 Thomas Rouma1,2,3†
Thomas Rouma1,2,3† Emeline Barbieux1,2,3†
Emeline Barbieux1,2,3† Lorenzo Bossi3,4,5†Katy Poncin1Sébastien Denanglaire3,5Yohannes Tafesse3,6,7
Lorenzo Bossi3,4,5†Katy Poncin1Sébastien Denanglaire3,5Yohannes Tafesse3,6,7 David Vermijlen3,6,7Sandrine Delbauve3,6
David Vermijlen3,6,7Sandrine Delbauve3,6 Véronique Flamand3,6Juliette Van Buylaere8,9
Véronique Flamand3,6Juliette Van Buylaere8,9 Charles Van der Henst8,9
Charles Van der Henst8,9 Xavier De Bolle1
Xavier De Bolle1 Fabienne Andris3,5‡
Fabienne Andris3,5‡ Eric Muraille1,2,3*‡
Eric Muraille1,2,3*‡- 1Unité de Recherche en Biologie des Microorganismes (URBM) - Laboratoire d’Immunologie et de Microbiologie, NAmur Research Institute for LIfe Sciences (NARILIS), University of Namur, Namur, Belgium
- 2Laboratoire de Parasitologie, Université Libre de Bruxelles (ULB), Gosselies, Belgium
- 3ULB Center for Research in Immunology (U-CRI), Gosselies, Belgium
- 4ImmunXperts SA, a Q² Solutions Company, Gosselies, Belgium
- 5Immunobiology Laboratory, ULB, Gosselies, Belgium
- 6Institute for Medical Immunology, ULB, Gosselies, Belgium
- 7Department of Pharmacotherapy and Pharmaceutics, ULB, Gosselies, Belgium
- 8Microbial Resistance and Drug Discovery, VIB-VUB Center for Structural Biology, VIB, Flanders Institute for Biotechnology, Brussels, Belgium
- 9Structural Biology Brussels, Vrije Universiteit Brussel (VUB), Brussels, Belgium
Background: Acinetobacter baumannii is an opportunistic bacterium that causes serious nosocomial infections, including pneumonia and bacteremia, especially in immunocompromised individuals.
Methods: Here, we first evaluated the protective efficacy of a vaccination protocol with the heat-killed (HK) A. baumannii strain LAC-4 in various models of immunodeficient C57BL/6 mice challenged with pulmonary infection by LAC-4. We then examined the ability of HK LAC-4 to stimulate peripheral blood mononuclear cells (PBMCs) from healthy donors in vitro.
Results: We observed that mice deficient in Th1 (IL-12p35-/-, IFN-γ-/-), Th17 (IL-17RA-/-) and T cells (δTCR-/-, TAP1-/-, CD3-/-) display higher susceptibility to LAC-4 infection, but that our protocol improves their resistance. In contrast, vaccinated B cell-deficient (MuMT-/-) mice appear unable to control the infection, demonstrating that humoral immunity is essential to vaccine protection. Vaccination of wild-type mice with an HK ΔitrA strain deficient for capsule production failed to induce protection, showing that protective antibodies are mainly directed against the capsule. Our vaccination protocol also confers increased protection in wild-type mice vaccinated and then treated with cyclophosphamide; an immunosuppressive drug described to strongly increase the susceptibility of mice to A. baumannii infection. Finally, we demonstrate that HK LAC-4 can induce the activation of human monocyte-derived dendritic cells and T lymphocytes from PBMCs of healthy donors, suggesting that it may activate the human adaptive immune system and induce a protective memory response against A. baumannii.
Conclusions: Overall, our results demonstrate that administration of HK bacteria can induce protective immunity against A. baumannii in both immuno-competent and immuno-compromised mice and that these HK bacteria can activate the human adaptive immune system.
1 Introduction
Acinetobacter baumannii is an aerobic Gram-negative coccobacillus and an opportunistic nosocomial pathogen (1, 2). It is responsible for a wide range of human infections, including ventilator-associated pneumonia, bloodstream infections, wound and burn infections, urinary tract infections, and meningitis. Prolonged hospital stays—especially in intensive care units (ICU)—mechanical ventilation, indwelling foreign devices, extended exposure to antimicrobial treatment and immunodepression have been linked to multidrug-resistant A. baumannii infections (3). A relatively recent development is the association of A. baumannii with infections following injuries sustained in conflict zones, such as those in Iraq and Afghanistan (4, 5). Some evidence suggests that the use of morphine — commonly administered after battlefield or crush injuries — may enhance A. baumannii infections, possibly due to its immunosuppressive effects (6).
The genome of A. baumannii presents great diversity, with 2,119 genes forming the core genome and > 16,000 the pan genome (7), and high plasticity due to a very high rate of horizontal gene transfer (8). As a consequence, A. baumannii has an extraordinary ability to rapidly acquire resistance to antibiotics (9). Multidrug resistant strains (MDR) of A. baumannii have been documented worldwide. Several isolates are resistant to all commercially available antibiotics (7, 10). A. baumannii produces a high molecular weight capsular polysaccharide (11) which surrounds the outer membrane and makes it highly resistant to desiccation, disinfectants (12, 13) and reactive oxygen species (14). Significant variation in capsular polysaccharide structures between A. baumannii isolates has been identified (11), with several hundred distinct capsule types, incorporating a vast variety of sugars (15). A. baumannii is also able to form biofilms on biotic and abiotic surfaces, which further increases its survival on medical equipment as well as its resistance to detergents (9).
These characteristics make A. baumannii a pathogen particularly well suited to the hospital environment and a danger to patients hospitalized for long periods. Recently, the incidence of MDR A. baumannii co-infection has increased in intensive care units due to prolonged hospitalization of COVID-19 patients (16, 17). For example, among hospitalized COVID-19 patients in Wuhan, China, nearly half of the individuals who developed secondary bacterial infections died, and A. baumannii accounted for 36% of those infections (18). This phenomenon could partly be a consequence of the administration of high doses of dexamethasone, a steroid well known for its immunosuppressive effects, to treat patients with Covid-19 (19).
Consequently, the World Health Organization has defined carbapenem-resistant A. baumannii as a Priority 1 ‘‘critical’’ organism, and ranked it the number 1 organism recommended for new research and development (20). It has also been placed on the ESKAPE Restricted Pathogen List and ranked as a top priority by the US Centers for Disease Control and Prevention (21).
Vaccination is one of the most effective measures for infection control and is likely to be unaffected by the drug resistance of the pathogen. However, the development of a vaccine against A. baumannii has been plagued with several specific problems. The great variability of the A. baumannii genome (8) and capsule (11, 15) makes the development of a subunit vaccine focusing the adaptive response on a small number of antigenic determinants risky. Moreover, little is known about the protective immune response to A. baumannii, which limits our ability to identify immune markers for selecting vaccine candidates based on the responses they elicit. Studies in mouse models show that intranasal inoculation of A. baumannii induces high production of pro-inflammatory cytokines and rapid recruitment of neutrophils to the lungs. Several pattern recognition receptors, such as TLR4 (22), TLR9 (23) and NOD2 (24) have been implicated in A. baumannii detection and control, suggesting that a rapid response to infection is critical to its control. Complement, macrophages and neutrophils have been demonstrated to play a key role (25, 26). On the other hand, A. baumannii possesses several virulence mechanisms that allow it to escape the immune system and resist environmental stress. In particular, its capsule plays an important role in protection against destruction by host cells and escape from the innate immune response (27, 28). In particular, very little is known about the protective adaptive response against A. baumannii infection. The role of Th1 and Th17 responses, as well as the importance of the humoral and cellular response, have been the subject of debate (29). Furthermore, in some mouse models of infection, the animal does not develop a protective memory response (30).
The A. baumannii strain LAC-4 (31) was clinically isolated from a nosocomial outbreak in Los Angeles County hospitals in 1997 (32). In mouse models, LAC-4 intranasal infections reliably reproduce the most relevant features of human pulmonary A. baumannii infection and the corresponding pathology. Within 24 hours post-infection, LAC-4 displays rapid bacterial replication in the lungs, significant extrapulmonary dissemination to the spleen and severe bacteremia (33). Intranasal immunization of wild-type mice with formalin-killed whole LAC-4 bacteria has been shown to induce the development of short-term immunity via training of alveolar macrophages (34) and long-term immunity against intranasal challenge with live LAC-4 (35).
In the present study, we attempted to determine whether a single intranasal vaccination with the heat killed LAC-4 strain protects mice genetically deficient for key elements of the immune response as well as mice treated with cyclophosphamide, an immunosuppressive drug frequently used in clinical settings to treat cancers and autoimmune diseases (36) and described to greatly increase susceptibility to infections by A. baumannii in mice (37). We also attempted to determine whether killed LAC-4 can induce activation of human dendritic cells and T lymphocytes.
2 Results
2.1 Immunization with a killed strain of AB5075 or LAC-4 induces protection against challenge with a live strain of A. baumannii
The LAC-4 (33) and AB5075 (38) strains of A. baumannii have previously been reported as highly virulent in mouse models and reproducing the main features of the pathology of a human pulmonary infection by A. baumannii. Interestingly, these strains are not closely related. They differ substantially in allelic variants of housekeeping genes and do not qualify as single- or double-locus variants of each other under either the Pasteur or Oxford schemes. LAC-4 is classified as Sequence Type (ST)10 and has not been assigned to any Global Clone (31). LAC-4 is sometimes referred to as Clonal Complex (CC)10, whereas AB5075 is classified as ST1 and belongs to Clonal Group I (39).
In our C57BL/6 intranasal infection model, we found that AB5075 exhibits lower virulence than LAC-4. Bacterial loads in the lungs at 24 hours post-infection were higher for LAC-4 (Supplementary Figure S1A), and this strain induced significant mortality at a dose of 2×107 CFU, whereas all mice survived the same dose of AB5075 (Supplementary Figure S2B). Therefore, we choose to use strain LAC-4 to test the acquisition of protective memory against A. baumannii infection.
A vaccine composed of 5x105 or 5x107 CFU of formalin-killed A. baumannii LAC-4 administered intranasally has been shown (35) to fully protect wild-type C57BL/6 mice against a challenge with 7x107 CFU of live LAC-4 that killed 80% of control unvaccinated mice. We choose to use a similar experimental model with a vaccine composed of heat-killed (HK) A. baumannii and tested whether intranasal or intraperitoneal administration of HK A. baumannii LAC-4 or AB5075 strains could induce long-lasting immunity against a semi lethal challenge with the live LAC-4 strain.
Wild-type C57BL/6 mice were inoculated intranasally or intraperitoneally with a solution of PBS (control unvaccinated group) or 108 CFU of HK AB5075 or HK LAC-4 in PBS. Six weeks later, 2x107 CFU of live LAC-4 was administered intranasally to all groups. Survival was evaluated in each group of mice up to 5 days post-challenge (Figure 1A). The bacteria count in the lungs (Figure 1B) and spleen (Figure 1C) of the mice was measured by counting the CFUs at 24 hours post-challenge.
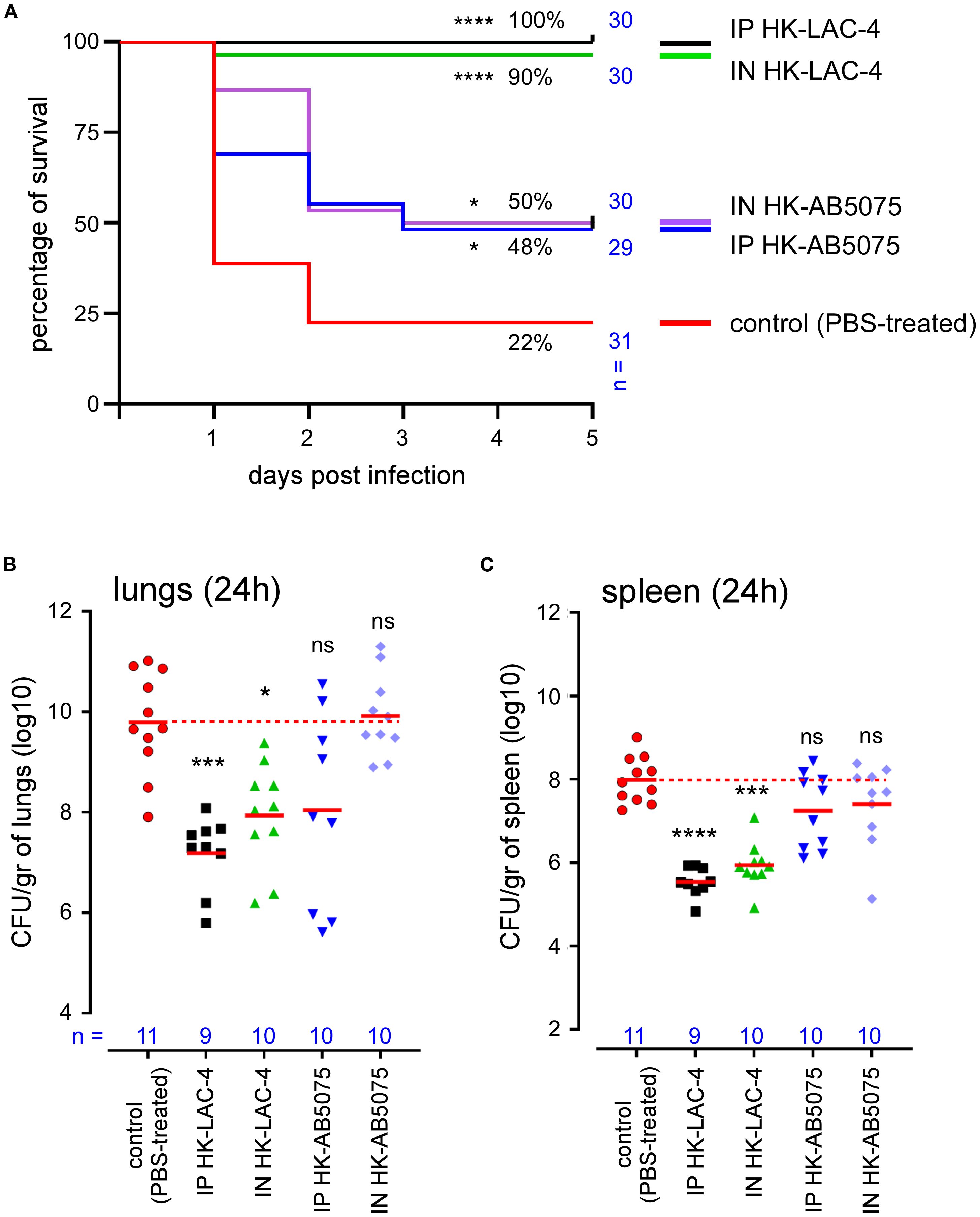
Figure 1. Intranasal immunization with heat-killed LAC-4 or AB5075 A. baumannii strains confer significant protection against challenge with live LAC-4 strain. Wild-type C57BL/6 mice were inoculated intranasally (IN) or intraperitoneally (IP) with PBS (control) or 108 CFU of heat-killed (HK) LAC-4 or HK 5075 A. baumannii strains in PBS. 6 weeks later, control and vaccinated mice were challenged intranasally with 2x107 CFU of live LAC-4 strain. (A) Survival rate. Over the 5 days after challenge, the fitness of infected mice was monitored. When the human endpoint was reached, the mice were euthanized. These data represent a pool of two independent experiments. n = number of mice per group. (B, C) CFU counts. 24 hours after challenge, some mice were sacrificed and the number of bacteria per gram in the lungs (B) and spleen (C) was evaluated by CFU counting. Red lines represent the geometric mean for each group of mice. Dotted red line represents the mean value of the control (unvaccinated) group. Significant differences between control (unvaccinated) and vaccinated groups are marked with asterisks: *p < 0.05, ***p < 0.001, ****p < 0.0001, in a log-rank (Mantel–Cox) test for survival curve and a (Wilcoxon-) Mann-Whitney post-test for CFU count.
