 Ana Pinheiro
Ana Pinheiro J. Ricardo Borges1,2,3
J. Ricardo Borges1,2,3 João Pedro Marques
João Pedro Marques Pedro José Esteves
Pedro José Esteves- 1CIBIO‐InBIO, Research Center in Biodiversity and Genetic Resources, University of Porto, Vairão, Portugal
- 2BIOPOLIS Program in Genomics, Biodiversity and Land Planning, Research Center in Biodiversity and genetic resources (CIBIO), Vairão, Portugal
- 3Department of Biology, Faculty of Sciences, University of Porto, Porto, Portugal
- 4CITS - Center of Investigation in Health Technologies, CESPU, Gandra, Portugal
Background: The Immunoglobulin Kappa Constant (IGKC) gene encodes the constant region of the immunoglobulin kappa light chain, a crucial component of antibodies. Despite its important biological role, the genetic information for this gene remains scarce, with data for only 16 mammal species (as of July 2025) fully characterized in the International ImMunoGeneTics information system (IMGT) database.
Results: Using genomic sequences from NCBI and Ensembl, we expanded this to 124 IGKC sequences across 104 mammals, including two monotremes, eight marsupials, and 94 placentals. We uncovered unusual evolutionary dynamics in lagomorphs, showing independent IGKC duplications in Ochotonidae and Leporidae, giving rise to rabbit IGKC1 and IGKC2. No conserved glycosylation sites were found, but 26 sequences from 14 species carried potentially N-glycosylated sites, including two new sites in lagomorphs. Selection analyses revealed pervasive purifying selection interspersed with codons under positive selection, while aBSREL identified episodic diversifying selection in several lineages.
Conclusions: Our study represents a significant contribution to the knowledge of the IGKC gene, substantially expanding on information available in IMGT. It highlights complex evolutionary trajectories, especially in lagomorphs. The presence of N-glycosylated sites suggests potential effects on antigen binding, stability, or half-life. The coexistence of purifying and episodic positive selection points to a balance between structural conservation and lineage-specific functional diversification.
1 Introduction
Antibodies are typically Y-shaped glycoproteins composed of two distinct types of polypeptide chains, two identical heavy (H), and two identical light (L) chains. Each chain has a variable (V) and a constant (C) region. The light (VL and CL) chains and the variable (VH) and first constant domain of the heavy chains (CH1) constitute the antibody regions that recognizes and binds antigens (Fab regions), while the remaining constant domains of the heavy chains constitute the region that assures an effector function (Fc region). In mammals, there are two Ig L chain isotypes, kappa (K) and lambda (λ), which are functionally indistinguishable. Each L chain is encoded by three genes, variable (VL), joining (JL), and constant (CL), and each isotype has its own set of VL, JL, and CL. The constant region of the kappa light chain (IGKC) gene encodes the constant region of the immunoglobulin kappa light chain, a protein that interacts with the κVL, and JL regions and contributes to the effector phase of humoral immunity by mediating the elimination of bound antigens.
Humans and mice have a single IGKC. In humans, the IGKC gene is located on chromosome 2 (2p11.2), while in mice it is located on chromosome 6 (6C1). In the European rabbit (Oryctolagus cuniculus), the IGKC and the joining region of the kappa light chain (IGKJ) have duplicated and originated two different kappa light chains (IGKC1 and IGKC2) (1, 2). This duplication, confirmed by genomic data obtained from the OryCun 2.0 assembly, is located on chromosome 2 (3).
The genetic variation of the rabbit kappa light chains has been studied in detail and shows unique features. The IGKC1 locus has an additional cysteine residue, C85.4 (International ImMunoGeneTics information system (IMGT) numbering), that links, through an extra disulfide bond, to IGKV1 C96 (IMGT numbering) (4–6). Additionally, the European rabbit IGKC1 has a unique glycosylation site, 85.1NLS86 (IMGT numbering). Glycosylation, the process of attaching sugar molecules (glycans) to proteins, plays a crucial role in the immune system. Glycosylation can affect protein folding, stability, and interactions with other molecules, including other proteins and receptors (7, 8). The degree of inter-allelic diversity of the rabbit IGKC1 revealed high amino acid differences (9–12), only similar to that currently observed at vertebrate MHC loci (13). The trans-species nature, another benchmark of MHC evolution (14), was also documented for the rabbit b-locus allotypes obtained in L. americanus and European rabbit (15).
The information available for IGKC in the IMGT database remains limited, with data for only 16 mammal species (as of July 2025; https://www.imgt.org/IMGTrepertoire/Proteins/index.php#B). Recently, advances in genome sequencing technologies, particularly next-generation sequencing (NGS), have enabled the rapid sequencing and comparison of genomes across a wide range of species. This comparative genomics approach allows us to identify pathways that are unique to certain species, providing insights into their evolutionary history and adaptations. Evolutionary studies are crucial for understanding the mechanisms driving evolutionary innovation. In essence, the increased genetic information revealed by new genome sequencing technologies provides a powerful tool to understand the dynamics and creative processes of evolution. In this work, we sought to expand the knowledge about the IGKC genes in mammals (monotremes, marsupials, and placentals). Mining public databases, we retrieved IGKC sequences from over 100 mammal species and performed natural selection analyses. The results represent a major contribution to the study of this gene and provide an important increment to the IMGT database.
2 Materials and methods
Mammalian IGKC sequences were obtained from publicly accessible databases using our standard methodology, which we have already used in several studies (e.g (16–19). In total, we gathered 123 sequences from monotremes (two), marsupials (seven), and placentals (114). The accession numbers of all the sequences used are listed in Supplementary Table S1. The 114 placental sequences represent 94 species, with most species represented by a single sequence, except for lagomorphs, where several sequences per species were obtained (Supplementary Table S1). The sequences were retrieved through BLASTn searches in NCBI’s GenBank (http://www.ncbi.nlm.nih.gov/genbank/) and Ensembl (https://www.ensembl.org/index.html) genome databases, using as queries the mammalian IGKC sequences available in the IMGT database, which represent true IGKC with high confidence. For Ensembl-derived sequences, we used the Homo sapiens IGKC gene (ENSG00000211592) as a query and retrieved the list of mammalian orthologues available under the “Orthologues” section. As of July 2025, a total of 49 one-to-one orthologues and one one-to-two orthologue (in Oryctolagus cuniculus) were available. Because our BLASTn queries retrieved only annotated CDS, with transcript evidence, the initial two nucleotides of the IGKC exon (derived from the J–C splice junction) were not included in all sequences. As these do not alter the reading frame or translation, we expect no effect on functional inference, though small phylogenetic biases cannot be excluded. All obtained sequences contained the typical stop codon at the 3′ end; this was excluded from our analysis.
Sequences were aligned with Clustal W (20) as implemented in BioEdit (21), followed by visual inspection and necessary manual corrections. The final nucleotide sequence alignment is provided in Supplementary Material: Data 1. Amino acids were translated from the nucleotide sequences. Codon numbering is according to the IMGT unique numbering (22). N-linked glycosylation sites were estimated using the online tool NetNGlyc 1.0 Server, with + indicating a potential to reach the 0.5 threshold and ++/+++ to reach the 0.75 threshold (23).
