 Amah Patricia Victorine Goran-Kouacou*‡
Amah Patricia Victorine Goran-Kouacou*‡ Oppong Richard Yéboah‡
Oppong Richard Yéboah‡ Yida Jocelyne Séri‡Aya Ursule Aniela Assi‡Adjoumanvoulé Honoré AdouSalimata MoussaBrou Doris OuraKoffi N’Guessan
Yida Jocelyne Séri‡Aya Ursule Aniela Assi‡Adjoumanvoulé Honoré AdouSalimata MoussaBrou Doris OuraKoffi N’Guessan Kouabla Liliane SiransySéry Romuald Dassé‡
Kouabla Liliane SiransySéry Romuald Dassé‡- Department of Immunology and Allergology, Faculty of Medicine, Félix Houphouët-Boigny University, Abidjan, Côte d’Ivoire
Background: Healthcare workers in resource-limited settings are frequently exposed to SARS-CoV-2, often with incomplete vaccine coverage. Yet, their adaptive immune responses remain poorly characterized in sub-Saharan Africa.
Methods: We conducted a multicenter study involving 36 healthcare workers with RT-PCR-confirmed SARS-CoV-2 infection (83.3% vaccinated) and 40 strictly unvaccinated, COVID-19-naïve controls from three university hospitals in Abidjan. Blood samples were collected on Days 0, 7, 14, 21, and 28. IgM and IgG were measured by ELFA (Mini VIDAS®), neutralizing antibodies with CHORUS TRIO®, lymphocyte subsets by flow cytometry, and Th1/Th2/Th17 cytokines using bead-based multiplex assays.
Results: Infected participants showed strong antibody production at baseline. IgG levels were closely linked to neutralizing activity (ρ = 0.83; p < 0.0001), and CD4+ T-cell counts correlated with IL-2 (ρ = 0.71; p < 0.0001). We observed early activation across Th1 (IFN-γ, TNF-α), Th2 (IL-4, IL-10), and Th17 (IL-17A) pathways. IL-17A levels were higher in asymptomatic individuals (p = 0.031). Over time, IgM and pro-inflammatory cytokines declined, while IgG remained stable and regulatory cytokines rose.
Conclusion: This cohort developed a broad immune response involving antibodies, T cells, and cytokines. The IL-17A pattern seen in asymptomatic cases may reflect effective mucosal control. These findings contribute essential data from a region where immune profiling remains limited.
1 Introduction
Since the emergence of SARS-CoV-2, healthcare workers (HCWs) have remained at high risk of infection, particularly in resource-limited settings where vaccine coverage is often incomplete (1, 2). Their repeated exposure makes them a key population for studying adaptive immune responses. Protective immunity against SARS-CoV-2 involves both humoral and cellular arms. Virus-specific antibodies, especially against the spike protein, contribute to neutralization (3), but their levels tend to decline over time (4). In parallel, CD4+ and CD8+ T-cell responses play essential roles in viral clearance and long-term protection (2, 5, 6). These responses are regulated by a complex cytokine environment that reflects T-helper polarization, including Th1 (e.g., IFN-γ, TNF-α), Th2 (e.g., IL-4, IL-10), and Th17 (e.g., IL-17A) pathways (7, 8). IL-17A, in particular, has been implicated in mucosal defense and may limit viral replication without systemic inflammation (9, 10). This cytokine pattern is especially relevant in asymptomatic cases (11, 12). In sub-Saharan Africa, immune dynamics may be shaped by region-specific factors such as exposure to diverse pathogens, chronic parasitic infections, or prior contact with endemic human coronaviruses (7). Yet despite these unique features, immunological studies in African populations remain scarce (13). To address this gap, we investigated post-infectious adaptive immunity in HCWs from Côte d’Ivoire, a population exposed to high SARS-CoV-2 circulation and heterogeneous vaccine access. We specifically analyzed the kinetics and coordination of virus-specific antibodies, lymphocyte subsets, and cytokine profiles, seeking to identify immune signatures associated with clinical presentation.
2 Materials and methods
2.1 Study design, setting, and participants
This investigation was conducted as part of a multicenter project on SARS-CoV-2 immune responses among healthcare workers in Côte d’Ivoire. The study was observational, descriptive, and longitudinal, carried out between January 2022 and June 2023 in three tertiary hospitals of Abidjan: the University Hospitals of Cocody, Treichville, and Angré. These facilities were selected for their high patient turnover and their diverse range of healthcare services, providing heterogeneous exposure conditions for healthcare staff.
The study population included medical, paramedical, and administrative personnel aged 18 years or older who provided written informed consent. Participants were stratified according to occupational exposure to SARS-CoV-2: low risk (administrative staff with no direct patient contact), intermediate risk (staff working with patients of unknown or suspected COVID-19 status), and high risk (staff with regular contact with confirmed COVID-19 cases, particularly in emergency and intensive care units).
A total of 275 participants were enrolled, including 36 SARS-CoV-2 RT-qPCR-positive cases and 40 uninfected controls followed longitudinally. All controls were unvaccinated and tested negative for SARS-CoV-2 by RT-qPCR at inclusion. Although asymptomatic infections could not be completely excluded, seronegative screening minimized potential misclassification bias. Although the cohort size was limited by logistical and financial constraints, stratification and random selection minimized recruitment bias. The modest number of confirmed cases was recognized as a potential limitation, possibly affecting statistical power and representativeness, and was explicitly considered when interpreting results.
2.2 Sampling and data collection
Baseline sampling (Day 0) consisted of venous blood collection (5 mL on EDTA and 5 mL on a dry tube) and nasopharyngeal swabbing. All samples were processed within two hours after collection. Serum and plasma were aliquoted to avoid repeated freeze-thaw cycles and stored at -80 °C until analysis.
Longitudinal follow-up was performed at Days 7, 14, 21, and 28 for RT-qPCR-positive participants and matched controls, allowing parallel evaluation of humoral, cellular, and cytokine dynamics. A standardized clinical and exposure questionnaire was administered at inclusion to capture sociodemographic characteristics, comorbidities, vaccination status, and occupational risk level.
2.3 Laboratory analyses
2.3.1 Viral RNA detection
Nasopharyngeal swabs placed in viral transport medium were analyzed for SARS-CoV-2 RNA using the KingFisher™ Duo Prime system (Thermo Fisher Scientific, Waltham, MA, USA) with the MagMAX™ Viral/Pathogen kit for automated extraction. Amplification was performed on a CFX96™ Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA) targeting the nucleocapsid (N) and RNA-dependent RNA polymerase (RdRp) genes. A cycle threshold (Ct) < 35 defined positivity. Each run included an internal control (cellular RNA), a certified positive control, and a negative control. Cycle threshold values were used qualitatively for case definition and not for viral load quantification.
2.3.2 Serological assays (IgM and IgG anti-RBD)
Detection of antibodies directed against the receptor-binding domain (RBD) of the SARS-CoV-2 Spike protein was performed on the Mini-VIDAS® analyzer (bioMérieux SA, Marcy-l’Étoile, France; Serial No. IVD7006414, Ref. 410417) using the VIDAS® SARS-CoV-2 IgM and VIDAS® SARS-CoV-2 IgG II kits. The assays rely on enzyme-linked fluorescent assay (ELFA) technology providing qualitative and semi-quantitative detection of specific antibodies.
