 Céline Fombellida-Lopez1
Céline Fombellida-Lopez1 Diego Aguilar Ortmans1
Diego Aguilar Ortmans1 Michel Moutschen2
Michel Moutschen2 Alexander O. Pasternak3†*
Alexander O. Pasternak3†* Gilles Darcis1,2†*
Gilles Darcis1,2†*- 1Immunology and Infectious Diseases Laboratory, Grappe Interdisciplinaire de Génoprotéomique Appliquée (GIGA)-Institute, University of Liège, Liège, Belgium
- 2General Internal Medicine and Infectious Diseases Department, University Hospital of Liège, Liège, Belgium
- 3Laboratory of Experimental Virology, Department of Medical Microbiology, Amsterdam University Medical Center (UMC), University of Amsterdam, Amsterdam, Netherlands
Introduction: Despite effective antiretroviral therapy (ART), people with HIV (PWH) experience persistent immune activation and inflammation, increasing the risk of non-AIDS-related comorbidities. The contribution of the HIV reservoir to this chronic inflammatory state remains debated. Understanding the relationship between HIV persistence, immune activation, and inflammation is crucial for optimizing long-term therapeutic strategies.
Methods: This study assessed HIV persistence, immune activation, and systemic inflammation in 49 PWH treated with the same dolutegravir-based triple ART regimen. HIV reservoir size and activity were evaluated by measuring total HIV DNA in peripheral blood mononuclear cells (PBMCs) and rectal tissue, cell-associated (CA) unspliced (US) HIV RNA, and residual viremia. Over 20 inflammatory biomarkers, including sCD14, IL-6, TNF-α, and CXCL10, were analyzed, along with comprehensive immune profiling using a 26-color spectral flow cytometry panel. Clinical parameters such as age, nadir CD4 count, and co-infections were also considered.
Results and discussion: Our findings showed a limited association between HIV persistence markers and systemic inflammation or immune activation. Compared to previous studies, participants had lower reservoir sizes and transcriptional activity, likely due to early ART initiation and prolonged suppression. Immune preservation was evident, with high CD4/CD8 ratios and reduced activation markers. These results challenge the idea that the HIV reservoir is the primary driver of chronic inflammation in PWH on a dolutegravir-based long-term ART. Instead, the reservoir may evolve toward a more transcriptionally silent and defective state, reducing its impact on systemic immune activation.
1 Introduction
Human immunodeficiency virus (HIV) infection remains a major global health issue, affecting an estimated 39.9 million people in 2023 (WHO, HIV and AIDS, Fact sheets, 22 July 2024 (1)). Antiretroviral therapy (ART) has transformed HIV infection into a manageable chronic condition and brought the life expectancy of people living with HIV (PWH) nearly in line with that of uninfected individuals (2, 3). However, ART is not curative and requires life-long adherence; while it effectively suppresses viral replication, it does not eradicate the virus from latent reservoirs or fully restore immune function (4–6). PWH often suffer from chronic immune activation and inflammation, increasing their risk of non-AIDS-related comorbidities, such as cardiovascular diseases, neurocognitive disorders, and metabolic syndrome (7, 8).
This persistent inflammatory state in ART-treated PWH is complex and multifactorial (9). HIV persists in viral reservoirs, especially within CD4+ T cells and monocytes (10, 11), where it can produce low levels of viral RNA, and even viral proteins, potentially triggering antiviral immune responses (12). Additionally, low-level viral replication in sanctuary sites, where ART do not fully penetrate, may contribute to ongoing inflammation (13, 14). Microbial translocation, which is the leakage of bacterial products from the gut into the bloodstream due to gut-associated lymphoid tissue (GALT) disruption by HIV, further stimulates immune responses and increases levels of inflammatory mediators, such as lipopolysaccharide (LPS) and soluble CD14 (sCD14) (15, 16). Microbial translocation is linked to thymic function failure, another crucial factor involved in HIV disease progression and immune dysregulation (17). Immune dysregulation, particularly in CD4+ and CD8+ T cells, adds to this pro-inflammatory environment. CD4+ T cell depletion drives the release of inflammatory cytokines (e.g., IL-6, TNF-α), which are closely associated with poorer clinical outcomes in individuals with chronic HIV infection (18). Moreover, ART-treated individuals often show increased immune exhaustion (PD-1, LAG-3) and activation markers (CD38, HLA-DR) on T cells. These markers correlate with larger HIV reservoirs and reduced T-cell function (19, 20). Monocytes and other immune cells also contribute significantly to the inflammatory milieu (21). Monocytes play a central role in sustaining chronic inflammation, as they release pro-inflammatory cytokines and chemokines, which further activate immune pathways (22).
Lastly, clinical factors like age, gender, nadir CD4 count, co-infections (e.g. CMV, hepatitis B, hepatitis C, EBV), and timing of ART initiation, also influence inflammation levels. For instance, older age is associated with poorer immune responses (23–25), a phenomenon increasingly referred to as “inflammageing”, which has become more evident as ART extends life expectancy in PWH (26). Similarly, gender differences have been noted, with the female gender being linked to higher levels of immune activation (27, 28).
Understanding the relationship between HIV persistence markers and immune activation is crucial, as the inflammatory environment not only drives T-cell proliferation but also perpetuates a vicious cycle that maintains and replenishes viral reservoirs (13).
In this study, we aim to investigate the associations between HIV persistence, inflammatory markers, clinical indicators, and immune cell populations in a cohort of 49 PWH treated with the same dolutegravir-based ART regimen. We analysed over 40 parameters, categorized into four main groups:
1. HIV Persistence Markers: Total HIV DNA in PBMCs and in rectal biopsies (GALT), cell-associated (CA) unspliced (US) HIV RNA in PBMCs, and residual viremia.
2. Plasma inflammatory Markers: A panel of more than 20 cytokines, chemokines, and cell adhesion/inflammatory response molecules, including sCD14, High-sensitivity CRP (hs-CRP), IL-6, TNF-α, IFN-γ, CXCL10, and ICAM-1.
3. Immune Cell Populations: Using a 26-color spectral flow cytometry panel, we performed immunophenotyping to identify various immune cell populations, including T cells, B cells, monocytes, NK cells, regulatory T cells (Tregs), mucosal-associated invariant T (MAIT) cells, gamma delta (γδ) T cells and innate lymphoid cells (ILCs). We also assessed markers of immune activation (HLA-DR and CD38) and immune exhaustion (PD-1 and TIGIT).
4. Clinical Indicators: Patient demographic and health data, including gender, age, BMI, smoking status, ethnicity, nadir CD4 count, CD4/CD8 ratio, pre-ART zenith plasma viral load, time to treatment (the period between diagnosis of HIV infection and the start of treatment) and time of suppressive ART (duration between the initiation of the current suppressive ART regimen and the first study visit).
This research seeks to deepen our understanding of the complex immune landscape in ART-treated PWH and its implications for managing HIV-associated inflammation and related comorbidities.
2 Materials and methods
2.1 Study design and participants
This study combined the baseline data from two open-label, interventional, monocentric, randomized, and controlled clinical trials conducted at the University Hospital of Liège, Belgium, involving a total of 49 HIV-1-infected adults on long-term ART. Both trials focused on individuals treated with a combination of dolutegravir (DTG, 50 mg), abacavir (ABC, 600 mg), and lamivudine (3TC, 300 mg) for at least two years, with sustained viral suppression.
In the first study (ClinicalTrials.gov identifier: NCT05351684), 13 participants were enrolled in a phase 2 trial assessing the impact of DTG intensification (an additional daily dose of 50 mg) compared to a control group that continued the standard DTG/ABC/3TC regimen. Eligibility criteria included a consistently suppressed plasma viral load below 20 HIV RNA copies/mL for at least 12 months prior to screening (allowing brief “blips”) and an absolute CD4+ T lymphocyte count above 200 cells/mm³. Ethics approval was granted by the Liège University Hospital-Faculty Ethics Committee (2018/228), and all participants provided informed consent. The second study (ClinicalTrials.gov identifier: NCT04034862) was a phase 3 trial involving 36 participants. This trial aimed to evaluate the effects of treatment simplification to dual therapy (50 mg DTG + 300 mg 3TC) versus continued triple therapy. Inclusion criteria were identical to the first study: a viral load consistently below 20 HIV-1 RNA copies/mL for the previous 12 months and a CD4+ T cell count above 200 cells/mm³. Ethics approval for this study was provided by the same ethics committee (2018/292), and each participant provided written informed consent.
Detailed inclusion and exclusion criteria for both studies are available in Supplementary Data 1 and 2.
