 Anjana Aravind1
Anjana Aravind1 Vineetha Shaji1
Vineetha Shaji1 Mukhtar Ahmed2Shobha Dagamajalu1Manavalan Vijayakumar3*
Mukhtar Ahmed2Shobha Dagamajalu1Manavalan Vijayakumar3* Thottethodi Subrahmanya Keshava Prasad1*
Thottethodi Subrahmanya Keshava Prasad1* Rajesh Raju1,4*
Rajesh Raju1,4*- 1Center for Systems Biology and Molecular Medicine, Yenepoya Research Centre, Yenepoya (Deemed to be University), Mangalore, Karnataka, India
- 2Department of Zoology, College of Science, King Saud University, Riyadh, Saudi Arabia
- 3Department of Surgical Oncology, Yenepoya Medical College, Yenepoya (Deemed to be University), Mangalore, Karnataka, India
- 4Center for Integrative Omics Data Science (CIODS), Yenepoya (Deemed to be University), Mangalore, Karnataka, India
Introduction: Interleukin-24 (IL-24) is a cytokine belonging to the IL-10 family with immunoregulatory properties. It is known to induce cellular responses in various disease conditions, including inflammatory, infectious diseases, and cancer, in a receptor-dependent and independent manner. IL-24 is induced by both immune and non-immune cells and acts on a vast array of target cells to regulate various biological processes. The antitumor and immunoregulatory properties of IL-24 support its application as a therapeutic agent in a vast array of pathological conditions. Characterizing the molecular events associated with its immunomodulatory properties is essential for evaluating its utility in cancer immunotherapy.
Methods: The data related to signaling events induced by IL-24 were then manually annotated and cataloged from the literature pertaining to IL-24 signaling. The cataloged molecular events were then manually drawn based on the topology as a signaling pathway map of IL-24.
Results: Based on the data mined from published literature, we assembled IL-24-regulated signaling events linked to its immunoregulatory and anti-tumor activities. The receptor-ligand interaction of IL-24 regulates multiple downstream signaling cascades, leading to antitumor activities alongside inducing inflammatory reactions and anti-infectious processes. We provide a signaling pathway map of IL-24 consisting of 433 mRNAs/proteins and their 613 signaling events.
Discussion: We believe this map would serve as a knowledge base for reference and one-go visualization of the signaling events identified to be induced by IL-24. Moreover, this map is made available (https://ciods.in/il24/) for the researchers to explore the role of IL-24 in various pathophysiological conditions through the gene set enrichment of transcriptomics or proteomics datasets from various disease conditions.
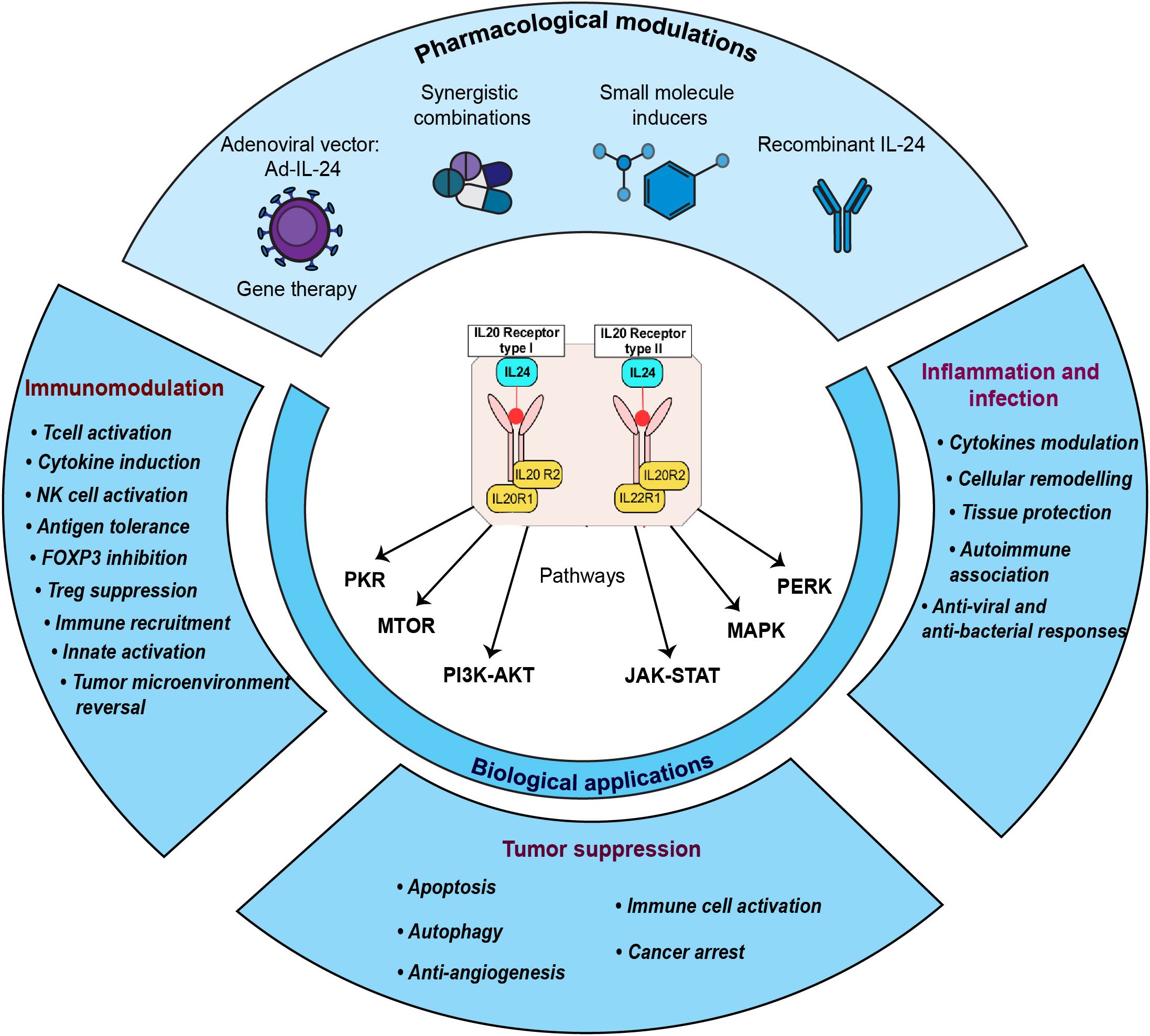
Graphical Abstract.
1 Introduction
Interleukin 24 (IL-24) is a multifunctional cytokine belonging to the IL-10 subfamily with immunomodulatory functions (1–3). It is also known as melanoma differentiation-associated gene-7 (MDA-7), as it was first isolated from growth-arrested and terminally differentiated human melanoma cells using subtractive hybridization (4). It is also known as ST16 (suppressor of tumorigenicity-16) and was renamed IL-24 after the discovery of cell surface receptors of MDA-7 (5–9). IL-24, with its unique cytokine properties, has dual functions as a pro- and anti-inflammatory agent and as a tumor suppressor cytokine (10, 11). It exhibits the ability to selectively induce cell death in various cancer cells (2, 10, 11). It is a pleiotropic cytokine studied in several inflammatory and cancer conditions, known for its characteristics to modulate immune cell activity with diverse functions depending on the target cell type and disease state (10). This secretory cytokine consists of 206 amino acid residues, with 51 amino acids serving as a signal peptide (12). The human IL-24 gene is located on chromosome 1, within a 195-kb cytokine cluster named the “IL-10 family gene cluster,” comprising four genes, IL-10, IL-19, IL-20, and IL-24. IL-24 shares a high sequence homology at the amino acid level across the species (9).
IL-24 is produced by immune cells of both lymphoid and myeloid lineages when activated with certain lipopolysaccharides or specific cytokines and non-immune cells, including melanocytes (1, 12). It originates mainly from activated monocytes and T helper 2 cells. IL-24 functions as a classical cytokine, exerting its effects through its cell-surface heterodimeric receptor complex (IL-20R1 and IL-20R2; IL-22R1 and IL-20R2). IL-20R1 and IL-22R1 pair with the IL-20R2 chain to form a heterodimeric receptor complex, thereby inducing cell signaling events (13, 14). The IL20R1 and IL22R1 expression levels determine the quantity of these two receptor complexes. The expression of the three subunits was consistent in the majority of the tissues and cellular conditions (6, 15). This heterodimeric receptor complex is a part of the IL-10 receptor family and is shared among cytokines within this family, which displays a 20-30% sequence homology (6–8). The utilization of shared receptors is a common mechanism within the cytokine family, leading to the cross-talk and precise regulation of cellular response in the immune system (9). Moreover, IL-24 may interact differently with type I and type II receptor heterodimers, exerting non-redundant and unique biological activities (15). In addition to receptor-ligand interaction-induced signaling events, IL-24 initiates multiple signaling events intracellularly, independent of its non-cognate receptors (16, 17).
IL-24 is a versatile cytokine that can regulate anti-tumor activity, immune response, tissue homeostasis, and host defense mechanisms (3, 10). This multifunctional protein affects broad types of cancer and has a crucial role in regulating cell proliferation, survival, tumor suppression, differentiation, and apoptosis-inducing properties (18, 19). It is well-known for its multi-pronged indirect anti-tumor activities, including the induction of the release of other cytokines, resulting in immunomodulatory activities (1, 20, 21). It can also regulate the proliferation and activation of CD4+ and CD8+ T cell populations (22, 23). Due to its remarkable tumor-suppressing ability, IL-24 shows greater potential for advanced targeted cancer therapy. Recent reports have demonstrated the immunotherapeutic potential of IL-24, with its immunomodulatory activity combined with the apoptotic and anti-angiogenic properties, strengthening its utility in cancer immunotherapy (24–28). A Phase I clinical trial using an adenovirus vector expressing IL-24 (Ad-IL-24) has documented the T cell activation and cytokine release for the eradication of tumor cells (28–36). The strategic utilization of IL-24 in cancer therapy leads to a promising method of precision medicine, navigating toward an effective approach in cancer treatment.
Despite the importance of IL-24-mediated signaling, comprehensive information on IL-24-mediated signaling events is currently limited. The KEGG and Reactome pathway databases only offer a general overview of the signaling modules regulated by IL-24. The potential roles of IL-24 as a cytokine in the form of a comprehensive signaling pathway map will encourage further investigation into IL-24-mediated cellular events. This may lead to unraveling specific roles of individual molecules and modules in various pathophysiological conditions, including cancer, inflammatory disorders, and other diseases. Previously, we compiled the signaling events of several other cytokines, including CCL18, CCL19, IL-11, IL-17, IL-19, and 1L-20, through the documentation of their cell-specific functional roles attributed to diverse experimental conditions (37–41). Similarly, the study-centric molecular reactions of IL-24 were assembled, adding them to the compendium of cytokine signaling pathway maps for gene set enrichment analyses.