We observed that all groups immunized with HK LAC-4 or HK AB5075 showed significantly improved survival compared to the unvaccinated controls (Figure 1A). As expected, immunization with LAC-4 provided stronger protection against LAC-4 infection than AB5075 immunization. Nevertheless, AB5075 immunization still conferred partial but statistically significant protection against LAC-4, indicating that vaccination with one strain of A. baumannii can induce cross-protection against genetically distant strains. Survival of HK LAC-4 groups correlated with bacterial control in both the lungs (Figure 1B) and spleen (Figure 1C). Interestingly, this is not the case with the protection conferred by HK AB5075, suggesting that survival is driven more by the nature of the inflammatory response than by the absolute bacterial burden in the organs.
2.2 Intranasal immunization with heat-killed LAC-4 induces cross protective A. baumannii antibodies and robust production of proinflammatory cytokines
Since protection against A. baumannii in mice vaccinated with formalin-killed LAC-4 was attributed to the presence of specific antibodies (35), we measured the presence of antibodies specific for LAC-4 and AB5075 in the serum of mice vaccinated intraperitoneally or intranasally with LAC-4 or AB5075. Our results show that immunization via the intraperitoneal route induces higher levels of specific IgM, IgG1 and IgG3 antibodies than immunization via the intranasal route (Figure 2). As expected, immunization induces a higher level of antibodies against the vaccination strain compared to the other strain. Nevertheless, immunization with LAC-4 induces a low but statistically significant level of antibodies able to recognize AB5075 and vice versa, which may explain the moderate cross protection we observed following a challenge with LAC-4.
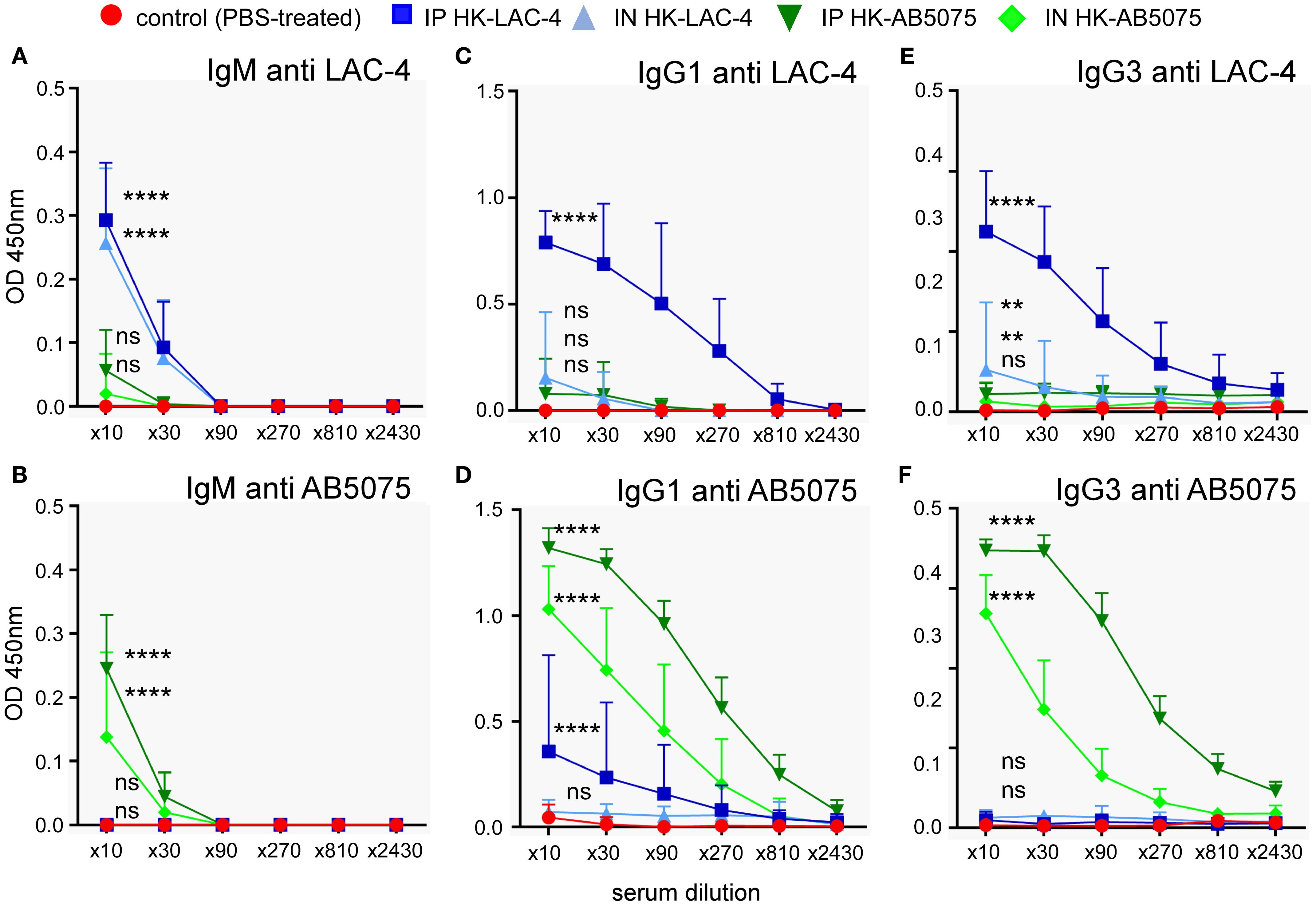
Figure 2. Humoral immune response induced by intranasal immunization with heat-killed A. baumannii. Wild-type C57BL/6 mice were inoculated intranasally with 108 CFU of heat-killed (HK) strains of Acinetobacter baumannii (LAC-4 in blue or AB5075 in green), as indicated. Serum was collected at 4 weeks post-vaccination, and ELISA was performed to determine the levels of IgM, IgG1 and IgG3 antibodies specific to LAC-4 (A, C, E) and AB5075 (B, D, F). The data represent the means ± SD of 8 mice per group. O.D., optical density. Significant differences between control (unvaccinated) and vaccinated groups are marked with asterisks: **p < 0.01, ****p < 0.0001, in a two-way Anova with Dunnett’s multiple comparisons test.
To better understand the nature of the protection conferred by the HK LAC-4 vaccine, we compared the production of proinflammatory cytokines induced by an intranasal infection with 2x106 CFU of live LAC-4 and the intranasal administration of 108 CFU of HK LAC-4. We administered a lower dose of live LAC-4 than dead LAC-4 because mice could not survive 24 hours after injection of 108 CFU of live LAC-4. The levels of various cytokines were measured at 4 and 24 hours post-intranasal inoculation in the lungs and the spleen by q-PCR reaction (Figure 3). We observed that HK LAC-4 induces innate proinflammatory cytokines e.g. IL-1, IL-6, TNF-α, as well as cytokines associated with both Th1 (IFN-γ) and Th17 (IL-17A, IL-17F and IL-22) responses. However, compared with live LAC-4, HK LAC-4 induces less IFN-γ but more IL-17F and IL-22, suggesting that HK LAC-4 would be particularly effective in inducing Th17 immunity.
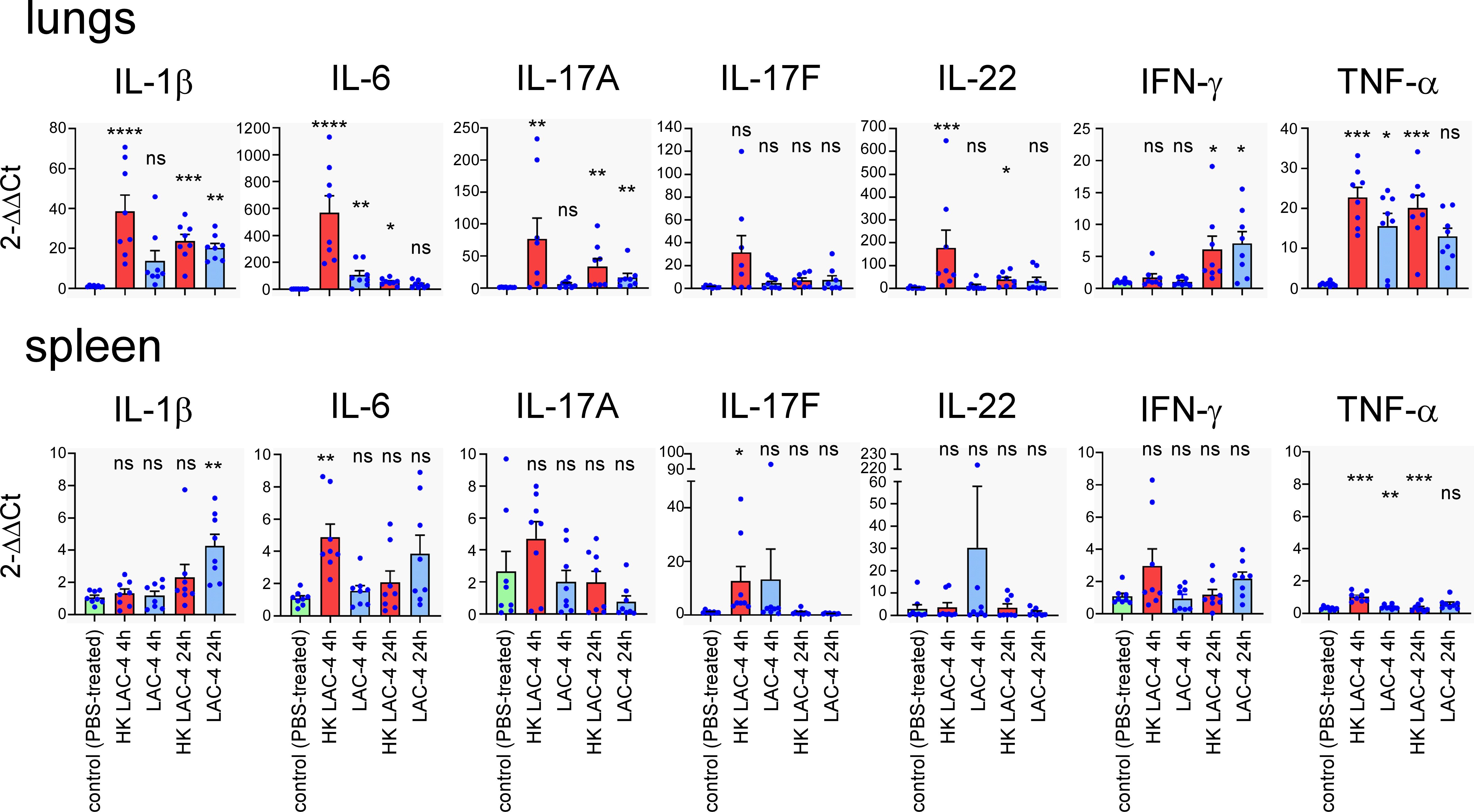
Figure 3. Intranasal administration of heat-killed A. baumannii LAC-4 strain induces the production of proinflammatory cytokines in the lungs and spleen. Wild-type C57BL/6 mice were inoculated intranasally with PBS (control), 108 CFU of heat-killed (HK) or 2x106 CFU of live A. baumannii LAC-4 strain in PBS, as indicated. At 4 and 24 hours post-infection, mice were sacrificed, lungs and spleen were collected. Expression levels of the indicated genes were assessed by quantitative RT-PCR and expressed as relative expression to RPL32 housekeeping mRNA. The naïve mice (number 1) condition was set to 1. The experiment was performed twice independently with 4 mice per group. The figure represents a pool of data from the 2 experiments, n=8 mice for each condition. Significant differences between control (unvaccinated) and vaccinated groups are marked with asterisks: *p < 0.05, **p < 0.01, **p < 0.001, ***p < 0.001, in a (Wilcoxon-) Mann-Whitney post-test.
2.3 Mice rendered genetically deficient in IL-17RA dependent response or γδT cells are highly susceptible to pulmonary A. baumannii infection
To determine which elements of the adaptive immune response are essential for control of an intranasal infection, we studied the survival of different strains of mice deficient for key elements of the immune system following infection with LAC-4.
Wild-type mice as well as mice rendered genetically deficient for the Th1 response (IL-12p35-/-, IFN-γ-/-), for the Th17 signaling (IL-17RA-/-) and for selected lymphocyte populations (δTCR-/-, TAP-1-/-, MHCII-/-, CD3-/-, MuMT-/-) were infected intranasally with 2x106 CFU of live LAC-4. Survival was measured in each group for 120 hours post-infection (Figure 4A). We observed that the absence of IL-17RA and δTCR has dramatic consequences. Almost all mice died within 24 hours in these groups (0 and 6% survival in the 17RA- and δTCR-KO groups, respectively). The levels of IL-17A, IL-17F, IL-22 and TNF-α were measured at 4 and 24 hours post-intranasal inoculation in the lungs of wild-type and δTCR-/- mice by q-PCR reaction (Supplementary Figure S2). We observed that mice deficient for δTCR do not produce fewer mRNA for these cytokines than wild-type mice, which suggests that the absence of γδ+T cells does not impair the production of these cytokines. A deficiency in TAP1 also induces significant early mortality with only 22% survival. CD3 deficiency (which affects all T cells) is associated with 46% survival of mice, while MHCII deficiency [which affects CD4+ T cells (40)] and MuMT [which affects B cells (41)] allows 76% and 80% survival, respectively. A statistical analysis of these results shows that the absence of CD3 significantly reduces the survival of the animals, whereas the deficiency of MHC-II or MuMT has no significant effects on it.
