 Delphine Béland1,2,3
Delphine Béland1,2,3 Mélissa Viens
Mélissa Viens Marie-Claude Bourgeois-Daigneault
Marie-Claude Bourgeois-Daigneault- 1Cancer Axis and Institut du cancer de Montréal, Centre Hospitalier de l’Université de Montréal (CHUM) Research Centre, Montreal, QC, Canada
- 2Immunopathology Axis, Centre Hospitalier de l’Université de Montréal (CHUM) Research Centre, Montreal, QC, Canada
- 3Department of Microbiology, Infectious Diseases and Immunology, Faculty of Medicine, University of Montreal, Montreal, QC, Canada
The proteasome (prosome, macropain) is a key cellular organelle responsible primarily for protein homeostasis, by degrading damaged or misfolded proteins. Proteasome-processed protein fragments can then be further trimmed and funneled to the major histocompatibility complex class I (MHC-I) antigen presentation pathway for cell surface display and immune recognition. Various types of proteasomes can be found in mammalian cells with different expression patterns and cleavage abilities. As such, the immunoproteasome (ImP) preferentially cleaves proteins to yield MHC-I-compatible fragments. It is constitutively expressed by some immune cells and can be induced by pro-inflammatory signals. Interestingly, it was also found to be expressed in multiple types of cancers and proteasome activity can be modulated by some cancer therapies. A better understanding of its impact on cancer progression, prognosis and treatment response is therefore needed to guide treatment decisions. In this review, we focus on the multiple roles of the ImP in cancer, including its interplay with the immune system, as well as its impact on patient outcomes.
1 Introduction
The constitutive 26S proteasome (CP) is a large intracellular proteolytic complex that is primarily known for its key role in protein homeostasis (1). Protein degradation by the CP allows for the recycling of misfolded and damaged proteins and also modulates various cellular functions by degrading key pathway regulators, as well as other components (2–4). Importantly, the CP also contributes to antigen presentation by major histocompatibility class I (MHC-I) molecules. Indeed, it cleaves proteins into peptides that are then fed into the pathway for loading onto MHC-I molecules, cell surface display and presentation to immune cells (5). Other types of proteasomes are also found in mammals. As such, the thymoproteasome, the spermatoproteasome and the ImP are expressed in different contexts. While the spermatoproteasome is present in the testes during spermatogenesis, and the thymoproteasome is expressed in cortical thymic cells, the ImP is expressed by some immune cells and also inducible by most cell types in inflammatory conditions (6). The ImP is also found in many cell types in the tumor microenvironment (TME), where it was described to impact disease outcomes (7). In this review, we discuss the various roles of the ImP through cancer progression (Figure 1).
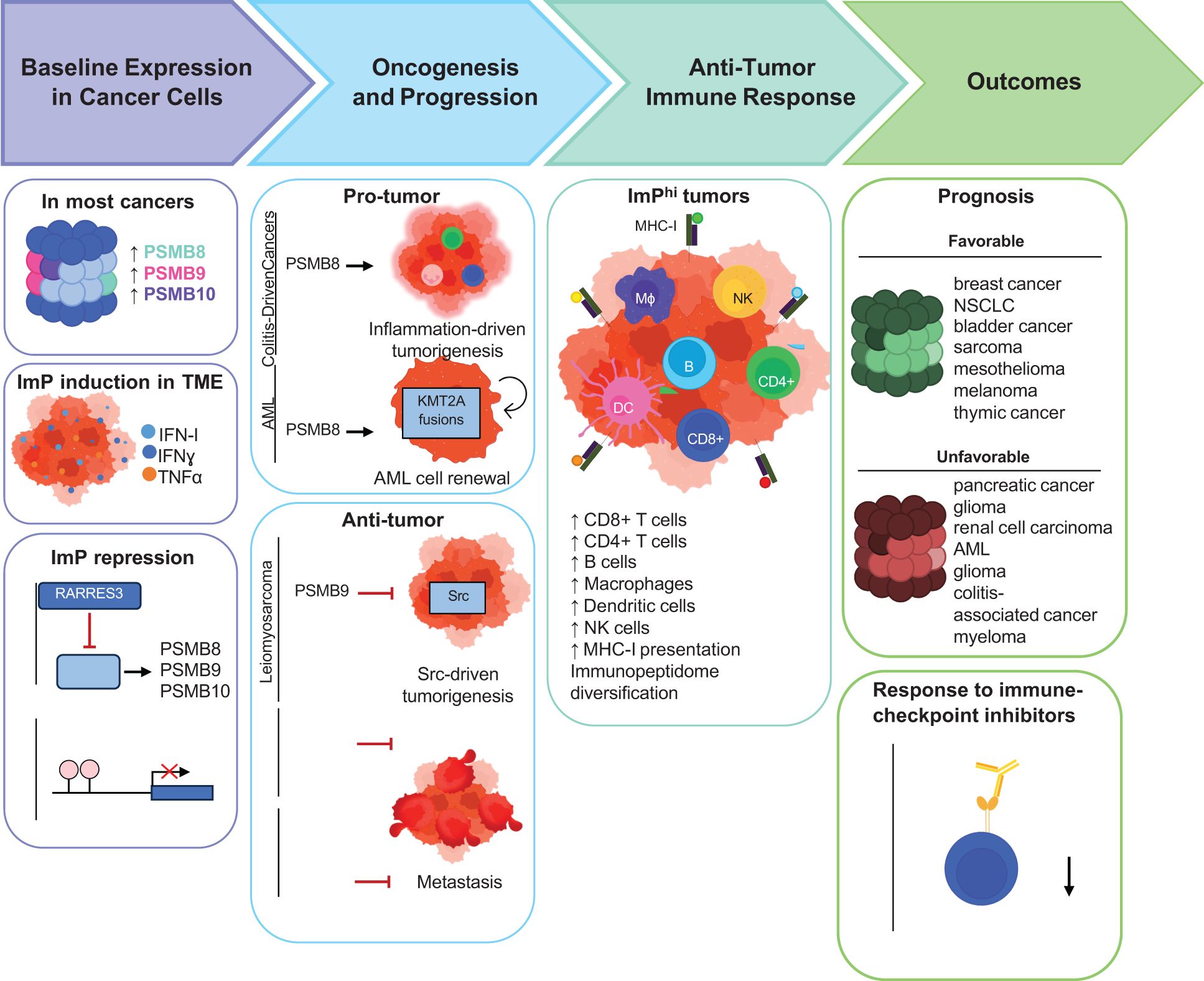
Figure 1. Schematic representation of the roles of the ImP across different stages of cancer progression. Most cancers exhibit increased ImP expression compared to healthy tissues while some others repress ImP expression. The known mechanisms by which cancers regulate ImP expression are depicted. The effects of the ImP in oncogenesis, anti-tumor immunity and outcomes are also outlined.
2 ImP structure
All proteasomes contain a barrel-shaped 20S core particle, which is composed of four heptameric rings. The two outer rings are composed of proteasome 20S subunit alpha 1 to 7, which are encoded by the proteasome 20S subunit alpha (PSMA) genes. The alpha rings are conserved across all proteasome subtypes and guard access to the inner catalytic chamber, where cleavage takes place (8). The two inner rings are composed of proteasome subunit beta type 1-7 (PSMB1-7), which have either structural (PSMB1-4) or catalytic roles. More specifically, PSMB5, 6 and 7, bear chymotrypsin-, caspase- and trypsin-like cleavage abilities, respectively (9). While chymotrypsin-like activity cleaves proteins after residues with hydrophobic side chains such as tyrosines, leucines, isoleucines and phenylalanines, caspase-like cleavage cuts after acidic residues like asparagines and glutamates and trypsin-like cleavages takes place after basic residues like arginines and lysines (10, 11). Notably, chymotrypsin-like cleavage yields protein fragments with optimal residues for anchoring to the MHC-I peptide-binding groove. For the ImP, the catalytic subunits are PSMB8 (also called low molecular mass peptide (LMP) 7 or ImP subunit β5 (PSβ5i)), PSMB9 (LMP2 or PSβ1i) and PSMB10 (multi-catalytic endopeptidase complex subunit 1 or PSβ2i) (12). While PSMB8 and 10 retain the activities of their CP homologs, PSMB9 provides additional chymotrypsin-like cleavage to the ImP instead of trypsin-like activity (Table 1) (12). As a result, the CP and ImP generate different repertoires of peptides, which can then be loaded onto MHC-I molecules (13). This surface display of the intracellular protein content allows for immune cells to identify and eliminate cells that are infected or mutated. This is especially important in the context of cancer where immunoediting allows for the elimination of malignant cells (14). As described below, the ImP was found to affect cancer progression and treatment responses in a variety of cancer models (15, 16). It is expressed by different cell types and its aberrant regulation can affect cellular function that can be either beneficial or detrimental to the disease, depending on the context.
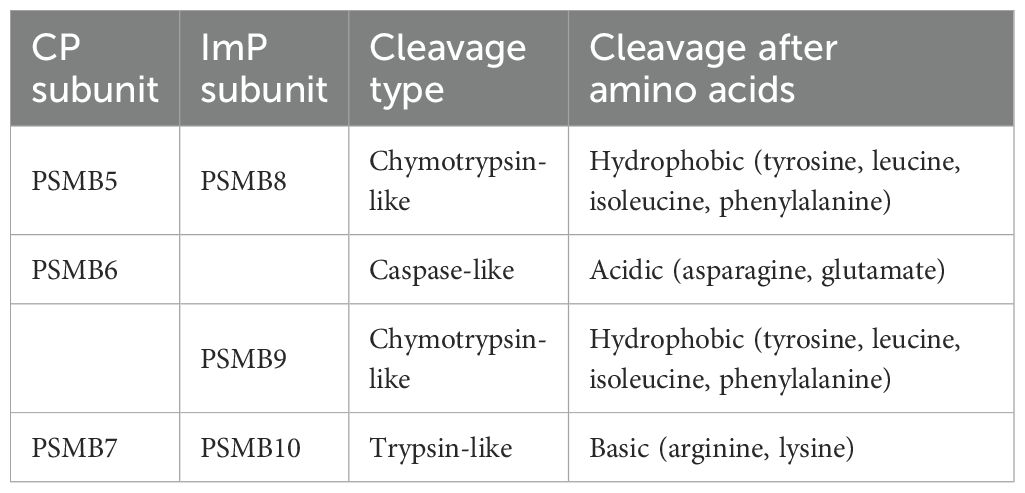
Table 1. Catalytic activities of CP and ImP subunits.
3 ImP regulation
Hematopoietic cells tend to have a proteasome pool that is skewed towards ImPs. Notably, the ImP is constitutively expressed by professional antigen-presenting cells such as dendritic cells, B cells and macrophages, therefore supporting its importance for antigen presentation (17). These cells, as well as T cells, NK cells, granulocytes, and monocytes, almost exclusively express the ImP (18). For non-immune cells, ImP expression can be induced by pro-inflammatory cytokines (19), with type-II interferon (IFNγ) being the most potent inducer (20). Type-I IFNs, as well as tumor necrosis factor α (TNFα) (20, 21) and various stress signals such as a heat shock, reactive oxygen species, nitric oxide and bacterial lipopolysaccharide can also trigger its expression (Figure 2) (22–25).
