 Ane Otaegui-Chivite1,2
Ane Otaegui-Chivite1,2 Miriam Gorostidi-Aicua1,2‡Laura Martins-Almeida1,2Ainhoa Alberro1,2‡Leire Romarate1‡Idoia Mendiburu1,3Amaya Álvarez de Arcaya4Maialen Arruti1,3Tamara Castillo-Triviño1,2,3
Miriam Gorostidi-Aicua1,2‡Laura Martins-Almeida1,2Ainhoa Alberro1,2‡Leire Romarate1‡Idoia Mendiburu1,3Amaya Álvarez de Arcaya4Maialen Arruti1,3Tamara Castillo-Triviño1,2,3 David Otaegui1,2*†
David Otaegui1,2*† Laura Moles1,2*†
Laura Moles1,2*†- 1Group of Neuroimmunilogy, Biogipuzkoa Health Research Institute, San Sebastián, Spain
- 2Center for Biomedical Research Network in Neurodegenerative Diseases (CIBER-CIBERNED-ISCIII), Madrid, Spain
- 3Neurology Department, Hospital Universitario Donostia, Osakidetza Basque Health Service, San Sebastián, Spain
- 4Neurology Department, Hospital Universitario Araba, Osakidetza Basque Health Service, Vitoria, Spain
Background: Multiple sclerosis (MS) is a complex immune-mediated disorder influenced by genetic, environmental, and microbial factors. Recent research has focused on the gut microbiota’s role in MS, yet limited studies have examined the fungal microbiota (mycobiota) in this context.
Methods: In this study, we characterized the gut mycobiota of individuals with MS from the Basque Country, identifying specific fungal taxa associated with MS risk factors, clinical severity, and dietary patterns.
Results: Our results revealed higher fungal diversity and richness in MS patients compared to controls, with significant enrichment of certain genera, including Saccharomyces, Torulaspora, and Malassezia. Malassezia demonstrated a strong association with increased disability, aligning with its previous identification in neurodegenerative conditions. Furthermore, we found that the presence of the MS-associated allele HLA-DRB1*1501 significantly influenced mycobiota composition and correlated with disability metrics. Additionally, we observed a complex interplay between plasma biomarkers (chitotriosidase and calprotectin) and specific fungal groups, with disease-specific correlations indicating potential interactions between the immune response and gut fungi. Notably, dietary fats showed a greater impact on mycobiota composition in MS patients than in controls, highlighting altered lipid metabolism in MS.
Conclusion: These findings provide new insights into the fungal component of the gut microbiota in MS and underscore its potential role in disease pathogenesis and progression. Our work suggests that fungal biomarkers, together with genetic and dietary factors, may help refine our understanding of MS and support the development of mycobiota-targeted therapies.
1 Introduction
Multiple Sclerosis (MS) is a complex, chronic and immune-mediated disease characterized by inflammation, demyelination and progressive neurodegeneration. It arises from an immune response against self-antigens in individuals with a genetic predisposition (1). MS primarily affects young adults, with women being three times more likely to develop the disease than men (2). While the etiology of MS remain unclear, genetic factors are believed to account for approximately one-third of the disease risk (3). Environmental factors and lifestyle choices are also recognized as additional contributors on MS susceptibility. In the last decade, significant emphasis has been placed on studying the gut microbiota in the context of autoimmunity and neurodegeneration. Several published research studies report an altered gut microbiota in individuals with MS (4, 5), indicating a role in the pathogenesis of the disease. These studies evaluate the influence of the gut microbiota on immune responses and its potential contribution to the development and progression of MS. In fact, changes in microbial populations are thought to affect the function of the immune system, the formation of chronic inflammation and the integrity of barriers, all of them implicated in the MS pathogenesis (6). While most studies today are focused on the bacterial component of the gut microbiome, relatively little is known about another fundamental part, the mycobiota (the fungal community in the gut), with potential to influence on both, disease progression and immune function. Many links between fungi and diseases involving chronic inflammation have been found recently. Dysbiosis of the fungal communities has been observed in inflammatory bowel syndrome (IBD) (7), with studies highlighting the impact of these alterations on the inflammatory response (8) and the identification of potential fungal markers (9). In the context of MS, early indications of fungal involvement were based on findings of elevated serological antibody titers specific to Candida in individuals with MS compared to controls (10, 11). Besides, the presence of specific enzyme activity of Candida albicans correlate with disease severity (12). DNA from different fungal species has been detected in the CNS of individuals with MS (13, 14), suggesting a fungal infection in cerebral tissue. Moreover, systemic infections by non-albicans Candida strains have been shown to influence both the immunological and clinical aspects of experimental autoimmune encephalomyelitis (EAE), a model of MS, further suggesting their possible relevance in MS development (15, 16).
Previous studies describe the human mycobiota as dynamic and variable between individuals. At least 75 genera and 267 species have been identified in the human gut (17, 18). Most studies agree on the dominance of the phyla Ascomycota and Basidiomycota, with smaller proportions of Mucoromycota and Zoopagomycota. Among these, three genera, Candida, Saccharomyces and Aspergillus, are considered the most prominent (10, 17, 19). However, like the microbiota, there is no consensus on what constitute a “healthy” mycobiota (17). Despite this lack of consensus, numerous publications associate dysbiosis of the intestinal mycobiota with various diseases, particularly those linked to chronic inflammation and neurological disorders, such as obesity, IBD, autism, schizophrenia, and Alzheimer’s disease (20–23).
The role of infections in MS remains a subject of active debated, as definite proof of autoimmunity is still lacking. Various pathogens have been associated with the development or exacerbation of MS, including bacteria such as Chlamydia pneumonia and endotoxins produced by Staphylococcus aureus, which act as superantigens. Viruses from the Herpesviridae family, including Epstein–Barr virus (EBV) and human herpes virus 6 (HHV-6), as well as human endogenous retroviruses, have also been implicated (24). Among them, EBV stands out as having the strongest link to MS. EBV infects approximately 90% of the general population during the first decade of life and remains in the body as a latent infection, primarily within memory B cells. This persistent presence has been closely tied to the development and progression of MS, although the exact mechanisms linking the virus to MS pathology remain under investigation. Leading hypothesis include the reactivation of EBV within memory B cells in the CNS, the cross-reactivity of anti-EBV antibodies with human proteins in the CNS and the activation of “forbidden” memory B cells that recognize CNS antigens (25). However, despite the strong link between EBV and MS, no single pathogen has been definitely accepted as causal agent of the disease. Fungal infections, though less explored, have also been cautiously examined in the literature as potential contributors to MS pathogenesis. Some studies have shown symptoms improvement with antifungal treatments and the presence of antibodies against fungi in both serum and cerebrospinal fluid (CSF) of MS patients (16). Moreover, several known MS risk factors are linked to an increased susceptibility to fungal infections. Fungal populations in the human gut, similar to bacterial communities, are closely correlated with age. However, fungi also exhibit notable differences based on sex. The primary sex-based distinction is the higher prevalence of the genus Candida in the gastrointestinal tract of women (22). Since Candida is a commensal microorganism of the female genitourinary tract, it is thought to serve as a “natural” reservoir for the gastrointestinal tract. Vertical transmission (mother to child) of Candida has been documented, but Candida is generally not detected in the gut until adolescence (26, 27), a period during which the first symptoms of MS often emerge (usually between the ages of 20 and 30). These observations suggest a potential connection between Candida colonization and the increased risk of MS, although further research is needed to clarify this connection.
The HLA-DRB1*15 allele group is the strongest genetic risk factor for MS (28). This gene variant plays a critical role in immune system regulation and antigen presentation, determining which antigens trigger a T cell-mediated immune response and supports the maturation of naive B cell. The specific alleles HLA-DRB1*1501 and HLA-DRB1*1503, which confer the greatest predisposition to MS, have also been linked to an increased incidence of infections suspected to have a fungal etiology, such as pulmonary sarcoidosis, uveitis, or allergic bronchopulmonary aspergillosis (16).
The potential fungal etiology of MS is also being explored based on the beneficial effect of dimethyl fumarate (DMF), an immunomodulatory drug for MS. The proposed mechanisms by which DMF modulates the immune response include its anti-inflammatory interaction with the nuclear factor (erythroid-derived 2)-like 2 (Nrf2) pathway. The efficacy of DMF has been demonstrated by its ability to reduce relapse rates by about 50% as compared with placebo and reduce the disability progression. However, the exact mode of action remains incompletely understood (29). Notably, DMF has long been used as an industrial fungicide to suppress mold growth, suggesting that its efficacy in alleviating MS symptoms may also stem from a direct antifungal effect (16).
Additionally, the presence of chitotriosidase in CSF is an important biomarker of MS. Chitotriosidase is produced by activated macrophages to hydrolyze chitin, a polysaccharide found in fungi and absent in prokaryotic or mammalian cells. The reason for elevated chitotriosidase in MS patients’ CSF is not fully understood, but it may be linked to the presence of fungi or a microbiota rich in fungal organisms (30, 31). Calprotectin is another key biomarker particularly elevated in the CSF of MS patients during disease relapses. Located in the cytosol of neutrophils, calprotectin is an antimicrobial protein with strongly activity against fungi, further supporting the hypothesis of a fungal involvement in MS (16).
This study seeks to investigate the composition of the gut mycobiota in MS patients across different treatment conditions and disease stages, compared to healthy controls. By integrating microbiota analysis with inflammatory markers such as calprotectin and chitotriosidase, as well as genetic data on the HLA-DRB1*15 alleles, we aim to uncover potential links between gut fungal communities, immune function, and genetic risk factors in MS. These findings may lead to the identification of new therapeutic targets and provide insights into how treatments like DMF affect both the gut mycobiome and MS progression.
2 Materials and methods
2.1 Participants and sample collection
We conducted a prospective observational case-control study involving 50 individuals diagnosed with Multiple Sclerosis (MS) from Hospital Universitario Donostia (Donostia-San Sebastian, Spain) and Hospital Universitario Araba (Vitoria-Gasteiz, Spain), between December 2021 and June 2023, alongside 25 healthy controls (HCs) of the same age range. The local ethics committee approved the research protocol (reference PI2020070) on July 14, 2021. Written informed consent was obtained from all participants.
To qualify for enrolment, MS participants needed confirmed MS diagnosis according to the revised McDonald criteria (32). Exclusion criteria included gastrointestinal or chronic infectious diseases, recent steroids use (within the last month), chemotherapy or antibiotics treatment (within the last three months), pregnancy, or being within six months postpartum. Additionally, individuals with MS must have been on the same disease modifying therapy (DMT) for at least the last three months. MS participants were either treatment-naïve or treated with Dimethyl fumarate (Tecfidera®). HCs were required to have no autoimmune, gastrointestinal or chronic infectious diseases and not be relative to MS subjects.
Participants were provided with materials and instructions for fecal sample collection, including a stool collector designed to adhere to the toilet, tubes to collect stool samples, a safety bag, and hydrated cold packs to maintain the required temperature. Upon submission of frozen fecal sample, blood was drawn via venipuncture for plasma and DNA extraction. Samples were store at -80°C until analysis. At recruitment, participants completed a food frequency questionnaire to assess their eating habits, and test included in the Multiple Sclerosis Functional Composite (MSFC) were performed to measure various levels of disability in patients. Relevant clinical and demographic data, along with key questionnaire results, are summarized in Table 1.
