohort. Asterisks indicate statistical significance level (*p<0.05, **p<0.01, ***p<0.001).
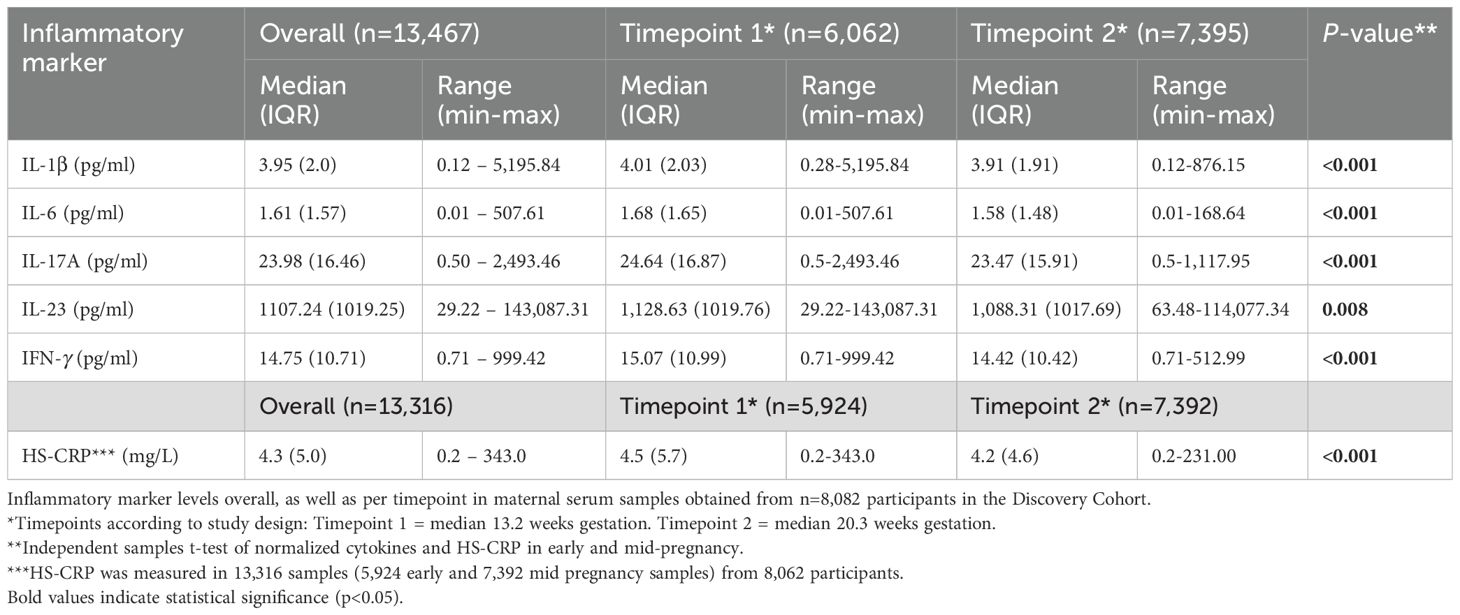
Table 2. Characteristics of inflammatory markers in the Discovery Cohort.
Replication Cohort I
In Replication Cohort I, blood samples were collected throughout gestation, at a median gestational age of 27.7 weeks (95% range: 6.9-40.6 weeks) (Figure 2E). Median inflammatory marker levels were 2.38 pg/mL for IL-1β, 1.24 pg/mL for IL-6, 9.85 pg/mL for IL-17A, and 16.9 mg/L for HS-CRP (Supplementary Table 1). The univariate analysis showed that group-level IL-1β (p=0.025) and IL-17A (p=<0.001) levels were lower in late pregnancy (>20 weeks), HS-CRP (p=<0.001) levels were higher in late pregnancy (>20 weeks) and IL-6 (p=0.066) levels were not different between early (<20 weeks) and late (>20 weeks) pregnancy (Figure 2F; Supplementary Table 1). Repeated measures were median 48 days apart among participants with a repeated measurement (n=541 participants, n=1,292 samples). The intra-individual correlation of inflammatory markers between early (<20 weeks) and late (>20 weeks) pregnancy in these participants was moderate to strong (r =0.34-0.70, p<0.001) (Figure 2G).
Replication Cohort II
In Replication Cohort II, blood samples were collected at 12, 20, and 28 weeks gestation, at a median gestational age of 20.2 weeks (95% range: 12.0-29.5 weeks) (Figure 2H). Median inflammatory marker levels were 2.14 pg/mL for IL-1β, 1.79 pg/mL for IL-6, 16.55 pg/mL for IL-17A, and 4.78 mg/L for HS-CRP (Supplementary Table 2). The univariate analysis showed that group-level inflammatory marker levels were not significantly different between three timepoints (Figure 2I, Supplementary Table 2). Repeated measures were median 50 days apart among participants with a repeated measurement (n=387 participants, n=970 samples). The intra-individual correlation of inflammatory markers between 12 and 20 weeks of pregnancy in these participants was strong (r =0.78-0.84, p<0.001) (Figure 2J) and similar between 12 and 28 weeks (r =0.71-0.84, p<0.001) and 20 and 28 weeks (r =0.76-0.86, p<0.001).
Construction of the maternal cytokine index
Given that inflammatory markers interact, and to capture systemic inflammatory marker changes, inflammatory markers were summarized in a maternal cytokine index in the Discovery Cohort. Cytokines IL-1β, IL-6, IL-17A, IL-23, and IFN-γ correlated moderately to strongly with each other at timepoint 1 and timepoint 2 (r=0.40 – 0.86; Figure 2K). HS-CRP showed a low correlation with the cytokines at both timepoints (Pearson’s r=-0.06 – 0.09). The low correlation of HS-CRP with cytokines was also seen in the two replication cohorts (Figure 2L). In a principal component analysis (PCA), standardized cytokines IL-1β, IL-6, IL-17A, IL-23, and IFN-γ loaded highly on the first principal component (PC) (84%, 66%, 89%, 77%, 89%, respectively), accounting for 47.64% of the variance (Figure 2M). Given the correlation structure among the inflammatory markers and the high loading of HS-CRP on the second PC, HS-CRP was excluded from the maternal cytokine index. The maternal cytokine index correlated strongly with individual cytokines at timepoint 1 (r=0.66 – 0.89) and timepoint 2 (r=0.64 – 0.89) and correlation between cytokines and the maternal cytokine index was higher compared to correlations among cytokines (Figure 2N).
Aim 1: mapping inflammatory marker dynamics throughout gestation
In the Discovery Cohort, gestational age (range 4.5-26.9 weeks) was significantly associated with HS-CRP (p<0.001) and the maternal cytokine index (p<0.001) in univariate analyses (Figure 3A). HS-CRP showed an increase in early pregnancy, followed by a gradual decrease (Figure 3A). The maternal cytokine index showed a gradual decrease throughout early and mid-pregnancy (Figure 3A). Individual cytokines displayed similar trajectories across gestational age (Figure 3A). In Replication Cohort I, gestational age at sample collection (range 3.7-42.1 weeks) was significantly associated with HS-CRP, IL-1β, IL-6, and IL-17A (p<0.001) in univariate analyses (Figure 3B). Similar to the Discovery Cohort, HS-CRP showed an increase in early pregnancy, followed by a gradual decrease. Cytokines showed a gradual decrease throughout early pregnancy, followed by an increase of IL-1β and IL-6, but not IL-17A, in the third trimester. In Replication Cohort II, gestational age at sample collection (range 9.4-42.0 weeks) was not significantly associated with inflammatory markers (HS-CRP (p=0.192), IL-1β (p=0.372), IL-6 (p=0.111), and IL-17A (p=0.102)) in univariate analyses (Figure 3C).
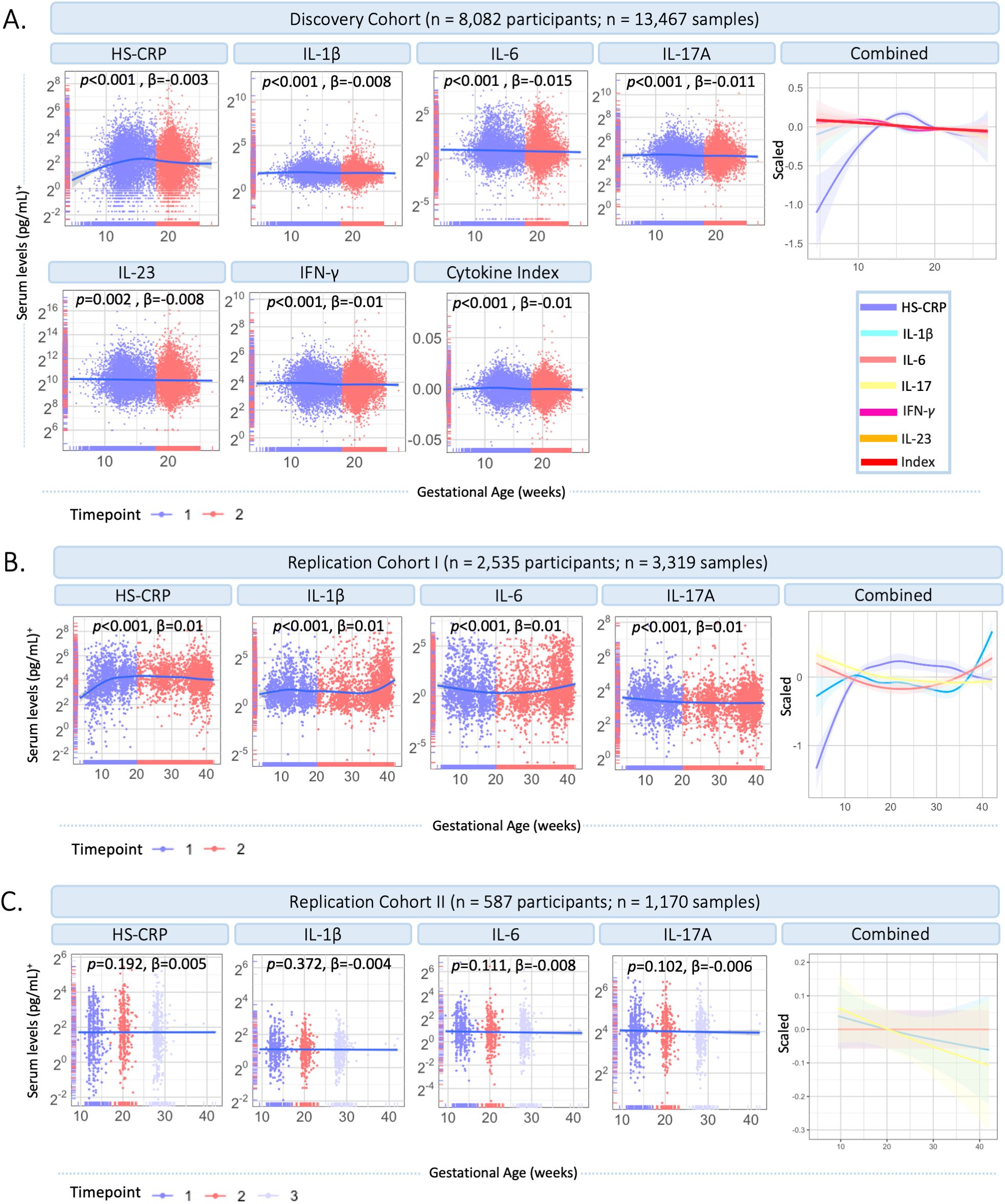
Figure 3. Inflammatory marker dynamics throughout gestation. (A) Inflammatory markers across gestational age in the Discovery Cohort (n=13,467 samples). Measurements are color coded by timepoint according to study design (timepoint 1 = median 13.2 weeks; timepoint 2 = 20.3 weeks). Inflammatory markers and the maternal cytokine index are scaled and combined in one plot (right). (B) Inflammatory markers across gestational age in Replication Cohort I (n=3,319 samples). Measurements are color coded by timepoint according to study design (timepoint 1 = 20 weeks gestation; timepoint 2 = >20 weeks gestation). Inflammatory markers are scaled and combined in one plot (right). (C) Inflammatory markers across gestational age in Replication Cohort II (n=1,170 samples). Measurements are color coded by timepoint according to study design (timepoint 1 = median 12 weeks; timepoint 2 = median 20 weeks; timepoint 3 = median 28 weeks). Inflammatory markers are scaled and combined in one plot (right). Legend: A general additive modeling (GAM) line was fitted for each marker indicated with the blue line and a 95% confidence interval on a log2 y-axis. P-values and effect sizes of the association with gestational age are provided. Color coding of inflammatory markers is identical in Figures A–C. +HS-CRP was measured in mg/L, cytokines were measured in pg/ml.
