 Rafael Tume1
Rafael Tume1 Samar El Sherbiny2
Samar El Sherbiny2 Roberto Bono2
Roberto Bono2 Thomas Gautier3Jean Paul Pais de Barros3,4
Thomas Gautier3Jean Paul Pais de Barros3,4 Tomás Meroño1,5*
Tomás Meroño1,5*- 1Biomarkers and Nutrimetabolomics Laboratory, Department de Nutrició, Ciències de l’Alimentació i Gastronomia, Nutrition and Food Safety Research Institute (INSA), Facultat de Farmàcia i Ciències de l’Alimentació, Universitat de Barcelona, Barcelona, Spain
- 2Department of Public Health and Pediatrics, University of Turin, Turin, Italy
- 3Center for Translational and Molecular Medicine (CTM) UMR1231, Inserm/Université Bourgogne Europe, Lipness Team, Dijon, France
- 4DiviOmics Platform, UMS 58 BioSand – Université Bourgogne Europe, Dijon, France
- 5Centro de Investigación Biomédica en Red de Fragilidad y Envejecimiento Saludable (CIBERFES), Instituto de Salud Carlos III, Madrid, Spain
Lipopolysaccharide (LPS) from Gram-negative bacteria has been one of the most studied pathogen-associated molecular patterns triggering rapid inflammatory reactions. However, evidence shows that not all LPS molecules are proinflammatory (“bad”), and that “good” LPS from gut commensal bacteria exert immunomodulatory actions. The Limulus amebocyte lysis test commonly used to quantify LPS in circulation, only targets “bad” LPS, when not inactivated by plasma components. Use of other methods showed healthy subjects featuring elevated levels of LPS (suggesting predominance of “good” or inactive LPS in circulation). This review aims to summarize the evidence supporting the higher abundance of “good” LPS coming from gut commensals of healthy individuals and their potential impact in human health.
Introduction
Lipopolysaccharides: structure, function, and heterogeneity
Lipopolysaccharides (LPS) are amphipathic glycoconjugates that serve as an essential component of the outer membrane of Gram-negative bacteria. In some cases, these bacteria can comprise up to approximately 65% of the intestinal microbiota, with the majority of LPS being derived from the Bacteroidota and Pseudomonadota (formerly Proteobacteria) phyla (1). LPS performs essential structural functions by providing integrity to the outer membrane and protecting the bacterial cell from adverse conditions (2). Additionally, LPS facilitates adhesion to surfaces through biofilm formation, increasing resistance to antibiotics and promoting colonization. One of its most notable characteristics is its thermal stability, as well as its key role in the pathogenicity of Gram-negative bacteria, distinguishing it from bacterial exotoxins. This is why LPS is also referred to as “endotoxin”. LPS is constantly released during bacterial division and cell lysis, making it a ubiquitous toxin in the environment.
Structure of LPS and LPS types
LPS exhibit structural variations among different bacteria and even among strains of the same species. However, in general terms, they are composed of the following main regions: (Figure 1):
● Lipid A: The hydrophobic portion anchored to the membrane, which serves as the primary virulence factor by binding to Toll-like receptor 4 (TLR4), triggering an inflammatory signaling cascade. It contains, among others, several acyl chains, including specific hydroxylated fatty acids allowing the branching of secondary acyl chains via ester bonds.
● Core oligosaccharide: An intermediate hydrophilic segment consisting of an inner and outer core, which connects lipid A to the O-antigen.
● O-antigen: A highly variable distal polysaccharide composed of repetitive units of up to eight distinct sugars. It is the main determinant of bacterial antigenicity and serotyping (e.g., Escherichia coli presents around 170 serotypes) (3).
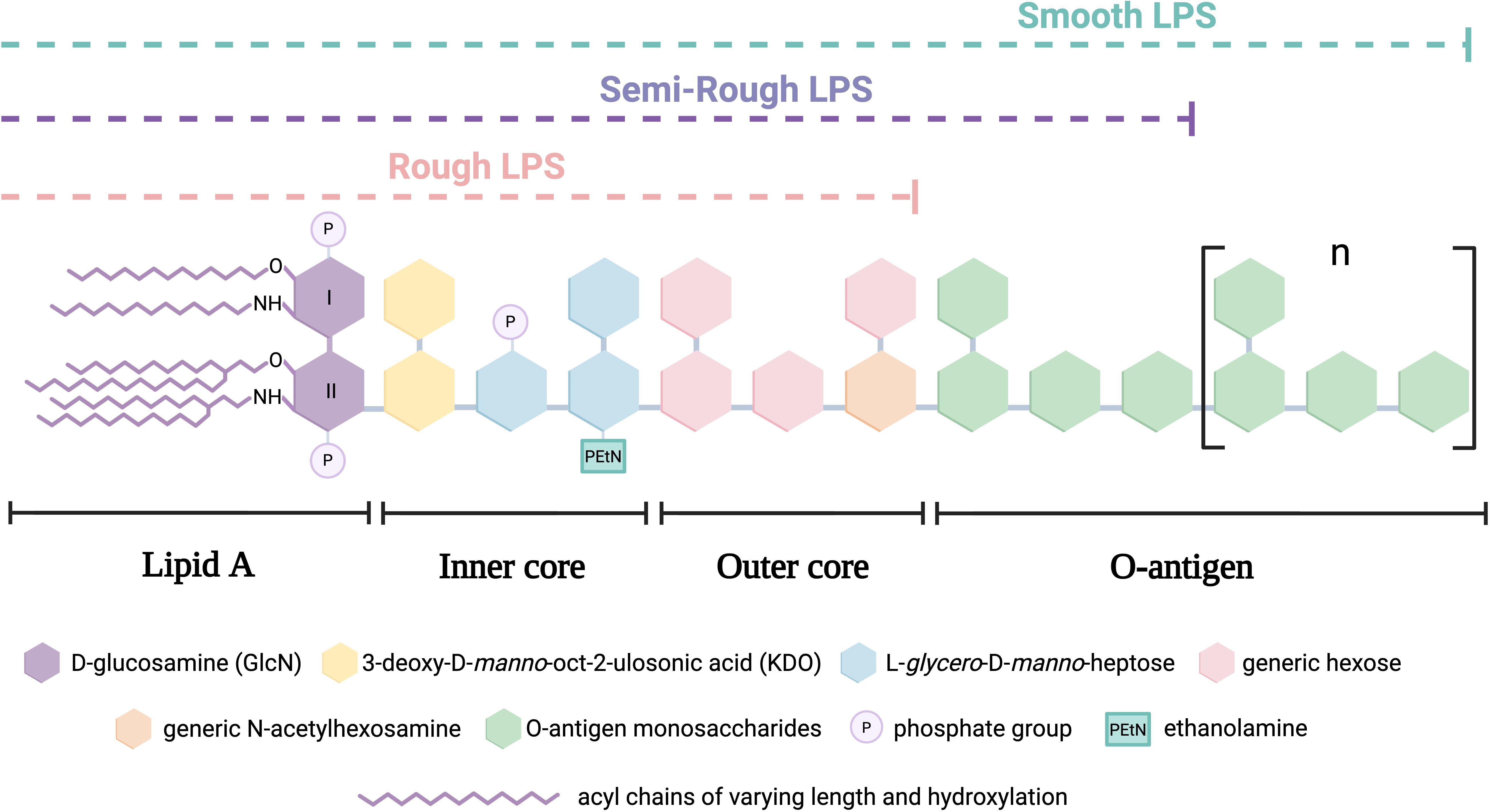
Figure 1. Schematic structure of canonical lipopolysaccharide (LPS) types in Gram-negative bacteria.
Additionally, LPS can be classified mainly into two types based on the presence and structure of the O-antigen:
● Smooth LPS (S-LPS): Contains the O-antigen, making it less susceptible to binding with plasma lipoproteins, favoring its circulation and prolonging its inflammatory activity.
