 Javier Carbone-Schellman1,2†
Javier Carbone-Schellman1,2† Javiera Fontecilla-Escobar1,3†
Javiera Fontecilla-Escobar1,3† Nicolás Sales-Salinas1William F. Chaparro-Pico4
Nicolás Sales-Salinas1William F. Chaparro-Pico4 Alfredo Molina-Berríos5
Alfredo Molina-Berríos5 María Celeste Ruete3Pablo A. González2,6
María Celeste Ruete3Pablo A. González2,6 Mayra A. Machuca4
Mayra A. Machuca4 Ma. Cecilia Opazo7,8
Ma. Cecilia Opazo7,8 Marcelo E. Ezquer1
Marcelo E. Ezquer1 Luisa F. Duarte1,2*
Luisa F. Duarte1,2*- 1Centro de Medicina Regenerativa, Facultad de Medicina, Clínica Alemana – Universidad del Desarrollo, Santiago, Chile
- 2Millennium Institute on Immunology and Immunotherapy, Santiago, Chile
- 3Laboratorio de Cannabinoides y Biología Espermática, Instituto de Histología y Embriología de Mendoza, Consejo Nacional de Investigaciones Científicas y Técnicas, Universidad Nacional de Cuyo, Mendoza, Argentina
- 4Escuela de Microbiología, Universidad Industrial de Santander, Bucaramanga, Colombia
- 5Institute for Research in Dental Sciences, Faculty of Dentistry, Universidad de Chile, Santiago, Chile
- 6Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile
- 7Instituto de Ciencias Naturales, Facultad de Medicina Veterinaria y Agronomía, Universidad de Las Américas, Santiago, Chile
- 8Centro de Investigación en Ciencias Biológicas y Químicas, Universidad de Las Américas, Santiago, Chile
Although significant progress has been made in medicine and antimicrobial research, viral infections continue to pose a critical global health challenge, particularly when they involve the central nervous system (CNS). Despite advances in vaccines, antiviral agents, and small molecule therapeutics, current strategies remain insufficient to address the complex consequences of many CNS infections fully. Notably, many viruses are neurotropic and can invade the CNS, triggering infectious neuroinflammation that often lead to chronic neurological disorders and lasting morbidity. Current therapeutic approaches are largely ineffective in preventing or reversing this long-term neurological damage, underscoring the urgent need for innovative prophylactic and therapeutic interventions. Mesenchymal stem cells (MSCs) have emerged as a promising strategy to counteract chronic neuroinflammation and promote tissue repair following viral CNS infections. This review provides a comprehensive overview of CNS viral infection and neuroinflammation, including epidemiology and pathophysiology, and critically examines the limitations of existing treatments, particularly their inability to mitigate persistent neurological sequelae. Furthermore, we summarize recent preclinical and clinical studies investigating the use of MSCs in the context of CNS viral infections, highlighting their immunomodulatory and neuroprotective mechanisms, and discuss the challenges and future directions for MSC-based therapies in clinical settings.
1 Introduction
Neuroinfection is a worldwide concern and an important cause of morbidity and mortality, characterized by pathogen invasion of the central nervous system (CNS), including the brain and spinal cord, which can lead to severe neurological damage and long-term disabilities due to chronic neuroinflammation (1).
Neuroinflammation is a physiological response induced as a defense mechanism against microbial infections or CNS injury. While this process can exert a protective role that contributes to pathogen clearance and the reestablishment of tissue homeostasis, it can also become persistent and exacerbated, often resulting in irreversible neurological damage and long-term cognitive or motor deficits, hearing loss, seizures, coma, and even death (2).
The CNS is protected by specialized protective structures, including the blood-brain barrier (BBB) and the blood-cerebrospinal fluid barrier (BCSFB), as well as resident immune cells such as microglia and astrocytes, all of which function to limit pathogen entry and protect the neural tissue (3). However, several pathogens have evolved strategies to bypass these defenses and provoke diseases like encephalitis, meningitis, and demyelinating pathologies (4). The incidence of infectious neuroinflammatory diseases remains challenging to determine due to the heterogeneity of etiological agents, limitations on epidemiological surveillance, and underreporting, especially regarding neurological complications and long-term outcomes. Nevertheless, it was estimated by the 2019 Global Burden of Disease Study that the total number of meningitis cases in all age groups reached 2.52 million, with an estimated 236 thousand deaths (5). On the other hand, in 2019, a total of 1,444,720 cases of encephalitis were estimated. Importantly, while encephalitis and meningitis represent the most severe clinical manifestations of neuroinfection, a larger proportion of affected individuals have milder symptoms, especially in the post-acute phase or chronic phases of infection (6).
Current treatments primarily target the causative agent, aiming to control the infection and, in some cases, attenuate the associated neuroinflammation (7, 8). However, these strategies often fail to address the chronic consequences of neuroinflammation, such as persistent neurological and cognitive impairments (9). Due the threat of emerging pathogens, antimicrobial resistance, and the limitations of current therapies in limiting the lifelong sequelae, the search for new venues for treating neuroinfection constitutes a critical public health problem. In this context, mesenchymal stem cell (MSC)-based therapies have emerged as a novel and promising approach to modulate neuroinflammatory responses and promote CNS repair (10–12). Preclinical and clinical studies support their benefits across diverse conditions, including cardiac injuries, autoimmune and neurodegenerative diseases such as Parkinson’s, Alzheimer’s, Huntington’s, and Multiple Sclerosis (MS) diseases (13–16). These findings have encouraged further exploration of MSC-based therapies as a prospective candidate to treat the detrimental effects of infection-induced neuroinflammation (15).
Although a wide array of microorganisms, including viruses, bacteria, fungi, and parasites, can cause CNS infections, this review will focus on viral pathogens, particularly those with well-reported neurotropism and high prevalence, such as human herpesviruses (herpes simplex virus type 1 and Epstein Barr virus), human retroviruses (human immunodeficiency virus type 1 and the human T-lymphotropic virus type 1), severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), and Zika Virus (1). In the following sections, we summarize the current state of the art and critically examine the potential of MSC to address the unmet clinical needs in the treatment of virus-induced neuroinflammatory conditions.
2 CNS viral infections and current treatment challenges
Several viruses can invade the CNS, leading to acute infection causing encephalitis, meningitis, among other diseases, all commonly accompanied by neuroinflammation. Despite advances in antiviral therapies and supportive care, effective treatments remain limited, particularly for managing the immune-mediated damage and persistent neurological symptoms that often follow viral clearance. This section provides an overview of the most relevant viral pathogens associated with CNS infections and examines the limitations of current therapeutic approaches used to combat their associated diseases.
2.1 Herpes simplex virus type 1 (HSV-1)
HSV-1 is a highly prevalent neurotropic virus that belongs to the Orthoherpesviridae family. It establishes lifelong latency in sensory neurons and can reactivate under stress or immune suppression (17, 18). HSV-1 infection of the CNS may lead to severe acute encephalitis (herpes simplex encephalitis, HSE), with a high mortality rate in untreated patients and high morbidity with neurological sequelae in survivors treated with the currently available antivirals. Notably, asymptomatic brain infection has also been reported in an important proportion of healthy individuals (>35%) and it is associated with residual chronic neuroinflammatory responses that are believed to lead to neurodegeneration (19).
During acute brain infection, neuronal and glial apoptosis and necrosis are observed, as well as BBB disruption, and exacerbated infiltration of innate immune cells into the CNS (20, 21). On the other hand, the long-term damages in the brain, are characterized by microglia activation, that lead to the release of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6, IL-1β, and chemokines C-X-C motif ligand (CXCL) 10 and C-C motif ligand (CCL) 5, that in turn recruit circulating lymphocytes to the CNS promoting chronic neuroinflammation (Figure 1A) (22, 23). This pro-inflammatory response has also been associated with tissue damage and neurodegeneration (24). Due to these characteristics, the contribution of HSV-associated neuroinflammation to the development of neurodegenerative disorders, such as MS, and Alzheimer’s disease, among others, has been widely considered (25–28).
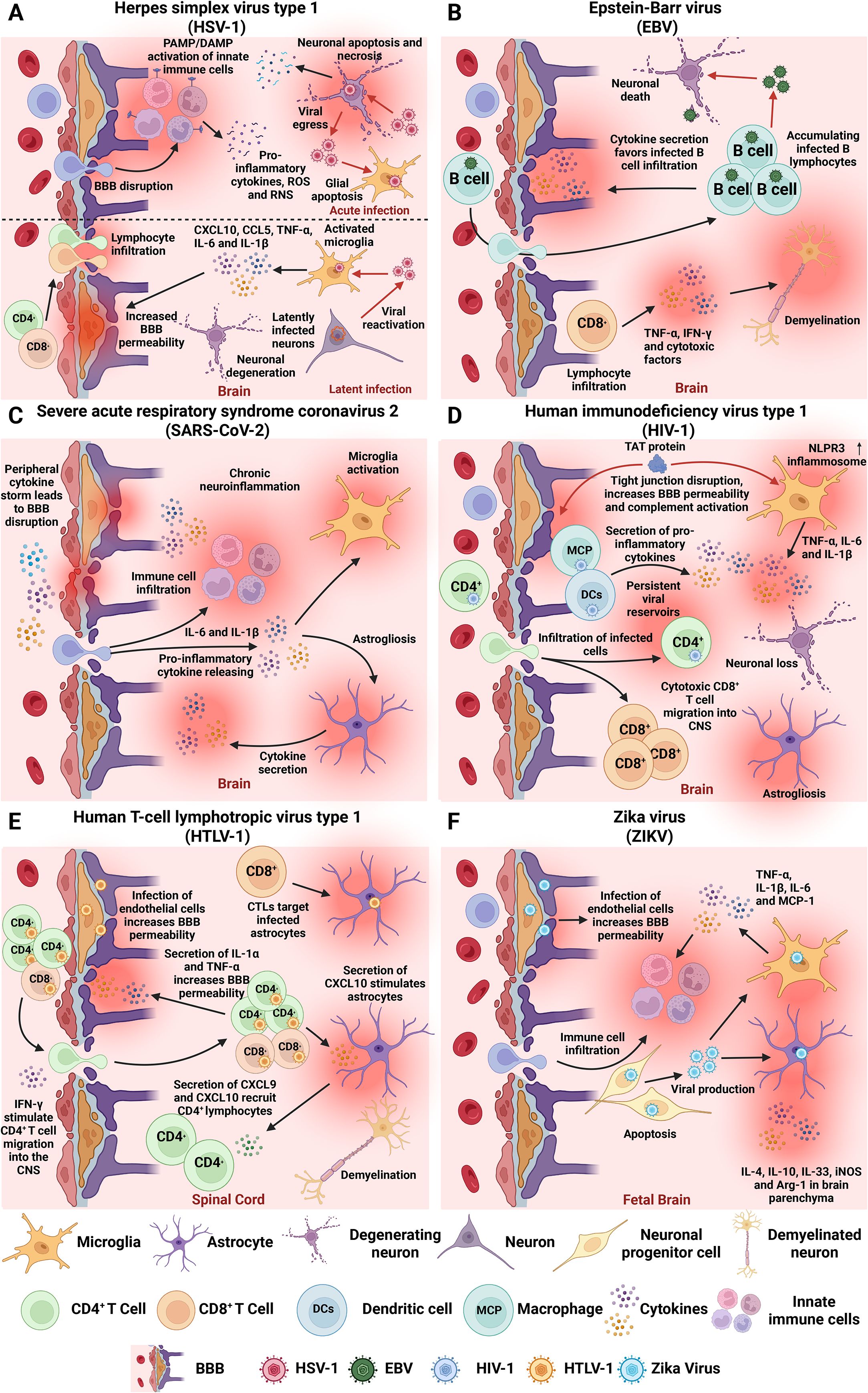
Figure 1. Neuroinflammation-associated hallmarks of viral infections in the CNS. Overview of key mechanisms by which prevalent human neurotropic viruses induce acute and/or chronic neuroinflammation, contributing to CNS dysfunction and long-term neuropathology. (A) HSV-1 can reach the CNS and cause an acute infection and herpes simplex encephalitis (HSE), characterized by neuronal and glial apoptosis and necrosis, BBB disruption, and massive innate immune cell infiltration. Latent infection is associated with sustained microglial activation and elevated levels of TNF-α, IL-6, IL-1β, CXCL10, and CCL5, promoting long-term neuroinflammation and tissue damage. (B) EBV establishes lifelong latency in B cells and has been associated with autoimmune and neurodegenerative disorders. The virus compromises BBB integrity, which facilitates CNS infiltration of infected B cells and triggers neuroinflammatory responses characterized by TNF-α and IFN-γ production, that can lead to chronic neuroinflammation and neurodegeneration. EBV-infected B cells and cytotoxic T cells accumulate in CNS tissues and can induce demyelination. (C) SARS-CoV-2 disrupts the BBB via systemic inflammation and cytokine storms. This leads to glial activation, increased IL-6 and IL-1β in CSF, and hippocampal injury. Other neuroinflammatory hallmarks include astrogliosis, microglial activation, and neuron damage. (D) HIV establishes reservoirs in CNS-resident macrophages and microglia early in infection. BBB disruption facilitates viral entry through a “Trojan Horse” manner into infected cells, enhanced by HIV Tat protein, which disrupts endothelial tight junctions and activates the complement cascade. Tat also triggers NLRP3 inflammasome activation and cytokine release by microglia. (E) HTLV-1 infects endothelial cells, promoting T cell infiltration and cytotoxic targeting of infected astrocytes. Infected immune cells infiltrate the spinal cord, where cytotoxic CD8+ T cells target HTLV-1-expressing cells and induce cytokine-mediated damage. (F) ZIKV virus crosses the placenta, infects neural progenitor cells, and induces apoptosis and impaired neurogenesis. ZIKV compromises BBB integrity and induces microglial activation with cytokine release. BBB, blood-brain barrier; CNS, central nervous system; IL, interleukin; IFN, interferon; TNF-α, tumor necrosis factor alpha.
Current antivirals like acyclovir (ACV) suppress viral replication but do not prevent long-term brain damage, and resistance, though rare, complicates treatment (29, 30). Unfortunately, nearly 50% of HSE patients treated with intravenous ACV show permanent sequelae after 1 year and 20% of mortality (29). Corticosteroids offer limited benefit due to immunosuppression risks (31, 32). Moreover, vaccine efforts remain unsuccessful (17, 33, 34), highlighting the urgent need for therapies that combine antiviral, immunomodulatory, and neuroprotective effects (35–37).
2.2 Epstein-Barr virus (EBV)
EBV, the gammaherpesvirus 4, causes infectious mononucleosis and is linked to carcinomas, lymphomas, and autoimmune diseases (38–41). After primary infection, it establishes lifelong latency in B cells, with periodic reactivation that can disrupt immune function (41, 42).
Importantly, EBV can invade the CNS, compromise the BBB integrity, and directly infect neurons leading to inflammation, tissue damage, and neurocognitive impairment (43, 44). It has been implicated in Alzheimer’s, Parkinson’s, and Multiple Sclerosis through mechanisms such as cytokine release, autoantibody cross-reactivity, and demyelination (Figure 1B) (45–54).
Despite growing evidence linking EBV to neurological disorders, its pathogenesis remains incompletely understood, and effective treatments are limited (55). Some compounds, such as cimetidine, have been used to treat chronic EBV reactivation and EBV associated carcinomas (56). Other molecules, including antiretrovirals, have shown anti-EBV activity and have been reported to induce long-term remission in some neurological disorders (57–61).
2.3 Severe acute respiratory syndorme coronavirus 2 (SARS-CoV-2)
SARS-CoV-2 is an RNA positive single-strand virus responsible for the COVID-19 pandemic, which has led to substantial global morbidity and mortality since its emergence in 2019 (62). It not only affects the respiratory system but also induces neurological damage. Some individuals develop lasting neurological symptoms, known as “Long COVID” or “Post-Acute Sequelae of SARS-CoV-2” (PASC) including cognitive deficits, delirium, and encephalopathy, that persist months after infection (63–65).
Neuropathological findings indicate that CNS injury is largely indirect, driven by peripheral cytokine storms that lead to hypoxia, BBB disruption, and microglial activation (Figure 1C) (66–69). Pro-inflammatory cytokines, such as IL-6 and IL-1β, have been detected in the cerebrospinal fluid (CSF) of patients with SARS-CoV-2 (70). Given the limited lymphatic drainage capability of the brain, these cytokines have been linked to neuropsychiatric symptoms (71). Thus, early treatment of the pro-inflammatory response against SARS-CoV-2 is crucial for reducing that symptoms and long-term sequelae, ultimately leading to the development of PASC (72).
Importantly, vaccination has reduced morbidity and mortality (73–79), and drugs such as remdesivir and tocilizumab can improve acute outcomes (80, 81), but no therapies currently prevent or treat PASC effectively (82), highlighting the need for new strategies targeting neuroinflammation and chronic sequelae.
2.4 Human immunodeficiency virus (HIV)
HIV is a single-strand RNA virus that establish latency in CD4+ T cells and reservoirs in macrophages and dendritic cells (DCs) leading to immune suppression and acquired immune deficiency syndrome (AIDS) (83). This loss of immune function increases susceptibility to opportunistic infections and malignancies (84). Besides its immunosuppressive effects, it has been described a spectrum of HIV-associated neurocognitive disorder (HAND), ranging from cognitive impairments to HIV-associated dementia, with up to 50% of patients affected (85).
Although HIV does not directly target the CNS, these neurological symptoms have been associated with virus infiltration through a “Trojan horse” manner due to the establishment of persistent reservoirs in perivascular macrophages and microglia (Figure 1D) (86). Moreover, although antiretroviral therapy (ART) can control HIV replication, the neurocognitive impairment and neuropathology persist in the CNS, as evidenced in post-mortem analysis of brain tissues with reported microglial activation, astrogliosis, neuronal loss, among other pathophysiological hallmarks of chronic neuroinflammation (87).