2.1 Molecular phylogenetic analyses
A maximum likelihood (ML) phylogenetic tree was constructed for the mammalian nucleotide alignment using IQ-TREE v3.0.1. The best-fit substitution model was TVM+F+I+G4, selected under the Bayesian Information Criterion (BIC), which we preferred over AIC/AICc due to its stronger penalization of model complexity, thus reducing the risk of over-parameterization in large datasets. Node support was estimated using 10,000 ultrafast bootstrap replicates. A tree was inferred, and the tree topology was compared to the accepted mammalian phylogeny.
To further investigate the lagomorphs’ IGKC evolution, we constructed an ML phylogenetic tree for the lagomorph amino acid alignment using IQ-TREE v3.0.1. The best-fit substitution model was WAG+R4, again selected under BIC for its balance between model fit and parsimony. Node support was estimated using 10,000 ultrafast bootstrap replicates.
2.2 Detection of positive selection
Positive selection was assessed using HyPhy package v2.5.75. Codon-based analyses were performed on the nucleotide alignment comprising 108 codons. Four site-based methods were applied: SLAC (single-likelihood ancestor counting), FEL (fixed effects likelihood), FUBAR (fast, unconstrained Bayesian approximation), and MEME (mixed effects model of evolution). These methods allowed the detection of pervasive and episodic selection acting across codon sites. Additionally, the aBSREL (adaptive branch-site random effects likelihood) model was used to detect episodic diversifying selection on individual branches of the phylogenetic tree.
Sites under pervasive positive and purifying selection were mapped onto the three-dimensional structure of the human IGKC protein, predicted by AlphaFold (UniProt entry P01834), using the high-confidence regions of the model for visualization.
3 Results
The obtained 123 IGKC sequences represent 105 mammalian species (95 placentals, seven marsupials, and two monotremes) (Figure 1). For the vast majority of mammalian species analyzed, we found only a single copy of this gene. The exception was lagomorphs, in which at least two sequences were recovered for each species. In our alignment, to the four IGKC2 and nine IGKC1 European rabbit alleles described in IMGT, we added two more previously published alleles, b4wc and b5wf (11), here named as rabbit IMGT IGKC1*10 and IMGT IGKC1*11, respectively. For Ochotona species, we obtained four novel sequences: two for O. princeps and two for O. curzoniae. Regarding Lepus americanus, we identified three new sequences in addition to the previously described L. americanus IGKC sequence (15). All sequences that we obtained contain a stop codon at the 3′ end, have high homology among them and the queries, and, as annotated CDS, have derived transcripts. Additionally, the diversity of genes that we found agrees with what is known about the IGKC; mammalian species have one IGKC except for the rabbit, which has two IGKC. The IGKC nucleotide ML tree generally conforms to the accepted mammalian phylogeny (24) (Figure 2). Within lagomorphs, the Ochotona sequences occupy a basal position (100 bootstrap support), while the rabbit and Lepus sequences form a second cluster (100 bootstrap support). Within this cluster, three groups are evident: (a) the rabbit IGKC2 cluster (99 bootstrap support), (b) the rabbit IGKC1 b9 alleles (IGKC1*4, IGKC1*5, and IGKC1*9) (100 bootstrap support), and (c) the remaining rabbit IGKC1 and the Lepus sequences (71 bootstrap support) (Figure 2).

Figure 1. Mammalian IGKC amino acid sequences alignment. The full IGKC amino acid sequences are depicted. The codon numbering is according to the IMGT unique numbering for the constant domain (22). Asterisks (*) above the numbering indicate the position of conserved Cys residues. For Lagomorphs, characteristic residues for IGKC1 are shaded in red, IGKC1 b9 alleles are shaded in yellow, IGKC2 are shaded in blue, IGKCLa3 are shaded in light blue, and Ochotonidae IGKC are shaded in gray. N-glycosylation sites identified using the online tool NetNGlyc 1.0 (23) are underlined in red. Dots (.) represent identity with the top sequence, and dashes (–) indicate gaps in the alignment. The codons identified as under pervasive positive selection, pervasive negative selection and episodic positive selection are shown under the alignment.
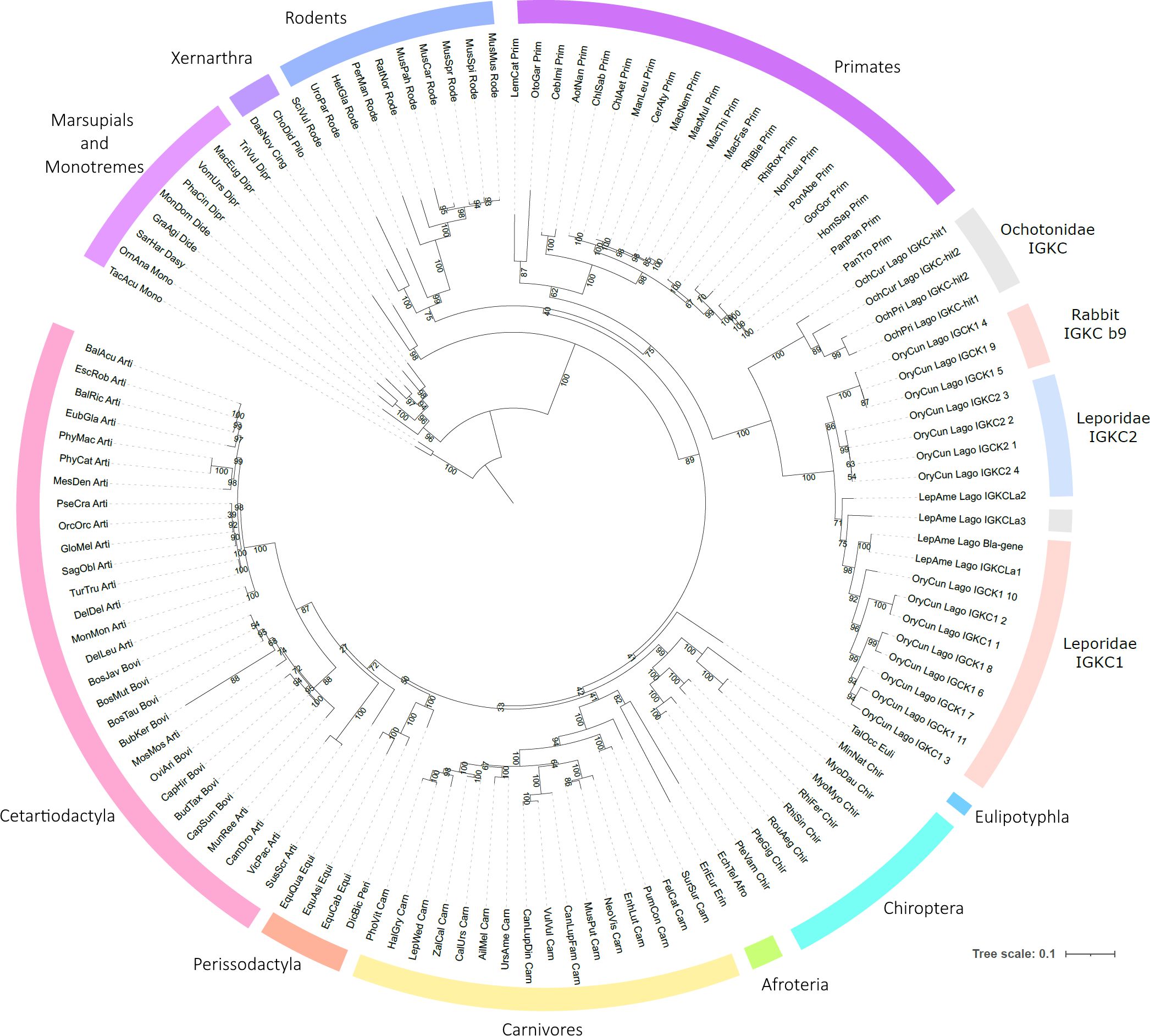
Figure 2. Phylogenetic tree of mammalian IGKC. The nucleotide phylogenetic tree was obtained using TVM+F+I+G4 as the best-fit substitution model, selected under the Bayesian Information Criterion (BIC). Node support was estimated using 10,000 ultrafast bootstrap replicates. All bootstrap values are shown, with values indicated near the most relevant branches. IGKC1 is shaded in pink, IGKC2 is shaded in blue, and indeterminate lagomorphs’ IGKC is shaded in gray.