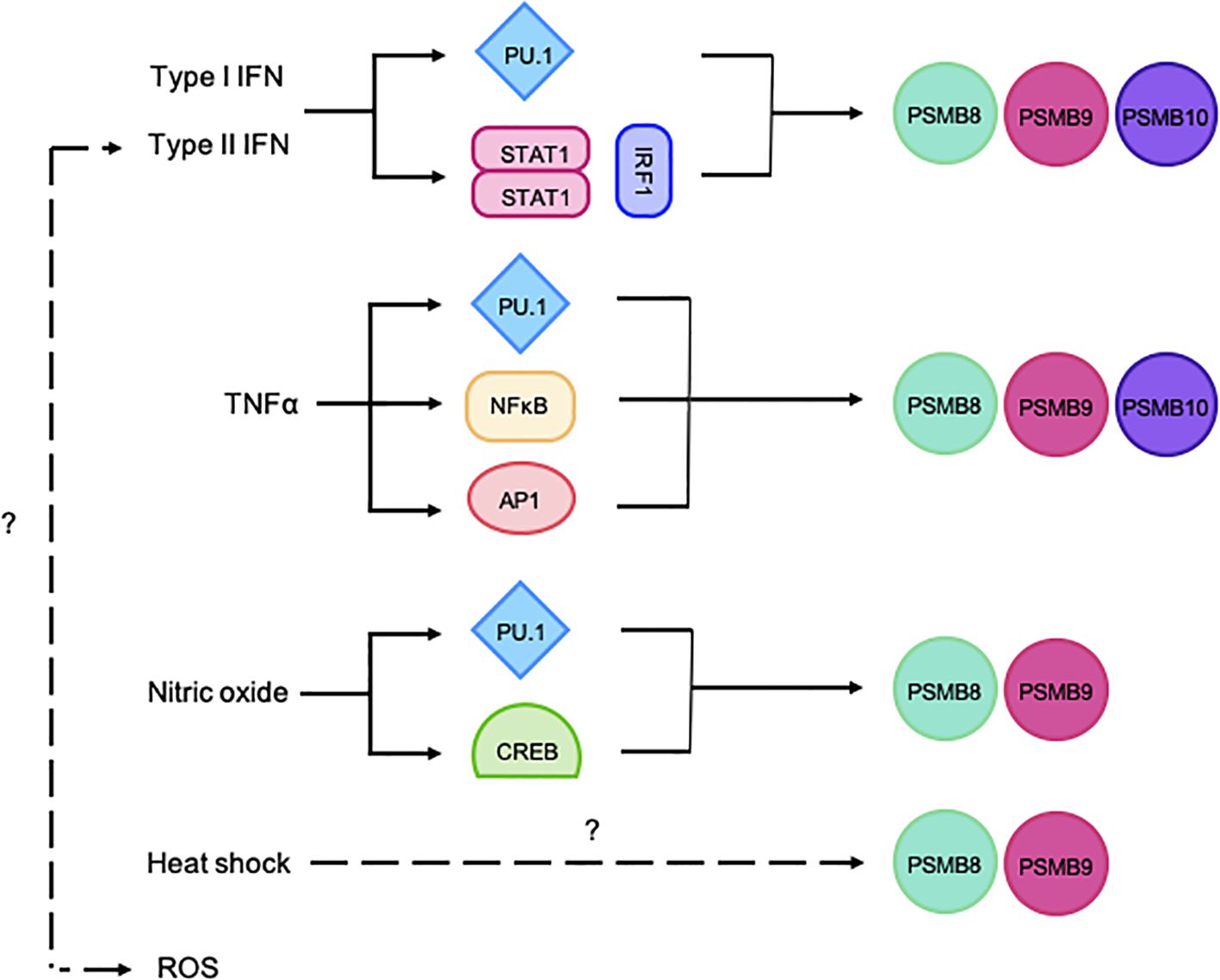
Figure 2. Schematic representation of the regulation of ImP subunit expression. The ImP subunits can be induced via PU.1, STAT1/IRF1, NF-κB, AP1 or CREB. The figure illustrates various stimuli that trigger the expression of PSMB8, 9 and/or 10.
PSMB8 and 9 genes are closely encoded in the MHC-II locus on chromosome 6 in humans or 17 in mice and are therefore co-regulated (12). Their promoters contains multiple sequence elements that are recognized by signal transduction and activator of transcription 1 (STAT1), as well as IFN regulatory factor 1 (IRF1), thus conferring IFN-responsiveness (26). Alternatively, PSMB8 and 9 transcription can also be induced by nuclear factor kappa-light-chain enhancer of activated B cells (NF-κB), cyclic adenosine monophosphate responsive element binding protein (CREB), zinc finger protein 268 (ZNF268), specificity protein 1 (SP1), as well as activator protein 1 (AP1/2), therefore allowing for their IFN-independent regulation (27, 28). PSMB10 is located on a different chromosome (8 in humans and 16 in mice), but can also be induced by STAT1, IRF1, NF-κB, SP1 and AP1/2 (12). Finally, IFNs, TNFα and nitric oxide can also induce the expression of all three ImP subunits via purine-rich box binding protein 1 (PU.1) (29–32) (Figure 2). When expressed, PSMB8, 9 and 10 are preferentially incorporated into the proteasome complex. Importantly, the subunits contain N-terminal pro-peptides that block their catalytic sites, which prevents aberrant protein cleavage (33). Once the complex is fully assembled, the pro-peptides are cleaved, and the ImP is catalytically active (3). Functional ImPs accomplish various functions that, in addition to shaping immune responses, also affect cancer onset, progression and treatment responses.
4 ImP functions within the tumor niche
The TME is a complex network that includes several cell types, stroma, as well as extracellular matrix. Given that the ImP is a cellular component that is not secreted, it is neither found in the stroma nor the extracellular matrix. Instead, it can be found in the cells of the TME, which include immune cells, endothelial cells, cancer-associated fibroblasts (CAFs) and tumor cells themselves (7, 34–36).
Immune cells infiltrate tumor tissues and act as critical modulators of immune surveillance (7, 37, 38). To avoid immune recognition, many cancers have therefore evolved defects in various components of the antigen presentation machinery. For example, non-small cell lung cancer (NSCLC), as well as pancreatic and esophagus squamous cell carcinomas downregulate ImP expression (39–41). Importantly, many studies are now describing the ImP as a biomarker of tumor immunogenicity (7, 42–44). For instance, in lower-grade glioma tumors, PSMB9 transcript levels were shown to correlate with increased gene signatures for CD8 and CD4 T cells, macrophages, as well as B cells (45). Similarly, Kumar et al. conducted a pan-cancer analysis of PSMB8, 9 and 10 expression and immune infiltration and found that PSMB8, 9 and 10 co-expression was linked to increased CD8 T cell, natural killer cell and activated dendritic cell gene signatures in most cancer types. Finally, it has been shown in myeloma, melanoma and clear cell renal cell carcinoma that activating the ImP increased the presentation of tumor-associated antigens, which resulted in enhanced anti-tumor activity (42, 46, 47).
Although the role of the ImP in antigen presentation is recognized as its main function, it also impacts immune cells in various other ways. For dendritic cells, ImP expression was shown to shape their cellular transcriptome. Indeed, PSMB8- and 10-deficient cells exhibited decreased activation of IRFs, STATs and NF-κB, which in turn prevented the expression of multiple-pro-inflammatory genes (48), which may impair the recruitment, activation and polarization of adaptive immune cells with anti-tumor activity within the TME (49).
For macrophages, high PSMB9 expression within the tumor was linked to increased phagocytosis (50), which could suggest enhanced tumor clearance and subsequent presentation of cancer antigens. For CD8 T cells, PSMB8, 9 and 10 co-expression was linked to increased activation and intra-tumor infiltration (49). For CD4 T cells, PSMB8 was shown to be critical for their activation by allowing for optimal MHC-II presentation (51). Furthermore, an ImP inhibitor was shown to favor the differentiation of T cells into regulatory T cells rather that Th1 or Th17 cells (52, 53). PSMB8 was also found to contribute to cell activation and protein homeostasis in both CD4 T cells and B cells (52).
For natural killer (NK) cells, which recognize missing self, their cytotoxic activity is increased against ImP-deficient cells because they express lower levels of surface MHC-I molecules. While this enhanced NK cell activity was studied in the context of influenza virus infection (54), the opposite was found in the context of cancer for which a single cell transcriptomics study revealed elevated activity of NK cells in melanoma tumors expressing high PSMB9 (50). Further work will be needed to better understand the importance of the ImP for NK cells in the context of cancer.
While ImP-mediated enhanced immune cell infiltration can be beneficial for immunogenic cancers, it can also be deleterious for inflammatory cancers such as colitis-induced colorectal carcinoma for which the recruitment and pro-tumoral polarization of immune cells sustain inflammation (55). For instance, PSMB8 was shown to increase the expression of chemokines that attract neutrophils, such as C-X-C motif ligand 1, 2 and 3, as well as vascular cell adhesion molecule 1, which was shown to support inflammation-driven tumorigenesis (55). The impact of the ImP on immune infiltration and cancer immunogenicity can also influence the response to therapy, as this infiltration was identified as important for the efficacy of immune checkpoint inhibitors (56, 57). For instance, NSCLC, melanoma, as well as muscle-invasive bladder, breast and thymic cancers expressing high levels of PSMB8, 9 and 10 were shown to respond better to the treatment (7, 58). PSMB9 was found to be more important in bladder cancer (59) and PSMB8 and 9 in melanoma (42).
Additional functions of the ImP in immune cells are summarized in Table 2. While some of these functions were described in non-cancer contexts and remain to be fully understood, they are likely to also take place within the TME (60, 61). Of note, while PSMB8 and 9 were the first ImP subunits to be discovered (62), PSMB10 was identified later (63) and remains overall less characterized. As such, additional functions in various immune cells will likely be uncovered in the future.