2 Methods
2.1 Data curation of IL-24 signaling events
To create a comprehensive IL-24 signaling pathway map, a literature survey was carried out on IL-24 signaling. The search terms used were “IL-24 AND Signaling,” and the articles were retrieved from PubMed. The abstract of research articles describing the signaling events induced by IL-24 was manually screened to select the relevant ones. The experimental datasets were limited to the signal transduction events induced by IL-24, excluding the knock-out and co-expression studies. The curated datasets encompass studies pertaining to diseases such as cancer, infectious, and inflammatory conditions.
2.2 Functional annotation and assembly of the signaling pathway data
The data related to signaling events induced by IL-24 were then manually annotated and cataloged according to the PathBuilder criteria (Figure 1) (42). IL-24-induced molecular events such as protein-protein interactions, protein expression, gene regulation events, activation/inhibition events, enzyme catalysis/post-translational modifications (PTM), molecular association, and protein translocation were curated from the research articles (43). The protein and gene expression events were cataloged for overexpressed (fold change >1.3, p-value <0.05) and downregulated (fold change <0.76, p-value <0.05) molecules, as devised in each of the studies. All possible PTM events, including acetylation, ubiquitination, denitrosylation, N-glycosylation, phosphorylation, and dephosphorylation, with their site-specific information, were annotated within the pathway map and are delineated with different colors. Further, the translocation events were categorized based on the transportation of proteins within the cellular organelles comprising the nucleus, plasma membrane, cytoplasm, mitochondria, and Golgi complex. The proteins and genes were mapped to their corresponding Entrez Gene ID to annotate the data in a standardized format. Besides the proteins, the involvement of small molecules or second messengers such as reactive oxygen species (ROS), Ca2+, and cAMP was also noted. Individual study-centric signaling events and their related disease conditions and cell type-specific reaction events were documented. The data also included the type of experiments performed to detect and validate the expression of downstream molecules of IL-24 ligand-receptor interaction and the cell line/tissue used. The data on the synergistic activities of IL-24 were curated using the same data annotation strategy and included information on the impact of drugs and other molecules of therapeutic potential.
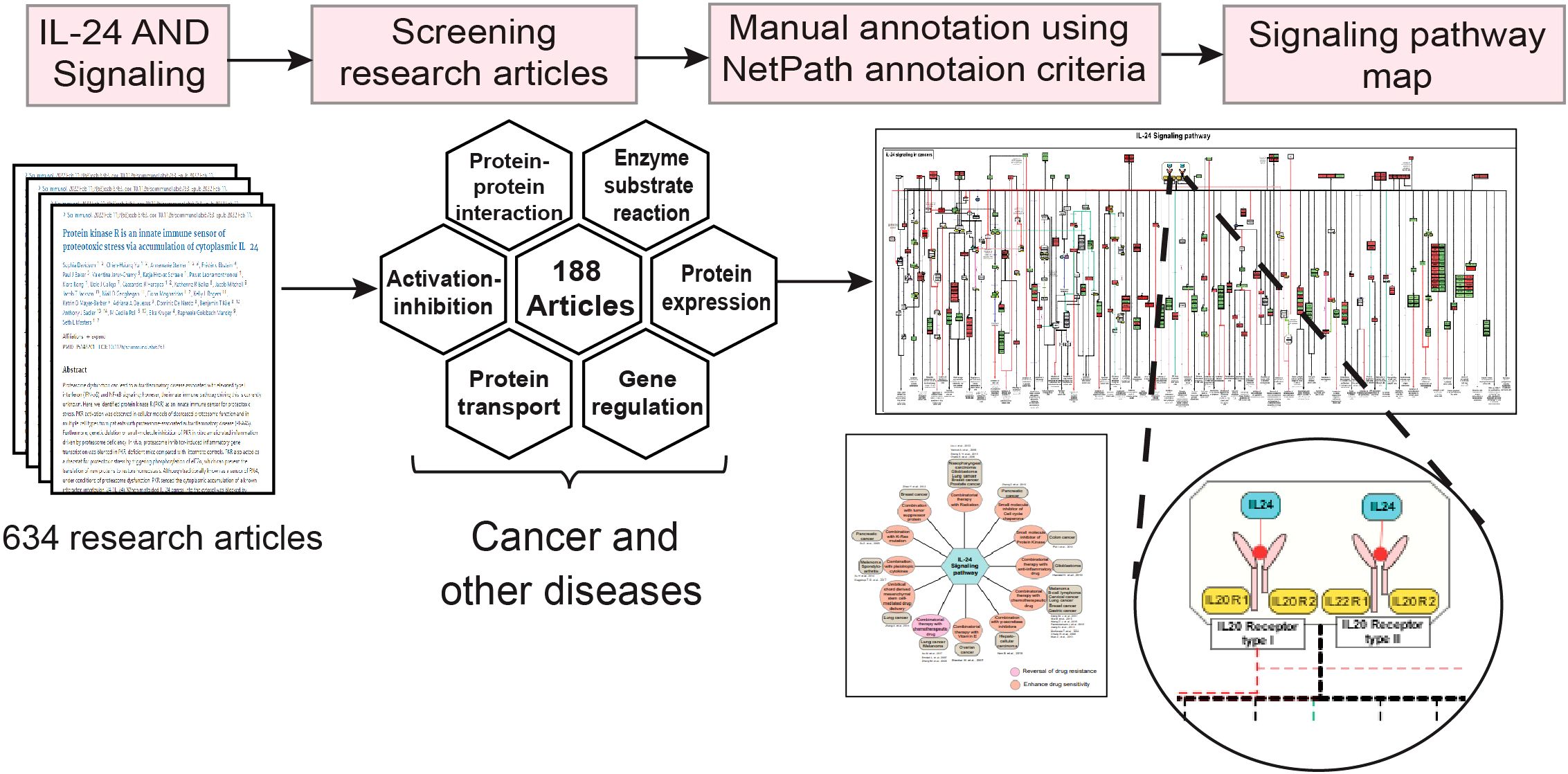
Figure 1. A detailed workflow outlining the methodology for the IL-24 signaling pathway data curation, annotation, and map development. The workflow depicts the sequential steps used to develop the IL-24 signaling pathway map. The literature related to IL-24 signaling in cancer, infectious, and inflammatory disease conditions was manually screened, followed by the curation and annotation of the signaling events according to the NetPath criteria. Further, with the annotated data, a detailed signaling pathway map was developed.
2.3 Development of the IL-24 network map
The annotated molecules, reactions, and disease-specific biological processes were manually drawn based on the topology as a signaling pathway map of IL-24 using the PathVisio (version 3) tool (44). The topology that represents the spatial order of the molecules was devised based on the directionality initiated by the IL-24 and its receptor interaction, as determined by studies using inhibitors or the established cascade of reactions.
2.4 Omics data integration and PPI mapping
The protein-protein interaction network analysis was performed with IL-24, its receptors, and downstream targets. The analysis was carried out using the tool STRING database with a confidence score of >0.7 (45). To enrich the functional clusters, the network was imported into the Cytoscape tool v 3.10.2, and the algorithm Molecular Complex Detection (MCODE) was applied (46, 47). MCODE identifies the clusters based on their local neighborhood density and expands from highly connected hubs. Additionally, publicly available omics datasets were analyzed to provide supporting evidence for the pathway map. The PBMC transcriptomic data were retrieved from the NCBI GEO DataSets and analyzed using GEO2r to assess the expression of IL-24 across various disease conditions. Similarly, differentially expressed proteins were obtained from the publicly available proteomic dataset, and the pathway enrichment analysis was performed using the Reactome database (48).
3 Results and discussion
To develop the IL-24 signaling pathway map, a total of 697 articles were screened in PubMed using the search criteria described in the methods section. A total of 341 molecules involved in IL-24-mediated 602 signal transduction events were manually vetted from the 191 studies. These events include 79 enzyme-substrate reactions, 15 protein-protein interactions, 76 activation/inhibition reactions, 10 protein translocation events, 262 protein expressions, and 171 gene regulation events identified from the annotations of these selected articles. Information on differentially regulated mRNA/proteins responding to IL-24 stimulation is also available. Among the 171 genes, 93 were overexpressed, and 78 were downregulated, whereas in 262 proteins, 132 were overexpressed, and 130 were downregulated. As far as we know, this is the first pathway resource that catalogs IL-24-mediated molecular reactions. The network map depicts the molecular events involved in a wide range of IL-24-induced processes, including host defense, tissue homeostasis, and immunological regulation in various physiological conditions.
The study-centric molecular reactions of IL-24 in various pathological conditions comprising cancer, infection, and other diseases were manually annotated, assembled, and represented as a comprehensive signaling pathway map (Figure 2). The detailed annotated data that were analyzed for building the IL-24 pathway map are provided in the Supplementary File S1. As supported by the assembled data, the functional role of IL-24 is facilitated either through the heterodimeric receptor complexes (IL-20R1 and IL-20R2; IL-22R1 and IL-20R2) or intracellularly, independent of the receptors. The receptor-mediated functional role of IL-24 was observed to be initiated through both of these receptor complexes in the majority of the investigated studies. The IL-24-IL-20R1/IL-20R2 and IL-24-IL-22R1/IL-20R2 ligand-receptor interactions are reported to mediate the cellular responses through multiple signaling modules encompassing JAK-STAT, PI3K-AKT, mTOR, CHOP, MAPK, PERK, and PKR. This heterodimeric receptor complex is also shared within the other IL-10 cytokine family members, allowing an overlapping core signaling mechanism, particularly involving the IL-22R1 and IL-20R2 and the JAK-STAT signaling pathway in epithelial cells, keratinocytes, and immune cells (49). Even though IL-24 is structurally related to other IL-10 family members and shares some signaling machinery, it is unique in its anti-tumor properties, endoplasmic reticulum (ER) stress, and reactive oxygen species (ROS) production that induce selective toxicity in cancer cells (11, 50). These intricate signaling networks contribute to the diverse physiological effects elicited by IL-24 in various biological contexts.