Notably, the viral trans-activator of transcription (Tat) protein facilitates CNS entry by disrupting endothelial tight junctions, activating complement, and recruiting monocytes (Figure 1D) (88). This, together with immune activation cause that macrophages release neurotoxins, cytokines, and metabolites, leading to brain damage (89, 90). Tat also activates the NLPR3 inflammasome in microglia, enhancing the release of TNF-α, IL-6 and IL-1β, thereby contributing to neuroinflammation (91, 92). Moreover, CD8+ infiltration into the brain can cause HIV CD8+ encephalitis (CD8E), a severe neurological manifestation which can lead to coma or even death if not treated properly (93).
While ART has significantly reduced mortality and incidence of HAND, milder cognitive symptoms persist in a large proportion of patients, indicating that ART alone is insufficient to fully prevent or reverse HIV-associated CNS injury (94–97), highlighting the need for therapies that target CNS inflammation and neuronal injury.
2.5 Human T-lymphotropic virus type 1 (HTLV-1)
HTLV-1, the first human retrovirus to be identified (98), affects between 5 and 10 million people worldwide (99) and is transmitted vertically, by non-protected sexual relationships, or through contamination with blood products (100–102).
In addition to its established role in adult T-cell leukemia (103), HTLV-1 is also responsible for HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP), a severe chronic inflammatory disease that involves the dysfunction of the spinal cord characterized by paraparesis, neurogenic bladder, and sensory disturbance of the legs (104–106). While HTLV-1 can infect multiple nucleated cells, the majority of the viral burden (≈95%) resides in CD4+ T cells (107, 108). This virus spreads from cell to cell via direct contact through the virological synapse, biofilm-like structures and cellular conduits (109).
In HAM/TSP pathology, T cell activation facilitates the migration of both CD4+ and CD8+ T lymphocytes from the peripheral blood by crossing the BBB. Within the CNS, HTLV-1-specific CD8+ T cells target HTLV-1 antigen-expressing cells, primarily infected CD4+ T cells and possibly infected glial cells (Figure 1E) (108), that in turn triggers the release of neurotoxic cytokines, contributing to tissue damage and demyelination (110, 111). After months or even years, the inflammatory process ends and, macroscopically, one of the signs found is the loss of spinal cord volume (112).
While no treatment is recommended in asymptomatic patients, corticosteroids can provide temporary symptom relief in progressive disease forms (113). On the other hand, antiretrovirals like zidovudine and lamivudine show inconsistent efficacy (114–117). Initial studies with Zidovudine reported both no clinical effect and improvements in patients (118, 119). Moreover, studies using drugs such as Lamivudine, Tenofovir, and Zidovudine + Lamivudine have shown either a reduction in viral load over a short period (120), in vitro activity (121), or conversely no decrease in viral load and no improvement in clinical symptoms (122). These findings highlight the need for novel therapeutic approaches capable of modulating the immune response, halting neurodegeneration, and improving functional outcomes in patients with HAM/TSP.
2.6 Zika virus (ZIKV)
ZIKV is a mosquito-borne human pathogen (123). This virus belongs to the Flaviviridae family, as do viruses such as Dengue virus and Yellow fever virus (124). This virus was linked to the 2015–2016 outbreak of congenital microcephaly, with thousands of cases of birth defects reported across the Americas (125–127).
Regarding the tropism of ZIKV for the CNS, studies have shown that the virus crosses the placenta and infects neural progenitor cells (NPCs) leading to apoptosis, disrupted neurogenesis and neurodevelopmental anomalies in the fetus (Figure 1F) (128). In adults, the infection can lead to neurological complications, such as Guillain‐Barre Syndrome, encephalitis, meningitis, myelitis and seizures (129). Severe brain infections are marked by presence of pro- and anti-inflammatory mediators, including IL-4, IL-10, IL-33, iNOS, and arginase 1 in brain parenchyma, suggesting a complex immune environment (130). Of these molecules, IL-33 is linked to necroptosis processes and may play a key role in ZIKV-induced neuronal injury (131).
ZIKV can also infect brain endothelial cells and compromise BBB integrity, facilitating viral entry and immune cell infiltration (132). Infection triggers a neuroinflammatory response characterized by the upregulation of cytokines and chemokines such as IL-6, TNF-α, IL-1β, and MCP-1 in microglia, promoting leukocyte recruitment and amplifying tissue damage (Figure 1F) (133).
Despite efforts in vaccine and antiviral development, no specific therapies are currently available to prevent or reverse ZIKV-induced neuroinflammation and its consequences.
2.7 Other viruses associated with chronic neuroinflammation
Japanese encephalitis virus (JEV), a mosquito-borne flavivirus, causes about 68,000 cases annually in Asia with 20–30% mortality (134, 135). During acute infection, JEV infects innate immune cells such as fibroblast, Langerhans cells, and macrophages, among others, activating antiviral pathways and secreting pro-inflammatory cytokines and chemokines (136). Once systemic replication is established, JEV can cross the BBB, likely facilitated by cytokine-induced permeability, and reaches the CNS (137). Inside the brain, JEV targets glial cells including microglia, inducing a pro-inflammatory state that leads to neuronal death and encephalitis (138–142). Clinical follow-up studies show that nearly half of survivors suffer long-term neurological sequelae, including cognitive deficits (143, 144). Importantly, no effective treatment exists (145). In 2021, a review comparing treatments against JE showed that only one of the twelve reports analyzed showed statistically significant positive outcomes after treatment (146). Minocycline showed only modest benefits without improving mortality or neurological outcomes compared to the placebo group (147).
Measles is a highly contagious infectious disease caused by Morbillivirus hominis formerly known as Measles virus (MeV) (148). This virus belongs to the Paramyxoviridae family and causes a disease with severe symptoms and complications and, in some cases, can lead to death (149). According to WHO, in 2018, nearly 142.000 measles-related deaths were reported, primarily among children under five (150). Subacute sclerosing panencephalitis (SSPE) is a chronic, progressive, and typically fatal infection of the CNS, caused by the persistence of MeV in the brain. Current therapies, such as interferon-α plus inosiplex have shown the highest rates of disease stabilization or improvement (151). However, continued research is needed to identify more accessible and effective treatments. Vaccination is the most effective preventive measure, yet vaccine hesitancy and incomplete coverage continue to drive outbreaks and severe complications (152–154).
In summary, current treatment strategies to combat CNS injury by viral infections are often insufficient likely due to the poor penetration across BBB, which restricts therapeutic efficacy at the site of chronic damage (Figures 2A-C). Additionally, for many neurotropic viruses, vaccines are absent or insufficient to prevent neurological sequelae, highlighting the urgent need for novel therapies that combine antiviral efficacy with immunomodulation to prevent or mitigate long-term neuroinflammatory damage.
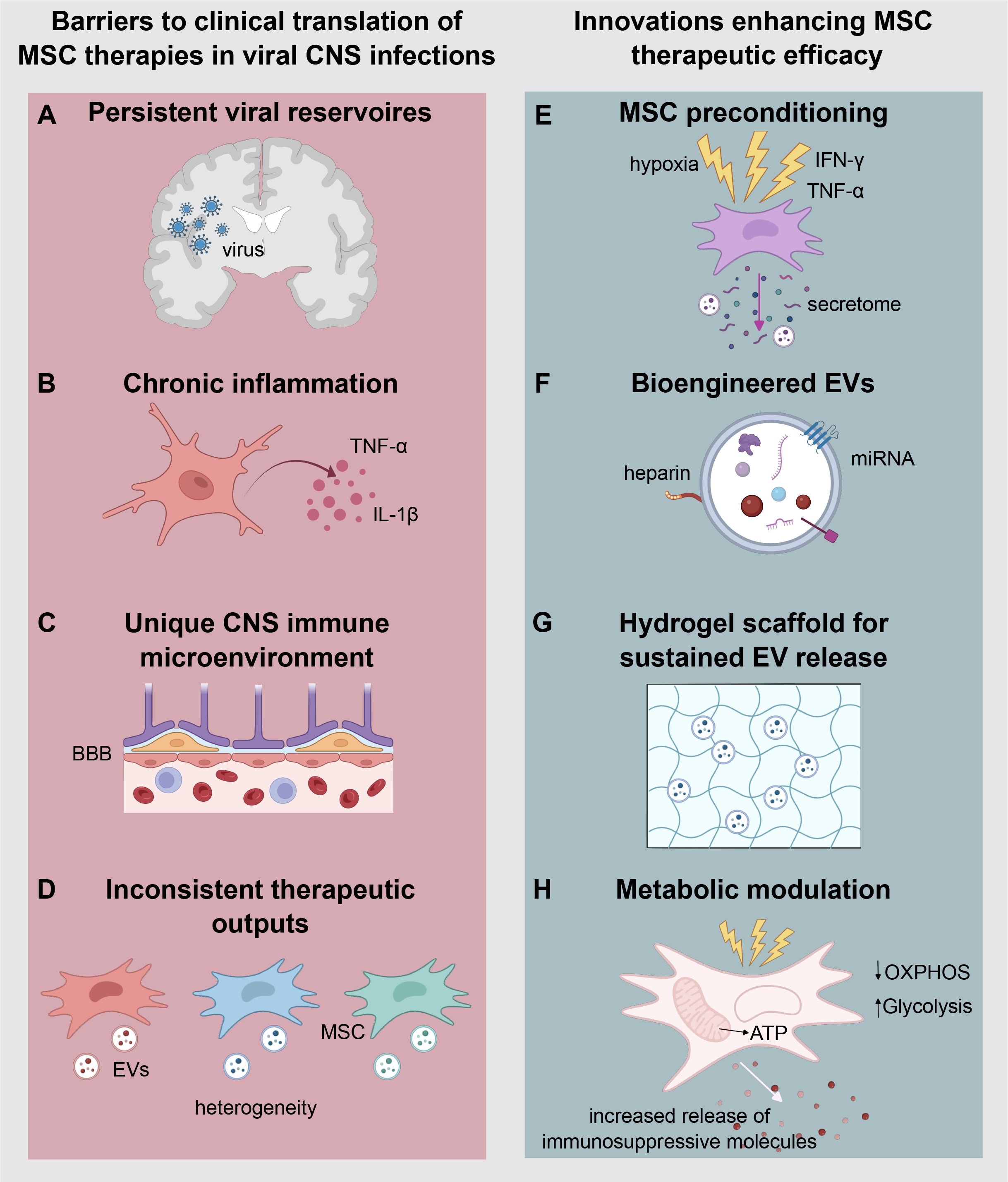
Figure 2. Barriers and emerging strategies for the clinical translation and enhancing therapeutic efficacy of mesenchymal stem cell therapies in viral central nervous system (CNS) infections. The left panel (A–D) illustrates key barriers to MSC-based therapeutic approaches in the context of viral CNS infections. (A) Persistent viral reservoirs promote chronic inflammation, which impairs the reparative function of MSCs. (B) Sustained production of pro-inflammatory cytokines such as TNF-α and IL-1β contributes to immune-mediated tissue damage. (C) The CNS immune microenvironment, characterized by the selective permeability of the blood-brain barrier (BBB), limits cell and therapeutic molecule access to the brain parenchyma. (D) MSC-based therapies are often hindered by inconsistent therapeutic outcomes due to donor heterogeneity and variability in the cargo of extracellular vesicles (EVs). The right panel (E–H) highlights emerging innovations aimed at overcoming these challenges and enhancing the therapeutic efficacy of MSCs. (E) MSC preconditioning using stimuli such as hypoxia, IFN-γ, or TNF-α enhances their anti-inflammatory and immunomodulatory secretome. (F) Bioengineered EVs, which incorporate targeted ligands and optimized miRNA content, improve brain targeting and functional delivery. (G) Hydrogels enable the sustained release of EVs, maintaining therapeutic concentrations in chronic inflammatory environments. (H) Metabolic modulation of MSCs promotes a shift from oxidative phosphorylation (OXPHOS) to Glycolysis, enhancing ATP production and driving the increased release of immunosuppressive molecules.
3 Mesenchymal stem cells as an alternative to treat neuroinflammation and promote CNS repair
MSC-based therapies, which include MSCs, extracellular vesicles (EVs), and their secretome, has demonstrated promising potential to reduce not only viral burden but also the associated neuroinflammatory response (14, 155, 156). In the next sections, we summarize the biological characteristics and immunomodulatory mechanisms of MSCs and their derivates, along with their applications in neuroinflammatory conditions.
3.1 Characteristics of mesenchymal stem cells and their secretome
MSCs are multipotent stromal cells with self-renewal capabilities and inherent tropism for injured tissues, including the inflamed CNS. According to the criteria established by the International Society for Cellular Therapy (ISCT), MSCs are characterized by adherence to plastic under standard culture conditions; expression of surface markers such as CD105, CD73, and CD90; and the absence of hematopoietic markers such as CD45, CD34, CD14, CD19, and HLA-DR (157, 158). Although MSCs can differentiate into osteoblasts, adipocytes, and chondroblasts in vitro, their therapeutic utility in vivo is primarily attributed to paracrine signaling rather than direct differentiation (159, 160).
MSCs are isolated from multiple tissue sources, each conferring distinct biological and clinical advantages. Bone marrow-derived MSCs (BM-MSCs) are the most extensively studied, with well-documented osteogenic and immunomodulatory properties; however, their isolation is invasive, and cell yield declines with donor age (161). Adipose-derived MSCs (AD-MSCs) are obtained through minimally invasive procedures, offering high cell yields and demonstrating therapeutic efficacy, with particular relevance in angiogenic applications (162). Umbilical cord-derived MSCs (UC-MSCs), particularly those from Wharton’s jelly, exhibit a primitive phenotype with enhanced proliferative capacity and lower immunogenicity, making them ideal for allogeneic transplantation (159). Emerging sources such as dental pulp, menstrual blood, and amniotic fluid are gaining attention due to their niche-specific advantages. For instance, dental pulp MSCs show strong neurogenic potential (163), while amniotic fluid-derived MSCs retain pluripotent-like properties (164).
The main therapeutic efficacy of MSCs is largely attributed to their secretome, which comprises a complex repertoire of growth factors such as brain-derived neurotrophic factor (BDNF) and vascular endothelial growth factor (VEGF), as well as immunoregulatory cytokines (e.g., IL-10, TGF-β), and EVs carrying regulatory miRNAs, mRNAs, and proteins that modulate immune and neuroinflammatory pathways which promote neuroprotection and tissue repair (165–167). EVs, especially exosomes, have garnered increasing interest by influencing cellular behavior without the risks associated with live cell therapies. EVs have shown anti-inflammatory and neuroprotective effects in various models of CNS diseases. Notably, EVs exhibit an enhanced capacity to cross the BBB compared to whole MSCs, thereby increasing their CNS bioavailability (168). Importantly, MSCs display a natural ability to migrate to sites of inflammation, a phenomenon guided by the expression of chemokine receptors such as CCR2, CXCR4, and CX3CR1 and their ligands secreted by inflamed tissues (169). These properties establish MSCs and their derivatives as strong candidates for cell-based therapies targeting the immune-mediated components of CNS viral infections.
3.2 Current applications of MSCs in neuroinflammation and CNS repair
MSCs secrete a diverse array of bioactive molecules, including cytokines, growth factors, and EVs loaded with regulatory microRNAs and proteins that collectively suppress neuroinflammation and promote tissue repair (155, 170–172). A key mediator, the tumor necrosis factor-inducible gene 6 protein (TSG-6), binds to CD44 receptors on microglia and macrophages, inhibiting TLR2/NF-κB signaling and reducing proinflammatory cytokines (e.g., TNF-α, IL-1β), and enhancing anti-inflammatory cytokines such as IL-10 and TGF-β (168, 173). This mechanism disrupts the feed-forward cycle of neuroinflammation, a hallmark in viral CNS complications, such as HAND and HSE (173). Secreted cytokines like IL-10, TGF-β, and PGE2 suppress pro-inflammatory Th1/Th17 responses and promote regulatory T cell expansion, which could attenuate cytokine-driven damage during viral CNS infections (174, 175), while neurotrophic factors such as BDNF, nerve growth factor (NGF), hepatocyte growth factor (HGF), and VEGF enhance neuronal survival and synaptic plasticity (176, 177).
Moreover, microRNAs-containing EVs (e.g., miR-134, miR-138-5p, miR-21-3p) reduce oxidative stress and neuronal apoptosis by modulating pathways such as the KDM6B–BMP2/BMF axis (178). These miRNAs have been shown to promote neuronal survival and functional recovery in experimental models of neuroinflammation (179, 180). Notably, miRNAs display greater stability and functionality when encapsulated in EVs, compared to the broader MSC secretome, where their half-life is significantly shorter due to enzymatic degradation (181).
Furthermore, MSCs and their EVs modulate the neuroimmune microenvironment by inducing microglial polarization from a proinflammatory M1 to an anti-inflammatory M2 phenotype (173). This phenotypic shift fosters a regenerative niche that supports synaptic repair and axonal regeneration. Regarding spinal cord injury models, MSC-EVs promote axonal regeneration by modulating the Rho-GTPase pathway, enhancing synaptic reconnection and functional recovery even during chronic stages (182, 183). Clinical trials in traumatic brain injury (TBI), have shown that intravenous MSC administration within 48 hours post-injury resulted in reduced plasma neurofilament light chain (NfL) levels, suggesting decreased axonal injury (184). The MATRIx phase II trial also showed greater white matter integrity and improved functional recovery with MSC therapy compared to placebo (185). In neuroinflammatory conditions such as MS and TBI, early-phase clinical trials have shown MSC infusions safety and some improvement in neurological function and reductions in systemic and CNS inflammation (186, 187).
Additionally, the MSC secretome plays a crucial role in modulating the neurogenic niche of the subventricular zone (SVZ), a key neurogenic area in the adult brain (188, 189). The SVZ harbors endogenous neural stem cells capable of proliferating, differentiating, and migrating to sites of injury or inflammation, thereby facilitating intrinsic brain repair (189–191). Factors secreted by MSCs and their EVs promote the proliferation and differentiation of these neural progenitor cells, enhancing neurogenesis and contributing to neuronal regeneration (189, 192). This paracrine effect complements the immunomodulatory functions of MSCs, expanding their therapeutic potential to repair neural damage in the context of chronic CNS inflammation, such as in viral infections (193).