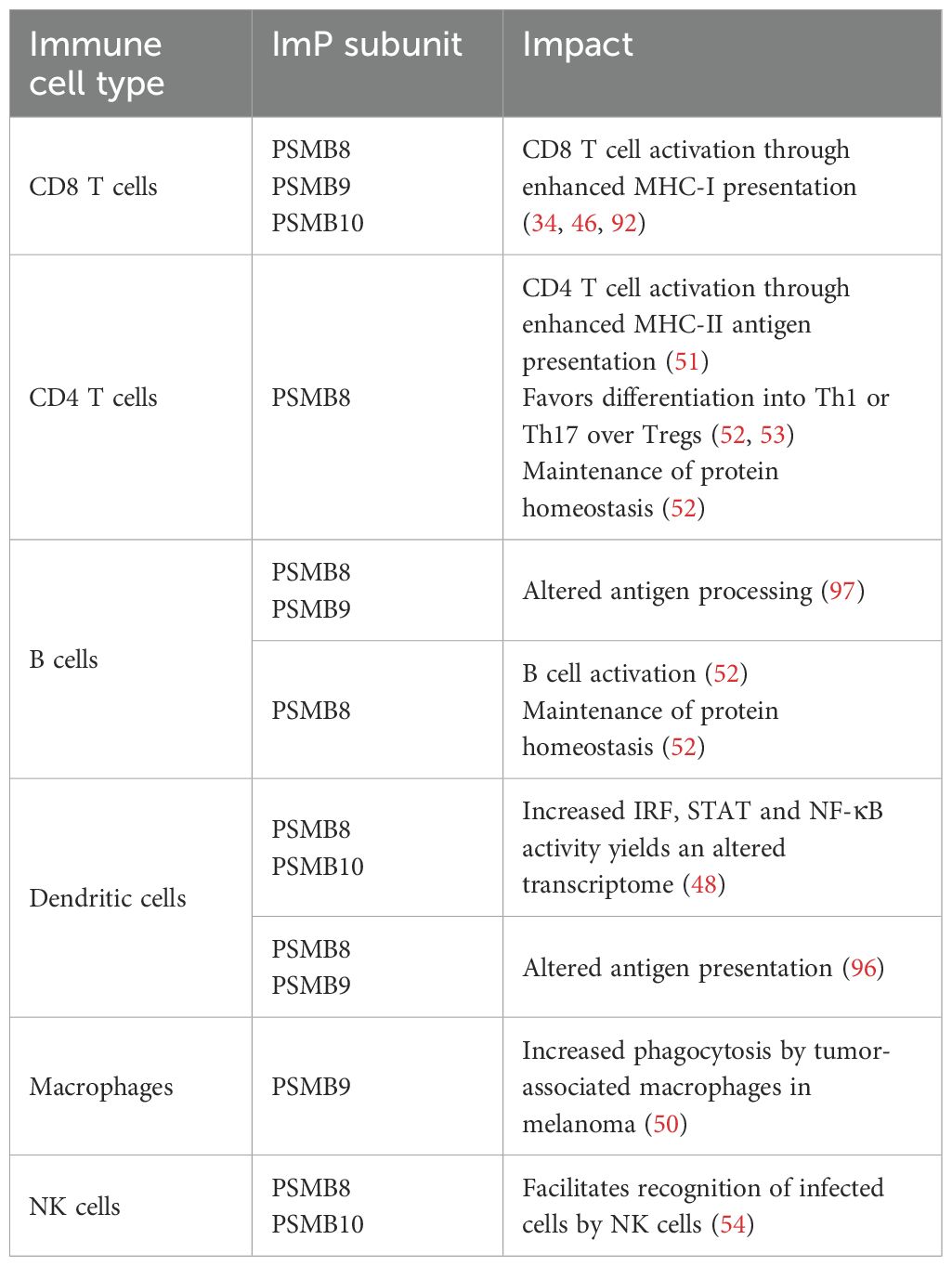
Table 2. Impact of ImP subunits on immune cells found within the TME.
For fibroblasts, the ImP can be induced by IFNγ and interestingly, this induction is lost in the context of senescence (64). Since senescence is often observed in cancer in response to treatments, CAFs might have defects that prevent ImP induction. Also, PSMB8-deficient fibroblasts of the skin have altered responses to oxidative stress, supporting an important role of the ImP in maintaining protein homeostasis (25). While we could not find any study specifically delineating the functions of the ImP in CAFs, its impacts in other types of fibroblasts might also translate in a cancer setting.
Similarly to CAFs, the impact of the ImP on tumor endothelial cells is understudied. In endothelial cells, the ImP can once again be induced by IFNγ (65) and ImP expression by these cells was shown to contribute to CD8 T cell activation and tissue infiltration in the context of hypertension (66). Once again, further studies are needed determine if this also occurs in tumor endothelial cells. As for Imp functions in tumor cells themselves, these are detailed in section 3.
5 ImP modulation in cancer
One hallmark of cancer is genetic instability (67). As such, cancer cells accumulate mutations, some of which perturb cellular processes and aberrant gene expression is often observed. Interestingly, the basal expression of PSMB8, 9 and 10 is often augmented in bladder, breast, head and neck, and subtypes of renal cancers compared to healthy tissues (45, 68). PSMB8 and 9 are also overexpressed in lung, pancreatic, stomach, colon, prostate, thyroid, liver, uterine, cervical, testicular, ovarian and rectal cancers, as well as in diffuse large B cell lymphoma, acute myeloid leukemia, glioma, glioblastoma, cholangioma and melanoma (45, 68–70). ImP expression can also be heterogenous within a tumor. For instance, a study found high expression of PSMB8 in only 20% of NSCLC cells (39). ImP expression can also vary across cancer subtypes. In breast cancer, PSMB8, 9 and 10 have been found to be expressed to higher levels in hormone-positive cancers compared to other subtypes (71). Given the impact of ImP expression on cancer cells, its regulation is often altered in malignant vs healthy cells.
Notably, ImP repression is often observed in cancer. In breast cancer, retinoic acid receptor responder protein 3, a known suppressor of lung metastasis (72), has been shown to downregulate PSMB8, 9 and 10 expression through IRF1 depletion (73). In acute promyelocytic leukemia, the promyelocytic leukemia-retinoic acid receptor-α fusion protein suppresses the function of PU.1, a key transcription factor that is required for ImP expression (Figure 2), and all-trans retinoic acid, which degrades the fusion protein, was shown to restore ImP expression and is seen as a promising therapeutic option of the patients (74). Another mechanism by which cancer cells prevent ImP expression is via epigenetic silencing. For instance, in mesenchymal NSCLC, STAT3 recruits methyltransferases that hyper-methylate the PSMB promoters and therefore prevent ImP expression (75). Likewise, acute myeloid leukemia of the M3 subtype show heavy DNA methylation in the region of PSMB promoters and expresses low ImP levels (68). Finally, CD28 co-stimulation interferes with DNA methylation of the ImP promoters in myeloma models, which prevents expression (76). The aberrant expression of the ImP by cancer cells affects the disease at all steps from carcinogenesis to dissemination and also modulates treatment responses. As such, a deeper understanding of its interplay with cancer is warranted in order to harness its activity to improve outcomes.
6 The ImP in oncogenesis and disease progression
The ImP plays cancer-specific roles at various stages of the disease (Table 3). While its role in cancer immunoediting is established (34, 77, 78), it also impacts tumor onset. As such, PSMB8 is a key driver of oncogenesis in many cancers, including colitis-induced cancers, which are associated with chronic inflammatory conditions (79, 80). Also, another group has demonstrated that PSMB8 knockout (KO) mice are resistant to chronic inflammation and fail to develop tumors upon exposure to carcinogens (55). Additionally, Leister et al. demonstrated that pro-tumorigenic factors such as cyclooxygenase-2, interleukin-6 and interleukin-1β were lower in ImP KO mice (49). Also, correlative studies predicted that PSMB8 has a carcinogenic role in lower-grade glioma, uveal melanoma and pancreatic adenocarcinoma (69). Additionally, in some subtypes of acute myeloid leukemia, genomic rearrangements involving the lysine methyltransferase 2A gene KMT2A yield fusion proteins that drive oncogenesis (81). Interestingly, a study found that inhibiting PSMB8 with the drug ONX-0914 inactivated KMT2A, therefore suggesting a pro-AML role for this ImP subunit (82). In hepatocellular carcinoma, PSMB8 was found to be co-expressed with zinc finger protein 655 a protein that supports cancer proliferation and tumour establishment (83, 84) and PSMB8 knock-down significantly reduced disease severity (85). Taken together, these studies support a pro-cancer role for the ImP in specific cancer types.
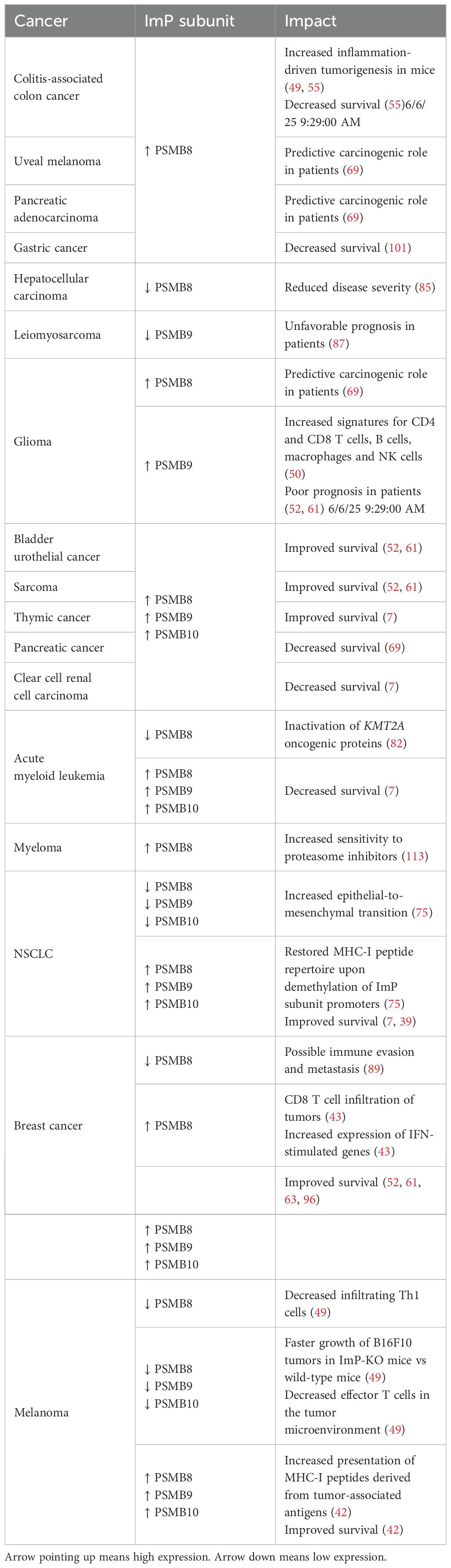
Table 3. Impacts of ImP expression in different cancer types.
Interestingly, the opposite was found for other indications. For instance, 36% of PSMB9 KO mice were found to spontaneously develop uterine leiomyosarcoma (86). For leiomyosarcoma, data from three different cohorts demonstrated that PSMB9low patients presented an increase in pathways driven by the Src proto-oncogene and a less favorable prognosis (87). Importantly, ImP expression also affects disease establishment and dissemination. In NSCLC, deficiencies in ImP subunits drive epithelial-to-mesenchymal transition (75), a phenomenon that contributes to tumor aggressiveness through metastasis (88). In breast cancer patient samples, PSMB8 was shown to be decreased in brain metastases, suggesting that its loss could play a role in metastasis (89). Altogether, these findings highlight the diverse roles of the ImP in cancer establishment and progression.