3.1 The special case: lagomorphs
We obtained IGKC genes for four lagomorph species: Ochotona prínceps, Ochotona curzoniae, Lepus americanus, and Oryctolagus cuniculus. The lagomorphs are divided into two families, Leporidae and Ochotonidae. Ochotonidae has a single genus, Ochotona (pikas), while Leporidae includes 10 genera, among them Lepus and Oryctolagus (25). The lagomorph IGKC amino acid tree shows a different pattern than the nucleotide phylogeny (Figures 2, 3, respectively). In the amino acid tree (Figure 3), the Ochotona sequences again occupy a basal position (100 bootstrap support), with rabbit and Lepus sequences forming a second cluster (96 bootstrap support). Within Leporidae, the L. americanus sequence IGKCLa3 occupies a basal position. Some IGKC1 alleles, the b9 alleles—IGKC1*4, IGKC1*5, and IGKC1*9—form a well-supported cluster (100 bootstrap support) grouping with IGKC2 (99 bootstrap support), apart from other IGKC1 alleles (80 bootstrap support). Previous phylogenetic analyses have also placed the IGCK1 b9 alleles closer to IGKC2 than to other IGKC1 (13). Although with a lower bootstrap support, L. americanus IGKCLa2 clusters with the IGKC2 and IGKC1 b9 alleles, while IGKCLa1 clusters with IGKC1 (58 and 64 bootstrap support, respectively).
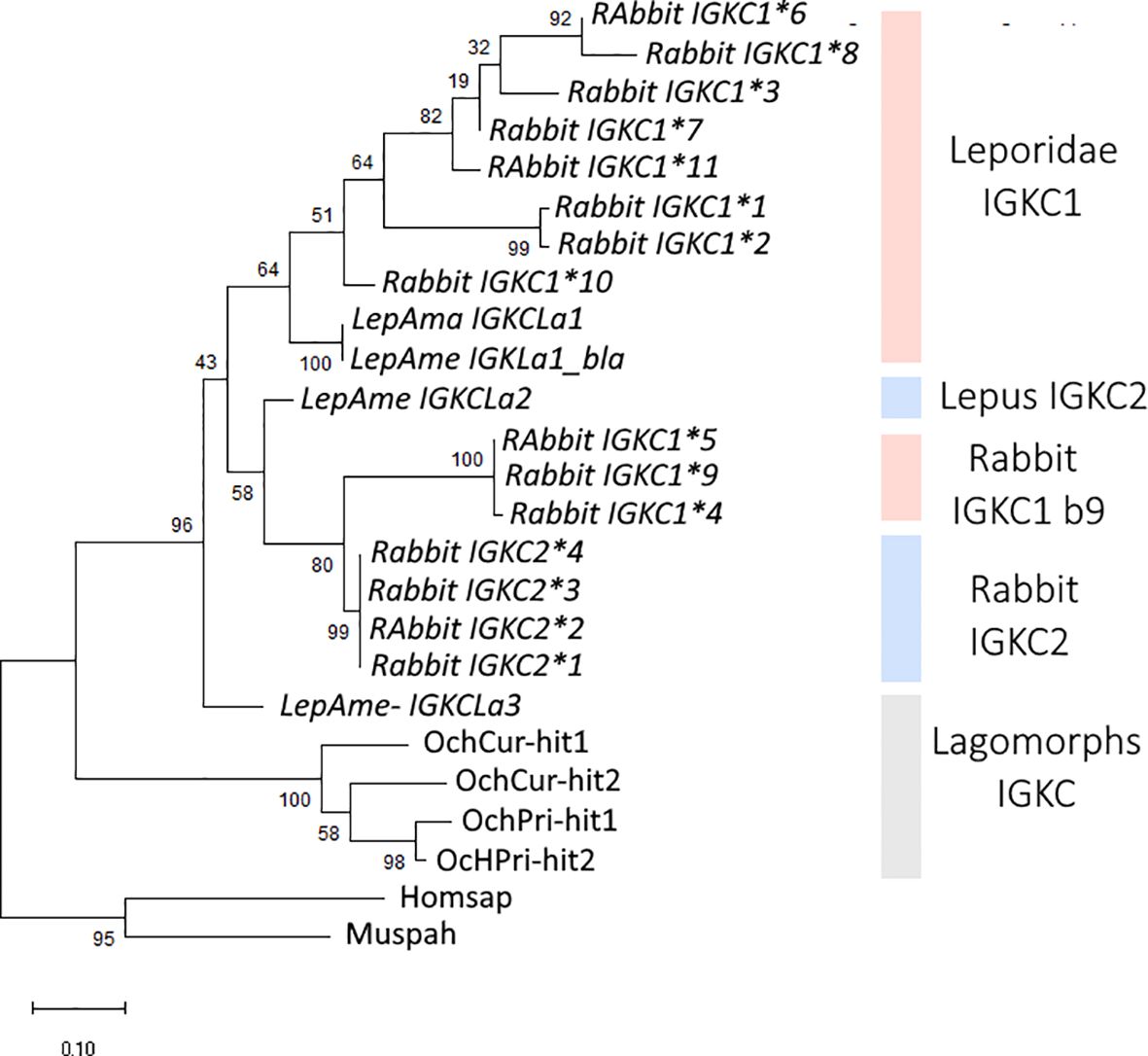
Figure 3. Phylogenetic tree of lagomorphs’ IGKC. The amino acid phylogenetic tree was obtained using WAG+R4 as the best-fit substitution model, selected under the Bayesian Information Criterion (BIC). Node support was estimated using 10,000 ultrafast bootstrap replicates. All bootstrap values are shown, with values indicated near the most relevant branches. IGKC1 is shaded in pink, IGKC2 is shaded in blue, and indeterminate lagomorphs’ IGKC is shaded in gray. The scale bar refers to the inferred amount of change per site along branches. Rabbit IGKC alleles are indicated with *n.
In both Ochotona species, we found two IGKC genes that were located chromosomally adjacent (Figure 4), similarly to the arrangement described for the European rabbit (3). The Ochotona IGKC sequences have characteristic residues distinguishing them from Leporidae IGKC: 24IADK27, 44GGV45.1, N99, I106, and L117 (Figure 1). These sequences also share rabbit IGKC2 residues N85.4 and 85.1SLS86 (Figure 1), supporting the view that IGKC2 predates IGKC1 (13).