IgM and IgG results were first obtained as index values (ratio of the sample Relative Fluorescence Value [RFV] to that of the calibrator), with an index ≥ 1.0 considered positive. For quantitative comparison, index values were converted to Binding Antibody Units per milliliter (BAU/mL) using the manufacturer-derived conversion factor 1 index = 20.33 BAU/mL, standardized to the 1st WHO International Standard (20/136). Accordingly, antibody titers ≥ 20 BAU/mL indicated seropositivity, and ≥ 250 BAU/mL represented a strong antibody response.
2.3.3 Neutralizing antibody assay
Neutralizing antibodies against the S1 subunit of the Spike protein were quantified using the CHORUS TRIO® semi-automated immunoanalytical system (Diesse Diagnostica Senese S.p.A., Siena, Italy; Serial No. 4341, P/N 81200) and the CHORUS SARS-CoV-2 “Neutralizing” Ab kit. The method is a competitive enzyme immunoassay in which antibodies in the sample compete with a labeled tracer for binding to the viral RBD/ACE2 complex. The degree of inhibition reflects neutralizing activity and is expressed in BAU/mL relative to the WHO International Standard (20/136). Results were classified as negative (< 20 BAU/mL), equivocal (20-49.9 BAU/mL), or positive (≥ 50 BAU/mL). Each analytical series included a stored master calibration curve, a certified positive control, and an internal negative control to ensure traceability and inter-assay reproducibility.
2.3.4 T-cell immunophenotyping
T-lymphocyte subsets were quantified on a BD FACSCanto™ II flow cytometer (BD Biosciences, San Jose, CA, USA; Serial No. V3389002039) equipped with 488 nm and 633 nm lasers. Whole blood (50 µL, EDTA-treated) was stained with monoclonal antibodies anti-CD3 (UCHT1-FITC, Ref. 555332), anti-CD4 (RPA-T4-APC, Ref. 555349), anti-CD8 (RPA-T8-PE, Ref. 555635), and anti-CD45 (2D1-PerCP-Cy5.5, Ref. 564105). After a 20-min incubation at room temperature, erythrocytes were lysed with BD FACS™ Lysing Solution and washed in PBS. Acquisition included ≥ 10,000 lymphocyte-gated events. Data were analyzed using FACSDiva™ v8.0 software (BD Biosciences). Absolute lymphocyte subset counts were derived by combining flow cytometry percentages with total lymphocyte counts obtained from concomitant full blood counts and expressed as cells/µL.
2.3.5 Cytokine profiling
Cytokines were quantified in serum using the BD™ CBA Human Th1/Th2/Th17 Cytokine Kit (Ref. 560484, BD Biosciences). The assay allows multiplex quantification of IL-2, IL-4, IL-6, IL-10, TNF-α, IFN-γ, and IL-17A by flow cytometry. Serum samples were centrifuged at 3,500 rpm for 5 min, heat-inactivated at 56 °C for 30 min, and diluted 1:4 before analysis. Fifty microliters of cytokine-capture beads were incubated with standards or samples and PE-labeled detection reagent for 3 h in the dark. Data were acquired on the BD FACSCanto™ II and analyzed using FCAP Array™ v3.0 software. Detection ranges (manufacturer data, 2022) were IL-2 (2-5,000 pg/mL), IL-4 (2-5,000), IL-6 (2.5-5,000), IL-10 (2.7-2,000), TNF-α (3.7-2,000), IFN-γ (3-5,000), and IL-17A (2-5,000 pg/mL).
2.4 Variables and data management
Variables included demographic characteristics, comorbidities, vaccination status, and occupational exposure level. Biological parameters included IgM and IgG anti-RBD, neutralizing antibodies, lymphocyte subsets, and cytokine concentrations. Data were double-entered into Microsoft Excel 2016, cross-checked for inconsistencies, and exported to SPSS v26.0 for analysis. All records were pseudonymized, and quality control ensured internal validity before database locking.
2.5 Statistical analysis
Data were analyzed using SPSS v26.0 (IBM Corp., Armonk, NY, USA). Quantitative variables were expressed as mean ± standard deviation (SD) or median (interquartile range, IQR), depending on distribution. Categorical variables were expressed as frequencies and percentages. Correlations between antibody titers (IgG, NAbs) and cytokine levels (IL-2, IFN-γ, TNF-α) were assessed using Spearman’s rank correlation coefficient (ρ). Group comparisons were performed using the Mann-Whitney U test for continuous variables and the Chi-square or Fisher’s exact test for categorical variables. Statistical significance was set at p < 0.05.
2.6 Ethical considerations
The study protocol was approved by the National Ethics Committee for Life Sciences and Health (CNESVS) of Côte d’Ivoire (Ref. 007-22/MSHP/CMU/CNESVS-km). All participants provided written informed consent prior to enrollment, and all procedures complied with the Declaration of Helsinki (2013) and national regulations on biomedical research ethics.
3 Results
3.1 Baseline characteristics of participants
Participant demographics and occupational data are summarized in Table 1. Cases and controls were comparable in median age (39.0 vs. 37.0 years), female proportion (63.9% vs. 67.5%), and median BMI (24.7 vs. 25.9 kg/m²). In contrast, hospital affiliation and occupational exposure level differed, with a higher proportion of high-risk exposure among cases (36.1% vs. 12.5%). All controls were unvaccinated, while 83.3% of cases had received at least one dose of a SARS-CoV-2 vaccine.

Table 1. Characteristics of participants.
3.2 Immune profiles at inclusion: cases vs. controls
At Day 0, cases had significantly higher levels of IgM, IgG, and neutralizing antibodies than controls (36.4 vs. 4.3 BAU/mL, 556.3 vs. 272.2 BAU/mL, and 1469.6 vs. 921.8 BAU/mL, respectively; p < 0.0001 for IgM and IgG, p = 0.0017 for NAbs; Table 2). CD4+ and total CD3+ T cell counts were also higher in cases (2368.4 vs. 1560.8 cells/µL and 3525.4 vs. 2949.0 cells/µL; p = 0.0054 and p = 0.0463), with no significant difference for CD8+ (p = 0.2418). Cytokine concentrations of IL-10, IFN-γ, TNF-α, IL-2, IL-4, and IL-17A were significantly higher in cases (p < 0.01 for all), while IL-6 showed no significant difference (14.9 vs. 18.8 pg/mL; p = 0.1988; Table 2).