● Rough LPS (R-LPS): Lacks the O-antigen and consists only of lipid A and the core oligosaccharide. It has a higher affinity for plasma lipoproteins, facilitating its neutralization and reducing its inflammatory effect. A related intermediate form, called semi-rough LPS, contains a single O-antigen repeat attached to the core and last, a specific subtype of R-LPS is lipooligosaccharide (LOS), characterized by the lack of O-antigen and a compact, structurally defined core oligosaccharide that serves as the main antigenic determinant (4).
Main determinants of LPS heterogeneity
To date, the structures of LPS from more than 50 bacterial species have been well characterized, revealing significant diversity in their structural modifications (5). Among these, LPS heterogeneity is determined by multiple structural factors, the most relevant being:
● Number and type of acyl chains in lipid A: The length, branching and degree of saturation of these chains influence immune system activation. Some variants can even modulate or inhibit immune activation (6–9).
● O-antigen length: Among structures sharing the same type of lipid A, a longer polysaccharide chain promotes the formation of stable aggregates in plasma, prolonging circulation time and enhancing inflammatory potential (10).
● Modifications with functional groups: Phosphate and ethanolamine groups can bind to Lipid A or the core oligosaccharide, altering interactions with immune cells and receptors (11).
Intestinal exposure and LPS translocation into the circulation
Under normal physiological conditions, very low levels of LPS can be found in plasma (12, 13). This is because a healthy intestine restricts the passage of LPS molecules into the bloodstream, despite an estimated 1 g of LPS being present in the intestinal lumen (14). In spite of this constant exposure to LPS, enterocytes and colonocytes exhibit a hyporesponsive state to these stimuli, partly explained by the low expression of Toll-like receptor 4 (TLR4) in these cells (15, 16). This intestinal tolerance mechanism minimizes inflammatory responses in the intestinal lumen (14). Additionally, exposure to LPS from gut microbiota plays a crucial role in immune system development and maturation, training regulatory T cells within the Gut-Associated Lymphoid Tissue (GALT) (17). An individual’s LPS profile depends on gut microbiota composition, which varies according to geographic and lifestyle factors, among others (18, 19).
Given this, LPS can enter systemic circulation through three main mechanisms:
● Infection by exogenous Gram-negative bacteria.
● Paracellular pathway, facilitated by increased intestinal permeability.
● Transcellular transport, where LPS is absorbed by enterocytes after dietary fat intake and secreted in association with chylomicrons at the basolateral membrane (13, 20, 21). Through the lymphatic system, chylomicron-borne LPS eventually reaches the bloodstream.
In relation to transcellular transport, it has been reported that after a single high-fat meal, plasma LPS levels increase, peaking 1–5 hours post-ingestion (12, 22, 23). Interestingly, despite high-fat diets being associated with an increase in Bacillota (formerly Firmicutes, Gram-positive bacteria) and a reduction in Bacteroidota (Gram-negative bacteria), elevated circulating LPS levels have been observed in humans under high-fat diets (24, 25) or other unhealthy dietary patterns (26, 27). These individuals can exhibit LPS levels 2 to 3 times higher compared to those on healthy diets. This phenomenon is referred to as “metabolic endotoxemia”, as LPS levels significantly rise without reaching the extreme levels observed in sepsis and it may have a prominent role in the development of cardiometabolic diseases (20, 28). In addition to diet, obesity also influences the magnitude of metabolic endotoxemia, with more pronounced increases in chylomicron-associated LPS levels as well as inflammatory cytokine production after a high fat meal in subjects with obesity (29). Another possible mechanism linking high-fat diets and LPS burden is related to altered intestinal permeability due to alteration in gut microbiome composition. By reducing Bifidobacterium levels, high-fat diets decrease glucagon-like peptide-2 (GLP-2) production, impairing intestinal barrier function and increasing paracellular LPS translocation (30, 31). This consequent endotoxemia can then promote the production of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β, further increasing intestinal permeability. This vicious cycle may contribute to metabolic dysfunction and low-grade chronic systemic inflammation (32, 33). Nonetheless, the relative contributions of transcellular and paracellular pathways for the total metabolic endotoxemia burden remain to be determined.
LPS metabolism: transport, biological activity, and elimination
Once in circulation, LPS can follow two main metabolic pathways:
● Immune activation: Binding to LPS-binding protein (LBP), which transfers LPS to the CD14/MD-2/TLR4 complex, triggering inflammatory signaling.
● Neutralization and elimination: Transfer of LPS to lipoproteins, mediated by LBP and phospholipid transfer protein (PLTP), followed by hepatic metabolism and biliary excretion.
Immune system activation via the CD14/MD-2/TLR4 complex
In an extracellular recognition mechanism, LPS initially binds to LBP. Then, LBP transfers LPS to the CD14 receptor, which exists in both soluble (sCD14) and membrane-bound forms (34). Since TLR4 cannot directly recognize LPS, CD14 facilitates its transfer to myeloid differentiation factor-2 (MD-2), stabilizing LPS binding and allowing TLR4 dimerization, which activates the signaling cascade (35). This CD14/MD-2/TLR4 complex is mainly present in immune cells such as monocytes, macrophages, and dendritic cells, though it is also detected in B cells, hepatocytes, adipocytes, and muscle cells, where it plays a role in inflammatory and metabolic responses.
The activation of this complex triggers two main signalling pathways:
1. The MyD88 pathway (rapid proinflammatory response): Activates myeloid differentiation primary response protein 88 (MyD88), leading to the activation of the NF-κB pathway and of the NLRP3 inflammasome (36), and eventually to the production of proinflammatory cytokines such as TNF-α, IL-6, and IL-1β (37).
2. The TRIF pathway (sustained antiviral response): Through the TIR-domain-containing adapter-inducing interferon-β (TRIF) adaptor protein, this cascade induces the production of IFN-β and other interferon-stimulated genes (ISGs), promoting an antiviral response and modulating inflammation in a more sustained manner (38, 39).
Intracellular immune recognition of LPS
In addition, recent evidence highlights an alternative intracellular recognition pathway for LPS. Certain immune cells, including macrophages, can internalize LPS through multiple mechanisms, such as endocytosis of outer membrane vesicles (OMVs) released by Gram-negative bacteria or direct uptake of LPS-protein complexes via endosomal pathways. Once inside the cytoplasm, LPS is recognized by inflammatory caspases, which bind directly to cytosolic LPS, triggering once again the activation of the NLRP3 inflammasome as well as the maturation of gasdermin D. This leads to the maturation and secretion of IL-1β and IL-18, as well as the gasdermin D-induced pyroptotic cell death (40), which serves as an additional immune defense mechanism. However, in conditions like sepsis, excessive activation of this pathway can contribute to systemic inflammation and tissue damage (41, 42).
Neutralization and elimination of LPS via lipoproteins
Since LPS are amphipathic phospholipids, in addition to binding to LBP, they can also bind to PLTP, facilitating their transfer to plasma lipoproteins, particularly HDL (43–45). This process limits LPS binding to TLR4, thereby exerting a protective effect against endotoxin-mediated inflammation (46, 47). It has also been reported that after 40 minutes of exposure, more than 50% of circulating LPS has been neutralized via this pathway, and that even after 10 minutes, LBP preferentially presents LPS to HDL rather than to CD14 in immune cells (48).
Once bound to HDL, LPS is transported to the liver, where it can be taken up mostly by hepatocytes or Kupffer cells. LPS uptake by hepatic cells was shown to involve scavenger receptor class-B type1 (SR-BI) which is the canonical receptor for HDL (49). In Kupffer cells, LPS undergoes processing through deacylation via acyl-oxyacyl hydrolase (AOAH). This enzyme is also active in other immune cells, such as macrophages and dendritic cells, which are present in various tissues, including the intestinal lamina propria. Finally, the resulting LPS degradation products, as well as remaining whole LPS, are excreted in the feces via bile, although this process can take several days (50).