7 ImP and anti-tumor immunity
Many reports describe the ImP influencing anti-tumor immunity (90–93). For instance, subcutaneous B16F10 melanoma tumors grow faster in ImP KO mice compared to their wild-type counterparts, a phenotype that was associated with decreased effector T cells within the TME and draining lymph nodes (49). In human breast cancer, high PSMB8 expression was associated with increased intra-tumoral CD8 T cell infiltration (43). The authors also found that PSMB8 expression correlated with increased expression of the IFN-stimulated genes myxovirus resistance gene A and protein kinase R, high mobility group nucleosome binding domain 1 and high mobility group box 1 danger-associated molecular patterns, therefore indicating a pro-immune state in the presence of PSMB8 (43). Finally, PSMB8 KO mice exhibit depleted Th1 CD4 T cells within melanoma tumors, which was shown to allow for faster progression (49). Given that T cells are important players of anti-tumor immunity, these effects of the ImP are likely to impact their capacity to recognize and eliminate cancer cells. To do so, T cells scan the antigenic peptides that are presented at the cell surface and specifically unleash their cytotoxic activity against the cells that are not recognized as self.
The collection of peptides that are presented at the cell surface by MHC molecules constitutes the peptide repertoire (or immunopeptidome) (94). Interestingly, dendritic cells from PSMB8/9/10 KO mice have been shown to be defective in the presentation of multiple epitopes (95). As such, the contribution of the ImP to the immunopeptidome in the context of cancer has emerged as a new avenue to increase anti-tumor immunity. Counter-intuitively, many studies reported that the ImP limited the presentation of some tumor epitopes by dendritic cells and B cells (96–98). These results, together with the established differences in the peptides yielded from protein degradation by the CP vs ImP further support an impact of the ImP on the peptide repertoire.
The peptide repertoire of cancer cells is important for their elimination by T cells, and is also a key factor in the efficacy of cancer vaccines. For a peptide vaccine targeting the cancer driver epidermal growth factor receptor variant III (EGFRvIII), a mutation in the peptide sequence enhanced ImP processing and translated into better therapeutic efficacy in the GL261-EGFRvIII glioblastoma model (99). Also, human cell lines of NSCLC were reported to have depleted repertoires of MHC-I peptides, which could be rescued by treatment with the DNA methyltransferase inhibitor 5-aza-2’-deoxycytidine (which demethylates ImP subunit promoters) or IFNγ, both of which increase Imp expression. Further peptide repertoire analyses and in vitro cell-mediated cytotoxicity assays confirmed the therapeutic potential of the approach (75). Altogether, these studies highlight the contribution of the ImP in shaping anti-tumor immune responses through peptide processing and presentation. As such, its expression affects cancer outcomes.
8 ImP and cancer outcomes
Given the ability of the ImP to enrich the tumor milieu with multiple key players of anti-tumor immunity, many groups have studied the link between ImP expression and the prognosis of cancer patients. In breast cancer, NSCLC, bladder urothelial cancer, sarcoma, mesothelioma, melanoma and thymic cancer, elevated levels of PSMB8, 9 or 10 mRNA correlate with improved survival (7, 39, 43, 45, 49, 69, 75). This has been very well studied in breast cancer for which PSMB8 expression was linked to better disease-free survival in patients presenting with lymph node metastases at the time of diagnosis (43). Furthermore, we previously published that triple-negative breast cancers had better prognoses when PSMB8 and 9 protein expression were detected in cancer cells (100). For some other cancers, ImP is associated with a worst prognosis. For example, PSMB8 expression decreased survival in gastric cancer (101) and PSMB9 expression correlated with poor outcomes in glioma (45). Co-expression of the three subunits was also linked to poor outcomes in pancreatic cancer, clear cell renal cell carcinoma and AML (7). These studies highlight the context-dependent effects of ImP expression in cancer.
From a therapeutic standpoint, ImP expression has also been reported to affect treatment responses, notably in the context of immunotherapies. In the last decade, immune checkpoint blockade has revolutionized cancer treatment (102, 103), and many studies have now shown that ImP expression is predictive of treatment response to immune checkpoint inhibitors in melanoma, NSCLC, breast, bladder and thymus cancer (7, 42, 58). Interestingly, ImP expression was found to be a superior at predicting treatment responses compared to the tumor mutational burden (42) and the expression of other IFNγ-induced gene (58). More work is still required to understand the molecular mechanisms linking ImP expression and immune checkpoint blockade sensitivity. The many impacts of ImP expression on the prognosis of cancer patients are summarized in Table 3.
For most hematological cancers, the impact of ImP expression remains poorly characterized (104). Interestingly, the ImP governs protein homeostasis and supports the survival of these cancers, notably because it constitutes the majority of the proteasome pool (105). This ImP dependency makes hematological malignancies ideal candidates for ImP inhibition as a treatment (104, 106–111). Accordingly, the ImP is gaining interest in the field of cancer therapy and its inhibition is being explored as a treatment for some cancers.
9 ImP inhibition in cancer
Inhibiting ImP activity is seen as a promising therapeutic avenue for many cancers (112, 113). The ImP inhibitors that are currently used in clinical and pre-clinical studies are summarized in Table 4. Proteasome inhibitors that target both the CP and the ImP were first explored and bortezomib, carfilzomib and ixazomib are currently approved by the Food and Drug Administration for treatment-refractory or relapsing multiple myeloma (112, 114, 115). These drugs target proteasome subunits with chymotrypsin-like activity (116–118) and worldwide, over 140 clinical trials are currently exploring the efficacy of these drugs against myelomas, lymphomas and leukemias. Notably, bortezomib is also approved in mantle cell lymphoma and is currently undergoing trials for other cancers although severe toxicities and the development of resistance have been reported (119).
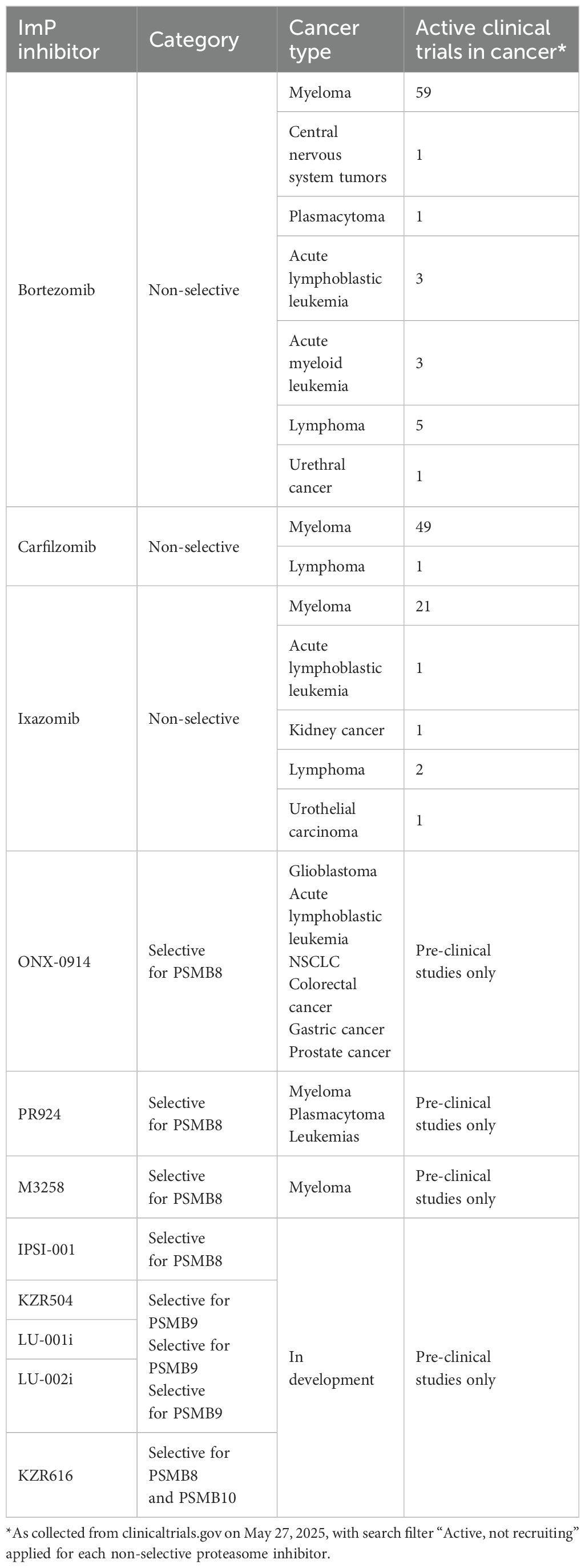
Table 4. Overview of current ImP inhibitor usage in clinical and pre-clinical studies.
As for inhibitors that specifically inhibit the ImP, few have been developed in the last decade. As such, ONX-0914, the most studied ImP inhibitor, blocks PSMB8 and has been reported to induce cell death in pre-clinical models of glioblastoma, acute lymphoblastic leukemia, NSCLC, colorectal, gastric and castration-resistant prostate cancers (55, 107, 120–125). PR-924 is another PSMB8 inhibitor with enhanced ImP selectivity and potency compared to ONX-0914 (126). It was shown to trigger apoptosis in multiple myeloma, prolonged survival in a plasmacytoma xenograft model and was also cytotoxic against bortezomib-resistant leukemia cell lines (127, 128). Another compound targeting PSMB8 is M3258, which was shown to efficiently induce the apoptosis of tumor cells in multiple myeloma xenograft models and presenting a favorable safety profile (110, 129). Additional ImP inhibitors have recently been developed, including IPSI-001, KZR504 and LU-001i, which all target PSMB9, as well as LU-002i which targets PSMB10 (130–133). The small molecule KZR616, which inhibits both PSMB8 and 10 (134), is the only selective ImP inhibitor to currently be tested clinically, although not for a cancer indication, but as a treatment for autoimmunity (NCT04628936, NCT04039477, NCT03393013, NCT04033926, NCT05569759, NCT05781750). The various clinical trials testing proteasome inhibitors against cancer are summarized in Table 4. As we deepen our understanding of the ImP in the context of cancer, it is likely that its therapeutic potential will be established, leading to additional clinical studies aiming at using ImP-selective inhibitors against cancer.
10 Concluding remarks
While the ImP was discovered almost 30 years ago, its role in cancer was only revealed in the last decade. We are currently expanding our knowledge of its dichotomous roles at the different stages of cancer progression, which will likely affect future therapies (Figure 3). Importantly, while most cancers exhibit increased ImP expression compared to healthy tissues, and many ImP inhibitors show promising results in pre-clinical models of the disease, the ImP also supports anti-tumor immunity, which is important for tumor control. This tug-of-war between dampening harmful ImP activity and preserving its pro-immune functions remains to be fully understood. Future research will likely aim at understanding this balance, uncovering optimal ImP targeting strategies based on cancer type, harnessing the potential to mount a favorable anti-tumor immune response, and combining Imp modulation with existing therapies.
