 Špela Konjar
Špela Konjar Evgen Benedik
Evgen Benedik Marko Šestan
Marko Šestan Marc Veldhoen
Marc Veldhoen Anže Županič
Anže Županič- 1Blood Transfusion Centre of Slovenia, Ljubljana, Slovenia
- 2Center for Proteomics, Faculty of Medicine, University of Rijeka, Rijeka, Croatia
- 3University Children’s Hospital, University Medical Centre Ljubljana, Ljubljana, Slovenia
- 4Biotechnical Faculty, University of Ljubljana, Ljubljana, Slovenia
- 5Department of Histology and Embryology, Faculty of Medicine, University of Rijeka, Rijeka, Croatia
- 6GIMM - Gulbenkian Institute for Molecular Medicine, Lisbon, Portugal
- 7Faculdade de Medicina da Universidade de Lisboa, Lisbon, Portugal
- 8Department of Biotechnology and Systems Biology, National Institute of Biology, Ljubljana, Slovenia
The global rise in inflammatory bowel disease (IBD) and other non-communicable diseases (NCDs) over the past five decades has coincided with the widespread adoption of a Western diet and lifestyle. These conditions, characterised by chronic inflammation, are shaped by complex interactions between genetic, environmental, immunological, and microbial factors. The Western diet rich in, refined sugars, unhealthy fats, ultra-processed foods and excess salt, is increasingly recognised as a major contributor to immune dysfunction, microbial dysbiosis, and compromised intestinal barrier integrity, all hallmarks of IBD. Systems biology offers a powerful framework for untangling the complexity of IBD by integrating large-scale biological data from various sources, leveraging computational modelling, high-throughput analyses, and network-based approaches to identify key regulatory pathways and molecular interactions driving disease progression. Complementary to this, nutritional epidemiology provides critical insights into the role of diet in IBD pathogenesis. By combining systems biology with nutritional epidemiology, researchers can move toward personalised dietary interventions and new therapeutic strategies, offering new opportunities for prevention and addressing the growing burden of IBD in societies adopting Western lifestyles. This review synthesise current findings and proposes integrated approaches for future precision prevention and treatment of IBD.
1 Introduction
The global incidence of inflammatory bowel disease (IBD), which includes Crohn’s disease (CD) and ulcerative colitis (UC), has been rising steadily over the past few decades, particularly in regions undergoing rapid Westernisation (1) (Figure 1). UC typically presents as a continuous inflammation in the colon, beginning in the distal colon and advancing towards the proximal colon up to the cecum. This inflammation may result in the development of ulcers and bleeding. CD can manifest as patchy lesions anywhere in the gastrointestinal tract, often accompanied by inflammation, stenosis, and/or fistulas (2). IBD is characterised by alternating phases: periods of remission or quiescence, during which there is no intestinal inflammation and periods of active disease marked by the re-emergence of inflammation and related symptoms. The precise origins of IBD phenotypes and its extraintestinal manifestation remain incompletely understood (3–7). Notably, early-onset paediatric CD is associated with more severe disease progression and may be more strongly influenced by early environmental exposures, particularly dietary factors and microbiota composition (8, 9).
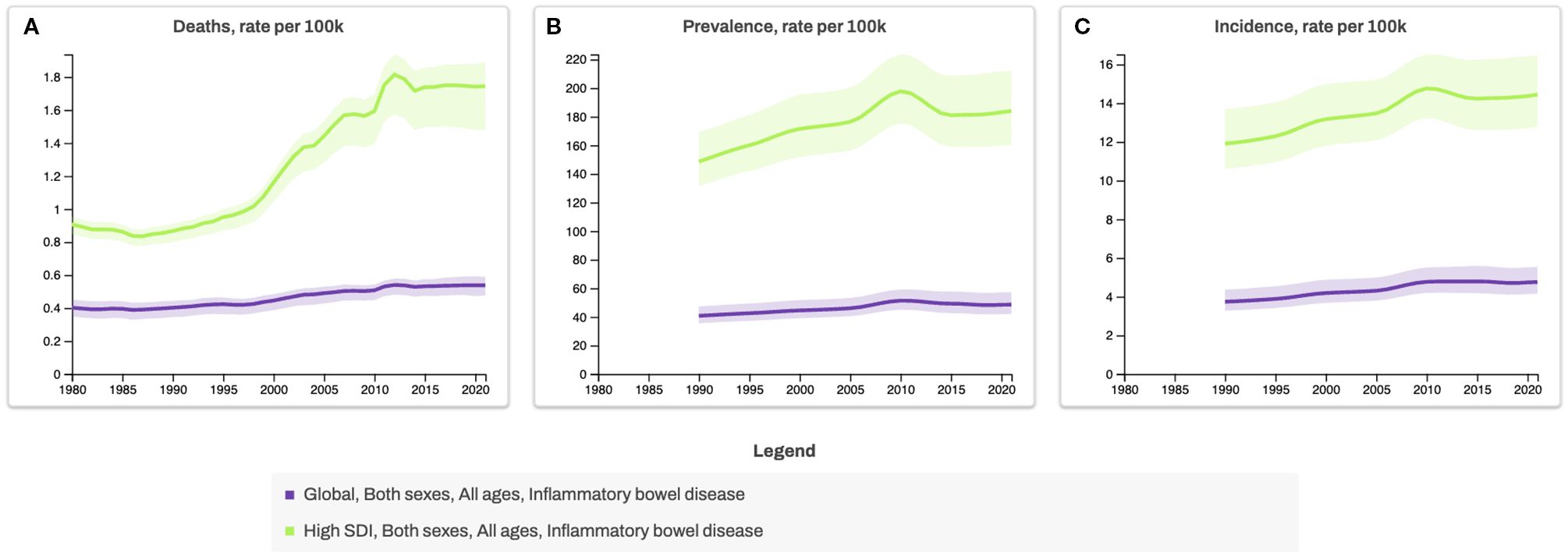
Figure 1. Global and High-Sociodemographic Index (SDI) Trends in Inflammatory Bowel Disease Mortality, Prevalence, and Incidence Rates, 1980–2020. Line graphs show age-standardized rates of (A) deaths, (B) prevalence, and (C) incidence of inflammatory bowel disease per 100,000 population for all ages and both sexes, stratified by SDI categories. The SDI is a composite measure from the Global Burden of Disease study that classifies countries by development level, based on income per capita, educational attainment, and fertility rate under age 25. The Global category represents the combined average across all countries, whereas High-SDI countries represent those with the highest development levels. *Data source for Figures 1, 2: Global Burden of Disease Collaborative Network. Global Burden of Disease Study 2021 (GBD 2021) Results. Seattle, United States: Institute for Health Metrics and Evaluation (IHME), 2022 (https://vizhub.healthdata.org/gbd-results, accessed 3 July 2025).
The prevalence of IBD is expected to rise to 1% in both developed and newly industrialised countries, mirroring the rise in Western-style dietary and lifestyle habits (10). With approximately 75% of patients experiencing persistent pattern of relapse this indicate the urgent need for a deeper understanding of these relapsing diseases (11–13).
While genetic predisposition plays a role in disease susceptibility, genetics alone is unlikely to explain the rapid increase in cases. Instead, environmental factors, particularly diet and lifestyle changes, are thought to be major drivers of IBD onset and progression. This aligns with broader epidemiological trends showing a parallel rise in other non-communicable diseases (NCDs), such as obesity, cardiovascular disease, type 2 diabetes and types of cancer, all of which share chronic inflammation as a common pathological feature (1) (Figures 1, 2). The Western diet, characterised by high consumption of refined carbohydrates, unhealthy fats, ultra-processed foods (UPFs), and excess salt, has emerged as a key contributor to systemic and gut-specific inflammation. Reinforcing the need to investigate dietary triggers in IBD pathogenesis.
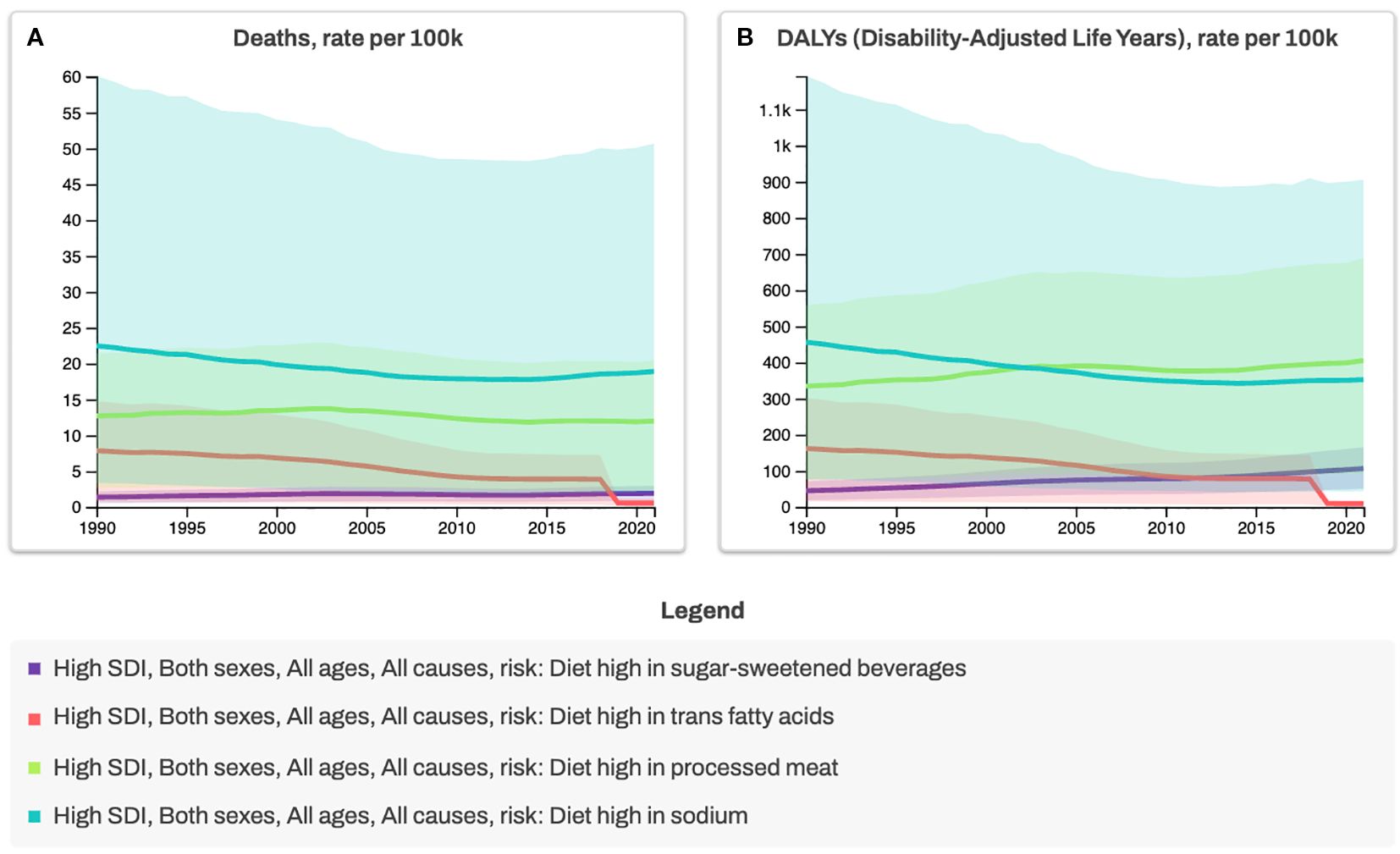
Figure 2. Trends in Mortality and Disability-Adjusted Life Years (DALYs) Attributable to Dietary Risk Factors in High-Sociodemographic Index (SDI) Countries, 1990–2020. Deaths (A) and diet-related DALYs (B) are shown as age-standardized rates per 100,000 population for all ages and both sexes. Analyses are stratified by four dietary risk factors: high intake of sugar-sweetened beverages, trans fatty acids, processed meat, and sodium. High- SDI countries represent nations with the highest income levels, greatest educational attainment, and lowest fertility rates, typically highly developed settings with advanced healthcare systems. Regulatory measures, such as trans fat bans (red line), illustrate the health benefits of reducing harmful dietary exposures and provide a model for comprehensive diet-related risk reduction strategies. *Data source for Figures 1, 2: Global Burden of Disease Collaborative Network. Global Burden of Disease Study 2021 (GBD 2021) Results. Seattle, United States: Institute for Health Metrics and Evaluation (IHME), 2022 (https://vizhub.healthdata.org/gbd-results, accessed 3 July 2025).