Although HDL may be the main lipoprotein class involved in LPS detoxification, triglyceride-rich lipoproteins (QM and VLDL), and LDL also play a significant role in this process. Indeed, all lipoprotein classes are able to bind LPS (44) and to alleviate the inflammatory response of LPS-treated macrophages in vitro (51, 52) and to protect against the noxious effects of LPS in animal models (53). In addition to the role of SR-BI in the clearance of HDL-bound LPS, the involvement of the LDL receptor (LDLR) in LPS detoxification and protection against infection has been demonstrated in LDLR-deficient mice (54) and in humans harboring mutations of PCSK9, the endogenous inhibitor of LDLR (55). Overall, these data show that the different lipoprotein classes offer complementary clearance pathways for circulating LPS, with possible interconnections between them.
LPS structural variations and their impact on immune activation
As previously described, the classical activation of the immune system by LPS is mediated through its interaction with the TLR4/MD-2 complex, with distinct immunological outcomes depending on structural variations in LPS. In the case of the degree of acylation of lipid A, a hexa-acylated lipid A, like the one from E. coli LPS, typically contains four primary and two secondary acyl chains and induce a strong inflammatory response. This is because the first five acyl chains are inserted deeply into a hydrophobic pocket within MD-2, while the sixth chain extends outward and facilitates the recruitment of a second TLR4 monomer. This interaction is essential for the dimerization of the TLR4/MD-2 complex, a prerequisite for downstream signaling activation (56). In contrast, tetra- and penta-acylated lipid A (such as those from gut commensals belonging to the Bacteroides genus) exhibit markedly reduced immune activation due to their limited ability to promote TLR4 dimerization, as the missing acyl chains impair the necessary structural interactions (1, 57, 58). However, not only the number but also the type of acyl chains (length and degree of saturation) influences bacterial activation of the immune system (35). For example, it has been observed that variants of lipid A from E. Coli with acyl chains of 12 to 14 carbons are more endotoxic than variants with longer chains of 16 to 18 carbons by an impairment of LPS interaction with the CD14/MD-2/TLR4 complex (59–61).
Additionally, the number of phosphate groups also plays an important role, as they increase the negative charge of LPS, facilitating its binding to proteins required for LPS presentation to TLR4 and also to TLR4 itself. Thus, hexa-acylated lipid A (like E. coli one) generally contains two negatively charged phosphate groups, which interact via ionic bonds with positively charged amino acid residues in the TLR4/MD-2 complex, thereby stabilizing it (62, 63). In contrast, LPS derived from Bacteroides vulgatus (a common gut commensal), in addition to being hypoacylated and containing long-chain fatty acids, is deficient in these critical phosphate modifications, which further reduces its ability to trigger a strong TLR4-mediated immune response, underlining the importance of this aspect for immune modulation (63). The differential signaling of lipid A depending on its degree of phosphorylation has been exploited, for example, in the design of monophosphoryl lipid A (MPLA). This is a semi-synthetic derivative of LPS from Salmonella, in which one of the two phosphate groups has been removed. As a result, it acts as a partial TLR4 agonist, primarily triggering MyD88-dependent signaling with minimal TRIF activation, thereby leading to reduced inflammation. Because of its safety and efficacy profile, MPLA has been approved by the FDA as an adjuvant in vaccines such as those against HPV and herpes zoster (64, 65). Also, gut cells present an intestinal isoenzyme of alkaline phosphatase (iALP) at the brush border which participates in the hydrolysis of the phosphate groups of the LPS molecule. Indeed, in animal models, overexpression of iALP has been associated with protection from atherosclerosis through inactivation of LPS inflammatory actions and preservation of the intestinal barrier function (66).
Many of the structural modifications discussed above result in what is known as weak agonistic LPS. Unlike strong agonists or pure antagonists, weak agonistic LPS induce low-level receptor signaling, which can promote anti-inflammatory effects. It is important to notice that similar LPS modifications are part of the immune evasion strategies exploited by certain pathogens such as Yersinia pestis (67), Francisella novicida (68), Porphyromonas gingivalis and Helicobacter Pylori (69).
A summary of the structural characteristics of LPS from different gut commensals is summarized in Table 1.
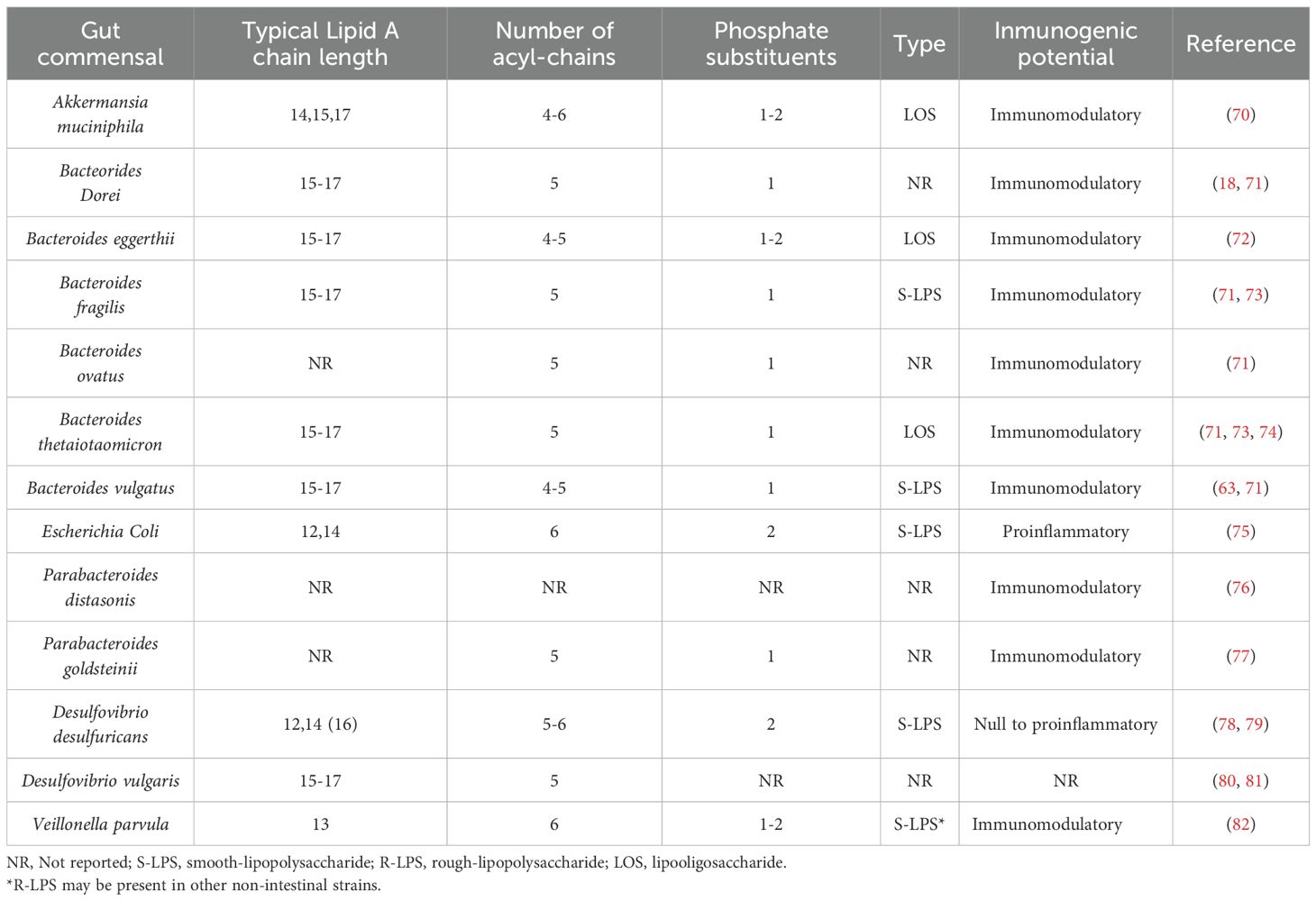
Table 1. Comparative Overview of LPS Structural features and immunogenic potential of Gut Commensals.