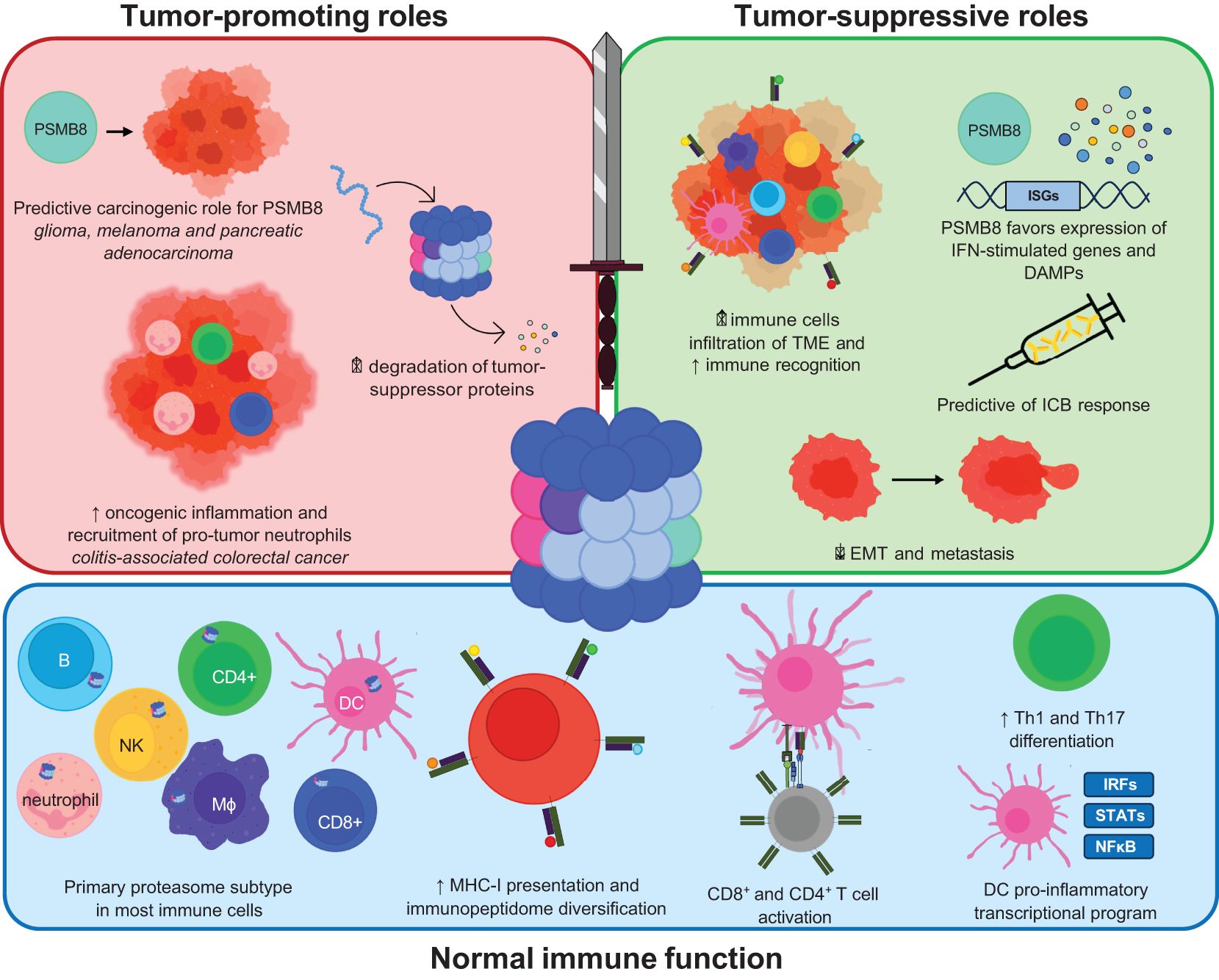
Figure 3. Schematic representation of the functional outcomes of ImP activity in normal immune function, tumor-promotion and tumor-suppression. The roles of the ImP in normal immune functions includes increased MHC-I presentation, immunopeptidome diversification, T cell activation, increased differentiation of CD4+ T cells into Th1 or Th17 cells or influencing the transcriptional program of dendritic cells. Amongst its tumor-promoting roles, the ImP has a predictive carcinogenic role in some cancers and degrades various tumor-suppressor proteins. The ImP also drives oncogenic inflammation in colitis-associated cancers. It may also increase immune cell infiltration in the TME and trigger danger-associated molecular patterns (DAMPs).
Author contributions
DB: Writing – original draft, Conceptualization, Writing – review & editing. MV: Writing – review & editing. EK: Writing – review & editing. M-CB-D: Supervision, Conceptualization, Writing – review & editing, Funding acquisition, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work is funded by grants from the “Institut du cancer de Montréal” and the Cole Foundation to M-CB-D. DB received scholarships from “Fonds de recherche du Québec – Santé” (FRQS), the Faculty of Medicine of the University of Montreal and “Institut du cancer de Montréal”. MV received scholarships from the Faculty of Medicine of the University of Montreal, University of Montreal Department of Microbiology, Infectious Diseases and Immunology and “Institut du cancer de Montréal”. EK received scholarships from the University of Montreal Department of Microbiology, Infectious Diseases and Immunology and “Institut du cancer de Montréal”. M-CB-D received salary awards from the FRQS and the Quebec Breast Cancer Foundation.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Nandi D, Tahiliani P, Kumar A, and Chandu D. The ubiquitin-proteasome system. J Biosci. (2006) 31:137–55. doi: 10.1007/BF02705243
2. Voutsadakis IA. The ubiquitin–proteasome system and signal transduction pathways regulating Epithelial Mesenchymal transition of cancer. J BioMed Sci. (2012) 19:1–13. doi: 10.1186/1423-0127-19-67
3. Rousseau A and Bertolotti A. Regulation of proteasome assembly and activity in health and disease. Nat Rev Mol Cell Biol. (2018) 19:697–712. doi: 10.1038/s41580-018-0040-z
4. Tu Y, Chen C, Pan J, Xu J, Zhou ZG, and Wang CY. The Ubiquitin Proteasome Pathway (UPP) in the regulation of cell cycle control and DNA damage repair and its implication in tumorigenesis. Int J Clin Exp Pathol. (2012) 5:726–38.
5. Pishesha N, Harmand TJ, and Ploegh HL. A guide to antigen processing and presentation. Nat Rev Immunol. (2022) 22:751–64. doi: 10.1038/s41577-022-00707-2
6. Kniepert A and Groettrup M. The unique functions of tissue-specific proteasomes. Trends Biochem Sci. (2014) 39:17–24. doi: 10.1016/j.tibs.2013.10.004
7. Kumar R, Dhaka B, Sahoo S, Jolly MK, and Sabarinathan R. Prognostic association of immunoproteasome expression in solid tumours is governed by the immediate immune environment. Mol Oncol. (2023) 17:1041–59. doi: 10.1002/1878-0261.13443
8. Smith DM, Chang SC, Park S, Finley D, Cheng Y, and Goldberg AL. Docking of the proteasomal ATPases’ carboxyl termini in the 20S proteasome’s alpha ring opens the gate for substrate entry. Mol Cell. (2007) 27:731–44. doi: 10.1016/j.molcel.2007.06.033
9. Bard JAM, Goodall EA, Greene ER, Jonsson E, Dong KC, and Martin A. Structure and function of the 26S proteasome. Annu Rev Biochem. (2018) 87:697–724. doi: 10.1146/annurev-biochem-062917-011931
10. Kasahara M. Role of immunoproteasomes and thymoproteasomes in health and disease. Pathol Int. (2021) 71:371–82. doi: 10.1111/pin.13088
11. Dick TP, Nussbaum AK, Deeg M, Heinemeyer W, Groll M, Schirle M, et al. Contribution of proteasomal β-subunits to the cleavage of peptide substrates analyzed with yeast mutants*. J Biol Chem. (1998) 273:25637–46. doi: 10.1074/jbc.273.40.25637
12. Ferrington DA and Gregerson DS. Immunoproteasomes: structure, function, and antigen presentation. Prog Mol Biol Transl Sci. (2012) 109:75–112. doi: 10.1016/B978-0-12-397863-9.00003-1
13. Angeles A, Fung G, and Luo H. Immune and non-immune functions of the immunoproteasome. Front Biosci Landmark Ed. (2012) 17:1904–16. doi: 10.2741/4027
14. Giles JR, Globig AM, Kaech SM, and Wherry EJ. CD8+ T cells in the cancer-immunity cycle. Immunity. (2023) 56:2231–53. doi: 10.1016/j.immuni.2023.09.005
15. Chen B, Zhu H, Yang B, and Cao J. The dichotomous role of immunoproteasome in cancer: Friend or foe? Acta Pharm Sin B. (2023) 13:1976–89. doi: 10.1016/j.apsb.2022.11.005
16. Tripathi SC, Vedpathak D, and Ostrin EJ. The functional and mechanistic roles of immunoproteasome subunits in cancer. Cells. (2021) 10:3587. doi: 10.3390/cells10123587
17. Eshof BLvd, Medfai L, Nolfi E, Wawrzyniuk M, and Sijts AJAM. The function of immunoproteasomes—An immunologists’ Perspective. Cells. (2021) 10:3360. doi: 10.3390/cells10123360
18. Inholz K, Anderl JL, Klawitter M, Goebel H, Maurits E, Kirk CJ, et al. Proteasome composition in immune cells implies special immune-cell-specific immunoproteasome function. Eur J Immunol. (2024) 54:e2350613. doi: 10.1002/eji.202350613
19. McCarthy MK and Weinberg JB. The immunoproteasome and viral infection: a complex regulator of inflammation. Front Microbiol. (2015) 6:21. doi: 10.3389/fmicb.2015.00021
20. Aki M, Shimbara N, Takashina M, Akiyama K, Kagawa S, Tamura T, et al. Interferon-gamma induces different subunit organizations and functional diversity of proteasomes. J Biochem (Tokyo). (1994) 115:257–69. doi: 10.1093/oxfordjournals.jbchem.a124327
21. Shin EC, Seifert U, Kato T, Rice CM, Feinstone SM, Kloetzel PM, et al. Virus-induced type I IFN stimulates generation of immunoproteasomes at the site of infection. J Clin Invest. (2006) 116:3006–14. doi: 10.1172/JCI29832
22. Callahan MK, Wohlfert EA, Ménoret A, and Srivastava PK. Heat shock up-regulates lmp2 and lmp7 and enhances presentation of immunoproteasome-dependent epitopes. J Immunol Baltim Md 1950. (2006) 177:8393–9. doi: 10.4049/jimmunol.177.12.8393
23. Kotamraju S, Matalon S, Matsunaga T, Shang T, Hickman-Davis JM, and Kalyanaraman B. Upregulation of immunoproteasomes by nitric oxide: potential antioxidative mechanism in endothelial cells. Free Radic Biol Med. (2006) 40:1034–44. doi: 10.1016/j.freeradbiomed.2005.10.052
24. Basler M, Christ M, Goebel H, and Groettrup M. Immunoproteasome upregulation is not required to control protein homeostasis during viral infection. J Immunol Baltim Md 1950. (2021) 206:1697–708. doi: 10.4049/jimmunol.2000822
25. Seifert U, Bialy LP, Ebstein F, Bech-Otschir D, Voigt A, Schröter F, et al. Immunoproteasomes preserve protein homeostasis upon interferon-induced oxidative stress. Cell. (2010) 142:613–24. doi: 10.1016/j.cell.2010.07.036
26. Chatterjee-Kishore M, Wright KL, Ting JP, and Stark GR. How Stat1 mediates constitutive gene expression: a complex of unphosphorylated Stat1 and IRF1 supports transcription of the LMP2 gene. EMBO J. (2000) 19:4111–22. doi: 10.1093/emboj/19.15.4111
27. James AB, Conway AM, and Morris BJ. Regulation of the neuronal proteasome by Zif268 (Egr1). J Neurosci Off J Soc Neurosci. (2006) 26:1624–34. doi: 10.1523/JNEUROSCI.4199-05.2006
28. Zhou P, Zanelli E, Smart M, and David C. Genomic organization and tissue expression of mouse proteasome gene Lmp-2. Genomics. (1993) 16:664–8. doi: 10.1006/geno.1993.1245
29. Meraro D, Gleit-Kielmanowicz M, Hauser H, and Levi BZ. IFN-stimulated gene 15 is synergistically activated through interactions between the myelocyte/lymphocyte-specific transcription factors, PU.1, IFN regulatory factor-8/IFN consensus sequence binding protein, and IFN regulatory factor-4: characterization of a new subtype of IFN-stimulated response element. J Immunol Baltim Md 1950. (2002) 168:6224–31. doi: 10.4049/jimmunol.168.12.6224