A growing body of research suggests that diet influence IBD pathogenesis through multiple mechanisms, including alterations in gut microbiota composition (dysbiosis), increased intestinal permeability, immune dysregulation, and psychological disorders affecting the gut-brain axis (11, 14–16) (Figures 3, 4). The gut microbiome plays a crucial role in maintaining intestinal homeostasis, influencing immune responses, and modulating inflammatory pathways. However, Western dietary patterns promote microbial imbalances that favour the increase of pro-inflammatory species while reducing beneficial bacteria. This shift in microbial composition is associated with a breakdown in gut barrier function, leading to increased permeability and facilitating the translocation of luminal antigens that can trigger mucosal inflammation (Figure 4). Despite accumulating evidence, pinpointing specific dietary triggers and their molecular effects remains challenging due to the complex interplay of genetic, microbial, and environmental factors. Traditional reductionist approaches to nutrition research, focusing on isolated nutrients or food components, often fail to capture these complexities. Instead, a systems-oriented perspective is needed to understand how diet influences gut health and disease risk at a broader level.
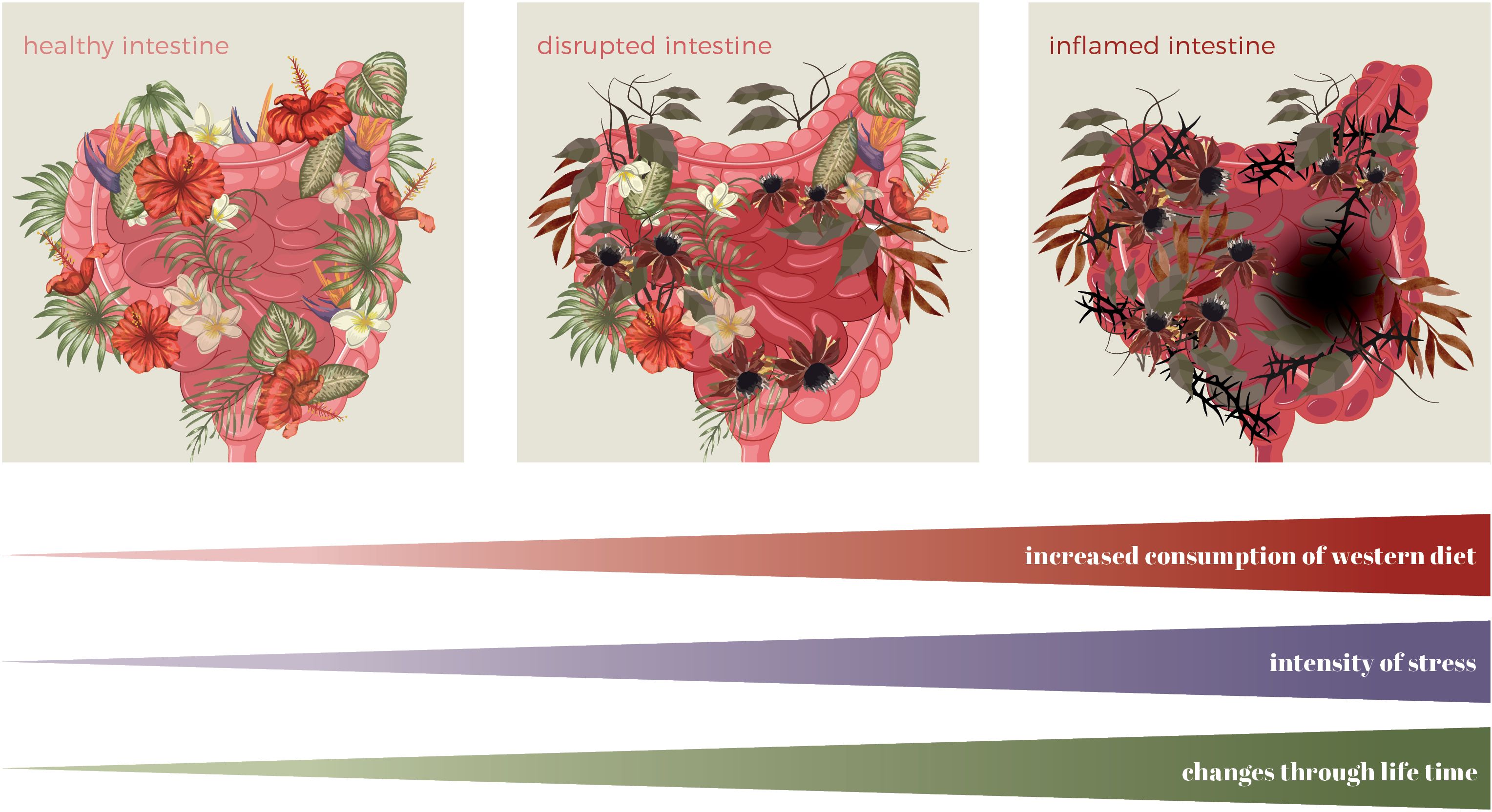
Figure 3. Changes in the intestine throughout a lifetime due to a Western diet and stress. Symbolic graphical presentation of intestinal landscape (microbiota=plants, intensity of inflammation=intensity of the colour of the intestine, lesion=black hole).
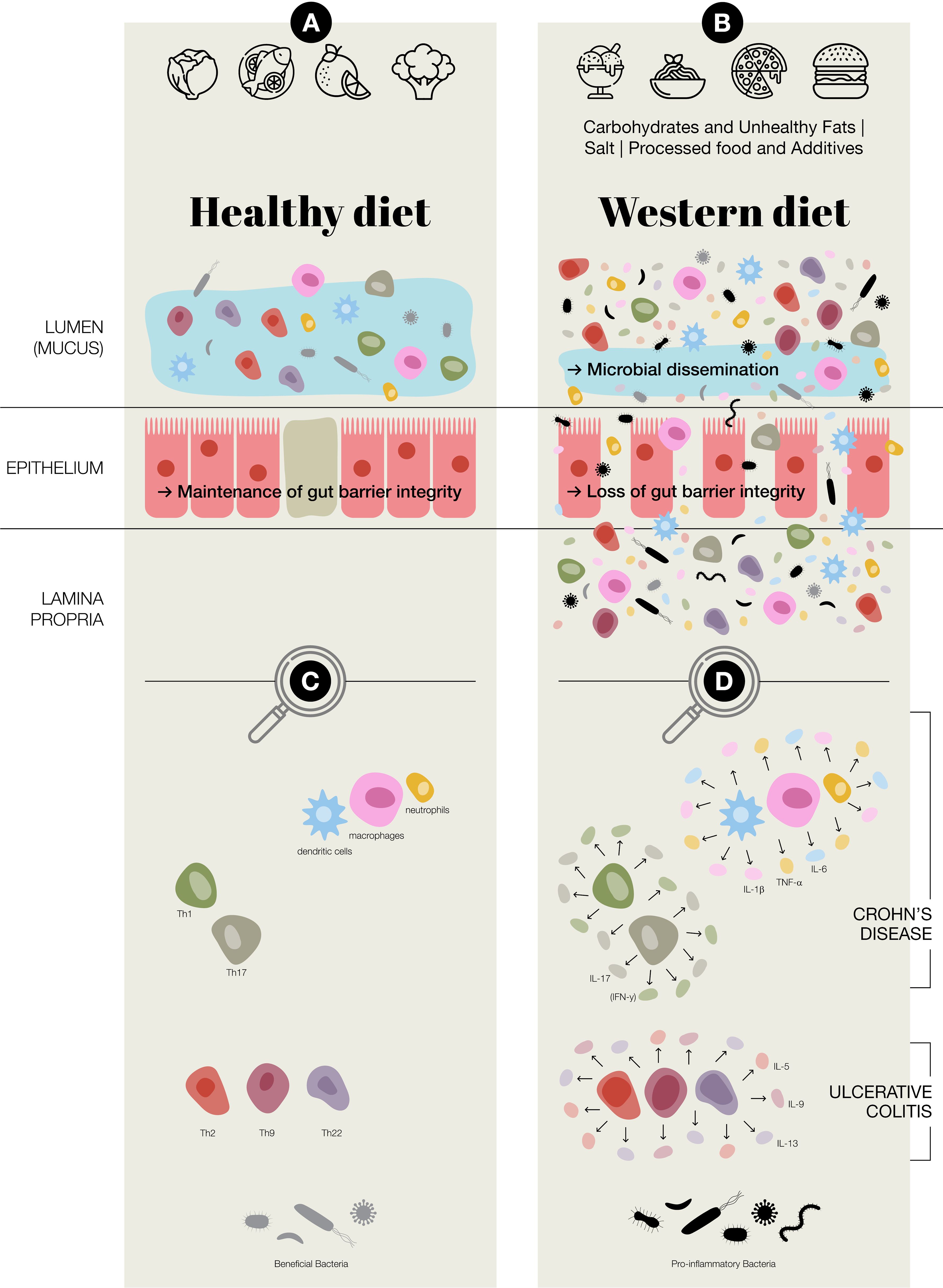
Figure 4. Potential of Western diet inducing chronic inflammation in the gut. (A) Healthy diet, maintenance of gut barrier integrity, normal immune regulation at the intestinal epithelial barrier, increased presence of beneficial bacteria; Faecalibacterium prausnitzii and Bifidobacterium (B) Western diet (Carbohydrates & Unhealthy Fats: alter gut microbiota, contribute to metabolic dysregulation and gut inflammation, increase intestinal permeability and endotoxemia. Processed food & Additives: disrupt gut microbial balance, erosion of the protective mucus layer, chronic low-grade inflammation. Salt: disrupting immune homeostasis, intestinal inflammation, alter the gut microbiota–immune system axis.); in general an aberrant activation of both innate and adaptive immune responses, loss of gut barrier integrity, a reduction in beneficial bacteria, such as Faecalibacterium prausnitzii and Bifidobacterium, and an increase in potentially pro-inflammatory species of bacteria, including adherent-invasive Escherichia coli and Ruminococcus gnavus (C) Predominate adaptive and innate immune cells present at intestinal epithelial barrier (D) Aberrant regulation/activation of predominate adaptive and innate immune cells in chronic inflammation in inflammatory bowel disease, which includes Crohn’s disease and ulcerative colitis.
Systems biology provides a powerful framework for studying IBD by integrating multi-omic datasets and modelling host–microbiome–immune interactions (17, 18). While computational models hold promise for biomarker discovery and therapeutic target identification, many remain hypothetical and require experimental validation (18). The increasing availability of biological samples, including blood, stool, and intestinal biopsies, facilitates multi-dimensional profiling in IBD research (17, 19).
There are many unresolved questions regarding the relationship between IBD and the Western diet. In this review, we examine how components of the Western diet alter the intestinal landscape, contributing to immune dysfunction, microbial dysbiosis, and impaired intestinal permeability. We explore how systems biology and nutritional epidemiology offer complementary tools to unravel the complex diet–IBD interface, by integrating cutting-edge research methodologies, that could in the near future contribute to the development of more effective prevention and treatment strategies.
2 The gut microbiota-immune system axis in IBD
The interplay between the gut microbiota and the immune system is a fundamental aspect of human health, particularly in maintaining intestinal homeostasis, thereby preventing chronic inflammation. Microbes continuously stimulate the host’s immune system. Immune tolerance to components of the microbiota prevents risk of intestinal inflammation (20, 21). A fibre-rich diet can encourage the growth of beneficial gut bacteria, helping to maintain a balanced gut ecosystem. This healthy microbiota, in turn, supports a well-regulated immune response in the intestines—an important factor in managing IBD (21, 22). However, in individuals with a dysfunctional or hyperactive immune system, this tolerance is undermined, and can lead to chronic enteritis and colitis. IBD, is characterised by a disrupted relationship between host immunity and the microbial environment of the gut (21, 23, 24). Changes in microbial composition and metabolites caused by dysbiosis affect also autophagy, while impaired autophagy disrupts the mucosal barrier and immune balance, worsening gut inflammation (25, 26). The dynamic interactions between the gut microbiota, epithelial barrier, and immune system drive disease pathogenesis (Figure 4) (27). Understanding these complex interactions is crucial for developing novel therapeutic strategies aimed at restoring gut homeostasis and mitigating chronic inflammation.
