 Laura E. Martínez-Gómez1
Laura E. Martínez-Gómez1 Carla I. Oropeza-Vélez2Maylin Almonte-Becerril3
Carla I. Oropeza-Vélez2Maylin Almonte-Becerril3 Leslie Chavez-Galan4
Leslie Chavez-Galan4 Carlos Martinez-Armenta1
Carlos Martinez-Armenta1 Rosa P. Vidal-Vázquez5
Rosa P. Vidal-Vázquez5 Juan P. Ramírez-Hinojosa5
Juan P. Ramírez-Hinojosa5 Paola Vázquez-Cárdenas5
Paola Vázquez-Cárdenas5 Diana Gómez-Martín6
Diana Gómez-Martín6 Gilberto Vargas-Alarcón7
Gilberto Vargas-Alarcón7 José M. Rodríguez-Pérez7
José M. Rodríguez-Pérez7 Lucero A. Ramón-Luing4
Lucero A. Ramón-Luing4 Julio Flores-Gonzalez4José G. Carrasco2
Julio Flores-Gonzalez4José G. Carrasco2 Ivette Cruz-Bautista8
Ivette Cruz-Bautista8 Mónica M. Mata-Miranda9
Mónica M. Mata-Miranda9 Gustavo J. Vázquez-Zapién9
Gustavo J. Vázquez-Zapién9 Adriana Martínez-Cuazitl9Nancy M. Parra-Torres3
Adriana Martínez-Cuazitl9Nancy M. Parra-Torres3 Felipe de J. Martínez-Ruiz10Dulce M. Zayago-Angeles10Ma. Luisa Ordoñez-Sánchez11Yayoi Segura-Kato11
Felipe de J. Martínez-Ruiz10Dulce M. Zayago-Angeles10Ma. Luisa Ordoñez-Sánchez11Yayoi Segura-Kato11 Carlos Suarez-Ahedo1Jessel Olea-Torres1
Carlos Suarez-Ahedo1Jessel Olea-Torres1 Brígida Herrera-López1
Brígida Herrera-López1 Carlos Pineda1
Carlos Pineda1 Gabriela A. Martínez-Nava1*
Gabriela A. Martínez-Nava1* Alberto G. López-Reyes1*
Alberto G. López-Reyes1*- 1Laboratorio de Gerociencias, Dirección General, Departamento de Reconstrucción Articular, Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra, Secretaría de Salud, Mexico City, Mexico
- 2Servicio de Medicina Interna de Hospital Petróleos Mexicanos PEMEX, Mexico City, Mexico
- 3Dirección Ejecutiva de Investigación y Estudios Avanzados (DEIEA), Universidad de la Salud (UNISA), Mexico City, Mexico
- 4Laboratory of Integrative Immunology, Instituto Nacional de Enfermedades Respiratorias Ismael Cosio Villegas, Mexico City, Mexico
- 5Centro de Innovación Médica Aplicada, Hospital General Dr. Manuel Gea González, Mexico City, Mexico
- 6Department of Immunology and Rheumatology, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Secretaría de Salud, Mexico City, Mexico
- 7Departamento de Biología Molecular y Endocrinología, Instituto Nacional de Cardiología Ignacio Chávez, Mexico City, Mexico
- 8Departamento de Endocrinología y Metabolismo, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Mexico, City, Mexico
- 9Laboratorio de Biología Celular y Tisular, Laboratorio de Embriología, Escuela Médico Militar, Universidad del Ejército y Fuerza Aérea, Mexico, City, Mexico
- 10Nuevo Hospital General Delegación Regional Sur de la Ciudad de México ISSSTE, Mexico City, Mexico
- 11Unidad de Biología Molecular y Medicina Genómica, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, Mexico City, Mexico
Background: Cytokines and chemokines are essential for establishing an appropriate immune response to severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2). Variations in the genes encoding cytokines and chemokines strongly influence the immune response to pathogenic challenges and disease outcomes. This study was conducted to investigate the associations between polymorphisms in the TNF-α, IL-6, IL-8, IL-10, and CCL5 genes and COVID-19 severity.
Methodology: We performed a cross-sectional study with a total of 627 unvaccinated COVID-19 patients were classified according to WHO disease severity. We evaluated the levels of IFN-α, IFN-γ, TNF-α, IL-1Ra, IL-2, IL-6, IL-7, IL-10, CCL2, CCL3, CXCL8, CXCL10 and GCSF in the serum and compared them among COVID-19 severity groups by Kruskal-Wallis test and stratified by polymorphism alleles. A logistic regression was performed to determine the association of the polymorphism and COVID-19 severity.
Results: This study revealed a significant increase in IL-2, IL-6 and CCL-2 levels in the deceased group. However, the IL-10 levels were higher in the moderate group than in the mild group. Logistic regression analysis revealed that five polymorphisms were associated with a higher risk of severe COVID-19: the TNF-α (rs1800610) A allele (OR=1.50; 95% CI: 1.01–2.24); the IL-6 (rs1800796) C allele (OR=1.64; 95% CI: 1.05–2.57); the IL-10 (rs1800871) T allele (OR=1.94; 95% CI: 1.24–3.04) and (rs1800872) A allele (OR=1.87; 95% CI: 1.21–2.89); and the CCL5 (rs3817656) G allele (OR= 1.64; 95% CI: 1.02–2.65).
Conclusion: Patients infected with SARS-CoV-2 who have the TNFα gene variant (rs1800629) are protected from developing COVID-19 moderate and severe outcomes, as well as from presenting low concentrations of some pro-inflammatory cytokines and chemokines. However, carriers of the IL-10 (rs1800872, rs1800871) and CCL-5 (rs2107538) gene variants were associated with patients who died from COVID-19. Of these, only the minor allele of CCL-5 was primarily associated with increased chemokines levels, as well as with some cytokines considered hallmarks of the cytokine storm.
1 Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is the etiologic agent of coronavirus disease 2019 (COVID-19). This disease has a broad spectrum of clinical presentations, ranging from asymptomatic to severe outcomes such as acute respiratory distress syndrome (ARDS). Severe outcomes are often marked by hyper inflammation and elevated levels of proinflammatory cytokines such as IFN-α, IFN-γ and, IL-6, as well as chemokines like monocyte chemotactic protein-3 (MCP-1/CCL-2) (1–3).
In addition, different risk factors involved in outcome severity have also been described, including older age, male sex, obesity, and cardiovascular disease, among others. However, these risk factors do not explain all severe cases; genetic host variation might influence the clinical outcomes of patients with COVID-19 (4). Genetic variations may affect innate and adaptive immune responses (4, 5).
Polymorphisms in genes encoding pro-inflammatory cytokines may play a role in the pathogenesis of severe COVID-19 and could even influence their production. Single nucleotide polymorphisms (SNPs) in the TNF-α, IL-6, IL-8, IL-10, CCL5 and CXCL6 genes have been found to be associated with COVID-19 severity, suggesting that SNPs could help explain COVID-19 outcomes (2, 6–12).
In this context, the rs1800610 polymorphism in the TNF-α gene has been associated with several chronic diseases, including cancer, and the development of chronic obstructive pulmonary disease (COPD), as well as other infectious disease like hepatitis B virus (HBV) infection (13–15).
Recent studies have also highlighted strong associations between SNPs of IL6 (rs2069840) and IL10 (rs1800872) genes and the severity of COVID-19. Notably, the CC genotype of IL-10 appears to play a protective role against severe disease progression, although these SNPs may also indicate increased susceptibility to inflammatory conditions in general (2, 16).