Although previous authors have employed different names to refer to immunomodulatory LPS (such as physiologic LPS, or anti-inflammatory LPS, etc.) (83–85), we consider for reaching a wider audience and to introduce the concept in this upbreak of gut microbiome research to classify them as “good” LPS as opposed to the largely known proinflammatory, “bad” LPS.
“Bad” vs. “Good” LPS: Gut commensals LPS and potential impact for human health
Evidence from metagenomic data of the Human Microbiome Project have confirmed that immune silencing via LPS is an intrinsic feature of the human gut microbiome (1). In healthy subjects, the majority of LPS primarily originates from bacteria of the phylum Bacteroidota (including genera such as Bacteroides, Parabacteroides, and Prevotella) rather than from Pseudomonadota, which includes Escherichia, Shigella, Enterobacter, among others (1, 86). Therefore, in opposition to what is commonly known about LPS, gut commensals LPS is not universally harmful, especially considering that most Gram-negative commensal bacteria are non-pathogenic and produce LPS, as well.
The mechanism that most probably drives these beneficial effects of LPS can be competition for TLR4 receptors. This seems to be exerted in two main ways: by directly competing with proinflammatory LPS in the binding with TLR4 or by preventing this binding by inhibiting their interaction (83). Additionally, some LPS are immunosilent, meaning they do not elicit any immune response, either pro or anti-inflammatory (1, 18).
LPS from Bacteroides members (likely the most abundant in the gut environment) are characterized by a hypoacylated lipid A structure (mainly penta-acylated), probably due to the absence of the late acyltransferase LpxM in the Raetz pathway, which normally adds the sixth acyl chain (71). These LPS are also monophosphorylated and contain 15–17-carbon acyl chains (5, 72, 74), all of which generally confer immunomodulatory effects. For example, administration of LPS from B. vulgatus demonstrated a direct anti-inflammatory effect by stimulating IL-10 production (63), while also reducing colon inflammation by inducing endotoxin tolerance (7). In combination with LPS from B. dorei, it may also confer protection against atherosclerosis (87). Similarly, LPS from B. fragilis and B. ovatus have been shown to improve intestinal inflammation and permeability in mice (88). In addition, Grombach et al. reported in a murine model that a microbiota dominated by Bacteroides, as opposed to one dominated by Enterobacteriaceae, induced lower levels of inflammation and immune activation in chemically induced colitis (61).
Regarding LPS from Parabacteroides, oral administration of a membrane fraction from P. distasonis, expected to contain LPS, not only exerted anti-inflammatory effects (89) but purified LPS from this species also reduced chemically induced acute colitis by inhibiting TNF-α production (76). Likewise, LPS from P. goldsteinii demonstrated an anti-inflammatory effect, reducing the development of induced chronic obstructive pulmonary disease (COPD) (77) and ameliorating H. pylori-related inflammation (90) presumably due to its hypoacylated lipid A structure (91). As for Prevotella, although approximately 40 species have been described, most belong to the oral microbiota, with Prevotella copri being the most prominent species in the gut. While its LPS has not been extensively characterized to date, current evidence suggests it may act as a pro-inflammatory molecule, as it is associated with increased LPS biosynthesis in the gut and has been linked to vascular calcification and metabolic liver disease (92, 93).
Although most bacteria within the Bacillota phylum are Gram-positive, there are exceptions such as Veillonella, a Gram-negative bacterium that therefore contains LPS. Oral administration of this bacterium provided protection against asthma in a mouse model of induced asthma. Moreover, higher levels of Veillonella have been reported in healthy children compared to those with asthma, with its LPS being identified as the main contributing factor (94).
Another interesting case is Akkermansia muciniphila, a Gram-negative bacterium to which several beneficial effects on the intestinal barrier have been attributed. This bacterium was recently approved by the EFSA as a novel food (95), specifically in its pasteurized form, highlighting that its beneficial effects may lie in its structural components rather than in its metabolic activity. In this context, its hypoacylated and monophosphorylated LOS, which lacks an O-antigen, favors an immunomodulatory response (70).
These findings indicate that weak agonistic LPS may serve as a potential therapeutic strategy providing an alternative to conventional treatments. Summary of the immunogenic potential of LPS from gut commensals characterized so far can be found at Table 1. These findings underscore not only the critical role of gut microbiota composition but also the therapeutic potential of microbiome-derived LPS in promoting immune homeostasis and mitigating inflammation in age-related diseases. In this regard, Gaber et al. (96) showed that in postmenopausal women gut microbiome alterations (with an imbalance favoring proinflammatory LPS producing bacteria, Escherichia coli, Shigella spp., and Klebsiella spp.) may contribute to visceral adiposity and that LPS from mice following a low-fat diet prevented some of the cardiometabolic alterations induced by a high-fat diet, underscoring the potential of “good” LPS for treatment of cardiometabolic diseases.
Approaches to LPS measurement in humans
From the above-mentioned, it arises the need for methods to characterize circulating LPS molecules at higher detail, reflecting the balance between “good” and “bad” LPS. LPS measurement is methodologically challenging, and several studies have shown the major limitation of the commonly used Limulus Amebocyte Lysate (LAL) test. LPS levels measured through LAL assays decrease in time because these assays measure the bioactive, free LPS molecules, which are rapidly detoxified by LBP, PLTP and lipoproteins in serum (97). Consequently, alternative biomarkers such as the levels of LBP, titers of antibodies against LPS (endoCab) or soluble CD-14 have been used. Nonetheless, these provide indirect evidence of higher exposure to LPS but neither contribute to characterize qualitative LPS alterations, nor to evaluate the balance between “good” and “bad” LPS.
Fatty acids in lipid A are 3-hydroxy fatty acids (3OH-FAs) of different chain lengths with a diverse acylation pattern. As already mentioned, the chain length of the acyl moieties composing the lipid A may be of relevance to explain differences among LPS immunogenic potential. For this reason, the measurement of esterified 3OH-FAs in circulation may be a proxy for LPS concentration, as these can only come LPS-Lipid A. GC/MS methods were initially used for the separation and quantification of these esterified 3OH-FAs (98). More recently, Pais-de-Barros et al. developed and optimized an HPLC/MS/MS methodology to detect and quantify esterified 3OH-FAs, obtained by calculating the difference between total and non-esterified (free) 3-OH-FAs, resulting in a LPS quantitative assay with increased sensitivity and accuracy (97). A recent study using this methodology showed that healthy controls exhibit higher concentrations of total esterified 3-OH-FAs than patients with end-stage renal disease (59). However, they identified that patients with end-stage renal disease exhibited a higher proportion of C12 and C14 3OH-FAs, which have been shown to produce a higher inflammatory activation in peripheral blood monocyte cells. Furthermore, studies done with this methodology show that among healthy subjects, the most prevalent esterified 3-hydroxy fatty acids are the C16 and C18 fatty acids. Therefore, this gives major evidence underscoring the importance of “good” LPS in human health. Alterations in LPS may therefore not only be quantitative, but most likely qualitative as well. These qualitative alterations may be relevant for disease development and progression and hold therapeutic potential.