Aim 2: drivers of maternal inflammatory markers
Environmental and genetic drivers of HS-CRP and the maternal cytokine index (univariate analyses)
To characterize drivers of the maternal inflammatory marker landscape, we assessed the association of multiple predictors with HS-CRP and the maternal cytokine index. In the Discovery Cohort (Figures 4A–C; Supplementary Table 3), most of the variance in HS-CRP (55.6%) and the maternal cytokine index (87.4%) is explained by within-individual effects. In addition, the variance in HS-CRP levels was partly driven by pre-pregnancy BMI (9.6%) (Figures 4A–C; Supplementary Table 3). Less than 1% of the variance in inflammatory markers was explained by pre-pregnancy characteristics and pregnancy circumstances (Figures 4A–C). These findings were replicated in both replication cohorts as the majority of the variance in HS-CRP (Replication Cohort I = 33.4%; Replication Cohort II = 58.1%) and cytokines (Replication Cohort I = 26.3-71.5%; Replication Cohort II = 74.7-84.2) was explained by within-individual effects (Figure 4D; Supplementary Table 3). Pre-pregnancy BMI contributed to the variance in HS-CRP in both Replication Cohort I (14.9%) and Replication Cohort II (16.1%) (Figure 4D; Supplementary Table 3). Less than 1% of the variance in inflammatory markers was explained by pre-pregnancy characteristics and pregnancy circumstances (Figure 4D). Across all cohorts, the remaining variance was attributed to residual factors (Discovery Cohort = 6.6%-32.1%; Replication Cohort I = 28%-72.9%; Replication Cohort II = 14.7%-25.1%) (Figures 4A–D; Supplementary Table 3). Across all cohorts, HS-CRP and pre-pregnancy BMI correlated moderately (r= 0.32 – 0.41) (Figure 4E). Within-individual concordance was further confirmed by individual trajectories among participants with repeated measurements in the Discovery Cohort and both Replication Cohorts (Supplementary Figures 4A–C). Based on the high intra-individual correlation of inflammatory markers and the lack of variance explained by pre-pregnancy characteristics and pregnancy circumstances, we further investigated whether HS-CRP levels were genetically determined. We constructed a PGS of CRP in all ancestries and European ancestries in the discovery cohort, both of which showed a moderate correlation (r =0.4) with serum HS-CRP levels (Figure 5A). Of the variance in HS-CRP, 14.1% was explained by the CRP PGS in all ancestries and 15.7% was explained by the CRP PGS in European ancestries (Figure 5B).
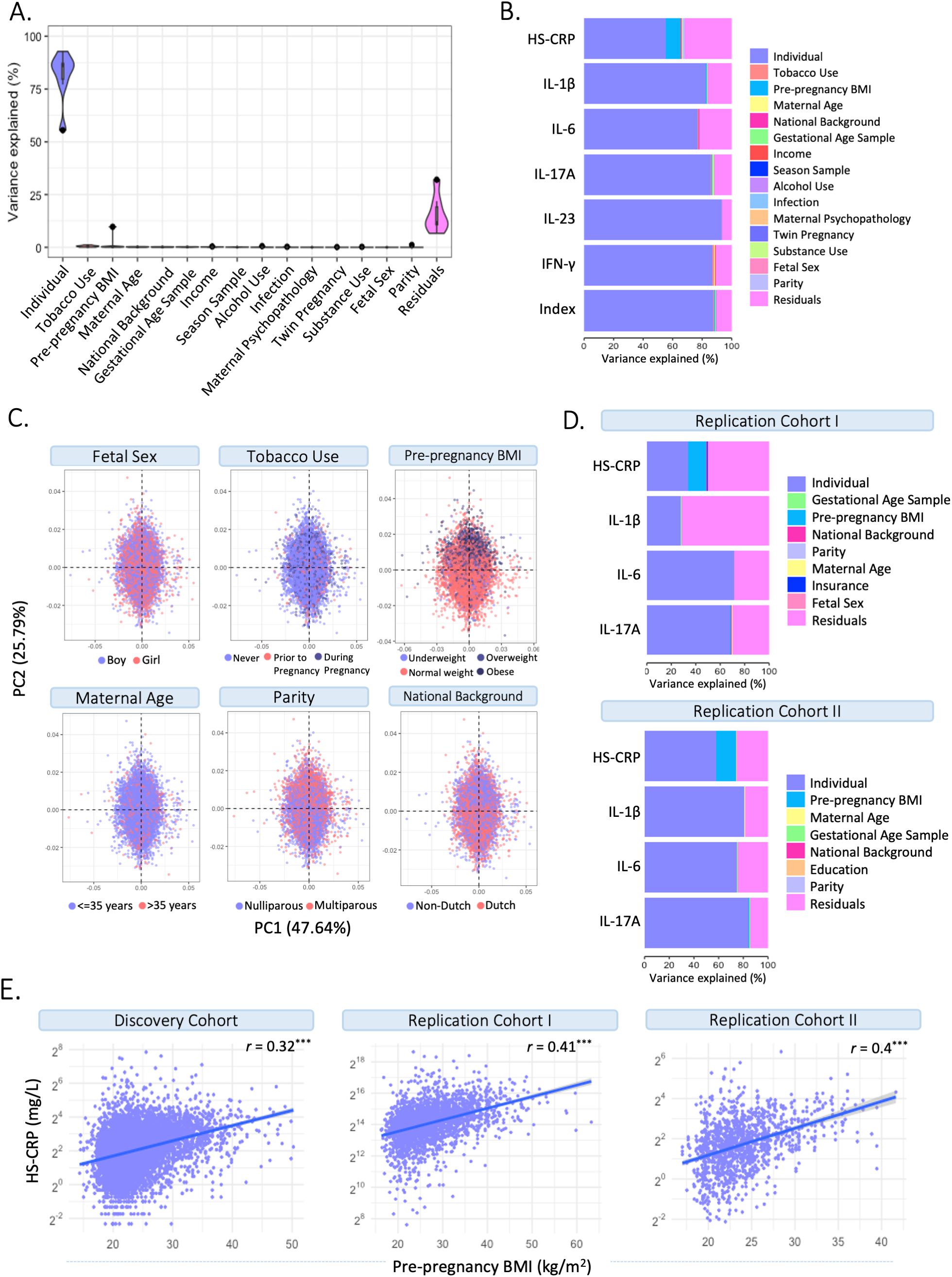
Figure 4. Identifying drivers of maternal inflammatory markers. (A) Variance partitioning analysis revealed the variance explained by potential drivers in HS-CRP, cytokines, and the maternal cytokine index. (B) Variance partitioning analysis revealed the variance explained by potential drivers in HS-CRP, cytokines, and the maternal cytokine index. (C) Distribution of potential drivers of inflammatory markers across Principal Component Analysis (PCA). (D) Variance partitioning analysis revealed the variance explained by potential drivers in inflammatory markers in Replication Cohort I (left) and Replication Cohort II (right). (E) HS-CRP and pre-pregnancy BMI show a moderate correlation (r = 0.32) in the Discovery Cohort and both Replication Cohort I (r = 0.41) and Replication Cohort II (r = 0.4). Asterisks indicate statistical significance level (*p<0.05, p<0.01, *p<0.001).
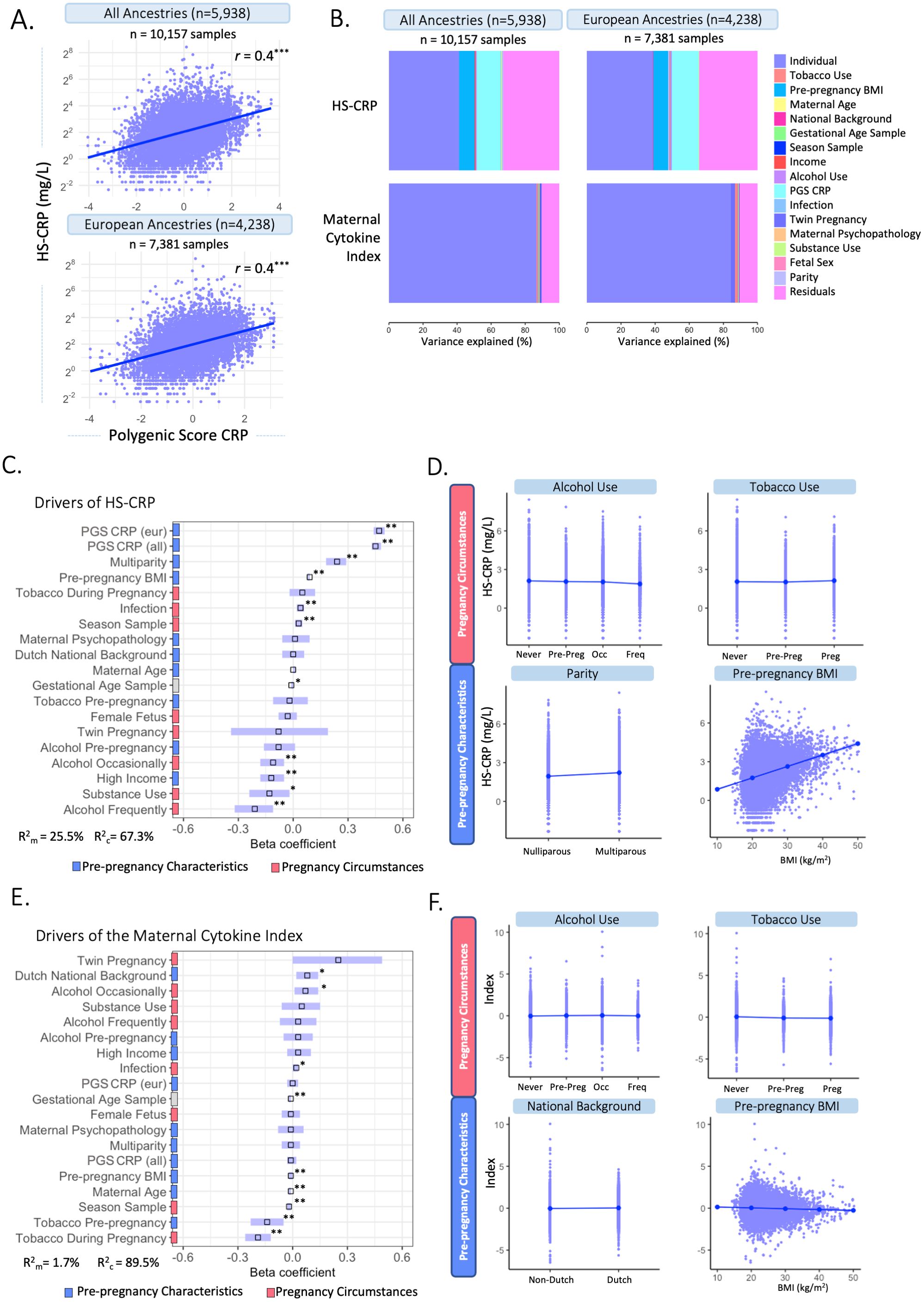
Figure 5. Multivariable associations between potential drivers and HS-CRP and the maternal cytokine index. (A) A polygenic Score (PGS) was computed for CRP among participants of all ancestries (n=5,938) and participants of European ancestry (n=4,238). CRP PGS were correlated with HS-CRP levels. Y-axis is on a log2 scale. (B) Variance explained by potential drivers in HS-CRP and the maternal cytokine index, including the PGS of all ancestries (left) and European ancestries (right). (C) Mixed effects linear regression model of potential drivers and their association with HS-CRP was performed among participants with a CRP PGS (n=5,938 participants, n=10,157 samples). Forest plot shows beta coefficients and 95% confidence interval of selected predictors with HS-CRP. (D) Visualization of the association of various pre-pregnancy characteristics and pregnancy circumstances with HS-CRP. (E) Mixed effects linear regression model of potential drivers and their association with the maternal cytokine index was performed among participants with a CRP PGS(n=5,938 participants, n=10,157 samples). Forest plot shows beta coefficients and 95% confidence interval of selected predictor. (F) The association between various pre-pregnancy characteristics and pregnancy circumstances with the maternal cytokine index. Legend: (C-F) Pre-pregnancy characteristics are indicated by blue; pregnancy circumstances are indicated by red. C&E: R2m indicates the marginal R squared of the mixed effects regression model, R2c indicates the conditional R-squared of the mixed effects regression model after adding individual as a random intercept. Asterisks indicate significance level after multiple testing correction (*p<0.05, **p<0.01, ***p<0.001). Units of continuous variables are as follows: maternal age (years); pre-pregnancy BMI (kg/m2); Infection score (sum score); maternal psychopathology (sum score); gestational age at sampling (weeks); PGS CRP European ancestry (polygenic score); PGS CRP all ancestry (polygenic score). Levels of categorical variables are as follows: Dutch national background vs non-Dutch national background; multiparity vs nulliparity; tobacco during pregnancy vs never smoked; tobacco pre-pregnancy vs never smoked; female fetus vs male fetus; twin pregnancy vs non-twin pregnancy; alcohol use pre-pregnancy vs never alcohol use; alcohol use occasionally vs never alcohol use; alcohol use frequently vs never alcohol use; high income (>€2200/month) vs low income (<€2,220/month); substance use vs no substance use. D&F: Alcohol categories are ‘never drank’, ‘continued drinking until pregnancy was known (pre-preg)’, ‘drank occasionally (occ)’ and ‘drank frequently (freq)’. Tobacco categories are ‘never smoked’, ‘continued tobacco use until pregnancy was known (pre-preg)’ and ‘continued tobacco use during pregnancy (preg)’.
Environmental and genetic drivers of HS-CRP (multivariable analyses)
Mixed effects linear regression models were performed to identify potential drivers of HS-CRP among participants with a CRP PGS (n=5,938 participants, n=10,157 samples). The CRP PGS was the strongest driver of HS-CRP among participants of all ancestry (β=0.45, 95% CI = 0.43; 0.48, p=0.003) and among participants of European ancestry (β=0.47, 95% CI = 0.44; 0.50, p=0.003) (Figure 5C). In addition, we found significant associations with pre-pregnancy characteristics BMI (β=0.09, 95% CI = 0.08; 0.09, p=0.003), high household income (β =-0.12, 95% CI =-0.18; -0.05, p=0.003), and multiparity (β =0.24, 95% CI = 0.18-0.29, p=0.003), as well as pregnancy circumstances, namely occasional and frequent alcohol use during pregnancy (β =-0.11, 95% CI = -0.18; -0.05, p= 0.003 and β =-0.21, 95% CI = -0.32; -0.11, p=0.003, respectively), season at first blood draw (β =0.03, 95% CI = 0.01; 0.05, p= 0.010) and infection score during pregnancy (β =0.04, 95% CI = 0.02; 0.06, p=0.003) in the Discovery Cohort (Figures 5C, D, Table 3). Maternal age (β =0.00, 95% CI = 0.00; 0.01, p=0.470), national background (β =0.00, 95% CI = -0.06; 0.06, p=0.945), maternal psychopathology (β =0.01, 95% CI = -0.06; 0.09, p=0.815), female fetus (β =-0.03, 95% CI = -0.08; 0.02, p=0.358), and twin pregnancy (β =-0.08, 95% CI = -0.34; 0.19, p=0.678) were not associated with HS-CRP (Figures 5C, D, Table 3). The marginal variance (i.e., variance explained by fixed effects) was 25.5%. Taking into account the individual, modeled as a random intercept, the conditional variance explained was 67.3%. In replication cohorts, pre-pregnancy BMI was significantly associated with increased HS-CRP levels in Replication Cohort I (β = 0.07, 95% CI = 0.06; 0.07, p=0.004) and Replication Cohort II (β = 0.14, 95% CI = 0.11; 0.16, p=0.009). White ethnicity was associated with decreased HS-CRP levels in Replication Cohort I (β = -0.14, 95% CI = -0.24; -0.05, p=0.005), but this was not the case for national background in Replication Cohort II (β = 0.15, 95% CI = -0.16; 0.46, p=0.623). Other maternal factors showed no significant associations with HS-CRP in Replication Cohort I and II (Supplementary Figures 5B, C, Supplementary Table 4).