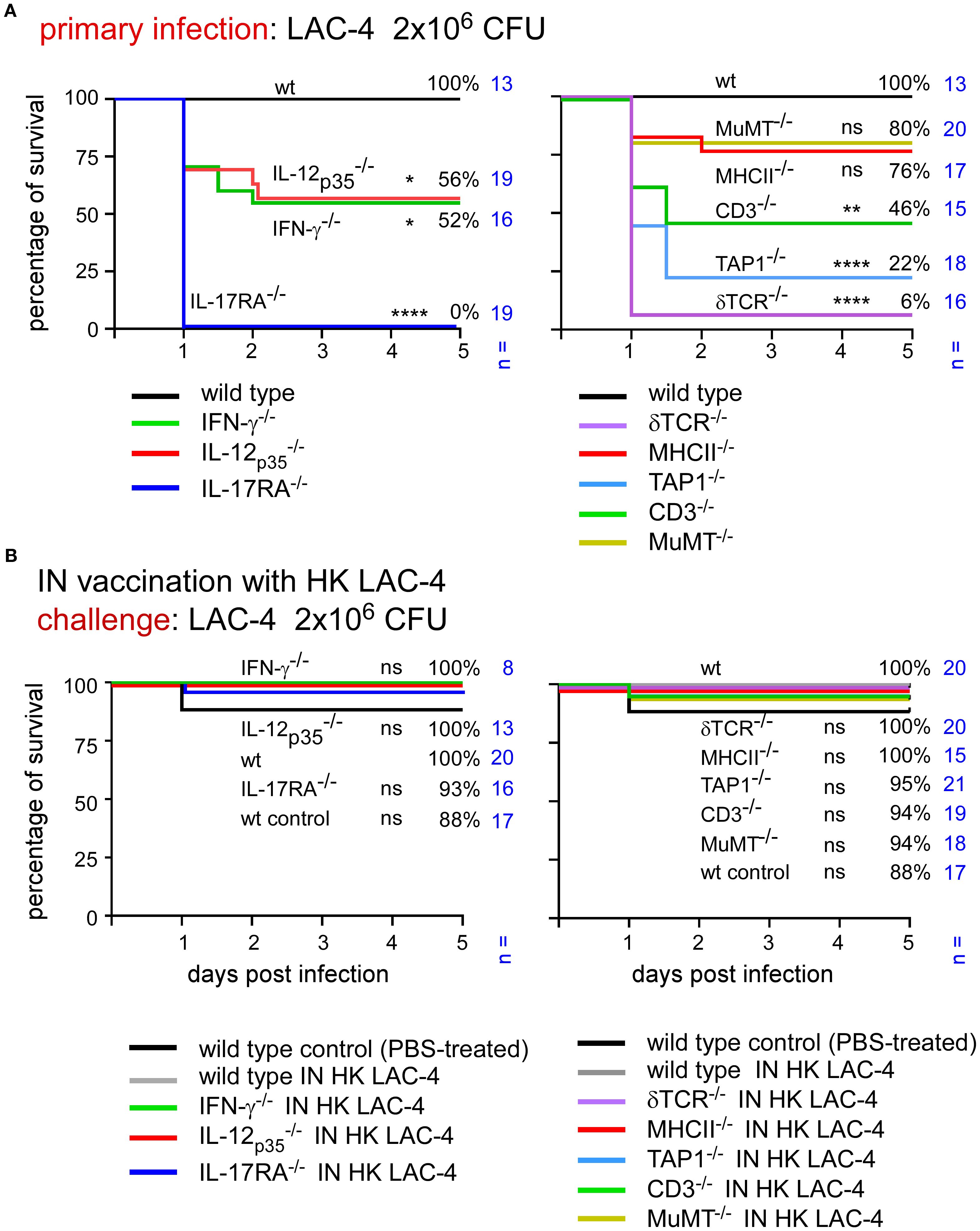
Figure 4. Immunization with HK LAC-4 protects wild-type mice and mice deficient for key elements of adaptive immune response against LAC-4 challenge. The figures present survival rates of wild-type and genetically deficient C57BL/6 mice measured over a period of 5 days after intranasal infection with A. baumannii LAC-4. When the human endpoint was reached, the mice were euthanized. (A) Mice were infected with 2x106 CFU of LAC-4. (B) Mice were treated intranasally with PBS (control) or 108 CFU of heat-killed (HK) LAC-4 in PBS. 6 weeks later, control and vaccinated mice were challenged intranasally with 2x106 CFU of live LAC-4. The data represent the pool of a minimum of two distinct experiments. n indicates the number of mice per group. Significant differences between wild-type mice (A) or vaccinated wild-type (B) and other groups are marked with asterisks: *p < 0.05, **p < 0.01, ****p < 0.0001, in a log-rank (Mantel–Cox) test.
Surprisingly, the CFU counts observed in the lungs and spleen of infected mice weakly correlated with survival of the mouse (Supplementary Figure S3). All groups of deficient mice had a higher level of bacteria in the lungs and spleen than wild-type mice and the differences were quite small, particularly in spleen, between the deficient mice. These results suggest that it is the nature of the inflammatory immune response rather than the number of bacteria in organs that determines the animal’s survival.
Taken together, these results suggest that γδ T cells, TAP-1 dependent lymphocyte populations and IL-17RA-dependent signaling play a very important role in the control of primary intranasal LAC-4 infection while the role of IL-12 and the IFN-γ dependent Th1 response as well as CD4+ T cells and B cells is more moderate.
2.4 Intranasal immunization with heat-killed LAC-4 induces long-term immune protection in a large panel of mice genetically deficient for key elements of the adaptive immune response
After identifying the genetic deficiencies leading to increased susceptibility to primary infection, we tested the capacity of intranasal immunization with HK LAC-4 to increase the survival of genetically immunodeficient mice against pulmonary LAC-4 infection.
Wild-type mice were inoculated intranasally with PBS (control, unvaccinated group) or 108 CFU of HK LAC-4 in PBS. The survival of these mice was compared with that of various genetically immunodeficient mice vaccinated with HK LAC-4 and subjected to an intranasal challenge with a dose of 2x106 CFU of live LAC-4 (Figure 4B). We observed that the survival rates in all groups of vaccinated immunodeficient mice were high (88-100%). This suggests that vaccination compensates for the absence of the Th1 or Th17 response as well as the TAP-1 and CD3 deficiencies.
To identify the key elements of protective memory immunity induced by HK LAC-4, we challenged vaccinated mice with a higher dose of live LAC-4, 2x107 CFU. At this dose, most unvaccinated mice, whether wild-type or genetically immunodeficient, succumb to infection within 48 hours (Figure 5A). Interestingly, the vaccine does not provide equivalent protection against a challenge with 2x107 CFU of live LAC-4 in all groups of immunodeficient mice (Figure 5B). MuMT-/- mice showed the lowest survival rate, at 5%, thus confirming the key role of antibodies in protective memory against A. baumannii. Vaccinated TAP1-/-, δTCR-/- and IL-17A-/-mice also show statistically significant reduction of survival, at 59%, 57% and 43% respectively, compared to vaccinated wild-type mice (94%). These findings demonstrate that vaccination cannot completely compensate for the absence of cell populations that depend on TAP1 and δTCR or of the IL-17RA-dependent Th17 response. In contrast, vaccine protection can be induced in the absence of CD3 or MHCII-dependent T cells as well as the IL-12p35 or IFN-γ dependent Th1 response.
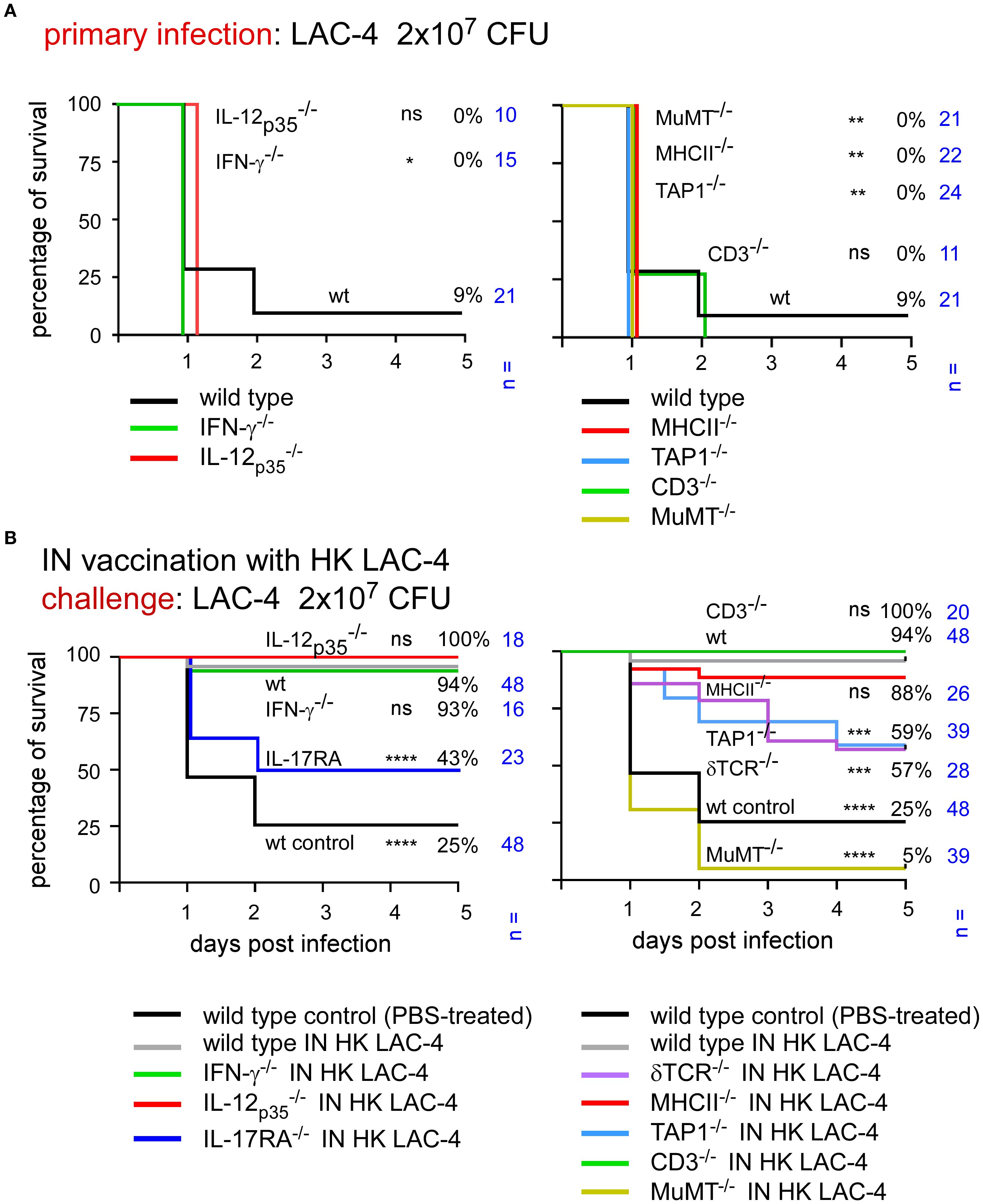
Figure 5. Immunization with HK LAC-4 fails to protect some deficient mice against high-dose LAC-4 challenge. The figures present the survival rates of wild-type and genetically deficient C57BL/6 mice measured over a period of 5 days after intranasal infection with A. baumannii LAC-4. When the human endpoint was reached, the mice were euthanized. (A) Mice were infected with 2x107 CFU of LAC-4. (B) Mice were treated intranasally with PBS (control) or 108 CFU of heat-killed (HK) LAC-4 in PBS. 6 weeks later, control and vaccinated mice were challenged intranasally with 2x107 CFU of live LAC-4. The data represent the pool of a minimum of two distinct experiments. n indicates the number of mice per group. Significant differences between wild-type mice (A) or vaccinated wild-type (B) and other groups are marked with asterisks: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001, in a log-rank (Mantel–Cox) test.
2.5 The bacterial capsule is essential for the induction of protective memory
Since the humoral response is a key element of protection against A. baumannii infection induced by HK LAC-4 in our experimental model, we attempted to identify the main antigenic targets of the protective antibodies. To this end, we produced a LAC-4 strain deficient for the itrA gene (as described in the Materials and Methods section), which is essential for bacterial capsule synthesis (42), and compared the vaccine capacity of this ΔitrA strain to that of the wild-type strain.
Wild-type C57BL/6 mice were vaccinated intranasally with 108 CFU of HK wild-type or ΔitrA LAC-4 strains. At 6 weeks post-vaccination, mice were infected with 2x107 CFU of live wild-type LAC-4 strain. As expected, most mice vaccinated with wild-type HK LAC-4 survived (89%) (Figure 6A). In striking contrast, mice vaccinated with HK ΔitrA LAC-4 had a very low survival rate (20%), close to that of unvaccinated mice (6%). Measurement by ELISA of the levels of antibodies specific to wild-type LAC-4 in the blood of vaccinated animals showed that mice vaccinated with HK ΔitrA did not have IgM, IgG1 and IgG3 antibodies recognizing wild-type LAC-4 (Figure 6B).

Figure 6. Vaccine produced with capsule-deficient bacteria fails to induce protection in mice. C57BL/6 mice were treated intranasally (IN) with PBS (control) or 108 CFU of heat-killed (HK) of wild-type LAC-4 or ΔitrA LAC-4 in PBS. (A) At 6 weeks post-vaccination, control and HK vaccinated mice were challenged intranasally with 2x107 CFU of live wild-type LAC-4 strain. The figure presents the survival rates of each group of mice measured over a period of 5 days after intranasal infection with A. baumannii LAC-4. When the human endpoint was reached, the mice were euthanized. The data represent the pool of two distinct experiments. n indicates the number of mice per group. (B) At 4 weeks post-vaccination, serum was collected, and ELISA was performed to determine the levels of IgM, IgG1 and IgG3 antibodies specific to LAC-4, as indicated. The data represent the means ± SD of 8 mice per group. O.D., optical density. (C) Antibodies produced by vaccinated mice were tested against bacterial samples of capsulated wild-type and non-capsulated ΔitrA LAC-4 strains. Pool of serums from unvaccinated mice was used for background control, while Coomassie Blue stain was used as loading control and to visualize proteinaceous content. Pool of serums from wild-type LAC-4 vaccinated mice targeted high molecular weight antigens in the capsulated strains only, as well as conserved lower molecular weight proteins in both strains. These results are representative of 3 independent experiments. Significant differences between groups (A) are marked with asterisks: *p < 0.05, ***p < 0.001, ****p < 0.0001, in a log-rank (Mantel–Cox) test.
To better identify the targets of protective antibodies against LAC-4 infection, we conducted Western blot analyses (Figure 6C) comparing antigens recognized by sera from mice vaccinated with PBS (control), wild-type, or capsule-deficient ΔitrA LAC-4 strains. Extracts from wild-type LAC-4 and ΔitrA LAC-4 were tested. Sera from mice immunized with wild-type LAC-4 recognized a large count of high–molecular-weight antigens in extract from wild-type LAC-4, whereas no corresponding signal was detected in capsule-deficient ΔitrA LAC-4 extract. This intense signal is not revealed by Coomassie blue staining. We also detected with serum from mice vaccinated with wild-type LAC-4 strains the presence of low molecular weight antigens in both extracts that are visible in Coomassie blue. Overall, these results demonstrate that protective antibodies induced by vaccination with HK wild-type LAC-4 are directed against non-proteinaceous capsule antigens and against some low molecular weight protein antigens.
2.6 Intranasal immunization with heat-killed LAC-4 partially protects cyclophosphamide-treated mice against challenge with A. baumannii
Since hospitalized patients at risk for A. baumannii infection are often immunocompromised (9), we tested the ability of intranasal vaccination with HK LAC-4 to protect cyclophosphamide-treated mice against intranasal infection with LAC-4. Wild-type C57BL/6 mice were inoculated intranasally with PBS (control) or 108 CFU of HK LAC-4. Five weeks later, half of the mice in each group were inoculated intraperitoneally with PBS or underwent cyclophosphamide treatment, as described in the Materials and Methods. One week post- cyclophosphamide treatment (6 weeks post-vaccination), all groups of mice received 2×106 CFU of live LAC-4 intranasally. Survival of the mice was measured up to 120 hours post-challenge.
As shown in Figure 7, in agreement with a previous study (37), cyclophosphamide treatment greatly reduced the survival of infected wild-type C57BL/6 mice. We observed that intraperitoneal immunization with HK LAC-4 significantly reduced mortality in mice treated with cyclophosphamide. Vaccine protection, however, was not complete in the cyclophosphamide-treated mice and the protection provided did not exceed 65%.