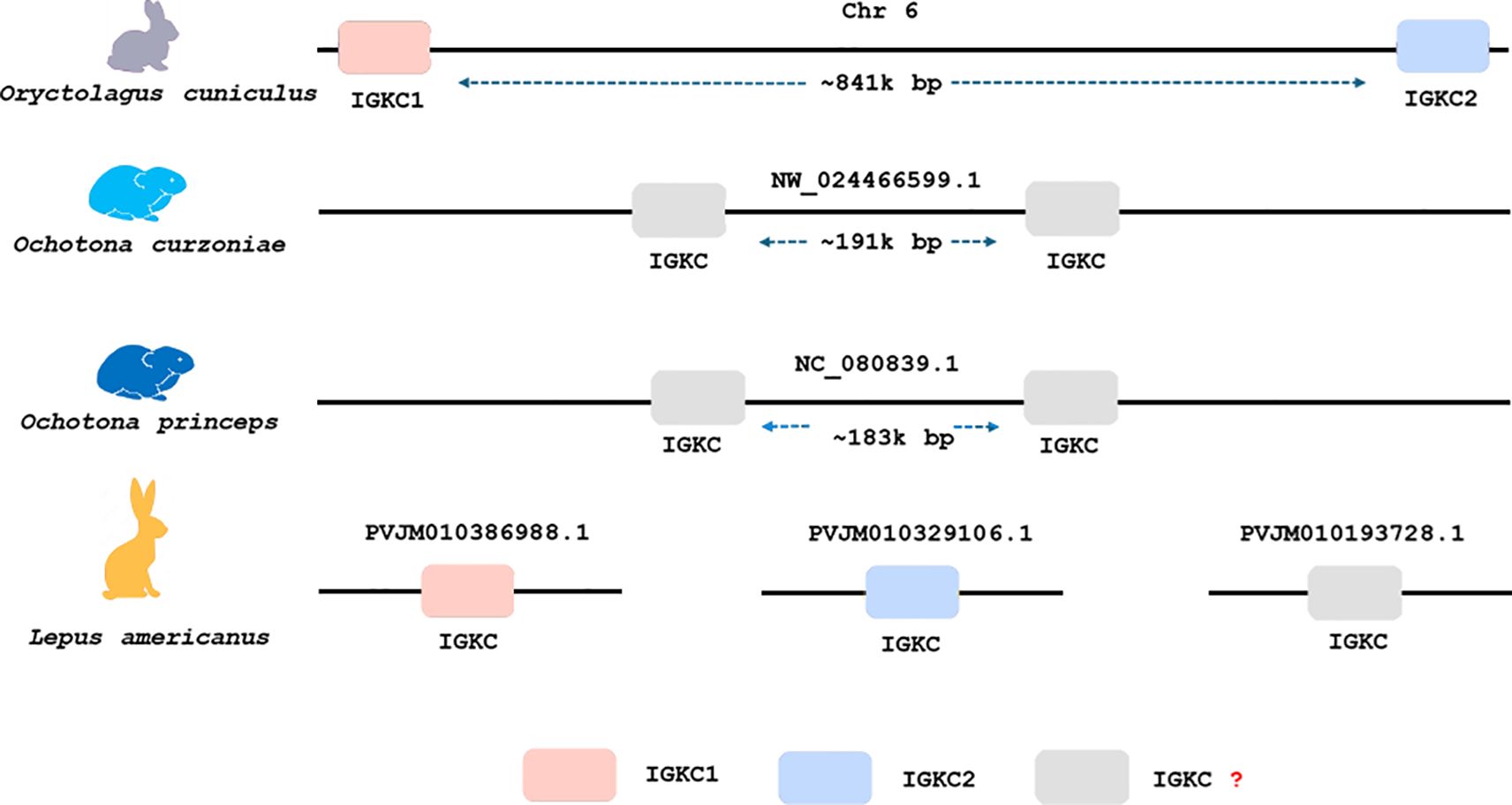
Figure 4. Genomic organization and copy number of IGKC genes in four lagomorph species. Each schematic represents one species: Oryctolagus cuniculus (European rabbit), Ochotona curzoniae (plateau pika), Ochotona princeps (American pika), and Lepus americanus (snowshoe hare), from top to bottom. The boxes indicate IGKC gene copies and their relative chromosomal positions based on BLASTn results.
For Lepus americanus, we identified three new sequences (Figure 1). As the genome assembly for this species is not a chromosome-level assembly, the three IGKC sequences are located on separate scaffolds, preventing us from determining whether they represent three gene copies or two genes with one allelic variant. The nucleotide and amino acid phylogenies (Figures 2 and 3, respectively) show two different scenarios for the leporid IGKC. In the nucleotide tree, all L. americanus sequences cluster with rabbit IGKC1 sequences (Figure 1), whereas in the amino acid tree L. americanus IGKCLa1 clusters with rabbit IGKC1 sequences and IGKCLa2 clusters with the rabbit IGKC2 sequences. The Lepus americanus IGKCLa1 shares two distinguishing features of the European rabbit IGKC1: C85.4 and 85.1NLS86. This shows that the rabbit IGKC1 novelty features, C85.4 and 85.1NLS86, that may have triggered the high allelic diversity observed in European rabbit IGKC1 (reviewed in (13)) were already present in L. americanus and emerged at least 12 million years ago (Oryctolagus–Lepus divergence time (26, 27);). The second L. americanus sequence, named here L. americanus IGKCLa2, surprisingly shares with the European rabbit IGKC2 residues N85.4 and 85.1SLS86 as well as 33SDIT36. The L. americanus IGKCLa3 shares some characteristic residues with the rabbit b9 alleles, IGKC1*4, IGKC1*5, and IGKC1*9, such as N33, 80NST82, and N110, but lacks the IGKC1 85.1NLS86 motif, having instead 85.1SLS86 like rabbit IGKC2 and unique features C29 and G85.4.
3.2 IGKC glycosylation sites
Glycosylation plays a major role in immunoglobulin structure and function. Heavy chain isotypes, IgA, IgG, IgE, IgD, and IgM, are known to be N-glycosylated to various extents (reviewed in (28), but IGKC glycosylation has not been described previously. We screened the mammalian IGKC sequences for N-linked glycosylation sites. No conserved sites of glycosylation were found for mammalian IGKC; however, 26 sequences, representing 14 species, are potentially N-glycosylated (Figure 1). Equus quagga and Equus asinus (Perissodactyla) (NetNGlyc threshold: ++), Orcinus orca (Cetartiodactyla) (NetNGlyc threshold: ++), and Miniopterus natalensis (Chiroptera) (NetNGlyc threshold: +) share the 18NAS20 glycosylation site, and five Carnivores have 101NFS103 (NetNGlyc threshold: +). Eschrichtius robustus IGKC has 16NGT18 (NetNGlyc threshold: +++) and Talpa occidentalis has 45.5NGS78 (NetNGlyc threshold: +).
In lagomorphs’ IGKC, a potential N-glycosylation site has been identified as a hallmark characteristic of IGKC1, 85.1NLS86 (15) (NetNGlyc threshold: ++). Our analysis reveals additional putative N-glycosylation sites. The rabbit b9 alleles, IGKC1*4, IGKC1*5, and IGKC1*9, are potentially glycosylated at 80NST82 and 85.1NLS86 (NetNGlyc threshold: ++). L. americanus IGKCLa1 is also potentially glycosylated at 85.1NLS86 (NetNGlyc threshold: +), while IGKCLa3 can be glycosylated at 80NST82 (NetNGlyc threshold: ++). All Ochotona sequences have a 84.2NXS/T84.4 motif, but our results indicate that only one O. curzoniae sequence is potentially glycosylated at this position, having the motif 84.2NCSD84.4 (NetNGlyc threshold: +).