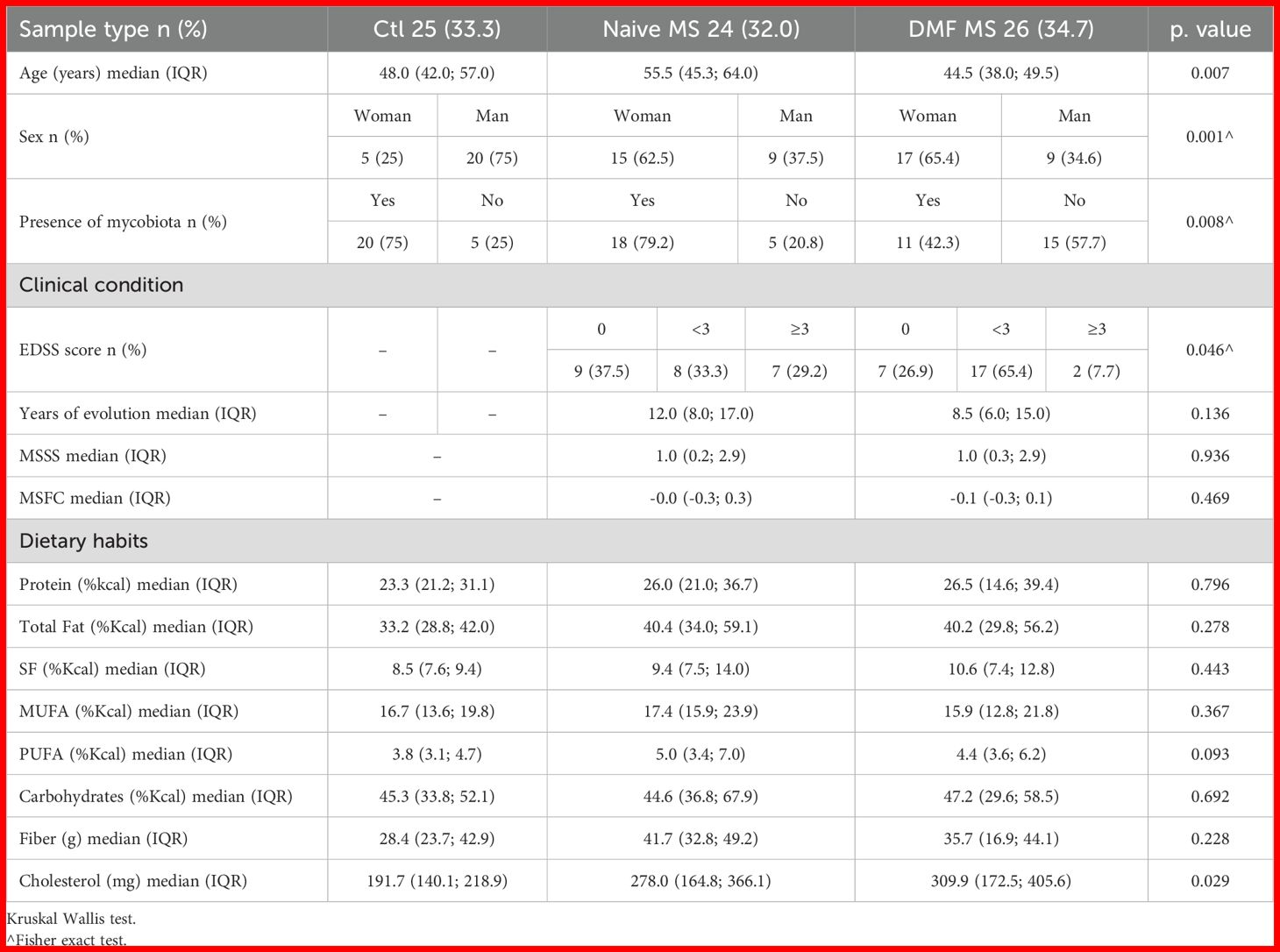
Table 1. Clinical and demographic data of the studied population and summary of their dietary habits.
2.2 DNA extraction and sequencing
Fecal samples were thawed on ice and diluted in 1X Dulbecco’s Phosphate Buffered Saline (DPBS) (Gibco, USA). DNA extraction was performed using both mechanical and enzymatic lysis, and the extraction kit QIAamp DNA Stool Mini Kit (Qiagen, Germany). Mechanical disruption involved bead-beating with 0.1 mm diameter zirconia/silica beads (Sigma, St.Louis, USA) using the Bead Ruptor 12 (OMNI International, Georgia, USA), conducted 1 minute three times. Enzymatic lysis of fungal cells was achieved with 0,1mg/ml zymolyase (MP Biomedicals, LLC, Illkirch-Graffenstaden, France). The extracted DNA underwent quality and quantity assessments. A negative control, extracted in parallel using an empty sterile tube (similar to those used for fecal samples) without any biological material, was included to evaluate the impact of reagents and potential contamination.
Amplification of the fungal ITS1 and ITS2 intergenic regions was performed using the primers ITS1 30F (5′- GTCCCTGCCCTTTGTACACA -3′) and ITS1 217R (5′- TTTCGCTGCGTTCTTCATCG -3′); as well as ITS86F (5’- GTGAATCATCGAATCTTTGAA -3’) and ITS4R (5’- TCCTCCGCTTATTGATATGC -3’), with AmpliTaq Gold™ DNA Polymerase (Applied Biosystems). The thermal cycling conditions were 95°C for 10 minutes, followed by 35 cycles of 95°C for 30 seconds, 55°C for 30 seconds, and 72°C for 1 minute. The PCR products from each fecal sample were pooled and sequence on Ion Torrent PGM (Life Technologies, MA, USA) using a 318 chip.
2.3 Data processing
ITS analysis was conducted using the QIIME2 microbiome bioinformatics platform. The pipeline performed in QIIME2 included the following steps: data import, quality filtering and denoising (using cutadapt to remove primers), ASV identification (via DADA2 denoise-pyro), diversity analysis (with reads rarefied to 10,000) and taxonomic classification using the UNITE database (through VSEARCH feature-classifier). Given the limited availability of pipelines for analyzing Ion Torrent sequences, the optimal approach for this analysis has been a topic of discussion within the QIIME2 community forum. The specific steps described here were selected based on recommendations and discussions in the forum (33–36). The pipeline designed and used for the microbiota analysis has been deposited on GitHub, https://github.com/MGorostidi/mbiome. (Gorostidi-Aicua, M. (2024) Mbiome. GitHub repository. https://github.com/MGorostidi/mbiome).
2.4 HLA-DRB1*15 allele genotyping
DNA was extracted from the buffy coat by the Basque Biobank (biobancovasco.org) using the FlexiGene DNA Kit (QIAGEN) and stored at -20˚C until use. Samples were thawed on ice prior to analysis.
Genotyping was performed using the TaqMan® SNP Genotyping Assay (Thermo Fisher). This assay employed a primer pair and two allele-specific probes, labeled FAM and VIC, to detect G and A nucleotides of the SNP rs3135388, respectively. Quantitative polymerase chain reaction (qPCR) was conducted using the TaqPath ProAmp Master Mix and 20X TaqMan SNP Genotyping Assay with 10ng of genomic DNA per well on 384-well plates. Reactions were carried out in a total volume of 5 µl using the CFX384 instrument (BioRad). The qPCR protocol included a pre-read step at 60°C for 30 seconds, an initial denaturalization at 95°C for 5 minutes, followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. Fluorescence detection occurred at 60°C. Control samples representing all possible genotypes and a negative template control (NTC) were included in each run.
2.5 Calprotectin and chitotriosidase ELISAs
Calprotectin concentration in plasma was measured with Human Calprotectin L1/S100-A8/A9 Complex ELISA Kit (Catalog# EH62RB, Invitrogen) following the manufacturer’s instructions. Plasma samples were diluted 1:400 to fit the standard curve of the kit. Plasma chitotriosidase levels were measured using the Human Chitotriosidase ELISA Kit (Catalog# EH105RB, Invitrogen) according to the manufacturer’s instructions. Plasma samples were diluted 1:80 to fit the standard curve provided by the kit.
2.6 Food Frequency Questionnaire analysis
The FFQ included 142 food items, with intake recorded in the following categories: never, hardly ever, monthly (1–3 times per month), weekly (1–6 times per week) and daily (1 to more than 6 times per day). The food items were based on the validated FFQ used in the PREDIMED study, with slight modifications (37). The questionnaire covered a wide range of foods typically consumed in a Mediterranean diet. The nutritional value of each item was calculated using food composition tables from the Universidad Complutense of Madrid (38), which provides detailed nutritional information for 960 food items, expressed per 100 grams of edible portion.
The percentage of calories provided by each macronutrient was calculated by converting grams to calories (4 Kcal/g for proteins and carbohydrates, and 9 Kcal/g for fat) and then determining the percentage relative to the total caloric intake.
The estimation of adequate/inadequate intake of the main macro and micronutrients has been based on WHO recommendations (39).
2.7 MSFC determination
Multiple Sclerosis Functional Composite (MSFC) is a multidimensional, three-component performance scale used to assess the degree of impairment in MS patients. Timed 25-Foot Walk (T25W) for leg function and ambulation, 9-Hole Peg Test (9HPT) for arm and hand function, and Symbol Digit Modalities Test (SDMT) for cognitive function. These tests were explained to participants and performed at the time of signing the documentation or during sample delivery.
Since the units of three variables measured by these tests are different, raw scores are converted to common metric, z-scores. The overall composite score is calculated as the average of the z-score from the three tests. Negative scores represent those patients with an evolution better than average and positive values indicate worse evolution.
2.8 Statistical analysis
Statistical analysis was done with R software version 4.3.1 (http://cran.r-project.org). Quantitative variables were expressed as mean and 95% coefficient interval. Categorical variables were expressed as numbers and percentages. Kruskal-Wallis test was used to compare between individuals with MS and controls in the quantitative variables, while Two Tailed Fisher exact test was used to compare groups in categorical variables. Pearson’s correlation was used to describe the association between the quantitative variables. All plots were performed in R environment. Correlation matrices were achieved by using the R corrplot package (v0.94) and significant correlations were calculated using Hmisc package (v5.1.3). ROC curves were performed with pROC R package (v1.18.5). AUC and cut point were calculated with the same package. PCA was performed using r packages ggfortify (v0.4.17) and ggplot2 (v3.5.1). P-values ≤ 0.05 were considered statistically significant.
3 Results
A total of 50 individuals with MS and 25 controls participated in the study. Participant ages ranged from 27 to 87 years across groups. The MS subjects treated with DMF were generally younger, with a median age of 44.5 years and older in naïve individuals (55.5 years as a median) versus controls (median age of 48 years) (p = 0.007). Following the trends in the incidence of the disease, women were more frequently represented in the MS group, while men were more common in the control group (p = 0.001).
MS treatment with DMF was associated with a significant reduction in the gut mycobiota presence; dropping from 79.2% in naïve group to 42.3% in DMF treated individuals. Mycobiota was present in 75% of controls (p = 0.008).
