 Mateusz Matwiejuk1*
Mateusz Matwiejuk1* Agnieszka Kulczyńska-Przybik2
Agnieszka Kulczyńska-Przybik2 Hanna Myśliwiec1
Hanna Myśliwiec1 Agnieszka Mikłosz3Adrian Chabowski3
Agnieszka Mikłosz3Adrian Chabowski3 Barbara Mroczko2Iwona Flisiak1
Barbara Mroczko2Iwona Flisiak1- 1Department of Dermatology and Venereology, Medical University of Bialystok, Bialystok, Poland
- 2Department of Neurodegeneration Diagnostics, Medical University of Bialystok, Bialystok, Poland
- 3Department of Physiology, Medical University of Bialystok, Bialystok, Poland
Atopic Dermatitis is a chronic skin condition characterized by inflammation and itching. It has a genetic component, but environmental factors also play a significant role. The immune system is overactive, leading to an abnormal inflammatory response. Literature data indicate that numerous proteins contribute to the development and progression of atopic dermatitis, like antimicrobial peptides, alarmins, autoantigens, cytokines, growth factors, and proteases. To synthesize current knowledge and identify the most promising contributors of AD pathogenesis a literature search was conducted using PubMed (1990–present), Google Scholar, and Embase, has been performed appropriate search terms. This narrative review summarizes the current knowledge on how elafin, chemerin, and nicotinamide phosphoribosyltransferase (visfatin/NAMPT) contribute to the pathophysiology of skin inflammation in atopic dermatitis. Recent discoveries have highlighted the importance of these proteins as important players in the functioning of the epidermal barrier. Importantly, some proteins exert anti-inflammatory effects (e.g., elafin), some pro-inflammatory effects, such as visfatin/NAMPT or chemerin, which exhibits both pro- and anti-inflammatory properties. This makes them intriguing candidates for modulating the complex inflammatory processes associated with atopic dermatitis. A deeper understanding of the role of these proteins may provide a basis for the development of appropriate treatments for atopic dermatitis. However, knowledge about the importance of these proteins in the pathological mechanisms of atopic dermatitis is still limited.
1 Introduction
1.1 Epidemiology
Atopic dermatitis (AD), also known as an atopic eczema, is the most common chronic, allergic, inflammatory skin disease worldwide (1). The significant burden that AD places on both healthcare funding and the quality of life of patients prompts a deeper understanding of the pathomechanisms of this disease (2). In 2021, the global number of pediatric AD cases reached 72.4 million, a 6.2% increase from 2000 (3). In adults, the prevalence of AD estimated between 3.4% in Israel to 33.7% in Thailand. Additionally, it was revealed that the prevalence was generally higher in females compared to males (4).
1.2 Symptoms
The “atopic march” describes the progression of atopic diseases, often starting with AD in infancy or early childhood, followed by the development of food allergies, allergic rhinitis (hay fever), and asthma (5). Eczematous skin, the type of inflammation seen in AD, is often described as redness, swelling, and oozing. The skin rashes and lesions appear in similar patterns on both sides of the body. Importantly, skin lesions in AD are characterized by age-dependent locations. The area’s most commonly affected by AD change with age: in infants these are: face, scalp, extensor surfaces of extremities (elbows, knees); in children: flexural areas (creases of elbows, knees, neck); and in adults: hands, feet, eyelids (6). Hanifin and Rajka Score are set of various criteria, which is a well-known and widely used method for diagnosing AD. To recognize AD is required to have three of four major criteria and three of twenty-three minor criteria (7). Moreover, AD is often associated with significant pruritus, which can significantly impact the quality of life of those who suffer from it. Scratching can further worsen the condition and impede healing (8).
1.3 Pathophysiology
The pathogenesis of AD comprises a complex interplay of factors, including impairments in both the innate and adaptive immune responses (9). These immune dysfunctions, combined with a compromised skin barrier, contribute to the inflammation and skin lesions characteristic of the condition. When the skin barrier is compromised, keratinocytes (the main cells of the epidermis) release increased amounts of certain cytokines, including thymic stromal lymphopoietin (TSLP) and interleukins such as IL-25 and IL-33. These cytokines play a role in activating the immune system, particularly T helper 2 (Th2) cells, and are involved in promoting type 2 immune responses Particularly TSLP initiates a cascade of events. Firstly, TSLP stimulates dendritic cells to express OX40L, which then interacts with OX40 on T cells, promoting Th2 cell differentiation. Secondly, this Th2 cell activation by TSLP, leads to the production of inflammatory cytokines like IL-4, IL-5, and IL-13, characteristic of type 2 inflammation (10). Furthermore, filaggrin (FLG) is a protein crucial for maintaining the skin’s barrier function. Mutations in the FLG gene are strongly associated with the development of AD. These mutations disrupt the skin barrier, leading to: an increased transepidermal water loss (TEWL) (presented as a dry, cracked skin); an increased penetration of allergens and irritants: (showed as triggering inflammation); and dysregulation of the immune system (11). Moreover, higher FLG mRNA levels were found in patients with severe AD compared to patients with moderate AD, suggesting that the body attempts to compensate for the weakened skin barrier by increasing FLG production (12).
Recent studies have shown that elafin (13), chemerin (14) and nicotinamide phosphoribosyltransferase (NAMPT)/visfatin (15) may be potential key players in the pathogenesis of AD. The potential mechanism of their action in AD is shown on the Figure 1. Briefly, elafin is a protein that acts as a serine protease inhibitor and is involved in the body’s defense against inflammation. Chemerin is involved in immune cell recruitment and inflammation, which are also characteristic of AD. In turn, NAMPT, also known as visfatin, is an enzyme involved in NAD+ biosynthesis and has pro-inflammatory properties. The link between these proteins and AD suggests that they may be novel targets for therapeutic intervention.