4 Advancing MSC-based therapies from bench to bedside for treating CNS viral pathologies
Although MSCs are not inherently antiviral agents, these cells and their derivatives may trigger a host response that indirectly suppresses viral replication helping to preserve the integrity of the BBB and reducing inflammation (194). In preclinical models, MSC-based treatments have shown to reduce viral titers and improve clinical outcomes (155, 195, 196). Moreover, clinical investigations are underway to validate the safety, tolerability, and efficacy of MSC-based therapies in multiple indications, as well as for determining optimal doses and delivery strategies. However, studies specifically targeting viral infections in the CNS remain limited and face significant regulatory and scalability challenges, as discussed in the following sections (177).
4.1 Preclinical evidence supporting the use of MSC-based therapies in viral infections and neuroinflammatory conditions
MSC co-cultured with JEV-infected microglia and neurons in vitro have shown the capacity to promote microglial polarization towards an anti-inflammatory M2 phenotype, enhance neuronal survival, and reduce viral replication (155). Consistent with this, mice infected with JEV and treated with BM-MSCs displayed a reduced mortality, neuroinflammation, and viral load, accompanied by decreased microglial activation, lower levels of pro-inflammatory cytokines such as TNF-α, IFN-γ, and CCL-2, preservation of the BBB, and increased expression of type I interferons (IFN-α/β) (155).
Similar findings have been reported for coxsackievirus B3 (CVB3)-induced myocarditis. Human BM-MSCs reduced apoptosis, oxidative stress, and viral replication, through mechanisms dependent on nitric oxide production induced by IFN-γ priming (196). Moreover, exosomes derived from human UC-MSCs alleviated inflammation and apoptosis in CVB3-infected human cardiomyocytes through activation of the AMPK/mTOR autophagy pathway, promoting cell survival and protein degradation (197). In murine models, intravenous administration of BM-MSCs or their EVs improved cardiac function, decreased myocardial apoptosis and inflammation, and lowered TNF-α (196–198).
Promising results also emerge in herpesviruses infections. In CMV-infected murine macrophages, exosomes from BM-MSCs shifted macrophage phenotype from pro-inflammatory M1 to anti-inflammatory M2 phenotypes and reduced pro-inflammatory cytokines (199). In murine models of CMV-induced pneumonia, MSC-exosomes significantly improved survival, and reduced weight loss, lung damage, inflammatory cell infiltration, and pulmonary fibrosis. These effects were associated with reduced NF-κB activation and suppression of the NLRP3 inflammasome (199). Moreover, conditioned media derived from bone marrow–isolated MSCs in mice has been shown to inhibit HSV-1 infection in vitro. Furthermore, administration of BM-MSCs in a lethal HSV-1 mouse model conferred 70% protection, compared with only 10% survival in untreated animals. MSC treatment was also associated with enhanced production of virus-neutralizing anti-HSV-1 antibodies and increased T-cell proliferation (200). Another study demonstrated that prior immunization with MSCs protected all mice from lethal HSV-1 infection, while genetically modified MSCs transfected with the Us6 gene encoding glycoprotein D conferred 65% protection compared to untreated mice that had 100% mortality (201). These findings highlight the unique capacity of MSCs to stimulate innate, adaptive, and protective immunity representing promising candidates for the development of next-generation cell-based vaccines against herpes and other viral infections.
Although CMV, HSV, and VZV can infect MSCs in vitro, viral replication is limited, possibly associated with constitutive expression of interferon-stimulated genes (ISG) (202, 203). In the same line, MSCs are non-permissive to EBV and HHV-6/7 infections (204). However, the direct antiviral or immunomodulatory effects of MSCs, their exosomes, or their secretome in the context of VZV, EBV, or HHV-6/7 infections remain largely unexplored. Further studies are needed to define the therapeutic potential of MSC-based interventions on the diseases caused by these viruses.
Notably, EVs derived from human Wharton’s Jelly MSCs, showed a dose-dependent inhibition of HSV-2 in Vero cells assays (205). At non-toxic doses, EVs reduced viral spread and cytopathic effects (205). Their antiviral mechanism may involve direct virucidal activity and interference with viral replication, possibly via antiviral microRNAs (205).
In HIV models, MSC therapy showed a dual effect. When were used on two latently infected U1 (monocytic) and ACH2 (T-cell) lines, MSCs could reactive latent virus via the PI3K and NF-κB pathways but also enhanced the efficacy of latency-reversing agents (LRAs) like panobinostat and Phorbol 12-myristate 13-acetate (PMA) (206). MSCs also demonstrated in vivo potential to restore immune balance, reduce neuroinflammation and modulate microglial activation, with possible benefits in HAND (207).
MSC-based therapies have been explored to mitigate cytokine storms and organ damage after SARS-CoV-2 infection. In preclinical models of acute respiratory distress syndrome (ARDS), MSCs improved oxygenation, reduced alveolar inflammation, and preserved endothelial integrity (208–210). MSC-derived EVs have also been proposed to treat COVID-19-associated neuroinflammation and stroke, through immune modulation and neuro-regenerative effects (211).
MSC and their secretome modulate multiple arms of antiviral immunity mechanisms that could in principle facilitate the viral clearance and disease resolution during acute infections. Indeed, they can produce type I interferons and secrete chemokines, that recruit and activate plasmacytoid, myeloid dendritic cells, and enhance NK and CD8+ T cell functions (212, 213). Once the infection has been resolved, MSCs shift toward anti-inflammatory phenotypes, releasing mediators such as TGF-β, indolamine 2,3-dioxygenase (IDO), IL-10, IL-1Ra, and PGE2, which control excessive immune activation and prevent cytokine storms (194, 203, 214). However, certain viruses, including HIV, HSV and hepatitis B virus (HBV), can infect MSC and potentially establish viral reservoirs, raising safety concerns. Therefore, donor MSCs must be rigorously screened for these pathogens prior to transplantation to minimize the risk of virus-associated complications and ensure the safety of MSC-based therapies.
In summary, while MSC show promising effects in acute viral infection by reducing viral load and dampening acute inflammation, evidence for their role in clearing persistent or latent infections is weaker. MSCs have been shown to reactivate latent HIV-1 in monocytic and T-cell models through activation of a noncanonical PI3K–NFκB–dependent pathway (206). Although this reactivation may represent a therapeutic leverage point, whether it leads to immune-mediated clearance in vivo remains to be determined. In persistent infections, MSCs could potentially serve as adjuvants to conventional antiviral therapy. Supporting this, exosomes derived from UC-MSCs have been reported to enhance the antiviral effects of interferon-α and telaprevir against hepatitis C in vitro, suggesting additive or even synergistic activity; these exosomes carry microRNAs that directly complement viral genomes within host cells (215).
4.2 Limited clinical evidence supporting MSC-based therapies for viral CNS infections
While preclinical studies suggest that MSCs can attenuate neuroinflammation, protect neurons, and indirectly suppress viral replication, clinical evidence in humans, particularly in cases of viral encephalitis and other CNS viral infections remains limited (216, 217). To date, most research has relied on rodent models or in vitro systems. Extrapolating these results to human applications requires caution, given substantial interspecies differences in immune response dynamics and CNS architecture (218).
Human clinical studies with MSCs have mostly focused on systemic viral infections, especially COVID-19, where MSCs are used for immunomodulation rather than direct viral clearance (186, 219). A search of the National Institutes of Health database (http://www.clinicaltrials.gov/), using “virus” as the condition/disease and “mesenchymal stem cells” as other terms, identified 107 MSC-based clinical trials. After excluding studies that did not involve MSCs for the treatment of viral infections and those targeting more than one virus, a total of 90 trials were tabulated in Figure 3. The distribution of studies by country revealed a predominance in China and the United States (Figure 3A). UC-MSC, BM-MSC and AD-MSC were the most frequently utilized cell sources (Figure 3B). The majority of trials conducted to date are phase 1 and phase 2 studies designed to assess safety and feasibility, with limited evidence available so far regarding therapeutic efficacy (Figure 3C). Numerous MSC-based studies have been registered for treatment of COVID-19 related pneumonia and acute respiratory distress syndrome (ARDS). Table 1 summarizes completed clinical studies involving MSC treatment for treating SARS-CoV-2 acute infections. MSC infusions demonstrated favorable safety and improved inflammatory profiles, supporting a substantial immunomodulatory potential in systemic viral infections that impact the CNS (238, 239).
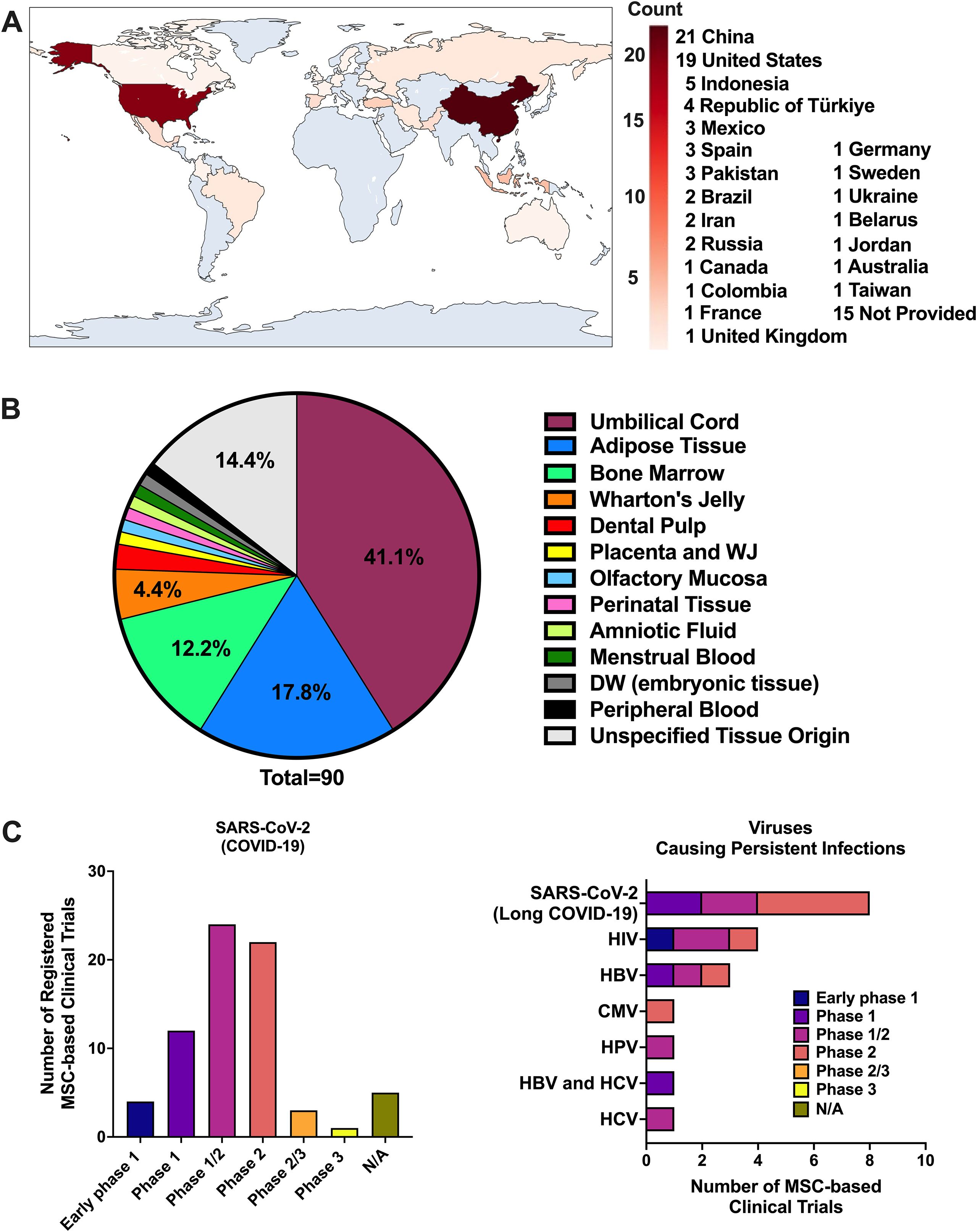
Figure 3. Summary of clinical trials on MSC-based therapies targeting viral infections. Overview of MSC-based clinical trials for viral infections. (A) Distribution of trials by country. (B) MSC sources used across studies. (C) Targeted viral pathogens, subdivided into SARS-CoV-2 acute infection/COVID-19 (left) and viruses associated with persistent infections, including long COVID due to SARS-CoV-2 (right). Data were obtained from the NIH ClinicalTrials.gov database by searching “virus” and “mesenchymal stem cells.” Of 107 identified studies, 90 were included after excluding those not using MSCs for viral infection treatment or targeting multiple viruses.
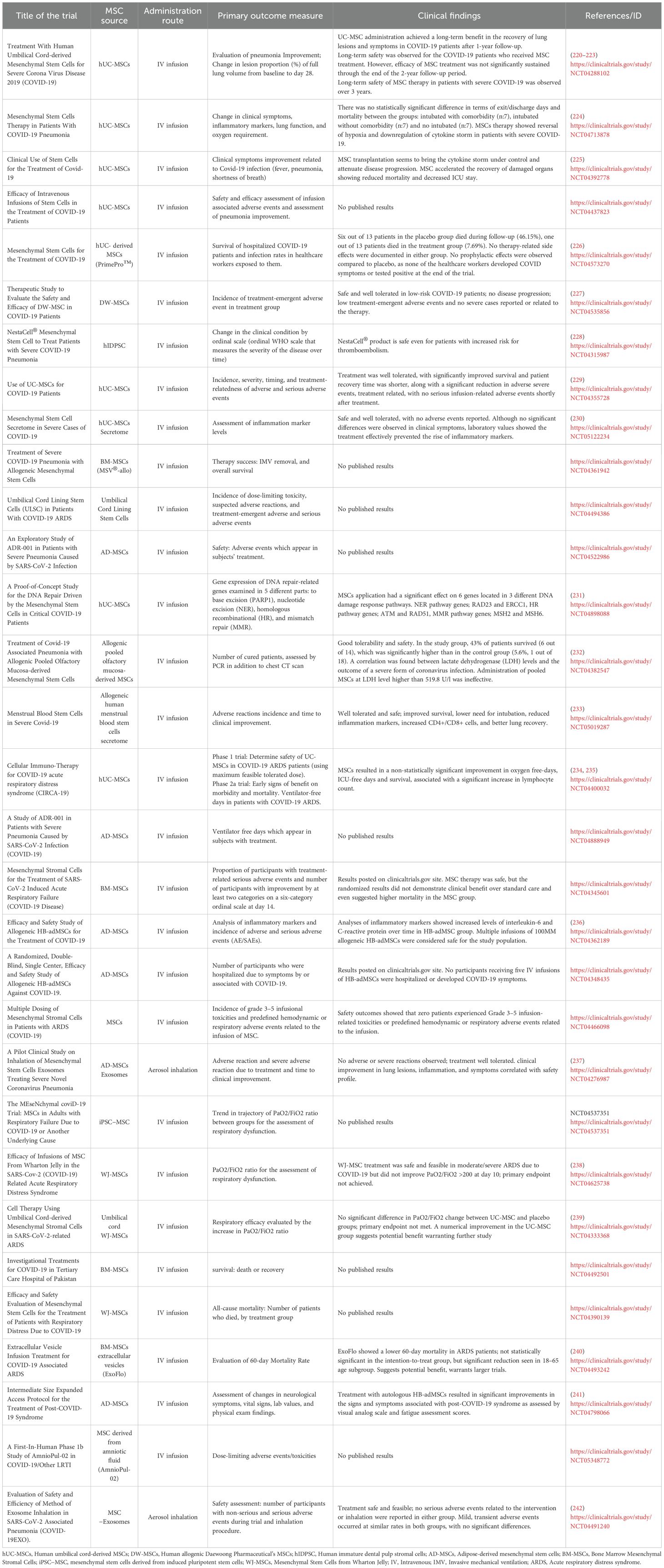
Table 1. Completed clinical studies on clinicaltrials.gov evaluating the use of MSC or their derivates on SARS-CoV-2 infection.
Although most studies focus on acute COVID-19, there is a growing interest in evaluating MSCs for persistent viral infections. A limited number of trials are investigating MSCs or their derivatives in chronic or latent infections and their associated complications, as summarized in Table 2, with some reporting encouraging outcomes. For instance, in HIV-positive individuals with poor immune recovery, allogeneic UC-MSC infusions increased CD4+ T-cell counts and enhanced HIV-specific responses without boosting viral load (246). However, these therapeutic effects appear to depend on the MSC source, as AD-MSC infusions failed to improve immune recovery in immune non-responder patients, showing no significant changes in lymphocyte subset phenotypes or in the inflammatory parameters analyzed (245).