Disability was assessed through EDSS scores, categorized as 0, <3 and ≥3. Slight differences in EDSS distribution were observed within MS subjects, being the intermediate form by far the predominant one in individuals treated with DMF (p = 0.046). However, no significant group differences were found for the Multiple Sclerosis Severity Score (MSSS) (p = 0.936) or the MS functional composite (MSFC) score (p = 0.469) (Table 1).
No differences were observed in the dietary habits of participants, except for cholesterol intake, which was higher among individuals with MS (p = 0.029).
3.1 Differential diet-fungus correlation pattern in individuals with MS suggests disease-specific macronutrient metabolism
The analysis of FFQs indicates that the dietary habits of individuals with MS are generally similar to those reported by controls, with the differences primarily limited to higher cholesterol consumption among those with MS (p= 0.029). Protein intake was within recommended levels for most participants, while total fat consumption, particularly of saturated fats, was excessive, and carbohydrates intake fell short of recommended values for the majority (Figure 1A).
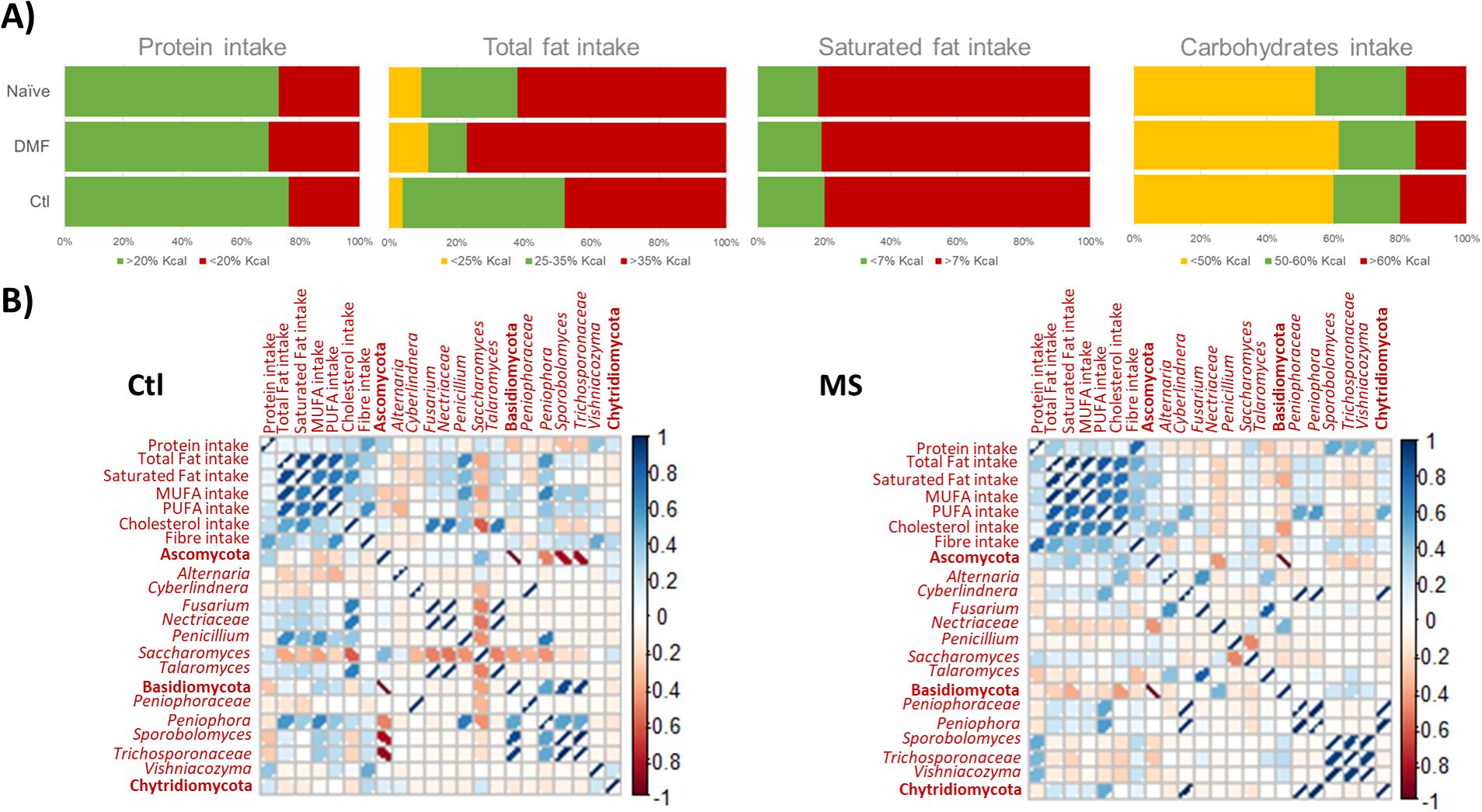
Figure 1. (A) Barr plot illustrating the intake of primary macronutrients among study participants, categorized by MS status and type of disease modifying therapy. Intake levels were evaluated against WHO recommend guidelines. (B) Pearson's correlation matrix depicting significant correlations between dietary components and fungal populations, with comparisons between the control (Ctl) and MS (MS) groups. Positive correlations are represented in blue, and negative correlations in red; the intensity of color and the width of ellipses correspond to the strength of the correlation.
Correlation patterns between major macronutrients and fungal populations differed between controls and individuals with MS (Figure 1B). Specifically, cholesterol intake was associated with levels of main fungal phyla, Ascomycota and Basidiomycota, in the MS group but not in controls. At more specific taxonomic levels, cholesterol intake was linked to increased levels of Talaromyces (p = 0.010), Nectriaceae (p = 0.006) and Fusarium (p = 0.010) in controls, and to Alternaria (p = 0.040) in MS patients. In controls, Saccharomyces levels decreased with higher intake of cholesterol (p = 0.060). The genus Peniophora correlated with total fat and MUFA intake in controls (p = 0.002) and with PUFA intake in individuals with MS (p = 0.005). Penicillium was correlated with total fats (p = 0.003), saturated fats (p = 0.046) and MUFA intake (p = 0.010) in controls, but no significant correlation was observed in MS patients. Additionally, Cyberlindnera was associated with PUFA ingestion in individuals with MS (p = 0.016) but not in controls. A positive correlation was observed between Vishniacozyma and fiber intake in controls (p = 0.024), whereas in MS patients, Vishniacozyma (p = 0.025), Sporobolomyces (p = 0.013) and Trichosporonaceae (p = 0.030) were associated with protein intake (Figure 1B).
3.2 The mycobiome of individuals with MS exhibits disease specific characteristics and is partially influenced by DMF treatment
The total DNA content per gram of feces ranged from 6.12 and 760.28 ng, with a mean of 133.24 ng/g, while no DNA was detected in the negative control. DNA quantity was standardized prior to amplification and again before sequencing.
The average total reads in the samples was 47598, with the number of genera ranging from 2 to 43. Negative controls exhibit similar total reads and Shannon diversity index to the samples; however, their Ascomycota/Basidiomycota ratio and taxonomic profile were distinctly different from those observed in the samples (Supplementary Figure 1). Most of the reads predominant in the negative control samples (except for Saccharomyces) were nearly undetectable in the samples (<0.5%). Therefore, we considered contamination background and ubiquitous DNA in reagents to be unlikely to affect the mycobiota composition in fecal samples.
The mycobiota composition did not show a clear clustering of the study groups. Nevertheless, the fungal genera Torulaspora and Debaryomyces were primarily associated with the MS mycobiota, particularly in treatment naïve participants, while Saccharomyces was characteristic of the control group. No distinct fungal taxa were identified in the mycobiota of patients treated with DMF (Figure 2A). Mycobiome detection was more frequent in controls than in people with MS, and DMF treatment determined a reduction in mycobiota presence. The Ascomycota phylum and the genus Saccharomyces predominated in the human mycobiota, with relative abundances ranging from 54.3% and 100% for Ascomycota and 0.01% and 99.96% for Saccharomyces across the samples (Figure 2B). A trend toward increased fungal diversity and richness was observed in people with MS, although no significant differences were found in the Ascomycota/Basidiomycota ratio between groups.
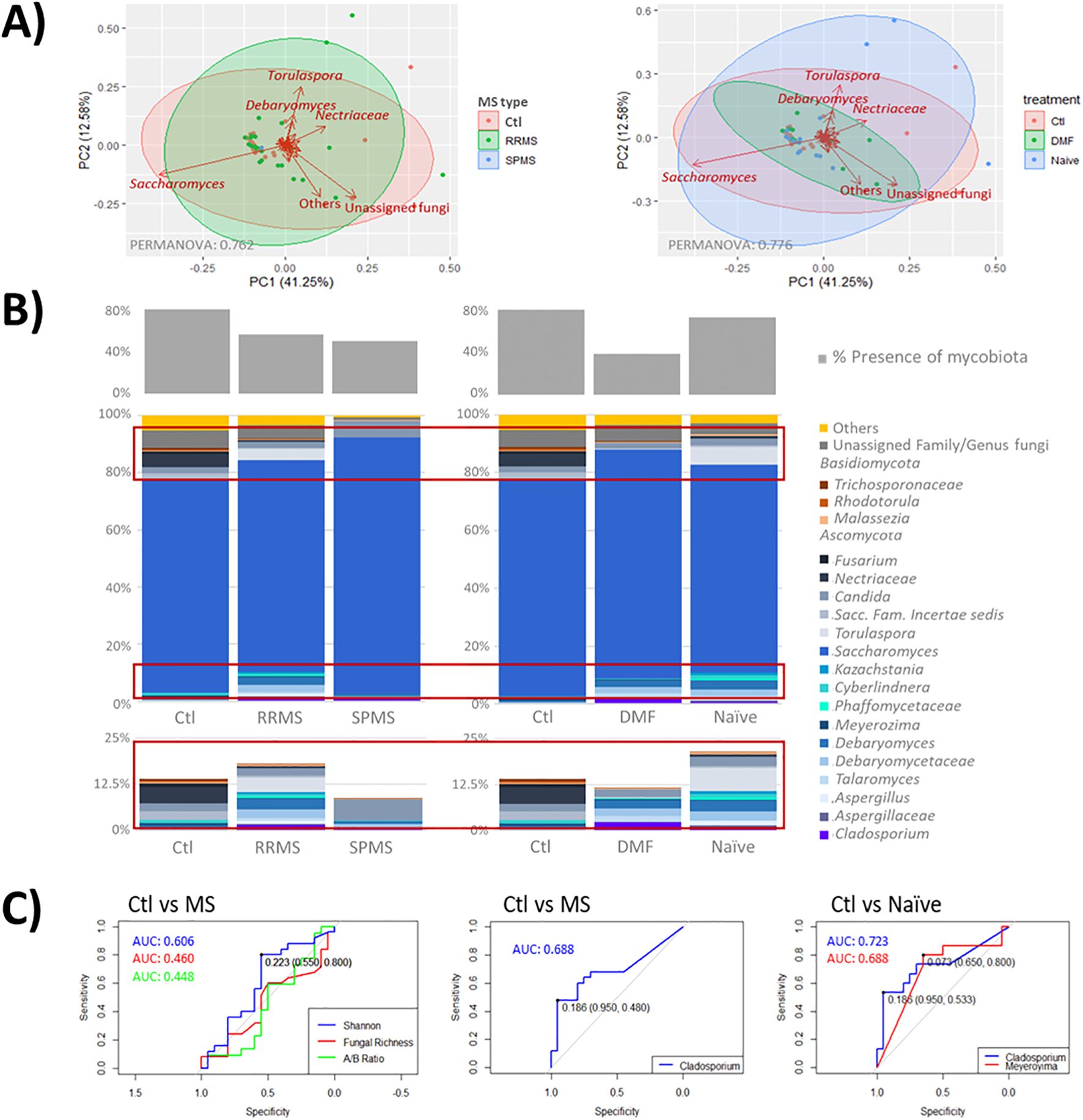
Figure 2. (A) Scatter plot representing samples grouped by disease type (left) and treatment status (right). Clustered samples were color-coded by group, with fungal groups that have the greatest influence on sample placement indicated by arrows. The bidimensional PCoA plot explains 53.83% of the variability among the samples. (B) Taxonomic analysis of mycobiota across samples and groups. The middle graph shows a taxonomy bar plot of the main phyla and genera (those accounting for >0.5%abundance and/or present in at least 10% of the samples). The upper graph illustrates the presence of mycobiota across the samples; with minority genera highlighted in red boxes are enlarged at the bottom. Note, where genus level resolution was not available, taxa are reported at the family level. (C) ROC curves illustrating the diagnostic performance of proposed mycobiome biomarkers (Cladosporium and Meyerozyma), with associated AUC values, compared to global parameters such as diversity (Shannon), fungal richness and the Ascomycota/Basidiomycota ratio.