2.1 Gut microbiota in IBD
The gut microbiota comprises of diverse communities of microorganisms, including bacteria, archaea, fungi, and viruses, which collectively perform essential functions in nutrient metabolism, immune modulation, and epithelial protection. In a healthy state, this microbial ecosystem is characterised by a balance between commensal and potentially pathogenic species, with dominant bacterial phyla including Firmicutes, Bacteroidetes, Actinobacteria, and Proteobacteria. These microorganisms contribute to gut health by fermenting dietary fibres into short-chain fatty acids (SCFAs), regulating mucosal immunity, and reinforcing the integrity of the epithelial barrier.
In IBD, however, a distinct shift in microbial composition, referred to as dysbiosis, has been consistently observed. This dysbiotic state is characterised by a reduction in beneficial bacteria, such as Faecalibacterium prausnitzii and Bifidobacterium (28), and an increase in potentially pro-inflammatory species, including adherent-invasive Escherichia coli and Ruminococcus gnavus (29). The loss of microbial diversity is particularly concerning as it is associated with impaired production of SCFAs, weakened mucosal protection, and heightened immune activation. Additionally, dysbiosis is not only a consequence but also a driver of disease progression, as an altered microbial landscape can influence immune cell differentiation and cytokine production, further promoting chronic inflammation (30, 31). Recent evidence suggests that dysbiosis may precede clinical disease onset, supporting its potential role as an early biomarker of IBD risk (17). Emerging evidence suggests that reduced microbial diversity and enrichment of pro-inflammatory taxa (e.g., Enterobacteriaceae) may serve as early predictors of IBD relapse (32).
Research in animal models and human studies has demonstrated a lasting imbalance in the gut microbiome in IBD (33–39). Reducing bacterial load with broad-spectrum antibiotics decreases the severity of colitis in IL-10 deficient mice (40). Another argument for the role of the microbiota is that CD lesions primarily occur in areas of the intestine with high bacterial populations such as ileum and colon (41).
2.1.1 Microbiota-derived metabolites and their immunomodulatory effects during IBD
In addition to shaping immune responses via direct interactions with immune receptors, the gut microbiota exerts profound effects on host immunity through the production of microbial metabolites. Among these, SCFAs, the most well-studied being butyrate, acetate, and propionate, play a critical role in maintaining intestinal homeostasis. Butyrate, in particular, has been shown to enhance epithelial barrier function by upregulating tight junction proteins and stimulating mucus production (42). Furthermore, SCFAs exhibit anti-inflammatory properties by modulating the activity of macrophages, dendritic cells, and regulatory T cells (Treg), thereby promoting immune tolerance and suppressing excessive inflammation (43). Clinical studies further suggest that maintaining SCFA production, particularly butyrate, is associated with prolonged remission periods in IBD patients (34).
However, in the dysbiotic state observed in IBD, the production of beneficial microbial metabolites is often diminished, while harmful metabolites such as hydrogen sulphide and trimethylamine-N-oxide (TMAO) are increased (44). Hydrogen sulphide, although produced in small amounts by commensal bacteria under normal conditions, becomes cytotoxic at higher concentrations, impairing mitochondrial respiration, disrupting epithelial barrier integrity, and inducing oxidative stress in intestinal tissues (45). Elevated hydrogen sulphide levels can promote epithelial injury and sustain inflammatory signalling. Beyond activating NOD-like receptor family, pyrin domain containing 3 (NLRP3) inflammasome pathways, trimethylamine-N-oxide has been implicated in promoting mitochondrial dysfunction, enhancing oxidative stress, and impairing epithelial cell renewal, thereby exacerbating barrier breakdown and chronic mucosal inflammation (46). The accumulation of these harmful metabolites thus represents a key mechanistic link between dysbiosis and the perpetuation of intestinal inflammation in IBD (47).
2.1.2 Microbiota and its impact on intestinal barrier dysfunction
Studies have shown that the microbiota of individuals with IBD is less functionally diverse, producing fewer SCFAs and more pro-inflammatory metabolites, which further exacerbate gut barrier disruption and immune dysregulation (48).
A key consequence of dysbiosis is its impact on the intestinal epithelial barrier, a complex structure that serves as the first line of defence against luminal pathogens and harmful antigens. The integrity of this barrier is maintained by tight junction proteins such as occluding, claudins and zonula occludens-1, which regulate paracellular permeability. In IBD, the Western diet has been implicated in disrupting tight junction assembly (49), leading to increased permeability and the phenomenon sometimes referred to as “leaky gut”, which is not a medical diagnosis. This reduction in barrier function facilitates the translocation of microbial-derived molecules, such as lipopolysaccharides and flagellin, which subsequently activate innate immune receptors and drive inflammatory cascades (50). The continuous exposure of the immune system to these microbial antigens can result in a state of hyperactivation, characterized by the excessive production of pro-inflammatory cytokines, including tumour necrosis factor-alpha (TNF-α) and interleukin (IL)-6, both of which are central to IBD pathogenesis (51). Additionally, oxidative stress caused by high-fat, low-fibre diets further compromise epithelial cell function, increasing susceptibility to inflammation and tissue damage (52, 53).
Emerging evidence suggests that dietary factors significantly influence barrier function, with Western dietary patterns rich in emulsifiers and artificial sweeteners further exacerbating epithelial dysfunction. Emulsifiers, such as polysorbate-80 and carboxymethylcellulose, have been shown to alter gut microbiota composition and promote low-grade inflammation, which can predispose individuals to barrier disruption (54). The increased permeability observed in IBD is also associated with alterations in the mucin layer, as reduced expression of mucin 2 (MUC2), a key mucin component, is frequently reported in patients with active disease (55).
2.2 Innate and adaptive immune responses in IBD
IBD is characterised by an aberrant activation of both innate and adaptive immune responses. The innate immune system, plays a crucial role in recognising and responding to microbial-associated molecular patterns through pathogen recognition receptors. In healthy individuals, innate immune responses are tightly regulated to prevent excessive inflammation. However, in IBD, innate immune cells, such as macrophages, dendritic cells, and neutrophils exhibit increased production of inflammatory mediators such as TNF-α, IL-1β, and IL-6 (56).
Neutrophils, in particular, are heavily recruited to the inflamed gut mucosa in IBD, where they release reactive oxygen species (ROS) and proteolytic enzymes that contribute to epithelial damage (57). Additionally, macrophages and dendritic cells in IBD patients display an altered cytokine profile, favouring the differentiation of pro-inflammatory T-helper (Th) cell subsets while suppressing regulatory mechanisms that could dampen inflammation (54, 58).
The adaptive immune system, which provides antigen-specific responses, also plays a central role in IBD pathogenesis. In CD, there is an overactivation of Th1 and Th17 cell immune responses, leading to excessive production of interferon-gamma (IFN-γ) and IL-17, both of which amplify inflammatory pathways and can drive tissue damage (59). Conversely, UC is primarily associated with a dysregulated Th2 cell response, characterised by increased secretion of IL-5 and IL-13, which disrupt epithelial barrier integrity and compromise the mucus layer (60).
Another key player in immune regulation is the population of Tregs, which are responsible for maintaining immune tolerance and suppressing excessive inflammation. In IBD, the balance between pro-inflammatory and regulatory immune responses is disrupted, with a reduction of Treg cells and an expansion of effector T cells. This imbalance results in persistent immune activation, prolonged tissue injury, and impaired mucosal healing. Importantly, the interaction between microbiota-derived metabolites and Treg cell differentiation and function is a growing area of research, with SCFAs playing a key role in promoting Treg expansion and function (61–64).
Significant challenges remain in elucidating the complex interplay within the gut microbiota-immune system axis during IBD. Key questions involve understanding precisely how and to what extent the homeostasis between the microbiota, the epithelial barrier, and the host immune system is disrupted (31, 47, 65–67). This includes determining the mechanisms driving the loss of microbial diversity and characterizing the functional consequences of altered microbial metabolite profiles during different disease stages (e.g., remission versus active disease). The inherent complexity arises from the multitude of interacting variables—spanning genetic, microbial, immune, and environmental factors—and their dynamic nature. Addressing these questions necessitates integrative approaches. As discussed further in Chapter 4, systems biology, which combines high-throughput ‘omics’ technologies with computational modelling, provides essential tools to dissect these interactions in both relevant animal models and human studies, paving the way for a more mechanistic understanding of IBD pathogenesis.
3 The role of diet in IBD
Inflammatory disorders are a common basis to NCDs and IBDs. NCDs include obesity, type 2 diabetes for which risk factor are carbohydrate and fats (68), different types of cancer for which ultra-processed food (UPF) is a risk factor (69) and cardiovascular diseases (CVD) for which a risk factor is salt (70). Given the epidemiological and mechanistic overlaps between IBD and NCDs, and the fact that these dietary risk factors are typical components of the Western diet, this chapter will explore how carbohydrates, fats, UPFs, and salt contribute to the development and exacerbation of IBD.
3.1 Carbohydrates and unhealthy fats as a risk factor for IBD
While the Western diet as a whole has been implicated in exacerbating IBD, individual dietary components exert specific effects on gut microbiota composition, immune responses, and epithelial integrity. Among the most concerning elements of this diet is the excessive consumption of refined carbohydrates, including sucrose and fructose, found in sugar-sweetened foods and beverages, and processed starches in energy-dense, nutrient-poor products, which contribute to metabolic dysregulation and gut inflammation (71). High-fructose corn syrup, a ubiquitous ingredient in processed foods and beverages, has been shown to increase gut permeability and oxidative stress, further aggravating inflammatory responses (71).
Carbohydrates include monosaccharides, disaccharides, oligosaccharides (typically comprising 3–10 units), and polysaccharides. Monosaccharides and disaccharides—often referred to as sugars—are generally soluble and are commonly consumed in the form of refined carbohydrates such as sucrose, fructose, and starch, found in energy-dense but nutrient-poor processed foods (72). Within the Western dietary pattern, refined sugars and processed grains (e.g. white flour and white bread) stand out for their detrimental impact on gut microbial composition and immune function (73).
The intake of refined sugars, particularly from sugar-sweetened beverages, has been shown to alter the gut microbiota, increasing the Firmicutes/Bacteroidetes ratio and reducing levels of beneficial butyrate-producing bacteria such as Lachnobacterium (74). These changes are linked to increased intestinal permeability and endotoxemia, which in turn promote local and systemic inflammation (75). Khan et al. investigated the effects of a high-sugar diet on colitis in rodent models and demonstrated that excessive intake of simple sugars exacerbated colitis in mice, whether administered before or after disease induction (76). Excessive sugar consumption has also been identified as a risk factor for obesity, type 2 diabetes, and cardiovascular disease (77). Furthermore, high sugar intake has been shown to induce physiological changes in the intestinal epithelium, including increased cell proliferation in crypts (78).
Fats, though an essential component of a balanced diet, can become harmful depending on their type and quantity. Unsaturated fats, often referred to as “healthy fats,” are associated with reduced risk of cardiovascular disease and metabolic syndrome (79, 80). In contrast, saturated and trans fats—prevalent in processed foods—are used to improve shelf life, texture, and flavours (81). These fats have been linked to intestinal dysfunction, including altered enteroendocrine cell activity, increased permeability, and mucosal inflammation (82). These effects contribute to chronic low-grade inflammation, a common feature underlying various NCDs (83).
Animal studies have demonstrated that high-fat diets significantly alter the gut microbiota, increasing the Firmicutes/Bacteroidetes ratio, reducing the antimicrobial function of Paneth cells, and elevating pro-inflammatory cytokines such as IFN-γ, TNF-α, IL-1β, and IL-6. These alterations promote bacterial translocation and systemic endotoxemia (72, 74).
The Western diet is often rich in fats associated with negative health outcomes, including refined seed oils (e.g. sunflower oil, palm oil), coconut oil, and processed foods high in omega-6 polyunsaturated fatty acids (84, 85). When consumed in excess, especially in the context of industrial food processing, these fatty acids may disrupt microbial homeostasis, promote the release of pro-inflammatory mediators, and compromise intestinal barrier integrity. Such alterations have been associated with an increased risk of IBD and colorectal cancer (86–90).
Likewise, diets rich in saturated and trans fats—particularly those derived from processed meats and fried foods—promote a microbial profile that favours pro-inflammatory species, thereby increasing susceptibility to intestinal inflammation (91). The consumption of red and processed meats has also been linked to the generation of harmful microbial metabolites, such as trimethylamine-N-oxide (TMAO), which contribute to both intestinal and systemic inflammation (92, 93).
3.2 Processed food and additives as a risk factor for IBD
The presence of food additives and emulsifiers in processed foods further complicates the dietary landscape of IBD. These substances may alter the composition and function of the gut microbiota, promoting the expansion of bacteria with pathogenic potential. In particular, dietary emulsifiers such as carboxymethylcellulose and polysorbate-80 have been shown to increase mucolytic bacterial activity, resulting in erosion of the protective mucus layer that separates the microbiota from the intestinal epithelium (53).