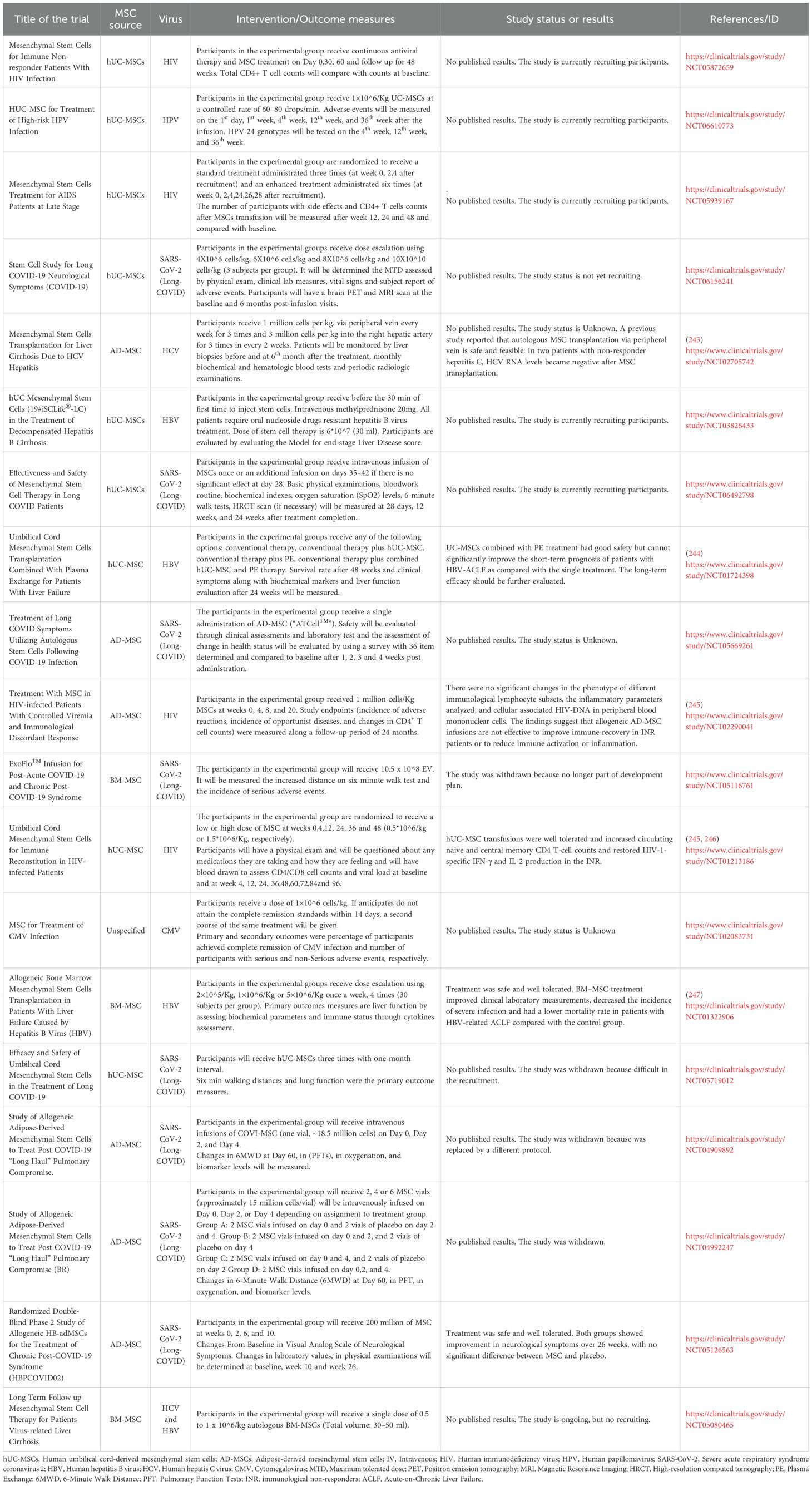
Table 2. Clinical studies registered on clinicaltrials.gov that evaluate the safety and efficacy of MSCs-based therapies against persistent viral infections.
In the case of HBV, one study reported that BM–MSC treatment improved clinical laboratory measurements, decreased the incidence of severe infection, and lowered mortality in patients with HBV-related acute-on-chronic liver failure (ACLF) compared with the control group (247). By contrast, combining UC-MSCs with plasma exchange (PE) demonstrated good safety for patients with liver failure but did not significantly improve the short-term prognosis compared with PE alone (244). Moreover, a case report described an HBV-related ACLF patient treated with repeated PE and UC-MSCs in combination with antiviral therapy (entecavir, ETV), who survived with marked improvements in hepatic function (248). In addition, a 24-month prospective study found that UC-MSC transplantation combined with PE and ETV was safe and effective, resulting in a higher cumulative survival rate than PE treatment alone (249).
While these findings highlight the immunomodulatory potential of MSCs for chronic viral infections, their effects on associated neurocognitive disorders remain unexplored in controlled clinical settings, with a clear translational gap between preclinical findings and human application. To date, only a single study has reported results in four patients with SSPE treated with MSCs, in which no clinical benefit was observed (250). Notably, a prospective, non-randomized observational registry study is currently underway to evaluate the clinical outcomes of patients with autoimmune or post-infectious neuroinflammatory syndromes receiving MSC-based therapies. This includes post-viral encephalopathies following measles, CMV, EBV, and SARS-CoV-2, as well as early-stage panencephalitis and chronic neuroimmune syndromes such as long COVID with CNS involvement (https://clinicaltrials.gov/study/NCT07145502). It is likely that in the coming years additional data will emerge, particularly from ongoing studies evaluating MSCs in long-COVID.
4.3 Emerging strategies to improve MSC therapeutic efficacy and overcome barriers to translational application
The clinical translation of MSC-based therapies in viral CNS infections faces several challenges including persistent viral reservoirs, chronic inflammation, and the specialized CNS immune microenvironment. Moreover, MSC experience variable efficacy and short-term activity in vivo (Figures 2A-D) (217, 251). Therefore, strategies to precondition or engineer MSCs are critical to enhance their therapeutic robustness and reproducibility (252) Emerging technologies are being developed to enhance MSC and EV functionality, improve delivery efficiency, and ensure consistent therapeutic outcomes (Figures 2E-H) (176, 198, 253).
Preconditioning approaches aim to “prime” MSCs by exposing them to specific environmental cues before transplantation, thereby activating protective and therapeutic pathways (254, 255). For instance, hypoxic preconditioning (1–5% O2) mimics the low oxygen tension of the native MSC niche, stabilizing hypoxia-inducible factors (HIFs) that upregulate glycolytic enzymes and promote a shift toward aerobic glycolysis (256, 257). This metabolic reprogramming enhances MSC survival, mitochondrial fitness, and resistance to oxidative stress, while boosting the secretion of trophic and angiogenic factors such as VEGF, BDNF, and HGF, thereby increasing regenerative and neuroprotective efficacy (256, 257). Similarly, inflammatory cytokine preconditioning using factors such as IFN-γ, TNF-α, or IL-1β upregulates key immunoregulatory molecules including IDO, TSG-6, and PD-L1, improving the MSC capacity to dampen immune responses, which could potentially inhibit viral-associated inflammation (Figure 2E) (258, 259).
In parallel, genetic engineering strategies are increasingly employed to overcome inherent limitations of MSCs (198, 260). Using viral vectors or CRISPR/Cas9-based systems, MSCs can be modified to overexpress therapeutic genes such as CXCR4 (to improved homing), IL-10, BDNF, or IFN-β, thereby enhancing their immunomodulatory, antiviral, and neurotrophic potential (170, 173, 261, 262). On the other hand, bioengineered EVs have been conjugated with heparin for enhancing brain targeting and MSC preconditioning with interferon γ or viral antigens for boosting IFN-β secretion while maintaining immunomodulatory capacity (Figure 2F) (263, 264).
Hydrogel scaffolds that sustain EV release may support long-term miRNA delivery, maintaining therapeutic thresholds in chronic infections (Figure 2G) (265, 266). Metabolic modulation represents another frontier, where exposure to agents that shift metabolism towards glycolysis by increasing the expression of certain glucose transporters can enhance MSC resistance to stress and alter the composition of their secretome toward a more anti-inflammatory and neuroprotective profile (Figure 2H) (267). Additionally, chemical preconditioning using small molecules (e.g., valproic acid, resveratrol) has also been explored to boost anti-apoptotic and antioxidant properties (268, 269).
Collectively, these bioengineering strategies, including hypoxic and inflammatory preconditioning, metabolic reprogramming, chemical priming, and genetic modification, are essential tools to optimize MSC function, especially under the hostile conditions of viral CNS infections. Their implementation could significantly improve the reproducibility, robustness, and clinical impact of MSC-based therapies.
5 Concluding remarks
Viral infections of the CNS represent a significant global health burden due to their capacity to induce persistent neuroinflammation and long-term neurological sequelae. Given the current state of evidence, future research should prioritize the design of well-powered randomized controlled trials targeting viral CNS infections, rather than extrapolating from systemic viral infections or non-viral neuroinflammatory disorders. Standardization of MSC manufacturing, dose regimens, and delivery routes will be essential to reduce variability and ensure reproducibility (270). Long-term follow-up studies will be critical to assess the durability of therapeutic benefits and to detect any delayed adverse events that may arise from MSC transplantation. Moreover, mechanistic studies in human subjects are needed to determine the precise contributions of MSCs to viral clearance, host immune responses modulation, and repair processes within the CNS (271). This knowledge will be pivotal for refining patient selection criteria and optimizing treatment regimens tailored to specific viral etiologies (194, 272). Emerging strategies, including artificial intelligence-driven secretome optimization and 3D bioprinting of MSC-laden scaffolds, aim to standardize production and enhance therapeutic consistency (273). Furthermore, combining MSC-EVs with nanotechnology-based delivery systems could improve CNS targeting, offering a translatable solution for viral infections like HAND or HSE. Combining MSC-based strategies with antiviral or immunomodulatory agents may offer synergistic benefits; however, this remains to be fully evaluated. While the therapeutic potential of MSC-based therapies is substantial, it is essential to approach this field with scientific rigor, and an interdisciplinary approach will be crucial to move MSC-based interventions from bench to bedside in the treatment of virus-induced neuroinflammatory diseases.
Author contributions
JC: Conceptualization, Writing – original draft, Writing – review & editing. JF: Writing – original draft, Writing – review & editing. NS: Writing – original draft. WC: Writing – original draft. AM: Writing – review & editing. MR: Writing – review & editing. PG: Writing – review & editing. MM: Writing – review & editing. MO: Writing – review & editing. ME: Writing – review & editing. LD: Writing – review & editing, Conceptualization, Supervision, Writing – original draft.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Authors are supported by the National Agency for Research and Development (ANID) through Fondo Nacional de Desarrollo Cientı́fico y Tecnológico (FONDECYT) grants ANID: ANID FONDECYT Iniciación grant#11241567 (LD), FONDECYT Regular grants #1240971 (PG),#1241130 (AM-B). Fondo de Fomento al Desarrollo Cientı́fico y Tecnológico FONDEF-ID24I10119 (ME). ANID - – Millennium Science Initiative Program – Millennium Institute on Immunology and Immunotherapy (MIII) ICN2021_045 (former ICN09_016; P09/016-F) (PG).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Nurmukanova V, Matsvay A, Gordukova M, and Shipulin G. Square the circle: diversity of viral pathogens causing neuro-infectious diseases. Viruses. (2024) 16:787. doi: 10.3390/v16050787
2. Tohidpour A, Morgun AV, Boitsova EB, Malinovskaya NA, Martynova GP, Khilazheva ED, et al. Neuroinflammation and infection: molecular mechanisms associated with dysfunction of neurovascular unit. Front Cell Infect Microbiol. (2017) 7:276. doi: 10.3389/fcimb.2017.00276
3. Ampie L and McGavern DB. Immunological defense of CNS barriers against infections. Immunity. (2022) 55:781–99. doi: 10.1016/j.immuni.2022.04.012
4. Dando SJ, Mackay-Sim A, Norton R, Currie BJ, St. John JA, Ekberg JAK, et al. Pathogens penetrating the central nervous system: Infection pathways and the cellular and molecular mechanisms of invasion. Clin Microbiol Rev. (2014) 27:691–726. doi: 10.1128/CMR.00118-13
5. Wang H, Zhao S, Wang S, Zheng Y, Wang S, Chen H, et al. Global magnitude of encephalitis burden and its evolving pattern over the past 30 years. J Infect. (2022) 84:777–87. doi: 10.1016/j.jinf.2022.04.026
6. Makkawi S, Alqurashi S, Hubayni W, Almahdawi S, Bahkali S, Alharbi A, et al. The clinical manifestations, risk factors, etiologies, and outcomes of adult patients with infectious meningitis and encephalitis: single center experience. Neurol Int. (2024) 16:966–75. doi: 10.3390/neurolint16050073
7. Sreeram S, Ye F, Garcia-Mesa Y, Nguyen K, El Sayed A, Leskov K, et al. The potential role of HIV-1 latency in promoting neuroinflammation and HIV-1-associated neurocognitive disorder. Trends Immunol. (2022) 43:630–9. doi: 10.1016/j.it.2022.06.003
8. Malik S, Sah R, Ahsan O, Muhammad K, and Waheed Y. Insights into the Novel Therapeutics and Vaccines against Herpes Simplex Virus. Vaccines (Basel). (2023) 11:325. doi: 10.3390/vaccines11020325
9. Yari N, Benardete EA, Chen W, Ambe SN, and Fonkem E. Outcomes with acyclovir treatment in herpes simplex encephalitis after surgery for solid CNS tumors: A case report and systematic review of the literature. Neuro Oncol Pract. (2019) 6(4):259–63. doi: 10.1093/nop/npz007
10. Mincheva G, Moreno-Manzano V, Felipo V, and Llansola M. Extracellular vesicles from mesenchymal stem cells improve neuroinflammation and neurotransmission in hippocampus and cognitive impairment in rats with mild liver damage and minimal hepatic encephalopathy. Stem Cell Res Ther. (2024) 15:472. doi: 10.1186/s13287-024-04076-6
11. Joshi HP, Jo HJ, Kim YH, An SB, Park CK, and Han I. Stem cell therapy for modulating neuroinflammation in neuropathic pain. Int J Mol Sci. (2021) 22:4853. doi: 10.3390/ijms22094853
12. Ge Y, Wu J, Zhang L, Huang N, and Luo Y. A new strategy for the regulation of neuroinflammation: exosomes derived from mesenchymal stem cells. Cell Mol Neurobiol. (2024) 44:24. doi: 10.1007/s10571-024-01460-x
13. Zaripova LN, Midgley A, Christmas SE, Beresford MW, Pain C, Baildam EM, et al. Mesenchymal stem cells in the pathogenesis and therapy of autoimmune and autoinflammatory diseases. Int J Mol Sci. (2023) 24:16040. doi: 10.3390/ijms242216040
14. Yasamineh S, Kalajahi HG, Yasamineh P, Gholizadeh O, Youshanlouei HR, Matloub SK, et al. Spotlight on therapeutic efficiency of mesenchymal stem cells in viral infections with a focus on COVID-19. Stem Cell Res Ther. (2022) 13:257. doi: 10.1186/s13287-022-02944-7
15. Islam MA, Alam SS, Kundu S, Ahmed S, Sultana S, Patar A, et al. Mesenchymal stem cell therapy in multiple sclerosis: A systematic review and meta-analysis. J Clin Med. (2023) 12:6311. doi: 10.3390/jcm12196311
16. Mattei V and Delle Monache S. Mesenchymal stem cells and their role in neurodegenerative diseases. Cells. (2024) 13:779. doi: 10.3390/cells13090779
17. Duarte LF, Carbone-Schellman J, Bueno SM, Kalergis AM, Riedel CA, and González PA. Tackling cutaneous herpes simplex virus disease with topical immunomodulators—a call to action. Clin Microbiol Rev. (2025) 38. doi: 10.1128/cmr.00147-24
18. Duarte LF, Reyes A, Farías MA, Riedel CA, Bueno SM, Kalergis AM, et al. Crosstalk between epithelial cells, neurons and immune mediators in HSV-1 skin infection. Front Immunol. (2021) 12:662234. doi: 10.3389/fimmu.2021.662234
19. Duarte LF, Farías MA, Álvarez DM, Bueno SM, Riedel CA, and González PA. Herpes simplex virus type 1 infection of the central nervous system: Insights into proposed interrelationships with neurodegenerative disorders. Front Cell Neurosci. (2019) 13:46. doi: 10.3389/fncel.2019.00046
20. Marques CP, Cheeran MC-J, Palmquist JM, Hu S, Urban SL, and Lokensgard JR. Prolonged microglial cell activation and lymphocyte infiltration following experimental herpes encephalitis. J Immunol. (2008) 181:6417–26. doi: 10.4049/jimmunol.181.9.6417
21. Terry RL, Getts DR, Deffrasnes C, van Vreden C, Campbell IL, and King NJC. Inflammatory monocytes and the pathogenesis of viral encephalitis. J Neuroinflamm. (2012) 9:270. doi: 10.1186/1742-2094-9-270
22. Marcocci ME, Napoletani G, Protto V, Kolesova O, Piacentini R, Li Puma DD, et al. Herpes simplex virus-1 in the brain: the dark side of a sneaky infection. Trends Microbiol. (2020) 28:808–20. doi: 10.1016/j.tim.2020.03.003
23. Mancini M and Vidal SM. Insights into the pathogenesis of herpes simplex encephalitis from mouse models. Mamm Genome. (2018) 29:425–45. doi: 10.1007/s00335-018-9772-5
24. Lokensgard JR, Hu S, Sheng W, vanOijen M, Cox D, Cheeran MCJ, et al. Robust expression of TNF-α, IL-1β, RANTES, and IP-10 by human microglial cells during nonproductive infection with herpes simplex virus. J Neurovirol. (2001) 7. doi: 10.1080/13550280152403254
25. Laval K and Enquist LW. The potential role of herpes simplex virus type 1 and neuroinflammation in the pathogenesis of Alzheimer’s disease. Front Neurol. (2021) 12:658695. doi: 10.3389/fneur.2021.658695
26. Duarte LF, Altamirano-Lagos MJ, Tabares-Guevara JH, Opazo MC, Díaz M, Navarrete R, et al. Asymptomatic herpes simplex virus type 1 infection causes an earlier onset and more severe experimental autoimmune encephalomyelitis. Front Immunol. (2021) 12:635257. doi: 10.3389/fimmu.2021.635257
27. Duarte LF, Gatica S, Castillo A, Kalergis AM, Bueno SM, Riedel CA, et al. Is there a role for herpes simplex virus type 1 in multiple sclerosis? Microbes Infect. (2023) 25:105084. doi: 10.1016/j.micinf.2022.105084
28. Duarte LF, Villalobos V, Farías MA, Rangel-Ramírez M, González-Madrid E, Navarro AJ, et al. Asymptomatic herpes simplex virus brain infection elicits cellular senescence phenotypes in the central nervous system of mice suffering multiple sclerosis-like disease. Commun Biol. (2024) 7:811. doi: 10.1038/s42003-024-06486-x