2.2 Methods
All methods used for the quantification of HIV persistence markers (including total HIV DNA in PBMCs and rectal biopsies, cell-associated HIV RNA in PBMCs, intact HIV DNA in PBMCs, and residual viremia), plasma inflammatory biomarkers, and immune cell populations are detailed in the original studies from which the data were derived (29, 30).
2.3 Statistical analysis
Statistical analyses were performed using R (version 4.2.2) and R studio (version 2023.6.1.524). Differences were tested for statistical significance using Mann-Whitney U tests for continuous variables and Fisher’s exact tests for categorical variables. Spearman’s rank method was used to determine the correlation coefficients. Correlograms were generated using the “corrplot” R package (version 0.95). For some variables, certain values were above or below a detection threshold set by the measuring instrument. For values below this threshold, a value between 0 and the threshold was randomly determined from a triangular distribution whose mode corresponds to the threshold value divided by 2. For values above a maximum detection threshold, the value of this threshold was used.
3 Results
3.1 Study design and participants
Forty-nine ART-suppressed participants, all receiving the same triple antiretroviral regimen consisting of 50 mg dolutegravir (DTG), 600 mg abacavir (ABC), and 300 mg lamivudine (3TC) for at least two years, were enrolled from two separate clinical trials (EDIT, NCT05351684 and IDOLTIB, NCT04034862) conducted at the University Hospital of Liège. The cohort consisted of thirteen participants from the EDIT study and thirty-six from the IDOLTIB study, all maintaining continuous suppressive treatment for a median of 7.62 years.
The baseline clinical characteristics of this combined cohort are presented in Table 1.
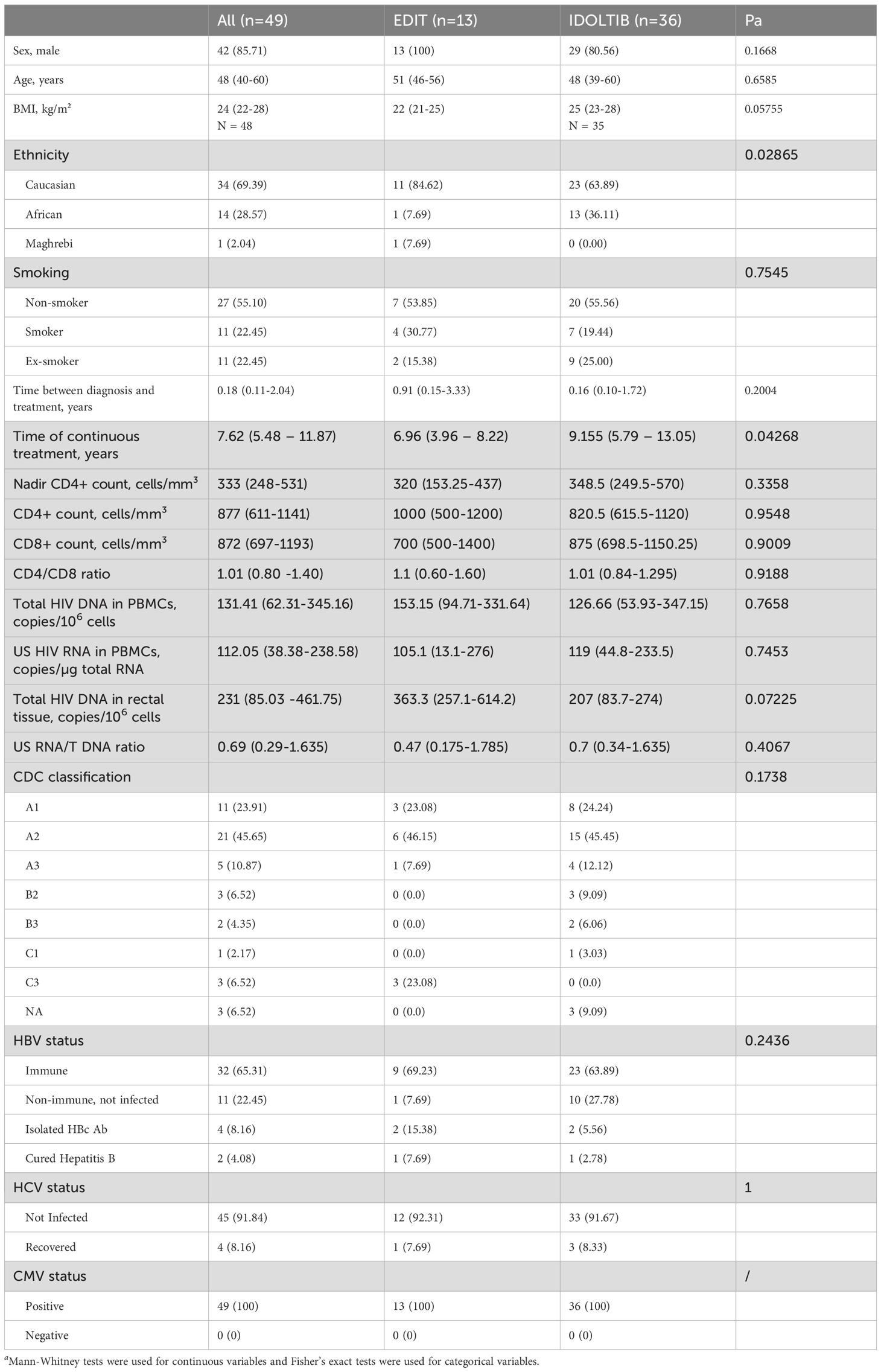
Table 1. Clinical characteristics of participants at baseline.
3.2 Associations between HIV persistence and inflammatory markers
The Spearman correlogram in Figure 1 demonstrates correlations between HIV persistence markers—including total HIV DNA (in PBMCs and rectal biopsies), unspliced (US) RNA (in PBMCs), and residual viremia—and various inflammatory markers.
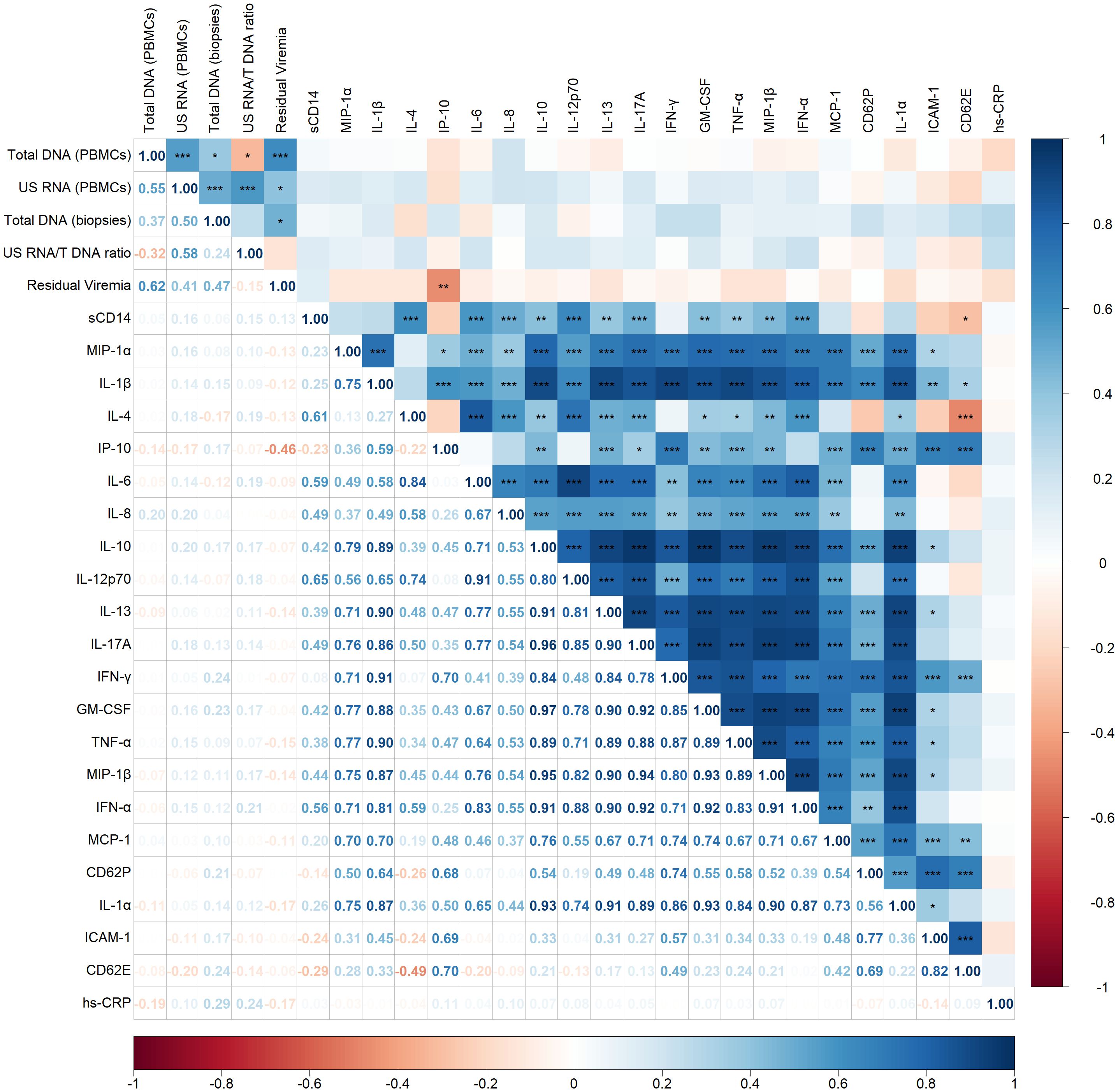
Figure 1. Spearman correlogram showing positive and negative correlations between HIV persistence and inflammatory markers. A heat map is used to indicate the strengths of associations between parameters with red representing negative correlations and blue representing positive correlations. The magnitude of the correlation coefficient, rho, indicates the degree of association, where values between 0.9 and 1.0 indicate very strong correlation, 0.7 to 0.9 indicate strong correlation, 0.5 to 0.7 indicate moderate correlation, 0.3 to 0.5 indicate weak correlation, and values below 0.3 indicate little to no linear correlation. Statistically significant correlations are marked with stars, where one star indicates p < 0.05, two stars indicate p < 0.01, and three stars indicate p < 0.001.