The thinning of this mucus barrier facilitates direct microbial–epithelial interactions, which can activate immune pathways and exacerbate inflammatory responses (66). Additionally, artificial sweeteners, widely used as sugar substitutes, have been implicated in metabolic dysregulation and shifts in microbial composition, potentially contributing to pro-inflammatory states within the gut (94).
Many food products within the Western diet are subjected to industrial processing techniques such as high-temperature cooking and extrusion. During these processing methods, harmful compounds such as advanced glycation end-products and other pro-inflammatory molecules can form through reactions between sugars, proteins, and lipids (95–97). Moreover, the Western diet is rich in UPFs, which typically contain a range of food additives, emulsifiers, preservatives, and synthetic colourings that may disrupt gut microbial balance and contribute to chronic low-grade inflammation (98). Synthetic food colorants, such as azo dyes (e.g. Red 40 and Yellow 6), are among the most commonly used additives in processed foods. Experimental evidence in genetically susceptible mouse models indicates that these compounds may induce colitis-like inflammation. Commensal bacteria, including Bacteroides ovatus and Enterococcus faecalis, have been shown to metabolize these dyes into 1-amino-2-naphthol-6-sulphonate sodium salt, a metabolite associated with colitis development (99).
Similarly, artificial sweeteners such as aspartame and sucralose have been shown to alter gut microbial composition, favouring the proliferation of pro-inflammatory bacterial species. Collectively, these dietary components can impair gut barrier integrity, enabling bacterial translocation and the passage of microbial antigens into the mucosal immune system, thereby exacerbating inflammation and mucosal damage (100).
However, there are challenges in interpreting data regarding the health effects of UPFs. The nutritional composition and quality of UPFs vary widely; for example, both seasoned canned tuna and ice cream are classified as UPFs despite their differing nutritional profiles (101). In a recent prospective cohort study, an association with CD risk was observed only when comparing extreme levels of UPF consumption (102). Furthermore, diets higher in UPF tend to also be high in fat and sugar and low in dietary fibre, making it difficult to determine whether health effects are due to ultra-processing itself or the overall nutritional composition. Although the precise mechanisms by which UPFs influence IBD risk are not yet fully understood, current clinical guidelines recommend that UPFs should be minimized in the diets of individuals with IBD or those at increased risk (103–105). Future studies employing systems biology approaches could help isolate the independent effects of food processing from overall nutrient composition.
3.3 Salt as a risk factor for IBD
Processed and preserved foods prevalent in the Western diet typically contain high levels of salt. However, the effects of this excessive salt intake on the human gut microbiota remain largely unknown (106). Several studies have linked UPFs to excessive salt consumption, which contributes to the development of hypertension and cardiovascular disease (107).
A high-salt diet (HSD) increased the abundance of Bifidobacterium and promoted greater gut permeability, which facilitated the intratumoural localization of Bifidobacterium. This, in turn, enhanced natural killer (NK) cell activity and contributed to tumour regression (108). Wilck et al. (2017) demonstrated that high-salt diets alter the gut microbiota–immune system axis by reducing populations of Lactobacillus spp., which may affect the production of SCFAs. This microbial shift was associated with enhanced activation of pro-inflammatory Th17 cells also limited metabolite availability cause cellular stress and enhance Th17 cell polarization (109, 110). Th17 cells play a pivotal role in IBD by producing pro-inflammatory cytokines such as IL-17A, IL-17F, and IL-22, which drive neutrophil recruitment, epithelial damage, and persistent inflammation (111). Excessive Th17 responses disrupt mucosal homeostasis, contributing to the chronic inflammatory milieu observed in CD and UC.
Similarly, Wu et al., 2013 showed that a modest increase in dietary salt concentration, promote IL-23 expression and enhance Th17 cell differentiation and pathogenic roles both in vitro and in vivo (112). An emerging body of evidence suggests that increased dietary salt intake, as a component of UPFs-rich diets, may contribute to the pathogenesis of IBD by disrupting immune homeostasis. Experimental models of colitis further support the role of high salt in exacerbating intestinal inflammation and promoting other immune-mediated disorders (113).
3.4 Components of diets that decrease symptoms of IBD
Conversely, some dietary components have been identified as protective factors in maintaining gut health and reducing IBD risk. Fibre-rich diets, promote the growth of beneficial gut bacteria and enhance the production of SCFAs, which are associated with reduced inflammation and improved gut barrier function (114).
Prebiotic fibres, such as inulin and resistant starch, selectively enhance the growth of commensal bacteria, leading to a more diverse and resilient microbiota. Unlike traditional diets, which support gut homeostasis through a diverse nutrient intake, the Western diet is reduced in key bioactive compounds such as dietary fibre and polyphenols (115).
The low intake of dietary fibre, has significant consequences for gut health, as fibre serves as a substrate for microbial fermentation. This process results in the generation of SCFAs—including butyrate, acetate, and propionate—which play a critical role in maintaining epithelial integrity, modulating immune responses, and supporting mucosal energy metabolism (116).
In addition to fibre, other bioactive dietary components such as polyphenols and omega-3 fatty acids have been recognized for their anti-inflammatory and immunomodulatory properties. Polyphenols, abundant in fruits, vegetables, and green tea, exert strong antioxidant effects and modulate inflammatory signalling pathways (117). Similarly, omega-3 fatty acids—commonly found in oily fish such as salmon and mackerel—have been shown to reduce the production of pro-inflammatory cytokines and enhance epithelial barrier function, offering potential therapeutic benefits for individuals with IBD (43).
The Western diet decreases microbiota diversity and consequently impact the composition of microbial metabolites. With metagenomics, metabolomics studies and development of models in systems biology the impact of diet and impact of individual component is already studied and will be extensively studied in the future (118). This will solidify which component of diet have predominate and harmful impact on intestinal microbiota and could be avoid in diets in the treatments of inflammatory disease such as NCDs and IBD.
4 Systems biology and nutritional epidemiology approaches in IBD and western diet
4.1 Systems biology in IBD research
High-throughput ‘omics’ technologies, including genomics, transcriptomics, proteomics, metabolomics, and metagenomics, has revolutionized IBD research by enabling integrated analyses of genetic, microbial, and immune dysregulation. Molecular profiling of patient samples has uncovered genetic risk loci, dysregulated pathways, metabolic alterations, and microbial imbalances, moving research beyond reductionist models. However, despite their promise, multi-omics-based systems approaches are rarely translated into clinical practice, primary due to challenges in data standardization, integration, and interpretation (18).
Genome-wide association studies (GWAS) represent a cornerstone of IBD genetics, identifying over 200 genetic loci linked to disease risk, particularly in pathways related to innate immunity, epithelial barrier function, and host-microbe interactions (17). While GWAS have been instrumental in mapping the genetic architecture of IBD, it’s important to note that the identified genetic predisposition explains only a portion of the overall disease variance, underscoring the significant contribution of environmental factors and gene-environment interactions.
Transcriptomic analyses, measuring gene expression levels, have provided critical insights into the molecular pathways perturbed in IBD. Studies analysing mucosal biopsies and circulating immune cells from IBD patients using bulk RNA sequencing have consistently revealed dysregulated pathways related to cytokine signalling, oxidative stress responses, and immune cell differentiation (19). More recently, the application of single-cell RNA sequencing (scRNA-seq) has added another layer of resolution. This powerful technique has allowed researchers to characterize the extensive heterogeneity of immune and stromal cell populations within the inflamed gut mucosa. Such studies have successfully uncovered distinct populations of pro-inflammatory cells, including specific subsets of Th17 cells, activated macrophages, and mucosa-associated invariant T (MAIT) cells, that are enriched or activated in patients with active IBD (19).
Metabolomics, the study of small molecule profiles in biological samples, complements transcriptomic and genomic data by revealing the functional metabolic outputs of the host and associated microbiota. Analyses of faecal, serum, and tissue metabolites from IBD patients have consistently identified significant alterations compared to healthy controls. Commonly observed changes include reduced levels of beneficial microbial-derived SCFAs like butyrate, alterations in bile acid profiles which impact fat digestion and signalling, and elevated markers indicative of increased oxidative stress within the intestinal environment (119). These metabolic shifts are often closely linked to the observed patterns of microbial dysbiosis and the compromised epithelial barrier function characteristic of IBD.
4.1.1 Analysis focused on individual through time periods of remission and active state of disease
Understanding the multifactorial nature of IBD requires an integrative approach that captures the dynamic interplay between genetic, environmental, microbial, and immunological factors. Understanding the molecular dynamics during remission and active disease phases in individual IBD patients represents a crucial future research avenue. Monitoring time-resolved omics profiles could reveal unique disease trajectories and support personalized interventions. Particularly to understand the dynamics of change through these time points in composition of clusters of transcriptomes, metabolome etc. There are already papers where omics blood sample studies of IBD patient were able to predispose clinical relapse in IBD patient (120). The other publication identified biomarkers associated with functional remission in IBD (18, 121). These are some of the first studies that still need further replication and validation.
At the same it would be important to focus on specific individuals to characterize how different responses develop through times of remission and active disease. This could help to develop dynamic models that would be unique to the individual and, which may benefit in personalized precision medicine treatments. It was already shown that different mouse strains can immunologically respond differently to the same pathogen in the intestine. C57BL6/J and BALB/c mice differ in their tuft cell response to the protozoa T. muris but not the helminth H. Polygyrus in the ileum (122). There is a possibility that individuals develop different, potentially pathological, intestinal responses to the same stimuli or, that IBD pathogenesis develops similarly between groups of people. These differences and similarities could be detected and characterized with systems biology approaches.
4.1.2 Specific versus complex, reductionist versus holistic
Omics data integration, network biology, and interactomes all reflect the inherent complexity, variability, and heterogeneity found in real-life physiological and pathological processes (123). Maintaining health amidst the constant challenges of growth, environmental influences, adaptation, nutrition, metabolism, immunity, behaviour, and both mental and physical demands require the precise coordination of countless physiological functions and their associated ‘omes´ (18, 124). The reductionist approach of seeking a single cause for complex diseases like IBD could be abandoned, as no single gene, environmental factor, microbial imbalance, or immune anomaly has been shown to cause typical CD or UC. Instead, a model that integrates all potential contributors offers a more effective path to understanding molecular mechanisms and identifying precise therapeutic targets (123). By integrating high-dimensional datasets systems biology provides a framework for identifying key molecular drivers and regulatory pathways involved in IBD pathogenesis (17).
With the three-predominant fronts of the intestinal landscape; microbiota, immune cells and epithelial the intestinal system is too complex to develop “clean” models. If we focus on a single omics method, analyses through time periods of remission and active disease and align the data with controls and develop through cluster the models with which we could gain useful, reliable and not too dispersed information. Initially the models can be developed in mice, which in comparison to studies in humans can be more interventional, and are environmentally and genetically controlled.
The understanding of IBD using appropriate models is not an easy reach. Such models, to be established and improved with reliable omics data, are difficult to develop due to the complexity of the multiple interactions, the dynamics of the disease evolution, the intricate anatomy, etc. Studying individual patients at several stages of disease and therapy using omics data is methodologically viable, incorporating sample collection in treatment protocols, documenting changes in medication, diet and lifestyle.
4.1.3 Omics analysis before and after treatment of IBD
The advent of therapies targeting leukocyte trafficking (anti-integrins) or cytokines (anti-TNF, anti-IL-12/23) has transformed the management of IBD, significantly improving clinical remission and endoscopic healing while reducing disease-related morbidity (125). However, only a small subset of patients maintains long-term remission despite an initial response.
In patients with CD single-cell RNA-seq was applied to ileal CD lesions to address whether cellular heterogeneity contributes to TNFα antibody treatment resistance. They found that a subset of patients expressed a unique cellular module in inflamed tissues that consisted of IgG-producing plasma cells, inflammatory mononuclear phagocytes, activated T cells, and stromal cells, which they named the GIMATS module. This showed the existence of two qualitatively distinct subsets of disease, with distinct responses to anti-TNFα therapy (19). In a paper by Lee et al., 2021 (126) they found serum signatures of immune proteins reflecting microbial diversity identified patients more likely to achieve remission with anti-TNFα therapy.
The prediction of response to therapy in IBD patients based on results of multi-omics analyses appears to be at a preliminary stage and the results vary depending on the number and combination of the omics included in the prediction modelling. Perhaps more importantly, it still remains to be verified whether any of the newly described omics-based biomarkers are indeed better than traditional ones, such as C-reactive protein and fecal calprotectin (18, 127).