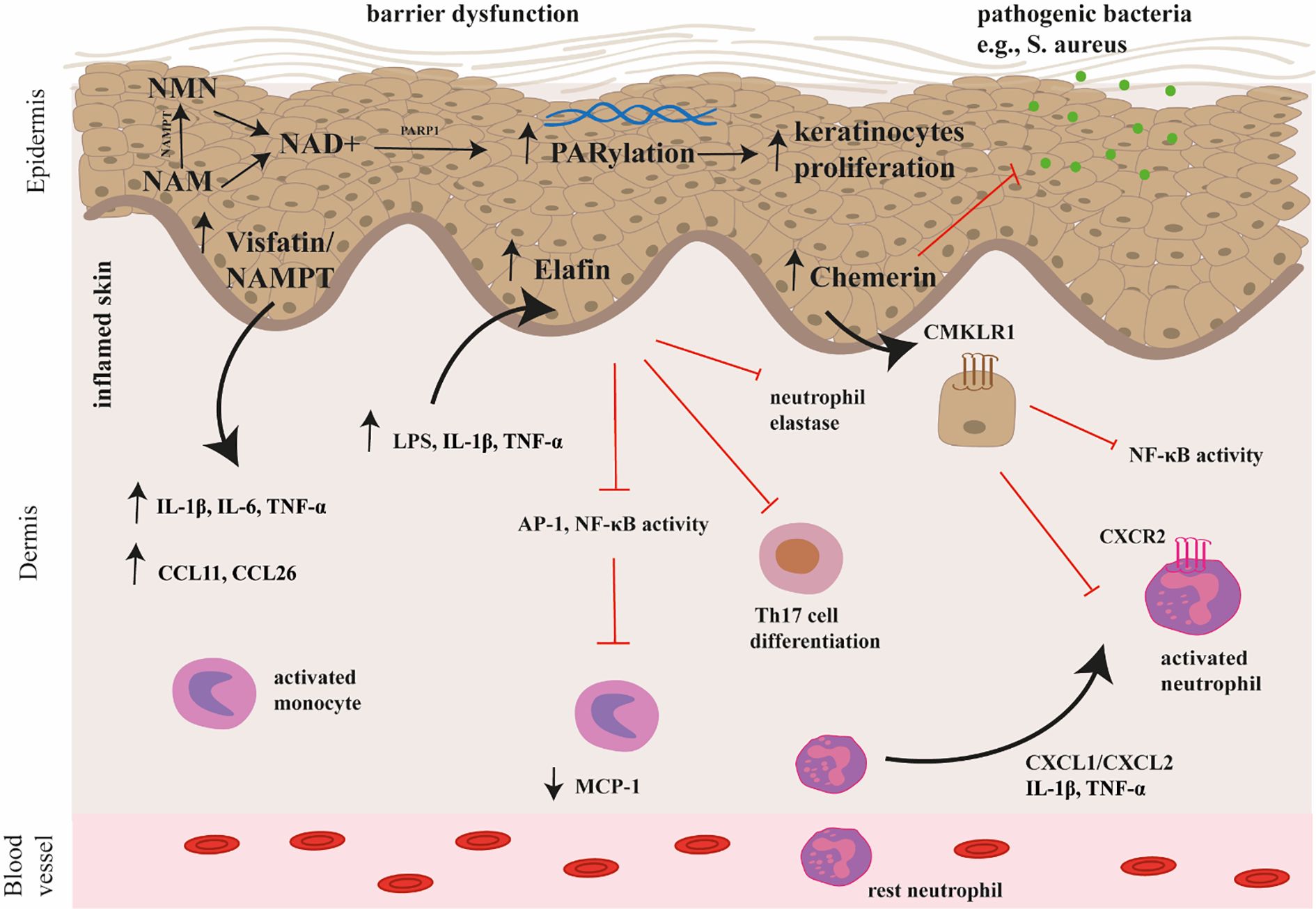
Figure 1. The potential mechanism of chemerin, elafin, and visfatin/NAMPT action in the pathogenesis of AD. Chemerin exerts antimicrobial activity and immunomodulatory effects through interactions with the CMKLR1 receptor. In particular, chemerin attracts neutrophils to leave inflammatory sites by interacting with CCRL2, which may participate in the spread of inflammation. Elafin reduces skin inflammation and tissue damage by inhibiting the activity of transcription factors (AP-1, NF-κB) involved in the production of pro-inflammatory cytokines and by inhibiting the activity of neutrophil elastase. Moreover, it affects the differentiation and function of Th17 cells. Visfatin/NAMPT is the rate limiting enzyme in the NAD+ salvage pathway, converting NAM into NMN, which is then converted to NAD+. Subsequently, poly(ADP-ribose) (PAR) polymerases (PARP) consume NAD+ during the PARylation process. Overactivation of PARP mediates skin inflammation and promotes keratinocyte proliferation. Visfatin/NAMPT increases the secretion of pro-inflammatory cytokines such as IL-1β, IL-6, TNF-α, as well as chemokines (e.g., CCL11, CCL26). AP-1, activator protein 1; CCL11, CC Motif Chemokine Ligand 11; CCL26, CC Motif Chemokine Ligand 26; CCRL2, C-C Chemokine receptor-like 2; CMKLR1, Chemerin Chemokine-Like Receptor 1; IL-1β, interleukin 1β, IL-6, interleukin 6; NAD+, nicotinamide adenine dinucleotide; NAM, nicotinamide; NMN, nicotinamide mononucleotide; PARP, poly(ADP-ribose) (PAR) polymerase; TNF-α, tumor necrosis factor α.
A comprehensive understanding of the role of chemerin, elafin, and visfatin in the pathogenesis of atopic dermatitis (AD) is essential for the development of novel and more effective therapeutic strategies. Although these proteins have been increasingly studied in recent years, their specific functions in AD remain incompletely understood, with some studies reporting conflicting findings. This narrative review aims to synthesize the current evidence regarding their involvement in AD, identify gaps in the existing knowledge, and propose future research directions. By clarifying the roles of these mediators in AD, this review seeks to offer timely and clinically relevant insights that may inform the development of targeted treatments and ultimately improve patient outcomes. While previous reviews have focused on single mediators, our study integrates evidence on three structurally and functionally distinct proteins implicated in AD. A literature search was conducted using PubMed (1990–present), Google Scholar, and Embase, employing appropriate search terms.