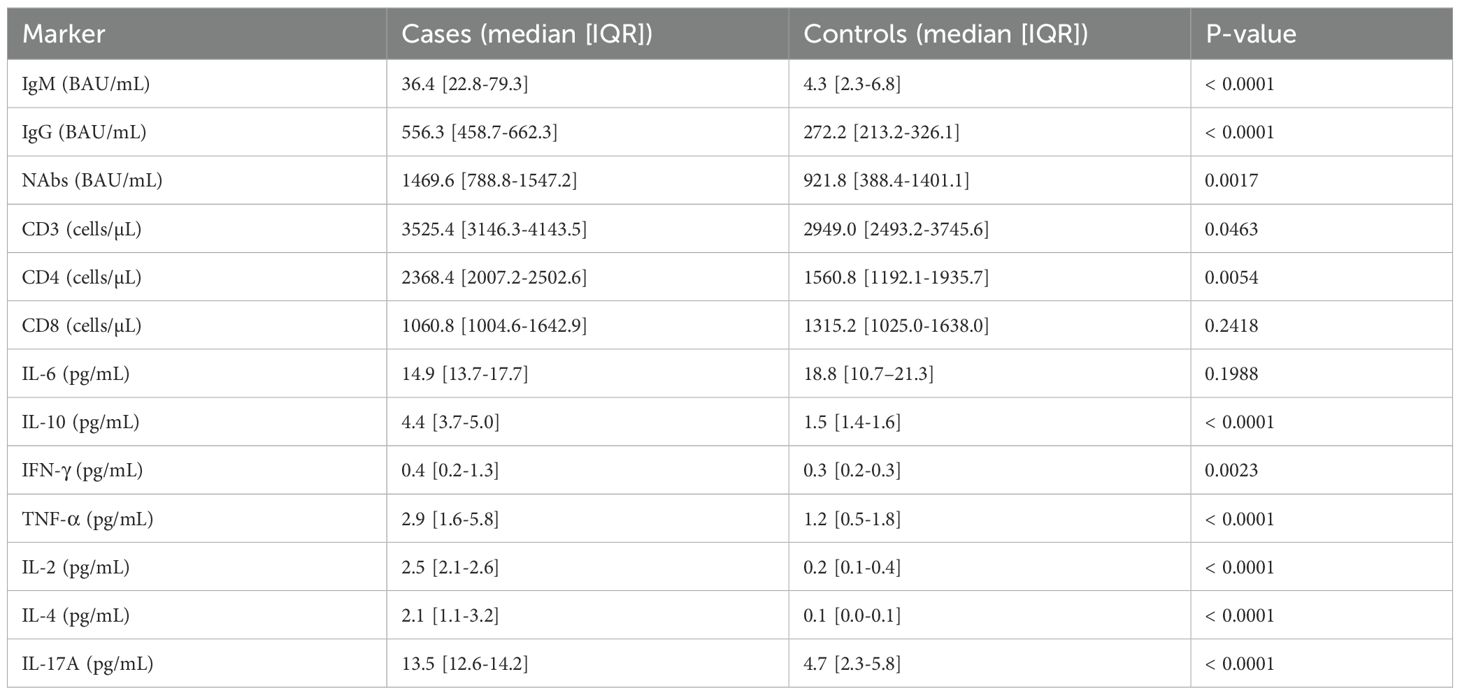
Table 2. Comparison of immune markers at day 0 between cases and controls.
3.3 Comparison by symptom status at inclusion
Among infected individuals, most immune markers showed no significant differences between symptomatic and asymptomatic subgroups. No statistical differences were observed for IgM, IgG, NAbs, CD4+, CD8+, IL-2, IL-4, IL-6, IL-10, IFN-γ, or TNF-α. Only IL-17A was significantly higher in asymptomatic individuals (13.8 vs. 12.9 pg/mL; p = 0.0311; Table 3).
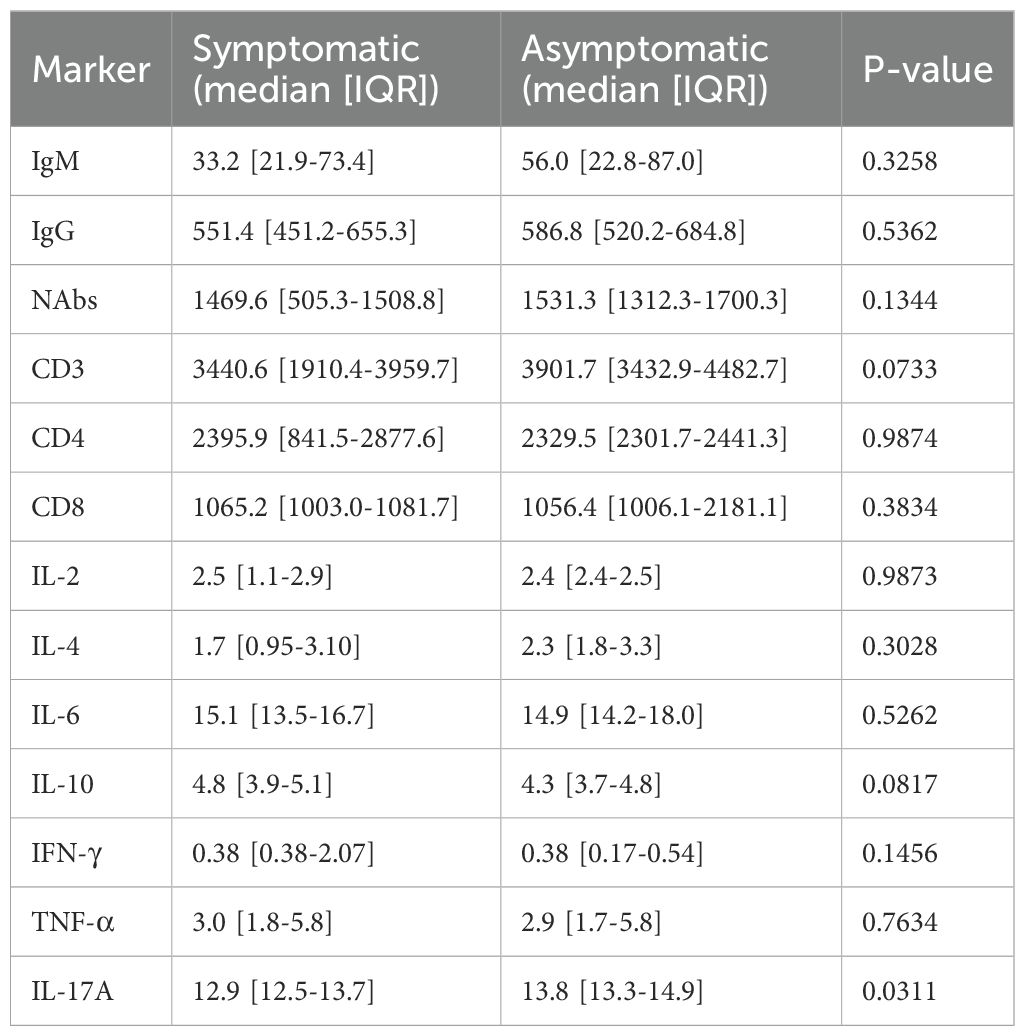
Table 3. Comparison of immune and cytokine markers at day 0 between symptomatic and asymptomatic cases.
3.4 Immune correlations at inclusion in cases
A strong positive correlation was observed between IgG and neutralizing antibodies (ρ = 0.83; p < 0.0001). Other significant correlations included CD4+ with IL-2 (ρ = 0.71; p < 0.0001), CD4+ with IL-10 (ρ = 0.46; p = 0.005), and CD4+ with TNF-α (ρ = 0.41; p = 0.013). IL-6 also correlated with both IL-10 and TNF-α, and a strong correlation was noted between IL-2 and IL-4 (ρ = 0.62; p < 0.0001; Table 4).