A study reported by Alvarez et al., identified an association between SNP rs2107538 of CCL5 and bronchiolitis caused by respiratory syncytial virus (RSV), including the increased need for mechanical ventilation in affected patients (17). This SNPs has also been linked to human immunodeficiency virus (HIV) infection (18). Furthermore, a meta-analysis by Areeshi et al., demonstrated a significant association between rs2107538 and increased susceptibility to tuberculosis (TB) (19).
The aim of this work was to evaluate the associations of polymorphisms in the TNF-α, IL-6, IL-8, IL-10, CCL5 and CXCL6 genes with COVID-19 outcomes and with the serum levels of IFN-α, IFN-γ, TNF-α, IL-1Ra, IL-6, IL-7, IL-10, CCL2, CCL3, CXCL8, CXCL10 and GCSF.
2 Materials and methods
2.1 Setting and participants
From June 2020 to March 2021, nonprobability sampling was performed with unvaccinated patients in a multicenter cross-sectional study from different Mexican governmental health care institutions. This study was approved by the ethics committee of the Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra (INR-LGII: 17/20). Informed consent was obtained from each participant. The inclusion criteria were as follows: either sex, age ≥18 years with clinical features of COVID-19, and a positive qRT–PCR test from a nasopharyngeal swab. The exclusion criteria were pregnancy, incomplete clinical records and individuals related to included participants. The participants were classified according to the WHO clinical progression scale and scored as follows: ambulatory mild disease (score of 1-3), hospitalized moderate disease (score of 4-5), severe disease (score of 6-9), and death (score of 10) (20).
2.2 Sample collection
Peripheral blood samples were collected at two days after hospital admission for COVID-19. One sample using EDTA anti-coagulant tubes was used for DNA extraction and other tube with clot activator/separator gel additives for serum collection. The latter was allowed to clot and centrifuged at 2,000 X G for 10 minutes to obtain the serum, which was stored at -80°C until further use. In the case of ambulatory patients, the samples were collected at the hospital´s triage, after the individual was confirmed to be positive by qRT-PCR test.
2.3 Selection and genotyping of SNPs
The polymorphisms of TNF-α (rs1800610 (+489 G/A), rs1800629 (-308 G/A), and rs3093664), IL-6 (rs1800796 (-572 G/C) and rs10499563 (-6331 T/C)), IL-8 (rs2227307), IL-10 (rs1800872 (-819 C/T) and rs1800071 (-592 C/A)), CXCL6 (rs4279174), and CCL5 (rs2107538 and rs3817656) were selected based on their previous scientific evidence of associations with different diseases in any population that included independent genetic studies from 2003–2020.
Genomic DNA was isolated using a specialized commercial kit (QIAmp DNA Blood Mini Kit, Qiagen, Hilden, Germany). The quality of the DNA samples was evaluated by the 260/280 nm absorbance ratio, and 1% agarose gels were stained with SYBR® Green (Invitrogen, CA, USA). The DNA concentration was subsequently quantified using a spectrophotometric method and adjusted to 20 ng/μl.
For genotyping, 10 ng/μl of genomic DNA was transferred into OpenArray plates, which previously contained the specific genotyping primers and probes, via the AccuFill system. Real-time PCR amplification was performed according to the supplier’s protocol via the OpenArray Platform through a Quant Studio 12K Flex System (Thermo Fisher Scientific, Waltham, MA, USA), and the results were analyzed using TaqMan Genotyper v1.6 software.
2.4 Soluble molecule evaluation
Serum IFN-α, IFN-γ, TNF-α, IL-1Ra, IL-2, IL-6, IL-7, IL-10, CCL2, CCL3, CXCL8, CXCL10 and GCSF levels were measured using a Human COVID-19 Cytokine Storm Panel 1 (14 plex) provided by Biolegend (Cat. No. 741088, Lot B377360) following the manufacturer´s instructions.
2.5 Statistical analysis
The normality of the variable distributions was evaluated. The Kruskal–Wallis test was used to compare nonparametric continuous variables among the studied groups, and the results are presented as the median and interquartile range (IQR). For the categorical variables, the chi-square test was performed. For all tests, a value of P <0.05 was considered to indicate statistical significance.
In addition, the linkage disequilibrium (LD) among all variants was determined using HaploView software V4.2. We tested 11 SNPs, Bonferroni test corrections was applied and correction of the P value was set at (α = 0.05/11) α = 0.004.
Logistic regression analysis adjusted for age, sex, hypertension status, type 2 diabetes status and obesity status was used to evaluate associations between genetic variants and the outcomes of COVID-19 patients. The final models were evaluated using the Hosmer–Lemeshow goodness-of-fit test. The analysis was performed using the STATA v.16 statistical package (StataCorp Texas, USA).
3 Results
3.1 Study population
In the present work, 627 subjects were included and stratified in 20% mild (n=123), 36% moderate (n=229), 26% severe (n=162) and 18% dead (n=113). Table 1 shows the features and clinical information of the study population. The median age of the total population was 52 years (IQR 43-63), 63% were males, and the principal comorbidities were obesity in 31% (n=197), hypertension in 30% (n=188) and type 2 diabetes mellitus in 31% (n=193).
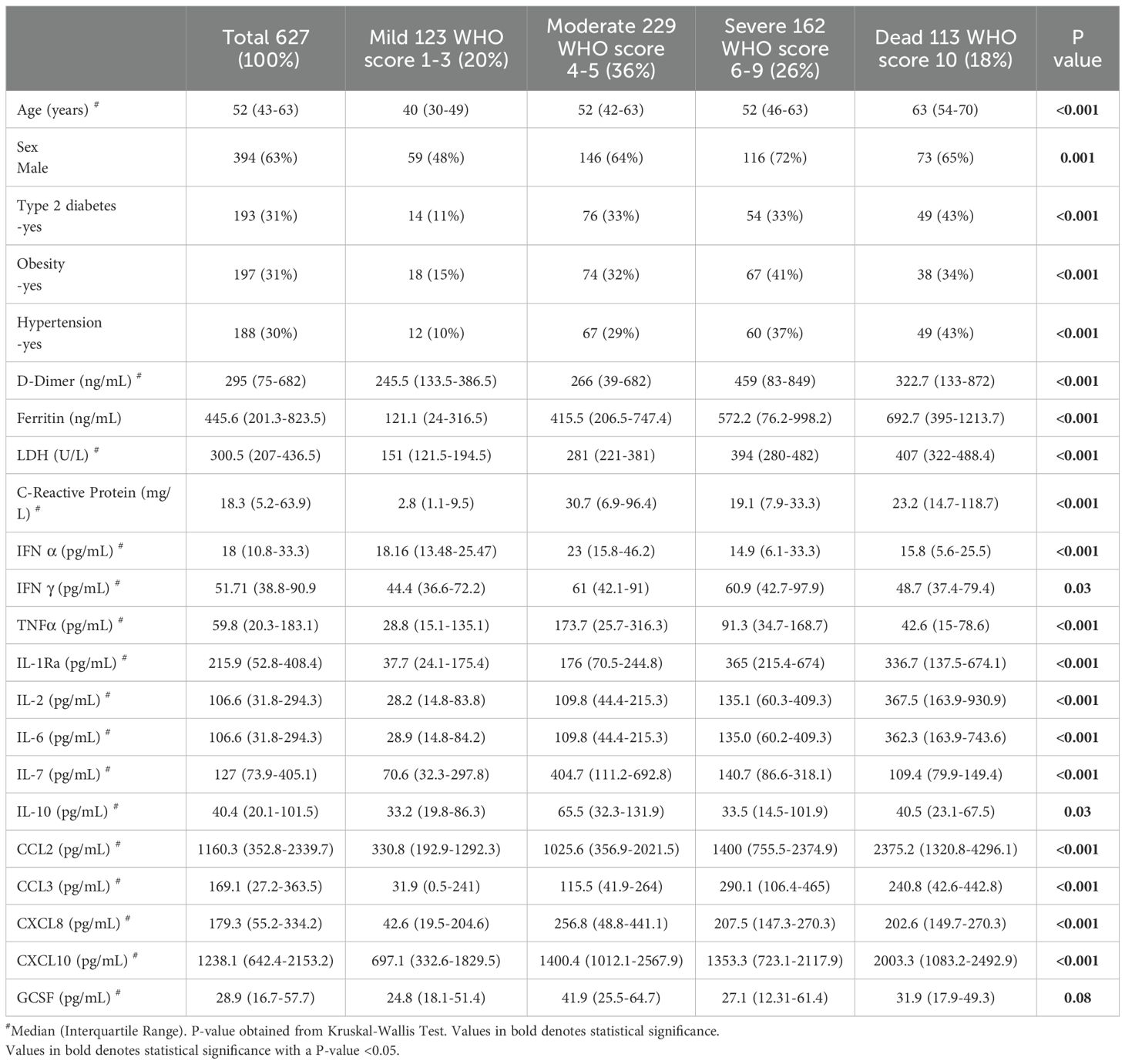
Table 1. Anthropometric and clinical characteristics of population study.