Moderate to strong positive correlations were observed among the HIV persistence markers. Specifically, total DNA in PBMCs correlated positively with US RNA (p < 0.001, rho = 0.55) and residual viremia (p < 0.001, rho = 0.62). A weak positive correlation was also noted between total DNA in PBMCs and in biopsies (p < 0.05, rho = 0.37). US RNA correlated positively with total DNA in biopsies (p < 0.001, rho = 0.50) and demonstrated a positive correlation with residual viremia (p < 0.05, rho = 0.41). Additionally, total DNA in biopsies positively correlated with residual viremia (p < 0.05, rho = 0.47).
Surprisingly, apart from a negative correlation observed between residual viremia and IP-10 (CXCL10) (p < 0.01, rho = -0.46), no other significant correlations were identified between virological and inflammatory markers. Most inflammatory markers displayed strong positive correlations with each other, reflecting a highly interconnected inflammatory network. For instance, sCD14, a marker of monocyte activation, exhibited strong positive correlations with several cytokines, including IL-4 (p < 0.001, rho = 0.61), IL-6 (p < 0.001, rho = 0.59), IL-8 (p < 0.001, rho = 0.49), IL-12p70 (p < 0.01, rho = 0.65), IL-17A (p < 0.01, rho = 0.49) and IFN-α (p < 0.001, rho = 0.56).
Regarding other inflammatory interactions, there was a negative correlation between IL-4 and CD62E (p < 0.001, rho = -0.49), indicating an inverse relationship between this anti-inflammatory cytokine and this endothelial cell activation marker.
3.3 Associations between HIV persistence markers and immune cell populations
Next, we examined correlations between HIV persistence markers and various immune cell populations including CD4 and CD8 T cells, B cells, monocytes and NK cells (Figure 2).
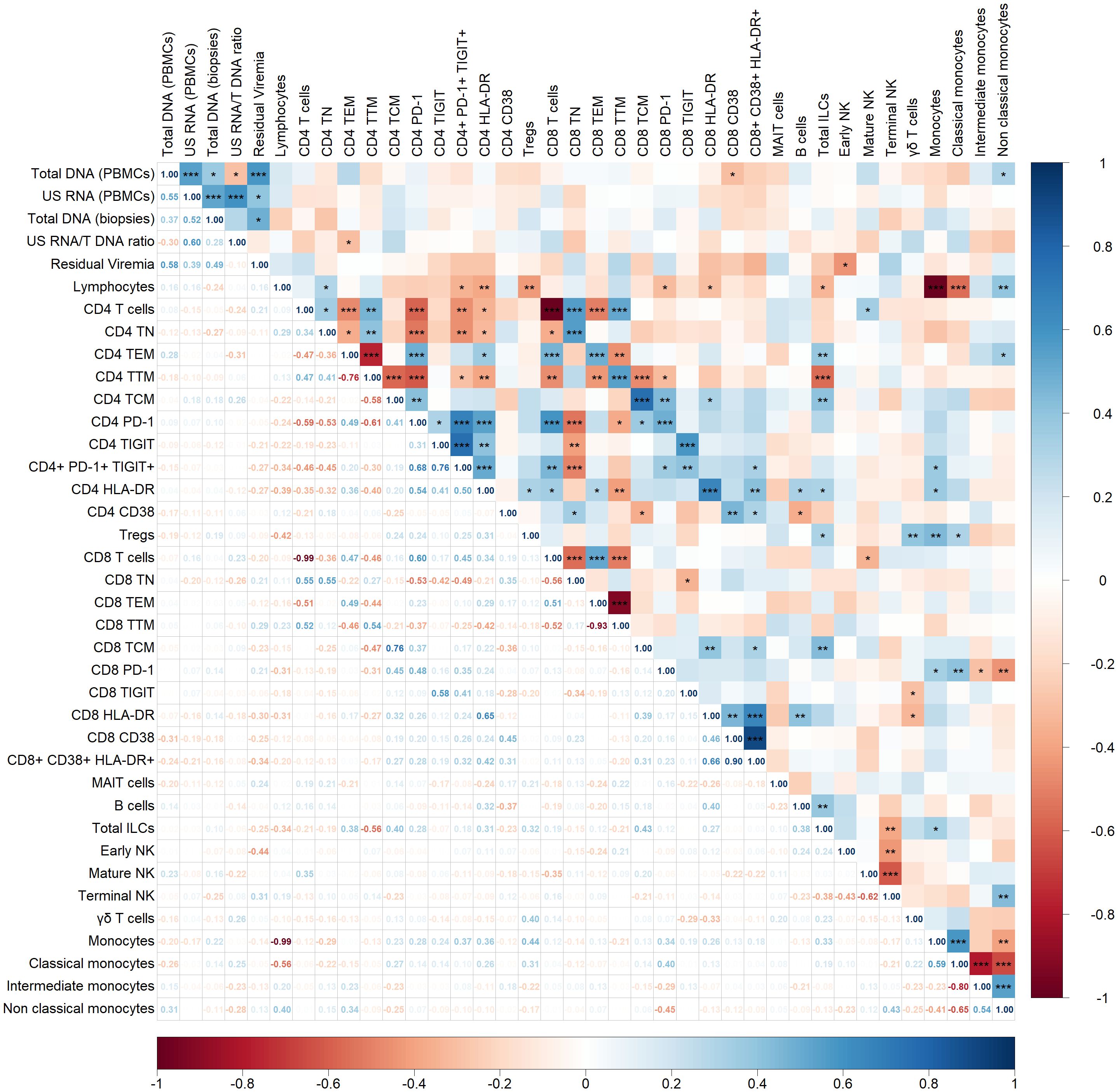
Figure 2. Spearman correlogram showing positive and negative correlations between HIV persistence markers and immune cell populations. A heat map is used to indicate the strengths of associations between parameters with red representing negative correlations and blue representing positive correlations. The magnitude of the correlation coefficient, rho, indicates the degree of association, where values between 0.9 and 1.0 indicate very strong correlation, 0.7 to 0.9 indicate strong correlation, 0.5 to 0.7 indicate moderate correlation, 0.3 to 0.5 indicate weak correlation, and values below 0.3 indicate little to no linear correlation. Statistically significant correlations are marked with stars, where one star indicates p < 0.05, two stars indicate p < 0.01, and three stars indicate p < 0.001.
Total DNA in PBMCs showed a statistically significant but weak negative correlation with CD8+ T cells expressing the activation marker CD38 (p < 0.05, rho = -0.31). Additionally, total DNA positively correlated with non-classical monocytes (p < 0.05, rho = 0.31). The US RNA/Total DNA ratio negatively correlated with CD4+ effector memory T cells (p < 0.05, rho = -0.31). Residual viremia demonstrated a negative correlation with early NK cell populations (p < 0.05, rho = -0.44). No other correlation or trend was observed between HIV persistence markers and immune cell populations in our cohort.