29. Gurgel Assis MS, Fernandes Pedrosa TC, de Moraes FS, Caldeira TG, Pereira GR, de Souza J, et al. Novel insights to enhance therapeutics with acyclovir in the management of herpes simplex encephalitis. J Pharm Sci. (2021) 110:1557–71. doi: 10.1016/j.xphs.2021.01.003
30. Álvarez DM, Castillo E, Duarte LF, Arriagada J, Corrales N, Farías MA, et al. Current antivirals and novel botanical molecules interfering with herpes simplex virus infection. Front Microbiol. (2020) 11:139. doi: 10.3389/fmicb.2020.00139
31. Reichardt SD, Amouret A, Muzzi C, Vettorazzi S, Tuckermann JP, Lühder F, et al. The role of glucocorticoids in inflammatory diseases. Cells. (2021) 10:2921. doi: 10.3390/cells10112921
32. Chastain DB, Spradlin M, Ahmad H, and Henao-Martínez AF. Unintended consequences: risk of opportunistic infections associated with long-term glucocorticoid therapies in adults. Clin Infect Dis. (2024) 78:e37–56. doi: 10.1093/cid/ciad474
33. Farías M, Duarte L, Tognarelli E, and González P. Herpes simplex virus interference with immunity: Focus on dendritic cells. Virulence. (2021) 12:2583–607. doi: 10.1080/21505594.2021.1980990
34. Farías MA, Cancino FA, Navarro AJ, Duarte LF, Soto AA, Tognarelli EI, et al. HSV-1 alters lipid metabolism and induces lipid droplet accumulation in functionally impaired mouse dendritic cells. iScience. (2025) 28:112441. doi: 10.1016/j.isci.2025.112441
35. Tabares-Guevara JH, Jaramillo JC, Ospina-Quintero L, Piedrahíta-Ochoa CA, García-Valencia N, Bautista-Erazo DE, et al. IL-10-dependent amelioration of chronic inflammatory disease by microdose subcutaneous delivery of a prototypic immunoregulatory small molecule. Front Immunol. (2021) 12:708955. doi: 10.3389/fimmu.2021.708955
36. Tognarelli EI, Duarte LF, Farías MA, Cancino FA, Corrales N, Ibáñez FJ, et al. Heme oxygenase-1 expression in dendritic cells contributes to protective immunity against herpes simplex virus skin infection. Antioxidants. (2023) 12:1170. doi: 10.3390/antiox12061170
37. Tognarelli EI, Retamal-Díaz A, Farías MA, Duarte LF, Palomino TF, Ibañez FJ, et al. Pharmacological inhibition of IRE-1 alpha activity in herpes simplex virus type 1 and type 2-infected dendritic cells enhances T cell activation. Front Immunol. (2022) 12:764861. doi: 10.3389/fimmu.2021.764861
38. Bjornevik K, Cortese M, Healy BC, Kuhle J, Mina MJ, Leng Y, et al. Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis. Sci (1979). (2022) 375:296–301. doi: 10.1126/science.abj8222
39. Hassani A, Reguraman N, Shehab S, and Khan G. Primary peripheral Epstein-Barr virus infection can lead to CNS infection and neuroinflammation in a rabbit model: implications for multiple sclerosis pathogenesis. Front Immunol. (2021) 12:764937. doi: 10.3389/fimmu.2021.764937
40. Soni N, Ora M, Singh R, Mehta P, Agarwal A, and Bathla G. Unpacking the CNS manifestations of Epstein-Barr virus: an imaging perspective. Am J Neuroradiol. (2023) 44:1002–8. doi: 10.3174/ajnr.A7945
41. Vrzalikova K, Sunmonu T, Reynolds G, and Murray P. Contribution of Epstein–Barr virus latent proteins to the pathogenesis of classical hodgkin lymphoma. Pathogens. (2018) 7:59. doi: 10.3390/pathogens7030059
42. Smatti MK, Al-Sadeq DW, Ali NH, Pintus G, Abou-Saleh H, and Nasrallah GK. Epstein-barr virus epidemiology, serology, and genetic variability of LMP-1 oncogene among healthy population: An update. Front Oncol. (2018) 8:211. doi: 10.3389/fonc.2018.00211
43. Zhang N, Zuo Y, Jiang L, Peng Y, Huang X, and Zuo L. Epstein-Barr virus and neurological diseases. Front Mol Biosci. (2022) 8:816098. doi: 10.3389/fmolb.2021.816098
44. Lee GH, Kim J, Kim HW, and Cho JW. Clinical significance of Epstein-Barr virus in the cerebrospinal fluid of immunocompetent patients. Clin Neurol Neurosurg. (2021) 202. doi: 10.1016/j.clineuro.2021.106507
45. Kleines M, Schiefer J, Stienen A, Blaum M, Ritter K, and Häusler M. Expanding the spectrum of neurological disease associated with Epstein-Barr virus activity. Eur J Clin Microbiol Infect Dis. (2011) 30:1561–9. doi: 10.1007/s10096-011-1261-7
46. Weinberg A, Li S, Palmer M, and Tyler KL. Quantitative CSF PCR in Epstein-Barr virus infections of the central nervous system. Ann Neurol. (2002) 52:543–8. doi: 10.1002/ana.10321
47. Jha HC, Mehta D, Lu J, El-Naccache D, Shukla SK, Kovacsics C, et al. Erratum: Gammaherpesvirus infection of human neuronal cells (mBio, (2015) 6(6), e01844-15, 10.1128/mBio.01844-15). mBio. (2021) 12:e01844-15. doi: 10.1128/mBio.02981-20
48. Rocamonde B, Hasan U, Mathieu C, and Dutartre H. Viral-induced neuroinflammation: Different mechanisms converging to similar exacerbated glial responses. Front Neurosci. (2023) 17:1108212. doi: 10.3389/fnins.2023.1108212
49. Tiwari D, Singh VK, Baral B, Pathak DK, Jayabalan J, Kumar R, et al. Indication of neurodegenerative cascade initiation by amyloid-like aggregate-forming EBV proteins and peptide in Alzheimer’s disease. ACS Chem Neurosci. (2021) 12:3957–67. doi: 10.1021/acschemneuro.1c00584
50. Gate D, Saligrama N, Leventhal O, Yang AC, Unger MS, Middeldorp J, et al. Clonally expanded CD8 T cells patrol the cerebrospinal fluid in Alzheimer’s disease. Nature. (2020) 577:399–404. doi: 10.1038/s41586-019-1895-7
51. Woulfe J, Gray MT, Ganesh MS, and Middeldorp JM. Human serum antibodies against EBV latent membrane protein 1 cross-react with a-synuclein. Neurol Neuroimmunol Neuroinflamm. (2016) 3:e239. doi: 10.1212/NXI.0000000000000239
52. Woulfe J, Hoogendoorn H, Tarnopolsky M, and Muñoz DG. Monoclonal antibodies against Epstein-Barr virus cross-react with α-synuclein in human brain. Neurology. (2000) 55:1398–401. doi: 10.1212/WNL.55.9.1398
53. Drosu N, Giovannoni G, Pohl D, Hawkes C, Lechner-Scott J, and Levy M. Probing the association between Multiple Sclerosis and Epstein Barr Virus from a therapeutic perspective. Mult Scler Relat Disord. (2021) 52. doi: 10.1016/j.msard.2021.103087
54. Munger KL, Levin LI, O’Reilly EJ, Falk KI, and Ascherio A. Anti-Epstein-Barr virus antibodies as serological markers of multiple sclerosis: A prospective study among United States military personnel. Multiple Sclerosis J. (2011) 17:1185–93. doi: 10.1177/1352458511408991
55. Ioannides ZA, Csurhes PA, Douglas NL, Mackenroth G, Swayne A, Thompson KM, et al. Sustained clinical improvement in a subset of patients with progressive multiple sclerosis treated with Epstein–Barr virus-specific T cell therapy. Front Neurol. (2021) 12:652811. doi: 10.3389/fneur.2021.652811
56. Knockaert DC, Visser LG, Ceuppens JL, Goubau PF, and Bobbaers HJ. Cimetidine is not more effective than placebo in acute infectious mononucleosis. Acta Clin Belg. (1993) 48:213–4. doi: 10.1080/17843286.1993.11718310
57. Gorres KL, Daigle D, Mohanram S, McInerney GE, Lyons DE, and Miller G. Valpromide inhibits Lytic cycle reactivation of epstein-Barr virus. mBio. (2016) 7:e00113. doi: 10.1128/mBio.00113-16
58. Charalampos S, Gontika M, Katsavos S, Velonakis G, Toulas P, Anagnostouli M, et al. Multiple sclerosis and subsequent human immunodeficiency virus infection: A case with the rare comorbidity, focus on novel treatment issues and review of the literature. In Vivo (Brooklyn). (2018) 31:1041–46. doi: 10.21873/invivo.11167
59. Li N, Yang L, Qi XK, Lin YX, Xie X, He GP, et al. BET bromodomain inhibitor JQ1 preferentially suppresses EBV-positive nasopharyngeal carcinoma cells partially through repressing c-Myc article. Cell Death Dis. (2018) 9:761. doi: 10.1038/s41419-018-0789-1
60. Keck KM, Moquin SA, He A, Fernandez SG, Somberg JJ, Liu SM, et al. Bromodomain and extraterminal inhibitors block the Epstein-Barr virus lytic cycle at two distinct steps. J Biol Chem. (2017) 292:13284–295. doi: 10.1074/jbc.M116.751644
61. Pennisi R, Trischitta P, Costa M, Venuti A, Tamburello MP, and Sciortino MT. Update of natural products and their derivatives targeting Epstein–Barr infection. Viruses. (2024) 16:124. doi: 10.3390/v16010124
62. Chung YS, Lam CY, Tan PH, Tsang HF, and Wong SCC. Comprehensive review of COVID-19: epidemiology, pathogenesis, advancement in diagnostic and detection techniques, and post-pandemic treatment strategies. Int J Mol Sci. (2024) 25:8155. doi: 10.3390/ijms25158155
63. Priyal, Sehgal V, Kapila S, Taneja R, Mehmi P, and Gulati N. Review of neurological manifestations of SARS-CoV-2. Cureus. (2023) 15(4):e38194. doi: 10.7759/cureus.38194
64. Efstathiou V, Stefanou M-I, Demetriou M, Siafakas N, Makris M, Tsivgoulis G, et al. Long COVID and neuropsychiatric manifestations (Review). Exp Ther Med. (2022) 23:363. doi: 10.3892/etm.2022.11290
65. Jaywant A, Vanderlind WM, Alexopoulos GS, Fridman CB, Perlis RH, and Gunning FM. Frequency and profile of objective cognitive deficits in hospitalized patients recovering from COVID-19. Neuropsychopharmacology. (2021) 46:2235–40. doi: 10.1038/s41386-021-00978-8
66. Lu Y, Li X, Geng D, Mei N, Wu PY, Huang CC, et al. Cerebral micro-structural changes in COVID-19 patients - an MRI-based 3-month follow-up study. EClinicalMedicine. (2020) 25:100484. doi: 10.1016/j.eclinm.2020.100484
67. Klein R, Soung A, Sissoko C, Nordvig A, Canoll P, Mariani M, et al. COVID-19 induces neuroinflammation and loss of hippocampal neurogenesis. Res Sq. (2021) 1:21. doi: 10.21203/rs.3.rs-1031824/v1
68. Najjar S, Najjar A, Chong DJ, Pramanik BK, Kirsch C, Kuzniecky RI, et al. Central nervous system complications associated with SARS-CoV-2 infection: integrative concepts of pathophysiology and case reports. J Neuroinflamm. (2020) 17. doi: 10.1186/s12974-020-01896-0
69. Matschke J, Lütgehetmann M, Hagel C, Sperhake JP, Schröder AS, Edler C, et al. Neuropathology of patients with COVID-19 in Germany: a post-mortem case series. Lancet Neurol. (2020) 19:919–29. doi: 10.1016/S1474-4422(20)30308-2
70. Bodro M, Compta Y, Llansó L, Esteller D, Doncel-Moriano A, Mesa A, et al. Increased CSF levels of IL-1β, IL-6, and ACE in SARS-CoV-2–associated encephalitis. Neurol Neuroimmunol Neuroinflamm. (2020) 7:e821. doi: 10.1212/NXI.0000000000000821
71. Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, et al. Erratum: Structural and functional features of central nervous system lymphatic vessels (Nature (2015) 523 (337-341) DOI: 10.1038/nature14432). Nature. (2016) 533. doi: 10.1038/nature16999
72. Yarlagadda A, Preston SL, Jeyadhas RP, Lang AE, Hammamieh R, and Clayton AH. Blood–brain barrier: Covid-19, pandemics, and cytokine norms. Innov Clin Neurosci. (2021) 18:21–3.
73. Watson OJ, Barnsley G, Toor J, Hogan AB, Winskill P, and Ghani AC. Global impact of the first year of COVID-19 vaccination: a mathematical modelling study. Lancet Infect Dis. (2022) 22:1293–302. doi: 10.1016/S1473-3099(22)00320-6
74. Martínez-Balboa Y, Rodríguez-Guilarte L, Méndez C, Ríos M, Rivera DB, Moreno-Tapia D, et al. Age and primary vaccination schedule impact humoral and cellular immunity with an inactivated SARS-CoV-2 vaccine. iScience. (2025) 28(8):113167. doi: 10.1016/j.isci.2025.113167
75. Duarte LF, Vázquez Y, Diethelm-Varela B, Pavez V, Berríos-Rojas R, Méndez C, et al. Differential severe acute respiratory syndrome coronavirus 2–specific humoral response in inactivated virus–vaccinated, convalescent, and breakthrough-infected subjects. J Infect Dis. (2023) 228:857–67. doi: 10.1093/infdis/jiad320
76. Soto JA, Melo-González F, Gutierrez-Vera C, Schultz BM, Berríos-Rojas RV, Rivera-Pérez D, et al. Inactivated vaccine-induced SARS-CoV-2 variant-specific immunity in children. mBio. (2022) 13:e0131122. doi: 10.1128/mbio.01311-22
77. Schultz BM, Melo-González F, Duarte LF, Gálvez NMS, Pacheco GA, Soto JA, et al. A booster dose of CoronaVac increases neutralizing antibodies and T cells that recognize delta and omicron variants of concern. mBio. (2022) 13:e0142322. doi: 10.1128/mbio.01423-22
78. Gálvez NM, Pacheco GA, Schultz BM, Melo-González F, Soto JA, Duarte LF, et al. Differences in the immune response elicited by two immunization schedules with an inactivated SARS-CoV-2 vaccine in a randomized phase 3 clinical trial. Elife. (2022) 11:e81477. doi: 10.7554/eLife.81477
79. Méndez C, Peñaloza HF, Schultz BM, Piña-Iturbe A, Ríos M, Moreno-Tapia D, et al. Humoral and cellular response induced by a second booster of an inactivated SARS-CoV-2 vaccine in adults. EBioMedicine. (2023) 91:104563. doi: 10.1016/j.ebiom.2023.104563
80. Sarhan RM, Harb HS, Abou Warda AE, Salem-Bekhit MM, Shakeel F, Alzahrani SA, et al. Efficacy of the early treatment with tocilizumab-hydroxychloroquine and tocilizumab-remdesivir in severe COVID-19 Patients. J Infect Public Health. (2022) 15:116–22. doi: 10.1016/j.jiph.2021.10.024
81. Vulturar D-M, Neag MA, Vesa Ştefan C, Maierean A-D, Gherman D, Buzoianu AD, et al. Therapeutic efficacy and outcomes of remdesivir versus remdesivir with tocilizumab in severe SARS-CoV-2 infection. Int J Mol Sci. (2022) 23:14462. doi: 10.3390/ijms232214462
82. Bonilla H, Peluso MJ, Rodgers K, Aberg JA, Patterson TF, Tamburro R, et al. Therapeutic trials for long COVID-19: A call to action from the interventions taskforce of the RECOVER initiative. Front Immunol. (2023) 14:1129459. doi: 10.3389/fimmu.2023.1129459
83. Bekker LG, Beyrer C, Mgodi N, Lewin SR, Delany-Moretlwe S, Taiwo B, et al. Author correction: HIV infection (Nature reviews disease primers, (2023), 9, 1, (42), 10.1038/s41572-023-00452-3). Nat Rev Dis Primers. (2023) 9. doi: 10.1038/s41572-023-00464-z
84. Chang CC, Crane M, Zhou J, Mina M, Post JJ, Cameron BA, et al. HIV and co-infections. Immunol Rev. (2013) 254:114–42. doi: 10.1111/imr.12063
85. Heaton RK, Franklin DR, Ellis RJ, McCutchan JA, Letendre SL, LeBlanc S, et al. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: Differences in rates, nature, and predictors. J Neurovirol. (2011) 17:3–16. doi: 10.1007/s13365-010-0006-1
86. Killingsworth L and Spudich S. Neuropathogenesis of HIV-1: insights from across the spectrum of acute through long-term treated infection. Semin Immunopathol. (2022) 44:709–24. doi: 10.1007/s00281-022-00953-5
87. Williams ME and Naudé PJW. The relationship between HIV-1 neuroinflammation, neurocognitive impairment and encephalitis pathology: A systematic review of studies investigating post-mortem brain tissue. Rev Med Virol. (2024) 34:e2519. doi: 10.1002/rmv.2519
88. Bethel-Brown C, Yao H, Hu G, and Buch S. Platelet-derived growth factor (PDGF)-BB-mediated induction of monocyte chemoattractant protein 1 in human astrocytes: Implications for HIV-associated neuroinflammation. J Neuroinflamm. (2012) 9. doi: 10.1186/1742-2094-9-262
89. Genis P, Jett M, Bernton EW, Boyle T, Gelbard HA, Dzenko K, et al. Cytokines and arachidonic metabolites produced during human immunodeficiency virus (HIV)-infected macrophage-astroglia interactions: implications for the neuropathogenesis of HIV disease. J Exp Med. (1992) 176:1703–18. doi: 10.1084/jem.176.6.1703
90. Yeh MW, Kaul M, Zheng J, Nottet HSLM, Thylin M, Gendelman HE, et al. Cytokine-stimulated, but not HIV-infected, human monocyte-derived macrophages produce neurotoxic levels of l -cysteine. J Immunol. (2000) 164:4265–70. doi: 10.4049/jimmunol.164.8.4265
91. Chivero ET, Guo M-L, Periyasamy P, Liao K, Callen SE, and Buch S. HIV-1 tat primes and activates microglial NLRP3 inflammasome-mediated neuroinflammation. J Neurosci. (2017) 37:3599–609. doi: 10.1523/JNEUROSCI.3045-16.2017
92. Thangaraj A, Periyasamy P, Liao K, Bendi VS, Callen S, Pendyala G, et al. HIV-1 TAT-mediated microglial activation: role of mitochondrial dysfunction and defective mitophagy. Autophagy. (2018) 14:1596–619. doi: 10.1080/15548627.2018.1476810