3.3 Positive selection analyses
The analysis of selection pressures acting on IGKC coding sequences identified multiple codon sites under pervasive and episodic positive selection. Seven codons (1.3, 35, 45.2, 82, 92, 101, and 103) were supported by at least two site-based methods (SLAC, FEL, and FUBAR), while MEME revealed episodic selection in 25 codons, including those previously identified. Codon 85.4, which corresponds to one of the key motifs distinguishing rabbit IGKC1, was flagged as under selection.
Widespread purifying selection was also detected, with 53 codons identified across SLAC and FEL as being subject to significant evolutionary constraint. These included codons located in highly conserved regions of the constant domain, such as residues 2, 21–23, 28–30, 81, 84.5, 85.3–87, 91, 93, 96, 99, 102, 104–106, and 115–126. These positions likely reflect structural or functional constraints essential to the immunoglobulin fold.
The sites under pervasive positive and purifying selection are distributed across the protein. The sites identified as under pervasive positive selection are generally located in exposed regions, in the loops or loop vicinity (Figure 5).
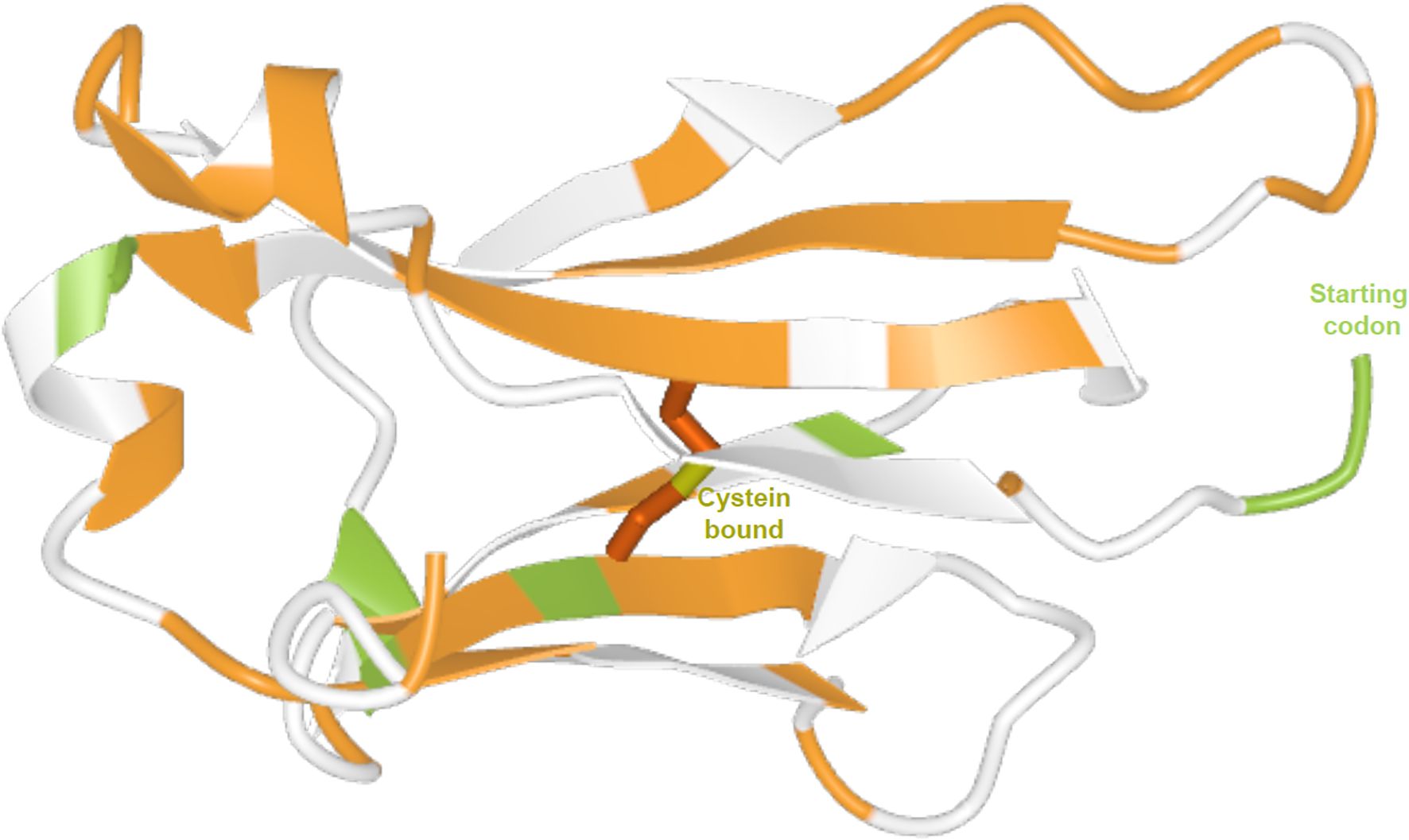
Figure 5. Three-dimensional structure of the human IGKC protein. Predicted by AlphaFold (UniProt entry P01834, model with very high confidence). Sites under pervasive positive selection are shown in green, and sites under purifying selection are shown in orange.
Branch-site analyses using aBSREL detected episodic diversifying selection in the terminal branches of Sarcophilus harrisii and Equus asinus as well as in several internal nodes: between Vulpes vulpes and Canis lupus dingo, between one copy of Ochotona curzoniae and the two O. princeps sequences, and in two clades within the rabbit IGKC1 group. These results support the notion that selective pressures are not uniformly distributed across mammalian lineages.
4 Discussion
Antibodies are hallmark molecules of the vertebrate immune system, playing a crucial role in protective immunity. The heavy chains, which determine the antibody isotypes, have been widely studied, with their genetic diversity, structure, function, and evolution well characterized (reviewed in, e.g (29–31)) and continue being studied as new evidences of clinical relevance continue emerging. On the contrary, the two light chains, kappa (K) and lambda (λ), have been comparatively neglected, and much less is known about their diversity and evolution. In this work, we sought to extend the knowledge on the evolution of mammalian IGKC by mining available genome assemblies and conducting natural selection analyses.
We recovered sequences for 104 mammalian species, enhancing our understanding of IGKC genetic diversity. We acknowledge that the accuracy of available genome assemblies is a limiting factor. Assembly errors, particularly in fragmented genomes such as Lepus americanus, may underlie some ambiguities in copy number or motif conservation. Nevertheless, synteny, motif analysis, and phylogenetic consistency strongly support our identifications. Our data revealed the duplication of this gene in lagomorphs and confirmed that such duplication is unique to this group. The IGKC was previously shown to be duplicated in the European rabbit, which carries two IGKC copies, IGKC1 and IGKC2 (1–3). The presence of two IGKC genes in Ochotona and Oryctolagus could be explained by a duplication in the lagomorph ancestor between 50 to 57.2 million years ago (26, 32, 33). In that scenario, one Ochotona gene would be expected to cluster with rabbit IGKC1 and the other with rabbit IGKC2. However, both the nucleotide and amino acid sequence phylogenies (Figures 2 and 3) place the Ochotona sequences together in a basal position relative to all lagomorph IGKC. This pattern suggests that the Ochotona IGKC duplication likely occurred in the Ochotona ancestor, after the Leporidae and Ochotonidae split. A second IGKC duplication would then have taken place in the Leporidae ancestor, leading to the emergence of rabbit IGKC1 and IGKC2. Despite the incongruences between the lagomorph IGKC nucleotide and amino acid trees, we consider the most likely scenario to be a duplication in the Leporidae ancestor, as supported by the amino acid data and shared diagnostic residues between Lepus and Oryctolagus sequences.