For clinical biomarkers, we observed statistical significance (p<0.001) among the groups, where ferritin, lactate dehydrogenase (LDH) and CRP levels tended to increase with the severity of the disease (Table 1).
3.2 Cytokine levels among severity COVID-19
Concerning IL-6 and CCL2, we observed a tendency toward higher levels (pg/mL) as the severity of the disease increased. Similarly, CXCL-10 showed the highest concentration in the deceased group, and IL-1Ra and CCL3 also presented the highest concentrations in the severe and deceased COVID-19 groups. Interestingly, the concentrations of IFN-α, IFN-γ, TNF-α, IL-2, IL-10, CXCL8 and GCSF were the highest in the moderate disease group (Table 1). We performed a Spearman correlation analysis with the WHO severity score and observed a moderate positive correlation with IL-6 (ρ=0.53), IL-1 (ρ=0.52) and CCL2 (ρ=0.43) (p ≤ 0.0001).
3.3 Allelic and genotypic frequencies and linkage disequilibrium
The allelic and genotypic frequencies are shown in Supplementary Table S1. We observed a significant difference in the distribution of the allele frequencies among the study groups for the IL-6 rs1800796 (-572 G/C) and rs10499563 (-6331 T/C); IL-10 rs1800871 (-819 T/C) and rs1800872 (-592 C/A); and CCL5 rs3817656 polymorphisms (Supplementary Table S1). The LD was calculated for all SNPs; however, only rs1800871 (-819 T/C) and rs1800872 (-592 C/A) of the IL-10 gene showed LD, with R2 values of 0.99 and D´=0.99, as did rs2107538 and rs3817656 of CCL5 (R2 values of 0.79 and D´=0.95).
3.4 Logistic regression analysis
In the Figure 1, showed the association of the SNPs, among COVID-19 severity. For the TNF-α gene, we observed an association of the rs1800610 risk allele with moderate disease (OR=1.5, 95% CI= 1.01-2.24, p=0.04), as well as a protective association of the rs1800629 minor allele with moderate disease (OR=0.45, 95% CI95% CI= 0.22-0.94, p=0.03) and severe outcomes (OR=0.34, 95% CI95% CI=0.14-0.83, p=0.02) (Supplementary Table S2). In severe outcomes, for IL-6 gene, we found an association for the rs1800796 (-572 G/C) C allele (OR of 1.64 (95% CI=1.05–2.57, p=0.03). Additionally, for the CC genotype of the codominant model, the OR was 2.83 (95% CI=1.11–7.26; p=0.03). In addition, for the CC of the recessive model, we found an OR of 2.44 (95% CI=1.01-5.87, p=0.04) (Supplementary Table S3). For rs1049953 (-6331T/C), we did not observe a significant association with any outcome.
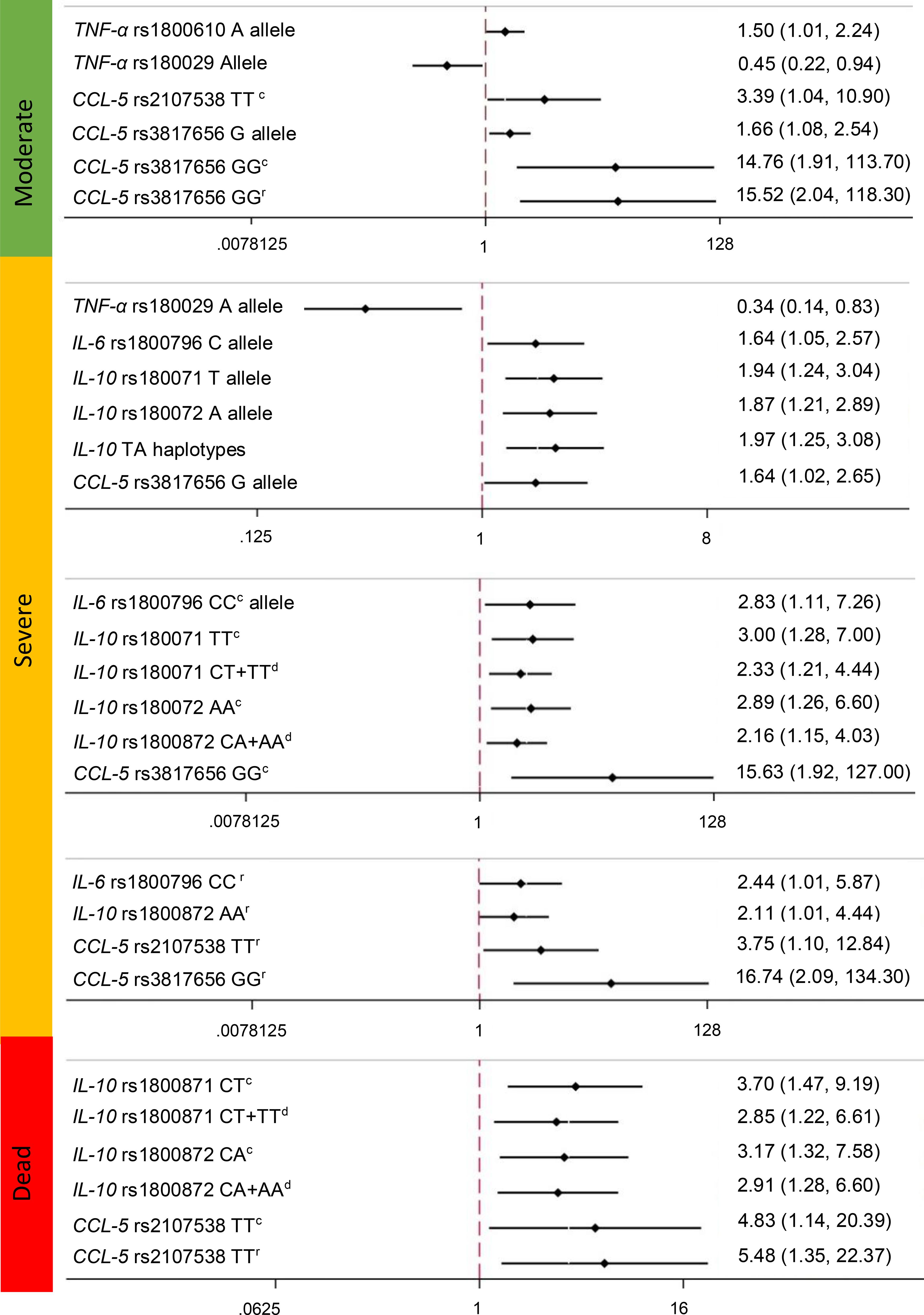
Figure 1. Association of the Polymorphisms of TNFα (rs1800610, rs1800629, rs3093664), IL-6 (rs1800796, rs10499563), IL-10 (rs1800872 and rs1800071), and CCL5 (rs2107538, rs3817656) genes and COVID-19 outcomes. c, Codominant; d, dominant; r:recesive. OR (CI). Association statistically significant differences among groups. The models were adjusted by the main covariates: type two diabetes, obesity, hypertension, sex and age.