Within immune cell subsets, several significant correlations were observed. Focusing on a statistically strong correlation with p < 0.001, positive correlations were noted within T cell subsets. For example, CD4+ T naïve cells positively correlated with CD8+ T naïve cells (rho = 0.55), and similar patterns were observed across effector memory (rho = 0.49), transitional memory (rho = 0.54), and central memory (rho = 0.76) subsets.
Immune exhaustion and activation markers also displayed positive correlations between CD4+ and CD8+ T cell subsets. For instance, PD-1 expression on CD4+ T cells correlated strongly with PD-1 expression on CD8+ T cells (rho = 0.48), and similar correlations were observed for TIGIT (rho = 0.58) and HLA-DR (rho = 0.65).
Finally, a correlation was observed among monocyte subsets, including classical, intermediate, and non-classical monocytes.
3.4 Associations between HIV persistence markers and clinical factors
Figure 3 illustrates the associations between HIV persistence markers and various clinical factors, including demographic characteristics (age and BMI), and parameters related to infection history and treatment (nadir CD4, CD4/CD8 ratio, zenith plasma viral load pre-ART, duration of suppressive ART, time to treatment initiation).
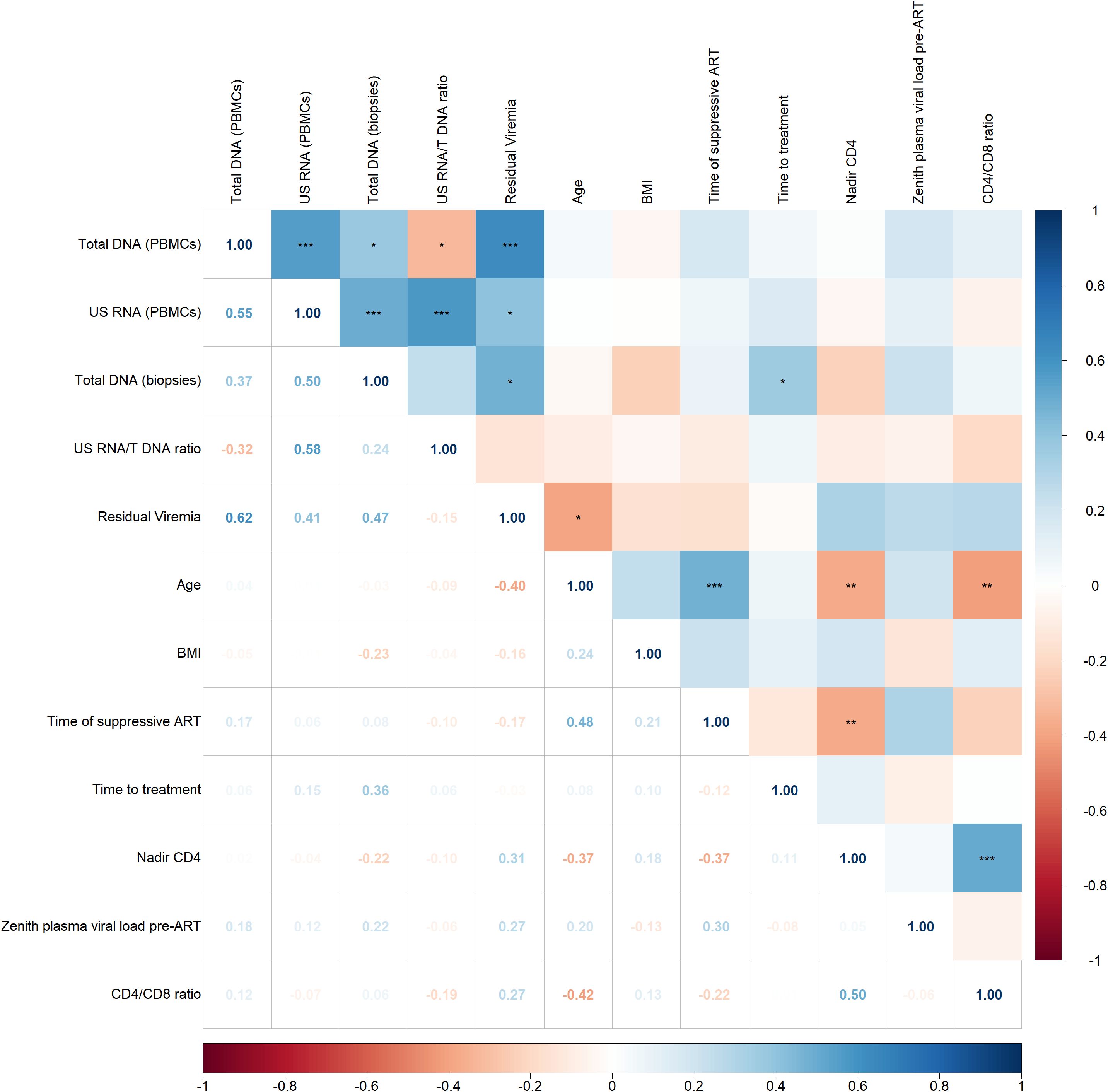
Figure 3. Spearman correlogram showing positive and negative correlations between HIV persistence markers and clinical factors. A heat map is used to indicate the strengths of associations between parameters with red representing negative correlations and blue representing positive correlations. The magnitude of the correlation coefficient, rho, indicates the degree of association, where values between 0.9 and 1.0 indicate very strong correlation, 0.7 to 0.9 indicate strong correlation, 0.5 to 0.7 indicate moderate correlation, 0.3 to 0.5 indicate weak correlation, and values below 0.3 indicate little to no linear correlation. Statistically significant correlations are marked with stars, where one star indicates p < 0.05, two stars indicate p < 0.01, and three stars indicate p < 0.001.
The correlogram reveals some statistically significant but weak correlations. Total HIV DNA in biopsies positively correlated with time on treatment (p < 0.05, rho = 0.36) and residual viremia negatively correlated with age (p < 0.05, rho = -0.40). No other significant correlations were observed between HIV persistence markers and clinical indicators.
Among clinical indicators and focusing on statistically strong correlations with p < 0.001, a significant positive correlation was found between nadir CD4 count and CD4/CD8 ratio (p < 0.001, rho = 0.50). Additionally, a positive correlation was observed between age and the duration of suppressive ART (p < 0.001, rho = 0.48). This reflects the expected trend that older individuals have been on suppressive ART for a longer period compared to younger individuals.
4 Discussion
In this study, we investigated the associations between HIV persistence markers, inflammatory markers, immune cell populations and clinical indicators in a cohort of 49 PWH on long-term DTG-based ART. The relationship between HIV persistence and immune activation has been a subject of debate, with the literature presenting conflicting findings.
Understanding the origins of persistent inflammation and immune activation in ART-treated PWH holds significant clinical implications. If inflammation is primarily driven by the HIV reservoir, therapeutic strategies should focus on reducing reservoir size or silencing its activity, for example, by inhibiting viral transcription (e.g., Tat inhibitors) or minimizing residual viral replication. Conversely, if the link between reservoirs and inflammation is weak, alternative drivers such as lifestyle factors (e.g., smoking, obesity), microbial translocation, co-infections, or other comorbidities must be prioritized. Identifying these drivers is essential for optimizing therapeutic interventions and improving long-term outcomes.
On the one hand, a significant portion of the literature supports a strong link between viral reservoirs and immune activation, suggesting an interdependent relationship between HIV persistence and inflammation (13). Early evidence from Chomont et al. demonstrated that HIV proviral DNA preferentially resides in CD4+ T cells expressing immune activation (PD-1) and proliferation (Ki67) markers (31). Further analyses across different compartments – blood, rectal tissue and lymph nodes – confirmed a positive correlation between the frequency of PD-1+ CD4+ T cells in blood and total HIV DNA, as well as between PD-1+ CD4+ T cells in rectal tissue and both integrated HIV DNA and CA US HIV RNA (32). Similarly, strong positive associations were observed in rectal tissue and lymph nodes between CD8+ T cells expressing HLA-DR and CD38 and both integrated HIV DNA and CA US HIV RNA. Later, Hatano et al. corroborated these findings, demonstrating significant associations between proviral DNA levels and the frequency of PD-1-expressing CD4+ T cells in blood samples. They further concluded that low CD4+ T cell count (<350 cells/mm3), despite suppressive therapy, was linked to higher CA RNA levels, proviral DNA levels and increased frequencies of CD4+ T cells expressing CD38, HLA-DR, CCR5 and/or PD-1 (33). The same group found a consistent association between CD4+ and CD8+ T cells expressing HLA-DR and the frequency of resting CD4+ T cells containing HIV DNA (34). In a more recent study conducted in 2021 by Olson et al., the focus shifted from total or integrated HIV DNA and CA RNA to intact proviral DNA (35). Interestingly, while intact proviral DNA did not correlate with plasma inflammation markers (e.g. D-dimer), a significant correlation was observed with intracellular HIV RNA. This finding suggests that the viral transcriptional activity, rather than the presence of intact DNA, may play a more important role in driving inflammation in ART-treated individuals. Furthermore, the study linked aging to reduced control over HIV-1 transcription, highlighting how age-related immune changes may exacerbate viral transcription and its inflammatory consequences in long-term ART-treated individuals. In line with these results, Scherpenisse et al. demonstrated that HIV-1 transcription at an early time point during ART correlates with markers of immune activation, exhaustion, and apoptosis (36).