4.2 Systems biology methodologies for integration and modelling
While individual omics technologies provide valuable information about specific molecular layers, a deeper understanding of IBD requires integrating these diverse data types to capture the complex interplay between different biological processes. Beyond single-omics studies, systems biology methodologies aim to integrate multi-layered data to construct predictive models of disease mechanisms and therapeutic responses. Key approaches include statistical integration, network-based analysis, and computational modelling.
4.2.1 Statistical multi-omics integration
This approach uses statistical methods to identify patterns, correlations, and associations across different omics datasets generated from the same samples or patient cohorts. The goal is often data-driven discovery of relationships without necessarily relying heavily on prior biological knowledge. Common techniques include:
● Multivariate statistics: Employing methods like Principal Component Analysis (PCA), Canonical Correlation Analysis (CCA), Partial Least Squares Discriminant Analysis (PLS-DA), and others to reduce data dimensionality and identify major axes of variation that span multiple omics layers (128). These can help distinguish between disease states or patient subgroups based on combined molecular profiles.
● Factor analysis methods: Tools like Multi-Omics Factor Analysis (MOFA+) aim to disentangle the sources of variation in multi-omics datasets, identifying latent factors that represent coordinated biological processes influencing multiple molecular layers simultaneously (129).
Examples in IBD research include studies integrating host transcriptomics with microbiome data to link gene expression patterns to microbial community structure (17) or combining multiple omics profiles from blood samples to identify signatures predictive of disease relapse (120).
4.2.2 Network-based integration and disease maps
Network-based approaches leverage prior biological knowledge, typically stored in pathway databases (like KEGG, Reactome) or interaction databases (like STRING), to interpret multi-omics data in a biological context. This involves mapping omics data onto existing molecular interaction networks (e.g., protein-protein interaction networks, metabolic pathways, gene regulatory networks) to identify perturbed subnetworks or pathways.
A powerful extension of this concept is the Disease Maps initiative (130). Disease maps are community-driven efforts to build comprehensive, curated, and computationally accessible representations of the molecular mechanisms underlying specific diseases. They integrate information from literature and databases into standardized graphical representations (often using Systems Biology Graphical Notation - SBGN), providing a detailed overview of disease pathophysiology. These maps can be used as knowledge hubs and visualization tools, for instance allowing multiomics data to be overlaid and analysed in the context of known pathways and serve as a basis for computational modelling.
Platforms like MINERVA (131) provide the infrastructure to host, visualize, explore, and analyse these complex disease maps online. While dedicated, comprehensive IBD disease maps are still under development compared to fields like COVID-19 (132). The framework holds immense potential. An IBD map could integrate pathways related to immune cell signalling (TNF, IL-23/Th17, JAK-STAT), epithelial barrier regulation, host-microbe interactions, and the influence of genetic risk factors and environmental triggers like diet, providing a powerful tool for hypothesis generation and data interpretation.
4.2.3 Computational modelling
Computational modelling aims to create mathematical or algorithmic representations of biological systems or subsystems to simulate their behaviour over time or under different conditions. These models often integrate data from multiple omics layers or are built upon the knowledge curated in network maps. Common modelling paradigms include:
● Ordinary Differential Equations (ODEs): Used to model the dynamics of molecular concentrations (e.g., cytokines, signalling intermediates) based on reaction rates, useful for simulating signalling pathways or metabolic fluxes.
● Agent-Based Models (ABMs): Simulate the behaviour and interactions of individual agents (e.g., immune cells, epithelial cells, bacteria) within a spatial context, useful for understanding tissue-level phenomena like immune cell infiltration or granuloma formation.
● Boolean (Logical) Models: Represent components (e.g., genes, proteins) as being either ON or OFF and use logical rules to describe interactions. These are particularly useful for modelling signaling networks derived from curated knowledge maps and predicting pathway activation states under perturbation, as demonstrated extensively in immunological contexts (133).
In the context of IBD, computational models offer promising tools to simulate the progression from intestinal homeostasis to inflammation, predict the effects of dietary changes on microbiota composition and subsequent immune responses, test therapeutic interventions in silico, and identify dynamic molecular patterns that could serve as early biomarkers for disease flares (18, 123). Although the development of complex, multi-scale models for IBD remains challenging due to the inherent heterogeneity of the disease and the dynamic interactions among genetic, microbial, immunological, and environmental factors (124), such approaches represent critical steps toward building predictive frameworks for understanding disease progression and personalizing therapy. Importantly, computational models primarily generate hypotheses that must be rigorously validated through experimental studies and clinical trials to ensure biological relevance and translational utility (16, 17).
4.3 Nutritional epidemiology and dietary interventions in IBD
Nutritional epidemiology helps identify dietary risk factors and informs prevention strategies. Randomised controlled trials (RCTs) in nutrition are limited by cost, compliance, and long-term adherence. Therefore, prospective cohort studies and case–control designs are essential for assessing associations between dietary patterns and disease outcomes.
Recent large-scale studies, such as the PURE cohort and the Nurses’ Health Study, show that high UPF intake correlates with increased IBD risk, especially CD. UPFs are rich in synthetic additives, refined sugars, and poor in fibre and micronutrients, all of which affect microbiota composition and immune regulation (98).
Conversely, adherence to anti-inflammatory dietary patterns, particularly the Mediterranean diet, is associated with improved outcomes. Rich in polyphenols, fibre, and unsaturated fats, the Mediterranean diet promotes microbial diversity, supports SCFA production, and reduces intestinal inflammation. Butyrate, the most abundant SCFA, exerts anti-inflammatory effects by promoting the differentiation of Tregs in the colon, inhibiting histone deacetylases, and suppressing pro-inflammatory cytokine expression, thereby strengthening mucosal immune tolerance (134). Additionally, specific therapeutic diets such as the specific carbohydrate diet, low fermentable oligo-, di-, monosaccharides and polyols diet, and CD exclusion diet are being explored for their utility in symptom management and disease modulation (135). Recent studies also suggest that partial enteral nutrition combined with targeted dietary interventions, such as an anti-inflammatory diet or a modified CD exclusion diet, can achieve comparable rates of clinical and endoscopic remission to exclusive enteral nutrition in paediatric CD, while offering improved tolerability (136, 137).
The convergence of systems biology with nutritional epidemiology enables the design of personalised dietary interventions tailored to an individual’s microbiome composition, immune profile, and metabolic status. Advances in machine learning and computational modelling further enhance the ability to predict individual responses to specific dietary interventions, paving the way for precision nutrition in IBD care (138). Pilot studies suggest that gut microbiota profiling can predict individual dietary responsiveness, enabling the development of targeted precision nutrition interventions for IBD patients (138, 139).
Emerging evidence highlights the critical importance of early-life nutrition in programming the gut microbiome and immune system. Breastfeeding, maternal fibre intake, and low exposure to emulsifiers during infancy are associated with enhanced microbial diversity and a lower risk of inflammatory diseases, including IBD later in life (140, 141). These findings underscore the need for preventive strategies beginning in early childhood.
5 Conclusions
Over the past several decades, the global rise in IBD has paralleled profound shifts towards Westernized dietary and lifestyle patterns. While genetic predisposition contributes to IBD susceptibility, growing evidence indicates that environmental triggers primarily diet, alterations in the microbiota, and immune dysregulation, play pivotal roles in disease onset and progression. These complexities necessitate interdisciplinary research strategies that move beyond reductionist models and embrace integrative approaches based on systems biology and nutritional epidemiology.
Systems biology has emerged as a transformative tool in IBD research, enabling the integration of multi-omic datasets to uncover novel disease mechanisms, identify biomarkers for early diagnosis, and support the development of precision medicine strategies. By combining genomic, transcriptomic, metabolomic, and microbiome data, researchers can construct predictive models that capture the dynamic interplay between host and environmental factors. These integrative models hold promise for improving patient stratification, optimising treatment strategies, and identifying novel therapeutic targets aimed at restoring gut homeostasis. However, successful clinical implementation of systems biology approaches requires overcoming key challenges, including the standardisation of data collection protocols, integration of heterogeneous datasets, and rigorous validation of computational predictions through experimental and clinical studies. In parallel, nutritional epidemiology has offered critical insights into the impact of long-term dietary exposures on IBD risk and disease progression. The association between Western dietary patterns, characterised by high consumption of UPFs, refined sugars, unhealthy fats, and food additives, and increased IBD incidence highlights the urgent need for dietary interventions as both a preventive and therapeutic strategies. In contrast, emerging evidence supports the protective effects of anti-inflammatory dietary patterns, such as the Mediterranean diet, which favourably modulate gut microbiota composition, enhance intestinal barrier integrity, and reduce systemic inflammation.
5.1 Future prospective
Looking forward, the convergence of systems biology with precision nutrition represents a transformative frontier in IBD research and clinical practice. Advances in machine learning and artificial intelligence will enable the development of personalised dietary recommendations based on an individual’s genetic profile, microbiome composition, and immune status. Future research should priorities elucidating the mechanistic links between specific dietary components, microbiota modulation, and immune responses in IBD pathogenesis. In parallel, the identification of reliable, clinically relevant biomarkers capable of predicting dietary responsiveness will be crucial for the personalization of nutritional interventions.
Another crucial area of investigation involves the impact of early-life nutritional exposures, including maternal diet, breastfeeding practices, and early childhood nutrition, on microbiota development, immune system programming, and long-term disease susceptibility. Given the ubiquity of food additives, emulsifiers, and artificial sweeteners in modern diets, their potential role in disrupting gut barrier function and promoting intestinal inflammation demands urgent and focused investigation. Although significant progress has been made in elucidating the complex interactions between diet, microbiota, and immune responses in IBD, numerous critical questions remain unanswered.
Future research must embrace interdisciplinary collaborations, combining expertise across gastroenterology, immunology, microbiology, bioinformatics, and nutritional sciences to develop comprehensive, patient-centred approaches. By leveraging the full potential of systems biology and nutritional epidemiology, the goal is to shift the paradigm of IBD care, from reactive disease management to proactive, personalized prevention, ultimately improving long-term outcomes and quality of life for individuals affected by this complex condition. Establishing causality remains a key challenge, and future progress will require large-scale, well-controlled, longitudinal studies to generate robust, evidence-based dietary guidelines for individuals at risk of, or living with, IBD.
Author contributions
ŠK: Writing – original draft, Writing – review & editing. EB: Writing – original draft, Writing – review & editing. MŠ: Writing – original draft, Writing – review & editing. MV: Writing – original draft, Writing – review & editing. AŽ: Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Špela Konjar was funded by Marie Sklodowska Curie action individual fellowship (Project Nu. 101027263), Evgen Benedik was financed by the Slovenian Research and Innovation Agency (research core funding Nos. P3-0395), Marko Šestan was funded by grant of the University of Rijeka (uniri-iskusni-biomed-23-105), Marc Veldhoen was supported by “La Caixa” Foundation and Anže Županič was funded by the Slovenian research agency, under the grant JR-2550, JR-3097, PR-1065.
Acknowledgments
The authors would like to thank professor Primož Rožman, MD, PhD, Blood Transfusion Centre of Slovenia, Ljubljana for the helpful discussion and advices. Additional we would like to thank to Pedro Fernandes, Gulbenkian Science Emeritus Scholar Lisbon, Portugal for discussions about the role of system biology in IBD and for some good suggestions and Kaja Mlakar-Agrež, graphic and media designer, Litija, Slovenia for providing the drawing for Figures 3 and 4.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
IBD, Inflammatory Bowel Disease; NCD, Non-Communicable Diseases; CD, Crohn’s Disease; UC, Ulcerative Colitis; SCFAs, Short-Chain Fatty Acids; UPFs, Ultra-Processed Foods.
References
1. Christ A, Lauterbach M, and Latz E. Western diet and the immune system: an inflammatory connection. Immunity. (2019) 51:794–811. doi: 10.1016/j.immuni.2019.09.020
2. Aldars-Garcia L, Marin AC, Chaparro M, and Gisbert JP. The interplay between immune system and microbiota in inflammatory bowel disease: A narrative review. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms22063076
3. Levine JS and Burakoff R. Extraintestinal manifestations of inflammatory bowel disease. Gastroenterol hepatology. (2011) 7:235–41.