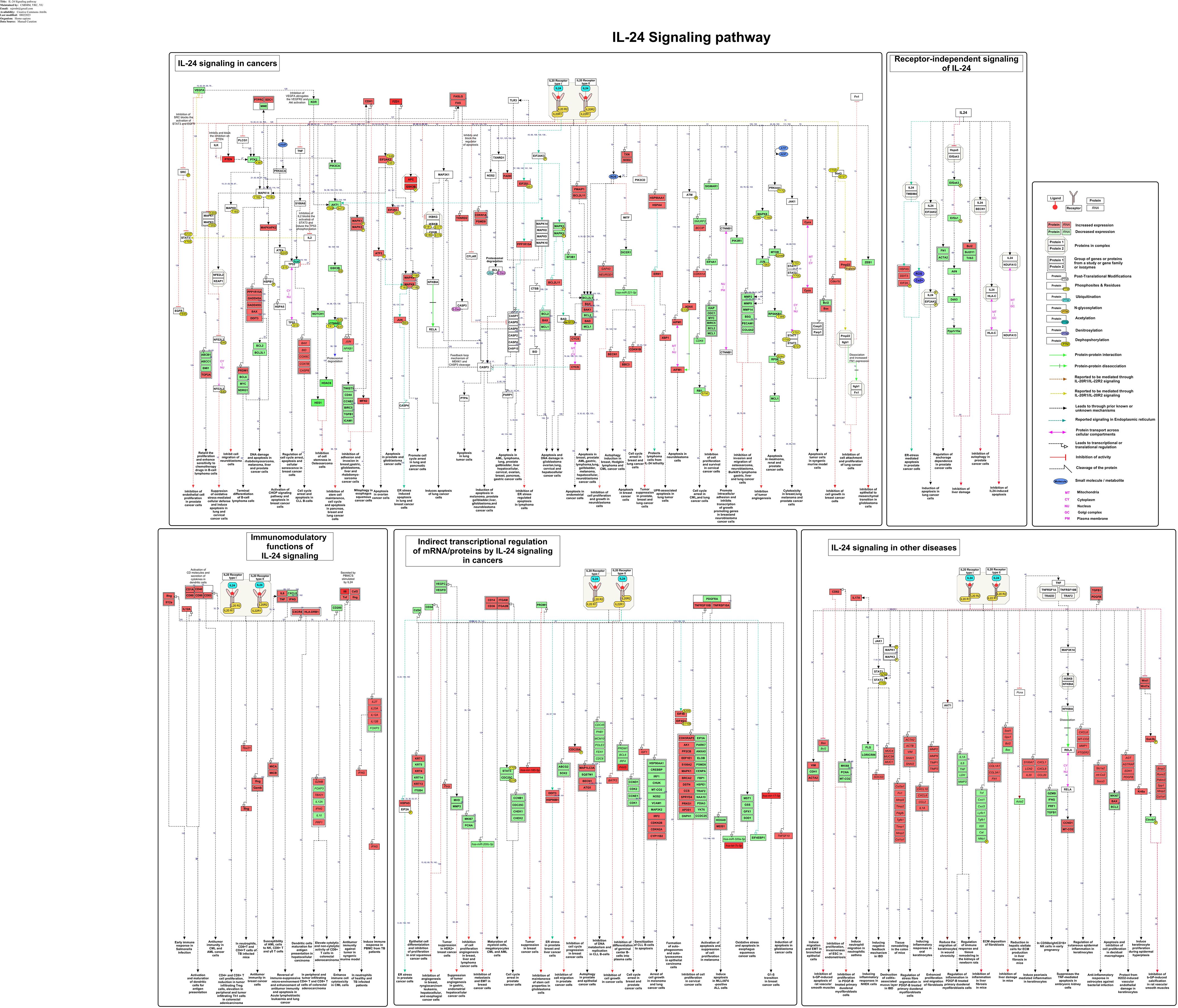
Figure 2. The schematic representation of IL-24-induced signaling reactions. The signaling pathway map shows molecules associated with ligand-receptor interactions and IL-24 downstream molecular events. The pathway comprises the signaling modules such as activation/inhibition, enzyme catalysis, protein/gene regulatory events, molecular association, and translocation events induced by IL-24 toward its anti-tumor and immunoregulatory properties. These signaling events are represented with a specific type of edge and are color-coded as mentioned in the pathway legend. The pathway also includes information about the site and residues of post-translational modifications. The functions/processes linked to each signaling event, as annotated from the curated study, are displayed at the bottom of the pathway.
3.1 Deciphering the role of IL-24 as a tumor suppressor cytokine
The distinctive functions of IL-24 in cancer include tumor-specific killing through an integrated effect of apoptosis, leading to a potent “bystander” anti-cancer activity, inhibition of cell proliferation, migration, and angiogenesis (18). This cytotoxic activity could cause either growth arrest or apoptosis through various cellular mechanisms solely in cancer cells at the supraphysiological levels of IL-24. Meanwhile, IL-24, at its natural physiological level, is recognized to function regularly to maintain its cellular processes and immune responses. A potential explanation for this selectively induced apoptosis is the intrinsic biochemical distinctions between normal and cancer cells. This encompasses factors such as high ROS production, altered metabolism, reprogrammed signaling pathways, and oxidative and endoplasmic reticulum (ER) stress (51). The bystander activity of IL-24 induces anti-cancer effects on IL-24 target cells and influences the neighboring untreated cells. This intriguing property makes IL-24 a promising candidate for cancer therapy as it amplifies its therapeutic potential beyond the directly treated cells. The IL-24 expression in tumor cells is primarily derived from tumor-infiltrating immune cells, such as T cells and NK cells, rather than from the tumor cells themselves. Moreover, the cells transduced with IL-24 can secrete other cytokines acting on the adjacent tumor cells, exerting their apoptotic and anti-proliferative effects (11). The tumor suppressive activity of IL-24 has been widely studied in a broad range of cancers, including oral, breast, lung, cervical, and colorectal carcinoma (22, 52–55).
3.1.1 Induction of apoptosis through IL-24 signaling axis
IL-24 modulates diverse apoptosis-related signaling pathways through the regulation of multiple apoptosis-effector proteins in various cancers. It promotes apoptosis through the extrinsic pathway, mediated by death receptors (56, 57). It also invokes the intrinsic pathway, driven by the expression of Ataxia Telangiectasia Mutated protein (ATM) and TP53. This intricate network stimulates pro-apoptotic and inhibits anti-apoptotic molecules, culminating in the activation of caspases, orchestrating the subsequent cell death across diverse cancer types (58–60). Additionally, it initiates apoptosis through oxidative and ER stress by triggering the release of ROS, activation of PKR (EIF2AK2) and multiple MAPKs, followed by the activation of ER stress markers, DNA damage-inducing proteins, and BiP (HSPA4) (51, 56, 61–63). Despite the activation of apoptosis, IL-24 is also known to induce autophagy-associated cell death by the regulation of miR-221 and Beclin 1 (BECN1) axis and direct activation of autophagy-related gene 5 (ATG5) (64, 65). Besides, IL-24 is well-known for the phosphorylation, activation, and nuclear translocation of the tumor suppressor protein TP53, resulting in cell proliferation arrest (66). Altogether, these combined actions culminate in the initiation of apoptosis within several cancer types.
3.1.2 Diverse mechanisms of IL-24-mediated anti-tumor activity
IL-24 exhibits potent anti-tumor activity, including anti-angiogenesis by suppressing endothelial cell markers such as vascular endothelial growth factors and CD34. It induces cell cycle arrest (G2-M phase) by modulating cell cycle proteins such as CDKs, cyclins, and cell division cycle proteins. Further, it inhibits cancer cell adhesion and invasion by downregulating matrix metalloproteinases (MMPs) and adhesion molecules (67–70). IL-24 has also proven to be an anti-proliferative molecule, substantiated by its ability to reduce the expression of proliferation markers, Marker of Proliferation Ki-67 (MKI67), and Proliferating Cell Nuclear Antigen (PCNA) (71, 72). In addition, IL-24 has a significant role in impeding cancer stem cell maintenance and epithelial-to-mesenchymal transition (EMT), reversing drug resistance, and inducing cellular senescence in cancer cells. Additionally, it involves the inhibition of both Notch and the Wnt/β-catenin signaling pathways with the subsequent proteasomal degradation of β-catenin (CTNNB1), and the ubiquitination and degradation of BCL2 (52, 71, 73–78).
3.2 Synergistic activities of IL-24
Beyond the intrinsic property of IL-24 as a cytotoxic and tumor-suppressant agent, numerous studies have investigated the utility of IL-24 with other drugs as a combinatorial drug administration approach in various diseases (Figure 3). Research findings have demonstrated that combining multiple chemotherapeutic agents and radiation with IL-24 in cancer treatment has amplified the effectiveness of chemotherapy and radiotherapy in numerous cancers (79–81). Also, studies have shown that IL-24 promotes apoptosis in various cancer types, boosting its anticancer activity when used in conjunction with some anti-inflammatory drugs, pleiotropic cytokines, including IL-20, IFNG, IL-6, vitamin E, gamma-secretase, and other small-molecule inhibitors (82–84).