In the nucleotide phylogeny, all three L. americanus IGKC sequences cluster with the rabbit IGKC1, except for IGKC1*4, IGKC1*5, and IGKC1*9 (Figure 1). In the amino acid tree, however, only the L. americanus IGKCLa1 maintains this position; IGKCLa1, but not IGKCLa2 and IGKCLa3, shares the two distinguishing features of the European rabbit IGKC1: C85.4 and 85.1NLS86. This indicates that these rabbit IGKC1 novelty features, 85.4C and 85.1NLS86, which may have triggered the high allelic diversity observed in European rabbit IGKC1 (reviewed in (13)), were already present in the Leporidae ancestor 12 million years ago (Oryctolagus–Lepus divergence time (26, 27)). The IGKCLa1 gene is identical to the L. americanus bla allele (Figure 1) described by Bouton and van der Loo (15). The bla allele is more closely related to the European rabbit b4wc allele (IGKC1*10) than the b4wc allele is to other European rabbit IGKC1 alleles (15). This pattern is an example of a trans-species polymorphism. Together with other examples also described between the European rabbit and Lepus species in IGHV (34, 35) and IGHA (36), these are some of the rare instances of trans-species polymorphisms described outside of MHC genes (14, 37). In the amino acid tree, L. americanus IGKCLa2 instead clusters with rabbit IGKC2. IGKCLa2 shares residues N85.4 and 85.1SLS86, as well as 33SDIT36, with the European rabbit IGKC2. The L. americanus IGKCLa3 shares some characteristic residues with the rabbit b9 alleles, IGKC1*4, IGKC1*5, and IGKC1*9, but lacks the IGKC1 85.1NLS86 motif, having instead 85.1SLS86 like rabbit IGKC2; it also has unique residues. The sharing of residues with IGKC1 and IGKC2 and its singularities cause it to adopt a basal position to other leporid IGKC.
Taken together, the most parsimonious explanation is that the second duplication in lagomorphs’ IGKC happened in the Leporidae ancestor, as reflected in the amino acid phylogeny and shared diagnostic residues between rabbit and Lepus sequences. The clustering of the Lepus IGKC with rabbit IGKC1 in the nucleotide tree is likely the result of high homology observed in several regions of the Lepus IGKC sequences, which are probably being homogenized through concerted evolution (see Supplementary Material: Data 1). Concerted evolution can explain incongruences between nucleotide and amino acid phylogenetic trees when gene conversion and unequal crossing-over events homogenize sequences within a multigene family, leading to a pattern where paralogous genes within a species are more similar to each other than to their orthologous counterparts in different species (38, 39).
Glycans are found attached to the Fc tail of all Ig isotypes comprising 2%–14% of the Ig molecular weight depending on the Ig isotype (40). The Fab arms may also carry N-glycans; IgA, IgE, and IgM heavy chains are glycosylated in the CH1 domain (40), and additionally, 15%–25% of circulating IgG are glycosylated on the V regions (VL or VH) (41). The glycosylation sites in the V regions are only acquired during antibody maturation through somatic hypermutation, and research indicates that selection pressure favors the acquisition of Fab N-glycosylation sites during B cell affinity maturation (28). The light chain C regions’ glycosylation has not, to our knowledge, been investigated. Our analysis reveals that putative N-glycosylation sites are present in ~21% of IGKC (26 out of 123 IGKC sequences carry N-glycosylation sites). N-linked glycans in the Fab region are known to influence antigen binding, increase antibody stability, or extend the antibody half-life (28, 42, 43). All identified sequences with N-glycosylation sites carry only one putative site, except for the European rabbit IGKC1 b9 alleles, IGKC1*4, IGKC1*5, and IGKC1*9, which carry two putative N-glycosylation sites, at 80NST82 and 85.1NLS86. The b9 alleles—IGKC1*4, IGKC1*5, and IGKC1*9—have been described to form an alternative bond between 84.5C and IGKJ in contrast to other IGKC1 alleles that form a bond between 84.5C and IGKV. A possible explanation for this alternative bond would be the presence of a glycan at 80NST82. The sharing of glycosylation sites between Lepus and rabbit further supports the common origin of IGKCLa1 and rabbit IGKC1. Intriguingly, the higher prevalence of IgG Fab glycans has been associated with the development of several autoimmune chronic diseases, namely, rheumatoid arthritis (RA), systemic lupus erythematosus, myasthenia gravis, pemphigus vulgaris, and ANCA-associated vasculitis (44, 45). The role of Fab glycans in autoimmunity is still unclear. On one hand, it has been shown that Fab glycans reduce the binding affinity for autoantigens (46, 47). Conversely, Fab glycans were found to enhance B cell receptor signaling and maintain its surface expression longer after antigenic stimulation (47). A shift toward the pro-inflammatory IgA2 subclass is also observed in RA patients with increased disease activity. IgA2 has twice as many N-glycosylation sites as IgA1, including a CH1 N-glycosylation site, and differences in their glycosylation pattern exist (48). Whether the glycan attached to the IgA2 CH1 N-glycosylation site has a direct influence on the pro-inflammatory nature of IgA2 has not been investigated.
4.1 Signatures of positive selection and functional relevance
The presence of codons under positive selection across diverse mammalian lineages indicates that IGKC, while structurally conserved, is subject to functional diversification. Several of the positively selected sites identified here lie within or adjacent to known functional motifs, including codon 85, which forms part of the unique glycosylation motif 85.1NLS86 in rabbit IGKC1. Its recurrent detection across SLAC, FEL, FUBAR, and MEME strongly suggests adaptive relevance. Similarly, branch-site tests identified episodic positive selection in specific taxa, including Sarcophilus harrisii, Equus asinus, and members of Ochotona and Oryctolagus, highlighting lineage-specific pressures potentially linked to immune challenges or structural innovation. In S. harrisii, this may reflect unique selective pressures associated with transmissible cancers, which have profoundly shaped the immune gene evolution in this species (49). In E. asinus, a domesticated species with a long history of close contact with humans and livestock, episodic selection may relate to exposure to diverse pathogen communities or to immune modulation under domestication (50).
In contrast, the strong signal of purifying selection across more than 50 codon sites points to a substantial functional constraint within much of the IGKC coding region. These conserved positions likely correspond to residues essential for maintaining the structural integrity of the kappa constant domain, particularly within β-strand regions and core elements of the immunoglobulin fold. The coexistence of strong purifying selection and episodic positive selection suggests a balance between preserving structural stability and allowing for lineage-specific functional adaptations, especially in clades where gene duplications or increased allelic diversity are observed.