30. Mazzi P, Donini M, Margotto D, Wientjes F, and Dusi S. IFN-gamma induces gp91phox expression in human monocytes via protein kinase C-dependent phosphorylation of PU.1. J Immunol Baltim Md 1950. (2004) 172:4941–7. doi: 10.4049/jimmunol.172.8.4941
31. Etzrodt M, Ahmed N, Hoppe PS, Loeffler D, Skylaki S, Hilsenbeck O, et al. Inflammatory signals directly instruct PU.1 in HSCs via TNF. Blood. (2019) 133:816–9. doi: 10.1182/blood-2018-02-832998
32. Niedbala W, Besnard AG, Nascimento DC, Donate PB, Sonego F, Yip E, et al. Nitric oxide enhances Th9 cell differentiation and airway inflammation. Nat Commun. (2014) 5:4575. doi: 10.1038/ncomms5575
33. De M, Jayarapu K, Elenich L, Monaco JJ, Colbert RA, and Griffin TA. Beta 2 subunit propeptides influence cooperative proteasome assembly. J Biol Chem. (2003) 278:6153–9. doi: 10.1074/jbc.M209292200
34. Xu H, van der Jeught K, Zhou Z, Zhang L, Yu T, Sun Y, et al. Atractylenolide I enhances responsiveness to immune checkpoint blockade therapy by activating tumor antigen presentation. J Clin Invest. (2021) 131:e146832, 146832. doi: 10.1172/JCI146832
35. Leone P, Malerba E, Susca N, Favoino E, Perosa F, Brunori G, et al. Endothelial cells in tumor microenvironment: insights and perspectives. Front Immunol. (2024) 15:1367875. doi: 10.3389/fimmu.2024.1367875
36. Tao L, Huang G, Song H, Chen Y, and Chen L. Cancer associated fibroblasts: An essential role in the tumor microenvironment. Oncol Lett. (2017) 14:2611. doi: 10.3892/ol.2017.6497
37. Pagès F, Galon J, Dieu-Nosjean MC, Tartour E, Sautès-Fridman C, and Fridman WH. Immune infiltration in human tumors: a prognostic factor that should not be ignored. Oncogene. (2010) 29:1093–102. doi: 10.1038/onc.2009.416
38. Dakal TC, George N, Xu C, Suravajhala P, and Kumar A. Predictive and prognostic relevance of tumor-infiltrating immune cells: tailoring personalized treatments against different cancer types. Cancers. (2024) 16:1626. doi: 10.3390/cancers16091626
39. Kiuchi T, Tomaru U, Ishizu A, Imagawa M, Iwasaki S, Suzuki A, et al. Expression of the immunoproteasome subunit β5i in non-small cell lung carcinomas. J Clin Pathol. (2021) 74:300–6. doi: 10.1136/jclinpath-2020-206618
40. Canel M, Sławińska AD, Lonergan DW, Kallor AA, Upstill-Goddard R, Davidson C, et al. FAK suppresses antigen processing and presentation to promote immune evasion in pancreatic cancer. Gut. (2023) 73:131–55. doi: 10.1136/gutjnl-2022-327927
41. Liu Q, Hao C, Su P, and Shi J. Down-regulation of HLA class I antigen-processing machinery components in esophageal squamous cell carcinomas: association with disease progression. Scand J Gastroenterol. (2009) 44:960–9. doi: 10.1080/00365520902998679
42. Kalaora S, Lee JS, Barnea E, Levy R, Greenberg P, Alon M, et al. Immunoproteasome expression is associated with better prognosis and response to checkpoint therapies in melanoma. Nat Commun. (2020) 11:896. doi: 10.1038/s41467-020-14639-9
43. Lee M, Song IH, Heo SH, Kim YA, Park IA, Bang WS, et al. Expression of immunoproteasome subunit LMP7 in breast cancer and its association with immune-related markers. Cancer Res Treat. (2019) 51:80–9. doi: 10.4143/crt.2017.500
44. Langguth M, Maranou E, Koskela SA, Elenius O, Kallionpää RE, Birkman EM, et al. TIMP-1 is an activator of MHC-I expression in myeloid dendritic cells with implications for tumor immunogenicity. Genes Immun. (2024) 25:188–200. doi: 10.1038/s41435-024-00274-7
45. Liu J, Yang X, Ji Q, Yang L, Li J, Long X, et al. Immune characteristics and prognosis analysis of the proteasome 20S subunit beta 9 in lower-grade gliomas. Front Oncol. (2022) 12:875131. doi: 10.3389/fonc.2022.875131
46. Rana PS, Ignatz-Hoover JJ, Guo C, Mosley AL, Malek E, Federov Y, et al. Immunoproteasome activation expands the MHC class I immunopeptidome, unmasks neoantigens, and enhances T-cell anti-myeloma activity. Mol Cancer Ther. (2024) 23:1743–60. doi: 10.1158/1535-7163.MCT-23-0931
47. Narukawa T, Yasuda S, Horinaka M, Taniguchi K, Tsujikawa T, Morita M, et al. The novel HDAC inhibitor OBP-801 promotes MHC class I presentation through LMP2 upregulation, enhancing the PD-1-targeting therapy in clear cell renal cell carcinoma. Cancers. (2024) 16:4058. doi: 10.3390/cancers16234058
48. de Verteuil DA, Rouette A, Hardy MP, Lavallée S, Trofimov A, Gaucher É, et al. Immunoproteasomes shape the transcriptome and regulate the function of dendritic cells. J Immunol. (2014) 193:1121–32. doi: 10.4049/jimmunol.1400871
49. Leister H, Luu M, Staudenraus D, Lopez Krol A, Mollenkopf HJ, Sharma A, et al. Pro- and antitumorigenic capacity of immunoproteasomes in shaping the tumor microenvironment. Cancer Immunol Res. (2021) 9:682–92. doi: 10.1158/2326-6066.CIR-20-0492
50. Hu X, Hu Z, Zhang H, Zhang N, Feng H, Jia X, et al. Deciphering the tumor-suppressive role of PSMB9 in melanoma through multi-omics and single-cell transcriptome analyses. Cancer Lett. (2024) 581:216466. doi: 10.1016/j.canlet.2023.216466
51. Zhang Y, Yang X, Bi T, Wu X, Wang L, Ren Y, et al. Targeted inhibition of the immunoproteasome blocks endothelial MHC class II antigen presentation to CD4+ T cells in chronic liver injury. Int Immunopharmacol. (2022) 107:108639. doi: 10.1016/j.intimp.2022.108639
52. Schmidt C, Berger T, Groettrup M, and Basler M. Immunoproteasome inhibition impairs T and B cell activation by restraining ERK signaling and proteostasis. Front Immunol. (2018) 9:2386. doi: 10.3389/fimmu.2018.02386
53. Kalim KW, Basler M, Kirk CJ, and Groettrup M. Immunoproteasome subunit LMP7 deficiency and inhibition suppresses Th1 and Th17 but enhances regulatory T cell differentiation. J Immunol Baltim Md 1950. (2012) 189:4182–93. doi: 10.4049/jimmunol.1201183
54. van Helden MJG, de Graaf N, Bekker CPJ, Boog CJP, Zaiss DMW, and Sijts AJAM. Immunoproteasome-Deficiency Has No Effects on NK Cell Education, but Confers Lymphocytes into Targets for NK Cells in Infected Wild-Type Mice. PloS One. (2011) 6:e23769. doi: 10.1371/journal.pone.0023769
55. Vachharajani N, Joeris T, Luu M, Hartmann S, Pautz S, Jenike E, et al. Prevention of colitis-associated cancer by selective targeting of immunoproteasome subunit LMP7. Oncotarget. (2017) 8:50447–59. doi: 10.18632/oncotarget.14579
56. Ye B, Fan J, Xue L, Zhuang Y, Luo P, Jiang A, et al. iMLGAM: Integrated Machine Learning and Genetic Algorithm-driven Multiomics analysis for pan-cancer immunotherapy response prediction. iMeta. (2025) 4:e70011. doi: 10.1002/imt2.70011
57. Zhang L, Cui Y, Zhou G, Zhang Z, and Zhang P. Leveraging mitochondrial-programmed cell death dynamics to enhance prognostic accuracy and immunotherapy efficacy in lung adenocarcinoma. J Immunother Cancer. (2024) 12:e010008. doi: 10.1136/jitc-2024-010008
58. Ravi A, Hellmann MD, Arniella MB, Holton M, Freeman SS, Naranbhai V, et al. Genomic and transcriptomic analysis of checkpoint blockade response in advanced non-small cell lung cancer. Nat Genet. (2023) 55:807–19. doi: 10.1038/s41588-023-01355-5
59. Wang X, You H, Zhang T, Li Y, Chen X, Basler M, et al. Immunoproteasome subunits are novel signatures for predicting efficacy of immunotherapy in muscle invasive bladder cancer. J Transl Med. (2025) 23:1–16. doi: 10.1186/s12967-025-06207-w
60. de Visser KE and Joyce JA. The evolving tumor microenvironment: From cancer initiation to metastatic outgrowth. Cancer Cell. (2023) 41:374–403. doi: 10.1016/j.ccell.2023.02.016
61. Ah Kioon MD, Pierides M, Pannellini T, Lin G, Nathan CF, and Barrat FJ. Noncytotoxic inhibition of the immunoproteasome regulates human immune cells in vitro and suppresses cutaneous inflammation in the mouse. J Immunol Baltim Md 1950. (2021) 206:1631–41. doi: 10.4049/jimmunol.2000951
62. Martinez CK and Monaco JJ. Homology of proteasome subunits to a major histocompatibility complex-linked LMP gene. Nature. (1991) 353:664–7. doi: 10.1038/353664a0
63. Nandi D, Jiang H, and Monaco JJ. Identification of MECL-1 (LMP-10) as the third IFN-gamma-inducible proteasome subunit. J Immunol Baltim Md 1950. (1996) 156:2361–4.