4. Roda G, Chien Ng S, Kotze PG, Argollo M, Panaccione R, Spinelli A, et al. Crohn's disease. Nat Rev Dis primers. (2020) 6:22. doi: 10.1038/s41572-020-0156-2
5. Khalili H, Chan SSM, Lochhead P, Ananthakrishnan AN, Hart AR, and Chan AT. The role of diet in the aetiopathogenesis of inflammatory bowel disease. Nat Rev Gastroenterol hepatology. (2018) 15:525–35. doi: 10.1038/s41575-018-0022-9
6. Chang JT. Pathophysiology of inflammatory bowel diseases. New Engl J Med. (2020) 383:2652–64. doi: 10.1056/NEJMra2002697
7. Lin L, Li Y, Zhou G, Wang Y, Li L, Han J, et al. Multi-omics analysis of western-style diet increased susceptibility to experimental colitis in mice. J Inflammation Res. (2022) 15:2523–37. doi: 10.2147/JIR.S361039
8. Vuijk SA, Camman AE, and de Ridder L. Considerations in paediatric and adolescent inflammatory bowel disease. J Crohn's colitis. (2024) 18:ii31–45. doi: 10.1093/ecco-jcc/jjae087
9. Chhibba T, Gros B, King JA, Windsor JW, Gorospe J, Leibovitzh H, et al. Environmental risk factors of inflammatory bowel disease: toward a strategy of preventative health. J Crohn's colitis. (2025) 19. doi: 10.1093/ecco-jcc/jjaf042
10. Ceballos D, Hernandez-Camba A, and Ramos L. Diet and microbiome in the beginning of the sequence of gut inflammation. World J Clin cases. (2021) 9:11122–47. doi: 10.12998/wjcc.v9.i36.11122
11. Adolph TE and Zhang J. Diet fuelling inflammatory bowel diseases: preclinical and clinical concepts. Gut. (2022) 71:2574–86. doi: 10.1136/gutjnl-2021-326575
12. Coward S, Clement F, Benchimol EI, Bernstein CN, Avina-Zubieta JA, Bitton A, et al. Past and future burden of inflammatory bowel diseases based on modeling of population-based data. Gastroenterology. (2019) 156:1345–53 e4. doi: 10.1053/j.gastro.2019.01.002
13. Kaplan GG and Windsor JW. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat Rev Gastroenterol hepatology. (2021) 18:56–66. doi: 10.1038/s41575-020-00360-x
14. Lavelle A and Sokol H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat Rev Gastroenterol hepatology. (2020) 17:223–37. doi: 10.1038/s41575-019-0258-z
15. Wang S, Zhou S, Han Z, Yu B, Xu Y, Lin Y, et al. From gut to brain: understanding the role of microbiota in inflammatory bowel disease. Front Immunol. (2024) 15:1384270. doi: 10.3389/fimmu.2024.1384270
16. Baumgart DC and Carding SR. Inflammatory bowel disease: cause and immunobiology. Lancet. (2007) 369:1627–40. doi: 10.1016/S0140-6736(07)60750-8
17. Lloyd-Price J, Arze C, Ananthakrishnan AN, Schirmer M, Avila-Pacheco J, Poon TW, et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature. (2019) 569:655–62. doi: 10.1038/s41586-019-1237-9
18. Fiocchi C. Omics and multi-omics in IBD: no integration, no breakthroughs. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms241914912
19. Martin JC, Chang C, Boschetti G, Ungaro R, Giri M, Grout JA, et al. Single-cell analysis of crohn's disease lesions identifies a pathogenic cellular module associated with resistance to anti-TNF therapy. Cell. (2019) 178:1493–508 e20. doi: 10.1016/j.cell.2019.08.008
20. Duchmann R, Kaiser I, Hermann E, Mayet W, Ewe K, and Meyer zum Buschenfelde KH. Tolerance exists towards resident intestinal flora but is broken in active inflammatory bowel disease (IBD). Clin Exp Immunol. (1995) 102:448–55. doi: 10.1111/j.1365-2249.1995.tb03836.x
21. Shan Y, Lee M, and Chang EB. The gut microbiome and inflammatory bowel diseases. Annu Rev Med. (2022) 73:455–68. doi: 10.1146/annurev-med-042320-021020
22. Subramanian A, J A, T T, Kumarasamy V, Begum MY, Sekar M, et al. Exploring the connections: autophagy, gut microbiota, and inflammatory bowel disease pathogenesis. J Inflammation Res. (2024) 17:10453–70. doi: 10.2147/JIR.S483958
23. Hegazy AN, West NR, Stubbington MJT, Wendt E, Suijker KIM, Datsi A, et al. Circulating and tissue-resident CD4(+) T cells with reactivity to intestinal microbiota are abundant in healthy individuals and function is altered during inflammation. Gastroenterology. (2017) 153:1320–37 e16. doi: 10.1053/j.gastro.2017.07.047
24. Saadawi A, Mair F, Rosenwald E, Hoces D, Slack E, and Kopf M. Investigating polyreactivity of CD4(+) T cells to the intestinal microbiota. Eur J Immunol. (2025) 55:e202451484. doi: 10.1002/eji.202451484
25. Subramanian A, Jahabardeen A, Thamaraikani T, and Vellapandian C. More on the interplay between gut microbiota, autophagy, and inflammatory bowel disease is needed. World J Gastroenterol. (2024) 30:3356–60. doi: 10.3748/wjg.v30.i27.3356
26. Larabi A, Barnich N, and Nguyen HTT. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy. (2020) 16:38–51. doi: 10.1080/15548627.2019.1635384
27. Didriksen BJ, Eshleman EM, and Alenghat T. Epithelial regulation of microbiota-immune cell dynamics. Mucosal Immunol. (2024) 17:303–13. doi: 10.1016/j.mucimm.2024.02.008
28. Joossens M, Huys G, Cnockaert M, De Preter V, Verbeke K, Rutgeerts P, et al. Dysbiosis of the faecal microbiota in patients with Crohn's disease and their unaffected relatives. Gut. (2011) 60:631–7. doi: 10.1136/gut.2010.223263
29. Tan C, Zheng Z, Wan X, Cao J, Wei R, and Duan J. The role of gut microbiota and amino metabolism in the effects of improvement of islet beta-cell function after modified jejunoileal bypass. Sci Rep. (2021) 11:4809. doi: 10.1038/s41598-021-84355-x
30. Kayama H, Okumura R, and Takeda K. Interaction between the microbiota, epithelia, and immune cells in the intestine. Annu Rev Immunol. (2020) 38:23–48. doi: 10.1146/annurev-immunol-070119-115104
31. Hou K, Wu ZX, Chen XY, Wang JQ, Zhang D, Xiao C, et al. Microbiota in health and diseases. Signal Transduct Target Ther. (2022) 7:135. doi: 10.1038/s41392-022-00974-4
32. Pascal V, Pozuelo M, Borruel N, Casellas F, Campos D, Santiago A, et al. A microbial signature for Crohn's disease. Gut. (2017) 66:813–22. doi: 10.1136/gutjnl-2016-313235
33. Kleessen B, Kroesen AJ, Buhr HJ, and Blaut M. Mucosal and invading bacteria in patients with inflammatory bowel disease compared with controls. Scandinavian J Gastroenterol. (2002) 37:1034–41. doi: 10.1080/003655202320378220
34. Machiels K, Joossens M, Sabino J, De Preter V, Arijs I, Eeckhaut V, et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut. (2014) 63:1275–83. doi: 10.1136/gutjnl-2013-304833
35. Vidal R, Ginard D, Khorrami S, Mora-Ruiz M, Munoz R, Hermoso M, et al. Crohn associated microbial communities associated to colonic mucosal biopsies in patients of the western Mediterranean. Systematic Appl Microbiol. (2015) 38:442–52. doi: 10.1016/j.syapm.2015.06.008
36. Pedamallu CS, Bhatt AS, Bullman S, Fowler S, Freeman SS, Durand J, et al. Metagenomic characterization of microbial communities in situ within the deeper layers of the ileum in crohn's disease. Cell Mol Gastroenterol hepatology. (2016) 2:563–6 e5. doi: 10.1016/j.jcmgh.2016.05.011
37. Khan I, Ullah N, Zha L, Bai Y, Khan A, Zhao T, et al. Alteration of gut microbiota in inflammatory bowel disease (IBD): cause or consequence? IBD treatment targeting the gut microbiome. Pathogens. (2019) 8. doi: 10.3390/pathogens8030126
38. Pittayanon R, Lau JT, Leontiadis GI, Tse F, Yuan Y, Surette M, et al. Differences in gut microbiota in patients with vs without inflammatory bowel diseases: A systematic review. Gastroenterology. (2020) 158:930–46 e1. doi: 10.1053/j.gastro.2019.11.294
39. Clooney AG, Eckenberger J, Laserna-Mendieta E, Sexton KA, Bernstein MT, Vagianos K, et al. Ranking microbiome variance in inflammatory bowel disease: a large longitudinal intercontinental study. Gut. (2021) 70:499–510. doi: 10.1136/gutjnl-2020-321106
40. Madsen KL, Doyle JS, Tavernini MM, Jewell LD, Rennie RP, and Fedorak RN. Antibiotic therapy attenuates colitis in interleukin 10 gene-deficient mice. Gastroenterology. (2000) 118:1094–105. doi: 10.1016/S0016-5085(00)70362-3
41. Baumgart DC and Sandborn WJ. Inflammatory bowel disease: clinical aspects and established and evolving therapies. Lancet. (2007) 369:1641–57. doi: 10.1016/S0140-6736(07)60751-X
42. Yan H and Ajuwon KM. Butyrate modifies intestinal barrier function in IPEC-J2 cells through a selective upregulation of tight junction proteins and activation of the Akt signaling pathway. PloS One. (2017) 12:e0179586. doi: 10.1371/journal.pone.0179586
43. Yan D, Ye S, He Y, Wang S, Xiao Y, Xiang X, et al. Fatty acids and lipid mediators in inflammatory bowel disease: from mechanism to treatment. Front Immunol. (2023) 14:1286667. doi: 10.3389/fimmu.2023.1286667
44. Yoo W, Zieba JK, Foegeding NJ, Torres TP, Shelton CD, Shealy NG, et al. High-fat diet-induced colonocyte dysfunction escalates microbiota-derived trimethylamine N-oxide. Science. (2021) 373:813–8. doi: 10.1126/science.aba3683
45. Ijssennagger N, van der Meer R, and van Mil SWC. Sulfide as a mucus barrier-breaker in inflammatory bowel disease? Trends Mol Med. (2016) 22:190–9. doi: 10.1016/j.molmed.2016.01.002
46. Chen ML, Zhu XH, Ran L, Lang HD, Yi L, and Mi MT. Trimethylamine-N-oxide induces vascular inflammation by activating the NLRP3 inflammasome through the SIRT3-SOD2-mtROS signaling pathway. J Am Heart Assoc. (2017) 6. doi: 10.1161/JAHA.117.006347
47. Caruso R, Lo BC, Chen GY, and Nunez G. Host-pathobiont interactions in Crohn's disease. Nat Rev Gastroenterol hepatology. (2025) 22:395–414. doi: 10.1038/s41575-024-00997-y
48. Shin Y, Han S, Kwon J, Ju S, Choi TG, Kang I, et al. Roles of short-chain fatty acids in inflammatory bowel disease. Nutrients. (2023) 15. doi: 10.3390/nu15204466
49. Cuzic S, Antolic M, Ognjenovic A, Stupin-Polancec D, Petrinic Grba A, Hrvacic B, et al. Claudins: beyond tight junctions in human IBD and murine models. Front Pharmacol. (2021) 12:682614. doi: 10.3389/fphar.2021.682614
50. Mehandru S and Colombel JF. The intestinal barrier, an arbitrator turned provocateur in IBD. Nat Rev Gastroenterol hepatology. (2021) 18:83–4. doi: 10.1038/s41575-020-00399-w
51. Paik J, Fierce Y, Treuting PM, Brabb T, and Maggio-Price L. High-fat diet-induced obesity exacerbates inflammatory bowel disease in genetically susceptible Mdr1a-/- male mice. J Nutr. (2013) 143:1240–7. doi: 10.3945/jn.113.174615
52. Lee JC, Lee HY, Kim TK, Kim MS, Park YM, Kim J, et al. Obesogenic diet-induced gut barrier dysfunction and pathobiont expansion aggravate experimental colitis. PloS One. (2017) 12:e0187515. doi: 10.1371/journal.pone.0187515