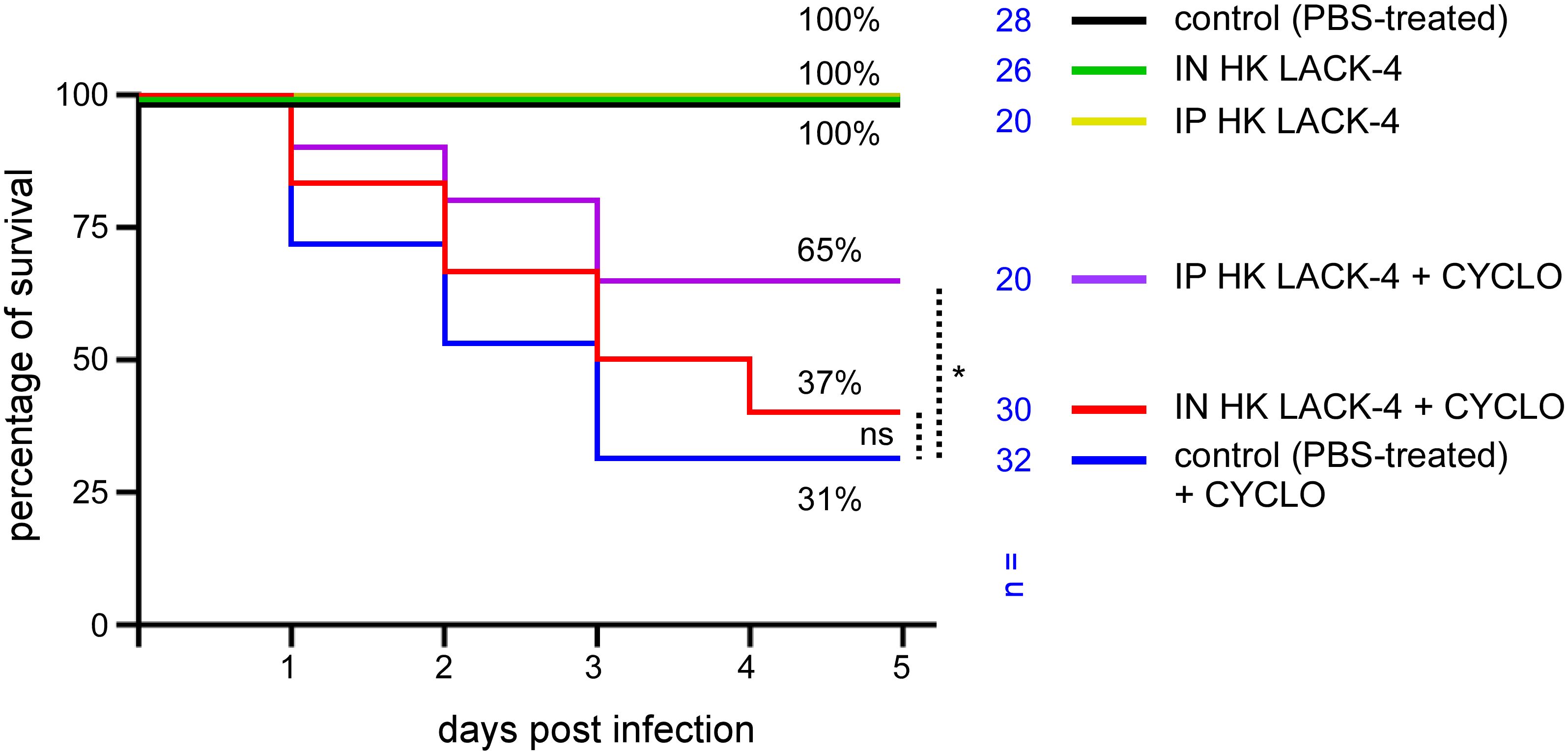
Figure 7. Immunization with HK LAC-4 confers partial protection against LAC-4 challenge to cyclophosphamide-treated wild-type C57BL/6 mice. Wild-type C57BL/6 mice were treated intranasally (IN) with PBS (control) or 108 CFU of heat-killed (HK) LAC-4 A. baumannii in PBS. 6 weeks later, control and HK vaccinated mice were challenged intranasally with 2x106 CFU of live LAC-4 strain. Cyclophosphamide (cyclo) was administrated intraperitoneally at 4 days (150 mg/kg) and 1 day (100 mg/kg) before the challenge. Picture shows the percent survival rate. Over the 5 days after challenge, the fitness of the infected mice was monitored. When the human endpoint was reached, the mice were euthanized. These data represent a pool of three independent experiments. n= number of mice for each condition. Significant differences between control (unvaccinated) + CYCLO and vaccinated groups + CYCLO are marked with asterisks: *p < 0.05, in a (Wilcoxon-) Mann-Whitney post-test.
2.7 Heat-killed LAC-4 activates human peripheral blood mononuclear cells
To evaluate the ability of HK LAC-4 to induce an adaptive immune response in humans, we measured in vitro the activation of dendritic cells and T lymphocytes from Peripheral Blood Mononuclear Cells (PBMC) of healthy donors in response to HK LAC-4.
Monocytes isolated from human PBMC were differentiated for 5 days to dendritic cells in the presence of IL-4+GM-CSF and then stimulated for 48 hours with 2x106 CFU/ml of HK A. baumannii LAC-4, HK Brucella melitensis, HK Escherichia coli or 1 µg/ml of LPS from E. coli. B. melitensis is a stealthy bacterium that expresses an atypical and poorly activating LPS (43). HK B. melitensis was used as a negative control in our experiments. We used HK E. coli and commercial LPS as positive controls. The expression of dendritic cell surface maturation markers was analyzed by flow cytometry. We observed that HK LAC-4 significantly up-regulated CD40, CD80, CD83 and CD86 maturation markers and down-regulated CD209 (a lectin receptor mostly expressed by immature DCs) (Figure 8A). Moreover, HK LAC-4 induced significant secretion of IL-23 and IL-12p70 in our experimental setting (Figure 8B).
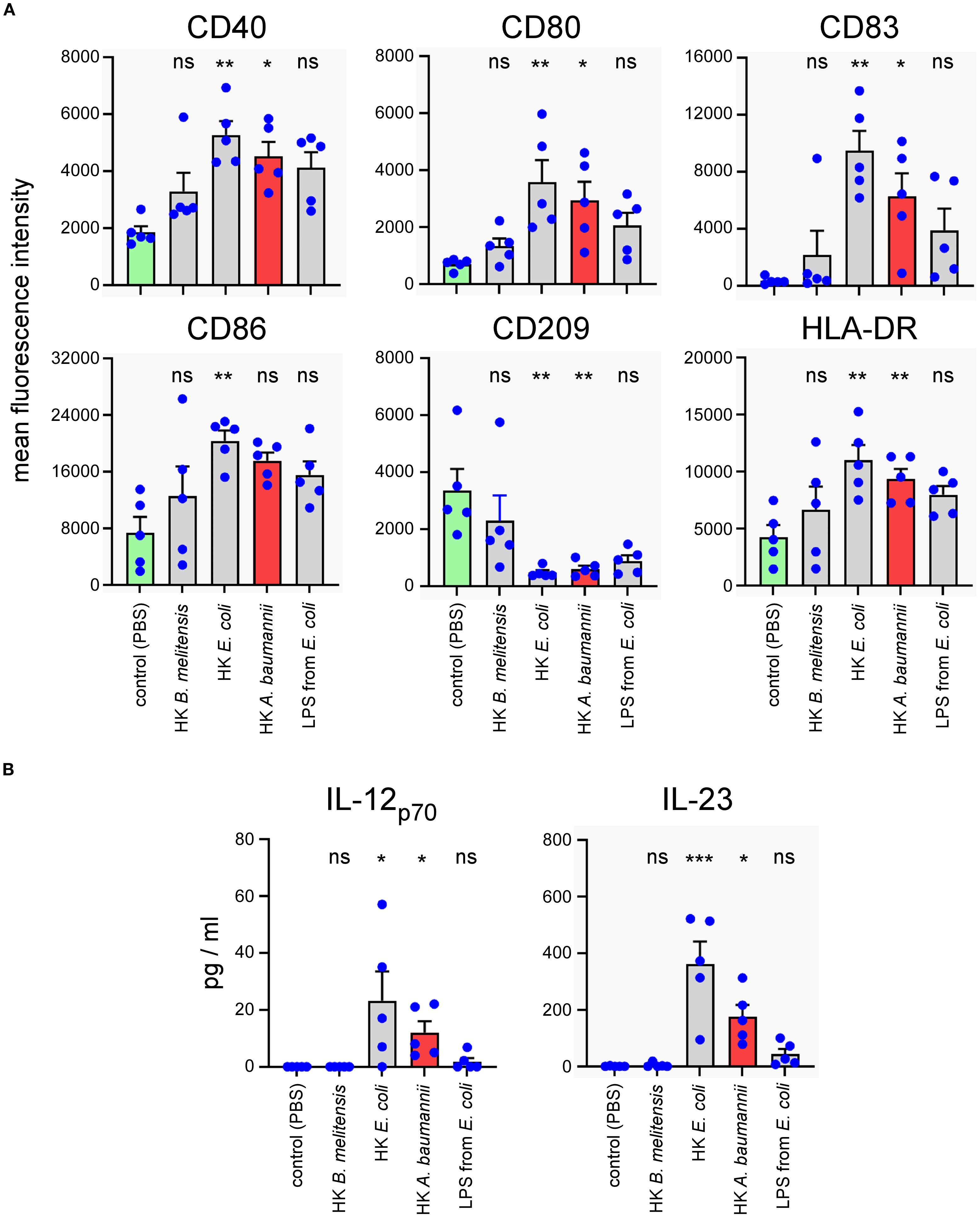
Figure 8. In vitro stimulation with heat-killed A. baumannii LAC-4 induces maturation of human monocyte-derived dendritic cells and the production of IL-23. Monocytes isolated from human PBMC were differentiated for 5 days to dendritic cells with IL-4+GM-CSF and then stimulated with PBS (negative control), heat-killed (HK) LAC-4 A. baumannii, HK B. melitensis, HK E. coli and LPS from E. coli for 48 hours. (A) Six maturation markers were analyzed by flow cytometry and are expressed here as the medium fluorescence intensity (MFI) reflecting the change in expression of the marker on the cell. (B) Supernatant from stimulated monocyte-derived dendritic cells was analyzed by ELISA to quantify the cytokines IL-12p70 and IL23. Columns represent the mean of five different donors +/- SEM. The stimulated groups are compared to the PBS-treated control group:*p<0.05; **p<0.01; ***p<0.001; determined by the non-parametric 1-way ANOVA with a multiple comparison.
To evaluate the capacity of HK LAC-4 to activate human T lymphocytes, total PBMC were stimulated for 7 days with 2x106 CFU/ml of HK A. baumannii LAC-4, HK B. melitensis or HK E. coli. We used 2 µg/ml CEF+CEFTA (pool of peptides-MHC class I and II-restricted T-cell epitopes from human Cytomegalovirus, Epstein Barr virus, Influenza virus, Tetanus toxin and Adenovirus 5) and 1 µg/ml of SEB (Staphylococcal enterotoxin B) as the positive control for CD4+ and CD8+ T cell proliferation and 10 µg/ml HMBPP (4-hydroxy-3-methyl-but-2-enyl pyrophosphate) as the positive control for γδ+ T cell proliferation. We observed that HK LAC-4 induced significant proliferation of CD4+ T cells, CD8+ T cells and γδ+ T cells (Figure 9A) as well as easily detectable secretion of IFN-γ, IL-17A and TNF-α (Figure 9B). To determine the cell source of these cytokines, we analyzed stimulated total PBMC by intracellular flow cytometry. Our results showed (Figure 10) that HK LAC-4 induced significant production of IFN-γ by CD8+ and γδ+ T cells, of IL-17A by CD4+, CD8+ and γδ + T cells and of TNF-α by CD4+ and γδ+ T cells.
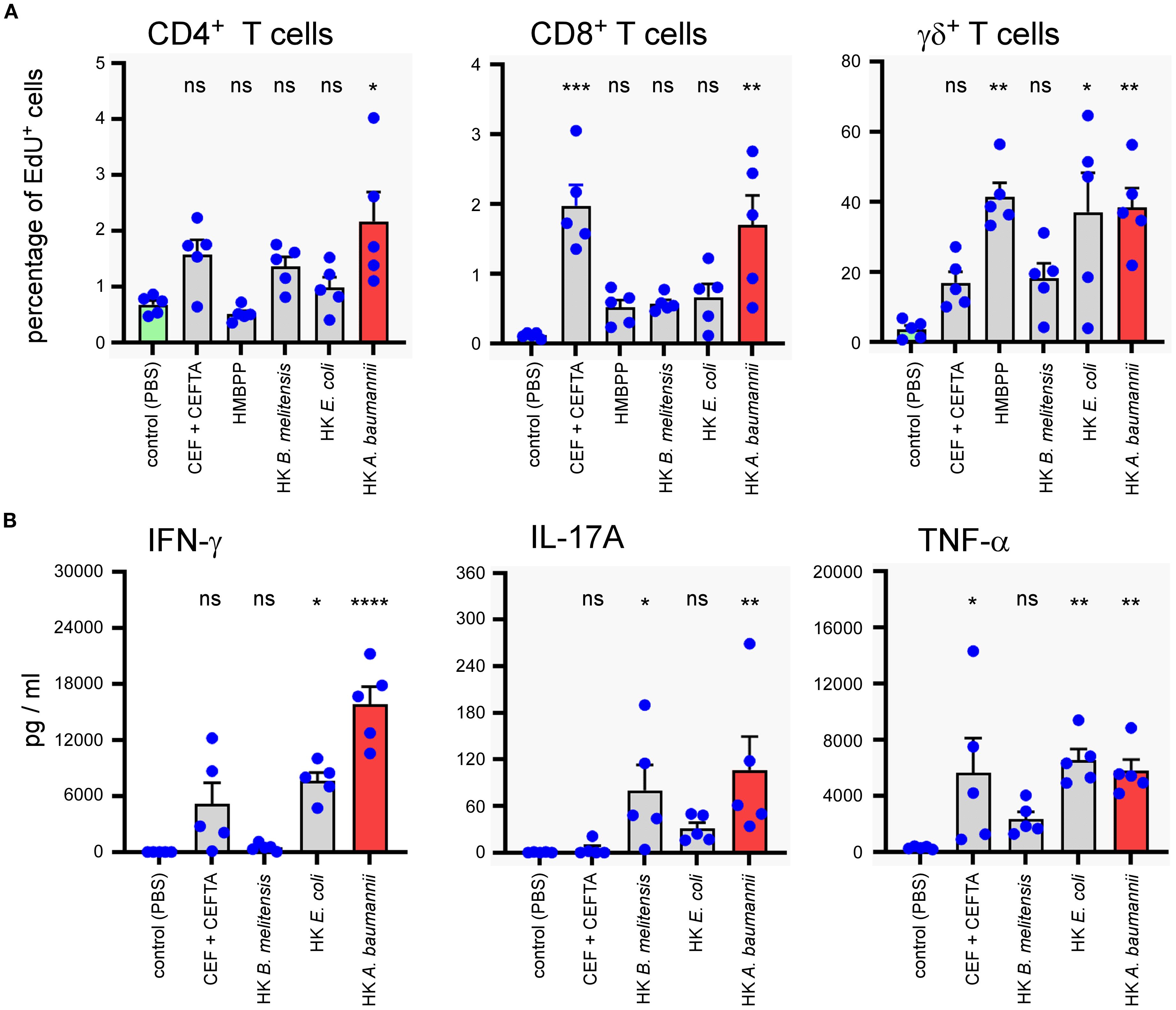
Figure 9. In vitro stimulation of human PBMC with heat-killed LAC-4 A. baumannii induces proliferation of T cells and production of IFN-γ, IL-17A and TNF-α. Total PBMC were stimulated for 7 days with PBS (negative control), CEF+CEFTA (pool of peptides-MHC class I and II-restricted T-cell epitopes) as the positive control for CD4 and CD8 T cell proliferation, HMBPP (4-hydroxy-3-methyl-but-2-enyl pyrophosphate) as the positive control for γδ T cell proliferation, heat-killed (HK) LAC-4 A. baumannii, HK B. melitensis and HK E. coli. (A) Proliferation of T cells is expressed by the percentage of cells incorporating EdU (5-ethynyl-2´-deoxyuridine). (B) Supernatant from stimulated PBMC was analyzed by ELISA for the quantification of IL-17A and with Luminex for IFN-γ and TNFα. Quantities are expressed in pg/ml. Columns represent the mean of five different donors +/- SEM. The stimulated groups are compared to the PBS-treated control group:*p<0.05; **p<0.01; ***p<0.001; determined by the non-parametric 1-way ANOVA with a multiple comparison.