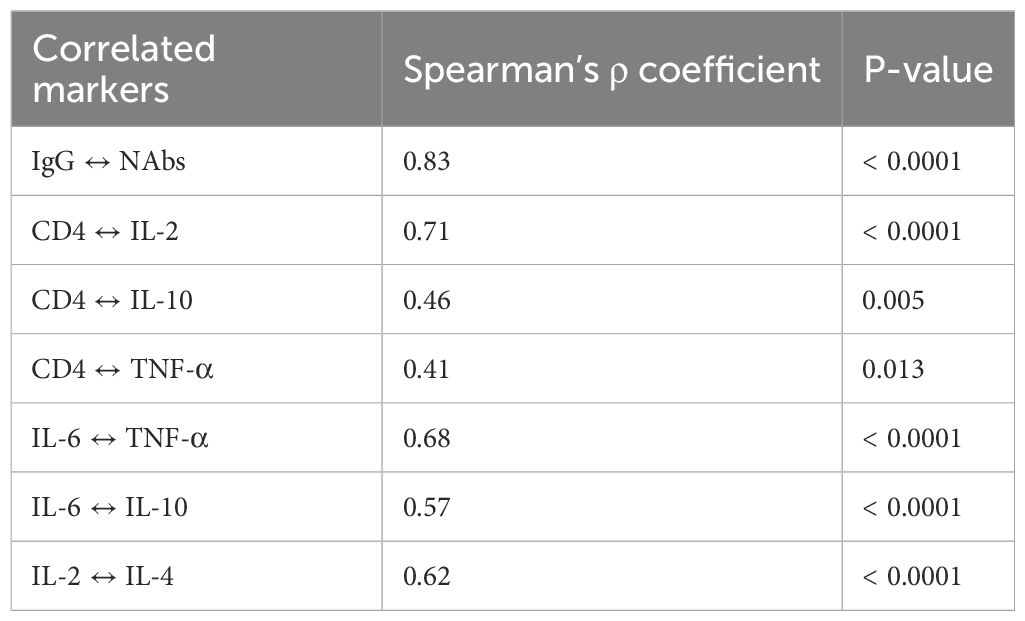
Table 4. Correlations between immune markers in positive cases at day 0.
3.5 Longitudinal evolution of immune markers in cases
IgM levels declined steadily from 36.4 to 2.3 BAU/mL between Day 0 and Day 28. IgG levels increased from 556.3 to 615.6 BAU/mL. Neutralizing antibodies decreased between Day 0 (1469.6 BAU/mL) and Day 14 (1172.6 BAU/mL), rose slightly at Day 21 (1250.2 BAU/mL), then dropped again at Day 28 (1186.1 BAU/mL; Figure 1). Total CD3+ T cells increased from 4868.5 to 6262.5 cells/μL. CD4+ T cells rose nonlinearly from 2395.9 to 3560.1 cells/μL between Day 0 and Day 14, dipped to 3011.3 at Day 21, and rose again to 3380.1 at Day 28. CD8+ T cells remained relatively stable between 999.9 and 1225.1 cells/μL (Figure 2). Pro-inflammatory cytokines such as IL-6 and TNF-α declined over time (IL-6: 14.9 to 9.6 pg/mL; TNF-α: 2.9 to 1.4 pg/mL), while regulatory cytokines IL-10, IL-2, and IL-4 showed modest increases from Day 7 to Day 28 (Figure 3).
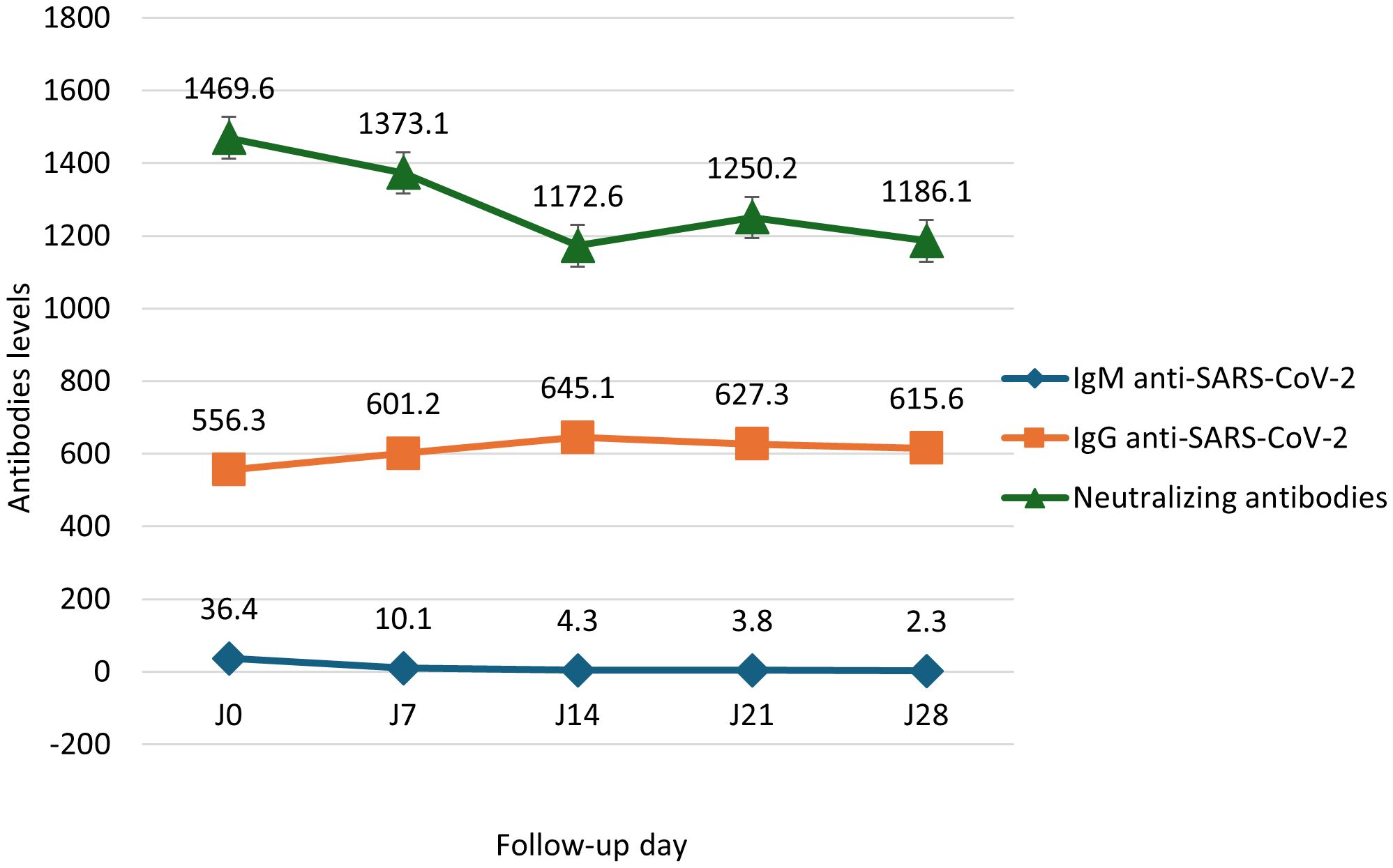
Figure 1. Median evolution of SARS-CoV-2–specific antibodies in positive cases (Day 0 to Day 28). Kinetics of SARS-CoV-2-specific antibody responses in COVID-19 patients. Levels of anti-SARS-CoV-2 IgM, IgG, and neutralizing antibodies were measured on days 0, 7, 14, 21, and 28 post-infection. IgM, IgG, and neutralizing antibody titers are expressed in Binding Antibody Units per milliliter (BAU/mL). Data are presented as medians with interquartile ranges (IQR); error bars represent the IQR.