53. Chassaing B, Koren O, Goodrich JK, Poole AC, Srinivasan S, Ley RE, et al. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature. (2015) 519:92–6. doi: 10.1038/nature14232
54. Na YR, Stakenborg M, Seok SH, and Matteoli G. Macrophages in intestinal inflammation and resolution: a potential therapeutic target in IBD. Nat Rev Gastroenterol hepatology. (2019) 16:531–43. doi: 10.1038/s41575-019-0172-4
55. Gulhane M, Murray L, Lourie R, Tong H, Sheng YH, Wang R, et al. High fat diets induce colonic epithelial cell stress and inflammation that is reversed by IL-22. Sci Rep. (2016) 6:28990. doi: 10.1038/srep28990
56. Garrido-Trigo A, Corraliza AM, Veny M, Dotti I, Melon-Ardanaz E, Rill A, et al. Macrophage and neutrophil heterogeneity at single-cell spatial resolution in human inflammatory bowel disease. Nat Commun. (2023) 14:4506. doi: 10.1038/s41467-023-40156-6
57. Prame Kumar K, Ooi JD, and Goldberg R. The interplay between the microbiota, diet and T regulatory cells in the preservation of the gut barrier in inflammatory bowel disease. Front Microbiol. (2023) 14:1291724. doi: 10.3389/fmicb.2023.1291724
58. Chen Y, Cui W, Li X, and Yang H. Interaction between commensal bacteria, immune response and the intestinal barrier in inflammatory bowel disease. Front Immunol. (2021) 12:761981. doi: 10.3389/fimmu.2021.761981
59. Cao H, Diao J, Liu H, Liu S, Liu J, Yuan J, et al. The pathogenicity and synergistic action of th1 and th17 cells in inflammatory bowel diseases. Inflammatory bowel diseases. (2023) 29:818–29. doi: 10.1093/ibd/izac199
60. Hu Y, Tang J, Xie Y, Xu W, Zhu W, Xia L, et al. Gegen Qinlian decoction ameliorates TNBS-induced ulcerative colitis by regulating Th2/Th1 and Tregs/Th17 cells balance, inhibiting NLRP3 inflammasome activation and reshaping gut microbiota. J Ethnopharmacol. (2024) 328:117956. doi: 10.1016/j.jep.2024.117956
61. Wang Y, Liu XP, Zhao ZB, Chen JH, and Yu CG. Expression of CD4+ forkhead box P3 (FOXP3)+ regulatory T cells in inflammatory bowel disease. J digestive diseases. (2011) 12:286–94. doi: 10.1111/j.1751-2980.2011.00505.x
62. Chaudhry A, Samstein RM, Treuting P, Liang Y, Pils MC, Heinrich JM, et al. Interleukin-10 signaling in regulatory T cells is required for suppression of Th17 cell-mediated inflammation. Immunity. (2011) 34:566–78. doi: 10.1016/j.immuni.2011.03.018
63. Van NT, Zhang K, Wigmore RM, Kennedy AI, DaSilva CR, Huang J, et al. Dietary L-Tryptophan consumption determines the number of colonic regulatory T cells and susceptibility to colitis via GPR15. Nat Commun. (2023) 14:7363. doi: 10.1038/s41467-023-43211-4
64. Voskens C, Stoica D, Rosenberg M, Vitali F, Zundler S, Ganslmayer M, et al. Autologous regulatory T-cell transfer in refractory ulcerative colitis with concomitant primary sclerosing cholangitis. Gut. (2023) 72:49–53. doi: 10.1136/gutjnl-2022-327075
65. Yoo JY, Groer M, Dutra SVO, Sarkar A, and McSkimming DI. Gut microbiota and immune system interactions. Microorganisms. (2020) 8. doi: 10.3390/microorganisms8101587
66. Di Sabatino A, Santacroce G, Rossi CM, Broglio G, and Lenti MV. Role of mucosal immunity and epithelial-vascular barrier in modulating gut homeostasis. Intern Emerg Med. (2023) 18:1635–46. doi: 10.1007/s11739-023-03329-1
67. Rio P, Gasbarrini A, Gambassi G, and Cianci R. Pollutants, microbiota and immune system: frenemies within the gut. Front Public Health. (2024) 12:1285186. doi: 10.3389/fpubh.2024.1285186
68. Barber TM, Hanson P, Kabisch S, Pfeiffer AFH, and Weickert MO. The low-carbohydrate diet: short-term metabolic efficacy versus longer-term limitations. Nutrients. (2021) 13. doi: 10.3390/nu13041187
69. Isaksen IM and Dankel SN. Ultra-processed food consumption and cancer risk: A systematic review and meta-analysis. Clin Nutr. (2023) 42:919–28. doi: 10.1016/j.clnu.2023.03.018
70. Ma H, Wang X, Li X, Heianza Y, and Qi L. Adding salt to foods and risk of cardiovascular disease. J Am Coll Cardiol. (2022) 80:2157–67. doi: 10.1016/j.jacc.2022.09.039
71. Cheng H, Zhou J, Sun Y, Zhan Q, and Zhang D. High fructose diet: A risk factor for immune system dysregulation. Hum Immunol. (2022) 83:538–46. doi: 10.1016/j.humimm.2022.03.007
72. Las Heras V, Melgar S, MacSharry J, and Gahan CGM. The influence of the western diet on microbiota and gastrointestinal immunity. Annu Rev Food Sci technology. (2022) 13:489–512. doi: 10.1146/annurev-food-052720-011032
73. Garcia-Montero C, Fraile-Martinez O, Gomez-Lahoz AM, Pekarek L, Castellanos AJ, Noguerales-Fraguas F, et al. Nutritional components in western diet versus mediterranean diet at the gut microbiota-immune system interplay. Implications for health and disease. Nutrients. (2021) 13. doi: 10.3390/nu13020699
74. Ramne S, Brunkwall L, Ericson U, Gray N, Kuhnle GGC, Nilsson PM, et al. Gut microbiota composition in relation to intake of added sugar, sugar-sweetened beverages and artificially sweetened beverages in the Malmo Offspring Study. Eur J Nutr. (2021) 60:2087–97. doi: 10.1007/s00394-020-02392-0
75. Jensen T, Abdelmalek MF, Sullivan S, Nadeau KJ, Green M, Roncal C, et al. Fructose and sugar: A major mediator of non-alcoholic fatty liver disease. J hepatology. (2018) 68:1063–75. doi: 10.1016/j.jhep.2018.01.019
76. Khan S, Waliullah S, Godfrey V, Khan MAW, Ramachandran RA, Cantarel BL, et al. Dietary simple sugars alter microbial ecology in the gut and promote colitis in mice. Sci Transl Med. (2020) 12. doi: 10.1126/scitranslmed.aay6218
77. Statovci D, Aguilera M, MacSharry J, and Melgar S. The impact of western diet and nutrients on the microbiota and immune response at mucosal interfaces. Front Immunol. (2017) 8:838. doi: 10.3389/fimmu.2017.00838
78. Zhou W, Ramachandran D, Mansouri A, and Dailey MJ. Glucose stimulates intestinal epithelial crypt proliferation by modulating cellular energy metabolism. J Cell Physiol. (2018) 233:3465–75. doi: 10.1002/jcp.26199
79. Gillingham LG, Harris-Janz S, and Jones PJ. Dietary monounsaturated fatty acids are protective against metabolic syndrome and cardiovascular disease risk factors. Lipids. (2011) 46:209–28. doi: 10.1007/s11745-010-3524-y
80. Shi F, Chowdhury R, Sofianopoulou E, Koulman A, Sun L, Steur M, et al. Association of circulating fatty acids with cardiovascular disease risk: analysis of individual-level data in three large prospective cohorts and updated meta-analysis. Eur J Prev Cardiol. (2025) 32:233–46. doi: 10.1093/eurjpc/zwae315
81. Waldman HS, Krings BM, Smith JW, and McAllister MJ. A shift toward a high-fat diet in the current metabolic paradigm: A new perspective. Nutrition. (2018) 46:33–5. doi: 10.1016/j.nut.2017.08.014
82. Ye L, Mueller O, Bagwell J, Bagnat M, Liddle RA, and Rawls JF. High fat diet induces microbiota-dependent silencing of enteroendocrine cells. eLife. (2019) 8. doi: 10.7554/eLife.48479
83. Netto Candido TL, Bressan J, and Alfenas RCG. Dysbiosis and metabolic endotoxemia induced by high-fat diet. Nutricion hospitalaria. (2018) 35:1432–40. doi: 10.20960/nh.1792
84. Sadeghi R, Norouzzadeh M, HasanRashedi M, Jamshidi S, Ahmadirad H, Alemrajabi M, et al. Dietary and circulating omega-6 fatty acids and their impact on cardiovascular disease, cancer risk, and mortality: a global meta-analysis of 150 cohorts and meta-regression. J Transl Med. (2025) 23:314. doi: 10.1186/s12967-025-06336-2
85. DiNicolantonio JJ and O'Keefe JH. Omega-6 vegetable oils as a driver of coronary heart disease: the oxidized linoleic acid hypothesis. Open Heart. (2018) 5:e000898. doi: 10.1136/openhrt-2018-000898
86. Kaliannan K, Wang B, Li XY, Kim KJ, and Kang JX. A host-microbiome interaction mediates the opposing effects of omega-6 and omega-3 fatty acids on metabolic endotoxemia. Sci Rep. (2015) 5:11276. doi: 10.1038/srep11276
87. Kadandale S, Marten R, and Smith R. The palm oil industry and noncommunicable diseases. Bull World Health Organization. (2019) 97:118–28. doi: 10.2471/BLT.18.220434
88. Martinez N, Prieto I, Hidalgo M, Segarra AB, Martinez-Rodriguez AM, Cobo A, et al. Refined versus extra virgin olive oil high-fat diet impact on intestinal microbiota of mice and its relation to different physiological variables. Microorganisms. (2019) 7. doi: 10.3390/microorganisms7020061
89. Ghezzal S, Postal BG, Quevrain E, Brot L, Seksik P, Leturque A, et al. Palmitic acid damages gut epithelium integrity and initiates inflammatory cytokine production. Biochim Biophys Acta Mol Cell Biol lipids. (2020) 1865:158530. doi: 10.1016/j.bbalip.2019.158530
90. Rodriguez-Garcia C, Sanchez-Quesada C, Algarra I, and Gaforio JJ. The high-fat diet based on extra-virgin olive oil causes dysbiosis linked to colorectal cancer prevention. Nutrients. (2020) 12. doi: 10.3390/nu12061705
91. Okamura T, Hashimoto Y, Majima S, Senmaru T, Ushigome E, Nakanishi N, et al. Trans fatty acid intake induces intestinal inflammation and impaired glucose tolerance. Front Immunol. (2021) 12:669672. doi: 10.3389/fimmu.2021.669672
92. Banno Y, Nomura M, Hara R, Asami M, Tanaka K, Mukai Y, et al. Trimethylamine N-oxide and risk of inflammatory bowel disease: A Mendelian randomization study. Med (Baltimore). (2023) 102:e34758. doi: 10.1097/MD.0000000000034758
93. Kul S, Caliskan Z, Guvenc TS, Celik FB, Sarmis A, Atici A, et al. Gut microbiota-derived metabolite trimethylamine N-oxide and biomarkers of inflammation are linked to endothelial and coronary microvascular function in patients with inflammatory bowel disease. Microvasc Res. (2023) 146:104458. doi: 10.1016/j.mvr.2022.104458
94. Suez J, Korem T, Zilberman-Schapira G, Segal E, and Elinav E. Non-caloric artificial sweeteners and the microbiome: findings and challenges. Gut Microbes. (2015) 6:149–55. doi: 10.1080/19490976.2015.1017700
95. Teodorowicz M, van Neerven J, and Savelkoul H. Food processing: the influence of the maillard reaction on immunogenicity and allergenicity of food proteins. Nutrients. (2017) 9. doi: 10.3390/nu9080835
96. Monteiro CA, Cannon G, Levy RB, Moubarac JC, Louzada ML, Rauber F, et al. Ultra-processed foods: what they are and how to identify them. Public Health Nutr. (2019) 22:936–41. doi: 10.1017/S1368980018003762
97. Nie C, Li Y, Qian H, Ying H, and Wang L. Advanced glycation end products in food and their effects on intestinal tract. Crit Rev Food Sci Nutr. (2022) 62:3103–15. doi: 10.1080/10408398.2020.1863904
98. Narula N, Wong ECL, Dehghan M, Mente A, Rangarajan S, Lanas F, et al. Association of ultra-processed food intake with risk of inflammatory bowel disease: prospective cohort study. Bmj. (2021) 374:n1554. doi: 10.1136/bmj.n1554