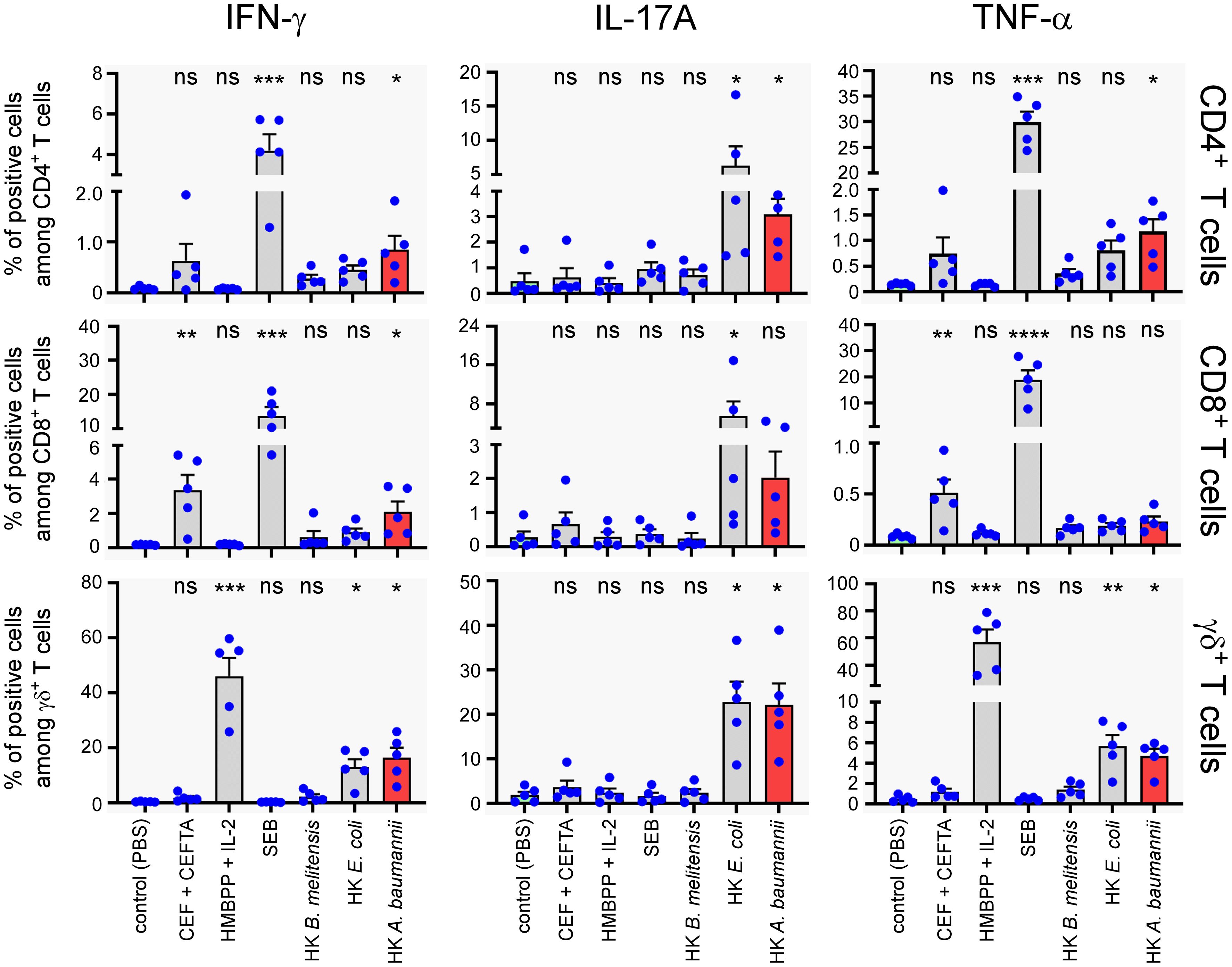
Figure 10. In vitro stimulation of human PBMC with heat-killed LAC-4 A. baumannii of human PBMC induces production of IFN-γ, IL-17A and TNF-α from three distinct T cell populations. Total PBMC were stimulated for 7 days with PBS (negative control), CEF+CEFTA (pool of peptides-MHC class I and II-restricted T-cell epitopes), HMBPP (4-hydroxy-3-methyl-but-2-enyl pyrophosphate) + IL-2, SEB, heat-killed (HK) LAC-4 A. baumannii, HK B. melitensis or HK E. coli. Brefeldin-A and Monensin were added for the last 5 hours before intracellular FACS staining. For stimulation with HMBPP + IL-2, the compounds were added for 5 hours in culture media treated cells, at the time of Brefeldin-A and Monensin blocking. Graphs represent the percentages of cells expressing IFN-γ, IL-17A or TNF-α in the three main T cell populations. Columns represent the mean of five different donors +/- SEM. The stimulated groups are compared to the PBS-treated control group: *p<0.05; **p<0.01; ***p<0.001; ****p<0.0001; ****p<0.0001 determined by the non-parametric 1-way ANOVA with a multiple comparison.
Overall, these results show that HK LAC-4 can induce in vitro dendritic cell maturation as well as activation of all T cell subsets, suggesting that it might be able to activate lymphocytes in patients.
3 Discussion
Given its growing significance as a public health threat, considerable efforts have been directed toward developing vaccines against A. baumannii. Numerous vaccine candidates have been evaluated in animal models, such as subunit (44, 45), DNA, Outer membrane vesicles, inactivated whole-cell and live attenuated vaccines (46). However, no vaccine candidates have advanced into clinical trials.
A. baumannii displays marked genetic and phenotypic diversity (8, 11) and causes opportunistic infections in individuals, some of whom experience varying degrees of disease- or treatment-related immunosuppression (3). These factors make it unlikely that subunit or live-attenuated vaccines would achieve widespread success in real-world settings. The plasticity of the A. baumannii genome means that an attenuated vaccine could recover its virulence and thus be fatal to immunocompromised individuals. This plasticity could also allow it to quickly escape the immunity conferred by a subunit vaccine which would be focused on a small number of antigenic determinants. Consequently, a whole inactivated vaccine could represent a promising solution by avoiding any risk of reversion of its virulence and providing immunization against many A. baumannii antigens.
Several whole inactivated vaccines, such as formalin-killed ATCC 19606 strain (47), formalin-killed LAC-4 strain (33) or γ-irradiated (48) and ultraviolet C–inactivated (49) AB5075 strain, have been shown to protect mice against intraperitoneal or intranasal A. baumannii infection. In this study, we test a vaccine composed of heat-inactivated LAC-4 bacteria. We chose this protocol of inactivation for its practicality, safety, and simplicity, so that it is easy to reproduce by all experimenters. Our choice was also informed by previous reports documenting that formalin treatment can alter protein presentation to T cells (50) and affect the immunogenicity of some vaccines, such as vaccine against pertussis toxin (51), poliovirus (52) and Pseudomonas aeruginosa and Staphylococcus aureus (53). Moreover, it has been showed in some models that heat-inactivated vaccines may elicit stronger immune responses than their formalin-inactivated counterparts (54).
In the present work, using heat-killed (HK) LAC-4 vaccine, we confirmed previous findings (35) demonstrating the protective efficacy of a formalin-inactivated LAC-4 strain against lethal lung infection with the hypervirulent LAC-4 strain in wild-type C57BL/6 mice. This protection was absent when using a vaccine prepared from the capsule-deficient LAC-4 ΔitrA strain, suggesting that the immune response targets mainly non-protein surface antigens. We observed that a HK vaccine composed of the AB5075 strain, which is genetically unrelated to LAC-4, could induce antibodies capable of recognizing LAC-4 antigens and a partial protection against LAC-4 infection. We also observed that the protection induced by the HK LAC-4 vaccine was critically dependent on B lymphocytes. Then, we further showed that a single dose of the LAC-4 vaccine can partially protect certain genetically immunodeficient mouse models susceptible to A. baumannii pulmonary infection, including IL-17RA-/-, γTCR-/-, TAP-1-/-, and CD3-/- mice, as well as wild-type mice rendered immunocompromised by cyclophosphamide treatment—an immunosuppressive drug widely used in patients with cancer or autoimmune diseases (36). Moreover, we demonstrated that the HK LAC-4 vaccine can activate human dendritic cells and T lymphocytes in vitro, suggesting its potential to stimulate adaptive immunity in patients. All of these results are summarized schematically in Figure 11.
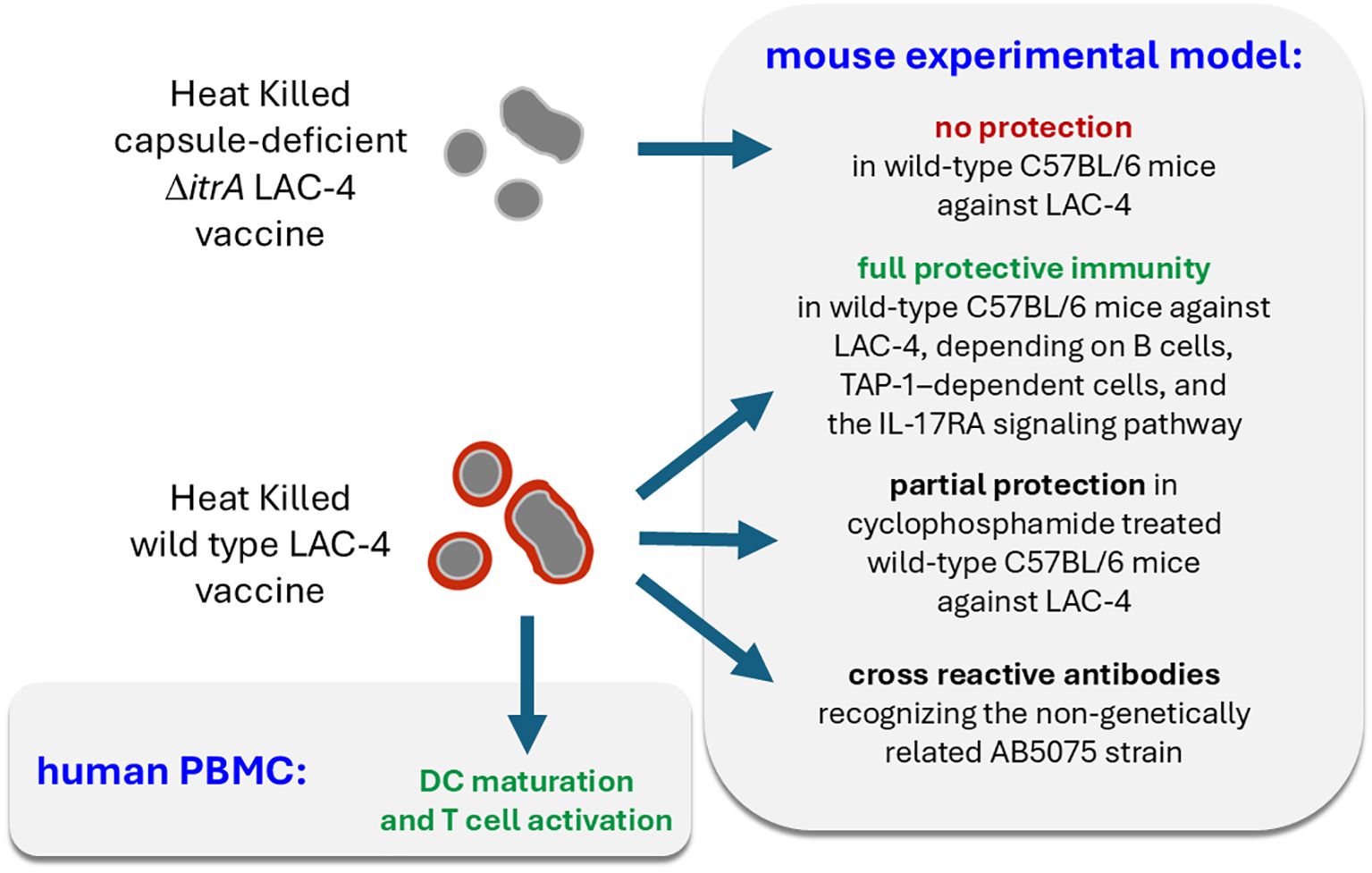
Figure 11. Schematic representation of the main results.
The precise mechanisms explaining the protective effect of the HK LAC-4 vaccine remain to be elucidated. Our vaccine fails to completely protect MuMT-/-, IL-17RA-/-, γTCR-/-, and TAP-1-/- mice against a high dose of LAC-4, suggesting that the IL-17RA-dependent signaling pathway and IgM, γTCR- and TAP-1-dependent cell populations are essential for optimal protection against A. baumannii.
The humoral response appears to be a key element of protection induced by the HK LAC-4 vaccine since MuMT deficiency in B cells completely abrogates protection in vaccinated mice. We have shown that HK LAC-4 vaccine induces IgM, IgG1 and IgG3 antibodies recognizing mainly the non-proteinaceous capsule antigens of the LAC-4 strain. A recent study (55) reported that a monoclonal antibody targeting cell-surface pseudaminic acid of A. baumannii exhibits direct bactericidal activity, supporting the possibility that antibodies directed against the capsule may confer protection. Interestingly, we have observed that antibodies induced by HK AB5075 not only recognize AB5075 but also cross-react with LAC-4. This cross-reaction confers significant protection as demonstrated by increased survival and reduced bacterial counts in the lungs and spleen of mice vaccinated with HK AB5075 and challenged with live LAC-4 compared to unvaccinated mice. These results suggests that a whole HK vaccine comprising a large panel of antigens could potentially confer protection against several different strains of A. baumannii. A promising vaccination strategy would be to create a multivalent inactivated vaccine combining several field strains among those most involved in human infections to ensure the broadest possible protection. However, the feasibility of this approach requires better identification of surface antigens that are shared between different strains and those that are not. Surface omics approaches (56), analyzing both proteins, sugars, and lipids exposed on the surface of A. baumannii strains, could help select the most interesting strains to combine in vaccines.
We attempted to protect naive wild-type mice using serum transfer from vaccinated mice. These transfers resulted in weak, statistically insignificant, and poorly reproducible protection in preliminary experiments. Therefore, we did not explore this path further. These results are similar to that obtained by KuoLee et al. (35), who demonstrated that the transfer of immune serum only prolongs survival by approximately 48 hours after infection with the LAC-4 strain. The difficulty in transferring protection via serum suggests that cellular components participate directly in protection. We observed that the cellular response induced by HK LAC-4 in lungs and spleen seems mainly directed towards the production of Th17 cytokines, such as IL-17A, IL-17F and IL-22, which are particularly important for inducing the production of antimicrobial peptides by the pulmonary mucosa (57).
Comparing the survival rates of different strains of mice genetically deficient for key elements of the adaptive response to LAC-4 infection opens up interesting new avenues of research. We observed that deficiencies in IL-17RA, δTCR and TAP-1 strongly reduce the survival of naïve mice and are associated with a loss of control of the multiplication of A. baumannii in the lungs, which has not been described previously to our knowledge. However, these results should be interpreted with caution. TAP-1 is involved in the association of cytosolic antigens with MHC-I and the absence of this molecule has been shown to prevent the development of CD8+ T cells (58). But its absence has also been found to have an effect on natural killer T cells (59) and on natural killers cells (60). Depletion of CD8+ T cells in C57BL/6 mice by administration of monoclonal antibody did not induce an increase of susceptibility to intranasal infection by A. baumannii (26). In contrast, depletion of NK1.1+ cells (a marker shared by NK and some NKT cells) in C57BL/6 mice increases mortality and reduces neutrophil recruitment during A. baumannii infection (61), suggesting that these cell types could participate in early defense against A. baumannii infection. IL-17RA recognizes IL-17A, IL-17E and IL-17F and has been implicated in the control of extracellular pathogens at the mucosal level (62), which could explain the high susceptibility of IL-17RA-/- mice observed in our infection model. As γδ T cells are known to produce IL-17A (63), we could hypothesize that a deficiency in γδ T cells results in lower production of IL-17A in mice infected with LAC-4. However, our data show that δTCR-/- mice produce as much IL-17A, IL-17F, IL-22 and TNF-α as wild-type mice. Furthermore, IL-17-/- C7BL/6 mice have been reported to be resistant to pulmonary LAC-4 infection (64). Thus, further investigation will be required to formulate definitive hypotheses explaining the susceptibility of IL-17RA signaling in the control of A. baumannii infection. This constitutes a priority for future research.