Key differences between individuals with MS and controls were primarily found in the minority taxa of the mycobiota (Figure 2B; Table 2). Specifically, reads from the genera Cladosporium (p = 0.022) and unassigned genera of the family Sclerotiniaceae (p = 0.039) were elevated in MS patients, while reads from Meyerozima (p = 0.043) and unassigned genera of the family Saccharomycetaceae (p = 0.039) were reduced. These differences were less pronounced in patients treated with DMF (Table 2).
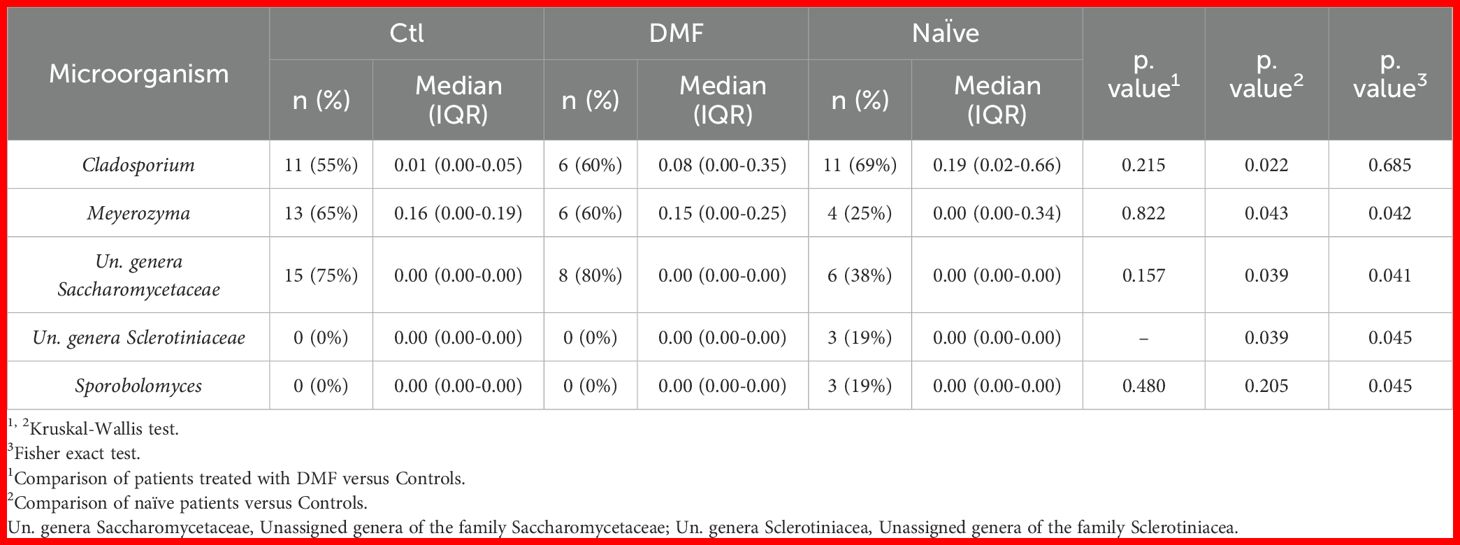
Table 2. Fungal groups detected on the mycobiota of the participants in the study.
Further analysis revealed that the presence of Meyerozima (p = 0.042) and unassigned genera of the family Saccharomycetaceae (p = 0.041) were less frequent in naïve MS patients, while the presence of unassigned genera of the family Sclerotiniaceae (p = 0.045) and the genus Sporobolomyces (p = 0.045) characterized the mycobiota of naïve patients (Table 2). Investigation of some of these genera as potential disease markers showed that Cladosporium abundance in the gut could predict MS with an AUC value of 0.688, while the combination of Cladosporium and Meyerozima levels differentiated between controls and naïve patients with AUC values of 0.723 and 0.688, respectively; higher than those obtained from general parameters (Figure 2C).
3.3 Mycobiota composition change with disease severity. Specific fungal population correlates with severity metrics of MS
All the severity metric used yielded similar results; Torulaspora was the fungal group most characterized of individuals with lower disability, while unassigned fungal genera predominated in those with higher disability levels. Although clustering was not statistically significant across any of the scales used, the EDSS scoring, categorized as <3 or ≥3, provided the most effective group separation (PERMANOVA: 0.072) (Figure 3A). Among specific microorganisms, those most strongly correlated with MS severity scores included Crassiperidium across all scales, as well as Aspergillus, Nectriaceae and Malassezia on the EDSS and MSSS scales (Figure 3B). Malassezia in the gut showed the highest predictive accuracy for MS severity on the EDSS and MSSS scales, with an AUC of 0.642. However, global mycobiome metrics, particularly alpha diversity measures, demonstrated even better predictive power, with Shannon Diversity index achieving an AUC of 0.700 (Figure 3C).
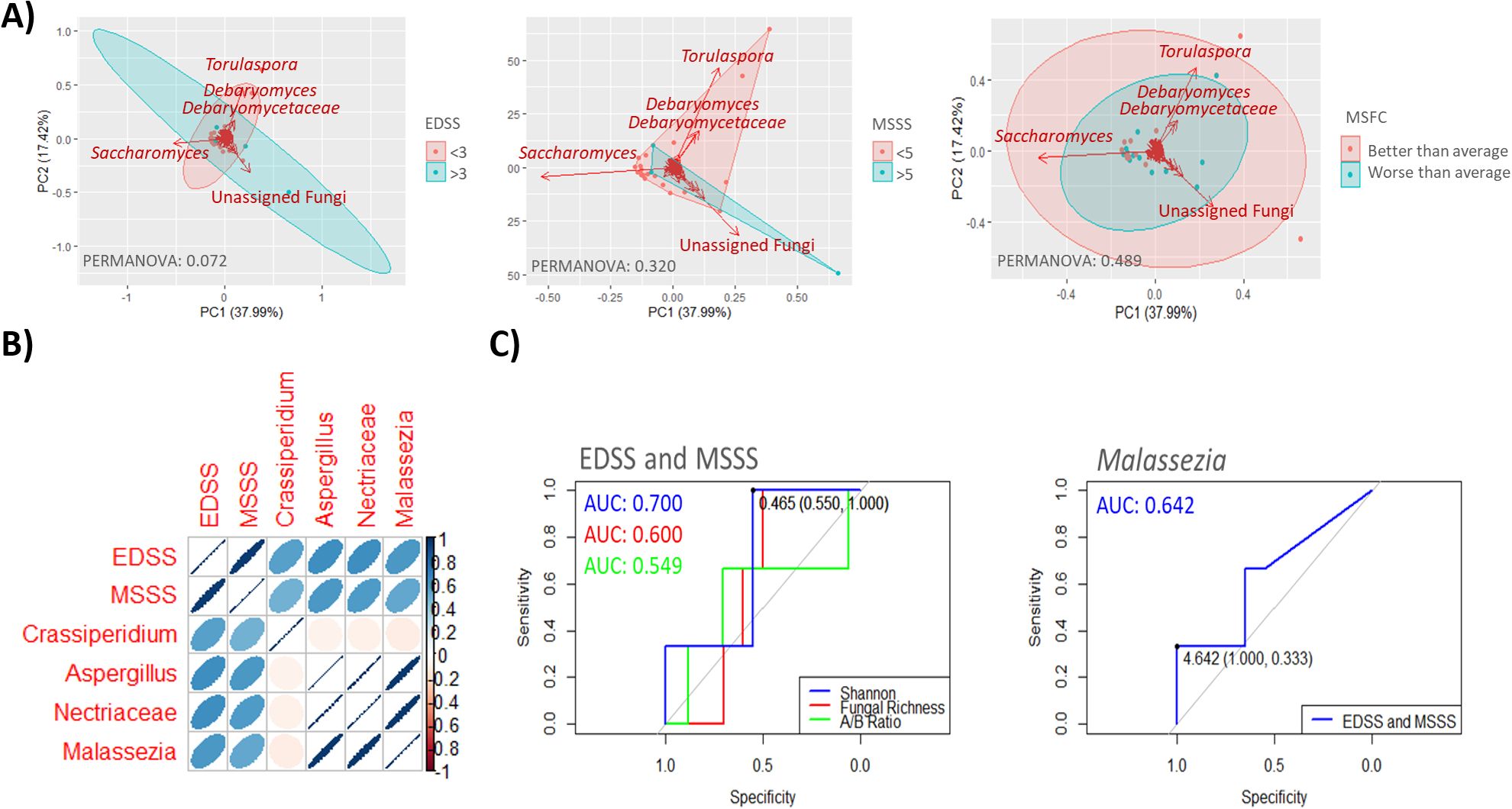
Figure 3. (A) Scatter plots showing sample groupings based on disease severity as determined by the EDSS score (≥3 or <3) (left), MSSS score (≥5 or <5) (middle) and MSFC score (better or worse than average) (right). Samples within clusters are shown in different colors, with arrows indicating the fungal groups most influential in determining sample placement. The bidimensional PCoA plot explains 55.41% of the sample variability. (B) Pearson correlation matrix depicting the relationship among four fungal genera significantly correlated (p < 0.05) with MS severity scales EDSS and MSSS. Color intensity reflects the correlation strength, with dark blue (1) indicating a complete positive correlation and dark red (−1) indicating a complete negative correlation. The shape of the ellipses denotes correlation strength, with narrower ellipses indicating stronger correlations and wider ellipses weaker correlation. (C) ROC curves illustrating the diagnostic performance of proposed mycobiome biomarkers, specifically Malassezia, in assessing severity as determined by EDSS or MSSS scores (right panel). Globally parameters such as diversity (Shannon), fungal richness and Ascomycota/Basidiomycota ratio are shown in the left panel.