99. Srour B, Kordahi MC, Bonazzi E, Deschasaux-Tanguy M, Touvier M, and Chassaing B. Ultra-processed foods and human health: from epidemiological evidence to mechanistic insights. Lancet Gastroenterol hepatology. (2022) 7:1128–40. doi: 10.1016/S2468-1253(22)00169-8
100. Basson AR, Katz J, Singh S, Celio F, Cominelli F, and Rodriguez-Palacios A. Sweets and inflammatory bowel disease: patients favor artificial sweeteners and diet foods/drinks over table sugar and consume less fruits/vegetables. Inflammatory bowel Dis. (2023) 29:1751–9. doi: 10.1093/ibd/izac272
101. de Oliveira PG, de Sousa JM, Assuncao DGF, de Araujo EKS, Bezerra DS, Dametto J, et al. Impacts of consumption of ultra-processed foods on the maternal-child health: A systematic review. Front Nutr. (2022) 9:821657. doi: 10.3389/fnut.2022.821657
102. Narula N, Chang NH, Mohammad D, Wong ECL, Ananthakrishnan AN, Chan SSM, et al. Food processing and risk of inflammatory bowel disease: A systematic review and meta-analysis. Clin Gastroenterol hepatology: Off Clin Pract J Am Gastroenterological Assoc. (2023) 21:2483–95 e1. doi: 10.1016/j.cgh.2023.01.012
103. Bischoff SC, Bager P, Escher J, Forbes A, Hebuterne X, Hvas CL, et al. ESPEN guideline on Clinical Nutrition in inflammatory bowel disease. Clin Nutr. (2023) 42:352–79. doi: 10.1016/j.clnu.2022.12.004
104. Godny L, Svolos V, Williams AJ, Czuber-Dochan W, Aloi M, Ibarra A, et al. Multidisciplinary perinatal care in IBD. J Crohn's colitis. (2023) 17:663–80. doi: 10.1093/ecco-jcc/jjac189
105. Halmos EP, Godny L, Vanderstappen J, Sarbagili-Shabat C, and Svolos V. Role of diet in prevention versus treatment of Crohn's disease and ulcerative colitis. Frontline Gastroenterol. (2024) 15:247–57. doi: 10.1136/flgastro-2023-102417
106. Newsome R, Yang Y, and Jobin C. Western diet influences on microbiome and carcinogenesis. Semin Immunol. (2023) 67:101756. doi: 10.1016/j.smim.2023.101756
107. He FJ, Jenner KH, and Macgregor GA. WASH-world action on salt and health. Kidney Int. (2010) 78:745–53. doi: 10.1038/ki.2010.280
108. Rizvi ZA, Dalal R, Sadhu S, Kumar Y, Kumar S, Gupta SK, et al. High-salt diet mediates interplay between NK cells and gut microbiota to induce potent tumor immunity. Sci Adv. (2021) 7:eabg5016. doi: 10.1126/sciadv.abg5016
109. Wilck N, Matus MG, Kearney SM, Olesen SW, Forslund K, Bartolomaeus H, et al. Salt-responsive gut commensal modulates T(H)17 axis and disease. Nature. (2017) 551:585–9. doi: 10.1038/nature24628
110. Brucklacher-Waldert V, Ferreira C, Stebegg M, Fesneau O, Innocentin S, Marie JC, et al. Cellular stress in the context of an inflammatory environment supports TGF-beta-independent T helper-17 differentiation. Cell Rep. (2017) 19:2357–70. doi: 10.1016/j.celrep.2017.05.052
111. Zhou L and Littman DR. Transcriptional regulatory networks in Th17 cell differentiation. Curr Opin Immunol. (2009) 21:146–52. doi: 10.1016/j.coi.2009.03.001
112. Wu C, Yosef N, Thalhamer T, Zhu C, Xiao S, Kishi Y, et al. Induction of pathogenic TH17 cells by inducible salt-sensing kinase SGK1. Nature. (2013) 496:513–7. doi: 10.1038/nature11984
113. Kuang R, O'Keefe SJD, Ramos Del Aguila de Rivers C, Koutroumpakis F, and Binion DG. Is salt at fault? Dietary salt consumption and inflammatory bowel disease. Inflammatory bowel diseases. (2023) 29:140–50. doi: 10.1093/ibd/izac058
114. Zhang F, Aschenbrenner D, Yoo JY, and Zuo T. The gut mycobiome in health, disease, and clinical applications in association with the gut bacterial microbiome assembly. Lancet Microbe. (2022) 3:e969–e83. doi: 10.1016/S2666-5247(22)00203-8
115. Maioli TU, Trindade LM, Souza A, Torres L, Andrade MER, Cardoso VN, et al. Non-pharmacologic strategies for the management of intestinal inflammation. BioMed Pharmacother. (2022) 145:112414. doi: 10.1016/j.biopha.2021.112414
116. Silva YP, Bernardi A, and Frozza RL. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol (Lausanne). (2020) 11:25. doi: 10.3389/fendo.2020.00025
117. Shapiro H, Singer P, Halpern Z, and Bruck R. Polyphenols in the treatment of inflammatory bowel disease and acute pancreatitis. Gut. (2007) 56:426–35. doi: 10.1136/gut.2006.094599
118. Arifuzzaman M, Collins N, Guo CJ, and Artis D. Nutritional regulation of microbiota-derived metabolites: Implications for immunity and inflammation. Immunity. (2024) 57:14–27. doi: 10.1016/j.immuni.2023.12.009
119. Ni J, Wu GD, Albenberg L, and Tomov VT. Gut microbiota and IBD: causation or correlation? Nat Rev Gastroenterol Hepatol. (2017) 14:573–84. doi: 10.1038/nrgastro.2017.88
120. Borren NZ, Plichta D, Joshi AD, Bonilla G, Sadreyev R, Vlamakis H, et al. Multi-"-omics" Profiling in patients with quiescent inflammatory bowel disease identifies biomarkers predicting relapse. Inflammatory bowel diseases. (2020) 26:1524–32. doi: 10.1093/ibd/izaa183
121. Janker L, Schuster D, Bortel P, Hagn G, Meier-Menches SM, Mohr T, et al. Multiomics-empowered deep phenotyping of ulcerative colitis identifies biomarker signatures reporting functional remission states. J Crohn's colitis. (2023) 17:1514–27. doi: 10.1093/ecco-jcc/jjad052
122. Howitt MR, Cao YG, Gologorsky MB, Li JA, Haber AL, Biton M, et al. The taste receptor TAS1R3 regulates small intestinal tuft cell homeostasis. Immunohorizons. (2020) 4:23–32. doi: 10.4049/immunohorizons.1900099
123. Fiocchi C and Iliopoulos D. IBD systems biology is here to stay. Inflammatory bowel diseases. (2021) 27:760–70. doi: 10.1093/ibd/izaa343
124. Karczewski KJ and Snyder MP. Integrative omics for health and disease. Nat Rev Genet. (2018) 19:299–310. doi: 10.1038/nrg.2018.4
125. Danese S, Vuitton L, and Peyrin-Biroulet L. Biologic agents for IBD: practical insights. Nat Rev Gastroenterol hepatology. (2015) 12:537–45. doi: 10.1038/nrgastro.2015.135
126. Lee JWJ, Plichta D, Hogstrom L, Borren NZ, Lau H, Gregory SM, et al. Multi-omics reveal microbial determinants impacting responses to biologic therapies in inflammatory bowel disease. Cell Host Microbe. (2021) 29:1294–304 e4. doi: 10.1016/j.chom.2021.06.019
127. Magro F, Rodrigues-Pinto E, Santos-Antunes J, Vilas-Boas F, Lopes S, Nunes A, et al. High C-reactive protein in Crohn's disease patients predicts nonresponse to infliximab treatment. J Crohn's colitis. (2014) 8:129–36. doi: 10.1016/j.crohns.2013.07.005
128. Zagorscak M, Abdelhakim L, Rodriguez-Granados NY, Siroka J, Ghatak A, Bleker C, et al. Integration of multi-omics data and deep phenotyping provides insights into responses to single and combined abiotic stress in potato. Plant Physiol. (2025) 197. doi: 10.1093/plphys/kiaf126
129. Argelaguet R, Velten B, Arnol D, Dietrich S, Zenz T, Marioni JC, et al. Multi-Omics Factor Analysis-a framework for unsupervised integration of multi-omics data sets. Mol Syst Biol. (2018) 14:e8124. doi: 10.15252/msb.20178124
130. Ostaszewski M, Gebel S, Kuperstein I, Mazein A, Zinovyev A, Dogrusoz U, et al. Community-driven roadmap for integrated disease maps. Brief Bioinform. (2019) 20:659–70. doi: 10.1093/bib/bby024
131. Gawron P, Ostaszewski M, Satagopam V, Gebel S, Mazein A, Kuzma M, et al. MINERVA-a platform for visualization and curation of molecular interaction networks. NPJ Syst Biol Appl. (2016) 2:16020. doi: 10.1038/npjsba.2016.20
132. Ostaszewski M, Niarakis A, Mazein A, Kuperstein I, Phair R, Orta-Resendiz A, et al. COVID-19 Disease Map, a computational knowledge repository of virus-host interaction mechanisms. Mol Syst Biol. (2021) 17:e10851. doi: 10.15252/msb.202110851
133. Laubenbacher R, Niarakis A, Helikar T, An G, Shapiro B, Malik-Sheriff RS, et al. Building digital twins of the human immune system: toward a roadmap. NPJ Digit Med. (2022) 5:64.
134. Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G, Takahashi D, et al. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature. (2013) 504:446–50. doi: 10.1038/nature12721
135. Li F, Armet AM, Korpela K, Liu J, Quevedo RM, Asnicar F, et al. Cardiometabolic benefits of a non-industrialized-type diet are linked to gut microbiome modulation. Cell. (2025) 188:1226–47 e18. doi: 10.1016/j.cell.2024.12.034
136. Urlep D, Benedik E, Brecelj J, and Orel R. Partial enteral nutrition induces clinical and endoscopic remission in active pediatric Crohn's disease: results of a prospective cohort study. Eur J Pediatr. (2020) 179:431–8. doi: 10.1007/s00431-019-03520-7
137. Urlep D, Orel R, Kunstek P, and Benedik E. Treatment of active crohn's disease in children using partial enteral nutrition combined with a modified crohn's disease exclusion diet: A pilot prospective cohort trial on clinical and endoscopic outcomes. Nutrients. (2023) 15. doi: 10.3390/nu15214676
138. Zeevi D, Korem T, Zmora N, Israeli D, Rothschild D, Weinberger A, et al. Personalized nutrition by prediction of glycemic responses. Cell. (2015) 163:1079–94. doi: 10.1016/j.cell.2015.11.001
139. Nieva C, Pryor J, Williams GM, Hoedt EC, Burns GL, Eslick GD, et al. The impact of dietary interventions on the microbiota in inflammatory bowel disease: A systematic review. J Crohn's colitis. (2024) 18:920–42. doi: 10.1093/ecco-jcc/jjad204
140. Nunez H, Nieto PA, Mars RA, Ghavami M, Sew Hoy C, and Sukhum K. Early life gut microbiome and its impact on childhood health and chronic conditions. Gut Microbes. (2025) 17:2463567. doi: 10.1080/19490976.2025.2463567
Keywords: IBD, western diet, immune cells, microbiota, systems biology
Citation: Konjar Š, Benedik E, Šestan M, Veldhoen M and Županič A (2025) Systems biology to unravel Western diet-associated triggers in inflammatory bowel disease. Front. Immunol. 16:1621334. doi: 10.3389/fimmu.2025.1621334
Received: 30 April 2025; Accepted: 17 September 2025;
Published: 10 October 2025.
Edited by:
Shahanshah Khan, University of Texas Southwestern Medical Center, United StatesReviewed by:
Arunkumar Subramanianm, Thanthai Roever College of Pharmacy, IndiaRahat Ali, Jamia Millia Islamia, India
Laura Ramos López, Canary Islands University Hospital, Spain
Copyright © 2025 Konjar, Benedik, Šestan, Veldhoen and Županič. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Špela Konjar, c3BlbGFfa29uamFyQHp0bS5zaQ==
†ORCID: Špela Konjar, orcid.org/0000-0002-8969-0233
Evgen Benedik, orcid.org/0000-0002-6954-1118
Marko Šestan, orcid.org/0000-0002-1473-6647
Marc Veldhoen, orcid.org/0000-0002-1478-9562
Anže Županič, orcid.org/0000-0003-3303-9086