Discussion-future insights
In addition to the possibility of designing LPS for use in postbiotics, probiotics, or vaccine adjuvants (60, 65), potential applications of LPS derived from non-commensal bacteria such as Rhodobacter capsulatus and Rhodobacter sphaeroides (99) which exhibit TLR4-antagonistic activity, as well as Pantoea agglomerans (100), whose LPS acts as a weak TLR4 agonist, are also being explored. Moreover, besides a potential immunoregulatory role of early-life exposure to specific LPS (18, 94), the balance between “good” and “bad” commensal LPS may shape the host environment for the development of systemic low-grade inflammation. The probable factors affecting this balance may require the concerted impairment of both gut microbiota composition, and intestinal barrier. The role of “good” LPS exerting immunomodulatory actions might have been disregarded in the bibliography as methods for demonstrating its presence in circulation have been recently developed and are not widely distributed. As increased intestinal permeability has been linked to several age-related chronic diseases, LPS translocation may, in this particular context, be detrimental. However, approaches to increase the pool of “good” LPS could be of particular use against age-related diseases. An association between “healthy” plant-based diets and lower “bad” LPS concentrations have been reported (26, 101). Last, next-generation probiotics based on species of the Bacteroides genus, showing immunomodulatory LPS, have received safety clearance by the European Commission and authorized for food processing and are under study (102). In terms of Public Health, which is increasingly targeting chronic inflammation for prevention of chronic diseases, the present review highlights how diet and gut microbiome eubiosis are of major relevance for promoting health.
Author contributions
RT: Writing – original draft, Writing – review & editing, Visualization. SS: Writing – original draft, Writing – review & editing, Visualization. RB: Writing – review & editing, Visualization. TG: Writing – review & editing, Visualization. JPPdB: Conceptualization, Writing – review & editing, Visualization. TM: Conceptualization, Writing – original draft, Writing – review & editing, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. RT received a grant from MICIU/AEI (Grant PRE2022-105209, 10.13039/501100011033), co-funded by the European Union - ESF+. Additional funds come from grant PID2021-128542OA-I00 funded by MICIU/AEI/10.13039/501100011033 and by “ERDF/EU A way of making Europe”. Additional support from INSA-UB Maria de Maeztu Unit of Excellence, (Grant CEX2021-001234-M) funded by MICIN/AEI/FEDER, UE.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. d’Hennezel E, Abubucker S, Murphy LO, and Cullen TW. Total lipopolysaccharide from the human gut microbiome silences toll-like receptor signaling. mSystems. (2017) 2. doi: 10.1128/mSystems.00046-17
2. Alexander C and Rietschel ET. Bacterial lipopolysaccharides and innate immunity. J Endotoxin Res. (2001) 7:167–202. doi: 10.1179/096805101101532675
3. Raetz CRH and Whitfield C. Lipopolysaccharide endotoxins. Annu Rev Biochem. (2002) 71:635–700. doi: 10.1146/annurev.biochem.71.110601.135414
4. Dardelle F, Phelip C, Darabi M, Kondakova T, Warnet X, Combret E, et al. Diversity, complexity, and specificity of bacterial lipopolysaccharide (LPS) structures impacting their detection and quantification. Int J Mol Sci. (2024) 25. doi: 10.3390/ijms25073927
5. Fux AC, Casonato Melo C, Michelini S, Swartzwelter BJ, Neusch A, Italiani P, et al. Heterogeneity of lipopolysaccharide as source of variability in bioassays and LPS-binding proteins as remedy. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms24098395
6. Saitoh SI, Akashi S, Yamada T, Tanimura N, Kobayashi M, Konno K, et al. Lipid A antagonist, lipid IVa, is distinct from lipid A in interaction with Toll-like receptor 4 (TLR4)-MD-2 and ligand-induced TLR4 oligomerization. Int Immunol. (2004) 16:961–9. doi: 10.1093/intimm/dxh097
7. Steimle A, Michaelis L, Di Lorenzo F, Kliem T, Münzner T, Maerz JK, et al. Weak agonistic LPS restores intestinal immune homeostasis. Mol Ther. (2019) 27:1974–91. doi: 10.1016/j.ymthe.2019.07.007
8. Ciesielska A, Matyjek M, and Kwiatkowska K. TLR4 and CD14 trafficking and its influence on LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. (2021) 78:1233–61. doi: 10.1007/s00018-020-03656-y
9. Heine H and Zamyatina A. Therapeutic targeting of TLR4 for inflammation, infection, and cancer: A perspective for disaccharide lipid A mimetics. Pharmaceuticals. (2023) 16. doi: 10.3390/ph16010023
10. Hardy E, Kamphuis T, Japaridze A, Wilschut JC, and Winterhalter M. Nanoaggregates of micropurified lipopolysaccharide identified using dynamic light scattering, zeta potential measurement, and TLR4 signaling activity. Anal Biochem. (2012) 430:203–13. doi: 10.1016/j.ab.2012.08.027
11. Narce M and Niot I. Fatty acids and lipopolysaccharides from health to disease. Biochimie. (2019) 159:1–2. doi: 10.1016/j.biochi.2019.02.008
12. Erridge C, Attina T, Spickett CM, and Webb DJ. A high-fat meal induces low-grade endotoxemia: evidence of a novel mechanism of postprandial inflammation. Am J Clin Nutr. (2007) 86:1–3. doi: 10.1093/ajcn/86.5.1286
13. Ghoshal S, Witta J, Zhong J, de Villiers W, and Eckhardt E. Chylomicrons promote intestinal absorption of lipopolysaccharides. J Lipid Res. (2009) 50:90–7. doi: 10.1194/jlr.M800156-JLR200
14. Gnauck A, Lentle RG, and Kruger MC. The characteristics and function of bacterial lipopolysaccharides and their endotoxic potential in humans. Int Rev Immunol. (2016) 35:189–218. doi: 10.3109/08830185.2015.1087518
15. Cuesta CM, Pascual M, Pérez-Moraga R, Rodríguez-Navarro I, García-García F, Ureña-Peralta JR, et al. Tlr4 deficiency affects the microbiome and reduces intestinal dysfunctions and inflammation in chronic alcohol-fed mice. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms222312830
16. Abreu MT. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat Rev Immunol. (2010) 10:131–43. doi: 10.1038/nri2707
17. Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Sci (1979). (2011) 331:337–41. doi: 10.1126/science.1198469
18. Vatanen T, Kostic AD, D’Hennezel E, Siljander H, Franzosa EA, Yassour M, et al. Variation in microbiome LPS immunogenicity contributes to autoimmunity in humans. Cell. (2016) 165:842–53. doi: 10.1016/j.cell.2016.04.007
19. Jha AR, Davenport ER, Gautam Y, Bhandari D, Tandukar S, Ng KM, et al. Gut microbiome transition across a lifestyle gradient in Himalaya. PloS Biol. (2018) 16. doi: 10.1371/journal.pbio.2005396