2 Discussion
2.1 Elafin – an overview
Elafin and its precursor, trappin-2, are potent inhibitors of human serine proteases, specifically those belonging to the chelonianin family (16). The protein is an epithelial host-defense protein that is absent in normal skin, but is significantly upregulated in keratinocytes in inflamed skin (17). Elafin, by inhibiting excessive protease activity, plays a crucial role in protecting the skin’s epidermal structure and integrity. This is crucial for maintaining the skin’s barrier function and preventing damage. Therefore, elafin may facilitate epidermal repair and regeneration after injury. Furthermore, elafin may influence the migration of polymorphonuclear leukocytes (PMNs), a type of white blood cell involved in inflammation and wound healing. By modulating PMN migration, elafin helps to ensure a controlled and effective inflammatory response (18). Elafin has a positive charge, which is important for its interactions with other molecules and its function. Proinflammatory cytokines like IL-1 and TNF-α stimulate the production of elafin, indicating its role in the inflammatory response (19). In summary, elafin has been shown to possess significant antimicrobial properties against bacteria, viruses, parasites, and fungi (20).
Elafin inhibits lipopolysaccharide (LPS)-induced production of monocyte chemotactic protein 1 (MCP-1) in monocytes by blocking the activation of activatior protein (AP-1) and NF-κB, which are key transcription factors in pro-inflammatory cytokine production, including those involved in Th17 differentiation. The suppression of AP-1 and NF-κB signaling could potentially lead to a downregulation of signals that promote Th17 differentiation and subsequent cytokine production (Figure 1) (21, 22).
2.1.1 Elafin’s role in the pathogenesis of atopic dermatitis
Current studies indicate that elafin could be a pivotal indicator of the AD (Table 1). Brunner et al. (23) found that in children with AD, blood levels of markers associated with Th2 (Chemokine (C-C motif) ligand 13 (CCL13), Chemokine (C-C motif) ligand 22 (CCL22)) and Th17 (elafin) were significantly higher compared to healthy children. The study found only weak correlations between BMI and inflammatory markers (elafin, IL-16, and IL-2RA). This suggests that while obesity contributes to inflammation, its impact on the specific inflammatory pathways involved in pediatric AD may be limited (23) (Table 1).

Table 1. Summary of the studies on elafin’s role in atopic dermatitis.
The study by Esaki et al. (24) found that children with AD exhibited significantly higher levels of Th17-related cytokines and antimicrobial peptides (like elafin) compared to adults with AD. Importantly, similar to the studies mentioned above, this study observed strong correlations between disease activity scores and markers associated with Th2 and Th17 pathways (including elafin). Precisely, AD severity was strongly associated with IL-17 (cytokine produced by Th17)-related elafin marker. This suggests that these immune pathways play a significant role in driving the severity of the disease (24) (Table 1).
Facheris et al. (25) found that individuals with pediatric-onset AD (POAD) persisting into adulthood have higher levels of inflammation and elafin in their affected skin compared to those with adult-onset atopic dermatitis (AOAD). These differences between POAD and AOAD regarding these markers were statistically significant, with a p-value less than 0.05 (25) (Table 1).
In line with previous findings, Brunner et al. (13) investigated the relationship between inflammatory markers and AD severity They found that serum levels of elafin are associated on the extent of AD skin disease. Importantly, these correlations were observed in both affected and unaffected skin of AD patients. On the other side, elafin was also linked to atherosclerosis development alongside other inflammatory markers like E-selectin, Chemokine CC motif ligand 7 (CCL7), IL16. The study also established a strong link between elafin and AD severity, but interestingly, no correlation was found between elafin levels and body mass index (BMI) (13) (Table 1).
Gittler et al. (26) revealed that Th2 and Th22 immune responses are progressively activated as AD transitions from acute to chronic phases. The study found a positive correlation between the SCORAD index and the expression of IL-22 mRNA in acute disease. This suggests that IL-22 may play a significant role in the severity of acute AD. While small increases in Th17-related cytokines (IL-17, IL-23p19, and IL-23p40) were observed, there were significant increases in the expression of genes regulated by elafin, IL-17, such as chemokine ligand 20 (CCL20), and lipocalin 2 (LCN2). This suggests that the Th17 pathway is also involved in acute AD, although to a lesser extent than Th2 and Th22 (26) (Table 1).
On the other hand, Lancto et al. (27) observed significantly lower transcript levels for elafin, secretory leukocyte proteinase inhibitor (SLPI), canine beta-defensin 1 (CBD-1), canine beta-defensin 103 (CBD-103), and canine beta-defensin 122 (CBD-122) in both lesional and non-lesional skin of dogs with AD compared to healthy skin. Significantly higher elafin expression was observed on the inner thigh compared to the scapula, axilla, and forehead. These findings highlighted that a deficiency in antimicrobial peptides, particularly elafin and SLPI, may contribute to the development and progression of atopic dermatitis and other inflammatory skin conditions in dogs. This could potentially lead to increased susceptibility to skin infections and exacerbation of the inflammatory response (27) (Table 1).
In contrast to the above-mentioned studies, Kamsteeg et al. (28) showed that elafin expression in the skin of patients with AD and allergic contact dermatitis (ACD) was variable and less intense compared to those with psoriasis. As expected, no elafin was detected in healthy skin (28) (Table 1).
Guttman-Yassky et al. (29) found that elafin, S100A7, S100A9, and cystatin A mRNAs were highly expressed in psoriasis skin lesions. While these genes were also expressed at lower levels in atopic dermatitis (AD) lesions compared to psoriasis, their expression was still higher in AD lesions than in healthy skin, suggesting a potential role in AD inflammation, albeit less prominent than in psoriasis (29) (Table 1).
Pavel et al. (30) showed that ASN002 represents a potential new treatment option for AD directly targeting the underlying inflammatory mechanisms of the disease. ASN002 is an oral medication that inhibits two key signaling pathways: Janus Kinase (JAK) and Spleen Tyrosine Kinase (SYK). In addition, its ability to modulate multiple signaling pathways also includes TH2, TH22, TH17, and TH1 pathways, all of which play a crucial role in AD (30) (Table 1).
The study by Jensen et al. (31) demonstrated that both pimecrolimus and betamethasone valerate treatments led to a decrease in elafin expression in the skin of patients with atopic dermatitis (AD). This suggests that both medications effectively reduce inflammation in AD (31) (Table 1).