64. Stratford FLL, Chondrogianni N, Trougakos IP, Gonos ES, and Rivett AJ. Proteasome response to interferon-gamma is altered in senescent human fibroblasts. FEBS Lett. (2006) 580:3989–94. doi: 10.1016/j.febslet.2006.06.029
65. Amersfoort J, Eelen G, and Carmeliet P. Immunomodulation by endothelial cells — partnering up with the immune system? Nat Rev Immunol. (2022) 22:576–88. doi: 10.1038/s41577-022-00694-4
66. de la Visitación N, Chen W, Krishnan J, Van Beusecum JP, Amarnath V, Hennen EM, et al. Immunoproteasomal processing of isoLG-adducted proteins is essential for hypertension. Circ Res. (2024) 134:1276–91. doi: 10.1161/CIRCRESAHA.124.324068
67. Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. (2022) 12:31–46. doi: 10.1158/2159-8290.CD-21-1059
68. Rouette A, Trofimov A, Haberl D, Boucher G, Lavallée VP, D’Angelo G, et al. Expression of immunoproteasome genes is regulated by cell-intrinsic and -extrinsic factors in human cancers. Sci Rep. (2016) 6:34019. doi: 10.1038/srep34019
69. Chen D, Jin C, Dong X, Wen J, Xia E, Wang Q, et al. Pan-cancer analysis of the prognostic and immunological role of PSMB8. Sci Rep. (2021) 11:20492. doi: 10.1038/s41598-021-99724-9
70. Luo Y, Ye Y, Saibaidoula Y, Zhang Y, and Chen Y. Multifaceted investigations of PSMB8 provides insights into prognostic prediction and immunological target in thyroid carcinoma. PloS One. (2025) 20:e0323013. doi: 10.1371/journal.pone.0323013
71. Kondakova I, Sereda E, Sidenko E, Vtorushin S, Vedernikova V, Burov A, et al. Association of proteasome activity and pool heterogeneity with markers determining the molecular subtypes of breast cancer. Cancers. (2025) 17:159. doi: 10.3390/cancers17010159
72. Morales M, Arenas EJ, Urosevic J, Guiu M, Fernández E, Planet E, et al. RARRES3 suppresses breast cancer lung metastasis by regulating adhesion and differentiation. EMBO Mol Med. (2014) 6:865–81. doi: 10.15252/emmm.201303675
73. Anderson AM, Kalimutho M, Harten S, Nanayakkara DM, Khanna KK, and Ragan MA. The metastasis suppressor RARRES3 as an endogenous inhibitor of the immunoproteasome expression in breast cancer cells. Sci Rep. (2017) 7:39873. doi: 10.1038/srep39873
74. Yang XW, Wang P, Liu JQ, Zhang H, Xi WD, Jia XH, et al. Coordinated regulation of the immunoproteasome subunits by PML/RARα and PU.1 in acute promyelocytic leukemia. Oncogene. (2014) 33:2700–8. doi: 10.1038/onc.2013.224
75. Tripathi SC, Peters HL, Taguchi A, Katayama H, Wang H, Momin A, et al. Immunoproteasome deficiency is a feature of non-small cell lung cancer with a mesenchymal phenotype and is associated with a poor outcome. Proc Natl Acad Sci U S A. (2016) 113:E1555–1564. doi: 10.1073/pnas.1521812113
76. Leone P, Malerba E, Prete M, Solimando AG, Croci GA, Ditonno P, et al. Immune escape of multiple myeloma cells results from low miR29b and the ensuing epigenetic silencing of proteasome genes. biomark Res. (2024) 12:43. doi: 10.1186/s40364-024-00592-y
77. Ma W, Vigneron N, Chapiro J, Stroobant V, Germeau C, Boon T, et al. A MAGE-C2 antigenic peptide processed by the immunoproteasome is recognized by cytolytic T cells isolated from a melanoma patient after successful immunotherapy. Int J Cancer. (2011) 129:2427–34. doi: 10.1002/ijc.v129.10
78. Aptsiauri N, Cabrera T, Garcia-Lora A, Lopez-Nevot MA, Ruiz-Cabello F, and Garrido F. MHC class I antigens and immune surveillance in transformed cells. Int Rev Cytol. (2007) 256:139–89. doi: 10.1016/S0074-7696(07)56005-5
79. Parang B, Barrett CW, and Williams CS. AOM/DSS model of colitis-associated cancer. Methods Mol Biol Clifton NJ. (2016) 1422:297–307. doi: 10.1007/978-1-4939-3603-8_26
80. Snider AJ, Bialkowska AB, Ghaleb AM, Yang VW, Obeid LM, and Hannun YA. Murine model for colitis-associated cancer of the colon. Methods Mol Biol Clifton NJ. (2016) 1438:245–54. doi: 10.1007/978-1-4939-3661-8_14
81. Górecki M, Kozioł I, Kopystecka A, Budzyńska J, Zawitkowska J, and Lejman M. Updates in KMT2A gene rearrangement in pediatric acute lymphoblastic leukemia. Biomedicines. (2023) 11:821. doi: 10.3390/biomedicines11030821
82. Tubío-Santamaría N, Jayavelu AK, Schnoeder TM, Eifert T, Hsu CJ, Perner F, et al. Immunoproteasome function maintains oncogenic gene expression in KMT2A-complex driven leukemia. Mol Cancer. (2023) 22:196. doi: 10.1186/s12943-023-01907-7
83. Shao Z, Li C, Wu Q, Zhang X, Dai Y, Li S, et al. ZNF655 accelerates progression of pancreatic cancer by promoting the binding of E2F1 and CDK1. Oncogenesis. (2022) 11:44. doi: 10.1038/s41389-022-00418-2
84. Teng Z, Yao J, Zhu L, Zhao L, and Chen G. ZNF655 is involved in development and progression of non-small-cell lung cancer. Life Sci. (2021) 280:119727. doi: 10.1016/j.lfs.2021.119727
85. Fan S, Liu Y, Lin Z, Zhang Y, Zhang N, Zhao Y, et al. ZNF655 promotes the progression of hepatocellular carcinoma through PSMB8. Cell Biol Int. (2023) 47:1535–46. doi: 10.1002/cbin.12050
86. Hayashi T and Faustman DL. Development of spontaneous uterine tumors in low molecular mass polypeptide-2 knockout mice1. Cancer Res. (2002) 62:24–7.
87. Maia Falcão R, Kokaraki G, De Wispelaere W, Amant F, De Souza GA, de Souza JES, et al. The expression of the immunoproteasome subunit PSMB9 is related to distinct molecular subtypes of uterine leiomyosarcoma. Cancers. (2022) 14:5007. doi: 10.3390/cancers14205007
88. Ribatti D, Tamma R, and Annese T. Epithelial-mesenchymal transition in cancer: A historical overview. Transl Oncol. (2020) 13:100773. doi: 10.1016/j.tranon.2020.100773
89. Adwal A, Kalita-de Croft P, Shakya R, Lim M, Kalaw E, Taege LD, et al. Tradeoff between metabolic i-proteasome addiction and immune evasion in triple-negative breast cancer. Life Sci Alliance. (2020) 3:e201900562. doi: 10.26508/lsa.201900562
90. Matsuda-Lennikov M, Ohigashi I, and Takahama Y. Tissue-specific proteasomes in generation of MHC class I peptides and CD8+ T cells. Curr Opin Immunol. (2022) 77:102217. doi: 10.1016/j.coi.2022.102217
91. Sijts EJ a. M and Kloetzel PM. The role of the proteasome in the generation of MHC class I ligands and immune responses. Cell Mol Life Sci CMLS. (2011) 68:1491–502. doi: 10.1007/s00018-011-0657-y
92. Wang X, Zhang H, Wang Y, Bramasole L, Guo K, Mourtada F, et al. DNA sensing via the cGAS/STING pathway activates the immunoproteasome and adaptive T-cell immunity. EMBO J. (2023) 42:e110597. doi: 10.15252/embj.2022110597
93. Morozov AV and Karpov VL. Proteasomes and several aspects of their heterogeneity relevant to cancer. Front Oncol. (2019) 9:761. doi: 10.3389/fonc.2019.00761
94. Hammer GE, Kanaseki T, and Shastri N. The final touches make perfect the peptide-MHC class I repertoire. Immunity. (2007) 26:397–406. doi: 10.1016/j.immuni.2007.04.003
95. Kincaid EZ, Che JW, York I, Escobar H, Reyes-Vargas E, Delgado JC, et al. Mice completely lacking immunoproteasomes show major changes in antigen presentation. Nat Immunol. (2011) 13:129–35. doi: 10.1038/ni.2203
96. Morel S, Lévy F, Burlet-Schiltz O, Brasseur F, Probst-Kepper M, Peitrequin AL, et al. Processing of some antigens by the standard proteasome but not by the immunoproteasome results in poor presentation by dendritic cells. Immunity. (2000) 12:107–17. doi: 10.1016/S1074-7613(00)80163-6
97. Anderson KS, Zeng W, Sasada T, Choi J, Riemer AB, Su M, et al. Impaired tumor antigen processing by immunoproteasome-expressing CD40-activated B cells and dendritic cells. Cancer Immunol Immunother CII. (2011) 60:857–67. doi: 10.1007/s00262-011-0995-5
98. Ito M, Koido S, Iwamoto T, Morimoto S, Fujiki F, Sugiyama H, et al. Enhancing the immunogenicity of Wilms tumor 1 epitope in mesothelioma cells with immunoproteasome inhibitors. PloS One. (2024) 19:e0308330. doi: 10.1371/journal.pone.0308330
99. Fidanza M, Gupta P, Sayana A, Shanker V, Pahlke SM, Vu B, et al. Enhancing proteasomal processing improves survival for a peptide vaccine used to treat glioblastoma. Sci Transl Med. (2021) 13:eaax4100. doi: 10.1126/scitranslmed.aax4100
100. Geoffroy K, Araripe Saraiva B, Viens M, Béland D, and Bourgeois-Daigneault MC. Increased expression of the immunoproteasome subunits PSMB8 and PSMB9 by cancer cells correlate with better outcomes for triple-negative breast cancers. Sci Rep. (2023) 13:2129. doi: 10.1038/s41598-023-28940-2
101. Kwon CH, Park HJ, Choi YR, Kim A, Kim HW, Choi JH, et al. PSMB8 and PBK as potential gastric cancer subtype-specific biomarkers associated with prognosis. Oncotarget. (2016) 7:21454–68. doi: 10.18632/oncotarget.v7i16
102. Darvin P, Toor SM, Sasidharan Nair V, and Elkord E. Immune checkpoint inhibitors: recent progress and potential biomarkers. Exp Mol Med. (2018) 50:1–11. doi: 10.1038/s12276-018-0191-1