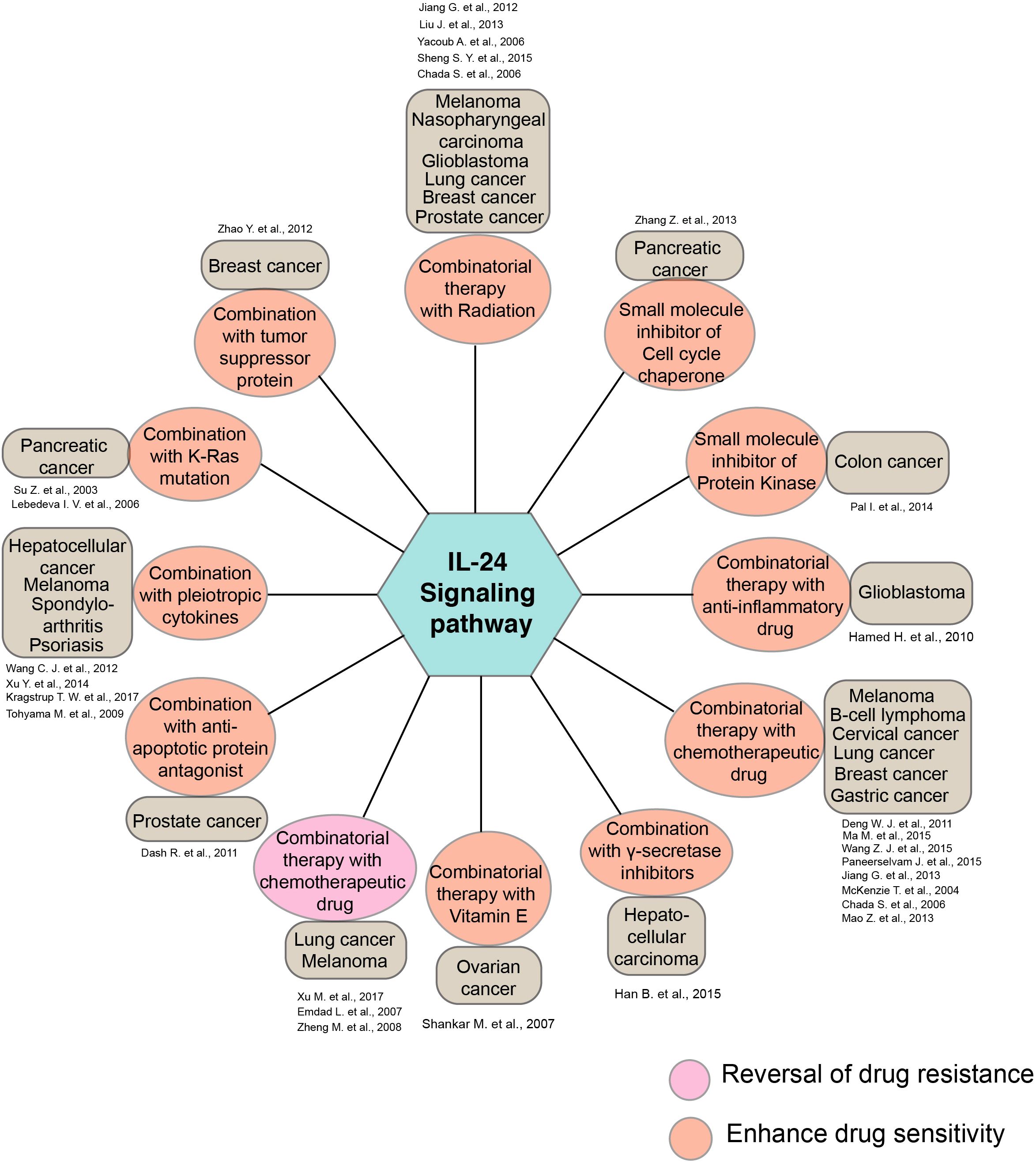
Figure 3. Diagrammatic illustration of the synergistic activity of IL-24 along with the various drugs and therapeutic agents. The diagram summarizes the synergistic action of IL-24 and other pharmacological agents in various disease conditions. Key drugs and the cancer conditions indicated in the study are represented along with the amplified therapeutic responses.
3.3 IL-24 signaling in inflammation and infectious diseases
IL-24 can be a potent therapeutic agent against viral and bacterial infections like influenza, Human immunodeficiency virus (HIV), Mycobacterium, and Salmonella (85). As well, in inflammatory conditions such as endometriosis, inflammatory bowel diseases (IBD), asthma, and psoriasis, IL-24 has been observed to modulate inflammation-associated events (86–90). Higher expression levels of IL-24 have always been associated with chronic inflammation and autoimmune disorders such as psoriasis, arthritis, vascular permeability, and IBD (91). In chronic inflammatory conditions such as asthma, IL-24 contributes to inflammation by promoting EMT of alveolar cells and neutrophils (89, 90). While in pathogenic infections, IL-24 induces an immune response through the secretion of cytokines such as IL-12, IL-23, IL-27, and IFNG (20, 92). IL-24 induces cytokine secretion by the activation of SOCS proteins through the JAK-STAT pathway. The SOCS proteins are known to play an important role in IL-24 signaling cascades for various inflammatory and infectious conditions (14).
The role of IL-24 in inflammatory diseases is context-specific, where it has been implicated in promoting inflammation in certain pathophysiological conditions. Similar to the case of cancer, IL-24 functions to reduce the proliferation and invasiveness of endometrial squamous cells (ESCs) in endometriosis, marked by the downregulation of MKI67 and PCNA. In addition, IL-24 also suppresses the macrophage-induced high viability and invasiveness of ESCs (88). Whereas, in IBD, recent studies have found that peripheral blood mononuclear cells (PBMCs) and epithelial cells secrete higher amounts of IL-24, resulting in the activation of extracellular matrix (ECM) and mucin proteins (collagens, fibronectins) contributing to the remodeling of the mucous layer in IBD (93, 94). In other inflammatory conditions, such as psoriasis, IL-24 is highly stimulated and induces the expression of the inflammatory mediators, including Psoriasin, chemokine ligands (87).
Furthermore, studies have proven that IL-24 can act as a protective agent and reverse the molecular events caused by the injuries induced by specific agents, such as H2O2 and β-glycerophosphate (95, 96). Similarly, in liver fibrosis, IL-24 functions to protect the liver from inflammation and injury. IL-24 protects the liver cells through the activation of anti-oxidant proteins and the reduction of ER stress-related DNA-damaging proteins, pro-apoptotic and pro-inflammatory proteins (97, 98). Also, its potential as a therapeutic agent in central nervous system autoimmunity has been recently investigated, revealing that IL-24 suppresses ocular inflammation by markedly inhibiting the Th1 and Th17 cells differentiation and cytokine secretion from the ocular infiltrating pathogenic Th1 and Th17 cells (99). In inflammatory conditions, IL-24 plays a dual role in Th17 cells by promoting mitochondrial recruitment of STAT3, enhancing oxidative phosphorylation, and dampening the nuclear transcriptional activity of STAT3, thereby regulating the IL-10 expression and restraining the Th17-mediated immunopathology (100). Additionally, IL-24 also acts as a downstream regulator in the IL-17A-mediated signaling loop within Th17 cells, controlling the expression of other Th17-associated cytokines such as IL-17F and GM-CSF (101). Thereby, IL-24 effectively modulates the balance between inflammation and regulation in autoimmune diseases by orchestrating Th17 cell function and cytokine expression. Therefore, the intricate involvement of IL-24 in modulating immune responses and inflammatory processes unravels its potential as a therapeutic target in a range of inflammatory conditions.
3.4 IL-24 as an immunoregulatory cytokine
IL-24 is widely recognized as an immunomodulatory cytokine with the property to enhance the tumor-specific immune response and disrupt the tolerance toward tumor antigens (102). Recent reports indicate that IL-24 can augment CD4+ and CD8+ T cell populations and reduce T regulatory cells within tumor-infiltrating lymphocytes (TILs), suggesting IL-24-mediated anti-tumor immune responses in colorectal adenocarcinoma (22). Also, Ma et al. (2016) have reported IL-24 mediates tumor suppression through increased percentages of CD45+CD4+ and CD45+CD8+ cells and decreased percentage of CD45+CD4+Foxp3+ cells in IL-24-treated colon cancer (102). The enhanced cytolytic activity of CD8+ T cells was marked by the IFNG, granzymes, and perforins secretion stimulated by IL-24 (22). Moreover, IL-24 markedly enhanced the mRNA expression of the transcription factor T-bet (Th1 transcriptional factor) to activate CD4+ T cells. It also inhibits the expression of a T regulatory cell activation marker, transcriptional factor Forkhead Box P3 (FOXP3), ultimately reducing the T regulatory cells in colorectal carcinoma (22, 102). Alongside, it also activates and enhances the functions of the cluster of differentiation 80 and 86 (CD80, CD86) proteins that are known for the transduction of secondary signals for T-cell activation. Furthermore, it also stimulates innate and adaptive immune responses in myelogenous leukemia cells through the production of ligands (MICA and MICB) for Killer Cell Lectin Like Receptor K1 (KLRK1) on NK and T cells (103).
Besides its intrinsic immunoregulatory functions, IL-24 can induce the secretion of other cytokines and modulate its activity in diverse pathological conditions (104, 105). IL-24 stimulates the secretion of IL-6, IL-12, IL-27, and IFNG in cancer and infectious disease conditions. Thereby, IL-24 plays a significant role in regulating the immune response against tumorigenesis by reversing the immunosuppressive tumor microenvironment and promoting anti-tumor immunity (103). Additionally, in infectious disease conditions, it stimulates the immune response through the recruitment of CD4+, CD8+ T cells, and neutrophils by the activation of T-Cell-Specific T-Box Transcription Factor (TBX21), leading to the secretion of IFNG and several other interleukins (106). This orchestrated immune activation by IL-24 plays a crucial role in combating pathogens, external antigens, and tumor cells, thereby promoting the host-defense mechanisms.
3.5 Therapeutic applications of IL-24
The demonstration and visualization of the mechanisms of IL-24-mediated immunoregulation facilitate and aid in developing and utilizing IL-24 therapeutic approaches. Recent reports have proven the efficacy of IL-24 in enhancing CAR-T cell therapy against cancer stem cells (CSCs) that exhibit therapeutic resistance (25). The study by Hu et al. (2021) has reported that IL-24 potentially enhances the vitality of T cells, which is a common cell resource for CAR-T products, and significantly inhibits the viability and proliferation of cancer cells in leukemia and lymphoma (26). In addition, it was found that IL24-potentiated T-cell expansion and increased tumor infiltration significantly inhibited the cancer progression. Therefore, arming T cells with IL-24 shows the capabilities of tumor eradication and T-cell expansion within the tumor microenvironment. This highlights a promising approach of cellular immunotherapy that inhibits tumor escape (27). Moreover, through the annotation of IL-24 signaling mechanisms, we identified that IL-24 has the potency to significantly suppress the expression and activity of the T regulatory cell transcription factor, FOXP3, in cancer cells and infectious disease conditions. This transcription factor is well known for activating T regulatory cells that impair the treatment response in cancer immunotherapy (107). Extensive research activities are carried out to suppress this transcription factor as a cancer therapeutic approach (108). However, the ability of IL-24 to specifically target and suppress the function of FOXP3 remains inadequately investigated. Therefore, with this signaling pathway map, we were able to showcase all possible signaling mechanisms, primarily in cancer as well as in other inflammatory and infectious diseases. IL-24 stands out as a potent therapeutic agent, with supporting evidence highlighting its potential in developing advanced and personalized cancer therapeutics.