Interestingly, for the IL-10 gene rs1800871 (-819 C/T) polymorphism, we observed a significant association between the risk allele and severe outcomes with an OR of 1.94 (95% CI=1.24-3.04, p=0.004). The TT genotype of the codominant model had an OR=3.00 (95% CI=1.28-7.00, p=0.01), and the CT+TT genotypes of the dominant model had an OR=2.33 (95% CI 1.21-4.44, p=0.01) for severe outcomes. In addition, for the deceased outcome, we observed a significant association with CT in the codominant model (OR=3.7 95% CI=1.47-9.19, p=0.005) and CT+TT in the dominant model (OR=2.85 95% CI=1.22-6.61, p=0.02). For rs1800872 (-592 C/A), the A allele was associated with severe and deceased outcomes, with ORs of 1.87 (95% CI=1.21-2.89, p=0.004) and 1.73 (95% CI=1.01-2.9, p=0.04), respectively. The AA genotype of the codominant model had an OR of 2.89 (95% CI=1.26-6.60, p=0.01), the CA+AA genotype of the dominant model had an OR=2.16 (95% CI=1.15-4.03, p=0.02), and the AA genotype of the recessive model had an OR=2.11 (95% CI =1.01-4.44, p=0.04), all of which were associated with severe outcomes. In addition, we observed a significant association between the CA genotype of the codominant model and the CA+AA genotypes of the dominant model with a deceased outcome, with ORs of 3.17 (95% CI=1.32-7.58, p=0.009) and 2.91 (95% CI=1.28-6.60, p=0.01), respectively (Figure 1).
We performed logistic regression with the haplotypes and found a significant association between the CT haplotype and severe outcomes, with an OR=1.97 (95% CI=1.25-3.08, p=0.003) (Supplementary Table S2).
For the CCL5 gene, the rs2107538 in the codominant model (AA genotype) was significantly associated with moderate (OR= 3.39 95% CI= 1.04-10.9, p=0.04) and deceased (OR= 4.83 95% CI= 1.14-20.39, p=0.03) outcomes. The recessive model (TT genotype) tended to increase the associations among the COVID-19 outcomes (Supplementary Table S2). Variant rs3817656, the G allele, was significantly associated with moderate (OR= 1.66 95% CI= 1.08-2.54, p=0.02) and severe (OR= 1.64 95% CI= 1.02-2.65, p=0.04) COVID-19 outcomes (Supplementary Table S2).
When we adjusted by Bonferroni correction, only the IL-10 gene rs1800871 (-819 C/T) polymorphism conserved the significant association between the T allele and severe outcome, with an OR of 1.94 (95% CI=1.24-3.04, p=0.004), as well as the association seen for the A allele of the rs1800872 polymorphism with severe outcome (OR=1.87 (95% CI=1.21-2.89, p=0.004).
3.5 Cytokine levels and SNPs
We performed a sub-analysis to determine the relationships between the concentrations of the cytokines previously mentioned with the SNPs. In this analysis, the carriers of the risk allele of the TNF-α rs1800629 polymorphism presented lower levels of IL-1Ra, IL-6, CCL-2, and CXCL10 than did the carriers of the major allele (Supplementary Table S3).
Interestingly, we observed differences for the rs1049953 (-6331T/C) polymorphism of the IL-6 gene concerning the level of IFN-α. In addition, we observed that carriers of the risk allele of this SNP had the highest serum concentrations of IL-6 and CCL-2 (Supplementary Table S3).
Similarly, for the CCL-5 rs2107538 polymorphism, carriers of the A allele had the highest serum levels of the IL-2, IL-6, IL-7, IL-10, CCL-2, GCSF, and CXCL10 cytokines. For rs3817656, carriers of the risk allele presented increased serum levels of the cytokines IL-2, IL-6, IL-7, CCL-2, GCSF, and CXCL8. These differences were also found to be significant for the CCL-5 haplotype AG, except the GCSF concentration, which did not reach significance (Supplementary Table S3).
4 Discussion
In COVID-19, severe outcomes have increased levels of pro-inflammatory and anti-inflammatory cytokines and chemokines, such as IL-6, TNF-α, IL-10, CCL2, CCL3, CXCL8, and CXCL10, among others (1, 3, 4). Since COVID-19 emerged, several studies across the world have reported some polymorphisms of different genes related to this disease (4, 5). SNPs in the TNF-α, IL-6, IL-8, IL-10, CCL5 and CXCL6 genes have been associated with COVID-19 outcomes and with serum cytokine and chemokine levels (3, 6).
We observed a trend toward increased CCL2 levels with the outcomes of the COVID-19 patients, as well as, of IL-1Ra and CCL3 levels being elevated in patients with severe disease. These observations are consistent to those reported by Torres-Ruiz et al. (2021), who reported a tendency for CCL2, CCL3, and other molecules to increase with disease severity (3, 6). Additionally, Dorgham et al. (2021) revealed increased levels of IFN-α and, TNF-α in patients who died from COVID-19, supporting the idea that inflammatory cytokines play a key role in disease progression (21, 22).
Several genome-wide association studies (GWASs) and candidate gene studies have identified SNPs associated with susceptibility to SARS-CoV-2 infection and severe COVID-19 (4, 5, 23).
TNF-α promotes detrimental tissue damage and gradual lung fibrosis, which results in pneumonia, pulmonary edema, and acute respiratory distress syndrome (21). In addition, the TNF-α gene is highly polymorphic, the rs1800610 is located in the first intron of the gene +489 (G/A), and rs1800629, which is located in the promoter region of the gene -308 (G/A). Our results revealed that only the rs1800610 A allele was significantly associated with the risk of a moderate COVID-19 outcome. This SNP in the Australian population was associated with the risk of developing breathing problems with bronchial exacerbation, suggesting that this SNP could contribute to disease progression. In addition, rs1800610 tends to increase cytokine concentrations in carriers of the A allele, and is related to prostate cancer and COPD development (7, 15). We were also able to demonstrate that the rs1800629 (-308 G/A) was found to be associated with protection against moderate and severe disease. The protective effect of rs1800629 (-308 G/A) is consistent with that reported by Heideri Nia et al., who showed that allele A decreases the risk of SARS‐CoV‐2 infection (24).
Interestingly, we identified a significant association between the A allele of the rs1800629 SNP with severe COVID-19. Due to the absence of AA genotype among individuals with mild, severe, and dead outcomes, standard genetic model analyses could not be performed. Nevertheless, rs1800629 has been previously reported to modulate the expression of key components within the inflammatory signaling cascade, which could influence the production of pro-inflammatory cytokines such as IL-6, TNF-α, IFN-γ, particularly in patients with metabolic syndrome (25, 26).
The role of IL-6 has been implicated in the regulation of genes and proteins involved in angiogenesis and immune cell recruitment (6). In addition to these roles, IL-6 exerts pleiotropic effects, contributing to a wide range of physiological and pathological processes, including inflammation, tumorigenesis, and immune modulation (8).