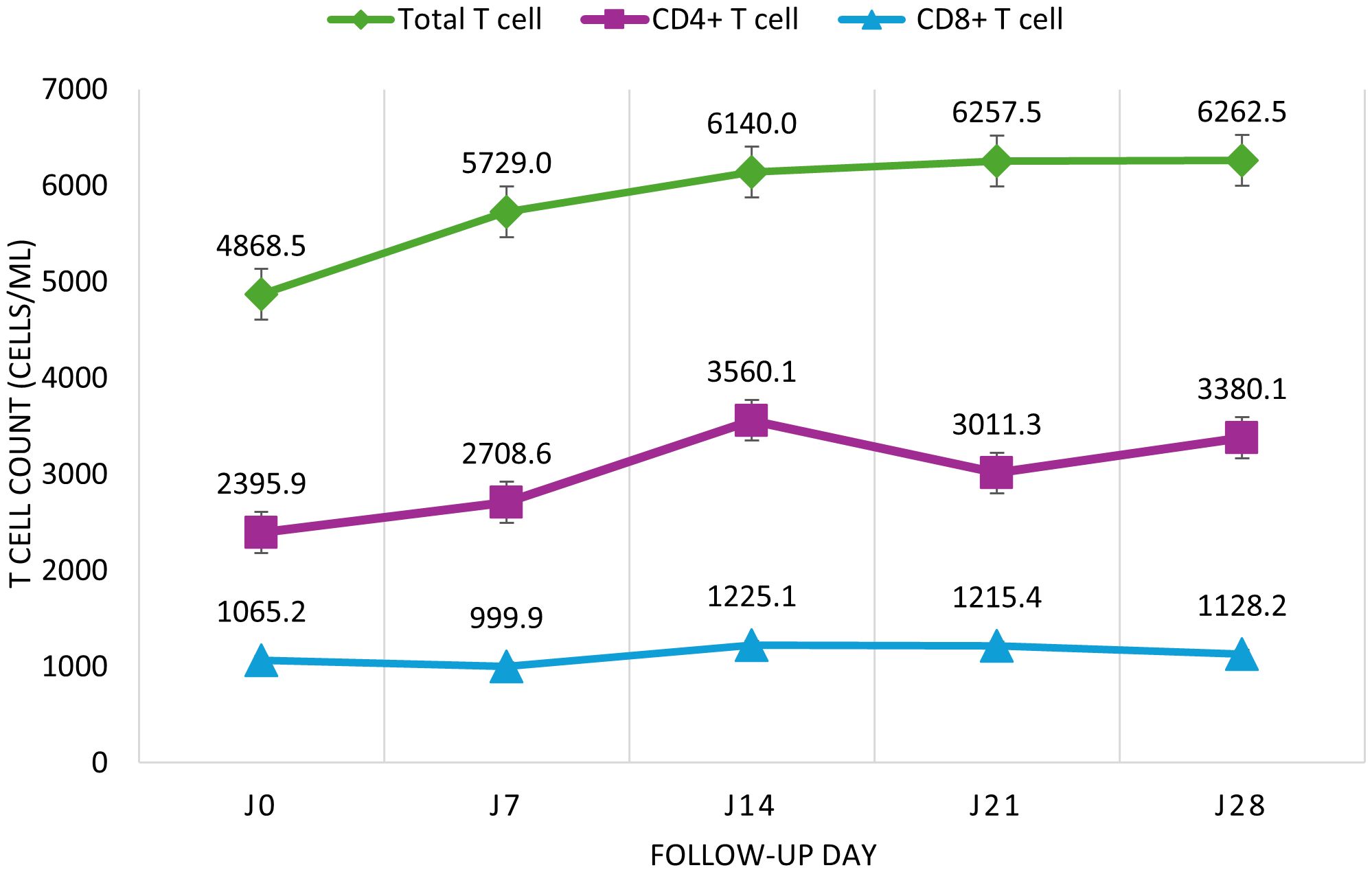
Figure 2. Median evolution of CD4+ and CD8+ T lymphocytes in COVID-19 cases from Day 0 to Day 28. Longitudinal evolution of T lymphocyte subpopulations in COVID-19 patients. Total T lymphocytes and CD4+/CD8+ subsets were quantified on days 0, 7, 14, 21, and 28 by flow cytometry. Cell counts are expressed in cells/µL. Data are presented as medians with interquartile ranges (IQR); error bars represent the IQR.