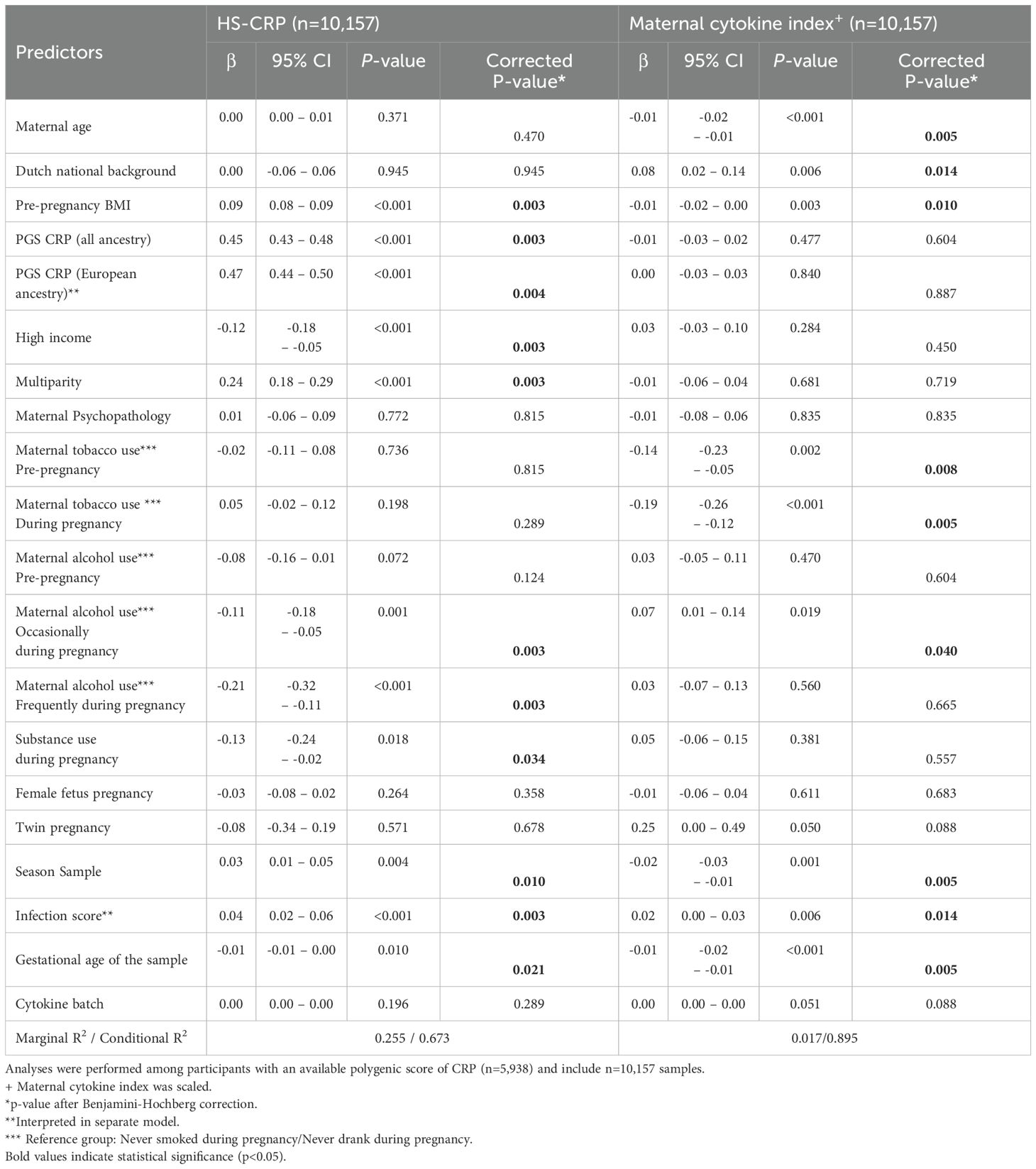
Table 3. Mixed effects models to investigate the association between selected predictors and HS-CRP and the maternal cytokine index.
Environmental and genetic drivers of the maternal cytokine index (multivariable analyses)
Next, we identified several significant associations between potential drivers of inflammation and the maternal cytokine index, including pre-pregnancy characteristics such as maternal age (β =-0.01, 95% CI = -0.02; -0.01, p=0.005), pre-pregnancy BMI (β =-0.01, 95% CI = -0.02; 0.00, p=0.003), and Dutch national background (β=0.08, 95% CI = 0.02; 0.14, p=0.014), as well as pregnancy circumstances including tobacco use pre-pregnancy and during pregnancy (β =-0.14, 95% CI = -0.23; -0.05, p=0.008; β =-0.19, 95% CI = -0.26; -0.12, p=0.005, respectively), occasional alcohol use during pregnancy (β =0.07, 95% CI = 0.01; 0.14, p= 0.040), season at first blood draw (β =-0.02, 95% CI = -0.03; -0.01, p=0.005), and infection score during pregnancy (β =0.02, 95% CI = 0.00 – 0.03, p= 0.014) in the Discovery Cohort (Figures 5E, F, Table 3). Multiparity (β = -0.01, 95% CI = -0.06; 0.04, p=0.719), maternal psychopathology (β = -0.01, 95% CI = -0.08; 0.06, p=0.835), substance use during pregnancy (β = 0.05, 95% CI = -0.06; 0.15, p=0.557), female fetus (β = -0.01, 95% CI = -0.06; 0.04, p=0.683), twin pregnancy (β = 0.25, 95% CI = 0.00; 0.49, p=0.088), and the CRP PGS for all ancestry (β = -0.01, 95% CI = -0.03; 0.02, p=0.604) and for European ancestry (β = 0.00, 95% CI = -0.03; 0.03, p=0.887) were not associated with the maternal cytokine index (Figures 5E, F, Table 3). The marginal variance (i.e., variance explained by fixed effects) was 1.7%. Taking into account the individual, modeled as a random intercept, the conditional variance explained was 89.5%.
Comparison pre-pregnancy and pregnancy
The contribution of the PRS of CRP to serum levels of HS-CRP, suggested a genetic contribution to inflammatory marker variance during pregnancy. Together with the high intra-individual correlation of cytokines, this led to the hypothesis that immune markers might not show major changes during pregnancy compared to preconception. We assessed HS-CRP and cytokine behavior in a pre-pregnancy cohort, the Generation R Next study. Samples were collected preconceptionally at median 11 weeks prior to conception (95% range: 74.5–0 weeks prior to conception, n=676), and during pregnancy at median 8.4 weeks gestation (95% range: 6.4-12.9 weeks, n=1,103) (Figure 6A). Median inflammatory marker levels were 4.82 pg/mL for IL-1β, 1.92 pg/mL for IL-6, 59.47 pg/mL for IL-17A, 1,209.5 pg/mL for IL-23, 18.89 pg/mL for IFN-γ, and 1.4 mg/L for HS-CRP (Supplementary Table 5). The univariate analysis showed that inflammatory marker levels were not significantly different between preconception and first trimester for IL-1β, IL-6, IL-17A, IL-23 and IFN-γ (Supplementary Table 5; Figure 6B). HS-CRP was significantly higher in the first trimester compared to preconception (Supplementary Table 5; Figure 6B). The intra-individual correlation of cytokines preconception and during pregnancy was strong (r=0.79-0.96, p<0.001) (Figure 6C). HS-CRP showed a moderate correlation between preconception and first trimester samples (r=0.45, <0.001) (Figure 6C). In the pre-pregnancy cohort, gestational age at sample collection (range 85 weeks prior to preconception - 17.6 weeks gestation) was significantly associated with HS-CRP (p<0.001), but not with IL-1β (p=0.360), IL-6 (p=0.530), IL-17A (0.313), IL-23 (p=0.376) and IFN-γ (p=0.110) in univariate analyses (Figures 6D, E). Similar to the Discovery Cohort, HS-CRP showed an increase in early pregnancy.
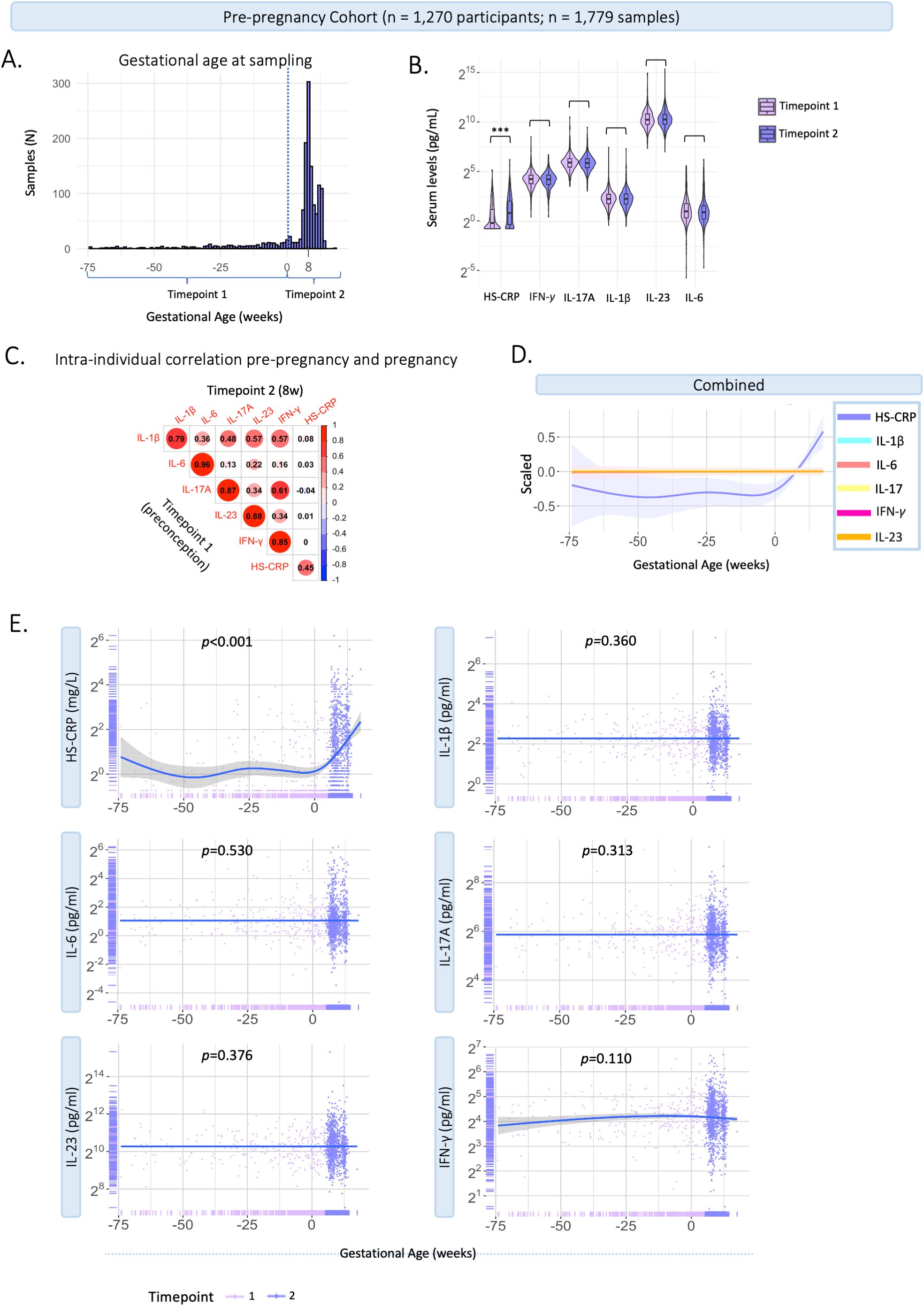
Figure 6. Characteristics of inflammatory markers pre-pregnancy and during pregnancy. Generation R Next cohort. (A) Distribution of gestational age (weeks) at sample collection (n=1,779 samples). The dotted line indicates the distinction between timepoint 1 (preconception samples), and timepoint 2 (trimester 1; median 8.4 weeks gestation). (B) Violin plots of inflammatory markers measured at timepoint 1 (preconceptionally; median 11.4 weeks before conception) and timepoint 2 (median 8.4 weeks gestation (95% range: 6.4-12.9 weeks). Cytokines (pg/mL) and HS-CRP (mg/L) are shown on a log2 axis. Univariate analysis at group-level show that cytokine levels were not significantly different between the first trimester and preconception. HS-CRP was significantly higher in the first trimester compared to preconception. (C) Intra-individual correlation between samples collected preconception (timepoint 1) and during pregnancy (timepoint 2) among participants with repeated measurements (n=395). Inflammatory markers were log2 transformed. Pearson correlation is shown. (D) Inflammatory markers are scaled and combined in one plot. (E) Inflammatory markers preconception and during pregnancy in the pre-pregnancy cohort (n=1,779 samples). Measurements are color coded by timepoint according to study design. A general additive modeling (GAM) line was fitted for each marker indicated with the blue line and a 95% confidence interval on a log2 y-axis. P-values indicate significance of the non-linear association with gestational age. HS-CRP was measured in mg/L, cytokines were measured in pg/ml. Asterisks indicate statistical significance level (*p<0.05, p<0.01, *p<0.001).