Genetic variations in the IL-6 promoter region, particularly the SNPs rs1800796 (-572 G/C) and rs10499563 (-6331 T/C), have been associated with immunological regulation of hepatocellular carcinoma and other inflammatory pathologies. These SNPs contribute to the functional regulation of expression of this gene (2, 27). We found that the C allele of the (-572 G/C) rs1800796 polymorphism was associated with severe COVID-19. However, Falahi et al. reported no association between the (-572 G/C) rs1800796 polymorphism and COVID-19 severity in the Kurdish population (28). However, their logistic regression analyses were not adjusted for major comorbidities and were conducted on a relatively small sample size, which may limit the reliability of their findings. By contrast, our study benefits from a larger sample size and incorporates adjustments for key comorbid conditions, thereby enhancing the robustness and validity of the results.
Karcioglu et al. investigated the correlation between rs1800796 (-572 G/C) frequency and incidence and mortality of COVID-19 across 23 countries. Their analysis did not reveal a statistically significant correlation. However, it is important to note that the dataset included two studies from Mexico, one involving patients with hip fractures and another involving individuals infected with HBV, neither of which focused on COVID-19 outcomes (29). Our study included individuals with severe COVID-19, and revealed genetic association with disease severity, supported by robust statistical models, showing the genetic contribution to this disease.
IL-8 has effects through binding to cognate G-protein-coupled CXC chemokine receptors, CXCR1 and CXCR2, which activate a phosphorylation cascade to trigger chemotaxis and neutrophil activation as part of the inflammatory response. In COVID-19, the plasma IL-8 concentration is increased, suggesting a significant role in cytokine release syndrome, multiorgan dysfunction, respiratory failure and shock (9, 30). SNPs in the IL-8 gene could influence the molecular mechanisms of COVID-19. Nevertheless, in our study, we did not find an association between rs2227307 and COVID-19 severity.
Previously, in Mexico, rs1800871 (-819 C/T) and rs1800872 (-592 C/A) were found not to be associated with COVID-19 severity. However, the sample size was small (193 subjects), which means that the statistical power was low (31). Abbood et al. (2023), in an Iranian population, reported that rs1800871 (-819 C/T) and r1800872 (-592 C/A) were related to COVID-19 mortality (32). Monroy-Muñiz et al. (2023) studied rs1800871 (-819 C/T) and r1800872 (-592 C/A) in a Mexican population and reported that the T and A alleles were associated with the risk of dengue infection (33). Our study reported that T and A alleles were associated to COVID-19 severity adjusted by Bonferroni test. The SNPs rs1800871 (-819 C/T) and rs1800872 (-592 C/A), of IL-10 gene, are located in the 5´-flanking region (7). In autoimmune diseases, like ankylosing spondylitis, increased levels of IL-10 in the serum have been described. Ling et al. (2021) reported that IL-10 serum levels are greater in patients admitted to the ICU than in those who are not (34). It has been reported that rs1800872 (-592 C/A) can reduce negative promoter function, and therefore modify IL-10 transcription (31, 32).
CXCL6 is a chemokine characterized by four conserved cysteine residues close to the amino terminus CXC motif; In lung inflammation, CXCL6 stimulates inflammation by recruiting and activating immune cells. However, in our study we did not observe a significant association between rs4279174 of the CXCL6 gene and COVID-19 severity. However, other variants in this gene may influence disease severity.
CCL5, also called RANTES, has been studied in many inflammatory diseases. Recently, the CCL5/CCR5 axis was reported to participate in the response to COVID-19, HIV and tuberculosis. Kouhpayeh et al. reported that the GA genotype in the dominant model of CCL5 (rs2107538) was associated with an increased risk of pulmonary tuberculosis (12). Our results revealed that this SNP in the recessive model was associated with COVID-19 severity. However, Pati et al. reported that this variant was associated with protection against SARS-CoV-2 infection, but the mechanisms are unknown and were not adjusted by confounding factors such as sex or age, among others (35). However, our analysis was adjusting for sex, age, type 2 diabetes and hypertension, we also observed higher concentrations of cytokines in the presence of the risk alleles. The rs21075238 is located at the binding site for GATA binding protein 2 (GATA2), which is involved in the transcriptional regulation of proinflammatory cytokines therefore, CCL5 might participate in the COVID-19 inflammatory process (36, 37).
CCL-2 participates in multiple lung inflammatory diseases, and its major mechanism involves mediating the migration of mononuclear cells into the microvasculature and airways. This could explain the monocyte and lymphocyte influx observed in ARDS, asthma and bronchiolitis obliterans syndrome (BOS) (38). A recent report by Abers et al. suggested that CCL-2 was associated with mortality due to COVID-19 (38). Pius-Sadowska et al. reported that lower concentrations of CCL-2 mRNA but higher concentrations of the protein in plasma were correlated with more severe courses of COVID-19, suggesting that CCL-2, like other cytokines, could be considered new prognostic factors for severe COVID-19 (39).
In our study, when stratifying serum cytokine concentrations by alleles of the SNPs investigated, we observed that carriers of the A allele of rs1800629 of TNF-α gene showed the lowest levels of IL-1Ra, IL-6, CCL2, and CXCL10, whereas carriers of the G allele showed higher levels. Consistent with our findings Szkpu M et al., reported that the G/G genotype of rs1800629, showed higher IL-6 levels in elderly women with metabolic syndrome. These observations support the hypothesis that autoinflammatory mechanisms may contribute significantly to the pathophysiology and complications of the disease. Furthermore, the A allele has been associated to increased transcriptional activity (26).
Interestingly, we observed higher levels of IFN-α, IL-6 and CCL2 with the C allele of the rs10499563 (-6331 T/C) of IL-6 gene. This SNPs is located near a distal regulatory sequence upstream of IL-6 expression via affecting the binding affinity of the transcription factor Oct-1 to its response element (Figure 2). Smith et al., demonstrated that this SNP is a significant predictor of increased IL-6 concentrations six hours after cardiac surgery. Other SNPs in close proximity to trs10499563 (-6331 T/C), such as rs18000795 and rs1800796 of IL-6 gene, have also been linked to elevated serum IL-6 levels. In our study, carriers of the variant allele of rs1800796 showed increased IL-6 levels, although this association did not reach statistical significance (40–42). Additionally, we observed increased levels of IFN-α, and CCL2 in patients with the variant alleles of the rs10499563 (-6331 T/C). These SNPs could contribute to increased IL-6 levels, which can increase CCL2 production and promote immune cell recruitment. While the impact of these SNPs on IFN-α is less clear, IL-6 can influence the broader immune environment, potentially enhancing IFN-α responses in viral infections (6, 23).