Another critical marker of HIV persistence, low-level residual viremia, has been associated with microbial translocation and systemic inflammation (37–39). Riddler et al. found that residual viremia was associated with higher CD8+ T cell counts and lower CD4/CD8 ratio in ART-treated individuals, suggesting incomplete immune recovery (40). In a study by Falasca et al., residual viremia showed a significant correlation with sCD14 levels, an established marker of microbial translocation and monocyte activation (41). However, this study found no correlations between HIV DNA, HIV RNA, and inflammatory markers, highlighting inconsistencies with previous findings. Finally, several studies on raltegravir intensification have reported reductions in HIV replication markers, such as 2-LTR circles or US HIV RNA, along with decreased immune activation levels (42–49).
On the other hand, some intensification studies have failed to replicate these findings (50–56) and several studies have not consistently found a strong relationship between HIV persistence markers and immune activation (35, 57–63). For example, Gandhi et al. proposed that inflammation and immune activation in ART-treated individuals might predominantly come from immunologic events occurring before ART initiation rather than being perpetuated by ongoing viral replication sustained by persistent low-level viremia during suppressive ART (63). Recently, Bailon et al. demonstrated that a larger reservoir size was initially associated with higher levels of soluble inflammatory biomarkers, as well as increased activation and exhaustion of CD4+ and CD8+ T cells (64). However, these associations disappeared upon achieving viral suppression, regardless of the antiretroviral regimen used (dual or triple therapy). Similarly, previous comparative studies between two-drug regimens (2DRs) and three-drug regimens (3DRs) have not shown consistent differences in inflammatory markers, residual viremia or T-cell activation and exhaustion markers (65–68).
Overall, our findings revealed limited association between HIV persistence markers – including both blood and tissue reservoirs – and systemic inflammation or immune activation. However, some of our observations aligned with previous research. Notably, we observed an association between NK cells and residual viremia, a relationship that has been documented in both adults and children (69–72). This finding underscores the pivotal role of NK cells as key effectors of the innate immune system, highlighting their potential contribution to the control and modulation of HIV reservoir cell dynamics.
Several factors could explain the lack of a strong link between HIV persistence and inflammation in our study. First, compared to earlier studies from Chomont and Hatano, our cohort exhibited relatively low reservoir sizes, with a median total HIV DNA of 131 copies/106 cells in PBMCs and 231 copies/106 cells in rectal tissue (31, 33). However, in comparison to the LoViret cohort described by Galvez et al., which reported exceptionally low reservoir levels in chronically treated HIV-1 infected individuals (<50 HIV-DNA copies/106 PBMC), the reservoir sizes in our cohort were comparatively higher (73). Reservoir activity, as measured by unspliced RNA levels, was also limited (median: 112 copies/µg total RNA). Although direct comparison with earlier studies is challenging due to differences in sampling methods (e.g., CD4-enriched populations vs. PBMCs) and techniques, these values are in line with limited reservoir size and activity in these participants. However, in comparison to a recent study of early treated adolescents living with HIV and receiving long-term suppressive ART, our cohort exhibited higher levels of CA US HIV RNA, as that study reported a median of 19 copies/106 PBMCs (74).
These discrepancies likely reflect key changes in the management of PWH, including the initiation of treatment regardless of CD4+ T cell count and advancements in ART regimens, particularly the widespread adoption of integrase inhibitor-based ART (75). The impact of these modifications in PWH care on HIV reservoir size and activity should not be underestimated. In recent cohorts, these changes have resulted in prolonged viral suppression, such as ours (median duration: 7.62 years), and treatment with more potent regimens such as the DTG-based regimen that was used by all participants of the present study.
Longitudinal studies evaluating the dynamics of HIV reservoir cells and the impact of the host immune system have recently emerged, alongside new technologies like the intact proviral DNA assay (IPDA) and near full-length individual proviral next-generation sequencing. These PCR-based methods have revealed significant differences in the longitudinal dynamics of intact versus defective proviruses. Intact proviruses decline more rapidly, likely due to the active immunological elimination of transcriptionally active reservoirs, which are more easily recognized and targeted by the immune system (76). Defective proviruses that encode proteins also exhibit a shorter half-life compared to those incapable of protein expression (77, 78). Over time, the HIV reservoir is thus progressively reshaped into a more silent and defective state under the persistent pressure of the immune system, which preferentially targets infected cells with higher residual proviral transcriptional activity (79).
We thus hypothesize that, in virally suppressed individuals, the contribution of the HIV reservoir to chronic immune activation and inflammation will diminish over time on suppressive ART.
In our cohort, this is likely reflected by a reservoir dominated by deeply latent or transcriptionally silent clones which contribute minimally to inflammation and immune activation. This aligns with the prolonged duration of viral suppression and the robust immune recovery observed in our participants (median CD4+ counts: 877 cells/mm3 and CD4/CD8 ratio ~ 1). In addition, all participants in this study were receiving an integrase inhibitor-based regimen. Integrase inhibitor-based therapies are known to achieve faster and probably stronger viral suppression compared to protease inhibitor or non-nucleoside reverse transcriptase inhibitor-based regimens (80–82).
These findings reinforce the idea that inflammation and immune activation in ART-treated PWH are influenced by a multifaceted interplay of factors extending beyond HIV persistence alone. Moreover, they suggest that the contribution of the viral reservoir to inflammation may vary substantially between individuals and even in the same individuals over time, depending on the clonal dynamics that dominate the proviral landscape. Such variability highlights the importance of considering both host and viral factors when evaluating the drivers of inflammation in ART-treated populations.
While our findings are informative, several limitations should be acknowledged. First, the modest sample size (n=49) may have limited statistical power to detect subtle correlations. Second, we did not evaluate the impact of common sexually transmitted infections (e.g., chlamydia, gonorrhoea, syphilis) or examine gut-microbiome dysbiosis in detail, even though emerging evidence links microbiome alterations to chronic inflammation and non-communicable diseases (83, 84). Third, cell-associated unspliced HIV RNA could not be measured reliably in rectal tissue. Fourth, key immunological features such as immune-senescence markers and the Th17/Treg balance were not assessed. Taken together, these constraints should be borne in mind when interpreting the study’s conclusions.
In conclusion, the narrative surrounding HIV persistence and its impact on inflammation has shifted. Earlier studies described a scenario where larger, more active reservoirs perpetuated immune activation and inflammation. In contrast, today’s PWH, treated earlier with more potent regimens, have smaller, less active reservoirs that exert a diminished impact on immune activation. This highlights the dynamic nature of reservoir evolution under long-term ART and underscores the importance of considering these shifts when interpreting contemporary findings.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by Liège University Hospital-Faculty Ethics Committee. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
CF-L: Writing – original draft. DA: Writing – review & editing. MM: Writing – review & editing. AP: Writing – review & editing. GD: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. CF-L and DA are FRIA fellows of the Belgian Fund for Scientific Research (Fonds de la Recherche Scientifique – FNRS) and are supported by the Fondation Léon Fredericq. AP acknowledges grant support from amfAR, The Foundation for AIDS Research (grant no. 1110680–77-RPRL), and from Partnership NWO-Dutch AIDS Fonds ‘HIV cure for everyone’ (grant no. KICH2.V4P.AF23.001). GD is supported by the Belgian Fund for Scientific Research (Fonds de la Recherche Scientifique –FNRS).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1628086/full#supplementary-material
References
1. WHO. Fact sheet, HIV and AIDS (2024). Available online at: https://www.who.int/news-room/fact-sheets/detail/hiv-aids (Accessed January 2025).