Sensitivity analyses
The PGS of CRP remained the largest predictor of HS-CRP in several sensitivity analyses in the Discovery Cohort, namely: i) excluding 163 samples (n=123 participants) with potential human anti-animal antibodies (HAAA), which can introduce technical interference in immunological assays (Supplementary Figures 2A, B, Supplementary Table 6); ii) excluding 855 samples of 495 participants with immune-related diseases (HIV, eczema, systemic lupus erythematosus (SLE), intestinal disorder, pre-gestation diabetes, multiple sclerosis, rheumatism) (Supplementary Figures 2C, D, Supplementary Table 7); and iii) excluding 174 outlier samples (n=147 participants) (Supplementary Figures 2E, F, Supplementary Table 8). Additionally, a sensitivity analysis was performed using the delta (change) between the two timepoints for HS-CRP and the maternal cytokine index. Infection score in early pregnancy (r =0.08, p<0.01) was correlated with the delta HS-CRP, and season (r =0.12, p=0.04) was correlated with the delta maternal cytokine index. Correlation coefficients between predictors and the HS-CRP delta (r =0.00-0.08) and maternal cytokine index delta (r =0.00-0.12) were low (Supplementary Figure 3A). The proportion of participants reporting an infection was significantly higher in the high-low HS-CRP group compared to the stable and low-high HS-CRP groups (Supplementary Figure 3B). Moreover, parity, season of first blood draw, and tobacco use during pregnancy significantly differed across categories of delta HS-CRP (Supplementary Figure 3B). Among maternal cytokine index delta categories, season of first blood draw differed significantly (Supplementary Figure 3C). A sensitivity analysis including only participants with a high absolute delta HS-CRP (5th and 95th quantiles) (n=435 participants; n=854 samples) no longer revealed significant associations between important drivers including multiparity, pre-pregnancy BMI, alcohol use, and household income with HS-CRP (Supplementary Figures 2G, H, Supplementary Table 9). The associations between predictors and individual cytokines in the Discovery Cohort are shown in Supplementary Figure 5A. White ethnicity, BMI, and maternal age were associated with IL-17A in Replication Cohort I. No significant associations of potential drivers were found with IL-1β and IL-6 in both Replication Cohorts (Supplementary Figures 5B, C).
Discussion
This study is the largest investigation to date of inflammatory markers during pregnancy in four multi-ethnic and socio-economically diverse pregnancy cohorts (>12,000 participants, The Netherlands and USA). Repeated measures enabled the thorough characterization of inflammatory marker patterns and the assessment of prominent inflammation drivers. Across cohorts, we consistently found strong correlations among cytokines, but no association between cytokines and HS-CRP. Results revealed high variability between individuals, yet a high intra-individual correlation of inflammatory markers measured at different time points during gestation as well as preconception using a unique pre-pregnancy cohort. Although we identified gestational age-dependent changes, results showed that gestational age at sampling explained less than 1% of the variance in inflammatory markers, similar to other factors including parity, tobacco use and fetal sex. Pre-pregnancy BMI and the polygenic score for CRP explained more than 9.6% and 14.1% of variance in HS-CRP levels, respectively.
Our results imply that selected cytokines (IL-1β, IL-6, IL-17A, IL-23, IFN-γ) share similar regulatory mechanisms, while HS-CRP is likely differentially regulated. Across the four cohorts, cytokines showed moderate to strong correlation with each other, but not with HS-CRP. These findings are in line with previous studies reporting a low correlation between HS-CRP and various inflammatory markers in early pregnancy (n=110-1,274) (22, 27, 28) and in a non-pregnant healthy female population (41). Additional evidence for differential regulation follows from distinct patterns of cytokines and HS-CRP, as early pregnancy confidence intervals did not overlap. Specifically, cytokines showed a gradual decrease throughout early pregnancy in the current study, in line with previous studies (27–29). Cytokine levels increased toward late pregnancy in Replication Cohort I, in line with upregulation of inflammatory pathways around parturition (42). HS-CRP deviated from the cytokine pattern and showed a gradual increase followed by a decrease throughout early gestation, in line with previous studies which also showed a distinct pattern for HS-CRP and a decrease around ~17 weeks (28) and ~24 weeks (27). Lastly, results revealed varying drivers of cytokines and HS-CRP as discussed below.
Precisely timed immune adaptations have been proposed as important regulating mechanisms in successful pregnancy (29). These immune adaptations are considered to reflect trimester-specific general anti- and pro-inflammatory states (27, 43). While our results showed gestational age-dependent inflammatory marker trajectories in two cohorts, we found no association with gestational age in Replication Cohort II. Across cohorts, less than 1% of the variance in inflammatory markers could be attributed to gestational age at sampling. Interestingly, when comparing pre-pregnancy and pregnancy immune markers, cytokines were also not significantly associated with gestational age, while HS-CRP is upregulated during pregnancy compared to preconception.
Inflammatory markers during pregnancy were found to be driven mainly by within-individual factors, as suggested by the high intra-individual correlation between repeated measures across cohorts. Rather than a pregnancy-specific phenomenon, this has been demonstrated in non-pregnant healthy and clinical populations as confirmed by longitudinal studies (n=10-250) indicating high intra-individual immune marker reproducibility in repeated samples ranging from 7 days to 25 years apart (44–47). We identified multiple pre-pregnancy and pregnancy specific drivers of peripheral HS-CRP levels, yet together these factors explained only 12% of variance in HS-CRP levels, the majority of which was captured by pre-pregnancy BMI (9.6%). The association with parity was not reported previously (27). Our finding that female fetal sex, maternal age and tobacco use were not associated with HS-CRP is in line with previous findings (27). The association between higher BMI and increased HS-CRP was replicated in two cohorts and confirms previous reports of an association between BMI and HS-CRP (27, 31, 48) but less so with other cytokines (49). Given that a prior study drew from a homogeneous population of normal singleton pregnancies excluding cases of immune disorders and extreme BMI (<18 or >40), the slightly higher impact of BMI on HS-CRP observed here is likely due to the use of a population-based cohort (27). Our findings are in line with non-pregnancy studies that showed a strong correlation between BMI and HS-CRP, but not IL-6 (50, 51). Several reviews have put forward CRP as a predictor of metabolic syndromes, non-alcoholic fatty liver disease and obesity, independent of inflammatory disease (48, 52). While it has been suggested that chronic subclinical inflammation is a consequence of obesity (53), other studies have showed that low-grade inflammation itself may cause insulin resistance which consequently also leads to obesity and metabolic disorders such as diabetes type 2 (54). The robust association between CRP and pre-pregnancy BMI in the current pregnant population supports the interaction between inflammatory and metabolic mechanisms. We additionally identified several pre-pregnancy and pregnancy specific drivers of the maternal cytokine index – yet the total variance in cytokines explained by these factors was only 1.5%. The significant impact of maternal age and tobacco use was not shown in a prior large study, possibly due to the population-based design of the current study compared to prior reports in a healthy pregnant population (27). Our finding that pre-pregnancy BMI was associated with increased inflammatory levels is in line with previous findings (21).
Bacterial and viral infections at time of blood sampling lead to upregulation of pro-inflammatory markers. In our cohorts, immune biomarkers were collected through research visits which likely caused healthy volunteer bias because ill patients may not attend. Due to the short half-life of immune markers, this limits detection of acute infection. Yet, our findings revealed that infection score in early pregnancy was correlated with HS-CRP in the first trimester and the maternal cytokine index between timepoints. In addition, we found that season of birth was correlated with these inflammatory markers, which is in line with prior findings of seasonal cytokine changes in mothers (55) and newborns (56, 57), possibly due to seasonality of viral exposures, vitamin D and allergen exposure (58). It should be noted that effect sizes were small in comparison to the more robust drivers such as BMI and the polygenic score of CRP.
Our findings put into perspective the role of previously identified inflammation drivers and suggest that additional intra-individual characteristics exist which impact maternal immune marker levels, including genetic mechanisms. We therefore analyzed the polygenic score of CRP, calculated based on a recent genome-wide association study (GWAS) of CRP (35), as a predictor of HS-CRP. Our analysis revealed that the polygenic score of CRP explained a remarkable 14.1% of the variance in serum HS-CRP in the current cohort, in comparison to 16.3% of the variance in the original GWAS loci (35). In line, we showed that preconception cytokines are strongly correlated with pregnancy cytokine levels, further supporting the notion of an individual setpoint. Together, our findings are suggestive of an individual inflammatory marker setpoint which is, in part, genetically driven. Interestingly, while the release of pro-inflammatory markers is considered an orchestrated event, and CRP production is stimulated by pro-inflammatory cytokines, the polygenic score of HS-CRP revealed no association with the maternal cytokine index.
Strengths of the current study include the combined sample size of more than 12,000 participants in a discovery cohort, two replication cohorts, and a pre-pregnancy cohort, each including the general obstetric population, hence providing generalizable results. In addition, inflammatory marker measurement was performed by the same company using identical assays in all cohorts, minimizing technical and qualitative variance. Additionally, inflammatory markers were measured preconceptionally, providing the unique opportunity to assess pregnancy-related inflammatory marker changes. Moreover, cytokines were assessed individually to account for individual effects as well as combined into a maternal cytokine index, to capture combined inflammatory activity of the cytokines (59). This study has several limitations. First, it should be noted that peripheral blood cytokine levels may not accurately reflect immune activity at localized sites such as the placenta or amniotic cavity. Hence, immune disruption at the tissue microenvironment may occur without inducing systemic changes and may not be detected systemically by peripheral cytokine levels. As well, the current study focused on a limited number of pro-inflammatory markers, and additional cytokines or immune cells were not included. Our study is restricted by the limited knowledge of the timing of self-reported infections, which, due to the timing of questionnaires during gestation could have occurred at any point between 0–12 weeks prior to biomarker assessment. In addition to the PGS of CRP, no other PGSes of included cytokines could be used due to lack of well-powered GWASes that show high variance explained by the PGS. Moreover, as not all cytokines were measured in the replication cohorts and to maximize comparability, we assessed individual cytokines rather than a maternal cytokine index in the replication cohorts. These showed similar patterns compared to the individual cytokines in the discovery cohort. We leveraged data from existing, large, high-quality pregnancy cohort, yet these were not originally designed for replication. While sampling and storage methods were consistent within cohorts and highly overlap between cohorts, we cannot preclude that methodological differences in study design and sample handling may have impacted our findings as the strength of within-person inferences may vary depending on the timing and number of repeated measures per individual. As well, variability in sampling number and timing limited the option to meta-analyze the four cohorts. Yet, the application of multiple replication cohorts allowed external validation of our findings. Lastly, given that the current findings stem from population-based pregnancy studies, their generalizability to specific subpopulations such as patients with chronic diseases is limited.
In conclusion, the current study mapped patterns and drivers of maternal inflammatory markers preconception and during pregnancy in the largest population to date. Our findings suggest the presence of an individual immune marker setpoint that is driven mostly by within-individual effects, including a genetic predisposition of CRP levels and a metabolic component. Pre-pregnancy BMI explained 9.6% of the variance in CRP, but less than 1% of the variance in cytokines. Other previously identified drivers, including tobacco use and parity, explained less than 1% of inflammatory marker variance. Additionally, our findings corroborate differential regulatory mechanisms of HS-CRP and cytokines, based on their low correlations, different trajectories, and distinct drivers throughout gestation. Our findings provide important considerations for marker selection and suggest that HS-CRP and cytokines may not be used interchangeably. As well, future studies should explore genetic and epigenetic components that explain the high intra-individual correlation in inflammatory marker levels, which could also lead to the identification of long-term, stable predictors of the immunological state of an individual. Future research should further investigate the association between systemic inflammatory marker levels and how this is related to localized immune mechanisms, e.g. in the placenta. Measuring peripheral inflammatory biomarkers has been proposed as potentially clinically relevant for capturing immune-related mechanisms underlying adverse pregnancy outcomes. However, taking together previous findings from us and others on the association between CRP and cytokines with adverse pregnancy outcomes (3, 60, 61), and our current findings showing largely stable trajectories with an individual setpoint, it is unlikely that these markers are reliable predictors of adverse pregnancy outcomes or effective tools for stratifying pregnant individuals based on immune-related mechanisms. At present, metabolic factors and clinical history offer more robust predictive insights.
Materials and methods
Study design and participants
Discovery cohort
The current study is embedded in the Generation R Study, a large-scale population-based prospective pregnancy cohort from early fetal life onward conducted in Rotterdam, the Netherlands. Pregnant women were recruited between April 2002 and January 2006 (62, 63). The enrollment procedure has been described elsewhere (64). The Generation R study was approved by the Medical Ethical Committee (MEC 198.782/2001/31) of the Erasmus Medical Center, Rotterdam, the Netherlands. All participants provided written informed consent. Inclusion criteria were that the woman needed to i) be pregnant, ii) live in Rotterdam, the Netherlands and iii) have a delivery date between April 2002 and January 2006. Of 9,778 participants enrolled during pregnancy, 72.3% were enrolled prior to 18 weeks gestation (n=7,069). Participants were excluded from the current analysis in case of induced abortion, non-live birth and loss to follow-up (n=149, 1.5%) and if no cytokine measurement at any point during gestation was available (n=1,644, 16.8%), if the assay showed low bead count or if there was insufficient sample volume available (n=53, 0.5%). A total of 8,082 participants were included in the current study. The study design is shown in Figure 1A.