Khattri et al. (32) suggested that ustekinumab, a biological drug, effectively targets and suppresses specific immune pathways involved in Atopic Dermatitis (AD) by inhibiting the activity of Th1 and Th17 immune cells. This inhibition is achieved by blocking the signaling pathways involving IL-12 and IL-23, potentially leading to a reduction in inflammation in conditions where these cells play a significant role (32) (Table 1).
Khattri et al. (33) observed that ciclosporin A treatment suppressed genes associated with Th2, Th22, and some Th17 immune responses. These genes include those involved in producing proteins like elafin, S100 Calcium Binding Protein A7-9 (S100A7-9), CC chemokine ligand 17 (CCL17), and S100As. In addition, this suppression of immune responses was correlated with reduced epidermal hyperplasia (skin thickening) and overall clinical improvement in AD patients. The study also noted that these changes in gene expression occurred in both lesional and non-lesional skin (33) (Table 1).
Czarnowicki et al. (34) described a study investigating the effects of petrolatum on AD skin. Its application significantly increased the expression of various antimicrobial peptides (S100 Calcium Binding Protein A8 (S100A8), S100 Calcium Binding Protein A9 (S100A9), CCL20, elafin, lipocalin 2, human β-defensin 2) and innate immune genes (IL-6, IL-8, and IL-1β) in the skin of patients with AD. This suggests that petrolatum may stimulate the skin’s natural defenses against infection. Moreover, petrolatum also stimulated the expression of key barrier differentiation markers (filaggrin and loricrin), which are essential for maintaining a healthy skin barrier. Additionally, it increased the thickness of the stratum corneum (the outermost layer of the skin), further enhancing the skin’s barrier function (34) (Table 1).
2.2 Chemerin – an overview
In 1997, chemerin was identified as a tazarotene-induced gene-2 (TIG-2) in psoriatic skin (35), Chemerin is initially secreted in an inactive precursor form known as prochemerin (36). Moreover, chemerin and its receptors (chemokine-like receptor 1 (CMKLR1) (36), G protein-coupled receptor 1 (GPR1) (37), C-C chemokine receptor-like 2 (CCRL2) (38) significantly contribute to various biological processes, like adipogenesis (39), osteoclastogenesis (40), angiogenesis (41), and skin inflammation (42). Regarding the role of chemerin in the skin, human keratinocytes cultured in the laboratory respond to specific microbial signals by altering the expression of chemerin and its receptors. Furthermore, chemerin has been found to be essential for the efficient clearance of bacteria in a skin infection model (43).
Chemerin has dual, or “chimeric,” effects on inflammation: pro-inflammatory via activation of the ChemR23 receptor or anti-inflammatory via CMKLR1. For example, activation of CMKLR1 has been linked to suppression of the transcription factor NF-κB and a shift of the macrophage phenotype toward a less inflammatory state. Chemerin has a chemotactic effect on immune cells, promoting cell migration in inflammatory conditions; on the other hand, it inhibits the action of pro-inflammatory cytokines such as interleukin 6 (IL-6) and tumor necrosis factor α (TNF-α), exerting anti-inflammatory effects. However, the precise balance of these effects in AD is still under investigation (44).
2.2.1 Chemerin’s role in the pathogenesis of atopic dermatitis
Literature data indicated that chemerin is engaged in pathological mechanisms underlying some skin diseases, including AD. Xiao et al. (14) discussed the role of IL-13 and toll-like receptor 2 (TLR2) in neurogenic inflammation, particularly in AD and itch. Pam3CSK4 (a TLR2 agonist) enhances the effects of IL-13 in cultured sensory neurons. This includes increased calcium signaling, a higher number of responding neurons, and the release of pro-inflammatory cytokines. In sensory neurons (mDRGs), pre-treatment with Pam3CSK4 significantly increased the secretion of cytokines (chemerin, TNFα, CCL17, and CCL20) in response to IL-13. This enhancement was significantly reduced in neurons with reduced IL-13Rα2 expression (achieved through IL-13Rα2 knockdown). Furthermore, these findings suggest that the interplay between IL-13 and TLR2 signaling pathways is crucial in driving neurogenic inflammation in AD. Summing up, targeting both IL-13-IL-13Rα2 and TLR2-IL-13Rα2 pathways could be a promising therapeutic strategy for treating AD and associated itch (14) (Table 2).

Table 2. Summary of the studies on chemerin’s role in atopic dermatitis.
Albanesi et al. (45) presented data on the distribution of chemerin and its potential role in AD. Very few chemerin-expressing cells were found in the dermis of AD skin lesions. This contrasts with findings in psoriasis, where chemerin plays a role in early lesion development. AD lesions showed minimal infiltration of plasmacytoid dendritic cells (pDCs). Importantly, there was no association between the presence of chemerin-expressing cells and pDCs or neutrophils in AD skin. Taken together these results suggest that the chemerin pathway may not play a significant role in the pathogenesis of AD, unlike in the pathogenesis of psoriasis (45) (Table 2).
2.3 Visfatin/NAMPT – an overview
Visfatin/NAMPT is a protein also known as pre-B-cell colony-enhancing factor (PBEF) and nicotinamide phosphoribosyltransferase (NAMPT), It is primarily secreted by visceral adipose tissue (46). Visfatin/NAMPT has a dual role, acting as both an intracellular enzyme and an extracellular protein. Inside cells, it acts as an enzyme that converts nicotinamide (NAM) into nicotinamide mononucleotide (NMN). However, in both intracellular and extracellular contexts, the produced NMN is then converted to nicotinamide adenine dinucleotide (NAD) by the enzyme nicotinamide/nicotinic acid mononucleotide adenyltransferase (NMNAT). NAD is a vital coenzyme involved in various cellular processes, including energy production, cellular signaling, and DNA repair (47). Visfatin/NAMPT enhances the production of pro-inflammatory cytokines, specifically IL-6 and IL-1β, in both human monocytes (48) and endothelial cells (49), supporting the classification of visfatin/NAMPT as an inflammatory adipokine.