93. Shenoy A, Marwaha PK, and Worku DA. CD8 encephalitis in HIV: A review of this emerging entity. J Clin Med. (2023) 12:770. doi: 10.3390/jcm12030770
94. Gannon P, Khan MZ, and Kolson DL. Current understanding of HIV-associated neurocognitive disorders pathogenesis. Curr Opin Neurol. (2011) 24:275–83. doi: 10.1097/WCO.0b013e32834695fb
95. Schmitt FA, Bigley JW, McKinnis R, Logue PE, Evans RW, and Drucker JL. Neuropsychological outcome of zidovudine (AZT) treatment of patients with AIDS and AIDS-related complex. New Engl J Med. (1988) 319:1573–8. doi: 10.1056/NEJM198812153192404
96. Robertson KR, Robertson WT, Ford S, Watson D, Fiscus S, Harp AG, et al. Highly active antiretroviral therapy improves neurocognitive functioning. JAIDS J Acquired Immune Deficiency Syndromes. (2004) 36:562–6. doi: 10.1097/00126334-200405010-00003
97. Anderson AM, Jang JH, Easley KA, Fuchs D, Gisslen M, Zetterberg H, et al. Cognitive and neuronal link with inflammation: A longitudinal study in people with and without HIV infection. JAIDS J Acquired Immune Deficiency Syndromes. (2020) 85:617–25. doi: 10.1097/QAI.0000000000002484
98. Matsuoka M and Jeang KT. Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nat Rev Cancer. (2007) 7:270–80. doi: 10.1038/nrc2111
99. Sampaio GCL, Ribeiro JR, De Almeida CN, Boa-Sorte N, Galvão-Castro B, Grassi MFR, et al. Human T cell lymphotropic virus type 1 global prevalence associated with the human development index: systematic review with meta-analysis. AIDS Res Hum Retroviruses. (2023) 39:145–65. doi: 10.1089/aid.2021.0230
100. Okochi K and Sato H. Transmission of ATLV (HTLV-I) through blood transfusion. Uirusu J Virol. (1986) 36:129–35. doi: 10.2222/jsv.36.195
101. Kaplan JE, Khabbaz RF, Murphy EL, Hermansen S, Roberts C, Lal R, et al. Male-to-female transmission of human T-cell lymphotropic virus types I and II: association with viral load. The Retrovirus Epidemiology Donor Study Group. J Acquir Immune Defic Syndr Hum Retrovirol. (1996) 12:193–201. doi: 10.1097/00042560-199606010-00014
102. Hino S. Establishment of the milk-borne transmission as a key factor for the peculiar endemicity of human T-lymphotropic virus type 1 (HTLV-1): The ATL prevention program nagasaki. Proc Jpn Acad Ser B Phys Biol Sci. (2011) 87:152–66. doi: 10.2183/pjab.87.152
103. Lepoutre V, Jain P, Quann K, Wigdahl B, and Khan ZK. Role of resident CNS cell populations in HTLV-1-associated neuroinflammatory disease. Front Biosci. (2009) 14:1152–68. doi: 10.2741/3300
104. Yamauchi J, Araya N, Yagishita N, Sato T, and Yamano Y. An update on human T-cell leukemia virus type I (HTLV-1)-associated myelopathy/tropical spastic paraparesis (HAM/TSP) focusing on clinical and laboratory biomarkers. Pharmacol Ther. (2021) 218:107669. doi: 10.1016/j.pharmthera.2020.107669
105. Osame M, Usuku K, Izumo S, Ijichi N, Amitani H, Igata A, et al. HTLV-I associated myelopathy, a new clinical entity. Lancet. (1986) 327:1031–2. doi: 10.1016/S0140-6736(86)91298-5
106. Gessain A, Vernant JC, Maurs L, Barin F, Gout O, Calender A, et al. Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet. (1985) 326:407–10. doi: 10.1016/S0140-6736(85)92734-5
107. Igakura T, Stinchcombe JC, Goon PKC, Taylor GP, Weber JN, Griffiths GM, et al. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Sci (1979). (2003) 299:1713–6. doi: 10.1126/science.1080115
108. Bangham CRM, Araujo A, Yamano Y, and Taylor GP. CORRIGENDUM to: HTLV-1-associated myelopathy/tropical spastic paraparesis (Nat. Rev. Dis. Primers, 15012; 10.1038/nrdp.2015.12). Nat Rev Dis Primers. (2015) 1:15012. doi: 10.1038/NRDP.2015.23
109. Omsland M, Pise-Masison C, Fujikawa D, Galli V, Fenizia C, Parks RW, et al. Inhibition of tunneling nanotube (TNT) formation and human T-cell leukemia virus type 1 (HTLV-1) transmission by cytarabine. Sci Rep. (2018) 8:11118. doi: 10.1038/s41598-018-29391-w
110. Zarei Ghobadi M, Emamzadeh R, Teymoori-Rad M, and Mozhgani S-H. Decoding pathogenesis factors involved in the progression of ATLL or HAM/TSP after infection by HTLV-1 through a systems virology study. Virol J. (2021) 18:175. doi: 10.1186/s12985-021-01643-8
111. Maher AK, Aristodemou A, Giang N, Tanaka Y, Bangham CRM, Taylor GP, et al. HTLV-1 induces an inflammatory CD4+CD8+ T cell population in HTLV-1–associated myelopathy. JCI Insight. (2024) 9:e173738. doi: 10.1172/jci.insight.173738
112. Shakudo M, Inoue Y, and Tsutada T. HTLV-I-associated myelopathy: Acute progression and atypical MR findings. Am J Neuroradiol. (1999) 20:1417–21.
113. Araujo A, Bangham CRM, Casseb J, Gotuzzo E, Jacobson S, Martin F, et al. Management of HAM/TSP: systematic review and consensus-based recommendations 2019. Neurol Clin Pract. (2021) 11:49–56. doi: 10.1212/CPJ.0000000000000832
114. Marino-Merlo F, Balestrieri E, Matteucci C, Mastino A, Grelli S, and Macchi B. Antiretroviral therapy in HTLV-1 infection: An updated overview. Pathogens. (2020) 9:342. doi: 10.3390/pathogens9050342
115. Nakashima H, Matsui T, Harada S, Kobayashi N, Matsuda A, Ueda T, et al. Inhibition of replication and cytopathic effect of human T cell lymphotropic virus type III/lymphadenopathy-associated virus by 3’-azido-3’-deoxythymidine in vitro. Antimicrob Agents Chemother. (1986) 30:933–7. doi: 10.1128/AAC.30.6.933
116. Matsushita S, Mitsuya H, Reitz MS, and Broder S. Pharmacological inhibition of in vitro infectivity of human T lymphotropic virus type I. J Clin Invest. (1987) 80:394–400. doi: 10.1172/JCI113085
117. Isono T, Ogawa K, and Seto A. Antiviral effect of zidovudine in the experimental model of adult T cell leukemia in rabbits. Leuk Res. (1990) 14:841–7. doi: 10.1016/0145-2126(90)90172-6
118. Sheremata WA, Benedict D, Squilacote DC, Sazant A, and DeFreitas E. High-dose zidovudine induction in HTLV-I-associated myelopathy. Neurology. (1993) 43:2125–9. doi: 10.1212/wnl.43.10.2125
119. Gout O, Gessain A, Iba-Zizen M, Kouzan S, Bolgert F, de Thé G, et al. The effect of zidovudine on chronic myelopathy associated with HTLV-1. J Neurol. (1991) 238:108–9. doi: 10.1007/BF00315691
120. Kawaoka Y. Equine H7N7 influenza A viruses are highly pathogenic in mice without adaptation: potential use as an animal model. J Virol. (1991) 65:3891–4. doi: 10.1128/jvi.65.7.3891-3894.1991
121. Balestrieri E, Sciortino MT, Mastino A, and Macchi B. Protective effect of the acyclic nucleoside phosphonate tenofovir toward human T-cell leukemia/lymphotropic virus type 1 infection of human peripheral blood mononuclear cells in vitro. Antiviral Res. (2005) 68:154–62. doi: 10.1016/j.antiviral.2005.09.001
122. Taylor GP, Goon P, Furukawa Y, Green H, Barfield A, Mosley A, et al. Zidovudine plus lamivudine in Human T-Lymphotropic Virus type-l-associated myelopathy: A randomised trial. Retrovirology. (2006) 3:63. doi: 10.1186/1742-4690-3-63
123. Pierson TC and Diamond MS. The emergence of Zika virus and its new clinical syndromes. Nature. (2018) 560:573–81. doi: 10.1038/s41586-018-0446-y
124. Sirohi D and Kuhn RJ. Zika virus structure, maturation, and receptors. J Infect Dis. (2017) 216:S935–944. doi: 10.1093/infdis/jix515
125. Rasmussen SA, Jamieson DJ, Honein MA, and Petersen LR. Zika virus and birth defects — Reviewing the evidence for causality. New Engl J Med. (2016) 374:1981–7. doi: 10.1056/nejmsr1604338
126. de Araújo TVB, Rodrigues LC, de Alencar Ximenes RA, de Barros Miranda-Filho D, Montarroyos UR, de Melo APL, et al. Association between Zika virus infection and microcephaly in Brazil, January to May, 2016: preliminary report of a case-control study. Lancet Infect Dis. (2016) 16:1356–63. doi: 10.1016/S1473-3099(16)30318-8
127. Zheng QL, Duan T, and Jin LP. Single-cell RNA expression profiling of ACE2 and AXL in the human maternal-Fetal interface. Reprod Dev Med. (2020) 4:7–10. doi: 10.4103/2096-2924.278679
128. Rolfe AJ, Bosco DB, Wang J, Nowakowski RS, Fan J, and Ren Y. Bioinformatic analysis reveals the expression of unique transcriptomic signatures in Zika virus infected human neural stem cells. Cell Biosci. (2016) 6:42. doi: 10.1186/s13578-016-0110-x
129. Muñoz LS, Parra B, and Pardo CA. Neurological implications of zika virus infection in adults. J Infect Dis. (2017) 216(suppl_10):S897–905. doi: 10.1093/infdis/jix511
130. Azevedo RSS, De Sousa JR, Araujo MTF, Martins Filho AJ, De Alcantara BN, Araujo FMC, et al. In situ immune response and mechanisms of cell damage in central nervous system of fatal cases microcephaly by Zika virus. Sci Rep. (2018) 8:1. doi: 10.1038/s41598-017-17765-5
131. Kaczmarek A, Vandenabeele P, and Krysko DV. Necroptosis: the release of damage-associated molecular patterns and its physiological relevance. Immunity. (2013) 38:209–23. doi: 10.1016/j.immuni.2013.02.003
132. Papa MP, Meuren LM, Coelho SVA, Lucas CG de O, Mustafá YM, Lemos Matassoli F, et al. Zika virus infects, activates, and crosses brain microvascular endothelial cells, without barrier disruption. Front Microbiol. (2017) 8:2557. doi: 10.3389/fmicb.2017.02557
133. Lum F-M, Low DKS, Fan Y, Tan JJL, Lee B, Chan JKY, et al. Zika virus infects human fetal brain microglia and induces inflammation. Clin Infect Dis. (2017) 64:914–20. doi: 10.1093/cid/ciw878
134. Srivastava KS, Jeswani V, Pal N, Bohra B, Vishwakarma V, Bapat AA, et al. Japanese encephalitis virus: an update on the potential antivirals and vaccines. Vaccines (Basel). (2023) 11:742. doi: 10.3390/vaccines11040742
135. Quan TM, Thao TTN, Duy NM, Nhat TM, and Clapham HE. Estimates of the global burden of Japanese encephalitis and the impact of vaccination from 2000-2015. Elife. (2020) 9:e51027. doi: 10.7554/eLife.51027
136. Chauhan S, Rathore DK, Sachan S, Lacroix-Desmazes S, Gupta N, Awasthi A, et al. Japanese encephalitis virus infected human monocyte-derived dendritic cells activate a transcriptional network leading to an antiviral inflammatory response. Front Immunol. (2021) 12:638694. doi: 10.3389/fimmu.2021.638694
137. Li F, Wang Y, Yu L, Cao S, Wang K, Yuan J, et al. Viral infection of the central nervous system and neuroinflammation precede blood-brain barrier disruption during Japanese encephalitis virus infection. J Virol. (2015) 89:5602–14. doi: 10.1128/jvi.00143-15
138. Chang CY, Wu CC, Wang JD, Li JR, Wang YY, Lin SY, et al. DHA attenuated Japanese Encephalitis virus infection-induced neuroinflammation and neuronal cell death in cultured rat Neuron/glia. Brain Behav Immun. (2021) 93:194–205. doi: 10.1016/j.bbi.2021.01.012
139. Ahmad F, Ahmad S, Husain A, Pandey N, Khubaib M, and Sharma R. Role of inflammatory cytokine burst in neuro-invasion of Japanese Encephalitis virus infection: an immunotherapeutic approaches. J Neurovirol. (2024) 30:251–65. doi: 10.1007/s13365-024-01212-z
140. Singh A, Kulshreshtha R, and Mathur A. Secretion of the chemokine interleukin-8 during Japanese encephalitis virus infection. J Med Microbiol. (2000) 49:607–12. doi: 10.1099/0022-1317-49-7-607
141. Bhowmick S, Duseja R, Das S, Appaiahgiri MB, Vrati S, and Basu A. Induction of IP-10 (CXCL10) in astrocytes following Japanese encephalitis. Neurosci Lett. (2007) 414:45–50. doi: 10.1016/j.neulet.2006.11.070
142. Khanna N, Agnihotri M, Mathur A, and Chaturvedi UC. Neutrophil chemotactic factor produced by Japanese encephalitis virus stimulated macrophages. Clin Exp Immunol. (1991) 86:299–303. doi: 10.1111/j.1365-2249.1991.tb05813.x
143. Ding D, Hong Z, Zhao S, Clemens JD, Zhou B, Wang B, et al. Long-term disability from acute childhood Japanese encephalitis in Shanghai, China. Am J Trop Med Hyg. (2007) 77:528–33. doi: 10.4269/ajtmh.2007.77.528
144. Yin Z, Wang X, Li L, Li H, Zhang X, Li J, et al. Neurological sequelae of hospitalized Japanese encephalitis cases in Gansu Province, China. Am Soc Trop Med Hyg. (2015) 92:1125–9. doi: 10.4269/ajtmh.14-0148
145. Laidlow TA, Johnston ES, Zadoks RN, Walsh M, Viana M, Wiley KE, et al. Scoping review of Japanese encephalitis virus transmission models. Transbound Emerg Dis. (2025) 2025:9880670. doi: 10.1155/tbed/9880670
146. Ajibowo AO, Ortiz JF, Alli A, Halan T, and Kolawole OA. Management of Japanese encephalitis: A current update. Cureus. (2021) 13(4):e14579. doi: 10.7759/cureus.14579
147. Singh DAK, Mehta DA, Kushwaha DKP, Pandey AK, Mittal DM, Sharma DB, et al. Minocycline trial in Japanese encephalitis: a double blind, randomized placebo study. Pediatr Rev: Int J Pediatr Res. (2016) 3:371–7. doi: 10.17511/ijpr.2016.i05.18
149. Moss WJ and Griffin DE. What’s going on with measles? J Virol. (2024) 98:e0075824. doi: 10.1128/jvi.00758-24
150. Mahase E. Measles: 142 000 people died in 2018, mostly aged under 5. BMJ. (2019) 367:l6830. doi: 10.1136/bmj.l6830
151. Gunasekaran PK and Saini AG. Subacute sclerosing panencephalitis. Semin Pediatr Neurol. (2025) 54:101207. doi: 10.1016/j.spen.2025.101207
152. Parums DV. A review of the resurgence of measles, a vaccine-preventable disease, as current concerns contrast with past hopes for measles elimination. Med Sci Monit. (2024) 30:e944436. doi: 10.12659/MSM.944436
153. Samia P, Oyieke K, Tunje D, Udwadia-Hegde A, Feemster K, Oncel I, et al. Options in the treatment of subacute sclerosing panencephalitis: implications for low resource areas. Curr Treat Options Neurol. (2022) 24:99–110. doi: 10.1007/s11940-022-00710-x
154. Xerri T, Darmanin N, Zammit MA, and Fsadni C. Complications of measles: a case series. BMJ Case Rep. (2020) 13:e232408. doi: 10.1136/bcr-2019-232408
155. Bian P, Ye C, Zheng X, Yang J, Ye W, Wang Y, et al. Mesenchymal stem cells alleviate Japanese encephalitis virus-induced neuroinflammation and mortality. Stem Cell Res Ther. (2017) 8:38. doi: 10.1186/s13287-017-0486-5
156. Harrell CR, Fellabaum C, Jovicic N, Djonov V, Arsenijevic N, and Volarevic V. Molecular mechanisms responsible for therapeutic potential of mesenchymal stem cell-derived secretome. Cells. (2019) 8:467. doi: 10.3390/cells8050467
157. Zhang X, Kuang Q, Xu J, Lin Q, Chi H, and Yu D. MSC-based cell therapy in neurological diseases: A concise review of the literature in pre-clinical and clinical research. Biomolecules. (2024) 14:538. doi: 10.3390/biom14050538
158. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I, Marini FC, Krause DS, et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. (2006) 8:315–7. doi: 10.1080/14653240600855905
159. Lv FJ, Tuan RS, Cheung KMC, and Leung VYL. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells. (2014) 32:1408–19. doi: 10.1002/stem.1681
160. de Vasconcellos MaChado C, da Silva Telles PD, and Nascimento ILO. Immunological characteristics of mesenchymal stem cells. Rev Bras Hematol Hemoter. (2013) 35:62–7. doi: 10.5581/1516-8484.20130017
161. Vizoso FJ, Eiro N, Cid S, Schneider J, and Perez-Fernandez R. Mesenchymal stem cell secretome: Toward cell-free therapeutic strategies in regenerative medicine. Int J Mol Sci. (2017) 18:1852. doi: 10.3390/ijms18091852
162. Harrell CR, Markovic BS, Fellabaum C, Arsenijevic A, and Volarevic V. Mesenchymal stem cell-based therapy of osteoarthritis: Current knowledge and future perspectives. Biomed Pharmacother. (2019) 109:2318–26. doi: 10.1016/j.biopha.2018.11.099
163. Petrenko Y, Vackova I, Kekulova K, Chudickova M, Koci Z, Turnovcova K, et al. A comparative analysis of multipotent mesenchymal stromal cells derived from different sources, with a focus on neuroregenerative potential. Sci Rep. (2020) 10. doi: 10.1038/s41598-020-61167-z