Results demonstrating partial protection in animals genetically deficient for key components of the immune response or animals treated with cyclophosphamide are particularly encouraging as immunocompromised patients are frequently infected by A. baumannii (9) and constitute an at-risk population during all epidemics. These populations not only suffer the most serious infections but are also poorly receptive to most vaccines and can act as reservoirs for pathogens, providing them opportunities to adapt to the host immune defense and to antibiotic treatments. For example, it is well known that immunocompromised patients are less well protected against SARS-CoV-2 by vaccines (65, 66). In addition, SARS-CoV-2 variants able to escape antiviral drugs (67) and adaptive immunity (68–70) are suspected to have emerged following chronic infection of immunocompromised individuals. This escape phenomenon is not limited to SARS-CoV-2, and has been documented with other viruses (71, 72) as well as with bacteria (73). It is therefore particularly important, both in the interest of patients and the wider population, to successfully develop vaccines that effectively protect immunocompromised individuals. Our study demonstrates that this is possible in the experimental model of pulmonary infection with A. baumannii.
A previous study by Dolery et al. (48) reported stronger protection than we observed in mice vaccinated with a gamma-irradiated A. baumannii strain, treated with cyclophosphamide, and subsequently infected intranasally with A. baumannii. Several differences between their model and ours likely account for this discrepancy. First, Dolery et al. used the AB5075 challenge strain, which is less virulent than the LAC-4 strain used in our experiments (see Supplementary Figure S1). Second, our vaccination protocol consisted of a single dose, whereas theirs involved three administrations. Third, in their study, the challenge occurred only two weeks after the final boost, compared with six weeks after vaccination in our model. Intranasal administration of inactivated LAC-4 has been shown to induce an innate immunity–mediated protective phase lasting at least seven days post-administration (34). Therefore, the stronger protection reported by Dolery et al. may result from the combination of multiple boosts, the short interval between the last boost and the challenge, and the lower virulence of the AB5075 strain.
Finally, as experimental mouse models often fail to predict the immunogenicity of a vaccine in humans, we analyzed in vitro the ability of HK LAC-4 to induce the activation of human monocyte-derived dendritic cells (DCs) and T cells. We found that HK LAC-4 is a potent activator, capable of inducing the expression of co-stimulatory signals on DCs and of inducing T cell proliferation and cytokine production, suggesting that HK LAC-4 could potentially activate these components of adaptive immune system of patients and induce the development of a protective memory response against A. baumannii.
Despite our promising findings, our study nevertheless has several important limitations. A. baumannii is known to exhibit enormous genetic and phenotypic variation. Our study focused on the LAC-4 strain which is described as hypervirulent in mice. Our results would be further substantiated if similar results were obtained with other strains of A. baumannii. In this study, we showed that HK LAC-4 is a very good activator of the murine and human immune systems. This is encouraging for the development of a vaccine but should elicit caution to its possible side effects. HK LAC-4 would likely be too proinflammatory to be used safely in humans. Strategies to enhance antigen exposure while minimizing inflammation include reducing the antigen dose, splitting doses (prime and boost), choosing a safer administration route (e.g., subcutaneous injection), and using a matrix for controlled antigen release. A recent study by Luzuriaga et al. (74) demonstrated that it was possible to inactivate bacteria and make them more immunogenic by biomimetically mineralizing them within a metal−organic framework. This process makes it possible to increase specific antibody levels induced as well as the level of protection against an extracellular bacterium such as the E. coli CFT073 strain responsible for urinary tract infections.
Taken together, our results improve our understanding of the protective mechanisms involved in the immune response against A. baumannii. They also suggest that the development of vaccines capable of protecting immunocompromised patients against opportunistic infections such as those caused by A. baumannii is possible.
4 Materials and methods
4.1 Ethics statement
The procedures used in this study and the handling of the mice complied with current European legislation (Directive 86/609/EEC). The Animal Welfare Committee of the Université de Namur (UNamur, Belgium) reviewed and approved the complete protocol for Acinetobacter baumannii infection (Permit Number: UN-LE-20/343, UN-LE-22/378). Whole blood for the isolation of human PBMC was collected from healthy donors as described in the ethical protocol/amendment IXP-001_V3 (Belgium; Reg. No. B6702014215858), protocol IXP-003_V1 (Belgium; Reg. No. B707201627607) or protocol IXP-004_V1 (The Netherlands; Reg. No. NL57912.075.16).
4.2 Mice, PBMC and bacterial strains
Wild-type C57BL/6 mice were obtained from Harlan (Bicester, UK). IFN-γ−/−, δTCR−/−, CD3ϵ−/−, MuMT−/− C57BL/6 mice were purchased from The Jackson Laboratory (Bar Harbor, ME). IL-12p35−/− C57BL/6 mice were obtained from Dr. B. Ryffel (University of Orleans, France). IL-17RA−/− C57BL/6 mice were obtained from Dr. K. Huygen (Belgian Scientific Institute for Public Health, Brussels, Belgium). TAP1−/− C57BL/6 mice and MHCII−/− C57BL/6 mice were acquired from Jörg Reimann (University of Ulm, Ulm, Germany). All wild-type and deficient mice used in this study were bred in the animal facility of the Gosselies campus of the Université Libre de Bruxelles (ULB, Belgium).
Cryopreserved primary PBMC were isolated from whole blood donated by healthy volunteers. Written informed consent was obtained from all patients enrolled in the study. All blood samples were tested and found negative for hepatitis B virus (HBV), hepatitis C virus (HCV), and human immunodeficiency virus (HIV). PBMC were separated from the blood by density gradient centrifugation and subsequently cryopreserved in fetal bovine serum (FBS), supplemented with 10% dimethyl sulfoxide, by controlled rate freezing. The PBMC were kept in cryogenic storage (−180 °C) until use.
A. baumannii LAC-4 (31, 32) (ASM78673v1 genome, https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_000786735.1/) was obtained from Wangxue Chen (Human Health Therapeutics Research Center, National Research Council of Canada, Ottawa, ON, Canada). A. baumannii AB5075 was isolated in 2008 from a combatant wound infection (38) and was obtained from Dr. C. Van der Henst (Structural Biology Brussels, Vrije Universiteit Brussel (VUB) and Microbial Resistance and Drug Discovery, VIB-VUB Center for Structural Biology, VIB, Flanders Institute for Biotechnology, Brussels, Belgium). A. baumannii was always handled in BSL-2 or BSL-3 containment at the Université Libre de Namur (UNamur, Namur).
4.3 Construction of the itrA gene deletion mutant
To generate the non-capsulated A. baumannii LAC-4 strain, the itrA3 gene (ABLAC_36890 from the LAC-4 reference genome CP007712) was obtained using the integrative plasmid pK18-apraR [pUC derivative (75)] generated using the following primers: itrA3_up_for_short: GCAAGTATCGAAGCAATATCAG; itrA3_up_rev_short:GGCCCAATTCGCCCTATAGTGAGTCGG
GATTAATGCGGTTAAGGAAATTACG; itrA3_down_rev_short: CCTAATTTTGGATGTTCTAAGCC; itrA3_down_for_short:CCGACTCACTATAGGGCGAATTGGGCCGCTCCAGATCAGTT
AGTAGG. Briefly, the pK18-ΔitrA3 was integrated into the parental wild-type LAC-4 strain and excised using the sucrose counter-selection marker as previously described (42). The lack of capsule of the LAC-4ΔitrA3 strain was confirmed by density gradients as previously described (76).
4.4 A. baumannii infection
Bacteria stored at -80°C in LB-glycerol 30% were inoculated in LB medium and incubated overnight at 37°C under agitation until the stationary phase. Then the culture was diluted to an OD600 (Optical Density measured at a wavelength of 600 nm) of 0.2 for AB5075 and 0.4 for LAC-4 and then was grown until an OD600 of 0.7-0.8. This log-phase culture was then centrifugated for 10 minutes at 5000 g and the pellet was washed twice with Phosphate-Buffered Saline (PBS). Then it was diluted to the target concentration prior to infection. The infectious dose was assessed by plating serial dilutions.
Mice were anesthetized with a cocktail of Xylazine (9 mg/kg) and Ketamine (36 mg/kg) in PBS before being inoculated by intranasal injection with the indicated dose of A. baumannii in 30 µl of PBS. Mice infected intraperitoneally were injected with the indicated dose in 500 µL. Control animals were inoculated with the same volume of PBS. The infectious doses were validated by plating serial dilutions of the inoculums. At the selected time after infection, mice were sacrificed by cervical dislocation. Immediately after sacrifice, the lungs and spleen were collected for bacterial count and qRT-PCR analyses.
4.5 Preparation of heat-killed bacteria
To carry out vaccinations as well as in vitro tests, different species and strains of bacteria were killed by heat. We used the previously described A. baumannii strains as well as Escherichia coli K12 MG1655 and B. melitensis strain 16M (Biotype1; ATCC 23456; American Type Culture Collection). To prepare Heat-Killed (HK) bacteria, overnight cultures of bacteria were grown under shaking at 37°C in 2YT media (Luria-Bertani (LB) broth 32 g/L with yeast extract 5 g/L and peptone 10 g/L) for B. melitensis and in LB media for A. baumannii and E. coli. B. melitensis and E. coli from an overnight liquid culture were washed twice in PBS (10 minutes at 3500 g for Brucella and 5000 g for E. coli). A. baumannii from an overnight liquid culture were washed twice in PBS (10 minutes at 5000 g) and diluted to an optical density of 0.4 in fresh LM media and growth for 90 minutes at 37°C under constant agitation. and then washed twice in PBS. After washing, the bacteria were resuspended to a concentration of 2x109 CFU/ml in PBS. This solution was heated at 80°C for 1 hour in a water bath. Following this treatment, the bacterial solution was stored at -20°C before use. Each batch of killed bacteria was verified to confirm the killing by growing it in an incubator at 37°C and counting the CFU after 24 hours for A. baumannii and E. coli and after 72 hours for B. melitensis.
4.6 Vaccination
C57BL/6 mice were unvaccinated or vaccinated by intranasal or intraperitoneal injection with 108 CFU of strains of the A. baumannii of interest, as indicated. Samples of blood for antibody detection were collected 4 weeks post-vaccination for analysis of the humoral response against A. baumannii. Six weeks post-vaccination, mice were challenged intranasally with live LAC-4 bacteria (2x106 or 2x107 CFU, as indicated). In some experiments, monohydrate cyclophosphamide (Sigma) diluted in PBS was administrated intraperitoneally at 4 days (150 mg/kg) and 1 day (100 mg/kg) before the challenge. Lungs and spleen were harvested 24 and 120 hours post-challenge and the CFU levels in each organ were counted.
4.7 Bacterial counting
Organs were homogenized in PBS/0.1% X-100 Triton (Sigma-Aldrich). We performed successive serial dilutions in PBS to obtain the most accurate bacterial count and plated them on LB medium. The CFU were counted after 1 day of incubation at 37°C.
4.8 RNA extraction from lungs and spleen
RNA from the whole lungs and spleen was extracted by the TRIzol™/Chloroform method. Briefly, the lungs and spleen were harvested from mice and ground in 1 mL of TRIzol™ (TRIzol™ Reagent - Invitrogen™) with a Polytron Homogenizer. After 5 minutes of incubation at room temperature, 200 µL of chloroform were added and the tube was mixed vigorously, then incubated for 2–3 minutes at room temperature. After centrifugation for 15 minutes (12,000 g at 4°C), the aqueous phase was recovered and RNA was extracted using the “NucleoSpin RNA Plus (Macherey-Nagel)” kit according to the manufacturer’s protocol.
4.9 qRT-PCR analysis of cytokines
The RNA was then reverse transcribed with Superscript II reverse transcriptase (Invitrogen) using hexamer random primers as described by the manufacturer. A condition without reverse transcriptase was also conducted in parallel as a negative control. cDNA was then mixed with SybrGreen mix (FastStart Universal SYBR Green Master (Rox) - Roche) and the appropriate primer sets and subjected to qRT-PCR in a LightCycler 480 (Roche). The forward and reverse primers used were 5’-GGCAACTGTTCCTGAACTCA-3’ and 5’-GGGTCCGTCAACTTCAAAGA-3’ for IL-1β, 5’-CCTGTCTATACCACTTCACA-3’ and 5’-CTCTTTTCTCATTTCCACGATTTCC-3’ for IL-6, 5’-GCCTCCCTCTCATCAGTTCTA-3’ and 5’-GCTACGACGTGGGCTACAG-3’ for TNF-α, 5’-TGCCAAGTTTGAGGTCAACA-3’ and 5’-GAATCAGCAGCGACTCCTTT-3’ for IFN-γ, 5’-ATCCCTCAAAGCTCAGCGTGTC-3’ and 5’-GGGTCTTCATTGCGGTGGAGAG-3’ for IL-17A, 5’-CCCTGGAGGATAACACTGTGA-3’ and 5’-AATTCACGTGGGACAGAAATG-3’ for IL-17F and 5’-CAGCAGCCATACATCGTCAA-3’ and 5’-GCCGGACATCTGTGTTGTTA-3’ for IL-22. Ribosomal Protein L32 (RPL32, forward 5’-ACATCGGTTATGGGAGCAAC-3’, reverse 5’-TCCAGCTCCTTGACATTGTG-3’) mRNA was used as the reference housekeeping gene for normalization. A total of 40 two-step cycles were performed as follows: 95°C for 15 seconds and 60°C for 1 minute preceded by 10 minutes at 95°C for activation. Melting curves were then performed to assess the primer specificity. The target mRNA fold change was calculated based on the 2-ΔΔCt formula, using the RPL32 gene as the reference gene and RNA from naïve mice as the standard condition. Four biological replicates were performed for each gene tested.
4.10 Enzyme-linked immunosorbent assay to detect anti-A. baumannii antibodies
The presence of Acinetobacter baumannii-specific murine IgM, IgG1 and IgG3 was determined by ELISA. Polystyrene plates (269620; Nunc) were coated with Heat-Killed (HK) A. baumannii LAC-4 or AB5075 at a dose of 107 CFU/mL, as indicated, and incubated overnight at 4°C. The plates were blocked for 2 hours at room temperature with 200 μl/well of PBS-1% Bovine Serum Albumin (BSA). Then, plates were incubated with 50 μl/well of plasma in serial dilutions in PBS-0.1% BSA. The plasma of uninfected mice and PBS were used as negative controls. After four washes with PBS, isotype-specific goat anti-mouse HRP conjugated antibodies were added (50 μl/well) at appropriate dilutions (anti-IgM from Sigma-Aldrich; anti IgG1 LO-MG1–13 HRPO and LO-anti IgG3 MG3–13 HRPO from LOIMEX). After 1 hour of incubation at room temperature, plates were washed four times in PBS and 100 μl/well of TMB substrate solution (BD OptEiA Kit) was added. After 15 minutes of incubation at room temperature in the dark, the enzyme reaction was stopped by adding 25 μl/well of 2 N H2SO4. The absorbance was measured at 450 nm.