3.6 Applications of the IL-24 signaling pathway map to gene expression analyses
The current pathway model serves as a tool that integrates molecular-level information derived from individual studies and does not represent a temporally orchestrated network within a single cell; rather, it represents standalone findings from each study. This applies to all pathway models available to date and is further augmented by the limitations in their effective visualization (37, 43, 109). In a way, most maps are static, considering the signaling network highly spatiotemporal and dynamic. However, the pathway model represents the collective information on the molecules and their reactions that are identified to be perturbed upon IL-24 stimulation into a computationally encoded form for incorporation into enrichment tools. These details, which are otherwise scattered across the literature, would be time-consuming for individual researchers to compile into pathway models. Additionally, the present study does not provide new in vivo or clinical validation. However, all the signaling events incorporated into the pathway were curated from experimentally validated studies, reflecting the available literature up to the time of publication. Even though challenged by the above limitations, the current model serves as a reference platform and database to update molecular data as more studies on IL-24 become available. We also aim to refer to notable studies in the map to ensure transparency and to support future updates as more data becomes available.
Furthermore, to demonstrate the utility of the annotated signaling pathway map serving as a framework for applying gene set enrichment analysis, we mapped it into the protein-protein interaction network comprising IL-24, its receptors, and downstream targets. The analysis highlighted the various hub genes, including key signaling modules such as JAK-STAT, SOCS, PI3K, PTK2, and MAPK, together with apoptosis and cell cycle regulator proteins (Figure 4). This emphasizes the role of the curated signaling map in the detection of enriched signaling modules and the selection of candidate molecules for further analysis. Additionally, we looked into the publicly available transcriptomic profiles of peripheral blood mononuclear cells in cancer and infectious disease conditions. We identified three transcriptomic datasets where IL-24 expression was detected (110–112). Notably, IL-24 was consistently downregulated in cancer and inflammatory conditions, together with the suppression of anti-tumor cytokine genes within the immunosuppressive microenvironment. Whereas, in the infectious disease condition of tuberculosis, IL-24 was found to be overexpressed, indicating its anti-infectious property. This reinforces the immunomodulatory role of IL-24 and represents the utility of this curated pathway map to elucidate disease-gene association and its expression changes for further functional analysis.

Figure 4. Protein-protein interaction network analysis. The figure represents the interaction network analysis of IL-24, its receptors, and downstream targets. The proteins are color-coded according to five distinct clusters based on the hub genes analysis. The hub genes are highlighted within each cluster, and the blue colored nodes depict the proteins that interact directly or indirectly with these hub genes.
Further, to complement the signaling pathway, we performed a pathway enrichment analysis with the differentially expressed proteins identified from the publicly available proteomic dataset of IL-24-treated cervical cancer cells (113). The analysis revealed significantly enriched pathways, including apoptosis, immunomodulatory pathways, and cell cycle regulations (Supplementary Figure S1). These enriched pathways align with the key signaling modules annotated in the pathway map of IL-24, further supporting the role of IL-24 in tumor suppression and immunoregulation. This integration demonstrates the utility of the application of this signaling pathway map in gene set enrichment analysis pipelines, offering a valuable resource for future studies in the analysis of high-throughput data.
4 Conclusions
A majority of studies have focused on the role of IL-24 in the pathophysiology of diseases, as this multi-functional cytokine serves as both a tumor suppressant and an agent with pro- and anti-inflammatory properties. IL-24 is very well known for its tumor-suppressive activity and is utilized as a therapeutic agent in cytokine therapy against a broad spectrum of cancers. The signaling pathway map of IL-24 provides a detailed illustration of its downstream molecular targets and their signaling mechanisms. Hence, this pathway map would accelerate clinical applications of IL-24 as a therapeutic target for various disorders, to its potential as a prominent therapeutic agent in cancer, and to reshape the tumor microenvironment.
The signaling pathway map developed in this study provides a framework to visualize the molecules and their reactions reported to be induced by IL-24 in various disease conditions. This comprehensive signaling pathway can help identify gaps in signaling events and investigate further molecular reactions, including dynamic protein complexes associated with IL-24 signaling across various pathophysiological conditions. In the current scenario, the involvement of IL-24 and its signaling components in various clinical/experimental global omics datasets is not ensured through gene set enrichment analysis approaches. This is primarily due to the lack of availability of its pathway model in the existing enrichment tools. Further enrichment of datasets derived from high-throughput approaches such as single-cell transcriptomics and proteomics may provide deeper insights into the role of IL-24 in health and disease, leading to an expansion of the pathway.
To enable its utility, the IL-24 map/model is made freely available in.gpml format for their conversion into other international standard exchange formats such as BioPAX, PSI-MI, and KEGGML for their incorporation into different enrichment analysis tools. To acquire its full potential, IL-24-mediated effects in diverse preclinical models should be validated for their efficacy across different biological conditions or engaged in studies toward patient stratification based on response to IL-24-targeted interventions. The combinatorial effects of IL-24 with other cytokines or their regulation under various disease conditions also demand further investigation. This model can be customized or enhanced by the users to engage them in the analysis of omics datasets by individual researchers. The IL-24 pathway model will be made available through the web portal (https://ciods.in/il24/) to keep the pathway updated and provide it to the users.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author contributions
AA: Data curation, Formal Analysis, Investigation, Methodology, Visualization, Writing – original draft. VS: Data curation, Investigation, Writing – original draft. MA: Funding acquisition, Project administration, Writing – review & editing. SD: Methodology, Writing – review & editing. MV: Conceptualization, Project administration, Resources, Supervision, Writing – review & editing. TP: Project administration, Resources, Supervision, Writing – review & editing. RR: Conceptualization, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research of this article. The authors acknowledge the Ongoing Research Funding Program, (ORF-2025-984), King Saud University, Riyadh, Saudi Arabia for the partial support for this study.
Acknowledgments
The authors acknowledge the support of the Department of Biotechnology, Government of India, to the Yenepoya (Deemed to be University) through the project on “Skill Development in Mass Spectrometry-based metabolomics technology BIC” (BT/PR40202/BTIS/137/53/2023). The authors thank Karnataka Biotechnology and Information Technology Services (KBITS), Government of Karnataka, India, for the support to the Center for Systems Biology and Molecular Medicine at Yenepoya (Deemed to be University) under the Biotechnology Skill Enhancement Programme in Multiomics Technology (BiSEP GO ITD 02 MDA 2017). Anjana Aravind is a recipient of the Senior Research Fellowship (08/652 (0002)/2019-EMR-1) from the Council of Scientific & Industrial Research (CSIR), Government of India.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1608101/full#supplementary-material
Supplementary Figure 1 | Pathway enrichment analysis of the differentially expressed proteins from IL-24-treated cancer cells. The diagram represents the enriched pathways and their associated proteins from the differentially expressed proteins identified from a publicly available dataset of IL-24-treated cervical cancer cells.
Supplementary Data Sheet 1 | A compendium of IL-24-mediated signaling events. The table provides the signaling events, the molecular events including activation/inhibition events, gene/protein regulations, enzyme/substrate reactions, protein-protein interaction, translocation events, and associated functional outcomes, together with the disease condition and experimental details. The data also provides the signaling events linked with the various pharmacological modulations of IL-24.
References
1. Caudell EG, Mumm JB, Poindexter N, Ekmekcioglu S, Mhashilkar AM, Yang XH, et al. The protein product of the tumor suppressor gene, melanoma differentiation-associated gene 7, exhibits immunostimulatory activity and is designated IL-24. J Immunol. (2002) 168:6041–6. doi: 10.4049/jimmunol.168.12.6041
2. Fisher PB. Is mda-7/IL-24 a “magic bullet” for cancer? Cancer Res. (2005) 65:10128–38. doi: 10.1158/0008-5472.CAN-05-3127
3. Zhong Y, Zhang X, and Chong W. Interleukin-24 immunobiology and its roles in inflammatory diseases. Int J Mol Sci. (2022) 23:627. doi: 10.3390/ijms23020627
4. Jiang H, Lin JJ, Su ZZ, Goldstein NI, and Fisher PB. Subtraction hybridization identifies a novel melanoma differentiation associated gene, mda-7, modulated during human melanoma differentiation, growth and progression. Oncogene. (1995) 11:2477–86.