Replication Cohort I
The first replication cohort is the Generation C Study conducted at the Mount Sinai Hospital in New York City, USA (65). In short, all pregnant individuals who received obstetrical care between April 2020 and February 2022 in the Mount Sinai Health System were eligible for participation. The study was approved by the Icahn School of Medicine at Mount Sinai Institutional Review Board (IRB-20-03352), reviewed by the US Centers for Disease Control and Prevention (CDC) and consistent with applicable federal law and CDC policy. All participants provided informed consent. In total, 3,157 participants were included in the Generation C Study. For the current study, participants were included who had given birth to a liveborn singleton infant and with at least one cytokine measurement at any point during gestation (n=2,535, 80.3%).
Replication Cohort II
The second replication cohort was the Brabant Study conducted at various midwife practices in the Netherlands (66). In short, pregnant individuals were recruited at 8–10 weeks gestation among midwifery practices in the South-East of the Netherlands between June 2018 and January 2023. All participants provided informed consent. The Brabant study was approved by the Medical Ethical Committee of the Máxima Medical Center Veldhoven (protocol number NL64091.015.17). Inclusion criteria included i) >18 years old, ii) pregnant, iii) understanding of the Dutch or English language, and iv) a first antenatal visit before 12 weeks gestation. In total, 2863 participants were included in the Brabant Study. High-sensitivity cytokine assays were performed in a subset of participants. For the current study, participants were included if at least one cytokine measurement at any point during gestation was available (n=587, 20.5%).
Pre-pregnancy Cohort
The Generation R Next study cohort is a population-based prospective cohort study from preconception onwards, conducted in Rotterdam, the Netherlands. Recruitment started in August 2017 and is currently ongoing. Participants of reproductive age planning to have children are recruited at general practitioners. Pregnant women are invited to the research center in the first and third trimester. The Generation R Next study was approved by the Medical Ethical Committee (MEC-2016-589, December 2016) of the Erasmus Medical Center, Rotterdam, the Netherlands. All participants provided written informed consent. A total of 1,270 participants were included in the current study with at least one cytokine measurement available preconception or in the first trimester.
Sample collection
Discovery cohort
In the Generation R Study, maternal venous blood samples were collected as part of routine prenatal visits at the midwife practice with the first blood sample collected at median 13.2 weeks (95% range: 9.6-17.6 weeks, n=6,072) and the second blood sample at median 20.4 weeks (95% range: 18.5-23.3 weeks, n=7,395), with an overall median of 19.2 weeks (95% range: 10.6 – 23.1 weeks, n=13,467). Blood was collected through ante-cubital venous puncture, temporarily stored at room temperature and transported to the regional laboratory (Star-MDC, Rotterdam, the Netherlands) within three hours of collection (63). Blood samples were centrifuged for 10 minutes, and the serum was aliquoted into 250ul into polypropylene Micronic tubes, and immediately stored at -80°C. Freeze-thaw cycles were avoided. Samples were bar coded with a unique laboratory number.
Replication Cohort I
In the Generation C Study (Replication Cohort I), blood specimens were obtained as part of routine blood draws during prenatal visits or at admission to the labor and delivery unit, ranging from one to six specimens collected from each participant. Blood samples were collected at median 27.7 weeks (95% range: 6.9-40.6 weeks, n=3,319). Blood was collected through ante-cubital venous puncture, temporarily stored at room temperature and processed within 4 hours (median = 1.6 hours; IQR = 1.7 hours). Blood samples were centrifuged for 10 minutes, aliquoted into 500 µl vials and stored at -80°C until further analysis. Freeze-thaw cycles were avoided. Samples were bar coded with a unique laboratory number.
Replication Cohort II
In the Brabant Study (Replication Cohort II), blood sample collection was performed by the regional organization responsible for blood collection for primary and secondary care laboratories (De Bloedafname). Blood samples were collected at three timepoints during gestation, namely at 12, 20 and 28 weeks, with an overall median of 20.2 weeks (95% range: 12-29.5 weeks, n=1,170). Blood samples were centrifuged for 10 minutes, and the serum was aliquoted into 250ul aliquots and stored at -80°C by the laboratory of clinical chemistry and hematology of the Máxima Medical Centre Veldhoven. Freeze-thaw cycles were avoided. Samples were bar coded with a unique laboratory number.
Pre-pregnancy Cohort
In the Generation R Next Study (Pre-pregnancy Cohort), blood sample collection was performed during an appointment at the research center prior to pregnancy at median 11.4 weeks prior to conception (95% range: 74.5-3.6 weeks prior to conception, n=676), and two blood samples during pregnancy in the first trimester at median 8.4 weeks gestation (95% range: 6.4-12.9 weeks, n=1,103) and in the third trimester at median 30.3 weeks gestation (95% range: 29-33.6 weeks, n=1,067). Blood samples were centrifuged for 10 minutes, and the serum was aliquoted into 250ul into polypropylene Micronic tubes, and immediately stored at -80°C. Freeze-thaw cycles were avoided. For the current analyses, only preconception and first trimester samples were included.
Inflammatory marker measurement
Discovery cohort
High-sensitivity C-reactive protein
A high sensitivity assay was used to measure HS-CRP to increase the sensitivity in the low range with improved precision at low CRP concentrations (67). HS-CRP assays were performed by the Department of Clinical Chemistry of the Erasmus MC, using an immunoturbidimetric assay on the Cobas 6000 analyzer (Roche Diagnostics). The lowest level of detection was 0.2 mg/L. The early and mid-pregnancy samples were run in two separate batches (2006 and 2022), using different assay kits and lot numbers. HS-CRP measurement of the first batch of mid-pregnancy blood samples has been described elsewhere (68).
Pilot: cytokine selection
In our search for factors that impact the maternal immune system during pregnancy, we first set out to construct a measure of maternal immune activation. Given that immune markers interact and are involved in specific pregnancy processes, it is of interest to quantify a panel of robust cytokines that have the potential to capture both low-grade and acute systemic inflammation. Quantification of 14 cytokines (GM-CSF, IL-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, IL-17A, IL-23, IFN-γ and TNF-α) as part of a pilot (n=100) revealed excellent performance with no values below the detection limit. Several outliers were excluded (Supplementary Figure 1B). The final panel of cytokines was selected based on several biological and statistical considerations. A data-driven approach was used to select markers that showed predicted values and associations in our pilot study, and markers that showed considerable variance. Clustering analyses showed correlations between cytokines including IL-1β, IL-2, IL-4, IL-6 IL-13, IL-17A and IFN-γ, while TNF-α and IL-8 did not correlate with other cytokines (Supplementary Figure 1A). Cytokines were differently distributed, with IL-6 showing values in the low range and IL-23 showing values in a higher range (Supplementary Figure 1C). Additionally, based on literature suggesting a role in pregnancy processes for IL-1β, IL-6, IL-17A, and IFN-γ (18–20), a final selection was made. Inflammatory cytokines IL-1β, IL-6, IL-17A, IL-23 and IFN-γ were measured in the entire cohort to establish pro-inflammatory marker patterns in early and mid-pregnancy (Supplementary Figure 1C).
Processing and quality assurance of cytokine data
Cytokine analyses were performed by Eve Technologies Corp., Calgary, Canada. A pilot study of 100 serum samples was performed using the Human High Sensitivity 14-Plex Discovery Assay which simultaneously quantified levels of 14 cytokines (granulocyte-macrophage (GM-) colony-stimulating factor (CSF), interferon (IFN)-γ, interleukin (IL)-1β, IL-2, IL-4, IL-5, IL-6, IL-8, IL-10, IL-12p70, IL-13, IL-17A, IL-23, and tumor necrosis factor (TNF)-α (pg/mL). Pilot results showed 100% detection within the standard curve, indicating high sensitivity and high assay performance. IL-1β, IL-6, IL-17A, IL-23, and IFN-γ were measured in the full Generation R cohort using the Human High Sensitivity T-Helper Cells Custom 5-plex assay from Millipore (Millipore, St. Charles, MO, USA) on the Luminex™ 100 system (Luminex, Austin, TX, USA). Serum samples were analyzed across 145 plates in 37 batches. Maternal and fetal characteristics including maternal age, BMI, fetal sex, parity, and birthweight were randomly distributed across plates (Supplementary Figure 1E). Several measures were taken to harbor the quality and limit variance based on laboratory conditions, namely: i) quality control (QC) samples were included in each assay session. All QC samples used in the study were from the same lot number, and reconstituted and aliquoted per kit protocol at the start of the study; ii) all analyses were run on the same kit and lot specific reagents (Kit Lot number 3891089; Detection Antibodies Lot number 3692805); iii) to minimize potential intra-assay variables, all analyses were run on identical Luminex instrument and calibration materials by the same technician, and iv) ancillary fluidic components including calibration material and sheath fluid were from the same lot number for the entire study and v) sample positions were re-oriented across assay sessions to evenly distribute any potential batch effects. Intra-assay variability was assessed by running the kit-supplied QC material in triplicate during each assay session. The coefficient of variability (CV) was 4.9%. Inter-assay variability was IL-1β: 8.8%, IL-6: 9.0%, IL-17A: 12.9%, IFNγ: 14.1%, IL-23: 8.3%, all well within the manufacturer’s range (20%). We found no effect of cytokine batch (Supplementary Figure 1D), however, to minimize technical variability, we included batch as a fixed effect in the main analyses. The observed analyte concentrations are generated based on cubic spline regression analyses using the expected concentration values of the standard, as defined in the manufacturer’s instructions for use, and the Fluorescence Intensity (FI), as suggested by Breen et al. (69). Cytokine concentrations were estimated from the FI using specific calibration curves for each analyte, constructed from standard samples in the reference lot. Less than 0.5% of the samples fell outside of the standard curve on the low end based on the cubic spline curve for that specific analyte. These values were substituted with the lowest measured value for that specific analyte (70). Immunoassay performance is shown in Supplementary Table 10. Cytokines IL-1β, IL-6, IL-17A, IL-23, IFN-γ and HS-CRP were log2 transformed, to satisfy the normality assumption for downstream analyses. Normalization resulted in well-aligned distributions across cohorts (Supplementary Figure 6).
Potential HAAA interference
Circulating human antibodies that are reactive with animal antibodies (human anti-animal antibodies, or HAAA), may pose a source of immunoassay interference. HAAA are high affinity, specific polyclonal antibodies produced by the human immune system against a specific animal immunogen (heterophile antigen), or due to non-iatrogenic causes such as pharmaceutical agents or vaccines (71). The most encountered HAAA are human anti-mouse antibodies (HAMA), considered to be present in about 5-10% of the population, with a potential increase in an inflammatory population as antibody-based therapeutics might be more frequent. In samples with HAMA, the anti-mouse antibodies can bind and cross-link capture antibodies to detection antibodies in the absence of the analyte, resulting in artificially high fluorescence values in immunoassays based on mouse-antibodies, such as the assay that we used (72). In the Milliplex assay, potential HAMA interference is indicated in samples with a median fluorescent intensity value above 500 across the five cytokines. Characteristics of inflammatory markers with and without HAAA samples are shown in Supplementary Table 11. However, as we are interested in states of immune activation, where increased cytokine levels are to be expected, we further investigated the behavior of samples with possible HAMA interference in a sensitivity analysis.
Replication Cohorts and Pre-pregnancy Cohort
In both replication cohorts and the pre-pregnancy cohort, IL-1β, IL-6 and IL-17A were assessed using the High Sensitivity T-cell Discovery Array 3-Plex (Millipore, St. Charles, MO, USA) at Eve Technologies using the Bio-Plex™ 200 system (Bio-Rad Laboratories, Inc., Hercules, CA, USA). For the pre-pregnancy cohort, IL-23 and IFN-y were also measured as part of this assay. For Replication Cohort I, HS-CRP was also analyzed as part of this assay. For Replication Cohort II, CRP was measured by the laboratory of clinical chemistry and hematology of the Máxima Medical Centre Veldhoven. Similar processing and quality assurance checks were performed as the discovery cohort. For the pre-pregnancy cohort, HS-CRP was measured by the Department of Clinical Chemistry of the Erasmus MC, similar to the Discovery Cohort.
Pre-pregnancy characteristics and pregnancy circumstances
Data collected at enrollment (pre-pregnancy characteristics)
Questionnaires at enrollment were used to assess maternal demographic variables, including maternal age at conception, pre-pregnancy body mass index (BMI) (kg/m2), education level (no education, primary education, secondary education), national background (Dutch, Non-Dutch), parity (nulliparous, multiparous), immune-related disease (HIV, eczema, systemic lupus erythematosus (SLE), intestinal disorder, pre-gestation diabetes, multiple sclerosis, rheumatism) and household income (low: <€2,220/month or high: >€2200/month, based on the average household income in Rotterdam in 2005). In addition, prenatal maternal psychopathology was assessed at enrollment using the Brief Symptom Inventory (73). A Global Severity Index (GSI) was calculated indicating prenatal maternal psychopathology with higher scores meaning more problems and referred to as ‘Maternal Psychopathology’ throughout the manuscript. These pre-existing maternal characteristics are referred to as pre-pregnancy characteristics.
Data collected during pregnancy (pregnancy circumstances)
Data on maternal tobacco use (never smoked, tobacco use pre-pregnancy, tobacco use during pregnancy), alcohol consumption (never drank, alcohol use pre-pregnancy, occasional alcohol use during pregnancy, frequent alcohol use during pregnancy) and substance use during pregnancy (yes/no), was obtained through questionnaires collected at three times during pregnancy. Questionnaire data obtained prior to or at the time of blood sample collection was included. Information on fetal sex was obtained from medical records. Gestational age of the sample was analyzed as a continuous variable, and further categorized into season following European references (spring, summer, fall, winter). These pregnancy-specific characteristics are referred to as pregnancy circumstances.