5 Conclusion
This work substantially increases the information available for the IGKC gene and makes an important contribution to updating the IMGT database. Our findings show that lagomorphs present a unique evolutionary pattern for the IGKC gene, distinct from that observed in other mammals. However, the evolutionary history of IGKC genes within leporids could be even more complex. Therefore, obtaining IGKC sequences for additional leporids will be vital to understand the full evolutionary history of this gene in lagomorphs. Though there are no conserved N-glycosylation sites in IGKC, we detected several N-glycosylation sites in different mammalian species, some shared between species, which suggests that these glycosylation sites may have an important biological role. The presence of codons under positive selection across diverse mammalian lineages indicates that IGKC, while structurally conserved, is subject to functional diversification. Furthermore, the detection of episodic positive selection in specific taxa highlights lineage-specific pressures potentially linked to immune challenges or structural innovation.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
AP: Writing – original draft, Writing – review & editing, Formal analysis, Methodology, Investigation. JRB: Data curation, Writing – review & editing. JPM: Formal analysis, Writing – original draft, Methodology, Writing – review & editing. PE: Investigation, Data curation, Writing – review & editing, Conceptualization, Supervision, Validation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. Work supported by the European Union’s Horizon 2020 Research and Innovation Programme under the Grant Agreement Number 857251.
Acknowledgments
BIOPOLIS—Enhancing the transference of scientific and technological knowledge through a new Centre of Excellence in Environmental Biology, Ecosystems and AgroBiodiversity (NORTE-01-0246-FEDER-000063).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1686094/full#supplementary-material
References
1. Kelus AS and Weiss S. Variant strain of rabbits lacking immunoglobulin κ polypeptide chain. Nature. (1977) 265:156–8. doi: 10.1038/265156a0
2. Hole NJ, Young-Cooper GO, and Mage RG. Mapping of the duplicated rabbit immunoglobulin kappa light chain locus. Eur J Immunol. (1991) 21:403–9. doi: 10.1002/eji.1830210223
3. Gertz EM, Schaffer AA, Agarwala R, Bonnet-Garnier A, Rogel-Gaillard C, Hayes H, et al. Accuracy and coverage assessment of Oryctolagus cuniculus (rabbit) genes encoding immunoglobulins in the whole genome sequence assembly (OryCun2.0) and localization of the IGH locus to chromosome 20. Immunogenetics. (2013) 65:749–62. doi: 10.1007/s00251-013-0722-9
4. McCartney-Francis N, Skurla RM Jr., Mage RG, and Bernstein KE. Kappa-chain allotypes and isotypes in the rabbit: cDNA sequences of clones encoding b9 suggest an evolutionary pathway and possible role of the interdomain disulfide bond in quantitative allotype expression. Proc Natl Acad Sci U S A. (1984) 81:1794–8. doi: 10.1073/pnas.81.6.1794
5. Poulsen K, Fraser KJ, and Haber E. An active derivative of rabbit antibody light chain composed of the constant and the variable domains held together only by a native disulfide bond. Proc Natl Acad Sci U S A. (1972) 69:2495–9. doi: 10.1073/pnas.69.9.2495
6. Zeeuws R and Strosberg AD. Proceedings: Extensive sequence differences in rabbit light chains of allotype b4 and b9. Arch Int Physiol Biochim. (1975) 83:205–6.
7. Pinho SS, Alves I, Gaifem J, and Rabinovich GA. Immune regulatory networks coordinated by glycans and glycan-binding proteins in autoimmunity and infection. Cell Mol Immunol. (2023) 20:1101–13. doi: 10.1038/s41423-023-01074-1
8. Rudd PM, Elliott T, Cresswell P, Wilson IA, and Dwek RA. Glycosylation and the immune system. Science. (2001) 291:2370–6. doi: 10.1126/science.291.5512.2370
9. Van der Loo W, Bousses P, Arthur CP, and Chapuis JL. Compensatory aspects of allele diversity at immunoglobulin loci: gene correlations in rabbit populations devoid of light chain diversity (Oryctolagus cuniculus L.; Kerguelen Islands). Genetics. (1996) 144:1181–94. doi: 10.1093/genetics/144.3.1181
10. van der Loo W, Ferrand N, and Soriguer RC. Estimation of gene diversity at the b locus of the constant region of the immunoglobulin light chain in natural populations of European rabbit (Oryctolagus cuniculus) in Portugal, Andalusia and on the Azorean Islands. Genetics. (1991) 127:789–99. doi: 10.1093/genetics/127.4.789
11. van der Loo W, Mougel F, Sanchez MS, Bouton C, Castien E, Fonseca A, et al. Cytonuclear disequilibria in wild populations of rabbit (Oryctolagus cuniculus L.) suggest unequal allele turnover rates at the b locus (IGKC1). Immunogenetics. (1999) 49:629–43.90490629.251[pii
12. Bernstein KE, Skurla RM Jr., and Mage RG. The sequences of rabbit kappa light chains of b4 and b5 allotypes differ more in their constant regions than in their 3’ untranslated regions. Nucleic Acids Res. (1983) 11:7205–14. doi: 10.1093/nar/11.20.7205
13. Pinheiro A, Neves F, Lemos de Matos A, Abrantes J, van der Loo W, Mage R, et al. An overview of the lagomorph immune system and its genetic diversity. Immunogenetics. (2016) 68:83–107. doi: 10.1007/s00251-015-0868-8
14. Figueroa F, Gúnther E, and Klein J. MHC polymorphism pre-dating speciation. Nature. (1988) 335:265–7. doi: 10.1038/335265a0
15. Bouton C and van der Loo W. The trans-species nature of rabbit b locus polymorphism is supported by studies on the snow-shoe hare. Immunogenetics. (1997) 45:444–6. doi: 10.1007/s002510050229
16. Côrte-Real JV, Pinheiro A, Sampson JM, Morrissey KA, Marques JP, Baldauf HM, et al. Deciphering the origins of guanylate binding proteins in mammals (Monotreme, Marsupials and Placentals). BMC Biol. (2025) 23:292. doi: 10.1186/s12915-025-02403-8
17. Pinheiro A, Borges JR, Corte-Real JV, and Esteves PJ. Evolution of guanylate binding protein genes shows a remarkable variability within bats (Chiroptera). Front Immunol. (2024) 15:1329098. doi: 10.3389/fimmu.2024.1329098
18. Matos MC, Pinheiro A, Melo-Ferreira J, Davis RS, and Esteves PJ. Evolution of fc receptor-like scavenger in mammals. Front Immunol. (2020) 11:590280. doi: 10.3389/fimmu.2020.590280
19. Fernandes AP, Agueda-Pinto A, Pinheiro A, Rebelo H, and Esteves PJ. Evolution of TRIM5 and TRIM22 in bats reveals a complex duplication process. Viruses. (2022) 14(2):345. doi: 10.3390/v14020345
20. Thompson JD, Higgins DG, and Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. (1994) 22:4673–80. doi: 10.1093/nar/22.22.4673
21. Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser. (1999) 41:3.
22. Lefranc MP, Pommie C, Kaas Q, Duprat E, Bosc N, Guiraudou D, et al. IMGT unique numbering for immunoglobulin and T cell receptor constant domains and Ig superfamily C-like domains. Dev Comp Immunol. (2005) 29:185–203. S0145-305X(04)00106-5
23. Gupta R and Brunak S. Prediction of glycosylation across the human proteome and the correlation to protein function. Pac Symp Biocomput. (2002), 310–22.
24. Esselstyn JA, Oliveros CH, Swanson MT, and Faircloth BC. Investigating difficult nodes in the placental mammal tree with expanded taxon sampling and thousands of ultraconserved elements. Genome Biol Evol. (2017) 9:2308–21. doi: 10.1093/gbe/evx168
25. Smith AT, Johnston CH, Alves PC, and Hackla K. Lagomorphs: pikas, rabbits, and hares of the world. In: Illustrations (Chiefly colour), colour maps. Johns Hopkins University Press, Baltimore (2018). p. 266.