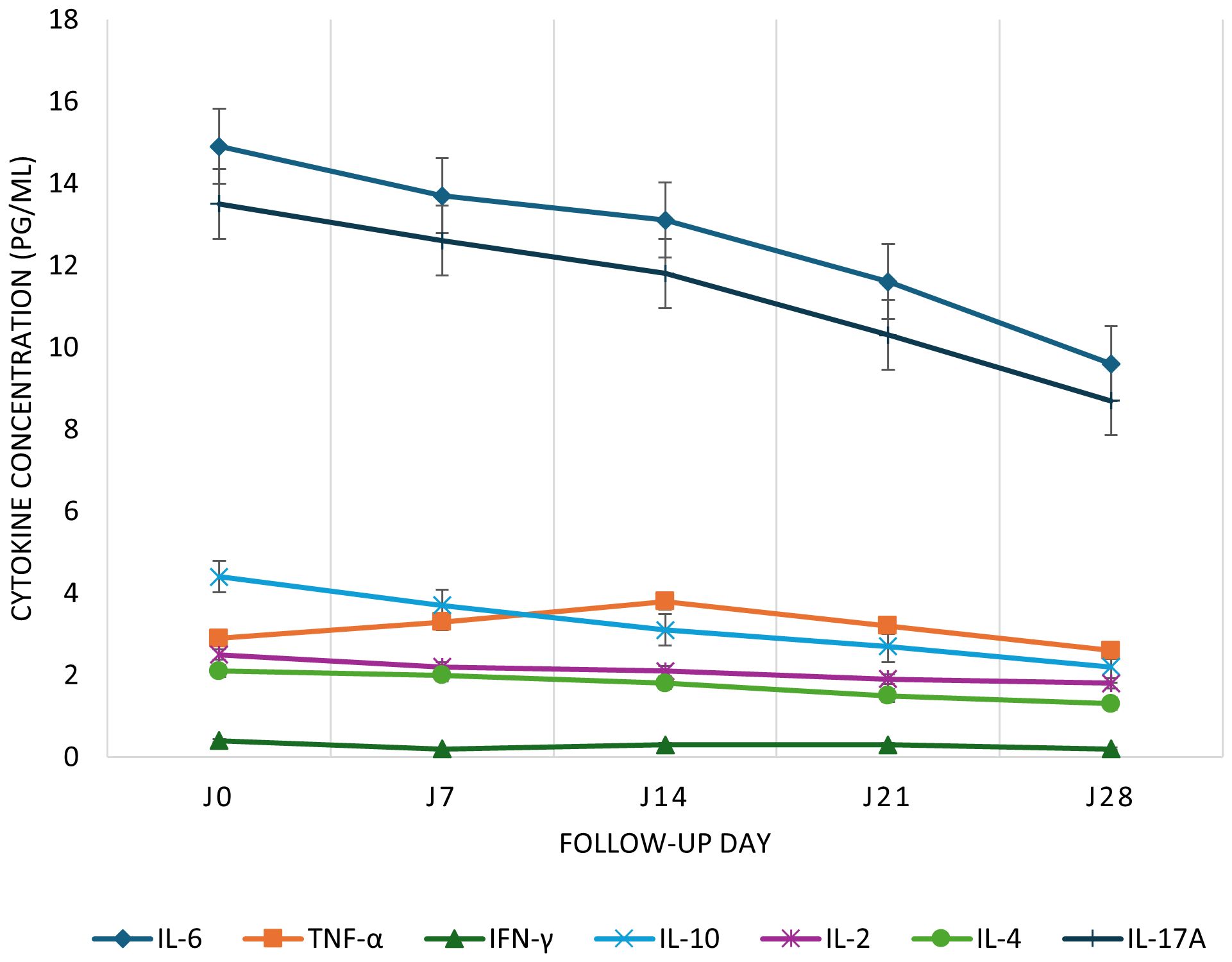
Figure 3. Kinetics of pro- and anti-inflammatory cytokines in COVID-19 cases from Day 0 to Day 28. Time course of plasma cytokine concentrations in SARS-CoV-2-infected patients. Plasma levels of IL-6, TNF-α, IFN-γ, IL-10, IL-2, IL-4, and IL-17A were measured at five time points during follow-up. Cytokine concentrations are expressed in pg/mL. Data are presented as medians with interquartile ranges (IQR); error bars represent the IQR.
4 Discussion
4.1 Humoral and neutralizing response
The observed humoral response is characterized by the production of specific IgG antibodies alongside neutralizing antibodies capable of preventing viral entry. In line with current knowledge, we observed a significant increase in antiviral IgG and detectable neutralizing activity from the second week post-infection (2, 14, 15). Neutralizing titers rose in parallel with IgG levels, peaking around week 4 (2, 14). This synchrony suggests that specific IgG directly contributes to viral neutralization, consistent with their documented role in viral clearance by blocking virus-cell binding (3, 4). We also demonstrated a positive correlation between IgG concentrations and neutralizing capacity, highlighting the importance of antibody quality in antiviral protection (4). This IgG-neutralizing antibody link has also been reported elsewhere, with both parameters rising in tandem about three weeks post-infection (14). Altogether, these findings confirm that high levels of specific IgG are closely associated with strong neutralizing activity, indicative of a robust and potentially protective humoral response. This includes some unvaccinated controls with IgG and neutralizing titers above threshold, likely due to undetected exposure or endemic coronavirus cross-reactivity.
4.2 Dominant CD4+ T Cell response and role of IL-2
The cellular arm of the adaptive immune response in our study is dominated by CD4+ T lymphocytes, with a more modest contribution from CD8+ T cells. CD4+ T cells were notable for their high IL-2 production, a cytokine essential for lymphocyte expansion and immune coordination. IL-2, mainly secreted by activated CD4+ helper T cells, plays a central role in promoting T and B cell proliferation and supporting NK cell activation (16). This T helper cytokine thus facilitates the coordinated rise of multi-compartment immunity. The observed strong correlation between activated CD4+ T cell frequency and IL-2 levels suggests a primarily Th1-type cellular response, focused on assisting other immune compartments. This Th1/IL-2-driven profile is frequently associated with effective immunity: in COVID-19, convalescent patients with favorable outcomes exhibit polyfunctional CD4+ T cells secreting IL-2, IFN-γ, and TNF-α (5, 11, 12), whereas severe cases often show loss of this functionality (17). Therefore, the predominance of IL-2-producing CD4+ T cells in our cohort may reflect a robust immune profile capable of supporting antibody production (via B cell help) and orchestrating the activation of other immune effectors.
4.3 Mixed cytokine profile (Th1/Th2/Th17)
Circulating cytokine measurements revealed a heterogeneous immune profile combining Th1, Th2, and Th17 features. In addition to canonical Th1 cytokines indicative of antiviral cellular responses (e.g., IFN-γ, IL-2, TNF-α), we simultaneously detected Th2-type cytokines (e.g., IL-4, IL-10) associated with humoral immunity and anti-inflammatory regulation, as well as the Th17 cytokine IL-17A. This combination suggests a non-polarized, multipronged T helper response. In SARS-CoV-2 infection, the virus has been reported to induce both proinflammatory Th1 and Th17 responses that may contribute to immunopathogenesis, while compensatory Th2 signaling may help mitigate excessive inflammation (18–21). The detection of IL-10 alongside IL-6 and TNF-α is consistent with previous reports highlighting the dual pro- and anti-inflammatory cytokine environment in moderate and severe COVID-19 (18) and reflects the regulatory role of IL-10 family cytokines in maintaining immune homeostasis (19). Our findings are consistent with such concurrent activation of multiple cytokine pathways. A mixed Th1/Th2/Th17 profile likely reflects the immune system’s effort to engage diverse arms of adaptive immunity to control infection while avoiding uncontrolled inflammation.
4.4 Specific role of IL-17A in asymptomatic individuals
A key finding of our study is the distinctive role of IL-17A in asymptomatic individuals. While IL-17A has been implicated in severe COVID-19 cases by fueling inflammation and cytokine storms (22, 23), our data suggest a different contribution in asymptomatic subjects. These individuals showed moderate but significant IL-17A production in the absence of clinical symptoms, implying a protective rather than pathogenic function. Recent studies have shown that some IL-17 family cytokines, such as epithelial-derived IL-17C, can locally boost barrier defenses and induce IL-17A expression to enhance innate immunity while limiting symptoms (24). Moreover, IL-17A, though identified as a marker of severe disease, is also induced during mild infections and remains elevated for up to 4 weeks post-infection (20, 24). It is therefore plausible that in asymptomatic individuals, a moderate and controlled Th17 response, marked by IL-17A, contributes to effective viral clearance (via neutrophil recruitment and mucosal barrier reinforcement) without causing tissue damage. This mechanistic interpretation aligns with the notion of infection tolerance seen in asymptomatic cases: these individuals may achieve viral control through efficient antiviral immunity while avoiding harmful inflammatory escalation (21, 24).