Infection score
Questionnaire data collected at three times during pregnancy was used to create a self-rated continuous infection score, as described elsewhere (74). An infection score of 0 indicates no infection was reported. An infection score of 1 and higher indicates the participant reported any one of the following infections at least once: upper respiratory infections (pharyngitis, rhinitis, sinusitis, ear infection), lower respiratory infections (pneumonia, bronchitis), gastrointestinal infections (diarrhea, enteritis), cystitis/pyelitis, dermatitis (boils, erysipelas), eye infections, herpes zoster, flu, sexually transmitted diseases (STD), and a period of fever (>38°C/100.4°F) in the 2–3 months prior to blood draw.
Replication Cohort I
In Replication Cohort I, information on maternal demographic variables was obtained through questionnaires collected at enrollment. These included maternal age at conception, pre-pregnancy body mass index (BMI) (kg/m2), national background (white, non-White), parity (nulliparous, multiparous) and insurance status (private/self-pay, public). These pre-existing maternal characteristics are referred to as pre-pregnancy characteristics. Information on fetal sex, twin pregnancy, birthweight and gestational age at birth was obtained from medical records. These pregnancy-specific characteristics are referred to as pregnancy circumstances.
Replication Cohort II
In Replication Cohort II, information on maternal demographic variables was obtained through questionnaires collected at each visit at 12, 20, and 28 weeks. These included maternal age at conception, pre-pregnancy body mass index (BMI) (kg/m2), national background (Dutch, non-Dutch), parity (nulliparous, multiparous), education level (no education, primary education, secondary education), immune-related disease (HIV, eczema, systemic lupus erythematosus (SLE), intestinal disorder, pre-gestation diabetes, multiple sclerosis, rheumatism) and employment status (employed, unemployed). These pre-existing maternal characteristics are referred to as pre-pregnancy characteristics. Information on fetal sex, birthweight and gestational age at birth was obtained from medical records. These pregnancy-specific characteristics are referred to as pregnancy circumstances.
Computation of polygenic score of CRP
Discovery cohort
Genotype data
Maternal samples were used for DNA extraction, as detailed elsewhere (63). Parents in Generation R were genotyped in two batches using the Illumina Global Screening Multi-Disease Array (GSA-MD) v2 (1,530 mothers in 2019/2020) and GSA-MD v3 (10,491 mothers and fathers in 2022) platforms. Both batches underwent extended quality control checks. Genotype call rate was checked in two rounds, the initial with a threshold of 95% and then with 97.5%. Single nucleotide variants (SNVs) that failed the one-sided HWE test (p ≤ 1*10-5) were removed, as well as samples with evidence of excess heterozygosity, gender mismatch, unexpected genetic duplicates and with unexpected familial bonds. After the extended quality control checks, the batches were merged, leaving 11,742 parents (including 7,256 mothers) and 660,868 SNVs. After merging the two batches, the unmapped single nucleotide polymorphisms (SNPs) were imputed to a reference panel (1000 Genomes Project - phase3 version 5, build 38) by integrating SHAPEIT and Minimac4 programs in an in-house Odyssey pipeline. The imputation quality threshold was set to 0.8 and the MAF threshold was set to 0.01.
CRP polygenic score
A recent large-scale gene-wide association study (GWAS) with 575,531 participants was used to construct a PGS of CRP based on imputed genotypic data (35). The summary statistics were obtained from the GWAS study catalog (ID: GCST90029070). The PGS was calculated for participants of any ancestry (n=5,938) and for participants of European descent (n=4,238) separately. LDpred2, the latest version of LDpred that offers better and more robust predictions, was used to calculate the PGS (75, 76). LDpred2 is implemented within the R-package bigsnpr (77) and derives PGSes based on summary statistics and Linkage Disequilibrium (LD) information from an external reference panel (76). The variants in the summary statistics were matched to the ones in the LD reference and the maternal genotypes. The final dataset included 1,044,979 SNPs. We used a genome-wide BED file with the maternal genotypes, HapMap3 variants with individual LD matrix in blocks and LDpred2-auto (one variant of LDpred2) to automatically estimate p (the proportion of causal variants) and h2 (the SNP heritability) from the summary statistics. After estimating h2, LDpred2-auto was run with 30 iterations. A sequence of 30 values from 10−4 to 0.2 equally spaced on a log scale were used as initial values for p. A shrinkage coefficient of 0.95 was used and effects sizes were forced to go through 0 first before changing sign in consecutive iterations to prevent instability of the Gibbs sampler as suggested by Privé (78). To get the final effects, only chains that passed the quality control filtering were used. The calculated score was then standardized to z-scores (mean 0, 1 SD) and residualized on the first 10 principal components. The R2 change between the null model and the model including the PGS of CRP is shown in Supplementary Table 12. A higher PGS of CRP indicates a higher genetic risk for elevated CRP levels. Said et al. (35) showed that the independent variants within the UK Biobank GWAS loci explained 16.3% of the variance in CRP levels. Prior studies have shown that the PGS of CRP is associated with cardiovascular outcomes and performs well in multiple cohorts (79, 80).
Replication Cohorts
No genotyping data was available.
Computation of maternal cytokine index
Discovery cohort
Cytokines are part of complex networks. To account for immune marker inter-relationships, a composite score of inflammatory markers was composed for each sample: the maternal cytokine index. Collapsing the inflammatory marker data into a maternal cytokine index allowed to capture the shared variance across cytokines, and is in line with prior approaches (38, 39). Principal component analysis (PCA) was used as a means of dimensionality reduction using the ‘prcomp’ function from the ‘stats’ package in R. The principal components represent linear combinations of the cytokine data. A higher loading of a cytokine on a principal component indicates increased contribution of that specific cytokine to the overall variability. In addition, PCA addresses multicollinearity by orthogonalizing the cytokines into uncorrelated principal components. This improves the stability and reliability of subsequent analyses. PCA revealed two distinct inflammatory marker patterns. On the one hand, cytokines IL-1β, IL-6, IL-17A, IL-23, and IFN-γ loaded highly on the first principal component (PC) (84%, 66%, 89%, 77%, 89%, respectively), accounting for 55% of the variance. In turn, CRP loaded highly on the second PC (98%). Additionally, IL-6, but not other immune markers, loaded on the third PC (68%). Adding the third PC did not substantially improve model fit and we did not consider the third PC to reflect a profile of biological relevance. An elbow plot suggested that two PCs should be taken into account. Hence, only the first two PCs were considered. Cytokines IL-1β, IL-6, IL-17A, IL-23, and IFN-γ were used to create the maternal cytokine index, defined as the eigenvalue, or singular value decomposition, aggregated across the cytokines, equivalent to the first principal component. Due to its high loading on the second PC, HS-CRP was not included in the maternal cytokine index and was assessed separately. The maternal cytokine index was included as a continuous score in analyses.
Replication Cohorts
Not all cytokines of the discovery cohort panel were measured in the replication cohorts and no maternal cytokine index was created in the replication cohorts. Individual cytokines were assessed to maximize comparability to the discovery cohort.
Statistical analysis
Analyses were performed using the R Statistical Software (version 4.1.2) (81). Visualizations were made with the ggplot2 package (82). Visual representation of study design was created using Biorender.
Demographics
Demographic characteristics were presented as means with standard deviation (SD) for normally distributed continuous data, medians with inter-quartile range (IQR) for non-normally distributed continuous data and as numbers (percentages) for categorical data.
Outlier analysis
Outlier samples for inflammatory markers were identified using unsupervised hierarchical clustering, based on Pearson coefficient and average distance metric, and principal component analysis (PCA). Samples more than three SD from the grand mean of the first PC were considered potential outlier samples. Considering that these samples might reflect levels of acute or chronic inflammation, they were included in the main analysis and excluded in a sensitivity analysis.
Missing data and imputation
In the Discovery Cohort, covariates with missing data included parity (1%), national background (5%), education (8%), alcohol use (10%), tobacco use (12%) and substance use (12%), pre-pregnancy BMI (18%), maternal psychopathology (22%) and household income (22%). In Replication Cohort I, missing data included insurance status (0.2%) and fetal sex (10.4%). In Replication Cohort II, missing data included education level (1.9%), parity (1.9%), employment status (2.0), tobacco use (2.2%), alcohol use (2.2%) and fetal sex (10.7%). Missing covariate data were imputed using the Mice package in R (100 iterations, 30 datasets) and analyses were conducted across the pooled datasets following Rubin’s rules.
Aim 1: mapping dynamics throughout gestation
Group-level inflammatory marker levels were compared between timepoints using an unpaired t-test. Correlations among inflammatory markers were assessed using Pearson’s r correlation coefficient across all samples, as well as at timepoint 1 and 2 separately. In addition, the correlation between inflammatory markers and the maternal cytokine index was assessed across all samples. Correlations of inflammatory markers between timepoint 1 and 2 were assessed pairwise among participants with repeated measures. Inflammatory marker trajectories were modeled with (non-)linear generalized additive models (GAM) (formula = y ~ s[x, bs = “cs”]) using the ‘ggplot’ package in R. Inflammatory markers were centered and scaled by subtracting the column means from each sample and dividing by their standard deviation using the ‘scale’ function in R. Next, variance partition analysis was performed to assess potential drivers of the variation in inflammatory marker expression using the ‘variancePartition’ package in R (83). Categorical predictors were modeled as random intercepts to account for variance across groups and continuous predictors were modeled as fixed effects to capture systematic variance in the outcome. Collinearity between predictors was assessed with the ‘fitVarPartModel’ function.
Aim 2: drivers of maternal inflammatory markers
Linear mixed-effects regression models were applied to investigate the association between potential drivers (exposures) and HS-CRP and the maternal cytokine index (outcomes) in separate models. Linear mixed models (LMM) were performed with log-normalized inflammatory markers as outcome variables. Gestational age at sampling and cytokine batch were included as fixed effects. Participant was included as random intercept to account for within-person variability over time, capturing intra-individual changes among participants with repeated measures. Three separate models were run; model 1 includes maternal age, pre-pregnancy BMI, PGS of CRP, national background, household income, parity, maternal psychopathology, tobacco use, alcohol use, substance use, fetal sex, and season of first blood draw as fixed effects. The infection score was interpreted from a separate model as we hypothesize that BMI may be on the pathway of pregnancy infection and inflammatory markers (Model 2) (84, 85). Models 1 and 2 were run in a subset of participants with complete CRP PGS for all ancestry (n=5,938 participants, n=10,157 samples). The PGS of European ancestry was interpreted from a separate model among participants with a complete PGS of European ancestry (n=4,238, n=7,381 samples) (Model 3).
The linear mixed effects regression model equations were:
- Model 1: Maternal cytokine index / HS-CRP / Individual cytokines ~ maternal age + pre-pregnancy BMI + CRP PGS (all ancestry) + national background + household income + parity + maternal psychopathology + tobacco use + alcohol use + substance use + fetal sex + season of first blood draw + cytokine batch + gestational age of the sample + (1 | participanti) + i.
- Model 2: Maternal cytokine index / HS-CRP / Individual cytokines ~ maternal age + pre-pregnancy BMI + CRP PGS (all ancestry) + national background + household income + parity + maternal psychopathology + tobacco use + alcohol use + substance use + fetal sex + season of first blood draw + infection score + cytokine batch + gestational age of the sample + (1 | participanti) + i.
- Model 3: Maternal cytokine index / HS-CRP / Individual cytokines ~ maternal age + pre-pregnancy BMI + CRP PGS (European ancestry) + national background + household income + parity + maternal psychopathology + tobacco use + alcohol use + substance use + fetal sex + season of first blood draw + cytokine batch + gestational age of the sample + (1 | participanti) + i.
The ‘i’ subscript indicates the participant. The R packages lme4 and lmertest were used to fit the models (86). Linear mixed-effects models were fit with the restricted maximum likelihood as estimation method. Assumptions were checked for all models. Residuals exhibited linearity, homoscedasticity and approximate normality. Collinearity between variables in the models was assessed based on the variance inflation factor (VIF). If the VIF>3, variables were considered collinear. All variables were below VIF =2. Benjamini-Hochberg correction was applied per outcome and per aim to correct for multiple testing (87). Both unadjusted and corrected p-values are shown in the tables. Statistical significance is considered if q<0.05 after multiple testing correction. P-values reported within the manuscript are corrected for multiple testing.
Delta between timepoints
The change in inflammatory marker levels between timepoints can be used to provide insight in inflammatory triggers that occurred between two timepoints. A continuous delta was computed for HS-CRP and the maternal cytokine index by subtracting the measurement of the first timepoint from the measurement of the second timepoint. Canonical correlation analysis was performed to assess the correlation between predictors and the delta HS-CRP and delta maternal cytokine index. Canonical correlation analysis was performed using the ‘variancePartition’ in R which allows the inclusion of both continuous and discrete variables in one formula. Additionally, participants were divided into three groups based on the HS-CRP delta and the maternal cytokine index delta. The groups were defined as the <5th, 5th-95th and >95th quantiles of the HS-CRP delta and maternal cytokine index delta, resulting in a high-low (high levels at T1 and low at T2; 5th quantile of the delta), stable, and low-high group (low levels at T2 and high at T2; 95th quantile of the delta). A sensitivity analysis was performed to determine the associations between predictors and HS-CRP and the maternal cytokine index among samples of participants with a high absolute HS-CRP delta (n=435 participants; n=854 samples).