20. Cani PD, Amar J, Iglesias MA, Poggi M, Knauf C, Bastelica D, et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes. (2007) 56:1761–72. doi: 10.2337/db06-1491
21. Boutagy NE, McMillan RP, Frisard MI, and Hulver MW. Metabolic endotoxemia with obesity: Is it real and is it relevant? Biochimie. (2016) 124:11–20. doi: 10.1016/j.biochi.2015.06.020
22. Ghanim H, Sia CL, Upadhyay M, Korzeniewski K, Viswanathan P, Abuaysheh S, et al. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and toll-like receptor expression. Am J Clin Nutr. (2010) 91:940–9. doi: 10.3945/ajcn.2009.28584
23. Anderson AS, Haynie KR, McMillan RP, Osterberg KL, Boutagy NE, Frisard MI, et al. Early skeletal muscle adaptations to short-term high-fat diet in humans before changes in insulin sensitivity. Obesity. (2015) 23:720–4. doi: 10.1002/oby.21031
24. Amar J, Burcelin R, Ruidavets JB, Cani PD, Fauvel J, Alessi MC, et al. Energy intake is associated with endotoxemia in apparently healthy men. Am J Clin Nutr. (2008) 87:1219–23. doi: 10.1093/ajcn/87.5.1219
25. Harte AL, Varma MC, Tripathi G, Mcgee KC, Al-Daghri NM, Al-Attas OS, et al. High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care. (2012) 35:375–82. doi: 10.2337/dc11-1593
26. André P, De Barros JPP, Merle BMJ, Samieri C, Helmer C, Delcourt C, et al. Mediterranean diet and prudent diet are both associated with low circulating esterified 3-hydroxy fatty acids, a proxy of LPS burden, among older adults. Am J Clin Nutr. (2021) 114:1080–91. doi: 10.1093/ajcn/nqab126
27. Ahola AJ, Lassenius MI, Forsblom C, Harjutsalo V, Lehto M, and Groop PH. Dietary patterns reflecting healthy food choices are associated with lower serum LPS activity. Sci Rep. (2017) 7. doi: 10.1038/s41598-017-06885-7
28. Opal SM, Scannon PJ, Vincent JL, White M, Carroll SF, Palardy JE, et al. Relationship between plasma levels of lipopolysaccharide (LPS) and LPS-binding protein in patients with severe sepsis and septic shock. J Infect Dis. (1999) 180:1584–89. doi: 10.1086/315093
29. Vors C, Pineau G, Drai J, Meugnier E, Pesenti S, Laville M, et al. Postprandial endotoxemia linked with chylomicrons and lipopolysaccharides handling in obese versus lean men: A lipid dose-effect trial. J Clin Endocrinol Metab. (2015) 100:3427–35. doi: 10.1210/jc.2015-2518
30. Cani PD, Possemiers S, Van De Wiele T, Guiot Y, Everard A, Rottier O, et al. Changes in gut microbiota control inflammation in obese mice through a mechanism involving GLP-2-driven improvement of gut permeability. Gut. (2009) 58:1091–103. doi: 10.1136/gut.2008.165886
31. Chae YR, Lee YR, Kim YS, and Park HY. Diet-induced gut dysbiosis and leaky gut syndrome. J Microbiol Biotechnol. (2024) 34:747–56. doi: 10.4014/jmb.2312.12031
32. Muccioli GG, Naslain D, Bäckhed F, Reigstad CS, Lambert DM, Delzenne NM, et al. The endocannabinoid system links gut microbiota to adipogenesis. Mol Syst Biol. (2010) 6. doi: 10.1038/msb.2010.46
33. Saad MJA, Santos A, and Prada PO. Linking gut microbiota and inflammation to obesity and insulin resistance. Physiology. (2016) 31:283–93. doi: 10.1152/physiol.00041.2015
34. Hailman E, Lichenstein HS, Wurfel MM, Miller DS, Johnson DA, Kelley M, et al. Lipopolysaccharide (LPS)-binding protein accelerates the binding of LPS to CD14. Am J Clin Nutr. (2007) 179:269–77. doi: 10.1084/jem.179.1.269
35. Park BS, Song DH, Kim HM, Choi BS, Lee H, and Lee JO. The structural basis of lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature. (2009) 458:1191–5. doi: 10.1038/nature07830
36. Bauernfeind FG, Horvath G, Stutz A, Alnemri ES, MacDonald K, Speert D, et al. Cutting edge: NF-κB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol. (2009) 183:787–91. doi: 10.4049/jimmunol.0901363
37. Kawai T, Takeuchi O, Fujita T, Inoue J, Mü hlradt PF, Sato S, et al. Lipopolysaccharide stimulates the myD88-independent pathway and results in activation of IFN-regulatory factor 3 and the expression of a subset of lipopolysaccharide-inducible genes. J Immunol. (2001) 167:5887–94. doi: 10.4049/jimmunol.167.10.5887
38. Wang H, Garcia CA, Rehani K, Cekic C, Alard P, Kinane DF, et al. IFN-β Production by TLR4-stimulated innate immune cells is negatively regulated by GSK3-β. J Immunol. (2008) 181:6797–802. doi: 10.4049/jimmunol.181.10.6797
39. Keck S, Müller I, Fejer G, Savic I, Tchaptchet S, Nielsen PJ, et al. Absence of TRIF signaling in lipopolysaccharide-stimulated murine mast cells. J Immunol. (2011) 186:5478–88. doi: 10.4049/jimmunol.1000458
40. Shi J, Zhao Y, Wang K, Shi X, Wang Y, Huang H, et al. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature. (2015) 526:660–5. doi: 10.1038/nature15514
41. Gabarin RS, Li M, Zimmel PA, Marshall JC, Li Y, and Zhang H. Intracellular and extracellular lipopolysaccharide signaling in sepsis: avenues for novel therapeutic strategies. J Innate Immun. (2021) 13:323–32. doi: 10.1159/000515740
42. Kerr AW, Efron PA, Larson SD, and Rincon JC. T-cell activation and LPS: A dangerous duo for organ dysfunction. J Leukocyte Biol. (2022) 112:219–20. doi: 10.1002/JLB.3CE0122-019R
43. Hailman E, Albers JJ, Wolfbauer G, Tu AY, and Wright SD. Neutralization and transfer of lipopolysaccharide by phospholipid transfer protein. J Biol Chem. (1996) 271:12172–8. doi: 10.1074/jbc.271.21.12172
44. Levels JHM, Abraham PR, Van den Ende A, and Van Deventer SJH. Distribution and kinetics of lipoprotein-bound endotoxin. Infect Immun. (2001) 69:2821–8. doi: 10.1128/IAI.69.5.2821-2828.2001
45. Levels JHM, Marquart JA, Abraham PR, Van Den Ende AE, Molhuizen HOF, Van Deventer SJH, et al. Lipopolysaccharide is transferred from high-density to low-density lipoproteins by lipopolysaccharide-binding protein and phospholipid transfer protein. Infect Immun. (2005) 73:2321–6. doi: 10.1128/IAI.73.4.2321-2326.2005
46. Harris HW, Grunfeld C, Feingold KR, and Rapp JH. Human very low density lipoproteins and chylomicrons can protect against endotoxin-induced death in mice. J Clin Invest. (1990) 86:696–702. doi: 10.1172/JCI114765
47. Pajkrt D, Doran JE, Koster E, Lerch EG, Arnet B, van der Poll T, et al. Antiinflammatory effects of reconstituted high-density lipoprotein during human endotoxemia. J Exp Med. (1996) 184:1601–8. doi: 10.1084/jem.184.5.1601
48. Wurfel MM, Kunitake ST, Lichenstein H, Kane JP, and Wright SD. Lipopolysaccharide (LPS)-binding protein is carried on lipoproteins and acts as a cofactor in the neutralization of LPS. J Exp Med. (1994) 180:1025–35. doi: 10.1084/jem.180.3.1025
49. Cai L, Ji A, De Beer FC, Tannock LR, and van der Westhuyzen DR. SR-BI protects against endotoxemia in mice through its roles in glucocorticoid production and hepatic clearance. J Clin Invest. (2008) 118:364–75. doi: 10.1172/JCI31539