2. Deeks SG, Lewin SR, and Havlir DV. The end of AIDS: HIV infection as a chronic disease. Lancet. (2013) 382:1525–33. doi: 10.1016/S0140-6736(13)61809-7
3. Trickey A, Sabin CA, Burkholder G, Crane H, d’Arminio Monforte A, Egger M, et al. Life expectancy after 2015 of adults with HIV on long-term antiretroviral therapy in Europe and North America: a collaborative analysis of cohort studies. Lancet HIV. (2023) 10:e295–307. doi: 10.1016/S2352-3018(23)00028-0
4. Hunt PW. HIV and inflammation: mechanisms and consequences. Curr HIV/AIDS Rep. (2012) 9:139–47. doi: 10.1007/s11904-012-0118-8
5. Wilson EMP and Sereti I. Immune restoration after antiretroviral therapy: the pitfalls of hasty or incomplete repairs. Immunol Rev. (2013) 254:343–54. doi: 10.1111/imr.12064
6. Yang X, Su B, Zhang X, Liu Y, Wu H, and Zhang T. Incomplete immune reconstitution in HIV/AIDS patients on antiretroviral therapy: Challenges of immunological non-responders. J Leukoc Biol. (2020) 107:597–612. doi: 10.1002/JLB.4MR1019-189R
7. Hileman CO and Funderburg NT. Inflammation, immune activation, and antiretroviral therapy in HIV. Curr HIV/AIDS Rep. (2017) 14:93–100. doi: 10.1007/s11904-017-0356-x
8. Sokoya T, Steel HC, Nieuwoudt M, and Rossouw TM. HIV as a cause of immune activation and immunosenescence. Mediators Inflammation. (2017) 2017:6825493. doi: 10.1155/2017/6825493
9. Younas M, Psomas C, Reynes J, and Corbeau P. Immune activation in the course of HIV-1 infection: Causes, phenotypes and persistence under therapy. HIV Med. (2016) 17:89–105. doi: 10.1111/hiv.12310
10. Siliciano JD and Siliciano RF. In vivo dynamics of the latent reservoir for HIV-1: new insights and implications for cure. Annu Rev Pathol. (2022) 17:271–94. doi: 10.1146/annurev-pathol-050520-112001
11. Pasternak AO and Berkhout B. HIV persistence: silence or resistance? Curr Opin Virol. (2023) 59:101301. doi: 10.1016/j.coviro.2023.101301
12. Klatt NR, Chomont N, Douek DC, and Deeks SG. Immune activation and HIV persistence: implications for curative approaches to HIV infection. Immunol Rev. (2013) 254:326–42. doi: 10.1111/imr.12065
13. Massanella M, Fromentin R, and Chomont N. Residual inflammation and viral reservoirs: alliance against an HIV cure. Curr Opin HIV AIDS. (2016) 11:234. doi: 10.1097/COH.0000000000000230
14. Martinez-Picado J and Deeks SG. Persistent HIV-1 replication during antiretroviral therapy. Curr Opin HIV AIDS. (2016) 11:417–23. doi: 10.1097/COH.0000000000000287
15. Zicari S, Sessa L, Cotugno N, Ruggiero A, Morrocchi E, Concato C, et al. Immune activation, inflammation, and non-AIDS co-morbidities in HIV-infected patients under long-term ART. Viruses. (2019) 11:200. doi: 10.3390/v11030200
16. Lv T, Cao W, and Li T. HIV-related immune activation and inflammation: current understanding and strategies. J Immunol Res. (2021) 2021:7316456. doi: 10.1155/2021/7316456
17. De Voeght A, Martens H, Renard C, Vaira D, Debruche M, Simonet J, et al. Exploring the link between innate immune activation and thymic function by measuring sCD14 and TRECs in HIV patients living in Belgium. PloS One. (2017) 12:e0185761. doi: 10.1371/journal.pone.0185761
18. Streeck H, Maestri A, Habermann D, Crowell TA, Esber AL, Son G, et al. Dissecting drivers of immune activation in chronic HIV-1 infection. EBioMedicine. (2022) 83:104182. doi: 10.1016/j.ebiom.2022.104182
19. Fromentin R, Bakeman W, Lawani MB, Khoury G, Hartogensis W, DaFonseca S, et al. CD4+ T cells expressing PD-1, TIGIT and LAG-3 contribute to HIV persistence during ART. PloS Pathog. (2016) 12:e1005761. doi: 10.1371/journal.ppat.1005761
20. Chiu CY, Schou MD, McMahon JH, Deeks SG, Fromentin R, Chomont N, et al. Soluble immune checkpoints as correlates for HIV persistence and T cell function in people with HIV on antiretroviral therapy. Front Immunol. (2023) 14:1123342. doi: 10.3389/fimmu.2023.1123342
21. Naidoo KK, Ndumnego OC, Ismail N, Dong KL, and Ndung’u T. Antigen presenting cells contribute to persistent immune activation despite antiretroviral therapy initiation during hyperacute HIV-1 infection. Front Immunol. (2021) 12. doi: 10.3389/fimmu.2021.738743
22. Muñoz-Muela E, Trujillo-Rodríguez M, Serna-Gallego A, Saborido-Alconchel A, Gasca-Capote C, Álvarez-Ríos A, et al. HIV-1-DNA/RNA and immunometabolism in monocytes: contribution to the chronic immune activation and inflammation in people with HIV-1. EBioMedicine. (2024) 108:105338. doi: 10.1016/j.ebiom.2024.105338
23. Guaraldi G, Orlando G, Zona S, Menozzi M, Carli F, Garlassi E, et al. Premature age-related comorbidities among HIV-infected persons compared with the general population. Clin Infect Dis. (2011) 53:1120–6. doi: 10.1093/cid/cir627
25. Chen J, Titanji K, Sheth AN, Gandhi R, McMahon D, Ofotokun I, et al. The effect of age on CD4+ T-cell recovery in HIV-suppressed adult participants: a sub-study from AIDS Clinical Trial Group (ACTG) A5321 and the Bone Loss and Immune Reconstitution (BLIR) study. Immun Ageing. (2022) 19:4. doi: 10.1186/s12979-021-00260-x
26. Mazzuti L, Turriziani O, and Mezzaroma I. The many faces of immune activation in hiv-1 infection: a multifactorial interconnection. Biomedicines. (2023) 11:159. doi: 10.3390/biomedicines11010159
27. Scully EP, Gandhi M, Johnston R, Hoh R, Lockhart A, Dobrowolski C, et al. Sex-based differences in human immunodeficiency virus type 1 reservoir activity and residual immune activation. J Infect Dis. (2019) 219:1084–94. doi: 10.1093/infdis/jiy617
28. Mihealsick E, Word A, and Scully EP. The impact of sex on HIV immunopathogenesis and therapeutic interventions. J Clin Invest. (2024) 134:e180075. doi: 10.1172/JCI180075
29. Fombellida-Lopez C, Valaitienė A, Winchester L, Maes N, Dellot P, Vanwinge C, et al. Doubling dolutegravir dosage reduces the viral reservoir in ART-treated people with HIV. (2009). doi: 10.7554/eLife.106931.1
30. Fombellida-Lopez C, Östling E, Aguilar Ortmans D, Dellot P, Marechal N, Vanwinge C, et al. Impact of Abacavir Withdrawal from a Triple ART Regimen on HIV Reservoir and Systemic Inflammation: A Phase 3 Randomised Controlled Trial. SSRN Scholarly Paper. (2025). doi: 10.2139/ssrn.5264475
31. Chomont N, El-Far M, Ancuta P, Trautmann L, Procopio FA, Yassine-Diab B, et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat Med. (2009) 15:893–900. doi: 10.1038/nm.1972
32. Khoury G, Fromentin R, Solomon A, Hartogensis W, Killian M, Hoh R, et al. Human immunodeficiency virus persistence and T-cell activation in blood, rectal, and lymph node tissue in human immunodeficiency virus–infected individuals receiving suppressive antiretroviral therapy. J Infect Dis. (2017) 215:911–9. doi: 10.1093/infdis/jix039
33. Hatano H, Jain V, Hunt PW, Lee T-H, Sinclair E, Do TD, et al. Cell-based measures of viral persistence are associated with immune activation and programmed cell death protein 1 (PD-1)–expressing CD4+ T cells. J Infect Dis. (2013) 208:50–6. doi: 10.1093/infdis/jis630
34. Cockerham LR, Siliciano JD, Sinclair E, O’Doherty U, Palmer S, Yukl SA, et al. CD4+ and CD8+ T cell activation are associated with HIV DNA in resting CD4+ T cells. PloS One. (2014) 9:e110731. doi: 10.1371/journal.pone.0110731