Sensitivity analyses
Several sensitivity analyses were performed: i) excluding samples with potential HAAA interference (n=123 participants; n=163 samples), to determine the associations without possible HAAA interference; ii) excluding samples of participants with immune-related diseases (defined as having any one of the following immune-related diseases: HIV, eczema, systemic lupus erythematosus (SLE), intestinal disorder, pre-gestation diabetes, multiple sclerosis, rheumatism), to determine the associations without potential noise due to inherent immune dysregulation (n=495 participants; n=855 samples); and iii) excluding samples considered outliers based on the outlier analysis described above (n=147 participants; n=174 samples), to determine the associations without cases that may unduly influence the inflammatory marker estimates; and iv) for each cytokine as an outcome, to determine associations between predictors and individual cytokines.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Medical Ethical Committee (MEC 198.782/2001/31) of the Erasmus Medical Center. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
FG: Conceptualization, Data curation, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing. AS: Conceptualization, Validation, Writing – original draft, Writing – review & editing. EI: Data curation, Writing – original draft, Writing – review & editing. A-SR: Conceptualization, Writing – original draft, Writing – review & editing. MGB: Supervision, Writing – original draft, Writing – review & editing. OD: Formal Analysis, Writing – original draft, Writing – review & editing. HEM: Writing – original draft, Writing – review & editing. ES: Writing – original draft, Writing – review & editing. MH: Writing – original draft, Writing – review & editing. RM: Conceptualization, Writing – original draft, Writing – review & editing. WL: Writing – original draft, Writing – review & editing. CC: Writing – original draft, Writing – review & editing. VP: Supervision, Writing – original draft, Writing – review & editing. MB: Conceptualization, Writing – original draft, Writing – review & editing. VB: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing. LW: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study is supported by the NIH (R01MH124776, VB). CC is supported by the European Union’s HorizonEurope Research and Innovation Programme (FAMILY, grant agreement No 101057529; HappyMums, grant agreement No 101057390). MH is supported by Stichting Volksbond Rotterdam and the Netherlands Organization for Health Research and Development (Aspasia grant agreement No 015.016.056; HappyMums, grant agreement No 101057390). The Brabant Study was gifted diagnostic kits from Roche Diagnostics.
Acknowledgments
We gratefully acknowledge the tremendous contribution of all participants of the Generation R Study, Generation C Study, Brabant Study and the Generation R Next Study pregnancy cohorts. We would like to thank the medical staff, biobank, and entire teams for their incredible efforts, including the recruitment, data collection and data analysis of the Generation R and Generation R Next cohort at the Erasmus Medical Center in Rotterdam, the Generation C cohort at the Icahn School of Medicine at Mount Sinai in New York City, and the Brabant Study cohort at Tilburg University in Tilburg. The Generation R infrastructure is located in the Erasmus MC, University Medical Center Rotterdam and functions in close collaboration with the School of Law and Faculty of Social Sciences of the Erasmus University Rotterdam, the Municipal Health Service Rotterdam area, Rotterdam, the Rotterdam Homecare Foundation, Rotterdam and the Stichting Trombosedienst and Artsenlaboratorium Rijnmond (STAR-MDC), Rotterdam. The general design of Generation R and Generation R Next is made possible by financial support from the Erasmus MC, Erasmus University Rotterdam, the Netherlands Organization for Health Research and Development and the Ministry of Health, Welfare and Sport.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
The content is solely the responsibility of the authors and does not necessarily represent the official views of the National Institutes of Health.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1561798/full#supplementary-material
References
1. Simpson E. Immunology: Why the baby isn’t thrown out. Curr Biol. (1996) 6:43–4. doi: 10.1016/S0960-9822(02)00418-9
2. Bilbo SD and Schwarz JM. Early-life programming of later-life brain and behavior: a critical role for the immune system. Front Behav Neurosci. (2009) 3:14. doi: 10.3389/neuro.08.014.2009
3. Spence T, Allsopp PJ, Yeates AJ, Mulhern MS, Strain JJ, and McSorley EM. Maternal serum cytokine concentrations in healthy pregnancy and preeclampsia. J Pregnancy. (2021) 2021:6649608. doi: 10.1155/2021/6649608
4. Hosseini A, Dolati S, Hashemi V, Abdollahpour-Alitappeh M, and Yousefi M. Regulatory T and T helper 17 cells: Their roles in preeclampsia. J Cell Physiol. (2018) 233:6561–73. doi: 10.1002/jcp.v233.9
5. Gilman-Sachs A, Dambaeva S, Salazar Garcia MD, Hussein Y, Kwak-Kim J, and Beaman K. Inflammation induced preterm labor and birth. J Reprod Immunol. (2018) 129:53–8. doi: 10.1016/j.jri.2018.06.029
6. Romero R, Espinoza J, Kusanovic JP, Gotsch F, Hassan S, Erez O, et al. The preterm parturition syndrome. BJOG Int J Obstet Gynaecol. (2006) 113 Suppl 3:17–42. doi: 10.1111/j.1471-0528.2006.01120.x
7. Wei SQ, Fraser W, and Luo ZC. Inflammatory cytokines and spontaneous preterm birth in asymptomatic women: a systematic review. Obstet Gynecol. (2010) 116:393–401. doi: 10.1097/AOG.0b013e3181e6dbc0
8. Sánchez AR, Kupp LI, Sheridan PJ, and Sánchez DR. Maternal chronic infection as a risk factor in preterm low birth weight infants: the link with periodontal infection. J Int Acad Periodontol. (2004) 6:89–94.
9. Ansariniya H, Zare F, Mosaffa N, Idali F, Shabani M, and Hadinedoushan H. Immunologic deviations in recurrent spontaneous abortion mouse model. Am J Reprod Immunol N Y N 1989. (2022) 88:e13631. doi: 10.1111/aji.13631
10. Ruocco MG, Chaouat G, Florez L, Bensussan A, and Klatzmann D. Regulatory T-cells in pregnancy: historical perspective, state of the art, and burning questions. Front Immunol. (2014) 5:389. doi: 10.3389/fimmu.2014.00389
11. Mellor AL, Sivakumar J, Chandler P, Smith K, Molina H, Mao D, et al. Prevention of T cell–driven complement activation and inflammation by tryptophan catabolism during pregnancy. Nat Immunol. (2001) 2:64–8. doi: 10.1038/83183
12. Krishnan L, Guilbert LJ, Wegmann TG, Belosevic M, and Mosmann TR. T helper 1 response against Leishmania major in pregnant C57BL/6 mice increases implantation failure and fetal resorptions. Correlation with increased IFN-gamma and TNF and reduced IL-10 production by placental cells. J Immunol Baltim Md 1950. (1996) 156:653–62. doi: 10.4049/jimmunol.156.2.653
13. Tsuda S, Nakashima A, Shima T, and Saito S. New paradigm in the role of regulatory T cells during pregnancy. Front Immunol. (2019) 10. doi: 10.3389/fimmu.2019.00573
14. Muzzio DO, Soldati R, Rolle L, Zygmunt M, Zenclussen AC, and Jensen F. B-1a B cells regulate T cell differentiation associated with pregnancy disturbances. Front Immunol. (2014) 5:6. doi: 10.3389/fimmu.2014.00006
15. Yoshizaki A, Miyagaki T, DiLillo DJ, Matsushita T, Horikawa M, Kountikov EI, et al. Regulatory B cells control T-cell autoimmunity through IL-21-dependent cognate interactions. Nature. (2012) 491:264–8. doi: 10.1038/nature11501
16. Girardi G, Yarilin D, Thurman JM, Holers VM, and Salmon JE. Complement activation induces dysregulation of angiogenic factors and causes fetal rejection and growth restriction. J Exp Med. (2006) 203:2165–75. doi: 10.1084/jem.20061022
17. Clark DA. Mouse is the new woman? Translational research in reproductive immunology. Semin Immunopathol. (2016) 38:651–68. doi: 10.1007/s00281-015-0553-x
18. Griffith OW, Chavan AR, Protopapas S, Maziarz J, Romero R, and Wagner GP. Embryo implantation evolved from an ancestral inflammatory attachment reaction. Proc Natl Acad Sci U S A. (2017) 114:E6566–75. doi: 10.1073/pnas.1701129114
19. Yockey LJ and Iwasaki A. Interferons and proinflammatory cytokines in pregnancy and fetal development. Immunity. (2018) 49:397–412. doi: 10.1016/j.immuni.2018.07.017
20. Ashkar AA, Black GP, Wei Q, He H, Liang L, Head JR, et al. Assessment of requirements for IL-15 and IFN regulatory factors in uterine NK cell differentiation and function during pregnancy. J Immunol Baltim Md 1950. (2003) 171:2937–44. doi: 10.4049/jimmunol.171.6.2937
21. Ross KM, Dunkel Schetter C, Carroll JE, Mancuso RA, Breen EC, Okun ML, et al. Inflammatory and immune marker trajectories from pregnancy to one-year post-birth. Cytokine. (2022) 149:155758. doi: 10.1016/j.cyto.2021.155758
22. Curry AE, Vogel I, Skogstrand K, Drews C, Schendel DE, Flanders WD, et al. Maternal plasma cytokines in early- and mid-gestation of normal human pregnancy and their association with maternal factors. J Reprod Immunol. (2008) 77:152–60. doi: 10.1016/j.jri.2007.06.051
23. Ferguson KK, McElrath TF, Chen YH, Mukherjee B, and Meeker JD. Longitudinal profiling of inflammatory cytokines and C-reactive protein during uncomplicated and preterm pregnancy. Am J Reprod Immunol N Y N 1989. (2014) 72:326–36. doi: 10.1111/aji.2014.72.issue-3
24. Ferguson KK, Meeker JD, McElrath TF, Mukherjee B, and Cantonwine DE. Repeated measures of inflammation and oxidative stress biomarkers in preeclamptic and normotensive pregnancies. Am J Obstet Gynecol. (2017) 216:527.e1–9. doi: 10.1016/j.ajog.2016.12.174
25. Kronborg CS, Gjedsted J, Vittinghus E, Hansen TK, Allen J, and Knudsen UB. Longitudinal measurement of cytokines in pre-eclamptic and normotensive pregnancies. Acta Obstet Gynecol Scand. (2011) 90:791–6. doi: 10.1111/j.1600-0412.2011.01134.x
26. Romero R, Erez O, Maymon E, Chaemsaithong P, Xu Z, Pacora P, et al. The maternal plasma proteome changes as a function of gestational age in normal pregnancy: a longitudinal study. Am J Obstet Gynecol. (2017) 217:67.e1–2. doi: 10.1016/j.ajog.2017.02.037
27. Jarmund AH, Giskeødegård GF, Ryssdal M, Steinkjer B, Stokkeland LMT, Madssen TS, et al. Cytokine patterns in maternal serum from first trimester to term and beyond. Front Immunol. (2021) 12:752660. doi: 10.3389/fimmu.2021.752660
28. Stokkeland LMT, Giskeødegård GF, Stridsklev S, Ryan L, Steinkjer B, Tangerås LH, et al. Serum cytokine patterns in first half of pregnancy. Cytokine. (2019) 119:188–96. doi: 10.1016/j.cyto.2019.03.013
29. Aghaeepour N, Ganio EA, Mcilwain D, Tsai AS, Tingle M, Van Gassen S, et al. An immune clock of human pregnancy. Sci Immunol. (2017) 2:eaan2946. doi: 10.1126/sciimmunol.aan2946
30. Reyes-Lagos JJ, Peña-Castillo MÁ, Echeverría JC, Pérez-Sánchez G, Álvarez-Herrera S, Becerril-Villanueva E, et al. Women serum concentrations of the IL-10 family of cytokines and IFN-γ Decrease from the third trimester of pregnancy to active labor. Neuroimmunomodulation. (2017) 24:162–70. doi: 10.1159/000480734
31. Christian LM and Porter K. Longitudinal changes in serum proinflammatory markers across pregnancy and postpartum: effects of maternal body mass index. Cytokine. (2014) 70:134–40. doi: 10.1016/j.cyto.2014.06.018
32. Enninga EAL, Nevala WK, Creedon DJ, Markovic SN, and Holtan SG. Fetal sex-based differences in maternal hormones, angiogenic factors, and immune mediators during pregnancy and the postpartum period. Am J Reprod Immunol. (2015) 73:251–62. doi: 10.1111/aji.2015.73.issue-3
33. Hedman AM, Lundholm C, Andolf E, Pershagen G, Fall T, and Almqvist C. Longitudinal plasma inflammatory proteome profiling during pregnancy in the Born into Life study. Sci Rep. (2020) 10:17819. doi: 10.1038/s41598-020-74722-5
34. D’Esposito V, Di Tolla MF, Lecce M, Cavalli F, Libutti M, Misso S, et al. Lifestyle and dietary habits affect plasma levels of specific cytokines in healthy subjects. Front Nutr. (2022) 9:913176. doi: 10.3389/fnut.2022.913176
35. Said S, Pazoki R, Karhunen V, Võsa U, Ligthart S, Bodinier B, et al. Genetic analysis of over half a million people characterises C-reactive protein loci. Nat Commun. (2022) 13:2198. doi: 10.1038/s41467-022-29650-5
36. Ramiro-Cortijo D, de la Calle M, Böger R, Hannemann J, Lüneburg N, López-Giménez MR, et al. Male fetal sex is associated with low maternal plasma anti-inflammatory cytokine profile in the first trimester of healthy pregnancies. Cytokine. (2020) 136:155290. doi: 10.1016/j.cyto.2020.155290