164. Lou S, Duan Y, Nie H, Cui X, Du J, and Yao Y. Mesenchymal stem cells: Biological characteristics and application in disease therapy. Biochimie. (2021) 185. doi: 10.1016/j.biochi.2021.03.003
165. Harrell CR, Jankovic MG, Fellabaum C, Volarevic A, Djonov V, Arsenijevic A, et al. Molecular mechanisms responsible for anti-inflammatory and immunosuppressive effects of mesenchymal stem cell-derived factors. Adv Exp Med Biol. (2019) 1084:187–206. doi: 10.1007/5584_2018_306
166. Elshaer SL, Bahram SH, Rajashekar P, Gangaraju R, and El-Remessy AB. Modulation of mesenchymal stem cells for enhanced therapeutic utility in ischemic vascular diseases. Int J Mol Sci. (2022) 23:249. doi: 10.3390/ijms23010249
167. Huang Y, Wu Q, and Tam PKH. Immunomodulatory mechanisms of mesenchymal stem cells and their potential clinical applications. Int J Mol Sci. (2022) 23:10023. doi: 10.3390/ijms231710023
168. Zhang X, Zhang Y, Peng X, Yang L, Miao J, Yue Y, et al. Targeting neuroinflammation in preterm white matter injury: therapeutic potential of mesenchymal stem cell-derived exosomes. Cell Mol Neurobiol. (2025) 45:23. doi: 10.1007/s10571-025-01540-6
169. Cuesta-Gomez N, Graham GJ, and Campbell JDM. Chemokines and their receptors: predictors of the therapeutic potential of mesenchymal stromal cells. J Transl Med. (2021) 19:156. doi: 10.1186/s12967-021-02822-5
170. Sarsenova M, Kim Y, Raziyeva K, Kazybay B, Ogay V, and Saparov A. Recent advances to enhance the immunomodulatory potential of mesenchymal stem cells. Front Immunol. (2022) 13:1010399. doi: 10.3389/fimmu.2022.1010399
171. Lee BC and Kang KS. Functional enhancement strategies for immunomodulation of mesenchymal stem cells and their therapeutic application. Stem Cell Res Ther. (2020) 11:397. doi: 10.1186/s13287-020-01920-3
172. Seo Y, Kang MJ, and Kim HS. Strategies to potentiate paracrine therapeutic efficacy of mesenchymal stem cells in inflammatory diseases. Int J Mol Sci. (2021) 22:3397. doi: 10.3390/ijms22073397
173. Gavasso S, Kråkenes T, Olsen H, Evjenth EC, Ytterdal M, Haugsøen JB, et al. The therapeutic mechanisms of mesenchymal stem cells in MS—A review focusing on neuroprotective properties. Int J Mol Sci. (2024) 25:1365. doi: 10.3390/ijms25031365
174. Yarahmadi A, Dorri Giv M, Hosseininejad R, Rezaie A, Mohammadi N, Afkhami H, et al. Mesenchymal stem cells and their extracellular vesicle therapy for neurological disorders: traumatic brain injury and beyond. Front Neurol. (2025) 16:1472679. doi: 10.3389/fneur.2025.1472679
175. Santillán-Guaján SM, Shahi MH, and Castresana JS. Mesenchymal-stem-cell-based therapy against gliomas. Cells. (2024) 13:617. doi: 10.3390/cells13070617
176. Yin T, Liu Y, Ji W, Zhuang J, Chen X, Gong B, et al. Engineered mesenchymal stem cell-derived extracellular vesicles: A state-of-the-art multifunctional weapon against Alzheimer’s disease. Theranostics. (2023) 13:1264–85. doi: 10.7150/thno.81860
177. Allegretta C, D’amico E, Manuti V, Avolio C, and Conese M. Mesenchymal stem cell-derived extracellular vesicles and their therapeutic use in central nervous system demyelinating disorders. Int J Mol Sci. (2022) 23:3829. doi: 10.3390/ijms23073829
178. Zhang Y, Liu J, Su M, Wang X, and Xie C. Exosomal microRNA-22-3p alleviates cerebral ischemic injury by modulating KDM6B/BMP2/BMF axis. Stem Cell Res Ther. (2021) 12:111. doi: 10.1186/s13287-020-02091-x
179. Korotkov A, Mills JD, Gorter JA, van Vliet EA, and Aronica E. Systematic review and meta-analysis of differentially expressed miRNAs in experimental and human temporal lobe epilepsy. Sci Rep. (2017) 7:11592. doi: 10.1038/s41598-017-11510-8
180. Garcia G, Pinto S, Ferreira S, Lopes D, Serrador MJ, Fernandes A, et al. Emerging role of miR-21-5p in neuron–glia dysregulation and exosome transfer using multiple models of Alzheimer’s disease. Cells. (2022) 11:3377. doi: 10.3390/cells11213377
181. Coenen-Stass AML, Pauwels MJ, Hanson B, Martin Perez C, Conceição M, Wood MJA, et al. Extracellular microRNAs exhibit sequence-dependent stability and cellular release kinetics. RNA Biol. (2019) 16:696–706. doi: 10.1080/15476286.2019.1582956
182. Liu Y, Zhuang X, Yu S, Yang N, Zeng J, Liu X, et al. Exosomes derived from stem cells from apical papilla promote craniofacial soft tissue regeneration by enhancing Cdc42-mediated vascularization. Stem Cell Res Ther. (2021) 12:76. doi: 10.1186/s13287-021-02151-w
183. Potter DR, Miyazawa BY, Gibb SL, Deng X, Togaratti PP, Croze RH, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate pulmonary vascular permeability and lung injury induced by hemorrhagic shock and trauma. J Trauma Acute Care Surg. (2018) 84:245–56. doi: 10.1097/TA.0000000000001744
184. Pischiutta F, Caruso E, Cavaleiro H, Salgado AJ, Loane DJ, and Zanier ER. Mesenchymal stromal cell secretome for traumatic brain injury: Focus on immunomodulatory action. Exp Neurol. (2022) 357:114199. doi: 10.1016/j.expneurol.2022.114199
185. Zanier ER, Pischiutta F, Rulli E, Vargiolu A, Elli F, Gritti P, et al. MesenchymAl stromal cells for Traumatic bRain Injury (MATRIx): a study protocol for a multicenter, double-blind, randomised, placebo-controlled phase II trial. Intensive Care Med Exp. (2023) 11:56. doi: 10.1186/s40635-023-00535-1
186. Mukai T, Sei K, and Nagamura-Inoue T. Mesenchymal stromal cells perspective: New potential therapeutic for the treatment of neurological diseases. Pharmaceutics. (2021) 13:1159. doi: 10.3390/pharmaceutics13081159
187. Vaheb S, Afshin S, Ghoshouni H, Ghaffary EM, Farzan M, Shaygannejad V, et al. Neurological efficacy and safety of mesenchymal stem cells (MSCs) therapy in people with multiple sclerosis (pwMS): An updated systematic review and meta-analysis. Mult Scler Relat Disord. (2024) 87:105681. doi: 10.1016/j.msard.2024.105681
188. Teixeira FG, Carvalho MM, Sousa N, and Salgado AJ. Mesenchymal stem cells secretome: A new paradigm for central nervous system regeneration? Cell Mol Life Sci. (2013) 70:3871–82. doi: 10.1007/s00018-013-1290-8
189. Salgado AJ, Sousa JC, Costa BM, Pires AO, Mateus-Pinheiro A, Teixeira FG, et al. Mesenchymal stem cells secretome as a modulator of the neurogenic niche: Basic insights and therapeutic opportunities. Front Cell Neurosci. (2015) 9:249. doi: 10.3389/fncel.2015.00249
190. Drago D, Cossetti C, Iraci N, Gaude E, Musco G, Bachi A, et al. The stem cell secretome and its role in brain repair. Biochimie. (2013) 95:2271–85. doi: 10.1016/j.biochi.2013.06.020
191. Łukowicz K, Grygier B, and Basta-Kaim A. Emerging role of neural stem/progenitor cell secretome in brain inflammatory response modulation. Pharmacol Rep. (2025) 77(4):907–20. doi: 10.1007/s43440-025-00733-6
192. Martins LF, Costa RO, Pedro JR, Aguiar P, Serra SC, Teixeira FG, et al. Mesenchymal stem cells secretome-induced axonal outgrowth is mediated by BDNF. Sci Rep. (2017) 7:4153. doi: 10.1038/s41598-017-03592-1
193. Baez-Jurado E, Hidalgo-Lanussa O, Barrera-Bailón B, Sahebkar A, Ashraf GM, Echeverria V, et al. Secretome of mesenchymal stem cells and its potential protective effects on brain pathologies. Mol Neurobiol. (2019) 56:6902–27. doi: 10.1007/s12035-019-1570-x
194. Shaw TD, Krasnodembskaya AD, Schroeder GN, Zumla A, Maeurer M, and O’Kane CM. Mesenchymal stromal cells: an antimicrobial and host-directed therapy for complex infectious diseases. Clin Microbiol Rev. (2021) 34:e0006421. doi: 10.1128/CMR.00064-21
195. Soni N, Tripathi A, Mukherjee S, Gupta S, Mohanty S, Basu A, et al. Bone marrow-derived extracellular vesicles modulate the abundance of infiltrating immune cells in the brain and exert an antiviral effect against the Japanese encephalitis virus. FASEB Bioadv. (2022) 4:798–815. doi: 10.1096/fba.2022-00071
196. Van Linthout S, Savvatis K, Miteva K, Peng J, Ringe J, Warstat K, et al. Mesenchymal stem cells improve murine acute coxsackievirus B3-induced myocarditis. Eur Heart J. (2011) 32:2168–78. doi: 10.1093/eurheartj/ehq467
197. Gu X, Li Y, Chen K, Wang X, Wang Z, Lian H, et al. Exosomes derived from umbilical cord mesenchymal stem cells alleviate viral myocarditis through activating AMPK/mTOR-mediated autophagy flux pathway. J Cell Mol Med. (2020) 24:7515–30. doi: 10.1111/jcmm.15378
198. Chen S, Sun F, Qian H, Xu W, and Jiang J. Preconditioning and engineering strategies for improving the efficacy of mesenchymal stem cell-derived exosomes in cell-free therapy. Stem Cells Int. (2022) 2022:1–18. doi: 10.1155/2022/1779346
199. Chen F, Chen Z, Wu HT, Chen XX, Zhan P, Wei ZY, et al. Mesenchymal stem cell-derived exosomes attenuate murine cytomegalovirus-infected pneumonia via NF-κB/NLRP3 signaling pathway. Viruses. (2024) 16:619. doi: 10.3390/v16040619
200. Klimova RR, Momotyuk ED, Demidova NA, Yarigina EI, and Kushch AA. Mesenchymal stem cells enhance immune response and protectmice against lethal herpes viral infection. Probl Virol. (2018) 63:261–7. doi: 10.18821/0507-4088-2018-63-6-261-267
201. Klimova RR, Demidova NA, Masalova OV, and Kushch AA. Preventive vaccination with mesenchymal stem cells protects mice from lethal infection caused by herpes simplex virus 1. Mol Biol. (2021) 55:413–23. doi: 10.1134/S0026893321020242
202. Lee RH, Pulin AA, Seo MJ, Kota DJ, Ylostalo J, Larson BL, et al. Intravenous hMSCs Improve Myocardial Infarction in Mice because Cells Embolized in Lung Are Activated to Secrete the Anti-inflammatory Protein TSG-6. Cell Stem Cell. (2009) 5:54–63. doi: 10.1016/j.stem.2009.05.003
203. Thanunchai M, Hongeng S, and Thitithanyanont A. Mesenchymal stromal cells and viral infection. Stem Cells Int. (2015) 2015:1–8. doi: 10.1155/2015/860950
204. Heinze A, Grebe B, Bremm M, Huenecke S, Munir TA, Graafen L, et al. The Synergistic Use of IL-15 and IL-21 for the Generation of NK Cells From CD3/CD19-Depleted Grafts Improves Their ex vivo Expansion and Cytotoxic Potential Against Neuroblastoma: Perspective for Optimized Immunotherapy Post Haploidentical Stem Cell Transplantation. Front Immunol. (2019) 10:2816. doi: 10.3389/fimmu.2019.02816
205. Al-Daim SA, Atta OM, Allayeh AK, Hussein HAM, Dawood MA, Khalifa EA, et al. Human wharton-jelly- mesenchymal stem cells microvesicles and exosomes as potential antiviral activity against herpes simplex type 2. J Community Med Public Health Rep. (2024) 5. doi: 10.38207/jcmphr/2024/mar05040332
206. Chandra PK, Gerlach SL, Wu C, Khurana N, Swientoniewski LT, Abdel-Mageed AB, et al. Mesenchymal stem cells are attracted to latent HIV-1-infected cells and enable virus reactivation via a non-canonical PI3K-NFκB signaling pathway. Sci Rep. (2018) 8:14702. doi: 10.1038/s41598-018-32657-y
207. Alipoor SD, Mortaz E, Garssen J, Movassaghi M, Mirsaeidi M, and Adcock IM. Exosomes and exosomal miRNA in respiratory diseases. Mediators Inflammation. (2016) 2016:5628404. doi: 10.1155/2016/5628404
208. Zheng G, Huang L, Tong H, Shu Q, Hu Y, Ge M, et al. Treatment of acute respiratory distress syndrome with allogeneic adipose-derived mesenchymal stem cells: A randomized, placebo-controlled pilot study. Respir Res. (2014) 15:39. doi: 10.1186/1465-9921-15-39
209. Batchinsky AI, Roberts TR, Antebi B, Necsoiu C, Choi JH, Herzig M, et al. Intravenous autologous bone marrow–derived mesenchymal stromal cells delay acute respiratory distress syndrome in swine. Am J Respir Crit Care Med. (2023) 208:1283–92. doi: 10.1164/rccm.202305-0865OC
210. Devaney J, Horie S, Masterson C, Elliman S, Barry F, O’Brien T, et al. Human mesenchymal stromal cells decrease the severity of acute lung injury induced by E. coli in the rat. Thorax. (2015) 70:625–35. doi: 10.1136/thoraxjnl-2015-206813
211. Varshney V, Gabble BC, Bishoyi AK, Varma P, Qahtan SA, Kashyap A, et al. Exploring exosome-based approaches for early diagnosis and treatment of neurodegenerative diseases. Mol Neurobiol. (2025). doi: 10.1007/s12035-025-05026-w
212. Chung E and Son Y. Crosstalk between mesenchymal stem cells and macrophages in tissue repair. Tissue Eng Regener Med. (2014) 11:431–8. doi: 10.1007/s13770-014-0072-1
213. Harrell C, Popovska Jovicic B, Djonov V, and Volarevic V. Molecular mechanisms responsible for mesenchymal stem cell-based treatment of viral diseases. Pathogens. (2021) 10:409. doi: 10.3390/pathogens10040409
214. Volarevic V, Gazdic M, Simovic Markovic B, Jovicic N, Djonov V, and Arsenijevic N. Mesenchymal stem cell-derived factors: Immuno-modulatory effects and therapeutic potential. BioFactors. (2017) 43:633–44. doi: 10.1002/biof.1374
215. Qian X, Xu C, Fang S, Zhao P, Wang Y, Liu H, et al. Exosomal microRNAs derived from umbilical mesenchymal stem cells inhibit hepatitis C virus infection. Stem Cells Transl Med. (2016) 5:1190–203. doi: 10.5966/sctm.2015-0348
216. León-Moreno LC, Reza-Zaldívar EE, Hernández-Sapiéns MA, Villafaña-Estarrón E, García-Martin M, Ojeda-Hernández DD, et al. Mesenchymal stem cell-based therapies in the post-acute neurological COVID syndrome: current landscape and opportunities. Biomolecules. (2024) 14:8. doi: 10.3390/biom14010008
217. Taechangam N, Kol A, Arzi B, and Borjesson DL. Multipotent stromal cells and viral interaction: current implications for therapy. Stem Cell Rev Rep. (2022) 18:214–27. doi: 10.1007/s12015-021-10224-9
218. Oliveira AG, Gonçalves M, Ferreira H, and M. Neves N. Growing evidence supporting the use of mesenchymal stem cell therapies in multiple sclerosis: A systematic review. Mult Scler Relat Disord. (2020) 38:101860. doi: 10.1016/j.msard.2019.101860
219. Hoang DM, Pham PT, Bach TQ, Ngo ATL, Nguyen QT, Phan TTK, et al. Stem cell-based therapy for human diseases. Signal Transduct Target Ther. (2022) 7:272. doi: 10.1038/s41392-022-01134-4
220. Shi L, Huang H, Lu X, Yan X, Jiang X, Xu R, et al. Effect of human umbilical cord-derived mesenchymal stem cells on lung damage in severe COVID-19 patients: a randomized, double-blind, placebo-controlled phase 2 trial. Signal Transduct Target Ther. (2021) 6:58. doi: 10.1038/s41392-021-00488-5
221. Shi L, Yuan X, Yao W, Wang S, Zhang C, Zhang B, et al. Human mesenchymal stem cells treatment for severe COVID-19: 1-year follow-up results of a randomized, double-blind, placebo-controlled trial. EBioMedicine. (2022) 75:103789. doi: 10.1016/j.ebiom.2021.103789
222. Yuan M-Q, Song L, Wang Z-R, Zhang Z-Y, Shi M, He J, et al. Long-term outcomes of mesenchymal stem cell therapy in severe COVID-19 patients: 3-year follow-up of a randomized, double-blind, placebo-controlled trial. Stem Cell Res Ther. (2025) 16:94. doi: 10.1186/s13287-025-04148-1
223. Li T-T, Zhang B, Fang H, Shi M, Yao W-Q, Li Y, et al. Human mesenchymal stem cell therapy in severe COVID-19 patients: 2-year follow-up results of a randomized, double-blind, placebo-controlled trial. EBioMedicine. (2023) 92:104600. doi: 10.1016/j.ebiom.2023.104600
224. Şahin AS, Kaya E, Turgut G, Dolay K, and Kocataş A. Mesenchymal stem cell therapy in COVID-19 pneumonia: A prospective, randomized clinical research. Turkiye Klinikleri J Med Sci. (2022) 42:5–13. doi: 10.5336/medsci.2021-85395
225. Adas G, Cukurova Z, Yasar KK, Yilmaz R, Isiksacan N, Kasapoglu P, et al. The systematic effect of mesenchymal stem cell therapy in critical COVID-19 patients: A prospective double controlled trial. Cell Transplant. (2021) 30:9636897211024942. doi: 10.1177/09636897211024942
226. von Schwarz ER, Busse N, Angelus KM, de la Torre A, Yu Y-CJ, Sung S, et al. Promising benefit of single mesenchymal stem cell injection in critically ill COVID-19 patients – A pilot phase randomized controlled study. Online Int Interdiscip Res J. (2011) 11.