5. Dumoutier L, Leemans C, Lejeune D, Kotenko SV, and Renauld JC. Cutting edge: STAT activation by IL-19, IL-20 and mda-7 through IL-20 receptor complexes of two types. J Immunol. (2001) 167:3545–9. doi: 10.4049/jimmunol.167.7.3545
6. Wang M, Tan Z, Zhang R, Kotenko SV, and Liang P. Interleukin 24 (MDA-7/MOB-5) signals through two heterodimeric receptors, IL-22R1/IL-20R2 and IL-20R1/IL-20R2. J Biol Chem. (2002) 277:7341–7. doi: 10.1074/jbc.M106043200
7. Parrish-Novak J, Xu W, Brender T, Yao L, Jones C, West J, et al. Interleukins 19, 20, and 24 signal through two distinct receptor complexes. Differences in receptor-ligand interactions mediate unique biological functions. J Biol Chem. (2002) 277:47517–23. doi: 10.1074/jbc.M205114200
8. Pestka S, Krause CD, Sarkar D, Walter MR, Shi Y, and Fisher PB. Interleukin-10 and related cytokines and receptors. Annu Rev Immunol. (2004) 22:929–79. doi: 10.1146/annurev.immunol.22.012703.104622
9. Wang M and Liang P. Interleukin-24 and its receptors. Immunology. (2005) 114:166–70. doi: 10.1111/j.1365-2567.2005.02094.x
10. Menezes ME, Bhoopathi P, Pradhan AK, Emdad L, Das SK, Guo C, et al. Role of MDA-7/IL-24 a multifunction protein in human diseases. Adv Cancer Res. (2018) 138:143–82. doi: 10.1016/bs.acr.2018.02.005
11. Modi J, Roy A, Pradhan AK, Kumar A, Talukdar S, Bhoopathi P, et al. Insights into the mechanisms of action of MDA-7/IL-24: A ubiquitous cancer-suppressing protein. Int J Mol Sci. (2021) 23:72. doi: 10.3390/ijms23010072
12. Huang EY, Madireddi MT, Gopalkrishnan RV, Leszczyniecka M, Su Z, Lebedeva IV, et al. Genomic structure, chromosomal localization and expression profile of a novel melanoma differentiation associated (mda-7) gene with cancer specific growth suppressing and apoptosis inducing properties. Oncogene. (2001) 20:7051–63. doi: 10.1038/sj.onc.1204897
13. Sarkar D, Su ZZ, Lebedeva IV, Sauane M, Gopalkrishnan RV, Dent P, et al. mda-7 (IL-24): signaling and functional roles. Biotechniques. (2002) Suppl:30–9. doi: 10.2144/Oct0204
14. Smith S, Lopez S, Kim A, Kasteri J, Olumuyide E, Punu K, et al. Interleukin 24: signal transduction pathways. Cancers (Basel). (2023) 15:3365. doi: 10.3390/cancers15133365
15. Logsdon NJ, Deshpande A, Harris BD, Rajashankar KR, and Walter MR. Structural basis for receptor sharing and activation by interleukin-20 receptor-2 (IL-20R2) binding cytokines. Proc Natl Acad Sci U S A. (2012) 109:12704–9. doi: 10.1073/pnas.1117551109
16. Sieger KA, Mhashilkar AM, Stewart A, Sutton RB, Strube RW, Chen SY, et al. The tumor suppressor activity of MDA-7/IL-24 is mediated by intracellular protein expression in NSCLC cells. Mol Ther. (2004) 9:355–67. doi: 10.1016/j.ymthe.2003.11.014
17. Sauane M, Lebedeva IV, Su ZZ, Choo HT, Randolph A, Valerie K, et al. Melanoma differentiation associated gene-7/interleukin-24 promotes tumor cell-specific apoptosis through both secretory and nonsecretory pathways. Cancer Res. (2004) 64:2988–93. doi: 10.1158/0008-5472.CAN-04-0200
18. Menezes ME, Bhatia S, Bhoopathi P, Das SK, Emdad L, Dasgupta S, et al. MDA-7/IL-24: multifunctional cancer killing cytokine. Adv Exp Med Biol. (2014) 818:127–53. doi: 10.1007/978‑1‑4471‑6458‑6_6
19. Gupta P, Su ZZ, Lebedeva IV, Sarkar D, Sauane M, Emdad L, et al. mda-7/IL-24: multifunctional cancer-specific apoptosis-inducing cytokine. Pharmacol Ther. (2006) 111:596–628. doi: 10.1016/j.pharmthera.2005.11.005
20. Ma Y, Chen H, Wang Q, Luo F, Yan J, and Zhang XL. IL-24 protects against Salmonella typhimurium infection by stimulating early neutrophil Th1 cytokine production, which in turn activates CD8+ T cells. Eur J Immunol. (2009) 39:3357–68. doi: 10.1002/eji.200939678
21. Gopalkrishnan RV, Sauane M, and Fisher PB. Cytokine and tumor cell apoptosis inducing activity of mda-7/IL-24. Int Immunopharmacol. (2004) 4:635–47. doi: 10.1016/j.intimp.2004.01.015
22. Zhang Y, Liu Y, and Xu Y. Interleukin-24 regulates T cell activity in patients with colorectal adenocarcinoma. Front Oncol. (2019) 9:1401. doi: 10.3389/fonc.2019.01401
23. Anuradha R, George PJ, Hanna LE, Kumaran P, Chandrasekaran V, Nutman TB, et al. Expansion of parasite-specific CD4+ and CD8+ T cells expressing IL-10 superfamily cytokine members and their regulation in human lymphatic filariasis. PloS Negl Trop Dis. (2014) 8:e2762. doi: 10.1371/journal.pntd.0002762
24. Persaud L, De Jesus D, Brannigan O, Richiez-Paredes M, Huaman J, Alvarado G, et al. Mechanism of action and applications of interleukin 24 in immunotherapy. Int J Mol Sci. (2016) 17:869. doi: 10.3390/ijms17060869
25. Zhang K, Hu W, Li F, Wen C, Zhou L, Zhang L, et al. IL-24 improves efficacy of CAR-T cell therapy by targeting stemness of tumor cells. Br J Cancer. (2024) 130:1337–47. doi: 10.1038/s41416-024-02601-1
26. Hu Q, Zhang Y, Wang P, Zhou M, Hu Z, Liu C, et al. IL-24 armored CAR19-T cells show enhanced antitumor activity and persistence. Signal Transduct Target Ther. (2021) 6:14. doi: 10.1038/s41392-020-00380-8
27. Liu Z, Guo C, Das SK, Yu X, Pradhan AK, Li X, et al. Engineering T cells to express tumoricidal MDA-7/IL24 enhances cancer immunotherapy. Cancer Res. (2021) 81:2429–41. doi: 10.1158/0008-5472.CAN-20-2604
28. Ramesh R, Ioannides CG, Roth JA, and Chada S. Adenovirus-mediated interleukin (IL)-24 immunotherapy for cancer. Methods Mol Biol. (2010) 651:241–70. doi: 10.1007/978-1-60761-786-0_14
29. Dent P, Yacoub A, Hamed HA, Park MA, Dash R, Bhutia SK, et al. The development of MDA-7/IL-24 as a cancer therapeutic. Pharmacol Ther. (2010) 128:375–84. doi: 10.1016/j.pharmthera.2010.08.001
30. Fisher PB, Gopalkrishnan RV, Chada S, Ramesh R, Grimm EA, Rosenfeld MR, et al. mda-7/IL-24, a novel cancer selective apoptosis inducing cytokine gene: from the laboratory into the clinic. Cancer Biol Ther. (2003) 2:S23–37. doi: 10.4161/cbt.458
31. Lebedeva IV, Sauane M, Gopalkrishnan RV, Sarkar D, Su ZZ, Gupta P, et al. mda-7/IL-24: exploiting cancer’s Achilles’ heel. Mol Ther. (2005) 11:4–18. doi: 10.1016/j.ymthe.2004.08.012
32. Cunningham CC, Chada S, Merritt JA, Tong A, Senzer N, Zhang Y, et al. Clinical and local biological effects of an intratumoral injection of mda-7 (IL24; INGN 241) in patients with advanced carcinoma: a phase I study. Mol Ther. (2005) 11:149–59. doi: 10.1016/j.ymthe.2004.09.019
33. Tong AW, Nemunaitis J, Su D, Zhang Y, Cunningham C, Senzer N, et al. Intratumoral injection of INGN 241, a nonreplicating adenovector expressing the melanoma-differentiation associated gene-7 (mda-7/IL24): biologic outcome in advanced cancer patients. Mol Ther. (2005) 11:160–72. doi: 10.1016/j.ymthe.2004.09.021
34. Lebedeva IV, Emdad L, Su ZZ, Gupta P, Sauane M, Sarkar D, et al. mda-7/IL-24, novel anticancer cytokine: focus on bystander antitumor, radiosensitization and antiangiogenic properties and overview of the phase I clinical experience (Review). Int J Oncol. (2007) 31:985–1007. doi: 10.3892/ijo.31.5.985
35. Inoue S, Shanker M, Miyahara R, Gopalan B, Patel S, Oida Y, et al. MDA-7/IL-24-based cancer gene therapy: translation from the laboratory to the clinic. Curr Gene Ther. (2006) 6:73–91. doi: 10.2174/156652306775515574
36. Fisher PB, Sarkar D, Lebedeva IV, Emdad L, Gupta P, Sauane M, et al. Melanoma differentiation associated gene-7/interleukin-24 (mda-7/IL-24): novel gene therapeutic for metastatic melanoma. Toxicol Appl Pharmacol. (2007) 224:300–7. doi: 10.1016/j.taap.2006.11.021
37. Aravind A, Palollathil A, Rex DAB, Kumar KMK, Vijayakumar M, Shetty R, et al. A multi-cellular molecular signaling and functional network map of C-C motif chemokine ligand 18 (CCL18): a chemokine with immunosuppressive and pro-tumor functions. J Cell Commun Signal. (2022) 16:293–300. doi: 10.1007/s12079-021-00633-3
38. Balakrishnan L, Soman S, Patil YB, Advani J, Thomas JK, Desai DV, et al. IL-11/IL11RA receptor mediated signaling: a web accessible knowledgebase. Cell Commun Adhes. (2013) 20:81–6. doi: 10.3109/15419061.2013.791683
39. Sharma J, Balakrishnan L, Datta KK, Sahasrabuddhe NA, Khan AA, Sahu A, et al. A knowledgebase resource for interleukin-17 family mediated signaling. J Cell Commun Signal. (2015) 9:291–6. doi: 10.1007/s12079-015-0297-3
40. Raju R, Gadakh S, Gopal P, George B, Advani J, Soman S, et al. Differential ligand-signaling network of CCL19/CCL21-CCR7 system. Database (Oxford). (2015) 2015:bav106. doi: 10.1093/database/bav106
41. Shaji V, Dagamajalu S, Sanjeev D, George M, Kanekar S, Prasad G, et al. Deciphering the receptor-mediated signaling pathways of interleukin-19 and interleukin-20. J Interferon Cytokine Res. (2024) 44(9):388–98.