37. Mitchell AM, Palettas M, and Christian LM. Fetal sex is associated with maternal stimulated cytokine production, but not serum cytokine levels, in human pregnancy. Brain Behav Immun. (2017) 60:32–7. doi: 10.1016/j.bbi.2016.06.015
38. Ross KM, Miller G, Culhane J, Grobman W, Simhan HN, Wadhwa PD, et al. Patterns of peripheral cytokine expression during pregnancy in two cohorts and associations with inflammatory markers in cord blood. Am J Reprod Immunol N Y N 1989. (2016) 76:406–14. doi: 10.1111/aji.12563
39. Keenan-Devlin LS, Caplan M, Freedman A, Kuchta K, Grobman W, Buss C, et al. Using principal component analysis to examine associations of early pregnancy inflammatory biomarker profiles and adverse birth outcomes. Am J Reprod Immunol N Y N 1989. (2021) 86:e13497. doi: 10.1111/aji.13497
40. Kooijman MN, Kruithof CJ, van Duijn CM, Duijts L, Franco OH, van IJzendoorn MH, et al. The Generation R Study: design and cohort update 2017. Eur J Epidemiol. (2016) 31:1243–64. doi: 10.1007/s10654-016-0224-9
41. Clendenen TV, Koenig KL, Arslan AA, Lukanova A, Berrino F, Gu Y, et al. Factors associated with inflammation markers, a cross-sectional analysis. Cytokine. (2011) 56:769–78. doi: 10.1016/j.cyto.2011.09.013
42. Norman JE, Bollapragada S, Yuan M, and Nelson SM. Inflammatory pathways in the mechanism of parturition. BMC Pregnancy Childbirth. (2007) 7:S7. doi: 10.1186/1471-2393-7-S1-S7
43. Mor G and Cardenas I. The immune system in pregnancy: A unique complexity. Am J Reprod Immunol N Y N 1989. (2010) 63:425–33. doi: 10.1111/j.1600-0897.2010.00836.x
44. Biancotto A, Wank A, Perl S, Cook W, Olnes MJ, Dagur PK, et al. Baseline levels and temporal stability of 27 multiplexed serum cytokine concentrations in healthy subjects. PloS One. (2013) 8:e76091. doi: 10.1371/journal.pone.0076091
45. Hofmann JN, Yu K, Bagni RK, Lan Q, Rothman N, and Purdue MP. Intra-individual variability over time in serum cytokine levels among participants in the Prostate, Lung, Colorectal and Ovarian Cancer Screening Trial. Cytokine. (2011) 56:145–8. doi: 10.1016/j.cyto.2011.06.012
46. McKay HS, Margolick JB, Martínez-Maza O, Lopez J, Phair J, Rappocciolo G, et al. Multiplex assay reliability and long-term intra-individual variation of serologic inflammatory biomarkers. Cytokine. (2017) 90:185–92. doi: 10.1016/j.cyto.2016.09.018
47. Wu D, Dinh TL, Bausk BP, and Walt DR. Long-term measurements of human inflammatory cytokines reveal complex baseline variations between individuals. Am J Pathol. (2017) 187:2620–6. doi: 10.1016/j.ajpath.2017.08.007
48. Pendeloski KPT, Ono E, Torloni MR, Mattar R, and Daher S. Maternal obesity and inflammatory mediators: A controversial association. Am J Reprod Immunol. (2017) 77:e12674. doi: 10.1111/aji.2017.77.issue-5
49. Friis CM, Frøslie KF, Røislien J, Voldner N, Godang K, Ueland T, et al. The interleukins IL-6 and IL-1Ra: a mediating role in the associations between BMI and birth weight? J Dev Orig Health Dis. (2010) 1:310–8. doi: 10.1017/s204017441000036x
50. Fried EI, von Stockert S, Haslbeck JMB, Lamers F, Schoevers RA, and Penninx BWJH. Using network analysis to examine links between individual depressive symptoms, inflammatory markers, and covariates. Psychol Med. (2020) 50:2682–90. doi: 10.1017/S0033291719002770
51. Gu Y, Zeleniuch-Jacquotte A, Linkov F, Koenig KL, Liu M, Velikokhatnaya L, et al. Reproducibility of serum cytokines and growth factors. Cytokine. (2009) 45:44–9. doi: 10.1016/j.cyto.2008.10.014
52. Choi J, Joseph L, and Pilote L. Obesity and C-reactive protein in various populations: a systematic review and meta-analysis. Obes Rev Off J Int Assoc Study Obes. (2013) 14:232–44. doi: 10.1111/obr.2013.14.issue-3
53. Esser N, Legrand-Poels S, Piette J, Scheen AJ, and Paquot N. Inflammation as a link between obesity, metabolic syndrome and type 2 diabetes. Diabetes Res Clin Pract. (2014) 105:141–50. doi: 10.1016/j.diabres.2014.04.006
54. Amin MN, Hussain MS, Sarwar MS, Rahman Moghal MM, Das A, Hossain MZ, et al. How the association between obesity and inflammation may lead to insulin resistance and cancer. Diabetes Metab Syndr. (2019) 13:1213–24. doi: 10.1016/j.dsx.2019.01.041
55. Tagoma A, Haller-Kikkatalo K, Roos K, Oras A, Kirss A, Ilonen J, et al. Interleukin-7, T helper 1, and regulatory T-cell activity-related cytokines are increased during the second trimester of healthy pregnancy compared to non-pregnant women. Am J Reprod Immunol N Y N 1989. (2019) 82:e13188. doi: 10.1111/aji.13188
56. Gold DR, Bloomberg GR, Cruikshank WW, Visness CM, Schwarz J, Kattan M, et al. Parental characteristics, somatic fetal growth, and season of birth influence innate and adaptive cord blood cytokine responses. J Allergy Clin Immunol. (2009) 124:1078. doi: 10.1016/j.jaci.2009.08.021
57. Garcia-Serna AM, Morales E, Cantero-Cano E, Norte-Muñoz M, Gil-Buendía MA, Velazquez-Marin J, et al. Cytokine production by newborns: influence of sex and season of birth. Pediatr Res. (2023) 93:526–34. doi: 10.1038/s41390-022-02153-1
58. Lendor C, Johnson A, Perzanowski M, Chew GL, Goldstein IF, Kelvin E, et al. Effects of winter birth season and prenatal cockroach and mouse allergen exposure on indoor allergen-specific cord blood mononuclear cell proliferation and cytokine production. Ann Allergy Asthma Immunol Off Publ Am Coll Allergy Asthma Immunol. (2008) 101:193–9. doi: 10.1016/S1081-1206(10)60209-8
59. Sanders AFP. From womb to brain: rethinking maternal immune activation and its long-term impact. Biol Psychiatry Cognit Neurosci Neuroimaging. (2025) 10:125–6. doi: 10.1016/j.bpsc.2024.12.012
60. Gigase FAJ, Boekhorst MMGBM, Suleri A, Rommel AS, Breen M, Muetzel RL, et al. Maternal immune activation during pregnancy and obstetric outcomes: A population-based cohort study. BJOG Int J Obstet Gynaecol. (2025). doi: 10.1111/1471-0528.18191
61. Hornaday KK, Stephenson NL, Canning MT, Tough SC, and Slater DM. Maternal cytokine profiles in second and early third trimester are not predictive of preterm birth. PloS One. (2024) 19:e0311721. doi: 10.1371/journal.pone.0311721
62. Jaddoe VWV, van Duijn CM, Franco OH, van der Heijden AJ, van Iizendoorn MH, de Jongste JC, et al. The Generation R Study: design and cohort update 2012. Eur J Epidemiol. (2012) 27:739–56. doi: 10.1007/s10654-012-9735-1
63. Kruithof CJ, Kooijman MN, van Duijn CM, Franco OH, de Jongste JC, Klaver CCW, et al. The generation R study: biobank update 2015. Eur J Epidemiol. (2014) 29:911–27. doi: 10.1007/s10654-014-9980-6
64. Jaddoe VWV, van Duijn CM, van der Heijden AJ, Mackenbach JP, Moll HA, Steegers EAP, et al. The Generation R Study: design and cohort update until the age of 4 years. Eur J Epidemiol. (2008) 23:801–11. doi: 10.1007/s10654-008-9309-4
65. Molenaar NM, Rommel AS, de Witte L, Dolan SM, Lieb W, Ibroci E, et al. SARS-CoV-2 during pregnancy and associated outcomes: Results from an ongoing prospective cohort. Paediatr Perinat Epidemiol. (2022) 36:466–75. doi: 10.1111/ppe.12812
66. Meems M, Hulsbosch L, Riem M, Meyers C, Pronk T, Broeren M, et al. The Brabant study: design of a large prospective perinatal cohort study among pregnant women investigating obstetric outcome from a biopsychosocial perspective. BMJ Open. (2020) 10:e038891. doi: 10.1136/bmjopen-2020-038891
67. Roberts WL, Moulton L, Law TC, Farrow G, Cooper-Anderson M, Savory J, et al. Evaluation of nine automated high-sensitivity C-reactive protein methods: implications for clinical and epidemiological applications. Part 2. Clin Chem. (2001) 47:418–25. doi: 10.1093/clinchem/47.3.418
68. Ernst GDS, de Jonge LL, Hofman A, Lindemans J, Russcher H, Steegers EAP, et al. C-reactive protein levels in early pregnancy, fetal growth patterns, and the risk for neonatal complications: the Generation R Study. Am J Obstet Gynecol. (2011) 205:132.e1–12. doi: 10.1016/j.ajog.2011.03.049
69. Breen EJ, Tan W, and Khan A. The statistical value of raw fluorescence signal in luminex xMAP based multiplex immunoassays. Sci Rep. (2016) 6:26996. doi: 10.1038/srep26996
70. Goldstein JM, Valido A, Lewandowski JP, Walker RG, Mills MJ, Messemer KA, et al. Variation in zygotic CRISPR/Cas9 gene editing outcomes generates novel reporter and deletion alleles at the Gdf11 locus. Sci Rep. (2019) 9:18613. doi: 10.1038/s41598-019-54766-y
71. Kricka LJ. Human anti-animal antibody interferences in immunological assays. Clin Chem. (1999) 45:942–56. doi: 10.1093/clinchem/45.7.942
73. de Beurs E and Zitman F. The Brief Symptom Inventory (BSI): Reliability and validity of a practical alternative to SCL-90. Maandbl Geestelijke Volksgezond. (2006) 61:120–41.
74. Suleri A, White T, Blok E, Cecil CAM, Reiss I, Jaddoe VWV, et al. The association between prenatal infection and adolescent behavior: investigating multiple prenatal, perinatal, and childhood second hits. J Am Acad Child Adolesc Psychiatry. (2023). 62(12):1340–50. doi: 10.1016/j.jaac.2023.06.009
75. Vilhjálmsson BJ, Yang J, Finucane HK, Gusev A, Lindström S, Ripke S, et al. Modeling linkage disequilibrium increases accuracy of polygenic risk scores. Am J Hum Genet. (2015) 97:576–92. doi: 10.1016/j.ajhg.2015.09.001
76. Privé F, Arbel J, and Vilhjálmsson BJ. LDpred2: better, faster, stronger. Bioinformatics. (2020) 36:5424–31. doi: 10.1093/bioinformatics/btaa1029
77. Privé F, Aschard H, Ziyatdinov A, and Blum MGB. Efficient analysis of large-scale genome-wide data with two R packages: bigstatsr and bigsnpr. Bioinformatics. (2018) 34:2781–7. doi: 10.1093/bioinformatics/bty185
78. Polygenic scores and inference using LDpred2. Available online at: https://privefl.github.io/bigsnpr/articles/LDpred2.html (Accessed January 15, 2024).
79. Aday AW, Bagheri M, Vaitinadin NS, Mosley JD, and Wang TJ. Polygenic risk score in comparison with C-reactive protein for predicting incident coronary heart disease. Atherosclerosis. (2023) 379:117194. doi: 10.1016/j.atherosclerosis.2023.117194
80. Kappelmann N, Czamara D, Rost N, Moser S, Schmoll V, Trastulla L, et al. Polygenic risk for immuno-metabolic markers and specific depressive symptoms: A multi-sample network analysis study. Brain Behav Immun. (2021) 95:256–68. doi: 10.1016/j.bbi.2021.03.024
81. R Core Team. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. (2020). Available online at: https://www.R-project.org/.
82. Wickham H. ggplot2: elegant Graphics for Data Analysis [Internet]. New York, NY: Springer (2009). doi: 10.1007/978-0-387-98141-3
83. Hoffman GE and SChadt EE. variancePartition: interpreting drivers of variation in complex gene expression studies. BMC Bioinf. (2016) 17:483. doi: 10.1186/s12859-016-1323-z
84. de Oliveira LC, Franco-Sena AB, Rebelo F, Farias DR, Lepsch J, Lima N da S, et al. Factors associated with maternal serum C-reactive protein throughout pregnancy: A longitudinal study in women of Rio de Janeiro, Brazil. Nutr Burbank Los Angel Cty Calif. (2015) 31:1103–8. doi: 10.1016/j.nut.2015.04.006
85. Barbaresko J, Koch M, Schulze MB, and Nöthlings U. Dietary pattern analysis and biomarkers of low-grade inflammation: a systematic literature review. Nutr Rev. (2013) 71:511–27. doi: 10.1111/nure.2013.71.issue-8
86. Bates D, Mächler M, Bolker B, and Walker S. Fitting linear mixed-effects models using lme4. J Stat Software. (2015) 67:1–48. doi: 10.18637/jss.v067.i01
Keywords: pregnancy, immunology, maternal immune activation, inflammatory marker dynamics, cohort study, intra-individual correlation
Citation: Gigase FAJ, Suleri A, Isaevska E, Rommel A-S, Boekhorst MGBM, Dmitrichenko O, El Marroun H, Steegers EAP, Hillegers MHJ, Muetzel RL, Lieb W, Cecil CAM, Pop VJM, Breen M, Bergink V and de Witte LD (2025) Inflammatory markers in pregnancy – identifying drivers in four large cohorts. Front. Immunol. 16:1561798. doi: 10.3389/fimmu.2025.1561798
Received: 16 January 2025; Accepted: 13 May 2025;
Published: 09 June 2025.
Edited by:
Grazisa Rossetti, IFOM - The FIRC Institute of Molecular Oncology, ItalyReviewed by:
Ivana Kawikova, National Institute of Mental Health, CzechiaAlexandra Sitarik, Henry Ford Health System, United States
Copyright © 2025 Gigase, Suleri, Isaevska, Rommel, Boekhorst, Dmitrichenko, El Marroun, Steegers, Hillegers, Muetzel, Lieb, Cecil, Pop, Breen, Bergink and de Witte. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frederieke A. J. Gigase, Zi5naWdhc2VAZXJhc211c21jLm5s
†These authors have contributed equally to this work