50. Shao B, Munford RS, Kitchens R, and Varley AW. Hepatic uptake and deacylation of the LPS in bloodborne LPS-lipoprotein complexes. Innate Immun. (2012) 18:825–33. doi: 10.1177/1753425912442431
51. Sprong T, Netea MG, van der Ley P, Verver-Jansen TJG, Jacobs LEH, Stalenhoef A, et al. Human lipoproteins have divergent neutralizing effects on E. coli LPS, N. meningitidis LPS, and complete Gram-negative bacteria. J Lipid Res. (2004) 45:742–9. doi: 10.1194/jlr.M300453-JLR200
52. Vreugdenhil ACE, Rousseau CH, Hartung T, Willem J, Greve M, Van ‘t Veer C, et al. Lipopolysaccharide (LPS)-binding protein mediates LPS detoxification by chylomicrons. J Immunol. (2003) 170:1399–405. doi: 10.4049/jimmunol.170.3.1399
53. Levine DM, Parker TS, Donnelly TM, Walsh A, and Rubin AL. In vivo protection against endotoxin by plasma high density lipoprotein (lipopolysaccharide/tumor necrosis factor a/infectious disease/transgenic/photoaffinity labeling). Proc Natl Acad Sci USA. (1993) 90:12040–44. doi: 10.1073/pnas.90.24.12040
54. Topchiy E, Cirstea M, Kong HJJ, Boyd JH, Wang Y, Russell JA, et al. Lipopolysaccharide is cleared from the circulation by hepatocytes via the low density lipoprotein receptor. PloS One. (2016) 11. doi: 10.1371/journal.pone.0155030
55. Walley KR, Thain KR, Russell JA, Reilly MP, Meyer NJ, Ferguson JF, et al. PCSK9 is a critical regulator of the innate immune response and septic shock outcome. Sci Transl Med. (2014) 6:258ra143. doi: 10.1126/scitranslmed.3008782
56. Fu Y, Kim H, Lee DS, Han AR, Heine H, Zamyatina A, et al. Structural insight into TLR4/MD-2 activation by synthetic LPS mimetics with distinct binding modes. Nat Commun. (2025) 16. doi: 10.1038/s41467-025-59550-3
57. Ohto U, Fukase K, Miyake K, and Shimizu T. Structural basis of species-specific endotoxin sensing by innate immune receptor TLR4/MD-2. Proc Natl Acad Sci U S A. (2012) 109:7421–6. doi: 10.1073/pnas.1201193109
58. Coats SR, Do CT, Karimi-Naser LM, Braham PH, and Darveau RP. Antagonistic lipopolysaccharides block E. coli lipopolysaccharide function at human TLR4 via interaction with the human MD-2 lipopolysaccharide binding site. Cell Microbiol. (2007) 9:1191–202. doi: 10.1111/j.1462-5822.2006.00859.x
59. Adda-Rezig H, Carron C, Pais de Barros JP, Choubley H, Charron E, Rérole AL, et al. New insights on end-stage renal disease and healthy individual gut bacterial translocation: different carbon composition of lipopolysaccharides and different impact on monocyte inflammatory response. Front Immunol. (2021) 12. doi: 10.3389/fimmu.2021.658404
60. Needham BD, Carroll SM, Giles DK, Georgiou G, Whiteley M, and Trent MS. Modulating the innate immune response by combinatorial engineering of endotoxin. Proc Natl Acad Sci U S A. (2013) 110:1464–9. doi: 10.1073/pnas.1218080110
61. Gronbach K, Flade I, Holst O, Lindner B, Ruscheweyh HJ, Wittmann A, et al. Endotoxicity of lipopolysaccharide as a determinant of T-cell-mediated colitis induction in mice. Gastroenterology. (2014) 146:765–75. doi: 10.1053/j.gastro.2013.11.033
62. Maeshima N and Fernandez RC. Recognition of lipid A variants by the TLR4-MD-2 receptor complex. Front Cell Infection Microbiol. (2013) 4. doi: 10.3389/fcimb.2013.00003
63. Di Lorenzo F, Pither MD, Martufi M, Scarinci I, Guzmán-Caldentey J, Łakomiec E, et al. Pairing bacteroides vulgatus LPS structure with its immunomodulatory effects on human cellular models. ACS Cent Sci. (2020) 6:1602–16. doi: 10.1021/acscentsci.0c00791
64. Cekic C, Casella CR, Eaves CA, Matsuzawa A, Ichijo H, and Mitchell TC. Selective activation of the p38 MAPK pathway by synthetic monophosphoryl lipid A. J Biol Chem. (2009) 284:31982–91. doi: 10.1074/jbc.M109.046383
65. Wang YQ, Bazin-Lee H, Evans JT, Casella CR, and Mitchell TC. MPL adjuvant contains competitive antagonists of human TLR4. Front Immunol. (2020) 11. doi: 10.3389/fimmu.2020.577823
66. Ghosh SS, Wang J, Yannie PJ, Cooper RC, Sandhu YK, Kakiyama G, et al. Over-expression of intestinal alkaline phosphatase attenuates atherosclerosis. Circ Res. (2021) 128:1646–59. doi: 10.1161/CIRCRESAHA.120.317144
67. Knirel YA, Lindner B, Vinogradov EV, Kocharova NA, Senchenkova SN, Shaikhutdinova RZ, et al. Temperature-dependent variations and intraspecies diversity of the structure of the lipopolysaccharide of Yersinia pestis. Biochemistry. (2005) 44:1731–43. doi: 10.1021/bi048430f
68. Okan NA and Kasper DL. The atypical lipopolysaccharide of Francisella. Carbohydr Res. (2013) 378:79–83. doi: 10.1016/j.carres.2013.06.015
69. Needham BD and Trent MS. Fortifying the barrier: The impact of lipid A remodelling on bacterial pathogenesis. Nat Rev Microbiol. (2013) 11:467–81. doi: 10.1038/nrmicro3047
70. Garcia-Vello P, Tytgat HLP, Elzinga J, Van Hul M, Plovier H, Tiemblo-Martin M, et al. The lipooligosaccharide of the gut symbiont Akkermansia muciniphila exhibits a remarkable structure and TLR signaling capacity. Nat Commun. (2024) 15:8411. doi: 10.1038/s41467-024-52683-x
71. Jacobson AN, Choudhury BP, and Fischbach MA. The biosynthesis of lipooligosaccharide from Bacteroides thetaiotaomicron. mBio. (2018) 9. doi: 10.1128/mBio.02289-17
72. Tiemblo Martín M, Coccimiglio M, Andretta E, De Simone Carone L, Bell A, Guerpe Amor T, et al. The human gut Bacteroides eggerthii expresses a new galactofuranose-containing lipooligosaccharide with weak immunostimulatory properties. Carbohydr Polym. (2025) 348. doi: 10.1016/j.carbpol.2024.122833
73. Berezow AB, Ernst RK, Coats SR, Braham PH, Karimi-Naser LM, and Darveau RP. The structurally similar, penta-acylated lipopolysaccharides of Porphyromonas gingivalis and Bacteroides elicit strikingly different innate immune responses. Microb Pathog. (2009) 47:68–77. doi: 10.1016/j.micpath.2009.04.015
74. Pither MD, Illiano A, Pagliuca C, Jacobson A, Mantova G, Stornaiuolo A, et al. Bacteroides thetaiotaomicron rough-type lipopolysaccharide: The chemical structure and the immunological activity. Carbohydr Polym. (2022) 297. doi: 10.1016/j.carbpol.2022.120040
75. Raetz CRH, Guan Z, Ingram BO, Six DA, Song F, Wang X, et al. Discovery of new biosynthetic pathways: The lipid A story. J Lipid Res. (2009) 50:S103–S108. doi: 10.1194/jlr.R800060-JLR200
76. Kverka M, Zakostelska Z, Klimesova K, Sokol D, Hudcovic T, Hrncir T, et al. Oral administration of Parabacteroides distasonis antigens attenuates experimental murine colitis through modulation of immunity and microbiota composition. Clin Exp Immunol. (2011) 163:250–9. doi: 10.1111/j.1365-2249.2010.04286.x