4.5 Immune kinetics over 28 days
Dynamic analysis of immune responses over 28 days revealed a well-coordinated sequence of phases. In the early days post-infection, innate immunity predominates, characterized by increased levels of inflammatory cytokines (e.g., IL-6 and TNF-α) in response to initial viral replication (1, 8). This acute phase is followed, from the second week onward, by the onset of specific adaptive immunity. Our data show that IgM and then IgG antibodies emerge during this period, with neutralizing activity becoming clearly detectable by 10–14 days post-infection (2, 14). Neutralizing titers, initially low in the first week, subsequently rise rapidly in parallel with specific IgG and typically peak around week 4 (2, 14, 15). Simultaneously, T cell responses emerge early: antigen-specific T cells can be detected by the end of the first week, with peak expansion occurring 1 to 2 weeks after infection (6, 25, 26). In our cohort, IL-2 and IFN-γ production by CD4+ T cells indicated strong cellular activation by the third week, suggesting that these cells reached maximal functionality at this stage. Finally, some cytokines such as IL-17A followed a later trajectory, with levels already elevated during the acute phase continuing to rise and remaining high through day 28 (24). This sequential pattern is consistent with longitudinal studies of COVID-19 describing a shift from early innate to robust adaptive immunity peaking between weeks 3 and 5 (2, 6, 25). In sum, our findings highlight an immune trajectory in which early innate responses prepare the ground, followed by humoral and cellular adaptive immunity reaching maximal strength by the end of the first month.
4.6 Cross-compartment correlations
Cross-analysis of immune compartments revealed significant correlations, underscoring coordinated orchestration of the anti-infective response. A strong correlation was observed between specific IgG levels and neutralizing activity: individuals with the highest IgG titers also showed the strongest neutralization capacity (4). This IgG-neutralizing antibody link confirms that humoral response quality (affinity and quantity of antibodies) largely determines neutralization efficacy. We also found a positive association between CD4+ T cell response magnitude and IL-2 production, consistent with the central role of helper T cells in IL-2 secretion to support cellular immunity (5, 16). In other words, stronger CD4+ T responses correspond to higher IL-2 availability, potentially amplifying effector lymphocyte expansion and antibody-secreting plasma cell generation. Furthermore, IL-6 and TNF-α levels were highly correlated, reflecting synchronous innate immune activation. These two mediators are often co-elevated in acute viral infections and act synergistically in inflammatory syndromes (1, 8). The observed IL-6-TNF correlation likely reflects shared upstream activation (e.g., via monocyte/macrophage pathways), a pattern frequently associated with more severe COVID-19 forms (27). Overall, these cross-compartment correlations emphasize that humoral, cellular, and cytokine responses do not operate in isolation but rather interact dynamically. Effective T cell responses facilitate antibody maturation (via CD4+ B cell help and IL-2 production), while innate inflammation modulates the context in which adaptive immunity unfolds. This multi-compartment coordination is key to a balanced and protective immune response.
4.7 Study limitations
This study has several limitations. First, the sample size was limited, which may reduce statistical power and the generalizability of findings. Second, the 28-day follow-up precluded assessment of longer-term immune durability, such as antibody titers beyond the first month or memory T cell persistence. Third, our analysis focused on selected immune compartments (IgG antibodies, CD4+ T cells, circulating cytokines) and did not assess potentially relevant actors such as cytotoxic CD8+ T cells, local innate immunity at infection sites, or mucosal IgA responses. These unmeasured elements could provide additional insights and merit future investigation. Finally, as an observational study centered on paucisymptomatic and asymptomatic cases, the reported correlations do not imply causality and may not be extrapolated to severe disease. Despite these limitations, our findings offer an integrated overview of the anti–SARS-CoV-2 immune response and pave the way for further investigations into mechanisms underpinning symptom-free infection and efficient viral resolution.
5 Conclusion
The immune responses observed in SARS-CoV-2-infected healthcare workers revealed a well-coordinated activation across humoral, cellular, and cytokine compartments. Sustained IgG and neutralizing antibody production, coupled with CD4+ T-cell expansion and cytokine modulation, reflected an adaptive response capable of controlling viral replication while minimizing inflammation. The immune profile of asymptomatic individuals suggests that effective viral clearance may occur through balanced, non-inflammatory mechanisms, possibly involving mucosal or Th17-mediated pathways. The detection of IgG and neutralizing antibodies in some uninfected controls likely reflects prior asymptomatic exposure or cross-reactive immunity. These findings provide valuable insights for improving immune surveillance and epidemic preparedness in African populations, while the limited cohort size underscores the need for larger longitudinal studies to confirm these observations.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by National Health Research Ethics Committee of Côte d’Ivoire. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AG-K: Conceptualization, Formal Analysis, Investigation, Methodology, Writing – original draft. OY: Data curation, Validation, Writing – review & editing. YS: Data curation, Validation, Writing – review & editing. AUAA: Data curation, Investigation, Resources, Writing – review & editing. AHA: Data curation, Investigation, Resources, Writing – review & editing. SM: Data curation, Investigation, Resources, Writing – review & editing. BO: Data curation, Investigation, Resources, Writing – review & editing. KS: Supervision, Writing – review & editing. KN’G: Supervision, Writing – review & editing. SRD: Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was supported by the Fonds pour la Science, la Technologie et l’Innovation (FONSTI), Côte d’Ivoire, as part of the national research program on COVID-19.
Acknowledgments
The authors thank the general management of the University Hospitals of Cocody, Treichville, and Angré for their institutional support, as well as all the healthcare professionals who generously agreed to participate in the study. Special thanks are extended to the teams of the Immunology Laboratory at the University Hospital of Cocody and the Faculty of Medicine of Abidjan for their technical assistance. This research was supported by the Fonds pour la Science, la Technologie et l’Innovation (FONSTI), Côte d’Ivoire, under the national COVID-19 research initiative.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Author disclaimer
The content of this publication is solely the responsibility of the authors and does not necessarily represent the official views of the funding body.