42. Kandasamy K, Keerthikumar S, Raju R, Keshava Prasad TS, Ramachandra YL, Mohan S, et al. PathBuilder–open source software for annotating and developing pathway resources. Bioinformatics. (2009) 25:2860–2. doi: 10.1093/bioinformatics/btp453
43. Kandasamy K, Mohan SS, Raju R, Keerthikumar S, Kumar GS, Venugopal AK, et al. NetPath: a public resource of curated signal transduction pathways. Genome Biol. (2010) 11:R3. doi: 10.1186/gb-2010-11-1-r3
44. Kutmon M, van Iersel MP, Bohler A, Kelder T, Nunes N, Pico AR, et al. PathVisio 3: an extendable pathway analysis toolbox. PloS Comput Biol. (2015) 11:e1004085. doi: 10.1371/journal.pcbi.1004085
45. Szklarczyk D, Kirsch R, Koutrouli M, Nastou K, Mehryary F, Hachilif R, et al. The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. (2023) 51:D638–D46. doi: 10.1093/nar/gkac1000
46. Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, et al. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. (2003) 13:2498–504. doi: 10.1101/gr.1239303
47. Bader GD and Hogue CW. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinf. (2003) 4:2. doi: 10.1186/1471-2105-4-2
48. Milacic M, Beavers D, Conley P, Gong C, Gillespie M, Griss J, et al. The reactome pathway knowledgebase 2024. Nucleic Acids Res. (2024) 52:D672–D8. doi: 10.1093/nar/gkad1025
49. Trivella DB, Ferreira-Junior JR, Dumoutier L, Renauld JC, and Polikarpov I. Structure and function of interleukin-22 and other members of the interleukin-10 family. Cell Mol Life Sci. (2010) 67:2909–35. doi: 10.1007/s00018-010-0380-0
50. Dash R, Bhutia SK, Azab B, Su ZZ, Quinn BA, Kegelmen TP, et al. mda-7/IL-24: a unique member of the IL-10 gene family promoting cancer-targeted toxicity. Cytokine Growth Factor Rev. (2010) 21:381–91. doi: 10.1016/j.cytogfr.2010.08.004
51. Do W, Herrera C, Mighty J, Shumskaya M, Redenti SM, and Sauane M. Sigma 1 Receptor plays a prominent role in IL-24-induced cancer-specific apoptosis. Biochem Biophys Res Commun. (2013) 439:215–20. doi: 10.1016/j.bbrc.2013.08.057
52. Persaud L, Mighty J, Zhong X, Francis A, Mendez M, Muharam H, et al. IL-24 Promotes Apoptosis through cAMP-Dependent PKA Pathways in Human Breast Cancer Cells. Int J Mol Sci. (2018) 19:3561. doi: 10.3390/ijms19113561
53. Oida Y, Gopalan B, Miyahara R, Branch CD, Chiao P, Chada S, et al. Inhibition of nuclear factor-kappaB augments antitumor activity of adenovirus-mediated melanoma differentiation-associated gene-7 against lung cancer cells via mitogen-activated protein kinase kinase kinase 1 activation. Mol Cancer Ther. (2007) 6:1440–9. doi: 10.1158/1535-7163.MCT-06-0374
54. Wang L, Feng Z, Wu H, Zhang S, Pu Y, Bian H, et al. Melanoma differentiation-associated gene-7/interleukin-24 as a potential prognostic biomarker and second primary Malignancy indicator in head and neck squamous cell carcinoma patients. Tumour Biol. (2014) 35:10977–85. doi: 10.1007/s13277-014-2392-0
55. Tian H, Zhang D, Gao Z, Li H, Zhang B, Zhang Q, et al. MDA-7/IL-24 inhibits Nrf2-mediated antioxidant response through activation of p38 pathway and inhibition of ERK pathway involved in cancer cell apoptosis. Cancer Gene Ther. (2014) 21:416–26. doi: 10.1038/cgt.2014.45
56. Gopalan B, Litvak A, Sharma S, Mhashilkar AM, Chada S, and Ramesh R. Activation of the Fas-FasL signaling pathway by MDA-7/IL-24 kills human ovarian cancer cells. Cancer Res. (2005) 65:3017–24. doi: 10.1158/0008-5472.CAN-04-3758
57. Tian H, Zhang DF, Zhang BF, Li HZ, Zhang Q, Li LT, et al. Melanoma differentiation associated gene-7/interleukin-24 induces caspase-3 denitrosylation to facilitate the activation of cancer cell apoptosis. J Interferon Cytokine Res. (2015) 35:157–67. doi: 10.1089/jir.2014.0061
58. Pataer A, Chada S, Hunt KK, Roth JA, and Swisher SG. Adenoviral melanoma differentiation-associated gene 7 induces apoptosis in lung cancer cells through mitochondrial permeability transition-independent cytochrome c release. J Thorac Cardiovasc Surg. (2003) 125:1328–35. doi: 10.1016/S0022-5223(02)73247-9
59. Valiyari S, Salami M, Mahdian R, Shokrgozar MA, Oloomi M, Mohammadi Farsani A, et al. sIL-24 peptide, a human interleukin-24 isoform, induces mitochondrial-mediated apoptosis in human cancer cells. Cancer Chemother Pharmacol. (2017) 80:451–9. doi: 10.1007/s00280-017-3370-1
60. Zerbini LF, Czibere A, Wang Y, Correa RG, Otu H, Joseph M, et al. A novel pathway involving melanoma differentiation associated gene-7/interleukin-24 mediates nonsteroidal anti-inflammatory drug-induced apoptosis and growth arrest of cancer cells. Cancer Res. (2006) 66:11922–31. doi: 10.1158/0008-5472.CAN-06-2068
61. Pradhan AK, Bhoopathi P, Talukdar S, Scheunemann D, Sarkar D, Cavenee WK, et al. MDA-7/IL-24 regulates the miRNA processing enzyme DICER through downregulation of MITF. Proc Natl Acad Sci U S A. (2019) 116:5687–92. doi: 10.1073/pnas.1819869116
62. Rastegari M, Shiri A, Behzad-Behbahani A, Rasoolian M, Zare F, Rafiei G, et al. The evaluation of tLyP-1-bound mda-7/IL-24 killing activity on a liver tumor cell line. Cancer Biother Radiopharm. (2021) 36:827–36. doi: 10.1089/cbr.2019.3080
63. Pataer A, Hu W, Xiaolin L, Chada S, Roth JA, Hunt KK, et al. Adenoviral endoplasmic reticulum-targeted mda-7/interleukin-24 vector enhances human cancer cell killing. Mol Cancer Ther. (2008) 7:2528–35. doi: 10.1158/1535-7163.MCT-08-0083
64. Pradhan AK, Talukdar S, Bhoopathi P, Shen XN, Emdad L, Das SK, et al. mda-7/IL-24 Mediates Cancer Cell-Specific Death via Regulation of miR-221 and the Beclin-1 Axis. Cancer Res. (2017) 77:949–59. doi: 10.1158/0008-5472.CAN-16-1731
65. Bhutia SK, Das SK, Azab B, Dash R, Su ZZ, Lee SG, et al. Autophagy switches to apoptosis in prostate cancer cells infected with melanoma differentiation associated gene-7/interleukin-24 (mda-7/IL-24). Autophagy. (2011) 7:1076–7. doi: 10.4161/auto.7.9.16163
66. Saeki T, Mhashilkar A, Chada S, Branch C, Roth JA, and Ramesh R. Tumor-suppressive effects by adenovirus-mediated mda-7 gene transfer in non-small cell lung cancer cell in vitro. Gene Ther. (2000) 7:2051–7. doi: 10.1038/sj.gt.3301330
67. Inoue S, Branch CD, Gallick GE, Chada S, and Ramesh R. Inhibition of Src kinase activity by Ad-mda7 suppresses vascular endothelial growth factor expression in prostate carcinoma cells. Mol Ther. (2005) 12:707–15. doi: 10.1016/j.ymthe.2005.05.015
68. Saito Y, Miyahara R, Gopalan B, Litvak A, Inoue S, Shanker M, et al. Selective induction of cell cycle arrest and apoptosis in human prostate cancer cells through adenoviral transfer of the melanoma differentiation-associated -7 (mda-7)/interleukin-24 (IL-24) gene. Cancer Gene Ther. (2005) 12:238–47. doi: 10.1038/sj.cgt.7700780
69. Zhuo B, Shi Y, Qin H, Sun Q, Li Z, Zhang F, et al. Interleukin-24 inhibits osteosarcoma cell migration and invasion via the JNK/c-Jun signaling pathways. Oncol Lett. (2017) 13:4505–11. doi: 10.3892/ol.2017.5990
70. Huo W, Li ZM, Zhu XM, Bao YM, and An LJ. MDA-7/IL-24 suppresses tumor adhesion and invasive potential in hepatocellular carcinoma cell lines. Oncol Rep. (2013) 30:986–92. doi: 10.3892/or.2013.2507
71. Hamed HA, Yacoub A, Park MA, Eulitt PJ, Dash R, Sarkar D, et al. Inhibition of multiple protective signaling pathways and Ad.5/3 delivery enhances mda-7/IL-24 therapy of Malignant glioma. Mol Ther. (2010) 18:1130–42. doi: 10.1038/mt.2010.29
72. Li QY, Shi Y, Huang DH, Yang T, Wang JH, Yan GH, et al. Cytokine-induced killer cells combined with dendritic cells inhibited liver cancer cells. Int J Clin Exp Med. (2015) 8:5601–10.
73. Lin T, Wang D, Chen J, Zhang Z, Zhao Y, Wu Z, et al. IL-24 inhibits the Malignancy of human glioblastoma cells via destabilization of Zeb1. Biol Chem. (2021) 402:839–48. doi: 10.1515/hsz-2020-0373
74. Bhutia SK, Das SK, Azab B, Menezes ME, Dent P, Wang XY, et al. Targeting breast cancer-initiating/stem cells with melanoma differentiation-associated gene-7/interleukin-24. Int J Cancer. (2013) 133:2726–36. doi: 10.1002/ijc.28289
75. Tian H, Wang J, Zhang B, Di J, Chen F, Li H, et al. MDA-7/IL-24 induces Bcl-2 denitrosylation and ubiquitin-degradation involved in cancer cell apoptosis. PloS One. (2012) 7:e37200. doi: 10.1371/journal.pone.0037200
76. Zhuo B, Wang X, Shen Y, Li J, Li S, Li Y, et al. Interleukin-24 inhibits the phenotype and tumorigenicity of cancer stem cell in osteosarcoma via downregulation Notch and Wnt/beta-catenin signaling. J Bone Oncol. (2021) 31:100403. doi: 10.1016/j.jbo.2021.100403
77. Emdad L, Lebedeva IV, Su ZZ, Sarkar D, Dent P, Curiel DT, et al. Melanoma differentiation associated gene-7/interleukin-24 reverses multidrug resistance in human colorectal cancer cells. Mol Cancer Ther. (2007) 6:2985–94. doi: 10.1158/1535-7163.MCT-07-0399
78. Fang P, Zhang X, Gao Y, Ding CR, Cui F, and Jiao SC. Reversal effect of melanoma differentiation associated gene-7/interleukin-24 on multidrug resistance in human hepatocellular carcinoma cells. Anat Rec (Hoboken). (2012) 295:1639–46. doi: 10.1002/ar.22551
79. Panneerselvam J, Jin J, Shanker M, Lauderdale J, Bates J, Wang Q, et al. IL-24 inhibits lung cancer cell migration and invasion by disrupting the SDF-1/CXCR4 signaling axis. PloS One. (2015) 10:e0122439. doi: 10.1371/journal.pone.0122439