26. Iracabal L, Barbosa MR, Selvatti AP, and Russo CAM. Molecular time estimates for the Lagomorpha diversification. PloS One. (2024) 19:e0307380. doi: 10.1371/journal.pone.0307380
27. Matthee CA, van Vuuren BJ, Bell D, and Robinson TJ. A molecular supermatrix of the rabbits and hares (Leporidae) allows for the identification of five intercontinental exchanges during the Miocene. Syst Biol. (2004) 53:433–47. doi: 10.1080/10635150490445715
28. Kristic J and Lauc G. The importance of IgG glycosylation-What did we learn after analyzing over 100,000 individuals. Immunol Rev. (2024) 328:143–70. doi: 10.1111/imr.13407
29. Pinheiro A, Lanning D, Alves PC, Mage RG, Knight KL, van der Loo W, et al. Molecular bases of genetic diversity and evolution of the immunoglobulin heavy chain variable region (IGHV) gene locus in leporids. Immunogenetics. (2011) 63:397–408. doi: 10.1007/s00251-011-0533-9
30. Sun Y, Huang T, Hammarstrom L, and Zhao Y. The immunoglobulins: new insights, implications, and applications. Annu Rev Anim Biosci. (2020) 8:145–69. doi: 10.1146/annurev-animal-021419-083720
31. Doron I, Kusakabe T, and Iliev ID. Immunoglobulins at the interface of the gut mycobiota and anti-fungal immunity. Semin Immunol. (2023) 67:101757. doi: 10.1016/j.smim.2023.101757
32. Springer MS, Murphy WJ, Eizirik E, and O’Brien SJ. Placental mammal diversification and the Cretaceous-Tertiary boundary. Proc Natl Acad Sci U S A. (2003) 100:1056–61. doi: 10.1073/pnas.0334222100
33. Ge D, Wen Z, Xia L, Zhang Z, Erbajeva M, Huang C, et al. Evolutionary history of lagomorphs in response to global environmental change. PloS One. (2013) 8:e59668. doi: 10.1371/journal.pone.0059668
34. Esteves PJ, Lanning D, Ferrand N, Knight KL, Zhai SK, and van der Loo W. The evolution of the immunoglobulin heavy chain variable region (IgVH) in Leporids: an unusual case of transspecies polymorphism. Immunogenetics. (2005) 57:874–82. doi: 10.1007/s00251-005-0022-0
35. Pinheiro A, de Mera IG, Alves PC, Gortazar C, de la Fuente J, and Esteves PJ. Sequencing of modern Lepus VDJ genes shows that the usage of VHn genes has been retained in both Oryctolagus and Lepus that diverged 12 million years ago. Immunogenetics. (2013) 65:777–84. doi: 10.1007/s00251-013-0728-3
36. Pinheiro A, de Sousa-Pereira P, and Esteves PJ. The IgA of hares (Lepus sp.) and rabbit confirms that the leporids IgA explosion is old and reveals a new case of trans-species polymorphism. Front Immunol. (2023) 14:1192460. doi: 10.3389/fimmu.2023.1192460
37. Million KM and Lively CM. Trans-specific polymorphism and the convergent evolution of supertypes in major histocompatibility complex class II genes in darters (Etheostoma). Ecol Evol. (2022) 12:e8485. doi: 10.1002/ece3.8485
38. Nei M and Rooney AP. Concerted and birth-and-death evolution of multigene families. Annu Rev Genet. (2005) 39:121–52. doi: 10.1146/annurev.genet.39.073003.112240
39. Carson AR and Scherer SW. Identifying concerted evolution and gene conversion in mammalian gene pairs lasting over 100 million years. BMC Evol Biol. (2009) 9:156. doi: 10.1186/1471-2148-9-156
40. de Haan N, Falck D, and Wuhrer M. Monitoring of immunoglobulin N- and O-glycosylation in health and disease. Glycobiology. (2020) 30:226–40. doi: 10.1093/glycob/cwz048
41. van de Bovenkamp FS, Hafkenscheid L, Rispens T, and Rombouts Y. The emerging importance of igG fab glycosylation in immunity. J Immunol. (2016) 196:1435–41. doi: 10.4049/jimmunol.1502136
42. Koers J, Derksen NIL, Ooijevaar-de Heer P, Nota B, van de Bovenkamp FS, Vidarsson G, et al. Biased N-Glycosylation Site Distribution and Acquisition across the Antibody V Region during B Cell Maturation. J Immunol. (2019) 202:2220–8. doi: 10.4049/jimmunol.1801622
43. Arnold JN, Wormald MR, Sim RB, Rudd PM, and Dwek RA. The impact of glycosylation on the biological function and structure of human immunoglobulins. Annu Rev Immunol. (2007) 25:21–50. doi: 10.1146/annurev.immunol.25.022106.141702
44. Koers J, Sciarrillo R, Derksen NIL, Vletter EM, Fillie-Grijpma YE, Raveling-Eelsing E, et al. Differences in IgG autoantibody Fab glycosylation across autoimmune diseases. J Allergy Clin Immunol. (2023) 151:1646–54. doi: 10.1016/j.jaci.2022.10.035
45. Valk AM, Koers J, Derksen NIL, Hogenboom L, van Kempen Z, Killestein J, et al. Elevated Fab glycosylation of autoantibodies maintained during B cell depletion therapy. Sci Rep. (2025) 15:14770. doi: 10.1038/s41598-025-99226-y
46. Sabouri Z, Schofield P, Horikawa K, Spierings E, Kipling D, Randall KL, et al. Redemption of autoantibodies on anergic B cells by variable-region glycosylation and mutation away from self-reactivity. Proc Natl Acad Sci U S A. (2014) 111:E2567–75. doi: 10.1073/pnas.1406974111
47. Kissel T, Ge C, Hafkenscheid L, Kwekkeboom JC, Slot LM, Cavallari M, et al. Surface Ig variable domain glycosylation affects autoantigen binding and acts as threshold for human autoreactive B cell activation. Sci Adv. (2022) 8:eabm1759. doi: 10.1126/sciadv.abm1759
48. Steffen U, Koeleman CA, Sokolova MV, Bang H, Kleyer A, Rech J, et al. IgA subclasses have different effector functions associated with distinct glycosylation profiles. Nat Commun. (2020) 11:120. doi: 10.1038/s41467-019-13992-8
49. Epstein B, Jones M, Hamede R, Hendricks S, McCallum H, Murchison EP, et al. Rapid evolutionary response to a transmissible cancer in Tasmanian devils. Nat Commun. (2016) 7:12684. doi: 10.1038/ncomms12684
Keywords: evolution, lagomorphs, IGKC, mammals, gene duplication
Citation: Pinheiro A, Borges JR, Marques JP and Esteves PJ (2025) The evolutionary history of IGKC in mammals reveals ancient duplications and remarkable divergence in lagomorphs. Front. Immunol. 16:1686094. doi: 10.3389/fimmu.2025.1686094
Received: 14 August 2025; Accepted: 26 September 2025;
Published: 15 October 2025.
Edited by:
Sofia Kossida, Université de Montpellier, FranceReviewed by:
Shamsa Batool, Université de Montpellier, FranceIfigeneia Sideri, Université de Montpellier, France
Copyright © 2025 Pinheiro, Borges, Marques and Esteves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro José Esteves, cGplc3RldmVzQGNpYmlvLnVwLnB0