3.4 Mycobiota alteration and its relation with risk of MS susceptibility
The mycobiota of individuals with MS demonstrated a significant correlation with the MS genetic risk factor HLA-DRB1*15 alleles. HLA genotyping revealed that individuals carrying the HLA-DRB1*1501 allele had a distinct mycobiota composition, characterized by the presence of Debaryomyces and Talaromyces genera. In contrast, the mycobiota of non-carriers was predominated by the genus Cladosporium (Figure 4A). Furthermore, carriers of the HLA-DRB1*1501 allele were more likely to exhibit more severe MS, as reflected by worse MSFC scores and higher EDSS scores. However, the overall composition of the gut mycobiota did not differ significantly between HLA-DRB1*1501 carriers and non-carriers (Figure 4B).
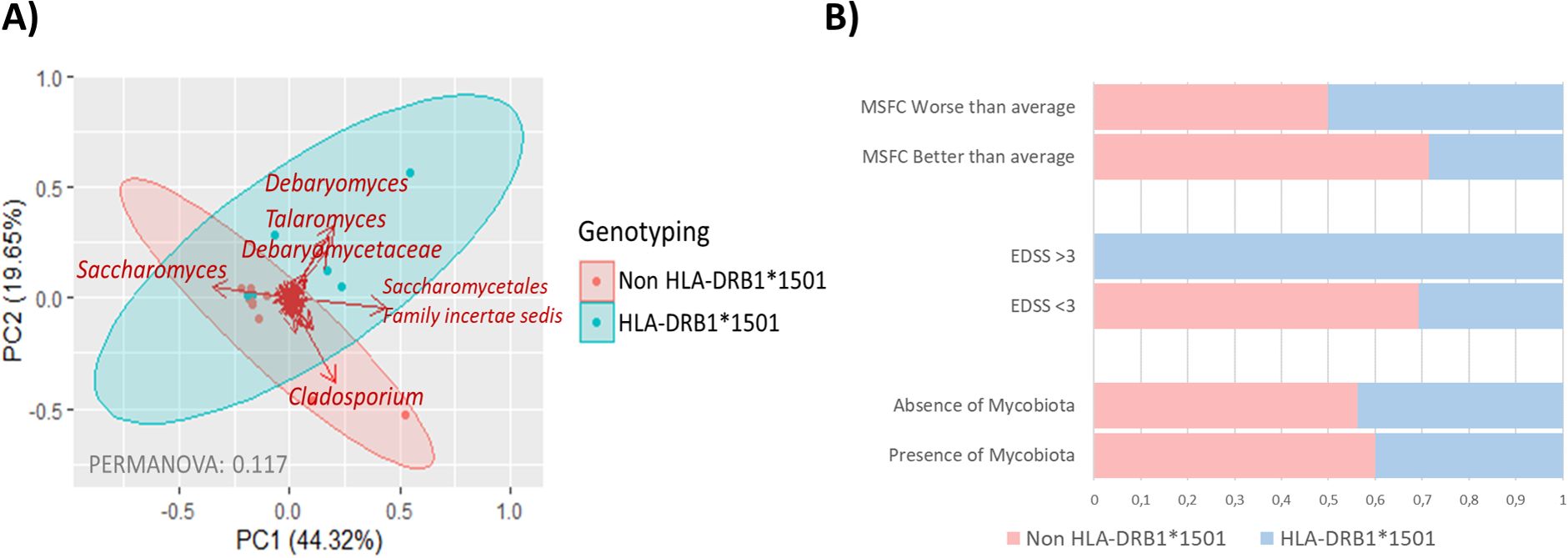
Figure 4. (A) Scatter plot illustrating sample groupings based on HLA-DRB1*15 allele genotyping, with carriers (HLA-DBR1*1501) represented in blue and non-carriers (HLA-DBR1*1501) in red. Arrows indicate the fungal groups most influential in determining samples placement. The bidimensional PCoA plot accounts for 63.97% of the variability in the samples. (B) Barr plot showing the distribution of HLA-DRB1*1501 allele carriers across MSFC and EDSS severity categories, alongside the presence of gut mycobiota in individuals with MS.
Plasma chitotriosidase levels did not differ significantly between individuals with MS and controls. Interestingly, in MS patients, chitotriosidase levels tended to increase with age (p = 0.027), a pattern not observed in controls. However, no significant correlation was observed between plasma chitotriosidase levels and MS severity metrics in the overall population, except for the higher levels in treatment naïve patients with worse prognosis, as indicated by a higher MSSS score (p = 0.028) (Table 3).
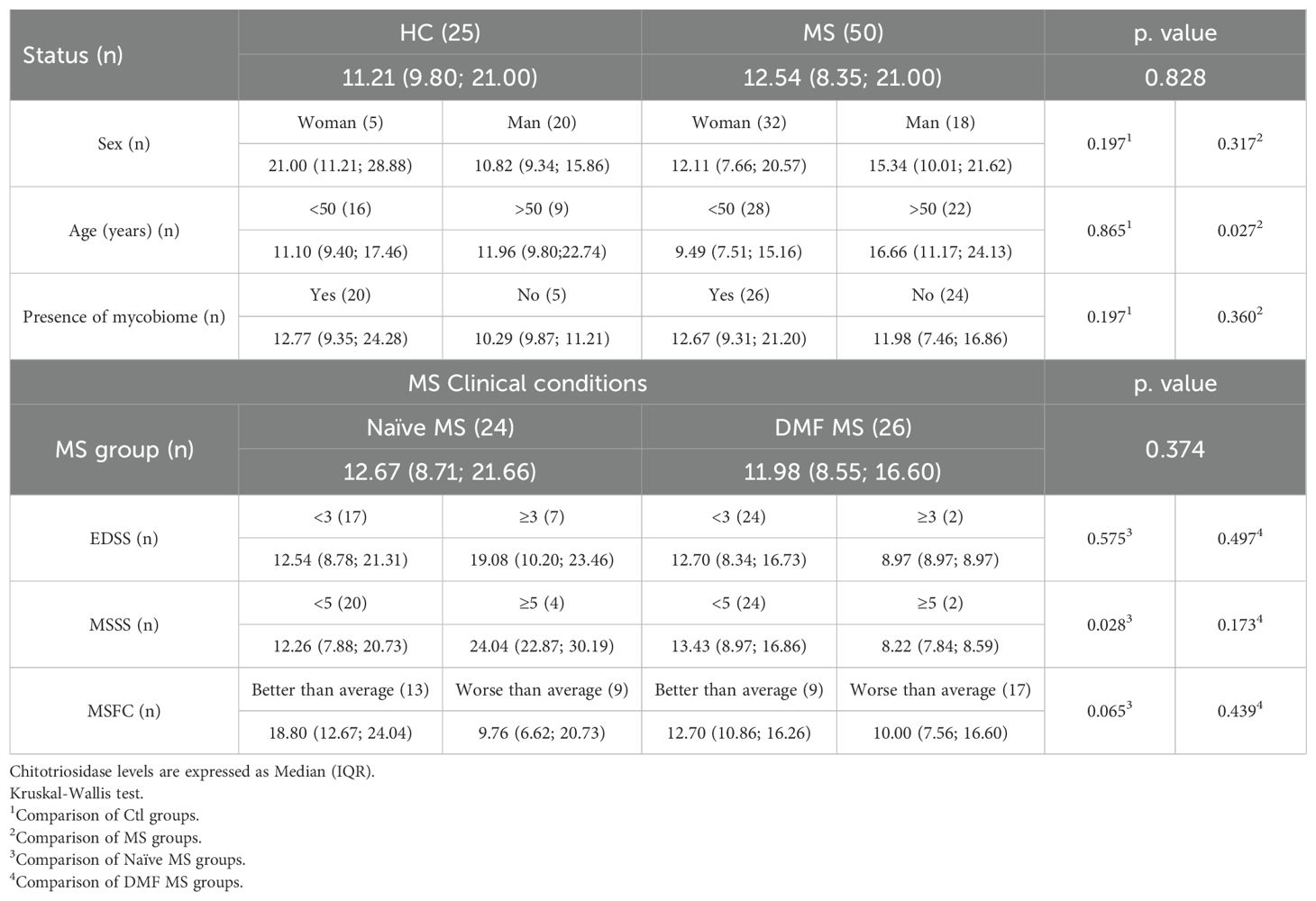
Table 3. Chitotriosidase plasma levels (ng/ml).
Plasma calprotectin levels do not show significant differences between individuals with MS and controls, nor were associated with age, sex, disease severity or treatment. However, a tendency for increased calprotectin levels was observed in men and in controls with the presence of gut mycobiota, a pattern not seen in individuals with MS (Table 4).
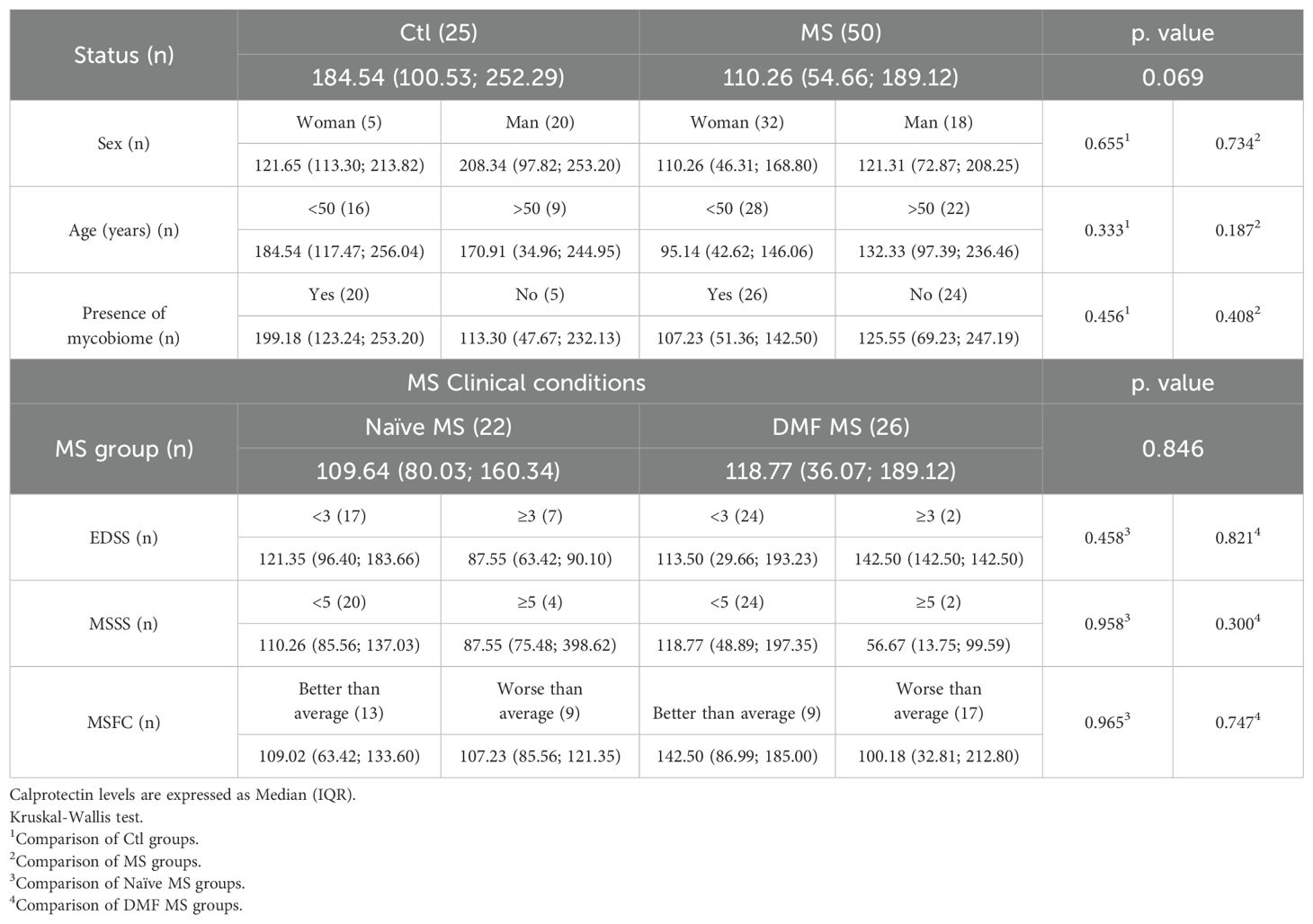
Table 4. Calprotectin plasma levels (ng/ml).