2.3.1 Visfatin/NAMPT role in the pathogenesis of atopic dermatitis
Arroyo et al. (15) found that visfatin/NAMPT, an enzyme crucial for NAD+ production, was significantly elevated in the spinous layer of the skin in AD patients. Some basal keratinocytes and dermal cells also showed increased visfatin/NAMPT levels. Moreover, patients with higher levels of visfatin/NAMPT and poly(ADP-ribose) (PAR) also exhibited increased levels of proliferating cell nuclear antigen (PCNA), a marker of cell proliferation. This suggests a link between visfatin/NAMPT activity and increased cell growth in AD skin (15) (Table 3).

Table 3. Summary of the studies on visfatin/NAMPT role in atopic dermatitis.
Suga et al. (50) showed that serum visfatin/NAMPT levels were significantly higher in patients with AD compared to healthy individuals. In AD patients, visfatin/NAMPT levels correlated with eosinophil counts, moreover, it was related to itch severity (measured by visual analogue scale (VAS)) and levels of certain inflammatory molecules (C-C motif chemokine 11 (CCL11) and C-C motif chemokine 26 (CCL26)). In addition, visfatin/NAMPT levels increased with AD severity (mild, moderate, severe). An adult-onset AD also showed significantly higher visfatin/NAMPT levels compared to classical childhood-onset AD. In conclusion, an increased visfatin/NAMPT level could be a biomarker for disease severity in AD (50) (Table 3).
In contrast to the abovementioned studies, Machura et al. (51) observed that serum visfatin/NAMPT level was significantly lower in children with AD compared to healthy controls. The ratio of visfatin/NAMPT levels to BMI was also lower in children with AD. However, visfatin/NAMPT levels were similar between normal-weight and obese children with AD. A strong positive correlation was found between visfatin/NAMPT concentration and triglyceride levels in children dealing with AD. The study suggests that visfatin/NAMPT levels could potentially be used to distinguish children with AD from healthy children (51) (Table 3).
2.4 Therapeutic potential
2.4.1 Elafin
Elafin, a serine protease inhibitor, holds clinical value in several areas due to its anti-inflammatory and anti-proteolytic properties. It is being investigated as a potential biomarker and therapeutic target in various inflammatory and neoplastic diseases, including inflammatory bowel disease (IBD), acute graft-versus-host disease (aGVHD), and cancers. Phase II trials are underway to investigate the therapeutic effects of elafin on post-operative inflammation and morbidity after major surgeries, such as esophagectomy (for esophageal cancer) (52), coronary artery bypass surgery (53) and pulmonary arterial hypertension (54).
2.4.2 Chemerin
Chemerin has emerged as a potential biomarker with clinical value in various diseases. It plays a role in inflammation, metabolism, and immune responses, and its levels in the body can be associated with conditions like obesity, cardiovascular disease, and certain cancers. Preclinical research in mice has explored modulating the chemerin/CMKLR1 axis, using chemerin-derived peptides or CMKLR1 inhibitors, for potential treatments for neuropathic pain and allergic airway inflammation. However, there is a lack of clinical trial data in humans (55).
2.4.3 Visfatin/NAMPT
Visfati/NAMPT is an adipokine with potential therapeutic applications, particularly in metabolic disorders and inflammatory conditions. It can influence various cellular processes, including glucose metabolism, inflammation, and angiogenesis. More than 100 patients were treated with CHS-828 (later known as GMX1778), a potent and specific NAD(+) inhibitor (NAMPT), in a phase I clinical trial (56). The drug was administered orally. The most common adverse events were thrombocytopenia and gastrointestinal (GI) effects such as diarrhea, vomiting, and esophagitis (57, 58). While visfatin holds promise as a therapeutic target, further research is needed to fully understand its complex mechanisms of action and to address potential toxicity concerns. Clinical trials are necessary to validate the safety and efficacy of visfatin-targeted therapies.
2.5 Conclusions
In summary, the available literature data indicate that elafin, chemerin, and visfatin/NAMPT play an important role in epidermal barrier function and inflammation in AD. Their potential stems from dual pro- and anti-inflammatory properties, suggesting a complex role in regulating the inflammatory balance in this dermatosis. For example, it was shown that chemerin restricts the growth of a variety of skin- associated bacteria but on the other hand promotes immune cells migration to the site of inflammation. Therefore, the usage of substances that can specifically regulate the activity of the studied proteins may offer a potential therapeutic approach for the treatment of AD. Available preclinical and early clinical studies have shown their potential safety and tolerability. However, long-term safety and efficacy need to be established in a larger patient population and reliable methods need to be developed to measure levels of these proteins for diagnostic purposes. If successful, these proteins could offer new therapeutic options for atopic dermatitis, potentially complementing or even surpassing existing treatment options.
2.6 Limitations
Although knowledge about chemerin, elafin, and visfatin/NAMPT in AD is growing, the inherent complexity and heterogeneity of the disease, combined with inconsistencies in study methodology, pose serious obstacles to drawing clear, comparable, and reproducible conclusions from scientific studies. There is a lack of uniform research methods and standards. Many studies differ significantly in terms of experimental design, such as in vitro experiments, animal models, or small-scale clinical trials, making it difficult to directly compare and analyze the results of different studies. Therefore, larger, longitudinal studies are needed in the future to verify these results and better assess the role of these proteins in the development and progression of AD.
Author contributions
MM: Investigation, Methodology, Project administration, Software, Visualization, Writing – original draft, Writing – review & editing. AK-P: Conceptualization, Formal Analysis, Resources, Supervision, Validation, Visualization, Writing – review & editing. HM: Conceptualization, Formal Analysis, Funding acquisition, Resources, Supervision, Visualization, Writing – review & editing. AM: Validation, Writing – review & editing. AC: Validation, Writing – review & editing. BM: Supervision, Writing – review & editing. IF: Supervision, Validation, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
CCL7, chemokine C-C motif ligand 7; BMI, body mass index; POAD, pediatric-onset AD; AOAD, adult-onset atopic dermatitis; JAK, Janus Kinase Inhibitor; SYK, Spleen Tyrosine Kinase; CCL13, chemokine C-C motif ligand 13; CCL22, Chemokine C-C motif ligand 22; CCL20, chemokine ligand 20; LCN2, lipocalin 2; S100A7-9, S100 Calcium Binding Protein A7-9; CCL17, CC chemokine ligand 17; S100A8, S100 Calcium Binding Protein A8; S100A9, S100 Calcium Binding Protein A9; SLPI, secretory leukocyte proteinase inhibitor; CBD-1, canine beta-defensin 1; CBD-103, canine beta-defensin 103; CBD-122, canine beta-defensin 122; ACD, allergic contact dermatitis; N1 – control group; N2 – study group; TLR2, toll-like receptor 2; TNFα, tumor necrosis factor alpha; pDCs, plasmacytoid dendritic cells; PAR, poly(ADP-ribose); PCNA, proliferating cell nuclear antigen; VAS, visual analogue scale; CCL11, C-C motif chemokine 11; CCL26, C-C motif chemokine 26.