227. Karyana M, Djaharuddin I, Rif’ati L, Arif M, Choi MK, Angginy N, et al. Safety of DW-MSC infusion in patients with low clinical risk COVID-19 infection: a randomized, double-blind, placebo-controlled trial. Stem Cell Res Ther. (2022) 13:134. doi: 10.1186/s13287-022-02812-4
228. Araldi RP, Prezoto BC, Gonzaga V, Policiquio B, Mendes TB, D’Amélio F, et al. Advanced cell therapy with low tissue factor loaded product NestaCell® does not confer thrombogenic risk for critically ill COVID-19 heparin-treated patients. Biomed Pharmacother. (2022) 149:112920. doi: 10.1016/j.biopha.2022.112920
229. Lanzoni G, Linetsky E, Correa D, Messinger Cayetano S, Alvarez RA, Kouroupis D, et al. Umbilical cord mesenchymal stem cells for COVID-19 acute respiratory distress syndrome: A double-blind, phase 1/2a, randomized controlled trial. Stem Cells Transl Med. (2021) 10:660–73. doi: 10.1002/sctm.20-0472
230. Abdullah M, Pawitan JA, Irawan C-R, Aditianingsih D, Liem IK, Sinto R, et al. Effectiveness and safety profile of mesenchymal stem cell secretome as a treatment for severe cases of COVID-19: a randomized controlled trial. F1000Res. (2022) 11:143. doi: 10.12688/f1000research.75580.2
231. Işıksaçan N, Adaş G, Kasapoğlu P, Çukurova Z, Yılmaz R, Kurt Yaşar K, et al. The effect of mesenchymal stem cells administration on DNA repair gene expressions in critically ill COVID-19 patients: prospective controlled study. Nucleosides Nucleotides Nucleic Acids. (2024) 43:1431–46. doi: 10.1080/15257770.2024.2327478
232. Hancharou AYE, Aliakseichyk SE, Rynda AH, Antonevich NG, and Pankratova Y. Clinical efficacy of the application of pooled mesenchimal stem cells of the olfactory lining for the treatment of patients with severe community-acquired pneumonia complicated by acute respiratory distress syndrome. J Grodno State Med Univ. (2023) 21:466–76. doi: 10.25298/2221-8785-2023-21-5-466-476
233. Fathi-Kazerooni M, Fattah-Ghazi S, Darzi M, Makarem J, Nasiri R, Salahshour F, et al. Safety and efficacy study of allogeneic human menstrual blood stromal cells secretome to treat severe COVID-19 patients: clinical trial phase I & II. Stem Cell Res Ther. (2022) 13:96. doi: 10.1186/s13287-022-02771-w
234. English S, Fergusson D, Lalu M, Thebaud B, Watpool I, Champagne J, et al. Results of the cellular immuno-therapy for covid-19 related acute respiratory distress syndrome (circa-phase i trial. Cytotherapy. (2021) 23:S22. doi: 10.1016/S1465324921002796
235. Stewart DJ, Fergusson D, Soliman K, Chassé M, Lalu M, Thébaud B, et al. Mesenchymal stromal cells may improve outcomes in severe COVID-19 related ARDS by normalizing SARS-CoV2-related lymphopenia. Trans Sci Eur Respir Soc. (2025) 11:TP314. doi: 10.1183/23120541.LSC-2025.TP314
236. de Dios C, Vij R, Kim H, Park H, and Chang D. Safety of multiple intravenous infusions of adipose-derived mesenchymal stem cells for hospitalized cases of COVID-19: a randomized controlled trial. Front Med (Lausanne). (2023) 10:1321303. doi: 10.3389/fmed.2023.1321303
237. Zhu Y-G, Shi M, Monsel A, Dai C, Dong X, Shen H, et al. Nebulized exosomes derived from allogenic adipose tissue mesenchymal stromal cells in patients with severe COVID-19: a pilot study. Stem Cell Res Ther. (2022) 13:220. doi: 10.1186/s13287-022-02900-5
238. Pochon C, Laroye C, Kimmoun A, Reppel L, Dhuyser A, Rousseau H, et al. Efficacy of Wharton Jelly Mesenchymal Stromal Cells infusions in moderate to severe SARS-Cov-2 related acute respiratory distress syndrome: a phase 2a double-blind randomized controlled trial. Front Med (Lausanne). (2023) 10:1224865. doi: 10.3389/fmed.2023.1224865
239. Monsel A, Hauw-Berlemont C, Mebarki M, Heming N, Mayaux J, Nguekap Tchoumba O, et al. Treatment of COVID-19-associated ARDS with mesenchymal stromal cells: a multicenter randomized double-blind trial. Crit Care. (2022) 26:48. doi: 10.1186/s13054-022-03930-4
240. Lightner AL, Sengupta V, Qian S, Ransom JT, Suzuki S, Park DJ, et al. Bone marrow mesenchymal stem cell-derived extracellular vesicle infusion for the treatment of respiratory failure from COVID-19. Chest. (2023) 164:1444–53. doi: 10.1016/j.chest.2023.06.024
241. Vij R, Kim H, Park H, Cheng T, Lotfi D, and Chang D. Adipose-derived, autologous mesenchymal stem cell therapy for patients with post-COVID-19 syndrome: an intermediate-size expanded access program. Stem Cell Res Ther. (2023) 14:287. doi: 10.1186/s13287-023-03522-1
242. Tiumina OV, Davydkin IL, Volchkov SE, Ovchinnikov PA, Bugakov AI, Sharonova DU, et al. Safety and efficiency of inhalation method for administration of small extracellular vesicles derived from multipotent mesenchymal stromal cells of human umbilical cord in SARS- CoV-2 associated pneumonia. Genes Cells. (2022) 17:19–25. doi: 10.23868/202205004
243. Kantarcioglu M, Demirci H, Avcu F, Karslioglu Y, Babayigit MA, Karaman B, et al. Efficacy of autologous mesenchymal stem cell transplantation in patients with liver cirrhosis. Turkish J Gastroenterol. (2015) 26:244–50. doi: 10.5152/tjg.2015.0074
244. Xu W, He H, Pan S, Chen Y, Zhang M, Zhu S, et al. Combination treatments of plasma exchange and umbilical cord-derived mesenchymal stem cell transplantation for patients with hepatitis B virus-related acute-on-chronic liver failure: A clinical trial in China. Stem Cells Int. (2019) 2019:1–10. doi: 10.1155/2019/4130757
245. Trujillo-Rodríguez M, Viciana P, Rivas-Jeremías I, Álvarez-Ríos AI, Ruiz-García A, Espinosa-Ibáñez O, et al. Mesenchymal stromal cells in human immunodeficiency virus-infected patients with discordant immune response: Early results of a phase I/II clinical trial. Stem Cells Transl Med. (2021) 10:534–41. doi: 10.1002/sctm.20-0213
246. Wang L, Zhang Z, Xu R, Wang X, Shu Z, Chen X, et al. Human umbilical cord mesenchymal stem cell transfusion in immune non-responders with AIDS: a multicenter randomized controlled trial. Signal Transduct Target Ther. (2021) 6:217. doi: 10.1038/s41392-021-00607-2
247. Lin B, Chen J, Qiu W, Wang K, Xie D, Chen X, et al. Allogeneic bone marrow–derived mesenchymal stromal cells for hepatitis B virus–related acute-on-chronic liver failure: A randomized controlled trial. Hepatology. (2017) 66:209–19. doi: 10.1002/hep.29189
248. Li YH and Wu HM. The strategy of combining antiviral agent, plasma exchange and transarterial mesenchymal stem cell transfusion in a patient with hepatitis B virus (HBV) related acute-on-chronic liver failure. Trans Med. (2016) 6. doi: 10.4172/2161-1025.1000173
249. Li Y-H, Xu Y, Wu H-M, Yang J, Yang L-H, and Yue-Meng W. Umbilical cord-derived mesenchymal stem cell transplantation in hepatitis B virus related acute-on-chronic liver failure treated with plasma exchange and entecavir: a 24-month prospective study. Stem Cell Rev Rep. (2016) 12:645–53. doi: 10.1007/s12015-016-9683-3
250. Kuşkonmaz B, Uçkan D, Yalnizoğlu D, Günel M, Karli Oğuz K, Konuşkan B, et al. Mesenchymal stem cell application in children with subacute sclerosing panencephalitis. Dev Med Child Neurol. (2015) 57:880–3. doi: 10.1111/dmcn.12747
251. Lebeau G, Ah-Pine F, Daniel M, Bedoui Y, Vagner D, Frumence E, et al. Perivascular mesenchymal stem/stromal cells, an immune privileged niche for viruses? Int J Mol Sci. (2022) 23:8038. doi: 10.3390/ijms23148038
252. de Wolf C, van de Bovenkamp M, and Hoefnagel M. Regulatory perspective on in vitro potency assays for human mesenchymal stromal cells used in immunotherapy. Cytotherapy. (2017) 19:784–97. doi: 10.1016/j.jcyt.2017.03.076
253. Shimizu Y, Ntege EH, Inoue Y, Matsuura N, Sunami H, and Sowa Y. Optimizing mesenchymal stem cell extracellular vesicles for chronic wound healing: Bioengineering, standardization, and safety. Regener Ther. (2024) 26:260–74. doi: 10.1016/j.reth.2024.06.001
254. Shi M, Li J, Liao L, Chen B, Li B, Chen L, et al. Regulation of CXCR4 expression in human mesenchymal stem cells by cytokine treatment: role in homing efficiency in NOD/SCID mice. Haematologica. (2007) 92:897–904. doi: 10.3324/haematol.10669
255. Hu X, Yu SP, Fraser JL, Lu Z, Ogle ME, Wang J-A, et al. Transplantation of hypoxia-preconditioned mesenchymal stem cells improves infarcted heart function via enhanced survival of implanted cells and angiogenesis. J Thorac Cardiovasc Surg. (2008) 135:799–808. doi: 10.1016/j.jtcvs.2007.07.071
256. Li H, Ji X-Q, Zhang S-M, and Bi R-H. Hypoxia and inflammatory factor preconditioning enhances the immunosuppressive properties of human umbilical cord mesenchymal stem cells. World J Stem Cells. (2023) 15:999–1016. doi: 10.4252/wjsc.v15.i11.999
257. Contreras-Lopez R, Elizondo-Vega R, Luque-Campos N, Torres MJ, Pradenas C, Tejedor G, et al. The ATP synthase inhibition induces an AMPK-dependent glycolytic switch of mesenchymal stem cells that enhances their immunotherapeutic potential. Theranostics. (2021) 11:445–60. doi: 10.7150/thno.51631
258. Sekenova A, Li Y, Issabekova A, Saparov A, and Ogay V. TNF-α Preconditioning improves the therapeutic efficacy of mesenchymal stem cells in an experimental model of atherosclerosis. Cells. (2023) 12:2262. doi: 10.3390/cells12182262
259. Tolstova T, Dotsenko E, Luzgina N, and Rusanov A. Preconditioning of mesenchymal stem cells enhances the neuroprotective effects of their conditioned medium in an Alzheimer’s disease in vitro model. Biomedicines. (2024) 12:2243. doi: 10.3390/biomedicines12102243
260. Zhang Z, Zhang Y, Gao F, Han S, Cheah KS, Tse H-F, et al. CRISPR/Cas9 genome-editing system in human stem cells: current status and future prospects. Mol Ther Nucleic Acids. (2017) 9:230–41. doi: 10.1016/j.omtn.2017.09.009
261. Mandegar MA, Huebsch N, Frolov EB, Shin E, Truong A, Olvera MP, et al. CRISPR interference efficiently induces specific and reversible gene silencing in human iPSCs. Cell Stem Cell. (2016) 18:541–53. doi: 10.1016/j.stem.2016.01.022
262. Guo J, Ma D, Huang R, Ming J, Ye M, Kee K, et al. An inducible CRISPR-ON system for controllable gene activation in human pluripotent stem cells. Protein Cell. (2017) 8:379–93. doi: 10.1007/s13238-016-0360-8
263. Bertolino GM, Maumus M, Jorgensen C, and Noël D. Recent advances in extracellular vesicle-based therapies using induced pluripotent stem cell-derived mesenchymal stromal cells. Biomedicines. (2022) 10:2281. doi: 10.3390/biomedicines10092281
264. Joshi BS and Zuhorn IS. Heparan sulfate proteoglycan-mediated dynamin-dependent transport of neural stem cell exosomes in an in vitro blood–brain barrier model. Eur J Neurosci. (2021) 53:706–19. doi: 10.1111/ejn.14974
265. Yin JQ, Zhu J, and Ankrum JA. Manufacturing of primed mesenchymal stromal cells for therapy. Nat BioMed Eng. (2019) 3:90–104. doi: 10.1038/s41551-018-0325-8
266. García JR, Quirós M, Han WM, O’Leary MN, Cox GN, Nusrat A, et al. IFN-γ-tethered hydrogels enhance mesenchymal stem cell-based immunomodulation and promote tissue repair. Biomaterials. (2019) 220:119403. doi: 10.1016/j.biomaterials.2019.119403
267. García-Guerrero CA, Fuentes P, Araya MJ, Djouad F, Luz-Crawford P, Vega-Letter AM, et al. How to enhance MSCs therapeutic properties? An insight on potentiation methods. Stem Cell Res Ther. (2024) 15:331. doi: 10.1186/s13287-024-03935-6
268. Liu C, Fan Y, Zhou L, Zhu H-Y, Song Y-C, Hu L, et al. Pretreatment of mesenchymal stem cells with angiotensin II enhances paracrine effects, angiogenesis, gap junction formation and therapeutic efficacy for myocardial infarction. Int J Cardiol. (2015) 188:22–32. doi: 10.1016/j.ijcard.2015.03.425
269. Killer MC, Nold P, Henkenius K, Fritz L, Riedlinger T, Barckhausen C, et al. Immunosuppressive capacity of mesenchymal stem cells correlates with metabolic activity and can be enhanced by valproic acid. Stem Cell Res Ther. (2017) 8:100. doi: 10.1186/s13287-017-0553-y
270. Levy O, Kuai R, Siren EMJ, Bhere D, Milton Y, Nissar N, et al. Shattering barriers toward clinically meaningful MSC therapies. Sci Adv. (2020) 6:eaba6884. doi: 10.1126/sciadv.aba6884
271. Guadix JA, López-Beas J, Clares B, Soriano-Ruiz JL, Zugaza JL, and Gálvez-Martín P. Principal criteria for evaluating the quality, safety and efficacy of hMSC-based products in clinical practice: current approaches and challenges. Pharmaceutics. (2019) 11:552. doi: 10.3390/pharmaceutics11110552
272. Guo BC, Wu KH, Chen CY, Lin WY, Chang YJ, Lee TA, et al. Mesenchymal stem cells in the treatment of COVID-19. Int J Mol Sci. (2023) 24:14800. doi: 10.3390/ijms241914800
Keywords: virus, mesenchymal stem cells, central nervous system (CNS), neuroinflammation, secretome, extracellular vesicle (EV), neurological and neurodegenerative diseases, persistent infections
Citation: Carbone-Schellman J, Fontecilla-Escobar J, Sales-Salinas N, Chaparro-Pico WF, Molina-Berríos A, Ruete MC, González PA, Machuca MA, Opazo MC, Ezquer ME and Duarte LF (2025) Therapeutic challenges in central nervous system viral infections: advancing mesenchymal stem cell-based strategies for treating neuroinflammation and promoting tissue repair. Front. Immunol. 16:1677433. doi: 10.3389/fimmu.2025.1677433
Received: 31 July 2025; Accepted: 29 September 2025;
Published: 29 October 2025.
Edited by:
Bernhard Ryffel, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Firoz Ahmad, Oklahoma State University, United StatesMin Yang, Jilin Agricultural Science and Technology College, China
Copyright © 2025 Carbone-Schellman, Fontecilla-Escobar, Sales-Salinas, Chaparro-Pico, Molina-Berríos, Ruete, González, Machuca, Opazo, Ezquer and Duarte. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luisa F. Duarte, bGR1YXJ0ZUB1ZGQuY2w=
†These authors have contributed equally to this work and share first authorship