Several fungal groups showed significant correlations with plasma calprotectin and chitotrisidase levels, with these correlations being disease specific. Notably, higher calprotectin levels in MS patients were associated with a gut mycobiota composition characterized by reduced Ascomycota and increased Basidiomycota. Both plasma calprotectin and chitotriosidase levels displayed a positive correlation with the fungal families Aspergillaceae and Nectriaceae, as well as the genus Malassezia in individuals with MS but not in controls (Figure 5A). The mycobiota of MS patients with plasma calprotectin levels either higher or lower than the average observed in controls showed significant differences (permanova 0.023). Specifically, the genera Saccharomyces and Aspergillus were predominant in patients with higher calprotectin levels, while Torulaspora and Debaryomyces characterized the mycobiota of those with lower calprotectin levels (Figure 5B). In contrast, no differences were observed in the mycobiota of MS patients when stratified by plasma chitotriosidase levels (Figure 5C).
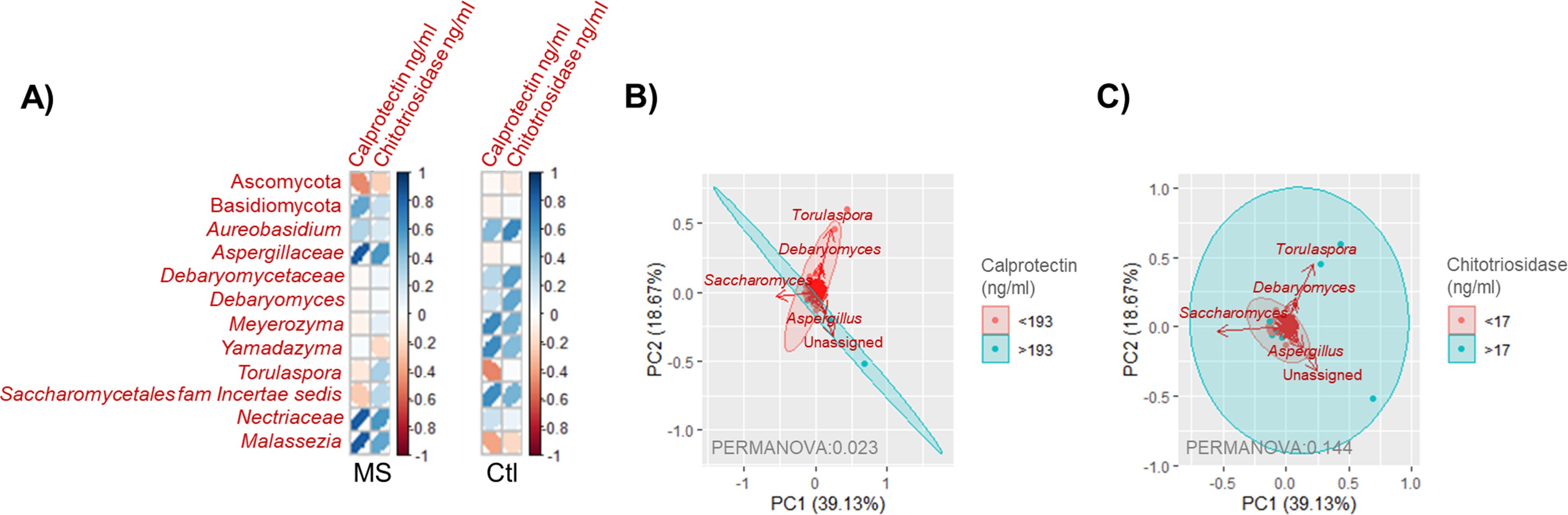
Figure 5. (A) Pearson correlation matrix showing fungal taxa significantly correlated (p < 0.05) with plasma calprotectin or chitotrisiosidase levels in individuals with MS or controls. Colors indicate the correlation strength, with dark blue representing a complete positive correlation (1) and dark red indicating a complete negative correlation (−1). The shape of the ellipses reflects correlation strength, with narrow ellipses indicating stronger correlations and wider ellipses weaker correlations. (B) Scatter plot representing samples of individuals with MS grouped by plasma calprotectin levels, categorized as <193 ≥193 ng/ml (the average concentration found in controls). Arrows indicate the fungal groups most influential in determining sample placement. The bidimensional PCoA plot explains 57.8% of the sample variability. (C) Scatter plot representing samples of individuals with MS grouped by plasma chitotriosidase levels, categorized as <17 ≥17 ng/ml (the average concentration found in controls). Arrows represent the fungal groups most influential in determining sample placement. The bidimensional PCoA plot explains 57.8% of the sample variability.
4 Discussion
Our previous studies have identified cross-kingdom interactions within the gut microbiota of individuals with MS from the Basque Country, including characterizing the bacterial gut microbiota in relation to disease modifying therapies for MS (40–42). In the current study, we expand our analysis by describing the fungal microbiota of this population and examining its associations with MS risk factors of different natures, as well as dietary habits.
The fungal microbiota of individuals with MS exhibited higher fungal diversity and richness, which is consistent with findings reported by other authors (43–45). While a general trend of increased Basidiomycota presence has been previously described in MS, we did not detect significant differences in the levels of Ascomycota or Basidiomycota in our cohort. However, we did observe a higher richness of the phylum Basidiomycota in individuals with MS compared to controls. The top five genera identified in our MS population were Saccharomyces, Torulaspora, Debaryomyces, Candida and Cladosporium. These findings align with previous studies, thought genera such as Xylaria, Penicillium, Agaricus, Pichia, Aspergillus and Malassezia have also been identified in other MS populations (43–45). Our analysis revealed that the main differences between individuals with MS and controls were found in minority genera, and no statistical differences in β-diversity parameters were observed.
We further explored the relationship between mycobiota composition and clinical metrics, finding several genera that positively correlated with EDSS and MSSS scores. Among these, Malassezia stood out as the most predictive genus for significant increases in disability, as measured by both EDSS (≥3) and MSSS (≥5) scores. This fungal genus, predominantly detected in women, has previously been linked to MS (46) and other neurodegenerative diseases (47–51). Malassezia is known for its affinity for lipids, and while it is typically a commensal of human skin and overgrowth on skin sebum, it has the potential to invade internal organ and cause infection, including the CNS, under certain conditions (50). The lipid rich environment of the brain (52), coupled with immune suppression and BBB disruption in MS, may facilitate the invasion of Malassezia into the CNS, potentially contributing to disease progression (47). Its presence has been identified on the CNS of individuals with Alzheimer (48) and MS (14).
Additionally, we found Aspergillus, a genus commonly associated with respiratory infections and known for mycotoxin production, also correlated with MS disability. Although CNS infections due to Aspergillus are rare, they can occur under specific conditions, especially in immunocompromised individuals (53, 54). The Nectriaceae family, which includes both saprophytic and pathogenic fungi, also correlated with disease severity, with the ubiquitous fungi Fusarium being of particular note. Fusarium has been documented as an opportunistic pathogen capable of invading the CNS, even in healthy individuals, though such infections are becoming more common, especially in immunocompromised patients (55).
Dietary habits in our study population revealed subtle differences between individuals with MS and controls. Notably, we observed distinct pattern of mycobiota composition associated with macronutrient intake, suggesting important metabolic differences in individuals with MS. In particular, cholesterol intake in controls was found to significantly influenced mycobiota composition, specifically in relation to fungal phyla abundances. Fats emerged as the most influential macronutrient in shaping mycobiota composition. Given the well-documented alterations in lipid metabolism in MS (56, 57), it is not surprising that dietary habits influence mycobiota composition differently in individuals with MS compared to controls.
We also explored the influence of MS genetic risk factors on mycobiota composition. The presence of the HLA-DRB1*1501 allele, a major genetic risk factor for MS (28), was found to significantly impact mycobiota composition, particularly in relation to MS related disability. The association between the HLA-DRB1*1501 allele and fungal infections has been widely discussed in the literature (58–60), and our findings further support its role in shaping both mycobiota composition and MS progression.
Our study also investigated the potential role of plasma chitotriosidase and calprotectin levels as biomarkers in MS. Previous research has report elevated levels of chitotriosidase in both the CNS (61) and plasma of MS patients (30). While we observed a slight increase in plasma chitotriosidase levels in MS patients, the differences were not statistically significant, though they were more pronounced in treatment naïve patients. These findings are consistent with those of Beliën et al., who linked increased chitotriosidase levels in CNS with microglia activation and proposed it as an early CSF biomarker for disability progression in MS (61). Interestingly, we observed marked gender and age related differences in chitotriosidase levels, particularly in MS patients. Although demographic parameters have not been extensively evaluated in relation to chitotriosidase levels, this warrant further investigation.
In contrast to studies reporting increased chitotriosidase levels with disease modifying treatments (e.g., β-interferons) (30), we found no significant effects of DMF, which has recognized antifungal activity, on chitotriosidase levels. This discrepancy may reflect the impact of DMF on both mycobiota composition and other immune related pathways. The correlation patterns between fungi, plasma chitotriosidase and calprotectin levels in individuals with MS and controls highlight the need for combined strategies to study microbiota composition, as the identification of potentially pathogenic species/strains may enhance our understanding of these differences.
Calprotectin, a calcium-binding protein involved in numerous biological processes, has been implicated in various inflammatory conditions, including MS. Previous studies have shown that calprotectin levels in CSF of MS patients are related to relapse rate (62) and plasma calprotectin levels are associated with active MS, as indicated by neurofilament light chain (NfL) levels (63). However, our study found no association between plasma calprotectin levels and disease status, disability, or progression. These results suggest that the influence of calprotectin may be more specific to CSF levels and disease activity rather than plasma levels. Interestingly, we did observe that plasma calprotectin levels influenced mycobiota composition (PERMANOVA: 0.023), aligning with previous findings suggesting a role for calprotectin in shaping the microbiota in inflammatory diseases (64).