4.11 Analysis of antigen profiles by Western blot
Bacterial cultures of A. baumannii were normalized at 2 x 109 bacteria/mL, then heat killed as previously described. Two milliliters of culture were concentrated in 300 µL of PBS containing the loading buffer (2% SDS, 5% β-mercaptoethanol, 10% glycerol, 0.03 M Tris HCl pH6.8) and samples were heated at 90 °C for 10 minutes before being loaded on 4-20% mini-PROTEAN acrylamide gels (BioRad), with 20 µL of samples per well. Migration was performed for 1 hour at 150V. After migration, one gel was kept for Coomassie Blue staining, while the others had their proteins transferred onto a nitrocellulose membrane which was blocked overnight in PBS supplemented with 0.05% Tween (PBS-T) and 5% milk. For primary antibodies, sera from 8 different vaccinated mice were pooled together and diluted 4 times in PBS-T supplemented with 1% milk. Incubation of the membranes with their respective primary antibodies were performed statically for 1 hour on clean petri dishes with drops of 300 µL for each membrane. Membranes were then washed in PBS-T 4 times for 5 min, before being incubated with secondary antibodies (polyclonal Rabbit anti-Mouse Ig/HRP, Agilent) diluted 5000 times in PBS-T supplemented with 1% milk. After 4 washings, proteins were revealed using Clarity Western ECL Substrate (Biorad) with Image Quant LAS 4000 (General Electric).
4.12 Human dendritic cell maturation assay
Peripheral Blood Mononuclear Cells (PBMC) from healthy human donors were thawed from the ImmunXperts biobank. Monocytes were isolated from PBMC using a MACS magnetic separation column (Miltenyi) following the manufacturer’s instructions and purity was evaluated by CD14 staining using a BD Lsrfortessa X-20 flow cytometer. Cells were then resuspended to a cellular density of 106cells/ml and plated on a tissue culture treated 24-well plate (1ml per well) in CellGenix DC medium (CellGenix) with added Gentamicin antibiotic (Sigma-Aldrich), IL-4 and GM-CSF for 5 days. At day 5, cells were stained for FACS analysis with several DC activation markers to assess their immature (iDC) state (CD14 Miltenyi, CD40 BD, CD80 BD, CD83 Miltenyi, CD86 Miltenyi, CD209 Miltenyi and HLA-DR BD). On the same day, the appropriate stimuli (various heat-killed bacteria and LPS from E. coli from Sigma-Aldrich) were added to the cell culture for 48 hours. At day 7, cells were stained for FACS analysis with the same markers used at day 5 to assess their mature state (mDC). Supernatant at day 7 was collected for further analysis.
4.13 ELISA to detect human cytokines
Supernatant from the cell culture was collected and stored at -80°C. After thawing, the presence of the cytokines IL-12p70, IL-23 and IL-17A was detected using LEGEND MAX™ Human ELISA kits (BioLegend) with pre-coated plates following the manufacturer’s instructions.
4.14 Luminex to detect human cytokines
Supernatant from cell culture was collected and stored at -80°C. After thawing, the presence of the cytokines IFN-γ and TNF-α was detected using the Luminex technology following the manufacturer’s instructions (MILLIPLEX MAP Human Cytokine/Chemokine kit, Merck).
4.15 Proliferation assay on human PBMC
Peripheral Blood Mononuclear Cells (PBMC) from healthy human donors were thawed from the ImmunXperts biobank and incubated for 6 days at 37°C with the appropriate stimuli: CEF+CEFTA (pool of 35 peptides-MHC class I and II-restricted T-cell epitopes from human Cytomegalovirus, Epstein Barr virus, Influenza virus, Tetanus toxin and Adenovirus 5 from Mabtech), HMBPP (4-Hydroxy-3-methylbut-2-enyl diphosphate lithium salt, Sigma-Aldrich) and various heat-killed bacteria. T cell proliferation was assessed by measuring the incorporation of 5-Ethynyl-2´-deoxyuridine (EdU). EdU is a thymidine analogue which is incorporated into the DNA of dividing cells during the S phase. On day 6, cell cultures were pulsed with EdU at 1 µM for approximately 16 hours. The next day, cell culture supernatants were collected and stored at -80°C for cytokine analysis. The remaining cells were fluorescently stained for viability and T cell surface markers (CD3 BD, CD4 BD, CD8 BD and γδTCR-Miltenyi), fixed, permeabilized and the incorporated EdU was stained with a fluorescent azide (Click-iT™ EdU Flow Cytometry Assay Kit, Invitrogen, Thermofisher Scientific). Cells were then acquired on a BD Lsrfortessa X-20.
4.16 Intracellular cytokine staining on human PBMC
Peripheral Blood Mononuclear Cells (PBMC) from healthy human donors were thawed from the ImmunXperts biobank and incubated for 7 days at 37°C with the appropriate stimuli. At day 7, part of the supernatant was collected, and the remaining cells were treated with Brefeldin A/Monensin (Sigma) for 4–5 hours at 37°C following the manufacturer’s instructions. For some conditions, cells were only stimulated for the last 4–5 hours with Brefeldin A/Monensin. Cells were then stained for viability and extracellular markers (CD3-BD, CD4-BD, CD8-BD and γδTCR-Miltenyi), fixed and permeabilized (BD Cytofix/Cytoperm™ Fixation/Permeabilization Kit) and intracellularly stained for the cytokines IFN-γ-BD, TNF-α-BD and IL-17A-eBiosciences. Cells were then acquired on a BD Fortessa X-20.
4.17 Statistical analysis
For comparisons of CFU counts between groups, statistical analyses were performed using the non-parametric one-way ANOVA followed by multiple comparison tests in GraphPad Prism. Survival curves were compared using the log-rank (Mantel–Cox) test in GraphPad Prism. To analyze ELISA data, we use two-way Anova with Dunnett’s multiple comparisons test in GraphPad Prism. A p-value < 0.05 was considered statistically significant (*p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Whole blood for the isolation of human PBMC was collected from healthy donors as described in the ethical protocol/amendment IXP-001_V3 (Belgium; Reg. No. B6702014215858), protocol IXP-003_V1 (Belgium; Reg. No. B707201627607) or protocol IXP-004_V1 (The Netherlands; Reg. No. NL57912.075.16). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study. The animal study was approved by Animal Welfare Committee of the Université de Namur (UNamur, Belgium). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
TR: Formal analysis, Investigation, Methodology, Writing – review & editing. EB: Formal analysis, Investigation, Methodology, Writing – review & editing. LB: Formal analysis, Investigation, Methodology, Writing – review & editing. KP: Methodology, Conceptualization, Formal analysis, Validation, Writing – review & editing. SéD: Formal analysis, Methodology, Validation, Writing – review & editing. YT: Methodology, Writing – review & editing. DV: Methodology, Writing – review & editing. SaD: Methodology, Writing – review & editing. VF: Writing – review & editing. JM: Resources, Writing – review & editing. CH: Funding acquisition, Methodology, Resources, Writing – review & editing. XB: Writing – review & editing. FA: Conceptualization, Formal analysis, Investigation, Methodology, Supervision, Validation, Writing – review & editing. EM: Conceptualization, Funding acquisition, Investigation, Methodology, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the Horizon Europe Project BAXERNA 2.0 (101080544) and by the Flanders Institute for Biotechnology (VIB). TR is supported by Horizon Europe Project BAXERNA 2.0 grant. EB hold FRIA PhD grants from the FRS-FNRS (Belgium). LB is supported by the European Union’s Horizon 2020 research and innovation program under the Marie Skłodowska-Curie grant agreement No 860325. FA is a Senior Research Associate from the FRS-FNRS (Belgium). EM is a Senior Research Associate from the FRS-FNRS (Belgium).
Conflict of interest
Author LB was affiliated with the company ImmunXperts SA, a Q2 Solutions Company during the study.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1641997/full#supplementary-material
Supplementary Figure 1 | Comparison of the virulence of LAC-4 and AB5075 strains in the C57BL/6 mouse intranasal infection model. Wild-type C57BL/6 mice were inoculated intranasally (IN) with 2x106 or 2x106 CFU of LAC-4 or HK 5075 A. baumannii strains. (A) At 4, 24 and 48 hours post infection, some mice were sacrificed and the number of bacteria per gram in the lungs and spleen was evaluated by CFU counting. Red lines represent the geometric mean for each group of mice. These data are representative of two independent experiments. (B) During the 120 hours following infection, the fitness of infected mice was monitored. When the human endpoint was reached, the mice were euthanized. These data represent a pool of two independent experiments. n = number of mice per group. Significant differences between groups are marked with asterisks: *p < 0.05, ***p < 0.001, ****p < 0.0001, in a log-rank (Mantel–Cox) test for survival curve and a (Wilcoxon-) Mann-Whitney post-test for CFU count.
Supplementary Figure 2 | Production of IL-17A, IL-17F, IL-22, and TNF-α is not reduced in δTCR-deficient mice in response to infection with A. baumannii LAC-4 strain. Wild-type and δTCR-/- C57BL/6 mice were inoculated intranasally with PBS (control) or 2x106 CFU of live A. baumannii LAC-4 strain in PBS, as indicated. At 4 and 24 hours post-infection, mice were sacrificed, and lungs were collected. Expression levels of the indicated genes were assessed by quantitative RT-PCR and expressed as relative expression to RPL32 housekeeping mRNA (log10). The naïve mice (number 1) condition was set to 1. n = 4 mice for each condition. Significant differences between groups (B) are marked with asterisks: ****p < 0.0001, in a (Wilcoxon-) Mann-Whitney post-test.
Supplementary Figure 3 | Evaluation of A. baumannii LAC-4 infection control in different stains of C57BL/6 mice. Wild-type and immunodeficient C57BL/6 mice were infected intranasally with LAC-4 A. baumannii (2.106 CFU) and sacrificed at 24 hours post-infection. The data represent the number of CFU/g of lungs or spleen, as indicated. Red lines represent the geometric mean. Dotted red line represents the mean of the results for wild-type mice. Significant differences between wild-type and deficient mice are marked with asterisks: *p < 0.1, **p < 0.01, ***p < 0.001, ****p < 0.0001, in a (Wilcoxon-) Mann-Whitney post-test.
References
1. Whiteway C, Breine A, Philippe C, and van der Henst C. Acinetobacter baumannii. Trends Microbiol. (2022) 30:199–200. doi: 10.1016/j.tim.2021.11.008
2. Sharma R and Lakhanpal D. Acinetobacter baumannii: A comprehensive review of global epidemiology, clinical implications, host interactions, mechanisms of antimicrobial resistance and mitigation strategies. Microb Pathog. (2025) 204:107605. doi: 10.1016/j.micpath.2025.107605
3. Munoz-Price LS and Weinstein RA. Acinetobacter infection. N Engl J Med. (2008) 358:1271–81. doi: 10.1056/nejmra070741
4. Peleg AY, Seifert H, and Paterson DL. Acinetobacter baumannii: Emergence of a successful pathogen. Clin Microbiol Rev. (2008) 21:538–82. doi: 10.1128/CMR.00058-07
5. Joly-Guillou ML. Clinical impact and pathogenicity of Acinetobacter. Clin Microbiol Infect. (2005) 11:868–73. doi: 10.1111/j.1469-0691.2005.01227.x
6. Breslow JM, Monroy MA, Daly JM, Meissler JJ, Gaughan J, Adler MW, et al. Morphine, but not trauma, sensitizes to systemic acinetobacter baumannii infection. J Neuroimmune Pharmacol. (2011) 6:551–65. doi: 10.1007/s11481-011-9303-6
7. Galac MR, Snesrud E, Lebreton F, Stam J, Julius M, Ong AC, et al. A diverse panel of clinical acinetobacter baumannii for research and development. Antimicrob Agents Chemother. (2020) 64:1–11. doi: 10.1128/AAC.00840-20
8. Rodrigues DLN, Morais-Rodrigues F, Hurtado R, Dos Santos RG, Costa DC, Barh D, et al. Pan-resistome insights into the multidrug resistance of Acinetobacter baumannii. Antibiotics. (2021) 10:3–5. doi: 10.3390/antibiotics10050596
9. Fournier PE, Richet H, and Weinstein RA. The epidemiology and control of acinetobacter baumannii in health care facilities. Clin Infect Dis. (2006) 42:692–9. doi: 10.1086/500202
10. Nowak J, Zander E, Stefanik D, Higgins PG, Roca I, Vila J, et al. High incidence of pandrug-resistant Acinetobacter baumannii isolates collected from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J Antimicrob Chemother. (2017) 72:3277–82. doi: 10.1093/jac/dkx322
11. Singh JK, Adams FG, and Brown MH. Diversity and function of capsular polysaccharide in Acinetobacter baumannii. Front Microbiol. (2019) 10:3301. doi: 10.3389/fmicb.2018.03301
12. Tipton KA, Chin C, Farokhyfar M, Weiss DS, and Rather PN. Role of capsule in resistance to disinfectants, host antimicrobials, and desiccation in acinetobacter baumannii. Antimicrob Agents Chemother (2018) 62:1–6. doi: 10.1128/AAC.01188-18
13. Chiang SR, Jung F, Tang HJ, Chen CH, Chen CC, Chou HY, et al. Desiccation and ethanol resistances of multidrug resistant Acinetobacter baumannii embedded in biofilm: The favorable antiseptic efficacy of combination chlorhexidine gluconate and ethanol. J Microbiol Immunol Infect. (2018) 51:770–7. doi: 10.1016/j.jmii.2017.02.003
14. Aranda J, Bardina C, Beceiro A, Rumbo S, Cabral MP, Barbé J, et al. Acinetobacter baumannii reca protein in repair of DNA damage, antimicrobial resistance, general stress response, and virulence. J Bacteriol. (2011) 193:3740–7. doi: 10.1128/JB.00389-11
15. Cahill SM, Hall RM, and Kenyon JJ. An update to the database for Acinetobacter baumannii capsular polysaccharide locus typing extends the extensive and diverse repertoire of genes found at and outside the K locus. Microb Genomics. (2022) 8:1–21. doi: 10.1099/mgen.0.000878
16. Wieland K, Chhatwal P, and Vonberg RP. Nosocomial outbreaks caused by Acinetobacter baumannii and Pseudomonas aeruginosa: Results of a systematic review. Am J Infect Control. (2018) 46:643–8. doi: 10.1016/j.ajic.2017.12.014
17. Rangel K, Chagas TPG, and De-Simone SG. Acinetobacter baumannii infections in times of COVID-19 pandemic. Pathogens. (2021) 10:1–13. doi: 10.3390/pathogens10081006
18. Karnoukh KI, Drozdov VN, Shikh EV, Zhilina SV, and Lazareva NB. Etiology and antimicrobial resistance of secondary bacterial infections in patients hospitalized with COVID-19: A retrospective analysis. Vestn Ross Akad Meditsinskikh Nauk. (2022) 77:25–32. doi: 10.15690/vramn1552