References
1. Peng W and Novak N. Pathogenesis of atopic dermatitis. Clin Exp Allergy. (2015) 45:566–74. doi: 10.1111/cea.12495
2. Nutten S. Atopic dermatitis: global epidemiology and risk factors. Ann Nutr Metab. (2015) 66 Suppl 1:8–16. doi: 10.1159/000370220
3. Jin L, Ge J, Cheng Y, Deng D, and Wan P. Worldwide prevalence of atopic dermatitis in children between 2000 and 2021: a systematic analysis. Ann Allergy Asthma Immunol. (2024). doi: 10.1016/j.anai.2024.12.005
4. Maspero J, De Paula Motta Rubini N, Zhang J, Sanclemente G, Amador JR, El Sayed MH, et al. Epidemiology of adult patients with atopic dermatitis in AWARE 1: A second international survey. World Allergy Organ J. (2023) 16:100724. doi: 10.1016/j.waojou.2022.100724
5. Thyssen JP, Halling AS, Schmid-Grendelmeier P, Guttman-Yassky E, and Silverberg JI. Comorbidities of atopic dermatitis-what does the evidence say? J Allergy Clin Immunol. (2023) 151:1155–62. doi: 10.1016/j.jaci.2022.12.002
6. Saeki H, Ohya Y, Arakawa H, Ichiyama S, Katsunuma T, Katoh N, et al. English version of clinical practice guidelines for the management of atopic dermatitis 2024. J Dermatol. (2024). doi: 10.1111/1346-8138.17544
7. Eichenfield LF, Tom WL, Chamlin SL, Feldman SR, Hanifin JM, Simpson EL, et al. Guidelines of care for the management of atopic dermatitis: section 1. Diagnosis and assessment of atopic dermatitis. J Am Acad Dermatol. (2014) 70:338–51. doi: 10.1016/j.jaad.2013.10.010
8. Darsow U, Pfab F, Valet M, Huss-Marp J, Behrendt H, Ring J, et al. Pruritus and atopic dermatitis. Clin Rev Allergy Immunol. (2011) 41:237–44. doi: 10.1007/s12016-010-8230-2
9. Malik K, Heitmiller KD, and Czarnowicki T. An update on the pathophysiology of atopic dermatitis. Dermatol Clin. (2017) 35:317–26. doi: 10.1016/j.det.2017.02.006
10. Nakahara T, Kido-Nakahara M, Tsuji G, and Furue M. Basics and recent advances in the pathophysiology of atopic dermatitis. J Dermatol. (2021) 48:130–9. doi: 10.1111/1346-8138.15664
11. Palmer CN, Irvine AD, Terron-Kwiatkowski A, Zhao Y, Liao H, Lee SP, et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat Genet. (2006) 38:441–6. doi: 10.1038/ng1767
12. Trzeciak M, Sakowicz-Burkiewicz M, Wesserling M, Dobaczewska D, Gleń J, Nowicki R, et al. Expression of cornified envelope proteins in skin and its relationship with atopic dermatitis phenotype. Acta Derm Venereol. (2017) 97:36–41. doi: 10.2340/00015555-2482
13. Brunner PM, Suárez-Fariñas M, He H, Malik K, Wen HC, Gonzalez J, et al. The atopic dermatitis blood signature is characterized by increases in inflammatory and cardiovascular risk proteins. Sci Rep. (2017) 7:8707. doi: 10.1038/s41598-017-09207-z
14. Xiao S, Lu Z, Steinhoff M, Li Y, Buhl T, Fischer M, et al. Innate immune regulates cutaneous sensory IL-13 receptor alpha 2 to promote atopic dermatitis. Brain Behav Immun. (2021) 98:28–39. doi: 10.1016/j.bbi.2021.08.211
15. Arroyo AB, Bernal-Carrión M, Cantón-Sandoval J, Cabas I, Corbalán-Vélez R, Martínez-Menchón T, et al. NAMPT and PARylation are involved in the pathogenesis of atopic dermatitis. Int J Mol Sci. (2023) 24:7992. doi: 10.3390/ijms24097992
16. Moreau T, Baranger K, Dadé S, Dallet-Choisy S, Guyot N, and Zani ML. Multifaceted roles of human elafin and secretory leukocyte proteinase inhibitor (SLPI), two serine protease inhibitors of the chelonianin family. Biochimie. (2008) 90:284–95. doi: 10.1016/j.biochi.2007.09.007
17. Doss RW, Mohammed EHA, Mohamed EH, and Abdellatif MEA. Elafin level in lichen planus. Arch Dermatol Res. (2024) 316:547. doi: 10.1007/s00403-024-03304-6
18. van Bergen BH, Andriessen MP, Spruijt KI, van de Kerkhof PC, and Schalkwijk J. Expression of SKALP/elafin during wound healing in human skin. Arch Dermatol Res. (1996) 288:458–62. doi: 10.1007/BF02505235
19. Schalkwijk J, Chang A, Janssen P, De Jongh GJ, and Mier PD. Skin-derived antileucoproteases (SKALPs): characterization of two new elastase inhibitors from psoriatic epidermis. Br J Dermatol. (1990) 122:631–41. doi: 10.1111/j.1365-2133.1990.tb07285.x
20. Simpson AJ, Maxwell AI, Govan JR, Haslett C, and Sallenave JM. Elafin (elastase-specific inhibitor) has anti-microbial activity against gram-positive and gram-negative respiratory pathogens. FEBS Lett. (1999) 452:309–13. doi: 10.1016/s0014-5793(99)00670-5
21. Guyot N, Butler MW, McNally P, Weldon S, Greene CM, Levine RL, et al. Elafin, an elastase-specific inhibitor, is cleaved by its cognate enzyme neutrophil elastase in sputum from individuals with cystic fibrosis. J Biol Chem. (2008) 283:32377–85. doi: 10.1074/jbc.M803707200