In conclusion, this study highlights the distinct characteristics of the fungal microbiota in individuals with MS, emphasizing its potential role in disease severity and progression. The associations between specific fungal genera, such as Malassezia and Aspergillus, and clinical metrics like EDSS and MSSS scores, underscore the relevance of fungal dynamics in MS pathology. Moreover, the interplay between genetic factors like HLA-DRB1*1501, dietary influences, and plasma biomarkers such as chitotriosidase and calprotectin further illustrates the complexity of host-mycobiota interactions in MS. The work of the International Multiple Sclerosis Microbiome Study (iMSMS) consortium has firmly established an association between the bacterial component of the microbiota and MS. However, microbial communities do not exist in isolation, fungal and bacterial populations are closely interconnected, and changes in one community can lead to reciprocal shift in the other. These interactions are further modulated by host factors, creating a dynamic and multidirectional network of influences. Understanding MS pathogenesis thus requires moving beyond single-kingdom analysis and embracing a holistic view of the microbiota as an integrated ecosystem. This study not only highlights novel associations between the fungal microbiota and clinical, genetic, and inflammatory features of MS, but also sets the groundwork for the systematic characterization of minority microbial communities, such as fungi, within the broader microbiome landscape. By establishing and applying a targeted methodology for the analysis of these often-overlooked components, we demonstrate the feasibility and relevance of integrating fungal data into microbiome research.
These findings provide a foundation for future research to explore the functional implications of fungal dysbiosis in MS and its potential as a therapeutic target or biomarker. A multidisciplinary approach combining microbiota profiling, biomarker analysis and clinical insights is essential to fully unravel the role of the mycobiota in MS pathogenesis and management.
Data availability statement
The data presented in the study are deposited in the NCBI SRA repository, accession numbers from SAMN49790753 to SAMN49790798 and BioProject PRJNA1286116.
Ethics statement
The studies involving humans were approved by Comité de Ética de la Investigación con Medicamentos de Euskadi. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributions
AO-C: Investigation, Writing – review & editing, Formal Analysis, Visualization, Writing – original draft. MG-A: Formal Analysis, Software, Data curation, Investigation, Writing – review & editing. LM-A: Investigation, Writing – review & editing. AA: Writing – review & editing, Investigation. LR: Methodology, Writing – review & editing. IM: Methodology, Writing – review & editing. AAA: Methodology, Writing – review & editing. MA: Writing – review & editing, Methodology. TC-T: Writing – review & editing, Methodology. DO: Project administration, Methodology, Resources, Writing – review & editing, Funding acquisition, Supervision, Conceptualization. LM: Visualization, Formal Analysis, Methodology, Investigation, Writing – review & editing, Project administration, Conceptualization, Supervision, Funding acquisition, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Department of Health of the Basque Government under Grants 2019111013 and 2022111029. LM was supported by the Sara Borrell contract (the National Institute of Health Carlos III). AO-C was supported by the Department of Education of the Basque Government and MG-A by the University of the Basque Country.
Acknowledgments
We would like to thank all the donors who participated in this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1625794/full#supplementary-material
Supplementary Figure 1 | Comparison of mycobiome profile in Negative controls (NC) and fecal samples. (A) Boxplot of the Ascomycota/Basidiomycota ratio. (B) Boxplot of the Shannon Diversity index. (C) Barr plot of the 10 more abundant taxa in the samples. Un. orders Dothideomycetes: Unassigned orders in the class Dothideomycetes; Un. genera Debaryomycetaceae: Unassigned genera in Debaryomycetaceae family; Un. genera Nectriaceae: Unassigned genera in Nectriaceae family; Un. families Pleosporales: Unassigned families in the order Pleosporales; Un. orders Sordariomycetes: Unassigned orders in the class Sordariomycetes; Un. classes Basidiomycota: Unassigned classes in the phyla Basidiomycota; Un. genera Schizoporaceae: Unassigned genera in Schizoporaceae family.
References
1. Kleinewietfeld M and Hafler DA. Regulatory T cells in autoimmune Neuroinflammation. Immunol Rev. (2014) 259:231–44. doi: 10.1111/imr.12169, PMID: 24712469
2. Koch-henriksen N, Thygesen LC, Stenager E, and Laursen B. Incidence of MS has increased markedly over six decades in Denmark particularly with late onset and in women. Neurology. (2018) 90:e1954–63. doi: 10.1212/WNL.0000000000005612, PMID: 29720546
3. Beecham AH, Patsopoulos NA, Xifara DK, Davis MF, Kemppinen A, Cotsapas C, et al. Analysis of immune-related loci identifies 48 new susceptibility variants for multiple sclerosis. Nat Genet. (2013) 45:1353–62. doi: 10.1038/ng.2770, PMID: 24076602
4. Ochoa-Repáraz J, Kirby TO, and Kasper LH. The gut microbiome and multiple sclerosis. Cold Spring Harb Perspect Med. (2018) 8:1–16. doi: 10.1101/cshperspect.a029017, PMID: 29311123
5. Ghezzi L, Cantoni C, Pinget GV, Zhou Y, and Piccio L. Targeting the gut to treat multiple sclerosis. J Clin Invest. (2021) 131:e143774. doi: 10.1172/JCI143774, PMID: 34196310
6. Wang X, Liang Z, Wang S, Ma D, Zhu M, and Feng J. Role of gut microbiota in multiple sclerosis and potential therapeutic implications. Curr Neuropharmacol. (2021) 20:1413–26. doi: 10.2174/1570159X19666210629145351, PMID: 34191698
7. Sokol H, Leducq V, Aschard H, Pham HP, Jegou S, Landman C, et al. Fungal microbiota dysbiosis in IBD. Gut. (2017) 66:1039–48. doi: 10.1136/gutjnl-2015-310746, PMID: 26843508
8. Wheeler ML, Limon JJ, Bar AS, Arditi M, Underhill DM, Iliev ID, et al. Short article immunological consequences of intestinal fungal short article immunological consequences of intestinal fungal dysbiosis. Cell Host Microbe. (2016) 19:865–73. doi: 10.1016/j.chom.2016.05.003, PMID: 27237365
9. Iliev ID. Mycobiota–host immune interactions in IBD: coming out of the shadows. Nat Rev Gastroenterol Hepatol. (2022) 19:91–2. doi: 10.1038/s41575-021-00541-2, PMID: 34815533
10. Forbes JD, Bernstein CN, Tremlett H, Van Domselaar G, and Knox NC. A fungal world: Could the gut mycobiome be involved in neurological disease? Front Microbiol. (2019) 10:1–13. doi: 10.3389/fmicb.2018.03249, PMID: 30687254
11. Benito-León J, Pisa D, Alonso R, Calleja P, Díaz-Sánchez M, and Carrasco L. Association between multiple sclerosis and Candida species: Evidence from a case-control study. Eur J Clin Microbiol Infect Dis. (2010) 29:1139–45. doi: 10.1007/s10096-010-0979-y, PMID: 20556470
12. Saroukolaei SA, Ghabaee M, Shokri H, Badiei A, and Ghourchian S. The role of Candida albicans in the severity of multiple sclerosis. Mycoses. (2016) 59:697–704. doi: 10.1111/myc.12489, PMID: 27061227
13. Pisa D, Alonso R, Jiménez-Jiménez FJ, and Carrasco L. Fungal infection in cerebrospinal fluid from some patients with multiple sclerosis. Eur J Clin Microbiol Infect Dis. (2013) 32:795–801. doi: 10.1007/s10096-012-1810-8, PMID: 23322279
14. Alonso R, Fernández-Fernández AM, Pisa D, and Carrasco L. Multiple sclerosis and mixed microbial infections. Direct identification of fungi and bacteria in nervous tissue. Neurobiol Dis. (2018) 117:42–61. doi: 10.1016/j.nbd.2018.05.022, PMID: 29859870
15. Fraga-Silva TFC, Mimura LAN, Marchetti CM, Chiuso-Minicucci F, Francą TGD, Zorzella-Pezavento SFG, et al. Experimental autoimmune encephalomyelitis development is aggravated by Candida albicans infection. J Immunol Res. (2015) 2015:635052. doi: 10.1155/2015/635052, PMID: 25969836
16. Benito-León J and Laurence M. The role of fungi in the etiology of multiple sclerosis. Front Neurol. (2017) 8:535. doi: 10.3389/fneur.2017.00535, PMID: 29085329
17. Hallen-Adams HE and Suhr MJ. Fungi in the healthy human gastrointestinal tract. Virulence. (2017) 8:352–8. doi: 10.1080/21505594.2016.1247140, PMID: 27736307
18. Zoll J, Snelders E, Verweij PE, and Melchers WJ. Next-generation sequencing in the mycology lab. Curr Fungal Infect Rep. (2016) 10:37–42. doi: 10.1007/s12281-016-0253-6, PMID: 27358660
19. Huseyin CE, O’Toole PW, Cotter PD, and Scanlan PD. Forgotten fungi-the gut mycobiome in human health and disease. FEMS Microbiol Rev. (2017) 41:479–511. doi: 10.1093/femsre/fuw047, PMID: 28430946
20. Ott SJ, Kühbacher T, Musfeldt M, Rosenstiel P, Hellmig S, Rehman A, et al. Fungi and inflammatory bowel diseases: Alterations of composition and diversity. Scand J Gastroenterol. (2008) 43:831–41. doi: 10.1080/00365520801935434, PMID: 18584522
21. Pisa Di, Alonso R, Fernández-Fernández AM, Rábano A, and Carrasco L. Polymicrobial infections in brain tissue from alzheimer’s disease patients. Sci Rep. (2017) 7:1–14. doi: 10.1038/s41598-017-05903-y, PMID: 28717130
22. Severance EG, Gressitt KL, Stallings CR, Katsafanas E, Schweinfurth LA, Savage CL, et al. Candida albicans exposures, sex specificity and cognitive deficits in schizophrenia and bipolar disorder. NPJ Schizophr. (2016) 2:16018. doi: 10.1038/npjschz.2016.18, PMID: 27336058
23. Strati F, Cavalieri D, Albanese D, De Felice C, Donati C, Hayek J, et al. New evidences on the altered gut microbiota in autism spectrum disorders. Microbiome. (2017) 5:1–11. doi: 10.1186/s40168-017-0242-1, PMID: 28222761
24. Marrodan M, Alessandro L, Farez MF, and Correale J. The role of infections in multiple sclerosis. Mult Scler J. (2019) 25:891–901. doi: 10.1177/1352458518823940, PMID: 30638421
25. Laurence M and Benito-León J. Epstein–Barr virus and multiple sclerosis: Updating Pender’s hypothesis. Mult Scler Relat Disord. (2017) 16:8–14. doi: 10.1016/j.msard.2017.05.009, PMID: 28755684
26. Strati F, Di Paola M, Stefanini I, Albanese D, Rizzetto L, Lionetti P, et al. Age and gender affect the composition of fungal population of the human gastrointestinal tract. Front Microbiol. (2016) 7:1227. doi: 10.3389/fmicb.2016.01227, PMID: 27536299
27. Ward TL, Knights D, and Gale CA. Infant fungal communities: current knowledge and research opportunities. BMC Med. (2017) 15:30. doi: 10.1186/s12916-017-0802-z, PMID: 28190400
28. Hollenbach JA and Oksenberg JR. The immunogenetics of multiple sclerosis: A comprehensive review. J Autoimmun. (2015) 64:13–25. doi: 10.1016/j.jaut.2015.06.010, PMID: 26142251
29. Linker RA and Gold R. Dimethyl fumarate for treatment of multiple sclerosis: Mechanism of action, effectiveness, and side effects. Curr Neurol Neurosci Rep. (2013) 13:394. doi: 10.1007/s11910-013-0394-8, PMID: 24061646
30. Comabella M, Domínguez C, Rio J, Martín-Gallán P, Vilches A, Vilarrasa N, et al. Plasma chitotriosidase activity in multiple sclerosis. Clin Immunol. (2009) 131:216–22. doi: 10.1016/j.clim.2008.12.004, PMID: 19176289
31. Elmonem MA, Van Den Heuvel LP, and Levtchenko EN. Immunomodulatory effects of chitotriosidase enzyme. Enzyme Res. (2016) 2016:17–20. doi: 10.1155/2016/2682680, PMID: 26881065
32. Thompson AJ, Banwell BL, Barkhof F, Carroll WM, Coetzee T, Comi G, et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. (2018) 17:162–73. doi: 10.1016/S1474-4422(17)30470-2, PMID: 29275977
33. QIIME2 forum. Available online at: https://forum.qiime2.org/t/cutadapt-in-ion-torrent-sequences/22165/10 (Accessed March 1, 2022).