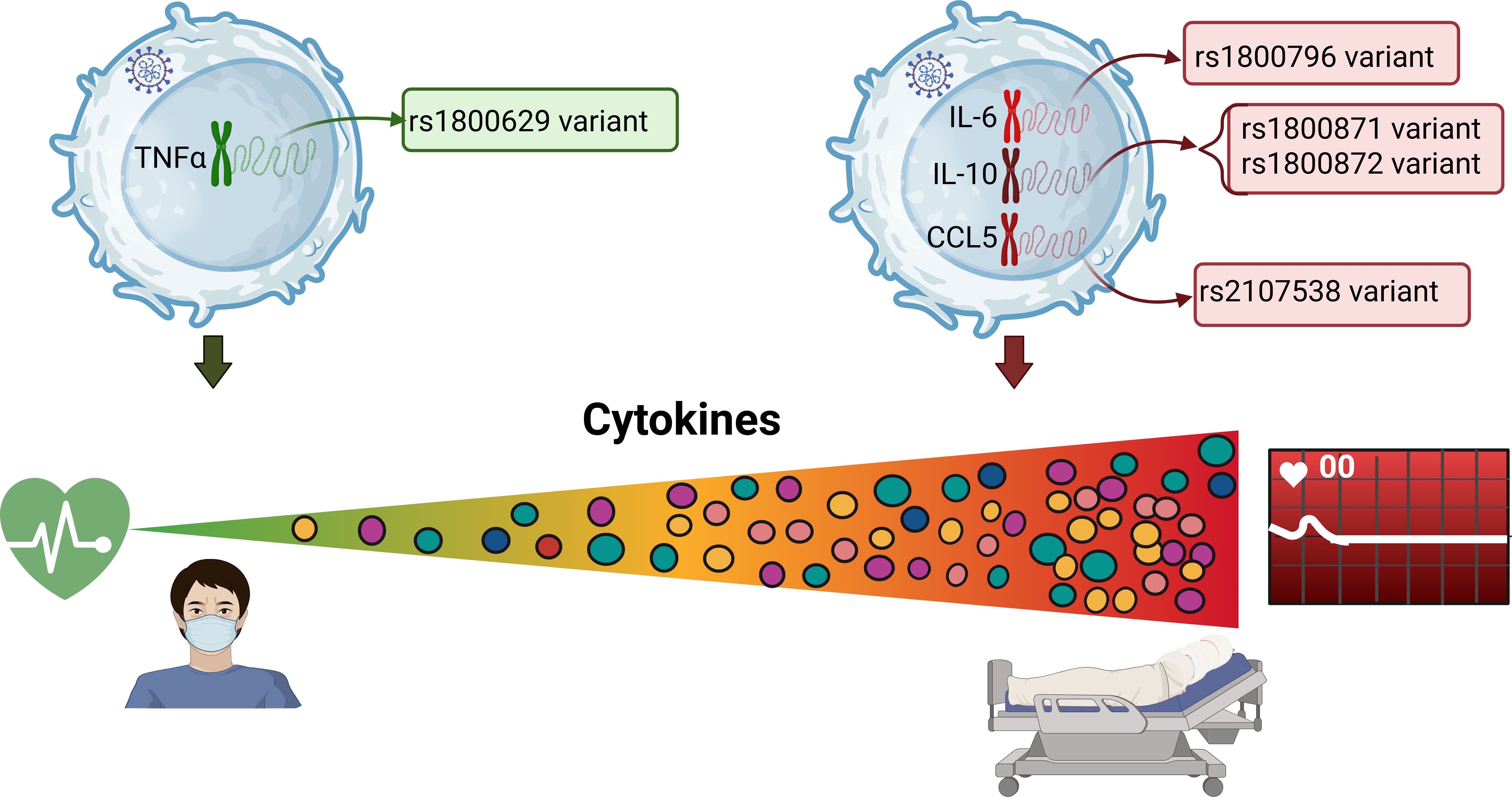
Figure 2. Association of the SNPs and cytokines, with COVID-19 severity.
We also observed elevated levels of IL2, IL-6, IL-10, CCL2, CXCL8, CXCL10 and GCSF in the T allele of the rs2107538 variant of CCL5 gene. Silva et al, reported that rs217538 has a positive regulatory impact on CCL5 protein production in HIV-1 infection. Additionally, Zhernakova reported that several studies have shown that rs2107538, located in the first intron, can influence the transcriptional activity of CCL5, potentially contributing to susceptibility to autoimmune diseases. However, these findings have primarily been derived from case-control studies (18, 43).
Genetic variability among different populations also help us understand the diversity of disease course and susceptibility to its complications. It is possible that others variants not evaluated in this study are in LD and could influence cytokine levels.
In clinical practice, the tendency of certain cytokines to increase with the severity of COVID-19 is a significant concern. Among them, IL-6 has emerged as a key driver of inflammation. Targeting IL-6 with specific therapies has become a potentially life-saving strategy for patients with severe COVID-19. There are three main types of IL-6 inhibitors, typically monoclonal antibodies, including siltuximab, tocilizumab and sarilumab. These drugs have been approved by the Food and Drug Administration (FDA), for use in various conditions. Further pharmacogenetics studies are needed to determine whether specific IL-6 gene variants could serve as prognostic markers for patients response to these therapies in COVID-19 (44, 45).
This is the first comprehensive study evaluating genetic profile and cytokine levels in unvaccinated Mexican COVID-19 patients, adjusted for main comorbidities. However, the study has several limitations, including the possibility that patients with other underlying conditions may have compromised immune system, as well as genetic substructure may influence the observed associations, or that the findings may only be representative of central Mexican populations.
Further investigations of other SNPs in these genes or others implicated in the mechanism of COVID-19 severity are necessary for developing prevention actions and precision medical therapies. It should be noted that our results are only representative of the Mexican-Mestizo population.
This study highlights the associations of the IL-6 rs1800796 (-572 G/C), IL-10 rs1800871 (-819 C/T), rs1800872 (-592 C/A) and CCL5 rs2107538 polymorphisms with fatal COVID-19 outcomes and elevation of cytokines (CCL-2, IL-6, IL-10 and CXC-L10) plays a crucial role in the pathogenesis of COVID-19. However, Patients infected with SARS-CoV-2 who have the TNF-α gene variant (rs1800629) are protected from developing COVID-19 moderate and severe outcomes, as well as from presenting low concentrations of some pro-inflammatory cytokines and chemokines (IL-6, CCL2, and CXCL-10).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Files. Further inquiries can be directed to the corresponding authors.
Ethics statement
The studies involving humans were approved by ethics committee of the Instituto Nacional de Rehabilitación Luis Guillermo Ibarra Ibarra (INR-LGII: 17/20). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
LM-G: Formal Analysis, Writing – original draft. CO-V: Writing – review & editing, Data curation, Visualization. MA-B: Writing – review & editing, Data curation, Visualization. LC-G: Visualization, Writing – review & editing, Data curation. CM-A: Data curation, Writing – review & editing, Visualization. RV-V: Data curation, Visualization, Writing – review & editing. JR-H: Data curation, Visualization, Writing – review & editing. PV-C: Resources, Investigation, Writing – review & editing. DG-M: Data curation, Writing – review & editing, Visualization. GV-A: Visualization, Data curation, Writing – review & editing. JR-P: Investigation, Writing – review & editing, Resources. LR-L: Investigation, Resources, Writing – review & editing. JF-G: Writing – review & editing, Investigation, Resources. JC: Writing – review & editing, Resources, Investigation. IC-B: Writing – review & editing, Methodology, Supervision. MM: Methodology, Supervision, Writing – review & editing. GV-Z: Methodology, Writing – review & editing, Supervision. AM-C: Supervision, Writing – review & editing, Methodology. P-T: Methodology, Writing – review & editing, Supervision. FM-R: Writing – review & editing, Supervision, Methodology. DZ-A: Software, Writing – review & editing, Validation. MO-S: Software, Validation, Writing – review & editing. YS-K: Validation, Software, Writing – review & editing. CS-A: Writing – review & editing, Validation, Software. JO-T: Validation, Writing – review & editing, Software. BH-L: Software, Validation, Writing – review & editing. CP: Writing – review & editing, Validation, Software. GM-N: Project administration, Writing – review & editing, Conceptualization. AL-R: Funding acquisition, Project administration, Writing – review & editing, Conceptualization.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by the Consejo Nacional de Ciencia y Tecnología; CONACYT 312513 SARS-COV 2.