35. Olson A, Coote C, Snyder-Cappione JE, Lin N, and Sagar M. HIV-1 transcription but not intact provirus levels are associated with systemic inflammation. J Infect Dis. (2021) 223:1934–42. doi: 10.1093/infdis/jiaa657
36. Scherpenisse M, Kootstra NA, Bakker M, Berkhout B, and Pasternak AO. Cell-associated HIV-1 unspliced-to-multiply-spliced RNA ratio at 12 weeks of ART predicts immune reconstitution on therapy. mBio. (2021) 12:e00099–21. doi: 10.1128/mBio.00099-21
37. Eastburn A, Scherzer R, Zolopa AR, Benson C, Tracy R, Do T, et al. Association of low level viremia with inflammation and mortality in HIV-infected adults. PloS One. (2011) 6:e26320. doi: 10.1371/journal.pone.0026320
38. Reus S, Portilla J, Sánchez-Payá J, Giner L, Francés R, Such J, et al. Low-level HIV viremia is associated with microbial translocation and inflammation. J Acquir Immune Defic Syndr. (2013) 62:129–34. doi: 10.1097/QAI.0b013e3182745ab0
39. Younas M, Psomas C, Reynes C, Cezar R, Kundura L, Portalès P, et al. Residual viremia is linked to a specific immune activation profile in HIV-1-infected adults under efficient antiretroviral therapy. Front Immunol. (2021) 12:663843. doi: 10.3389/fimmu.2021.663843
40. Riddler SA, Aga E, Bosch RJ, Bastow B, Bedison M, Vagratian D, et al. Continued slow decay of the residual plasma viremia level in HIV-1-infected adults receiving long-term antiretroviral therapy. J Infect Dis. (2016) 213:556–60. doi: 10.1093/infdis/jiv433
41. Falasca F, Carlo DD, Vito CD, Bon I, d’Ettorre G, Fantauzzi A, et al. Evaluation of HIV-DNA and inflammatory markers in HIV-infected individuals with different viral load patterns. BMC Infect Dis. (2017) 17:581. doi: 10.1186/s12879-017-2676-2
42. Buzon MJ, Massanella M, Llibre JM, Esteve A, Dahl V, Puertas MC, et al. HIV-1 replication and immune dynamics are affected by raltegravir intensification of HAART-suppressed subjects. Nat Med. (2010) 16:460–5. doi: 10.1038/nm.2111
43. Yukl SA, Shergill AK, McQuaid K, Gianella S, Lampiris H, Hare CB, et al. Effect of raltegravir-containing intensification on HIV burden and T-cell activation in multiple gut sites of HIV-positive adults on suppressive antiretroviral therapy. AIDS. (2010) 24:2451–60. doi: 10.1097/QAD.0b013e32833ef7bb
44. Hatano H, Strain MC, Scherzer R, Bacchetti P, Wentworth D, Hoh R, et al. Increase in 2-long terminal repeat circles and decrease in D-dimer after raltegravir intensification in patients with treated HIV infection: a randomized, placebo-controlled trial. J Infect Dis. (2013) 208:1436–42. doi: 10.1093/infdis/jit453
45. Llibre JM, Buzón MJ, Massanella M, Esteve A, Dahl V, Puertas MC, et al. Treatment intensification with raltegravir in subjects with sustained HIV-1 viraemia suppression: a randomized 48-week study. Antivir Ther. (2012) 17:355–64. doi: 10.3851/IMP1917
46. Vallejo A, Gutierrez C, Hernandez-Novoa B, Diaz L, Madrid N, Abad-Fernandez M, et al. The effect of intensification with raltegravir on the HIV-1 reservoir of latently infected memory CD4 T cells in suppressed patients. AIDS. (2012) 26:1885–94. doi: 10.1097/QAD.0b013e3283584521
47. Gutiérrez C, Díaz L, Vallejo A, Hernández-Novoa B, Abad M, Madrid N, et al. Intensification of antiretroviral therapy with a CCR5 antagonist in patients with chronic HIV-1 infection: effect on T cells latently infected. PloS One. (2011) 6:e27864. doi: 10.1371/journal.pone.0027864
48. Puertas MC, Massanella M, Llibre JM, Ballestero M, Buzon MJ, Ouchi D, et al. Intensification of a raltegravir-based regimen with maraviroc in early HIV-1 infection. AIDS. (2014) 28:325–34. doi: 10.1097/QAD.0000000000000066
49. Puertas MC, Gómez-Mora E, Santos JR, Molto J, Urrea V, Morón-López S, et al. Impact of intensification with raltegravir on HIV-1-infected individuals receiving monotherapy with boosted PIs. J Antimicrob Chemother. (2018) 73:1940–8. doi: 10.1093/jac/dky106
50. Dinoso JB, Kim SY, Wiegand AM, Palmer SE, Gange SJ, Cranmer L, et al. Treatment intensification does not reduce residual HIV-1 viremia in patients on highly active antiretroviral therapy. Proc Natl Acad Sci U.S.A. (2009) 106:9403–8. doi: 10.1073/pnas.0903107106
51. McMahon D, Jones J, Wiegand A, Gange SJ, Kearney M, Palmer S, et al. Short-course raltegravir intensification does not reduce persistent low-level viremia in patients with HIV-1 suppression during receipt of combination antiretroviral therapy. Clin Infect Dis. (2010) 50:912–9. doi: 10.1086/650749
52. Gandhi RT, Zheng L, Bosch RJ, Chan ES, Margolis DM, Read S, et al. The effect of raltegravir intensification on low-level residual viremia in HIV-infected patients on antiretroviral therapy: a randomized controlled trial. PloS Med. (2010) 7:e1000321. doi: 10.1371/journal.pmed.1000321
53. Gandhi RT, Coombs RW, Chan ES, Bosch RJ, Zheng L, Margolis DM, et al. No effect of raltegravir intensification on viral replication markers in the blood of HIV-1-infected patients receiving antiretroviral therapy. J Acquir Immune Defic Syndr. (2012) 59:229–35. doi: 10.1097/QAI.0b013e31823fd1f2
54. Cillo AR, Hilldorfer BB, Lalama CM, McKinnon JE, Coombs RW, Tenorio AR, et al. Virologic and immunologic effects of adding maraviroc to suppressive antiretroviral therapy in individuals with suboptimal CD4+ T-cell recovery. AIDS. (2015) 29:2121–9. doi: 10.1097/QAD.0000000000000810
55. Chaillon A, Gianella S, Lada SM, Perez-Santiago J, Jordan P, Ignacio C, et al. Size, composition, and evolution of HIV DNA populations during early antiretroviral therapy and intensification with maraviroc. J Virol. (2018) 92:e01589–17. doi: 10.1128/JVI.01589-17
56. Rasmussen TA, McMahon JH, Chang JJ, Audsley J, Rhodes A, Tennakoon S, et al. The effect of antiretroviral intensification with dolutegravir on residual virus replication in HIV-infected individuals: a randomised, placebo-controlled, double-blind trial. Lancet HIV. (2018) 5:e221–30. doi: 10.1016/S2352-3018(18)30040-7
57. Steel A, Cox AE, Shamji MH, John L, Nelson M, Henderson DC, et al. HIV-1 viral replication below 50 copies/ml in patients on antiretroviral therapy is not associated with CD8+ T-cell activation. Antivir Ther. (2007) 12:971–5. doi: 10.1177/135965350701200613
58. Chun T-W, Murray D, Justement JS, Hallahan CW, Moir S, Kovacs C, et al. Relationship between residual plasma viremia and the size of HIV proviral DNA reservoirs in infected individuals receiving effective antiretroviral therapy. J Infect Dis. (2011) 204:135–8. doi: 10.1093/infdis/jir208
59. Mexas AM, Graf EH, Pace MJ, Yu JJ, Papasavvas E, Azzoni L, et al. Concurrent measures of total and integrated HIV DNA monitor reservoirs and ongoing replication in eradication trials. AIDS. (2012) 26:2295–306. doi: 10.1097/QAD.0b013e32835a5c2f
60. Allavena C, Rodallec A, Sécher S, Reliquet V, Baffoin S, André-Garnier E, et al. Evaluation of residual viremia and quantitation of soluble CD14 in a large cohort of HIV-infected adults on a long-term non-nucleoside reverse transcriptase inhibitor-based regimen. J Med Virol. (2013) 85:1878–82. doi: 10.1002/jmv.23679