34. QIIME2 forum. Available online at: https://forum.qiime2.org/t/dada2-denoise-pyro-for-ion-torrent/17605 (Accessed Nov 26, 2020).
35. QIIME2 forum. Available online at: https://forum.qiime2.org/t/rarefying-when-having-multiple-runs/22087/18 (Accessed March 1, 2022).
36. QIIME2 forum. Available online at: https://forum.qiime2.org/t/classifier-for-ion-torrent-data/3675/8 (Accessed March 15, 2022).
37. Fernández-Ballart JD, Piñol JL, Zazpe I, Corella D, Carrasco P, Toledo E, et al. Relative validity of a semi-quantitative food-frequency questionnaire in an elderly Mediterranean population of Spain. Br J Nutr. (2010) 103:1808–16. doi: 10.1017/S0007114509993837, PMID: 20102675
38. Ortega RM, López-Sobaler AM, Andrés P, and Aparicio A. Food nutritional composition. A tool for the design and evaluation of food and diets. Departament of Nutrition and Food Science. Comlutense University of Madrid. Madrid. (2021).
39. WHO. Healthy diet. (2020). Available online at: https://www.who.int/news-room/fact-sheets/detail/healthy-diet (Accessed April 29, 2020).
40. Gorostidi-Aicua M, Reparaz I, Otaegui-Chivite A, García K, Romarate L, Álvarez de Arcaya A, et al. Bacteria-fungi interactions in multiple sclerosis. Microorganisms. (2024) 12(5):872. doi: 10.3390/microorganisms12050872, PMID: 38792701
41. Moles L, Delgado S, Gorostidi-Aicua M, Sepúlveda L, Alberro A, Iparraguirre L, et al. Microbial dysbiosis and lack of SCFA production in a Spanish cohort of patients with multiple sclerosis. Front Immunol. (2022) 13:960761. doi: 10.3389/fimmu.2022.960761, PMID: 36325343
42. Moles L, Otaegui-Chivite A, Gorostidi-Aicua M, Romarate L, Mendiburu I, Crespillo-Velasco H, et al. Microbiota modulation by teriflunomide therapy in people with multiple sclerosis: An observational case-control study. Neurotherapeutics. (2024) 21:e00457. doi: 10.1016/j.neurot.2024.e00457, PMID: 39406600
43. Yadav M, Ali S, Shrode RL, Shahi SK, Jensen SN, Hoang J, et al. Multiple sclerosis patients have an altered gut mycobiome and increased fungal to bacterial richness. PloS One. (2022) 17:e0264556. doi: 10.1371/journal.pone.0264556, PMID: 35472144
44. Shah S, Locca A, Dorsett Y, Cantoni C, Ghezzi L, Lin Q, et al. Alterations of the gut mycobiome in patients with MS. EBioMedicine. (2021) 71:103557. doi: 10.1016/j.ebiom.2021.103557, PMID: 34455391
45. Gargano F, Guerrera G, Piras E, Serafini B, Di Paola M, Rizzetto L, et al. Proinflammatory mucosal-associated invariant CD8+ T cells react to gut flora yeasts and infiltrate multiple sclerosis brain. Front Immunol. (2022) 13:890298. doi: 10.3389/fimmu.2022.890298, PMID: 35979352
46. Benito-León J and Laurence M. Malassezia in the central nervous system and multiple sclerosis. Infection. (2019) 47:135–6. doi: 10.1007/s15010-018-1196-3, PMID: 30120719
47. Fernández-Espejo E. Microorganisms associated with increased risk of Parkinson’s disease. Neurologia. (2023) 38:495–503. doi: 10.1016/j.nrleng.2020.08.023, PMID: 35644845
48. Alonso R, Pisa D, Aguado B, and Carrasco L. Identification of fungal species in brain tissue from alzheimer’s disease by next-generation sequencing. J Alzheimer’s Dis. (2017) 58:55–67. doi: 10.3233/JAD-170058, PMID: 28387676
49. Han X, Bedarf J, Proske-Schmitz S, Schmitt I, and Wüllner U. Increased diversity of Malassezia species on the skin of Parkinson’s disease patients. Front Aging Neurosci. (2023) 15:1268751. doi: 10.3389/fnagi.2023.1268751, PMID: 37854034
50. Laurence M, Benito-León J, and Calon F. Malassezia and parkinson’s disease. Front Neurol. (2019) 10:758. doi: 10.3389/fneur.2019.00758, PMID: 31396143
51. Phuna ZX and Madhavan P. A closer look at the mycobiome in Alzheimer’s disease: Fungal species, pathogenesis and transmission. Eur J Neurosci. (2022) 55:1291–321. doi: 10.1111/ejn.15599, PMID: 35048439
52. Yoon JH, Seo Y, Jo YS, Lee S, Cho E, Cazenave-Gassiot A, et al. Brain lipidomics: From functional landscape to clinical significance. Sci Adv. (2022) 8:1–14. doi: 10.1126/sciadv.adc9317, PMID: 36112688
53. London F, Stanciu-Pop C, and Mulquin N. Chronic cavitary pulmonary aspergillosis in a teriflunomide treated MS patient. Clin Neurol Nuerosurgery. (2024) 236:108125. doi: 10.1016/j.clineuro.2024.108125, PMID: 38246031
54. Janardan A, Prokhoda P, Razzak AN, Jethwa T, and Paudel HR. A complex case of aspergillus infection of the brain and its future medical implications. Cureus. (2022) 14:e29756. doi: 10.7759/cureus.29756, PMID: 36324347
55. Strong N and Ostrosky-Zeichner L. Fusarium species central nervous system infection. Curr Opin Infect Dis. (2024) 37:185–91. doi: 10.1097/QCO.0000000000001009, PMID: 38518108
56. Pineda-Torra I, Siddique S, Waddington KE, Farrell R, and Jury EC. Disrupted lipid metabolism in multiple sclerosis: A role for liver X receptors? Front Endocrinol (Lausanne). (2021) 12:639757. doi: 10.3389/fendo.2021.639757, PMID: 33927692
57. Podbielska M, O’keeffe J, and Pokryszko-Dragan A. New insights into multiple sclerosis mechanisms: Lipids on the track to control inflammation and neurodegeneration. Int J Mol Sci. (2021) 22:7319. doi: 10.3390/ijms22147319, PMID: 34298940
58. Chauhan B, Santiago L, Hutcheson PS, Schwartz HJ, Spitznagel E, Castro M, et al. Evidence for the involvement of two different MHC class II regions in susceptibility or protection in allergic bronchopulmonary aspergillosis. J Allergy Clin Immunol. (2000) 106:723–9. doi: 10.1067/mai.2000.109913, PMID: 11031343
59. Voorter CEM, Drent M, and Van Den Berg-Loonen EM. Severe pulmonary sarcoidosis is strongly associated with the haplotype HLA-DQB1*0602-DRB1*150101. Hum Immunol. (2005) 66:826–35. doi: 10.1016/j.humimm.2005.04.003, PMID: 16112030
60. Tang WM, Pulido JS, Eckels DD, Han DP, Mieler WF, and Pierce K. The association of HLA-DR15 and intermediate uveitis. Am J Ophthalmol. (1997) 123:70–5. doi: 10.1016/S0002-9394(14)70994-8, PMID: 9186099
61. Beliën J, Swinnen S, D’hondt R, de Juan LV, Dedoncker N, Matthys P, et al. CHIT1 at diagnosis predicts faster disability progression and reflects early microglial activation in multiple sclerosis. Nat Commun. (2024) 15:1–15. doi: 10.1038/s41467-024-49312-y, PMID: 38866782
62. Berg-Hansen P, Vandvik B, Fagerhol M, and Holmøy T. Calprotectin levels in the cerebrospinal fluid reflect disease activity in multiple sclerosis. J Neuroimmunol. (2009) 216:98–102. doi: 10.1016/j.jneuroim.2009.09.006, PMID: 19800696
63. Olsson A, Gustavsen S, Hasselbalch IC, Langkilde AR, Sellebjerg F, Oturai AB, et al. Biomarkers of inflammation and epithelial barrier function in multiple sclerosis. Mult Scler Relat Disord. (2020) 46:102520. doi: 10.1016/j.msard.2020.102520, PMID: 32980645
Keywords: mycobiome, multiple sclerosis, chitotriosidase, calprotectin, HLA-DRB1*1501
Citation: Otaegui-Chivite A, Gorostidi-Aicua M, Martins-Almeida L, Alberro A, Romarate L, Mendiburu I, Álvarez de Arcaya A, Arruti M, Castillo-Triviño T, Otaegui D and Moles L (2025) Exploring the mycobiota in multiple sclerosis: its influence on disease development and progression. Front. Immunol. 16:1625794. doi: 10.3389/fimmu.2025.1625794
Received: 09 May 2025; Accepted: 20 June 2025;
Published: 23 July 2025.
Edited by:
Ingo Kleiter, Marianne-Strauss-Klinik, GermanyReviewed by:
Luis Del Carpio-Orantes, Mexican Social Security Institute, MexicoAndreia Barroso, IQVIA, Brazil
Copyright © 2025 Otaegui-Chivite, Gorostidi-Aicua, Martins-Almeida, Alberro, Romarate, Mendiburu, Álvarez de Arcaya, Arruti, Castillo-Triviño, Otaegui and Moles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David Otaegui, RGF2aWQub3RhZWd1aUBiaW8tZ2lwdXprb2EuZXVz; Laura Moles, TGF1cmEubW9sZXNhbGVncmVAYmlvLWdpcHV6a29hLmV1cw==
†These authors have contributed equally to this work and share last authorship
‡ORCID: Miriam Gorostidi-Aicua, orcid.org/0000-0002-4868-3280
Ainhoa Alberro, orcid.org/0000-0002-9985-5812
Leire Romarate, orcid.org/0000-0002-5412-6413