19. Russo A, Gavaruzzi F, Ceccarelli G, Borrazzo C, Oliva A, Alessandri F, et al. Multidrug-resistant Acinetobacter baumannii infections in COVID-19 patients hospitalized in intensive care unit. Infection. (2022) 50:83–92. doi: 10.1007/s15010-021-01643-4
20. Tacconelli E, Carrara E, Savoldi A, Harbarth S, Mendelson M, Monnet DL, et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect Dis. (2018) 18:318–27. doi: 10.1016/S1473-3099(17)30753-3
21. Rice LB. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J Infect Dis. (2008) 197:1079–81. doi: 10.1086/533452
22. Knapp S, Wieland CW, Florquin S, Pantophlet R, Dijkshoorn L, Tshimbalanga N, et al. Differential roles of CD14 and Toll-like receptors 4 and 2 in murine Acinetobacter pneumonia. Am J Respir Crit Care Med. (2006) 173:122–9. doi: 10.1164/rccm.200505-730OC
23. Noto MJ, Boyd KL, Burns WJ, Varga MG, Peek RM, and Skaar EP. Toll-like receptor 9 contributes to defense against Acinetobacter baumannii infection. Infect Immun. (2015) 83:4134–41. doi: 10.1128/IAI.00410-15
24. Kale SD, Dikshit N, Kumar P, Balamuralidhar V, Khameneh HJ, Bin Abdul Malik N, et al. Nod2 is required for the early innate immune clearance of Acinetobacter baumannii from the lungs. Sci Rep. (2017) 7:1–9. doi: 10.1038/s41598-017-17653-y
25. Bruhn KW, Pantapalangkoor P, Nielsen T, Tan B, Junus J, Hujer KM, et al. Host fate is rapidly determined by innate effector-microbial interactions during acinetobacter baumannii bacteremia. J Infect Dis. (2015) 211:1296–305. doi: 10.1093/infdis/jiu593
26. Van Faassen H, KuoLee R, Harris G, Zhao X, Conlan JW, and Chen W. Neutrophils play an important role in host resistance to respiratory infection with Acinetobacter baumannii in mice. Infect Immun. (2007) 75:5597–608. doi: 10.1128/IAI.00762-07
27. Talyansky Y, Nielsen TB, Yan J, Carlino-Macdonald U, Di Venanzio G, Chakravorty S, et al. Capsule carbohydrate structure determines virulence in Acinetobacter baumannii. PloS Pathog. (2021) 17:1–24. doi: 10.1371/JOURNAL.PPAT.1009291
28. Akoolo L, Pires S, Kim J, and Parker D. The Capsule of Acinetobacter baumannii Protects against the Innate Immune Response. J Innate Immun. (2022) 14 (5): 543–54. doi: 10.1159/000522232
29. Morris FC, Dexter C, Kostoulias X, Uddin MI, and Peleg AY. The mechanisms of disease caused by acinetobacter baumannii. Front Microbiol. (2019) 10:1601. doi: 10.3389/fmicb.2019.01601
30. Qiu H, Li Z, KuoLee R, Harris G, Gao X, Yan H, et al. Host resistance to intranasal Acinetobacter baumannii reinfection in mice. Pathog Dis. (2016) 74:12–5. doi: 10.1093/femspd/ftw048
31. Ou HY, Kuang SN, He X, Molgora BM, Ewing PJ, Deng Z, et al. Complete genome sequence of hypervirulent and outbreak-associated Acinetobacter baumannii strain LAC-4: Epidemiology, resistance genetic determinants and potential virulence factors. Sci Rep. (2015) 5:1–13. doi: 10.1038/srep08643
32. Valentine SC, Contreras D, Tan S, Real LJ, Chu S, and Xu HH. Phenotypic and molecular characterization of Acinetobacter baumannii clinical isolates from nosocomial outbreaks in Los Angeles County, California. J Clin Microbiol. (2008) 46:2499–507. doi: 10.1128/JCM.00367-08
33. Harris G, Lee RK, Lam CK, Kanzaki G, Patel GB, Xu HH, et al. A Mouse model of acinetobacter baumannii-associated pneumonia using a clinically isolated hypervirulent strain. Antimicrob Agents Chemother. (2013) 57:3601–13. doi: 10.1128/AAC.00944-13
34. Gu H, Zeng X, Peng L, Xiang C, Zhou Y, Zhang X, et al. Vaccination induces rapid protection against bacterial pneumonia via training alveolar macrophage in mice. Elife. (2021) 10:1–19. doi: 10.7554/eLife.69951
35. Kuolee R, Harris G, Yan H, Xu HH, Conlan WJ, Patel GB, et al. Intranasal immunization protects against Acinetobacterbaumannii-associated pneumonia in mice. Vaccine. (2014) 33:260–7. doi: 10.1016/j.vaccine.2014.02.083
36. Ahlmann M and Hempel G. The effect of cyclophosphamide on the immune system: implications for clinical cancer therapy. Cancer Chemother Pharmacol. (2016) 78:661–71. doi: 10.1007/s00280-016-3152-1
37. MaNepalli S, Gandhi JA, Ekhar VV, Asplund MB, Coelho C, and Martinez LR. Characterization of a cyclophosphamide-induced murine model of immunosuppression to study Acinetobacter baumannii pathogenesis. J Med Microbiol. (2013) 62:1747–54. doi: 10.1099/jmm.0.060004-0
38. Jacobs AC, Thompson MG, and Black CC. AB5075, a highly virulent isolate of acinetobacter baumannii, as a model strain for the evaluation of pathogenesis and antimicrobial treatments. MBio. (2014) 5:1–10. doi: 10.1128/mBio.01076-14.Editor
39. Tan SYY, Chua SL, Liu Y, Høiby N, Andersen LP, Givskov M, et al. Comparative genomic analysis of rapid evolution of an extreme-drug- resistant Acinetobacter baumannii clone. Genome Biol Evol. (2013) 5:807–18. doi: 10.1093/gbe/evt047
40. Cosgrove D, Gray D, Dierich A, Kaufman J, Lemeur M, Benoist C, et al. Mice lacking MHC class II molecules. Cell. (1991) 66:1051–66. doi: 10.1016/0092-8674(91)90448–8
41. Kitamura D, Roes J, Kühn R, and Rajewsky K. A B cell-deficient mouse by targeted disruption of the membrane exon of the immunoglobulin mu chain gene. Nature. (1991) 350:423–6. doi: 10.1038/350423a0
42. Whiteway C, Valcek A, Philippe C, Strazisar M, De Pooter T, Mateus I, et al. Scarless excision of an insertion sequence restores capsule production and virulence in Acinetobacter baumannii. ISME J. (2022) 16:1473–7. doi: 10.1038/s41396-021-01179-3
43. Lapaque N, Moriyon I, Moreno E, and Gorvel JP. Brucella lipopolysaccharide acts as a virulence factor. Curr Opin Microbiol. (2005) 8:60–6. doi: 10.1016/j.mib.2004.12.003
44. Lau YT and Tan HS. Acinetobacter baumannii subunit vaccines: recent progress and challenges. Crit Rev Microbiol. (2024) 50:434–49. doi: 10.1080/1040841X.2023.2215303
45. Yang N, Jin X, Zhu C, Gao F, Weng Z, Du X, et al. Subunit vaccines for Acinetobacter baumannii. Front Immunol. (2023) 13:1088130. doi: 10.3389/fimmu.2022.1088130
46. Hu Y, Zhang X, Deng S, Yue C, Jia X, and Lyu Y. Non-antibiotic prevention and treatment against Acinetobacter baumannii infection: Are vaccines and adjuvants effective strategies? Front Microbiol. (2023) 14:1049917. doi: 10.3389/fmicb.2023.1049917
47. McConnell MJ and Pachón J. Active and passive immunization against Acinetobacter baumannii using an inactivated whole cell vaccine. Vaccine. (2010) 29:1–5. doi: 10.1016/j.vaccine.2010.10.052
48. Dollery SJ, Zurawski DV, Gaidamakova EK, Matrosova VY, Tobin JK, Wiggins TJ, et al. Radiation-inactivated Acinetobacter baumannii vaccine candidates. Vaccines. (2021) 9:1–16. doi: 10.3390/vaccines9020096
49. Dollery SJ, Zurawski DV, Bushnell RV, Tobin JK, Wiggins TJ, MacLeod DA, et al. Whole-cell vaccine candidates induce a protective response against virulent Acinetobacter baumannii. Front Immunol. (2022) 13:941010. doi: 10.3389/fimmu.2022.941010
50. Di Tommaso A, De Magistris MT, Bugnoli M, Marsili I, Rappuoli R, and Abrignani S. Formaldehyde treatment of proteins can constrain presentation to T cells by limiting antigen processing. Infect Immun. (1994) 62:1830–4. doi: 10.1128/iai.62.5.1830-1834.1994
51. Ibsen PH. The effect of formaldehyde, hydrogen peroxide and genetic detoxification of pertussis toxin on epitope recognition by murine monoclonal antibodies. Vaccine. (1996) 14:359–68. doi: 10.1016/0264-410X(95)00230-X
52. Tano Y, Shimizu H, Martin J, Nishimura Y, Simizu B, and Miyamura T. Antigenic characterization of a formalin-inactivated poliovirus vaccine derived from live-attenuated Sabin strains. Vaccine. (2007) 25:7041–6. doi: 10.1016/j.vaccine.2007.07.060
53. Fan Y, Mu Y, Lu L, Tian Y, Yuan F, Zhou B, et al. Hydrogen peroxide-inactivated bacteria induces potent humoral and cellular immune responses and releases nucleic acids. Int Immunopharmacol. (2019) 69:389–97. doi: 10.1016/j.intimp.2019.01.055
54. Mai TT, Kayansamruaj P, Taengphu S, Senapin S, Costa JZ, del-Pozo J, et al. Efficacy of heat-killed and formalin-killed vaccines against Tilapia tilapinevirus in juvenile Nile tilapia (Oreochromis niloticus). J Fish Dis. (2021) 44:2097–109. doi: 10.1111/jfd.13523
55. Yang X, Wei R, Liu H, Wei T, Zeng P, Cheung YC, et al. Discovery of a monoclonal antibody that targets cell-surface pseudaminic acid of acinetobacter baumannii with direct bactericidal effect. ACS Cent Sci. (2024) 10:439–46. doi: 10.1021/acscentsci.3c01507
56. Leung KK, Schaefer K, Lin Z, Yao Z, and Wells JA. Engineered proteins and chemical tools to probe the cell surface proteome. Chem Rev. (2025) 125:4069–110. doi: 10.1021/acs.chemrev.4c00554
57. Kolls JK, McCray PB, and Chan YR. Cytokine-mediated regulation of antimicrobial proteins. Nat Rev Immunol. (2008) 8:829–35. doi: 10.1038/nri2433
58. Van Kaer L, Ashton-Rickardt PG, Ploegh HL, and Tonegawa S. TAP1 mutant mice are deficient in antigen presentation, surface class I molecules, and CD4-8+ T cells. Cell. (1992) 71:1205–14. doi: 10.1016/s0092-8674(05)80068-6
59. Gourapura RJ, Khan MA, Gallo RM, Shaji D, Liu J, and Brutkiewicz RR. Forming a complex with MHC class I molecules interferes with mouse CD1d functional expression. PloS One. (2013) 8:1–11. doi: 10.1371/journal.pone.0072867
60. Goldszmid RS, Bafica A, Jankovic D, Feng CG, Caspar P, Winkler-Pickett R, et al. TAP-1 indirectly regulates CD4+ T cell priming in Toxoplasma gondii infection by controlling NK cell IFN-γ production. J Exp Med. (2007) 204:2591–602. doi: 10.1084/jem.20070634
61. Tsuchiya T, Nakao N, Yamamoto S, Hirai Y, Miyamoto K, and Tsujibo H. NK1.1 + cells regulate neutrophil migration in mice with Acinetobacter baumannii pneumonia. Microbiol Immunol. (2012) 56:107–16. doi: 10.1111/j.1348-0421.2011.00402.x
62. Gaffen SL. Structure and signalling in the IL-17 receptor family. Nat Rev Immunol. (2009) 9:556–67. doi: 10.1038/nri2586
63. O’Brien RL, Roark CL, and Born WK. IL-17-producing γδ T cells. Eur J Immunol. (2009) 39:662–6. doi: 10.1002/eji.200839120
64. Zhou Y, Xiang C, Wang N, Zhang X, Xie Y, Yang H, et al. Acinetobacter baumannii reinforces the pathogenesis by promoting IL-17 production in a mouse pneumonia model. Med Microbiol Immunol. (2023) 212:65–73. doi: 10.1007/s00430-022-00757-2
65. Lee ARYB, Wong SY, Chai LYA, Lee SC, Lee MX, Muthiah MD, et al. Efficacy of covid-19 vaccines in immunocompromised patients: systematic review and meta-analysis. BMJ. (2022) 376:e068632. doi: 10.1136/bmj-2021-068632
66. Shoham S, Batista C, Ben Amor Y, Ergonul O, Hassanain M, Hotez P, et al. Vaccines and therapeutics for immunocompromised patients with COVID-19. eClinicalMedicine. (2023) 59:101965. doi: 10.1016/j.eclinm.2023.101965
67. Gliga S, Lubke N, Killer A, Gruell H, Walker A, Dilthey AT, et al. Rapid selection of sotrovimab escape variants in severe acute respiratory syndrome coronavirus 2 omicron-infected immunocompromised patients. Clin Infect Dis. (2023) 76:408–15. doi: 10.1093/cid/ciac802
68. Weigang S, Fuchs J, Zimmer G, Schnepf D, Kern L, Beer J, et al. Within-host evolution of SARS-CoV-2 in an immunosuppressed COVID-19 patient as a source of immune escape variants. Nat Commun. (2021) 12:1–12. doi: 10.1038/s41467-021-26602-3
69. Wilkinson SAJ, Richter A, Casey A, Osman H, Mirza JD, Stockton J, et al. Recurrent SARS-CoV-2 mutations in immunodeficient patients. Virus Evol. (2022) 8:veac050. doi: 10.1093/ve/veac050
70. Borges V, Isidro J, Cunha M, Cochicho D, Martins L, Banha L, et al. Long-term evolution of SARS-coV-2 in an immunocompromised patient with non-hodgkin lymphoma. mSphere. (2021) 6:e0024421. doi: 10.1128/mSphere.00244-21
71. Greninger AL, Rybkina K, Lin MJ, Drew-Bear J, Marcink TC, Shean RC, et al. Human parainfluenza virus evolution during lung infection of immunocompromised individuals promotes viral persistence. J Clin Invest. (2021) 131:1–14. doi: 10.1172/JCI150506
72. Van Beek J, De Graaf M, Smits S, Schapendonk CME, Verjans GMGM, Vennema H, et al. Whole-genome next-generation sequencing to study within-host evolution of norovirus (NoV) among immunocompromised patients with chronic noV infection. J Infect Dis. (2017) 216:1513–24. doi: 10.1093/infdis/jix520
73. Klemm EJ, Gkrania-Klotsas E, Hadfield J, Forbester JL, Harris SR, Hale C, et al. Emergence of host-adapted Salmonella Enteritidis through rapid evolution in an immunocompromised host. Nat Microbiol. (2016) 1:15023. doi: 10.1038/nmicrobiol.2015.23
74. Luzuriaga MA, Herbert FC, Brohlin OR, Gadhvi J, Howlett T, Shahrivarkevishahi A, et al. Metal-organic framework encapsulated whole-cell vaccines enhance humoral immunity against bacterial infection. ACS Nano. (2021) 15:17426–38. doi: 10.1021/acsnano.1c03092
75. Pridmore RD. New and versatile cloning vectors with kanamycin-resistance marker. Gene. (1987) 56:309–12. doi: 10.1016/0378-1119(87)90149-1
Keywords: Acinetobacter baumannii, LAC-4 strain, pulmonary infection, whole body inactivated vaccine, immunodeficient mice, peripheral blood mononuclear cells of patients
Citation: Rouma T, Barbieux E, Bossi L, Poncin K, Denanglaire S, Tafesse Y, Vermijlen D, Delbauve S, Flamand V, Van Buylaere J, Van der Henst C, De Bolle X, Andris F and Muraille E (2025) Humoral response induced after intranasal vaccination with heat inactivated Acinetobacter baumannii protects immunodeficient mice against hypervirulent LAC-4 strain. Front. Immunol. 16:1641997. doi: 10.3389/fimmu.2025.1641997
Received: 05 June 2025; Accepted: 04 September 2025;
Published: 22 September 2025.
Edited by:
Piero Pileri, Toscana Life Sciences, ItalyReviewed by:
Pitchaipillai Sankar Ganesh, Saveetha Dental College And Hospitals, IndiaMegan Grund, Cleveland Clinic, United States
Gregory Tobin, Biological Mimetics, United States
Copyright © 2025 Rouma, Barbieux, Bossi, Poncin, Denanglaire, Tafesse, Vermijlen, Delbauve, Flamand, Van Buylaere, Van der Henst, De Bolle, Andris and Muraille. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Eric Muraille, ZW11cmFpbGxlQGhvdG1haWwuY29t
†These authors have contributed equally to this work and share first authorship
‡These authors have contributed equally to this work and share last authorship