80. Deng WG, Kwon J, Ekmekcioglu S, Poindexter NJ, and Grimm EA. IL-24 gene transfer sensitizes melanoma cells to erlotinib through modulation of the Apaf-1 and Akt signaling pathways. Melanoma Res. (2011) 21:44–56. doi: 10.1097/CMR.0b013e3283382155
81. Liu J, Zhang Y, Sun P, Xie Y, Xiang J, and Yang J. Enhanced therapeutic efficacy of adenovirus-mediated interleukin-24 gene therapy combined with ionizing radiotherapy for nasopharyngeal carcinoma. Oncol Rep. (2013) 30:1165–74. doi: 10.3892/or.2013.2550
82. Pal I, Sarkar S, Rajput S, Dey KK, Chakraborty S, Dash R, et al. BI-69A11 enhances susceptibility of colon cancer cells to mda-7/IL-24-induced growth inhibition by targeting Akt. Br J Cancer. (2014) 111:101–11. doi: 10.1038/bjc.2014.227
83. Shanker M, Gopalan B, Patel S, Bocangel D, Chada S, and Ramesh R. Vitamin E succinate in combination with mda-7 results in enhanced human ovarian tumor cell killing through modulation of extrinsic and intrinsic apoptotic pathways. Cancer Lett. (2007) 254:217–26. doi: 10.1016/j.canlet.2007.03.004
84. Hamed HA, Yacoub A, Park MA, Eulitt P, Sarkar D, Dimitrie IP, et al. OSU-03012 enhances Ad.7-induced GBM cell killing via ER stress and autophagy and by decreasing expression of mitochondrial protective proteins. Cancer Biol Ther. (2010) 9:526–36. doi: 10.4161/cbt.9.7.11116
85. Seong RK, Choi YK, and Shin OS. MDA7/IL-24 is an anti-viral factor that inhibits influenza virus replication. J Microbiol. (2016) 54:695–700. doi: 10.1007/s12275-016-6383-2
86. Onody A, Veres-Szekely A, Pap D, Rokonay R, Szebeni B, Sziksz E, et al. Interleukin-24 regulates mucosal remodeling in inflammatory bowel diseases. J Transl Med. (2021) 19:237. doi: 10.1186/s12967-021-02890-7
87. Kumari S, Bonnet MC, Ulvmar MH, Wolk K, Karagianni N, Witte E, et al. Tumor necrosis factor receptor signaling in keratinocytes triggers interleukin-24-dependent psoriasis-like skin inflammation in mice. Immunity. (2013) 39:899–911. doi: 10.1016/j.immuni.2013.10.009
88. Shao J, Zhang B, Yu JJ, Wei CY, Zhou WJ, Chang KK, et al. Macrophages promote the growth and invasion of endometrial stromal cells by downregulating IL-24 in endometriosis. Reproduction. (2016) 152:673–82. doi: 10.1530/REP-16-0278
89. Feng KN, Meng P, Zhang M, Zou XL, Li S, Huang CQ, et al. IL-24 contributes to neutrophilic asthma in an IL-17A-dependent manner and is suppressed by IL-37. Allergy Asthma Immunol Res. (2022) 14:505–27. doi: 10.4168/aair.2022.14.5.505
90. Feng KN, Meng P, Zou XL, Zhang M, Li HK, Yang HL, et al. IL-37 protects against airway remodeling by reversing bronchial epithelial-mesenchymal transition via IL-24 signaling pathway in chronic asthma. Respir Res. (2022) 23:244. doi: 10.1186/s12931-022-02167-7
91. Mitamura Y, Nunomura S, Furue M, and Izuhara K. IL-24: A new player in the pathogenesis of pro-inflammatory and allergic skin diseases. Allergol Int. (2020) 69:405–11. doi: 10.1016/j.alit.2019.12.003
92. Wu B, Huang C, Kato-Maeda M, Hopewell PC, Daley CL, Krensky AM, et al. IL-24 modulates IFN-gamma expression in patients with tuberculosis. Immunol Lett. (2008) 117:57–62. doi: 10.1016/j.imlet.2007.11.018
93. Rokonay R, Veres-Szekely A, Szebeni B, Pap D, Lippai R, Beres NJ, et al. Role of IL-24 in the mucosal remodeling of children with coeliac disease. J Transl Med. (2020) 18:36. doi: 10.1186/s12967-020-02221-2
94. Andoh A, Shioya M, Nishida A, Bamba S, Tsujikawa T, Kim-Mitsuyama S, et al. Expression of IL-24, an activator of the JAK1/STAT3/SOCS3 cascade, is enhanced in inflammatory bowel disease. J Immunol. (2009) 183:687–95. doi: 10.4049/jimmunol.0804169
95. Lee KM, Kang HA, Park M, Lee HY, Choi HR, Yun CH, et al. Interleukin-24 attenuates beta-glycerophosphate-induced calcification of vascular smooth muscle cells by inhibiting apoptosis, the expression of calcification and osteoblastic markers, and the Wnt/beta-catenin pathway. Biochem Biophys Res Commun. (2012) 428:50–5. doi: 10.1016/j.bbrc.2012.09.145
96. Wang Z, Wang Y, Chen Y, and Lv J. The IL-24 gene protects human umbilical vein endothelial cells against H(2)O(2)-induced injury and may be useful as a treatment for cardiovascular disease. Int J Mol Med. (2016) 37:581–92. doi: 10.3892/ijmm.2016.2466
97. Wang HH, Huang JH, Sue MH, Ho WC, Hsu YH, Chang KC, et al. Interleukin-24 protects against liver injury in mouse models. EBioMedicine. (2021) 64:103213. doi: 10.1016/j.ebiom.2021.103213
98. Wang J, Hu B, Zhao Z, Zhang H, Zhang H, Zhao Z, et al. Intracellular XBP1-IL-24 axis dismantles cytotoxic unfolded protein response in the liver. Cell Death Dis. (2020) 11:17. doi: 10.1038/s41419-019-2209-6
99. Zhang X, Hu C, Zhong Y, Qiao D, Chi W, Shen H, et al. Multifunctional interleukin-24 resolves neuroretina autoimmunity via diverse mechanisms. Int J Mol Sci. (2022) 23:11988. doi: 10.3390/ijms231911988
100. Sie C, Kant R, Peter C, Muschaweckh A, Pfaller M, Nirschl L, et al. IL-24 intrinsically regulates Th17 cell pathogenicity in mice. J Exp Med. (2022) 219:e20212443. doi: 10.1084/jem.20212443
101. Chong WP, Mattapallil MJ, Raychaudhuri K, Bing SJ, Wu S, Zhong Y, et al. The cytokine IL-17A limits th17 pathogenicity via a negative feedback loop driven by autocrine induction of IL-24. Immunity. (2020) 53:384–97 e5. doi: 10.1016/j.immuni.2020.06.022
102. Ma YF, Ren Y, Wu CJ, Zhao XH, Xu H, Wu DZ, et al. Interleukin (IL)-24 transforms the tumor microenvironment and induces anticancer immunity in a murine model of colon cancer. Mol Immunol. (2016) 75:11–20. doi: 10.1016/j.molimm.2016.05.010
103. Yu X, Miao J, Xia W, and Gu ZJ. Immunogenicity moderation effect of interleukin-24 on myelogenous leukemia cells. Anticancer Drugs. (2018) 29:353–63. doi: 10.1097/CAD.0000000000000606
104. Miyahara R, Banerjee S, Kawano K, Efferson C, Tsuda N, Miyahara Y, et al. Melanoma differentiation-associated gene-7 (mda-7)/interleukin (IL)-24 induces anticancer immunity in a syngeneic murine model. Cancer Gene Ther. (2006) 13:753–61. doi: 10.1038/sj.cgt.7700954
105. Yan S, Zhang H, Xie Y, Sheng W, Xiang J, Ye Z, et al. Recombinant human interleukin-24 suppresses gastric carcinoma cell growth in vitro and in vivo. Cancer Invest. (2010) 28:85–93. doi: 10.3109/07357900903095672
106. Ma Y, Chen HD, Wang Y, Wang Q, Li Y, Zhao Y, et al. Interleukin 24 as a novel potential cytokine immunotherapy for the treatment of Mycobacterium tuberculosis infection. Microbes Infect. (2011) 13:1099–110. doi: 10.1016/j.micinf.2011.06.012
107. Moreno Ayala MA, Gottardo MF, Imsen M, Asad AS, Bal de Kier Joffe E, Casares N, et al. Therapeutic blockade of Foxp3 in experimental breast cancer models. Breast Cancer Res Treat. (2017) 166:393–405. doi: 10.1007/s10549-017-4414-2
108. Saleh R and Elkord E. FoxP3(+) T regulatory cells in cancer: Prognostic biomarkers and therapeutic targets. Cancer Lett. (2020) 490:174–85. doi: 10.1016/j.canlet.2020.07.022
109. Abhinand CS, Raju R, Soumya SJ, Arya PS, and Sudhakaran PR. VEGF-A/VEGFR2 signaling network in endothelial cells relevant to angiogenesis. J Cell Commun Signal. (2016) 10:347–54. doi: 10.1007/s12079-016-0352-8
110. Cvetkovic J, Jacobi RHJ, Miranda-Bedate A, Pham N, Kutmon M, Groot J, et al. Human monocytes exposed to SARS-coV-2 display features of innate immune memory producing high levels of CXCL10 upon restimulation. J Innate Immun. (2023) 15:911–24. doi: 10.1159/000535120
111. Marotel M, Villard M, Drouillard A, Tout I, Besson L, Allatif O, et al. Peripheral natural killer cells in chronic hepatitis B patients display multiple molecular features of T cell exhaustion. Elife. (2021) 10:e60095. doi: 10.7554/eLife.60095.sa2
112. Pontrelli P, Rascio F, Zaza G, Accetturo M, Simone S, Infante B, et al. Interleukin-27 is a potential marker for the onset of post-transplant Malignancies. Nephrol Dial Transplant. (2019) 34:157–66. doi: 10.1093/ndt/gfy206
Keywords: tumor suppressor cytokine, immune cells, therapeutic agent, immuno-stimulation, signaling pathway
Citation: Aravind A, Shaji V, Ahmed M, Dagamajalu S, Vijayakumar M, Prasad TSK and Raju R (2025) An assembled molecular signaling map of interleukin-24: a resource to decipher its multifunctional immunoregulatory role in pathophysiological conditions. Front. Immunol. 16:1608101. doi: 10.3389/fimmu.2025.1608101
Received: 11 April 2025; Accepted: 10 June 2025;
Published: 30 June 2025.
Edited by:
Juan Bautista De Sanctis, Palacký University Olomouc, CzechiaReviewed by:
Paul B. Fisher, Virginia Commonwealth University, United StatesWai Po Chong, Hong Kong Baptist University, China
Moira Sauane, The City University of New York, United States
Copyright © 2025 Aravind, Shaji, Ahmed, Dagamajalu, Vijayakumar, Prasad and Raju. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rajesh Raju, cmFqcnJuYnRAZ21haWwuY29t; cmFqZXNocmFqdUB5ZW5lcG95YS5lZHUuaW4=; Thottethodi Subrahmanya Keshava Prasad, dHNrcHJhc2FkQGdtYWlsLmNvbQ==; a2VzaGF2QHllbmVwb3lhLmVkdS5pbg==; Manavalan Vijayakumar, dmljZWNoYW5jZWxsb3JAeWVuZXBveWEuZWR1Lmlu; bXZpamFpMjAxM0BnbWFpbC5jb20=