References
1. Arunachalam PS, Wimmers F, Mok CKP, Perera RAPM, Scott M, Hagan T, et al. Systems biological assessment of immunity to mild versus severe COVID-19 infection in humans. Science. (2020) 369:1210–20. doi: 10.1126/science.abc6261
2. Dan JM, Mateus J, Kato Y, Hastie KM, Yu ED, Faliti CE, et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science. (2021) 371:eabf4063. doi: 10.1126/science.abf4063
3. Premkumar L, Segovia-Chumbez B, Jadi R, Martinez DR, Raut R, Markmann A, et al. The receptor binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV-2 patients. Sci Immunol. (2020) 5:eabc8413. doi: 10.1126/sciimmunol.abc8413
4. Khoury DS, Cromer D, Reynaldi A, Schlub TE, Wheatley AK, Juno JA, et al. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat Med. (2021) 27:1205–11. doi: 10.1038/s41591-021-01377-8
5. Sette A and Crotty S. Adaptive immunity to SARS-coV-2 and COVID-19. Cell. (2021) 184:861–80. doi: 10.1016/j.cell.2021.01.007
6. Zuo J, Dowell AC, Pearce H, Verma K, Long HM, Begum J, et al. Robust SARS-CoV-2-specific T cell immunity is maintained at 6 months following primary infection. Nat Immunol. (2021) 22:620–26. doi: 10.1038/s41590-021-00902-8
7. Mathew D, Giles JR, Baxter AE, Oldridge DA, Greenplate AR, Wu JE, et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science. (2020) 369:eabc8511. doi: 10.1126/science.abc8511
8. Lucas C, Wong P, Klein J, Castro TBR, Silva J, Sundaram M, et al. Longitudinal analyses reveal immunological misfiring in severe COVID-19. Nature. (2020) 584:463–69. doi: 10.1038/s41586-020-2588-y
9. Liu Y, Zhang C, Huang F, Yang Y, Wang F, Yuan J, et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Natl Sci Rev. (2020) 7:1003–11. doi: 10.1093/nsr/nwaa037
10. Zhang Q, Bastard P, Liu Z, Le Pen J, Moncada-Velez M, Chen J, et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science. (2020) 370:eabd4570. doi: 10.1126/science.abd4570
11. Sekine T, Perez-Potti A, Rivera-Ballesteros O, Strålin K, Gorin JB, Olsson A, et al. Robust T cell immunity in convalescent individuals with asymptomatic or mild COVID-19. Cell. (2020) 183:158–68.e14. doi: 10.1016/j.cell.2020.08.017
12. Neidleman J, Luo X, Frouard J, Xie G, Gill G, Stein ES, et al. SARS-CoV-2-specific T cells exhibit phenotypic features of helper function, lack of terminal differentiation, and high proliferation potential. Cell Rep Med. (2020) 1:100081. doi: 10.1016/j.xcrm.2020.100081
13. Cose S, Bagaya B, Nerima B, Joloba M, Kambugu A, Tweyongyere R, et al. Immunology in Africa. Trop Med Int Health. (2015) 20:1771–77. doi: 10.1111/tmi.12599
14. Gaebler C, Wang Z, Lorenzi JCC, Muecksch F, Finkin S, Tokuyama M, et al. Evolution of antibody immunity to SARS-CoV-2. Nature. (2021) 591:639–44. doi: 10.1038/s41586-021-03207-w
15. Rodda LB, Netland J, Shehata L, Pruner KB, Morawski PA, Thouvenel CD, et al. Functional SARS-CoV-2-specific immune memory persists after mild COVID-19. Cell. (2021) 184:169–83.e17. doi: 10.1016/j.cell.2020.11.029
16. Boyman O and Sprent J. The role of interleukin-2 during homeostasis and activation of the immune system. Nat Rev Immunol. (2012) 12:180–90. doi: 10.1038/nri3156
17. Meckiff BJ, Ramírez-Suástegui C, Fajardo V, Chee SJ, Kusnadi A, Simon H, et al. Imbalance of regulatory and cytotoxic SARS-CoV-2-reactive CD4+ T cells in COVID-19. Cell. (2020) 183:1340–53.e16. doi: 10.1016/j.cell.2020.10.001
18. Chen G, Wu D, Guo W, Cao Y, Huang D, Wang H, et al. Clinical and immunological features of severe and moderate coronavirus disease 2019. J Clin Invest. (2020) 130:2620–29. doi: 10.1172/JCI137244
19. Ouyang W, Rutz S, Crellin NK, Valdez PA, and Hymowitz SG. Regulation and functions of the IL-10 family of cytokines in inflammation and disease. Annu Rev Immunol. (2011) 29:71–109. doi: 10.1146/annurev-immunol-031210-101312
20. Zhu Q, Xu Y, Wang T, and Xie F. Innate and adaptive immune response in SARS-CoV-2 infection-current perspectives. Front Immunol. (2022) 13:1053437. doi: 10.3389/fimmu.2022.1053437
21. Bhardwaj A, Sapra L, Saini C, Azam Z, Mishra PK, Verma B, et al. COVID-19: immunology, immunopathogenesis and potential therapies. Int Rev Immunol. (2022) 41:171–206. doi: 10.1080/08830185.2021.1883600
22. Del Valle DM, Kim-Schulze S, Huang HH, Beckmann ND, Nirenberg S, Wang B, et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat Med. (2020) 26:1636–43. doi: 10.1038/s41591-020-1051-9
23. Tang Y, Liu J, Zhang D, Xu Z, Ji J, and Wen C. Cytokine storm in COVID-19: the current evidence and treatment strategies. Front Immunol. (2020) 11:1708. doi: 10.3389/fimmu.2020.01708
24. Tjan LH, Furukawa K, Nagano T, Kiriu T, Nishimura M, Arii J, et al. Early differences in cytokine production by severity of coronavirus disease 2019. J Infect Dis. (2021) 223:1145–49. doi: 10.1093/infdis/jiab005
25. Rydyznski Moderbacher C, Ramirez SI, Dan JM, Grifoni A, Hastie KM, Weiskopf D, et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell. (2020) 183:996–1012.e19. doi: 10.1016/j.cell.2020.09.038
26. Peng Y, Mentzer AJ, Liu G, Yao X, Yin Z, Dong D, et al. Broad and strong memory CD4+ and CD8+ T cells induced by SARS-CoV-2 in UK convalescent individuals following COVID-19. Nat Immunol. (2020) 21:1336–45. doi: 10.1038/s41590-020-0782-6
Keywords: SARS-CoV-2, COVID-19, healthcare workers, humoral immunity, T-cell response, cytokines, neutralizing antibodies, sub-Saharan Africa
Citation: Goran-Kouacou APV, Yéboah OR, Séri YJ, Assi AUA, Adou AH, Moussa S, Oura BD, N’Guessan K, Siransy KL and Dassé SR (2025) Coordinated kinetics of humoral, T-cell, and cytokine responses to SARS-COV-2 in West African healthcare workers: a multicenter longitudinal study in university hospitals of Abidjan, Côte d’Ivoire. Front. Immunol. 16:1669517. doi: 10.3389/fimmu.2025.1669517
Received: 19 July 2025; Accepted: 03 November 2025;
Published: 21 November 2025.
Edited by:
Jingxin Li, Jiangsu Provincial Center for Disease Control and Prevention, ChinaReviewed by:
Pawel Zmora, Polish Academy of Sciences, PolandYuxin Chen, Nanjing Drum Tower Hospital Clinical College of Nanjing Medical University, China
Copyright © 2025 Goran-Kouacou, Yéboah, Séri, Assi, Adou, Moussa, Oura, N’Guessan, Siransy and Dassé. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amah Patricia Victorine Goran-Kouacou, a291YWNvdS5hbWFoQGdtYWlsLmNvbQ==
‡ORCID: Amah Patricia Victorine Goran-Kouacou, orcid.org/0009-0006-7767-8022
Oppong Richard Yéboah, orcid.org/0009-0004-4555-2148
Yida Jocelyne Séri, orcid.org/0009-0007-2088-1158
Aya Ursule Aniela Assi, orcid.org/0009-0000-4802-1888
Séry Romuald Dassé, orcid.org/0009-0006-4898-8830