Acknowledgments
We are grateful for the extraordinary efforts of health care workers, who sacrificed their lives while saving patients.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1641285/full#supplementary-material
References
1. Ghazavi A, Ganji A, Keshavarzian N, Rabiemajd S, and Mosayebi G. Cytokine profile and disease severity in patients with COVID-19. Cytokine. (2021) 137:155323. doi: 10.1016/j.cyto.2020.155323
2. Yessenbayeva A, Apsalikov B, Massabayeva M, Kazymov M, Shakhanova A, Mussazhanova Z, et al. Biomarkers of immunothrombosis and polymorphisms of IL2, IL6, and IL10 genes as predictors of the severity of COVID-19 in a Kazakh population. PloS One. (2023) 18:e0288139. doi: 10.1371/journal.pone.0288139
3. Zanza C, Romenskaya T, Manetti AC, Franceschi F, La Russa R, Bertozzi G, et al. Cytokine storm in COVID-19: immunopathogenesis and therapy. Med (Kaunas). (2022) 58. doi: 10.3390/medicina58020144
4. Fricke-Galindo I and Falfán-Valencia R. Genetics insight for COVID-19 susceptibility and severity: A review. Front Immunol. (2021) 12:622176. doi: 10.3389/fimmu.2021.622176
5. Chakravarty S. COVID-19: the effect of host genetic variations on host-virus interactions. J Proteome Res. (2021) 20:139–53. doi: 10.1021/acs.jproteome.0c00637
6. Santa Cruz A, Mendes-Frias A, Oliveira AI, Dias L, Matos AR, Carvalho A, et al. Interleukin-6 is a biomarker for the development of fatal severe acute respiratory syndrome coronavirus 2 pneumonia. Front Immunol. (2021) 12:613422. doi: 10.3389/fimmu.2021.613422
7. Santiago AM, da Silva Graça Amoras E, Queiroz MAF, da Silva Conde SRS, Cayres-Vallinoto IMV, Ishak R, et al. TNFA -308G>A and IL10 -1082A>G polymorphisms seem to be predictive biomarkers of chronic HCV infection. BMC Infect Dis. (2021) 21:1133. doi: 10.1186/s12879-021-06835-9
8. Song J, Park DW, Moon S, Cho HJ, Park JH, Seok H, et al. Diagnostic and prognostic value of interleukin-6, pentraxin 3, and procalcitonin levels among sepsis and septic shock patients: a prospective controlled study according to the Sepsis-3 definitions. BMC Infect Dis. (2019) 19:968. doi: 10.1186/s12879-019-4618-7
9. Grębowski R, Saluk J, Bijak M, Szemraj J, and Wigner P. Variability, expression, and methylation of IL-6 and IL-8 genes in bladder cancer pathophysiology. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms24076266
10. Lu L, Zhang H, Dauphars DJ, and He YW. A potential role of interleukin 10 in COVID-19 pathogenesis. Trends Immunol. (2021) 42:3–5. doi: 10.1016/j.it.2020.10.012
11. Korbecki J, Kojder K, Kapczuk P, Kupnicka P, Gawrońska-Szklarz B, Gutowska I, et al. The effect of hypoxia on the expression of CXC chemokines and CXC chemokine receptors-A review of literature. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms22020843
12. Kouhpayeh HR, Taheri M, Baziboroon M, Naderi M, Bahari G, and Hashemi M. CCL5 rs2107538 polymorphism increased the risk of tuberculosis in a sample of Iranian population. Prague Med Rep. (2016) 117:90–7. doi: 10.14712/23362936.2016.9
13. Danforth KN, Rodriguez C, Hayes RB, Sakoda LC, Huang WY, Yu K, et al. TNF polymorphisms and prostate cancer risk. Prostate. (2008) 68:400–7. doi: 10.1002/pros.20694
14. Talaat RM, Abdelkhalek MS, El-Maadawy EA, Abdel-Mageed WS, El-Shenawy SZ, and Osman MA. Association of TNF-Alpha gene polymorphisms and susceptibility to hepatitis B virus infection in Egyptians. Hum Immunol. (2017) 78:739–46. doi: 10.1016/j.humimm.2017.10.006
15. Yu S, Xue M, Yan Z, Song B, Hong H, and Gao X. Correlation between TNF-α -308 and +489 gene polymorphism and acute exacerbation of chronic obstructive pulmonary diseases. BioMed Res Int. (2021) 2021:6661281. doi: 10.1155/2021/6661281
16. Rizvi S, Rizvi SMS, Raza ST, Abbas M, Fatima K, Zaidi ZH, et al. Implication of single nucleotide polymorphisms in Interleukin-10 gene (rs1800896 and rs1800872) with severity of COVID-19. Egyptian J Med Hum Genet. (2022) 23:145. doi: 10.1186/s43042-022-00344-3
17. Alvarez AE, Marson FAL, Bertuzzo CS, Bastos JCS, Baracat ECE, Brandão MB, et al. Association between single nucleotide polymorphisms in TLR4, TLR2, TLR9, VDR, NOS2 and CCL5 genes with acute viral bronchiolitis. Gene. (2018) 645:7–17. doi: 10.1016/j.gene.2017.12.022
18. Silva MJA, Marinho RL, Dos Santos PAS, Dos Santos CS, Ribeiro LR, Rodrigues YC, et al. The association between CCL5/RANTES SNPs and susceptibility to HIV-1 infection: A meta-analysis. Viruses. (2023) 15. doi: 10.3390/v15091958
19. Areeshi MY, Mandal RK, Panda AK, and Haque S. A meta-analysis of the association between the CC chemokine ligand 5 (CCL5) -403 G>A gene polymorphism and tuberculosis susceptibility. PloS One. (2013) 8:e72139. doi: 10.1371/journal.pone.0072139
20. WHO Working Group on the Clinical Characterisation and Management of COVID-19 infection. A minimal common outcome measure set for COVID-19 clinical research. Lancet Infect Dis. (2020) 20:e192–e7. doi: 10.1016/S1473-3099(20)30483-7
21. Dorgham K, Quentric P, Gökkaya M, Marot S, Parizot C, Sauce D, et al. Distinct cytokine profiles associated with COVID-19 severity and mortality. J Allergy Clin Immunol. (2021) 147:2098–107. doi: 10.1016/j.jaci.2021.03.047
22. García-Ramírez RA, Ramírez-Venegas A, Quintana-Carrillo R, Camarena ÁE, Falfán-Valencia R, and Mejía-Aranguré JM. TNF, IL6, and IL1B polymorphisms are associated with severe influenza A (H1N1) virus infection in the mexican population. PloS One. (2015) 10:e0144832. doi: 10.1371/journal.pone.0144832
23. Choudhri Y, Maslove DM, and Rauh MJ. COVID-19 and the genetics of inflammation. Crit Care Med. (2023) 51:817–25. doi: 10.1097/CCM.0000000000005843
24. Heidari Nia M, Rokni M, Mirinejad S, Kargar M, Rahdar S, Sargazi S, et al. Association of polymorphisms in tumor necrosis factors with SARS-CoV-2 infection and mortality rate: A case-control study and in silico analyses. J Med Virol. (2022) 94:1502–12. doi: 10.1002/jmv.27477
25. Janssen I, Powell LH, Crawford S, Lasley B, and Sutton-Tyrrell K. Menopause and the metabolic syndrome: the Study of Women’s Health Across the Nation. Arch Intern Med. (2008) 168:1568–75. doi: 10.1001/archinte.168.14.1568
26. Szkup M, Chełmecka E, Lubkowska A, Owczarek AJ, and Grochans E. The influence of the TNFα rs1800629 polymorphism on some inflammatory biomarkers in 45-60-year-old women with metabolic syndrome. Aging (Albany NY). (2018) 10:2935–43. doi: 10.18632/aging.101600