77. Lai HC, Lin TL, Chen TW, Kuo YL, Chang CJ, Wu TR, et al. Gut microbiota modulates COPD pathogenesis: Role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut. (2022) 71:309–21. doi: 10.1136/gutjnl-2020-322599
78. Lodowska J, Wolny D, Jaworska-Kik M, Kurkiewicz S, Dzierzewicz Z, and Węglarz L. The chemical composition of endotoxin isolated from intestinal strain of Desulfovibrio desulfuricans. Sci World J. (2012) 2012. doi: 10.1100/2012/647352
79. Zhang-Sun W, Augusto LA, Zhao L, and Caroff M. Desulfovibrio desulfuricans isolates from the gut of a single individual: Structural and biological lipid A characterization. FEBS Lett. (2015) 589:165–71. doi: 10.1016/j.febslet.2014.11.042
80. Edlund A, Nichols PD, Roffey R, and White DC. Extractable and lipopolysaccharide fatty acid and hydroxy acid profiles from Desulfovibrio species. J Lipid Res. (1985) 26:982–8. doi: 10.1016/S0022-2275(20)34302-9
81. Opiyo SO, Pardy RL, Moriyama H, and Moriyama EN. Evolution of the Kdo2-lipid A biosynthesis in bacteria. BMC Evol Biol. (2010) 10. doi: 10.1186/1471-2148-10-362
82. Pither MD, Andretta E, Rocca G, Balzarini F, Matamoros-Recio A, Colicchio R, et al. Deciphering the chemical language of the immunomodulatory properties of veillonella parvula lipopolysaccharide. Angewandte Chemie - Int Edition. (2024) 63. doi: 10.1002/anie.202401541
83. Lin TL, Shu CC, Chen YM, Lu JJ, Wu TS, Lai WF, et al. Like cures like: pharmacological activity of anti-inflammatory lipopolysaccharides from gut microbiome. Front Pharmacol. (2020) 11. doi: 10.3389/fphar.2020.00554
84. Jiang W and Banks WA. Viewpoint: Is lipopolysaccharide a hormone or a vitamin? Brain Behavior Immun. (2023) 114:1–2. doi: 10.1016/j.bbi.2023.07.018
85. Di Lorenzo F, De Castro C, Silipo A, and Molinaro A. Lipopolysaccharide structures of Gram-negative populations in the gut microbiota and effects on host interactions. FEMS Microbiol Rev. (2019) 43. doi: 10.1093/femsre/fuz002
86. Huttenhower C, Gevers D, Knight R, Abubucker S, Badger JH, Chinwalla AT, et al. Structure, function and diversity of the healthy human microbiome. Nature. (2012) 486:207–14. doi: 10.1038/nature11234
87. Yoshida N, Emoto T, Yamashita T, Watanabe H, Hayashi T, Tabata T, et al. Bacteroides vulgatus and Bacteroides dorei Reduce Gut Microbial Lipopolysaccharide Production and Inhibit Atherosclerosis. Circulation. (2018) 138:2486–98. doi: 10.1161/CIRCULATIONAHA.118.033714
88. Tan H, Zhao J, Zhang H, Zhai Q, and Chen W. Novel strains of Bacteroides fragilis and Bacteroides ovatus alleviate the LPS-induced inflammation in mice. Appl Microbiol Biotechnol. (2019) 103:2353–65. doi: 10.1007/s00253-019-09617-1
89. Koh GY, Kane A, Lee K, Xu Q, Wu X, Roper J, et al. Parabacteroides distasonis attenuates toll-like receptor 4 signaling and Akt activation and blocks colon tumor formation in high-fat diet-fed azoxymethane-treated mice. Int J Cancer. (2018) 143:1797–805. doi: 10.1002/ijc.31559
90. Lai CH, Lin TL, Huang MZ, Li SW, Wu HY, Chiu YF, et al. Gut Commensal Parabacteroides goldsteinii MTS01 Alters Gut Microbiota Composition and Reduces Cholesterol to Mitigate Helicobacter pylori-Induced Pathogenesis. Front Immunol. (2022) 13. doi: 10.3389/fimmu.2022.916848
91. Lin TL, Chen WJ, Hung CM, Wong YL, Lu CC, and Lai HC. Characterization and safety evaluation of autoclaved gut commensal parabacteroides goldsteinii RV-01. Int J Mol Sci. (2024) 25. doi: 10.3390/ijms252312660
92. Yuan H, Wu X, Wang X, Zhou JY, and Park S. Microbial dysbiosis linked to metabolic dysfunction-associated fatty liver disease in asians: prevotella copri promotes lipopolysaccharide biosynthesis and network instability in the prevotella enterotype. Int J Mol Sci. (2024) 25. doi: 10.3390/ijms25042183
93. Hao QY, Yan J, Wei JT, Zeng YH, Feng LY, Que DD, et al. Prevotella copri promotes vascular calcification via lipopolysaccharide through activation of NF-κB signaling pathway. Gut Microbes. (2024) 16. doi: 10.1080/19490976.2024.2351532
94. Arrieta MC, Stiemsma LT, Dimitriu PA, Thorson L, Russell S, Yurist-Doutsch S, et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci Transl Med. (2015) 7. doi: 10.1126/scitranslmed.aab2271
95. Turck D, Bohn T, Castenmiller J, De Henauw S, Hirsch-Ernst KI, Maciuk A, et al. Safety of pasteurised Akkermansia muciniphila as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. (2021) 19. doi: 10.2903/j.efsa.2021.6780
96. Gaber M, Wilson AS, Millen AE, Hovey KM, LaMonte MJ, Wactawski-Wende J, et al. Visceral adiposity in postmenopausal women is associated with a pro-inflammatory gut microbiome and immunogenic metabolic endotoxemia. Microbiome. (2024) 12:192. doi: 10.1186/s40168-024-01901-1
97. De Barros JPP, Gautier T, Sali W, Adrie C, Choubley H, Charron E, et al. Quantitative lipopolysaccharide analysis using HPLC/MS/MS and its combination with the limulus amebocyte lysate assay. J Lipid Res. (2015) 56:1363–9. doi: 10.1194/jlr.D059725
98. Maitra SK, Schotztf MC, Yoshikawa TT, and Guze LB. Determination of lipid A and endotoxin in serum by mass spectroscopy (ft-hydroxymyristic acid/gas chromatography-mass spectrometry/Gram-negative sepsis). Med Sci. (1978) 75:3993–97. doi: 10.1073/pnas.75.8.3993
99. Barochia A, Solomon S, Cui X, Natanson C, and Eichacker PQ. Eritoran tetrasodium (E5564) treatment for sepsis: Review of preclinical and clinical studies. Expert Opin Drug Metab Toxicol. (2011) 7:479–94. doi: 10.1517/17425255.2011.558190
100. Morishima A and Inagawa H. Clinical effects of orally administered lipopolysaccharide derived from pantoea agglomerans on Malignant tumors. Anticancer Res. (2016) 36:3747–52.
101. Hornero-Ramirez H, Morisette A, Marcotte B, Penhoat A, Lecomte B, Panthu B, et al. Multifunctional dietary approach reduces intestinal inflammation in relation with changes in gut microbiota composition in subjects at cardiometabolic risk: the SINFONI project. Gut Microbes. (2025) 17:2438823. doi: 10.1080/19490976.2024.2438823
Keywords: lipopolysaccharide (endotoxin), inflammation, intestinal permeability, nutrition, gut microbiota
Citation: Tume R, El Sherbiny S, Bono R, Gautier T, Pais de Barros JP and Meroño T (2025) The balance between proinflammatory, “bad”, and immunomodulatory, “good”, lipopolysaccharide for understanding gut-derived systemic inflammation. Front. Immunol. 16:1588129. doi: 10.3389/fimmu.2025.1588129
Received: 05 March 2025; Accepted: 17 June 2025;
Published: 09 July 2025.
Edited by:
Jeffrey J. Pu, Upstate Medical University, United StatesReviewed by:
Flaviana Di Lorenzo, University of Naples Federico II, ItalyCopyright © 2025 Tume, El Sherbiny, Bono, Gautier, Pais de Barros and Meroño. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tomás Meroño, dG9tYXNtZXJvbm9AdWIuZWR1