61. Poizot-Martin I, Faucher O, Obry-Roguet V, Nicolino-Brunet C, Ronot-Bregigeon S, Dignat-George F, et al. Lack of correlation between the size of HIV proviral DNA reservoir and the level of immune activation in HIV-infected patients with a sustained undetectable HIV viral load for 10 years. J Clin Virol. (2013) 57:351–5. doi: 10.1016/j.jcv.2013.04.007
62. Guihot A, Dentone C, Assoumou L, Parizot C, Calin R, Seang S, et al. Residual immune activation in combined antiretroviral therapy-treated patients with maximally suppressed viremia. AIDS. (2016) 30:327–30. doi: 10.1097/QAD.0000000000000815
63. Gandhi RT, McMahon DK, Bosch RJ, Lalama CM, Cyktor JC, Macatangay BJ, et al. Levels of HIV-1 persistence on antiretroviral therapy are not associated with markers of inflammation or activation. PloS Pathog. (2017) 13:e1006285. doi: 10.1371/journal.ppat.1006285
64. Bailón L, Puertas MC, García-Guerrero MC, Moraes-Cardoso I, Aparicio E, Alarcón-Soto Y, et al. Impact of dolutegravir plus lamivudine as first-line antiretroviral treatment on HIV-1 reservoir and inflammatory markers in peripheral blood. J Infect Dis. (2024) 231:jiae530. doi: 10.1093/infdis/jiae530
65. Vassallo M, Durant J, Fabre R, Lotte L, Sindt A, Puchois A, et al. Inflammatory markers after switching to a dual drug regimen in HIV-infected subjects: A two-year follow-up. Viruses. (2022) 14:927. doi: 10.3390/v14050927
66. Wang R, Underwood M, Llibre JM, Bernal Morell E, Brinson C, Sanz Moreno J, et al. Very-low-level viremia, inflammatory biomarkers, and associated baseline variables: three-year results of the randomized TANGO study. Open Forum Infect Dis. (2024) 11:ofad626. doi: 10.1093/ofid/ofad626
67. Llibre JM, Cahn PE, Lo J, Barber TJ, Mussini C, van Welzen BJ, et al. Changes in inflammatory and atherogenesis biomarkers with the 2-drug regimen dolutegravir plus lamivudine in antiretroviral therapy-experienced, virologically suppressed people with HIV-1: A systematic literature review. Open Forum Infect Dis. (2022) 9:ofac068. doi: 10.1093/ofid/ofac068
68. Llibre JM, López Cortés LF, Aylott A, Wynne B, Matthews J, Van Solingen-Ristea R, et al. Brief report: evaluation of inflammation and atherogenesis biomarkers through 148 weeks postswitch to dolutegravir and rilpivirine in SWORD-1/SWORD-2. J Acquir Immune Defic Syndr. (2022) 91:73–8. doi: 10.1097/QAI.0000000000003019
69. Marras F, Casabianca A, Bozzano F, Ascierto ML, Orlandi C, Di Biagio A, et al. Control of the HIV-1 DNA reservoir is associated in vivo and in vitro with NKp46/NKp30 (CD335 CD337) inducibility and interferon gamma production by transcriptionally unique NK cells. J Virol. (2017) 91:e00647–17. doi: 10.1128/JVI.00647-17
70. Hartana CA, Garcia-Broncano P, Rassadkina Y, Lian X, Jiang C, Einkauf KB, et al. Immune correlates of HIV-1 reservoir cell decline in early-treated infants. Cell Rep. (2022) 40:111126. doi: 10.1016/j.celrep.2022.111126
71. Garcia-Broncano P, Maddali S, Einkauf KB, Jiang C, Gao C, Chevalier J, et al. Early antiretroviral therapy in neonates with HIV-1 infection restricts viral reservoir size and induces a distinct innate immune profile. Sci Transl Med. (2019) 11:eaax7350. doi: 10.1126/scitranslmed.aax7350
72. Ivison GT, Vendrame E, Martínez-Colón GJ, Ranganath T, Vergara R, Zhao NQ, et al. Natural killer cell receptors and ligands are associated with markers of HIV-1 persistence in chronically infected ART suppressed patients. Front Cell Infect Microbiol. (2022) 12:757846. doi: 10.3389/fcimb.2022.757846
73. Gálvez C, Urrea V, Dalmau J, Jimenez M, Clotet B, Monceaux V, et al. Extremely low viral reservoir in treated chronically HIV-1-infected individuals. EBioMedicine. (2020) 57:102830. doi: 10.1016/j.ebiom.2020.102830
74. Gärtner K, Domínguez-Rodríguez S, Heaney J, Gkouleli T, Grant P, Dorgham K, et al. Low unspliced cell-associated HIV RNA in early treated adolescents living with HIV on long suppressive ART. Front Immunol. (2024) 15. doi: 10.3389/fimmu.2024.1334236
75. González-Navarro I, Urrea V, Gálvez C, Garcia-Guerrero MDC, Morón-López S, Puertas MC, et al. Assessing advances in three decades of clinical antiretroviral therapy on the HIV-1 reservoir. J Clin Invest. (2024) 135:e183952. doi: 10.1172/JCI183952
76. Lian X, Seiger KW, Parsons EM, Gao C, Sun W, Gladkov GT, et al. Progressive transformation of the HIV-1 reservoir cell profile over two decades of antiviral therapy. Cell Host Microbe. (2023) 31:83–96.e5. doi: 10.1016/j.chom.2022.12.002
77. Pinzone MR, VanBelzen DJ, Weissman S, Bertuccio MP, Cannon L, Venanzi-Rullo E, et al. Longitudinal HIV sequencing reveals reservoir expression leading to decay which is obscured by clonal expansion. Nat Commun. (2019) 10:728. doi: 10.1038/s41467-019-08431-7
78. Cho A, Gaebler C, Olveira T, Ramos V, Saad M, Lorenzi JCC, et al. Longitudinal clonal dynamics of HIV-1 latent reservoirs measured by combination quadruplex polymerase chain reaction and sequencing. Proc Natl Acad Sci U.S.A. (2022) 119:e2117630119. doi: 10.1073/pnas.2117630119
79. Armani-Tourret M, Bone B, Tan TS, Sun W, Bellefroid M, Struyve T, et al. Immune targeting of HIV-1 reservoir cells: a path to elimination strategies and cure. Nat Rev Microbiol. (2024) 22:328–44. doi: 10.1038/s41579-024-01010-8
80. Wiesmann F, Braun P, Knickmann M, and Knechten H. Low level HIV viremia is more frequent under protease-inhibitor containing firstline therapy than under NNRTI-regimens. J Int AIDS Soc. (2014) 17:19828. doi: 10.7448/IAS.17.4.19828
81. Darcis G, Maes N, Pasternak AO, Sauvage A-S, Frippiat F, Meuris C, et al. Detectability of HIV residual viremia despite therapy is highly associated with treatment with a protease inhibitor-based combination antiretroviral therapy. Antimicrob Agents Chemother. (2020) 64:e01902–19. doi: 10.1128/AAC.01902-19
82. Darcis G, Berkhout B, and Pasternak AO. Differences in HIV markers between infected individuals treated with different ART regimens: implications for the persistence of viral reservoirs. Viruses. (2020) 12:489. doi: 10.3390/v12050489
83. Ishizaka A, Koga M, Mizutani T, Suzuki Y, Matano T, Yotsuyanagi H, et al. Sustained gut dysbiosis and intestinal inflammation show correlation with weight gain in person with chronic HIV infection on antiretroviral therapy. BMC Microbiol. (2024) 24:274. doi: 10.1186/s12866-024-03431-0
Keywords: HIV reservoir, inflammation, immune activation, correlation, antiretroviral therapy
Citation: Fombellida-Lopez C, Aguilar Ortmans D, Moutschen M, Pasternak AO and Darcis G (2025) No associations between HIV reservoir and inflammation in long-term virally suppressed dolutegravir-based ART-treated individuals. Front. Immunol. 16:1628086. doi: 10.3389/fimmu.2025.1628086
Received: 14 May 2025; Accepted: 09 July 2025;
Published: 28 July 2025.
Edited by:
Hiromi Imamichi, National Institute of Allergy and Infectious Diseases (NIH), United StatesReviewed by:
Marta Catalfamo, Georgetown University Medical Center, United StatesOmbretta Turriziani, Sapienza University of Rome, Italy
Copyright © 2025 Fombellida-Lopez, Aguilar Ortmans, Moutschen, Pasternak and Darcis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexander O. Pasternak, YS5vLnBhc3Rlcm5ha0BhbXN0ZXJkYW11bWMubmw=; Gilles Darcis, Z2RhcmNpc0BjaHVsaWVnZS5iZQ==
†These authors have contributed equally to this work and share last authorship