22. Chen L, Deng H, Cui H, Fang J, Zuo Z, Deng J, et al. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget. (2017) 9:7204–18. doi: 10.18632/oncotarget.23208
23. Brunner PM, He H, Pavel AB, Czarnowicki T, Lefferdink R, Erickson T, et al. The blood proteomic signature of early-onset pediatric atopic dermatitis shows systemic inflammation and is distinct from adult long-standing disease. J Am Acad Dermatol. (2019) 81:510–9. doi: 10.1016/j.jaad.2019.04.036
24. Esaki H, Brunner PM, Renert-Yuval Y, Czarnowicki T, Huynh T, Tran G, et al. Early-onset pediatric atopic dermatitis is TH2 but also TH17 polarized in skin. J Allergy Clin Immunol. (2016) 138:1639–51. doi: 10.1016/j.jaci.2016.07.013
25. Facheris P, Da Rosa JC, Pagan AD, Angelov M, Del Duca E, Rabinowitz G, et al. Age of onset defines two distinct profiles of atopic dermatitis in adults. Allergy. (2023) 78:2202–14. doi: 10.1111/all.15741
26. Gittler JK, Shemer A, Suárez-Fariñas M, Fuentes-Duculan J, Gulewicz KJ, Wang CQ, et al. Progressive activation of T(H)2/T(H)22 cytokines and selective epidermal proteins characterizes acute and chronic atopic dermatitis. J Allergy Clin Immunol. (2012) 130:1344–54. doi: 10.1016/j.jaci.2012.07.012
27. Lancto CA, Torres SM, Hendrickson JA, Martins KV, and Rutherford MS. Altered expression of antimicrobial peptide genes in the skin of dogs with atopic dermatitis and other inflammatory skin conditions. Vet Dermatol. (2013) 24:414–21. doi: 10.1111/vde.12034
28. Kamsteeg M, Jansen PA, van Vlijmen-Willems IM, van Erp PE, Rodijk-Olthuis D, van der Valk PG, et al. Molecular diagnostics of psoriasis, atopic dermatitis, allergic contact dermatitis and irritant contact dermatitis. Br J Dermatol. (2010) 162:568–78. doi: 10.1111/j.1365-2133.2009.09547.x
29. Guttman-Yassky E, Lowes MA, Fuentes-Duculan J, Zaba LC, Cardinale I, Nograles KE, et al. Low expression of the IL-23/Th17 pathway in atopic dermatitis compared to psoriasis. J Immunol. (2008) 181:7420–7. doi: 10.4049/jimmunol.181.10.7420
30. Pavel AB, Song T, Kim HJ, Del Duca E, Krueger JG, Dubin C, et al. Oral Janus kinase/SYK inhibition (ASN002) suppresses inflammation and improves epidermal barrier markers in patients with atopic dermatitis. J Allergy Clin Immunol. (2019) 144:1011–24. doi: 10.1016/j.jaci.2019.07.013
31. Jensen JM, Scherer A, Wanke C, Bräutigam M, Bongiovanni S, Letzkus M, et al. Gene expression is differently affected by pimecrolimus and betamethasone in lesional skin of atopic dermatitis. Allergy. (2012) 67:413–23. doi: 10.1111/j.1398-9995.2011.02747.x
32. Khattri S, Brunner PM, Garcet S, Finney R, Cohen SR, Oliva M, et al. Efficacy and safety of ustekinumab treatment in adults with moderate-to-severe atopic dermatitis. Exp Dermatol. (2017) 26:28–35. doi: 10.1111/exd.13112
33. Khattri S, Shemer A, Rozenblit M, Dhingra N, Czarnowicki T, Finney R, et al. Cyclosporine in patients with atopic dermatitis modulates activated inflammatory pathways and reverses epidermal pathology. J Allergy Clin Immunol. (2014) 133:1626–34. doi: 10.1016/j.jaci.2014.03.003
34. Czarnowicki T, Malajian D, Khattri S, Correa da Rosa J, Dutt R, Finney R, et al. Petrolatum: Barrier repair and antimicrobial responses underlying this “inert” moisturizer. J Allergy Clin Immunol. (2016) 137:1091–102. doi: 10.1016/j.jaci.2015.08.013
35. Nagpal S, Patel S, Jacobe H, DiSepio D, Ghosn C, Malhotra M, et al. Tazarotene-induced gene 2 (TIG2), a novel retinoid-responsive gene in skin. J Invest Dermatol. (1997) 109:91–5. doi: 10.1111/1523-1747.ep12276660
36. Wittamer V, Franssen JD, Vulcano M, Mirjolet JF, Le Poul E, Migeotte I, et al. Specific recruitment of antigen-presenting cells by chemerin, a novel processed ligand from human inflammatory fluids. J Exp Med. (2003) 198:977–85. doi: 10.1084/jem.20030382
37. Barnea G, Strapps W, Herrada G, Berman Y, Ong J, Kloss B, et al. The genetic design of signaling cascades to record receptor activation. Proc Natl Acad Sci U S A. (2008) 105:64–9. doi: 10.1073/pnas.0710487105
38. Zabel BA, Nakae S, Zúñiga L, Kim JY, Ohyama T, Alt C, et al. Mast cell-expressed orphan receptor CCRL2 binds chemerin and is required for optimal induction of IgE-mediated passive cutaneous anaphylaxis. J Exp Med. (2008) 205:2207–20. doi: 10.1084/jem.20080300
39. Goralski KB, McCarthy TC, Hanniman EA, Zabel BA, Butcher EC, Parlee SD, et al. Chemerin, a novel adipokine that regulates adipogenesis and adipocyte metabolism. J Biol Chem. (2007) 282:28175–88. doi: 10.1074/jbc.M700793200