27. Aleagha OE, Oltulu P, and Sadeghi M. Association between interleukin 6 polymorphisms (rs1800796, rs1800795, rs2069837, rs17147230, and rs1800797) and hepatocellular carcinoma susceptibility: a meta-analysis. Clin Exp Hepatol. (2020) 6:359–66. doi: 10.5114/ceh.2020.102171
28. Falahi S, Zamanian MH, Feizollahi P, Rezaiemanesh A, Salari F, Mahmoudi Z, et al. Evaluation of the relationship between IL-6 gene single nucleotide polymorphisms and the severity of COVID-19 in an Iranian population. Cytokine. (2022) 154:155889. doi: 10.1016/j.cyto.2022.155889
29. Karcioglu Batur L and Hekim N. Correlation between interleukin gene polymorphisms and current prevalence and mortality rates due to novel coronavirus disease 2019 (COVID-2019) in 23 countries. J Med Virol. (2021) 93:5853–63. doi: 10.1002/jmv.27127
30. Cesta MC, Zippoli M, Marsiglia C, Gavioli EM, Mantelli F, Allegretti M, et al. The role of interleukin-8 in lung inflammation and injury: implications for the management of COVID-19 and hyperinflammatory acute respiratory distress syndrome. Front Pharmacol. (2021) 12:808797. doi: 10.3389/fphar.2021.808797
31. Avendaño-Félix M, Ochoa-Ramírez LA, Ramos-Payán R, Aguilar-Medina M, Ayala-Ham A, Rendón-Aguilar H, et al. Lack of effects of the genetic polymorphisms of interleukin-10 in clinical outcomes of COVID-19. Viral Immunol. (2021) 34:567–72. doi: 10.1089/vim.2021.0022
32. Abbood SJA, Anvari E, and Fateh A. Association between interleukin-10 gene polymorphisms (rs1800871, rs1800872, and rs1800896) and severity of infection in different SARS-CoV-2 variants. Hum Genomics. (2023) 17:19. doi: 10.1186/s40246-023-00468-6
33. Eloisa Monroy-Muñoz I, Esteban Muñoz-Medina J, Manuel Fragoso J, Esperanza Santacruz-Tinoco C, Sevilla-Montoya R, Hidalgo-Bravo A, et al. Genetic polymorphisms rs1800871 and rs1800872 of IL-10 gene are associated with dengue infection, especially with serotype 1 and DwoWS in Mexican population. Cytokine. (2023) 166:156194. doi: 10.1016/j.cyto.2023.156194
34. Ling L, Chen Z, Lui G, Wong CK, Wong WT, Ng RWY, et al. Longitudinal cytokine profile in patients with mild to critical COVID-19. Front Immunol. (2021) 12:763292. doi: 10.3389/fimmu.2021.763292
35. Pati A, Nayak N, Sarangi S, Barik D, Kumar Nahak S, Padhi S, et al. CCL5 rs2107538 variant is associated with protection against SARS-coV-2 infection and related mortality: A population-based study. J Infect Dis. (2022) 225:1865–6. doi: 10.1093/infdis/jiac026
36. Katsumura KR, Ong IM, DeVilbiss AW, Sanalkumar R, and Bresnick EH. GATA factor-dependent positive-feedback circuit in acute myeloid leukemia cells. Cell Rep. (2016) 16:2428–41. doi: 10.1016/j.celrep.2016.07.058
37. Wen D, Du X, Nie SP, Dong JZ, and Ma CS. Association between RANTES gene polymorphisms and asthma: a meta-analysis. PloS One. (2014) 9:e90460. doi: 10.1371/journal.pone.0090460
38. Abers MS, Delmonte OM, Ricotta EE, Fintzi J, Fink DL, de Jesus AAA, et al. An immune-based biomarker signature is associated with mortality in COVID-19 patients. JCI Insight. (2021) 6. doi: 10.1172/jci.insight.144455
39. Pius-Sadowska E, Niedźwiedź A, Kulig P, Baumert B, Sobuś A, Rogińska D, et al. CXCL8, CCL2, and CMV seropositivity as new prognostic factors for a severe COVID-19 course. Int J Mol Sci. (2022) 23. doi: 10.3390/ijms231911338
40. Bank S, Andersen PS, Burisch J, Pedersen N, Roug S, Galsgaard J, et al. Associations between functional polymorphisms in the NFκB signaling pathway and response to anti-TNF treatment in Danish patients with inflammatory bowel disease. Pharmacogenom J. (2014) 14:526–34. doi: 10.1038/tpj.2014.19
41. Smith AJ, D’Aiuto F, Palmen J, Cooper JA, Samuel J, Thompson S, et al. Association of serum interleukin-6 concentration with a functional IL6 -6331T>C polymorphism. Clin Chem. (2008) 54:841–50. doi: 10.1373/clinchem.2007.098608
42. Xia C, Zhu W, Huang C, Lou G, Ye B, Chen F, et al. Genetic polymorphisms of interleukin-6 influence the development of hepatitis B virus-related liver cirrhosis in the Han Chinese population. Infect Genet Evol. (2020) 84:104331. doi: 10.1016/j.meegid.2020.104331
43. Zhernakova A, Alizadeh BZ, Eerligh P, Hanifi-Moghaddam P, Schloot NC, Diosdado B, et al. Genetic variants of RANTES are associated with serum RANTES level and protection for type 1 diabetes. Genes Immun. (2006) 7:544–9. doi: 10.1038/sj.gene.6364326
44. Grebenciucova E and VanHaerents S. Interleukin 6: at the interface of human health and disease. Front Immunol. (2023) 14:1255533. doi: 10.3389/fimmu.2023.1255533
Keywords: COVID-19, IL-6, IL-10, CCL-2, polymorphism, SARS-CoV-2
Citation: Martínez-Gómez LE, Oropeza-Vélez CI, Almonte-Becerril M, Chavez-Galan L, Martinez-Armenta C, Vidal-Vázquez RP, Ramírez-Hinojosa JP, Vázquez-Cárdenas P, Gómez-Martín D, Vargas-Alarcón G, Rodríguez-Pérez JM, Ramón-Luing LA, Flores-Gonzalez J, Carrasco JG, Cruz-Bautista I, Mata-Miranda MM, Vázquez-Zapién GJ, Martínez-Cuazitl A, Parra-Torres NM, Martínez-Ruiz FdJ, Zayago-Angeles DM, Ordoñez-Sánchez ML, Segura-Kato Y, Suarez-Ahedo C, Olea-Torres J, Herrera-López B, Pineda C, Martínez-Nava GA and López-Reyes AG (2025) Association of pro-inflammatory and anti-inflammatory cytokine polymorphisms with COVID-19 severity in unvaccinated patients. Front. Immunol. 16:1641285. doi: 10.3389/fimmu.2025.1641285
Received: 04 June 2025; Accepted: 28 July 2025;
Published: 13 August 2025.
Edited by:
Patricia Maria Lourenço Dutra, Universidade do Estado do Rio de Janeiro, BrazilReviewed by:
Kokouvi Kassegne, Shanghai Jiao Tong University, ChinaNurhafiza Zainal, University of Malaya, Malaysia
Copyright © 2025 Martínez-Gómez, Oropeza-Vélez, Almonte-Becerril, Chavez-Galan, Martinez-Armenta, Vidal-Vázquez, Ramírez-Hinojosa, Vázquez-Cárdenas, Gómez-Martín, Vargas-Alarcón, Rodríguez-Pérez, Ramón-Luing, Flores-Gonzalez, Carrasco, Cruz-Bautista, Mata-Miranda, Vázquez-Zapién, Martínez-Cuazitl, Parra-Torres, Martínez-Ruiz, Zayago-Angeles, Ordoñez-Sánchez, Segura-Kato, Suarez-Ahedo, Olea-Torres, Herrera-López, Pineda, Martínez-Nava and López-Reyes. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriela A. Martínez-Nava, YW1lcmlhLmp1c3RpY2VAZ21haWwuY29t; Alberto G. López-Reyes, YWxsb3JleUB5YWhvby5jb20=