103. Madden K and Kasler MK. Immune checkpoint inhibitors in lung cancer and melanoma. Semin Oncol Nurs. (2019) 35:150932. doi: 10.1016/j.soncn.2019.08.011
104. Besse A, Kraus M, Mendez-Lopez M, Maurits E, Overkleeft HS, Driessen C, et al. Immunoproteasome activity in chronic lymphocytic leukemia as a target of the immunoproteasome-selective inhibitors. Cells. (2022) 11:838. doi: 10.3390/cells11050838
105. Niewerth D, Franke NE, Jansen G, Assaraf YG, van Meerloo J, Kirk CJ, et al. Higher ratio immune versus constitutive proteasome level as novel indicator of sensitivity of pediatric acute leukemia cells to proteasome inhibitors. Haematologica. (2013) 98:1896–904. doi: 10.3324/haematol.2013.092411
106. Besse L, Besse A, Kraus M, Maurits E, Overkleeft HS, Bornhauser B, et al. High Immunoproteasome Activity and sXBP1 in Pediatric Precursor B-ALL Predicts Sensitivity towards Proteasome Inhibitors. Cells. (2021) 10:2853. doi: 10.3390/cells10112853
107. Jenkins TW, Downey-Kopyscinski SL, Fields JL, Rahme GJ, Colley WC, Israel MA, et al. Activity of immunoproteasome inhibitor ONX-0914 in acute lymphoblastic leukemia expressing MLL-AF4 fusion protein. Sci Rep. (2021) 11:10883. doi: 10.1038/s41598-021-90451-9
108. Jenkins TW, Fitzgerald JE, Park J, Wilson AM, Berry KL, Wong KS, et al. Highly specific Immunoproteasome inhibitor M3258 induces proteotoxic stress and apoptosis in KMT2A::AFF1 driven acute lymphoblastic leukemia. Sci Rep. (2025) 15:17284. doi: 10.1038/s41598-025-01657-0
109. Zhang L, Pham LV, Newberry KJ, Ou Z, Liang R, Qian J, et al. In vitro and in vivo therapeutic efficacy of carfilzomib in mantle cell lymphoma: targeting the immunoproteasome. Mol Cancer Ther. (2013) 12:2494–504. doi: 10.1158/1535-7163.MCT-13-0156
110. Sanderson MP, Friese-Hamim M, Walter-Bausch G, Busch M, Gaus S, Musil D, et al. M3258 is a selective inhibitor of the immunoproteasome subunit LMP7 (β5i) delivering efficacy in multiple myeloma models. Mol Cancer Ther. (2021) 20:1378–87. doi: 10.1158/1535-7163.MCT-21-0005
111. Breczko W, Lemancewicz D, Dzięcioł J, Kłoczko J, and Bołkun Ł. High immunoproteasome concentration in the plasma of patients with newly diagnosed multiple myeloma treated with bortezomib is predictive of longer OS. Adv Med Sci. (2021) 66:21–7. doi: 10.1016/j.advms.2020.11.003
112. Xi J, Zhuang R, Kong L, He R, Zhu H, and Zhang J. Immunoproteasome-selective inhibitors: An overview of recent developments as potential drugs for hematologic Malignancies and autoimmune diseases. Eur J Med Chem. (2019) 182:111646. doi: 10.1016/j.ejmech.2019.111646
113. Ettari R, Zappalà M, Grasso S, Musolino C, Innao V, and Allegra A. Immunoproteasome-selective and non-selective inhibitors: A promising approach for the treatment of multiple myeloma. Pharmacol Ther. (2018) 182:176–92. doi: 10.1016/j.pharmthera.2017.09.001
114. Miller Z, Lee W, and Kim KB. The immunoproteasome as a therapeutic target for hematological Malignancies. Curr Cancer Drug Targets. (2014) 14:537–48. doi: 10.2174/1568009614666140723113139
115. Kortuem KM and Stewart AK. Carfilzomib. Blood. (2013) 121:893–7. doi: 10.1182/blood-2012-10-459883
116. Lü S and Wang J. The resistance mechanisms of proteasome inhibitor bortezomib. biomark Res. (2013) 1:13. doi: 10.1186/2050-7771-1-13
117. Kuhn DJ, Chen Q, Voorhees PM, Strader JS, Shenk KD, Sun CM, et al. Potent activity of carfilzomib, a novel, irreversible inhibitor of the ubiquitin-proteasome pathway, against preclinical models of multiple myeloma. Blood. (2007) 110:3281–90. doi: 10.1182/blood-2007-01-065888
118. Richardson PG, Baz R, Wang M, Jakubowiak AJ, Laubach JP, Harvey RD, et al. Phase 1 study of twice-weekly ixazomib, an oral proteasome inhibitor, in relapsed/refractory multiple myeloma patients. Blood. (2014) 124:1038–46. doi: 10.1182/blood-2014-01-548826
119. Cengiz Seval G and Beksac M. The safety of bortezomib for the treatment of multiple myeloma. Expert Opin Drug Saf. (2018) 17:953–62. doi: 10.1080/14740338.2018.1513487
120. Koerner J, Brunner T, and Groettrup M. Inhibition and deficiency of the immunoproteasome subunit LMP7 suppress the development and progression of colorectal carcinoma in mice. Oncotarget. (2017) 8:50873–88. doi: 10.18632/oncotarget.15141
121. Monittola F, Bianchi M, Nasoni MG, Luchetti F, Magnani M, and Crinelli R. Gastric cancer cell types display distinct proteasome/immunoproteasome patterns associated with migration and resistance to proteasome inhibitors. J Cancer Res Clin Oncol. (2023) 149:10085–97. doi: 10.1007/s00432-023-04948-z
122. Li Y, Dong S, Tamaskar A, Wang H, Zhao J, Ma H, et al. Proteasome inhibitors diminish c-met expression and induce cell death in non-small cell lung cancer cells. Oncol Res. (2020) 28:497–507. doi: 10.3727/096504020X15929939001042
123. Li J, Liu N, Zhou H, Xian P, Song Y, Tang X, et al. Immunoproteasome inhibition prevents progression of castration-resistant prostate cancer. Br J Cancer. (2023) 128:1377–90. doi: 10.1038/s41416-022-02129-2
124. Chang HH, Lin YH, Chen TM, Tsai YL, Lai CR, Tsai WC, et al. ONX-0914 induces apoptosis and autophagy with p53 regulation in human glioblastoma cells. Cancers. (2022) 14:5712. doi: 10.3390/cancers14225712
125. Koerner J, Horvath D, Oliveri F, Li J, and Basler M. Suppression of prostate cancer and amelioration of the immunosuppressive tumor microenvironment through selective immunoproteasome inhibition. Oncoimmunology. (2023) 12:2156091. doi: 10.1080/2162402X.2022.2156091
126. Huber EM, Basler M, Schwab R, Heinemeyer W, Kirk CJ, Groettrup M, et al. Immuno- and constitutive proteasome crystal structures reveal differences in substrate and inhibitor specificity. Cell. (2012) 148:727–38. doi: 10.1016/j.cell.2011.12.030
127. Singh AV, Bandi M, Aujay MA, Kirk CJ, Hark DE, Raje N, et al. PR-924, a selective inhibitor of the immunoproteasome subunit LMP-7, blocks multiple myeloma cell growth both in vitro and in vivo. Br J Haematol. (2011) 152:155–63. doi: 10.1111/j.1365-2141.2010.08491.x
128. Niewerth D, van Meerloo J, Jansen G, Assaraf YG, Hendrickx TC, Kirk CJ, et al. Anti-leukemic activity and mechanisms underlying resistance to the novel immunoproteasome inhibitor PR-924. Biochem Pharmacol. (2014) 89:43–51. doi: 10.1016/j.bcp.2014.02.005
129. Sloot W, Glaser N, Hansen A, Hellmann J, Jaeckel S, Johannes S, et al. Improved nonclinical safety profile of a novel, highly selective inhibitor of the immunoproteasome subunit LMP7 (M3258). Toxicol Appl Pharmacol. (2021) 429:115695. doi: 10.1016/j.taap.2021.115695
130. Kuzina ES, Kudriaeva AA, Maltseva DV, and Belogurov AA. Peptidyl aldehyde specifically interacts with immunosubunit β1i proteasome: in vitro and in vivo effects. Bull Exp Biol Med. (2016) 161:69–71. doi: 10.1007/s10517-016-3347-9
131. Johnson HWB, Anderl JL, Bradley EK, Bui J, Jones J, Arastu-Kapur S, et al. Discovery of highly selective inhibitors of the immunoproteasome low molecular mass polypeptide 2 (LMP2) subunit. ACS Med Chem Lett. (2017) 8:413–7. doi: 10.1021/acsmedchemlett.6b00496
132. de Bruin G, Huber EM, Xin BT, van Rooden EJ, Al-Ayed K, Kim KB, et al. Structure-based design of β1i or β5i specific inhibitors of human immunoproteasomes. J Med Chem. (2014) 57:6197–209. doi: 10.1021/jm500716s
133. Xin BT, Huber EM, de Bruin G, Heinemeyer W, Maurits E, Espinal C, et al. Structure-based design of inhibitors selective for human proteasome β2c or β2i subunits. J Med Chem. (2019) 62:1626–42. doi: 10.1021/acs.jmedchem.8b01884
134. Johnson HWB, Lowe E, Anderl JL, Fan A, Muchamuel T, Bowers S, et al. Required immunoproteasome subunit inhibition profile for anti-inflammatory efficacy and clinical candidate KZR-616 ((2 S,3 R)- N-(( S)-3-(Cyclopent-1-en-1-yl)-1-(( R)-2-methyloxiran-2-yl)-1-oxopropan-2-yl)-3-hydroxy-3-(4-methoxyphenyl)-2-(( S)-2-(2-morpholinoacetamido)propanamido)propenamide). J Med Chem. (2018) 61:11127–43. doi: 10.1021/acs.jmedchem.8b01201
Keywords: immunoproteasome, cancer, oncogenesis, anti-tumor immunity, clinical outcome
Citation: Béland D, Viens M, Kalin EM and Bourgeois-Daigneault M-C (2025) From oncogenesis to prognosis: the roles of the immunoproteasome in cancer. Front. Immunol. 16:1603816. doi: 10.3389/fimmu.2025.1603816
Received: 01 April 2025; Accepted: 23 June 2025;
Published: 08 July 2025.
Edited by:
Chunhua Song, The Ohio State University, United StatesReviewed by:
Byung-Gyu Kim, Case Western Reserve University, United StatesJiacheng Jin, The Ohio State University, United States
Copyright © 2025 Béland, Viens, Kalin and Bourgeois-Daigneault. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie-Claude Bourgeois-Daigneault, bWFyaWUtY2xhdWRlLmJvdXJnZW9pcy1kYWlnbmVhdWx0QHVtb250cmVhbC5jYQ==