40. Muruganandan S, Roman AA, and Sinal CJ. Role of chemerin/CMKLR1 signaling in adipogenesis and osteoblastogenesis of bone marrow stem cells. J Bone Miner Res. (2010) 25:222–34. doi: 10.1359/jbmr.091106
41. Kaur J, Adya R, Tan BK, Chen J, and Randeva HS. Identification of chemerin receptor (ChemR23) in human endothelial cells: chemerin-induced endothelial angiogenesis. Biochem Biophys Res Commun. (2010) 391:1762–8. doi: 10.1016/j.bbrc.2009.12.150
42. Schultz S, Saalbach A, Heiker JT, Meier R, Zellmann T, Simon JC, et al. Proteolytic activation of prochemerin by kallikrein 7 breaks an ionic linkage and results in C-terminal rearrangement. Biochem J. (2013) 452:271–80. doi: 10.1042/BJ20121880
43. Banas M, Zegar A, Kwitniewski M, Zabieglo K, Marczynska J, Kapinska-Mrowiecka M, et al. The expression and regulation of chemerin in the epidermis. PloS One. (2015) 10:e0117830. doi: 10.1371/journal.pone.0117830
44. Zhang X, Weiß T, Cheng MH, Chen S, Ambrosius CK, Czerniak AS, et al. Structural basis of CMKLR1 signaling induced by chemerin9. bioRxiv. (2023) 10:2023. doi: 10.1101/2023.06.09.544295
45. Albanesi C, Scarponi C, Pallotta S, Daniele R, Bosisio D, Madonna S, et al. Chemerin expression marks early psoriatic skin lesions and correlates with plasmacytoid dendritic cell recruitment. J Exp Med. (2009) 206:249–58. doi: 10.1084/jem.20080129
46. Ismail SA and Mohamed SA. Serum levels of visfatin and omentin-1 in patients with psoriasis and their relation to disease severity. Br J Dermatol. (2012) 167:436–9. doi: 10.1111/j.1365-2133.2012.10980.x
47. Revollo JR, Körner A, Mills KF, Satoh A, Wang T, Garten A, et al. Nampt/PBEF/Visfatin regulates insulin secretion in beta cells as a systemic NAD biosynthetic enzyme. Cell Metab. (2007) 6:363–75. doi: 10.1016/j.cmet.2007.09.003
48. Moschen AR, Kaser A, Enrich B, Mosheimer B, Theurl M, Niederegger H, et al. Visfatin, an adipocytokine with proinflammatory and immunomodulating properties. J Immunol. (2007) 178:1748–58. doi: 10.4049/jimmunol.178.3.1748
49. Kim JY, Bae YH, Bae MK, Kim SR, Park HJ, Wee HJ, et al. Visfatin through STAT3 activation enhances IL-6 expression that promotes endothelial angiogenesis. Biochim Biophys Acta. (2009) 1793:1759–67. doi: 10.1016/j.bbamcr.2009.09.006
50. Suga H, Sugaya M, Miyagaki T, Kawaguchi M, Morimura S, Kai H, et al. Serum visfatin levels in patients with atopic dermatitis and cutaneous T-cell lymphoma. Eur J Dermatol. (2013) 23:629–35. doi: 10.1684/ejd.2013.2107
51. Machura E, Szczepanska M, Ziora K, Ziora D, Swietochowska E, Barc-Czarnecka M, et al. Evaluation of adipokines: apelin, visfatin, and resistin in children with atopic dermatitis. Mediators Inflamm. (2013) 2013:760691. doi: 10.1155/2013/760691
52. Shaw L and Wiedow O. Therapeutic potential of human elafin. Biochem Soc Trans. (2011) 39:1450–4. doi: 10.1042/BST0391450
53. Alam SR, Lewis SC, Zamvar V, Pessotto R, Dweck MR, Krishan A, et al. Perioperative elafin for ischaemia-reperfusion injury during coronary artery bypass graft surgery: a randomised-controlled trial. Heart. (2015) 101:1639–45. doi: 10.1136/heartjnl-2015-307745
55. Doyle JR, Krishnaji ST, Zhu G, Xu ZZ, Heller D, Ji RR, et al. Development of a membrane-anchored chemerin receptor agonist as a novel modulator of allergic airway inflammation and neuropathic pain. J Biol Chem. (2014) 289:13385–96. doi: 10.1074/jbc.M113.522680
56. Watson M, Roulston A, Bélec L, Billot X, Marcellus R, Bédard D, et al. The small molecule GMX1778 is a potent inhibitor of NAD+ biosynthesis: strategy for enhanced therapy in nicotinic acid phosphoribosyltransferase 1-deficient tumors. Mol Cell Biol. (2009) 29:5872–88. doi: 10.1128/MCB.00112-09
57. Hovstadius P, Larsson R, Jonsson E, Skov T, Kissmeyer AM, Krasilnikoff K, et al. A Phase I study of CHS 828 in patients with solid tumor Malignancy. Clin Cancer Res. (2002) 8:2843–50.
Keywords: atopic dermatitis, chemerin, elafin, visfatin, NAMPT
Citation: Matwiejuk M, Kulczyńska-Przybik A, Myśliwiec H, Mikłosz A, Chabowski A, Mroczko B and Flisiak I (2025) The role of chemerin, elafin, and visfatin in the pathogenesis of atopic dermatitis. Front. Immunol. 16:1628163. doi: 10.3389/fimmu.2025.1628163
Received: 13 May 2025; Accepted: 21 July 2025;
Published: 03 September 2025.
Edited by:
Manoj Kumar Tembhre, All India Institute of Medical Sciences, IndiaReviewed by:
Xianjun Xiao, Chengdu University of Traditional Chinese Medicine, ChinaXinling Bi, Changhai Hospital, China
Copyright © 2025 Matwiejuk, Kulczyńska-Przybik, Myśliwiec, Mikłosz, Chabowski, Mroczko and Flisiak. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mateusz Matwiejuk, bWF0ZXVzei5tYXR3aWVqdWtAdW1iLmVkdS5wbA==