 Joao Carlos Batista-Liz1†
Joao Carlos Batista-Liz1† María Sebastián Mora-Gil1†
María Sebastián Mora-Gil1† Mónica Renuncio García2María Teresa Leonardo3Ana Peñalba3Ligia Gabrie1,4Rafael Gálvez Sánchez1,4Luis Martín-Penagos1,5
Mónica Renuncio García2María Teresa Leonardo3Ana Peñalba3Ligia Gabrie1,4Rafael Gálvez Sánchez1,4Luis Martín-Penagos1,5 Javier Narvaez6Belén Sevilla-Pérez7Raquel Ríos Fernández8José Luis Callejas-Rubio8
Javier Narvaez6Belén Sevilla-Pérez7Raquel Ríos Fernández8José Luis Callejas-Rubio8 Luis Caminal-Montero9Paz Collado10José Javier Pérez Venegas11María José Rodríguez Valls11Diego De Árgila12Patricia Quiroga Colina13
Luis Caminal-Montero9Paz Collado10José Javier Pérez Venegas11María José Rodríguez Valls11Diego De Árgila12Patricia Quiroga Colina13 Esther Francisca Vicente Rabaneda13Esteban Rubio14Manuel León Luque14Juan María Blanco-Madrigal15
Esther Francisca Vicente Rabaneda13Esteban Rubio14Manuel León Luque14Juan María Blanco-Madrigal15 Eva Galíndez-Agirregoikoa15
Eva Galíndez-Agirregoikoa15 Javier Gonzalo Ocejo-Vinyals16,17
Javier Gonzalo Ocejo-Vinyals16,17 Ricardo Blanco1,4
Ricardo Blanco1,4 Verónica Pulito-Cueto1*‡
Verónica Pulito-Cueto1*‡ Raquel López-Mejías1*‡
Raquel López-Mejías1*‡- 1Immunopathology Group, Instituto de Investigación Marqués de Valdecilla (IDIVAL), Santander, Spain
- 2Immunology Unit, Complejo Asistencial Universitario de León, León, Spain
- 3Division of Paediatrics, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 4Division of Rheumatology, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 5Division of Nephrology, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 6Division of Rheumatology, Hospital Universitario de Bellvitge, Barcelona, Spain
- 7Division of Paediatrics, Hospital Universitario Clínico San Cecilio, Granada, Spain
- 8Systemic Autoimmune Disease Unit, Hospital Universitario Clínico San Cecilio, Instituto de Investigación Biosanitaria Ibs.GRANADA, Granada, Spain
- 9Internal Medicine Department, Hospital Universitario Central de Asturias, Instituto de Investigación Sanitaria del Principado de Asturias (ISPA), Oviedo, Spain
- 10Division of Rheumatology, Hospital Universitario Severo Ochoa, Madrid, Spain
- 11Division of Rheumatology, Hospital General de Jerez, Jerez de la Frontera, Spain
- 12Division of Dermatology, Hospital Universitario de La Princesa, Madrid, Spain
- 13Division of Rheumatology, Hospital Universitario de La Princesa, Instituto de Investigaciones Sanitarias (IIS)-Princesa, Universidad Autónoma de Madrid, Madrid, Spain
- 14Division of Rheumatology, Hospital Universitario Virgen del Rocío, Sevilla, Spain
- 15Division of Rheumatology, Hospital Universitario de Basurto, Bilbao, Spain
- 16Department of Immunology, Hospital Universitario Marqués de Valdecilla, Santander, Spain
- 17Infectious Diseases and Clinical Microbiology Group, Instituto de Investigación Marqués de Valdecilla (IDIVAL), Santander, Spain
Introduction: Immunoglobulin A vasculitis (IgAV) is an inflammatory disease mediated by B cells. Nuclear factor kappa B (NF-κB) is essential for B-cell development and maturation and plays a key role in autoimmunity and inflammation. In particular, the NF-κB canonical activation pathway genes NFKB1 (encoding NF-κB1) and NFKBIA (encoding NF-κB inhibitor alpha) have been identified as risk loci for several immune-mediated diseases, but their role in IgAV remains unclear. This study aimed to determine whether NFKB1 and NFKBIA represent novel genetic risk factors for IgAV pathogenesis.
Methods: The NFKB1 promoter variant −94 ins/del ATTG (rs28362491), six tag NFKB1 polymorphisms (rs77830930, rs1598856, rs7340881, rs4648055, rs4648090, and rs230547), and seven tag NFKBIA variants (rs3138055, rs696, rs1022714, rs2233419, rs2233415, rs1050851, and rs1957106) were genotyped in 343 Caucasian IgAV patients and 764 healthy, ethnically matched controls using TaqMan probes. Patients were stratified according to age at disease onset and the presence or absence of renal, articular, and gastrointestinal manifestations. Genotype, allele, and haplotype frequencies were compared between patients and controls, as well as across clinical subgroups.
Results: No statistically significant differences were found in genotype or allele frequencies of NFKB1 or NFKBIA between IgAV patients and healthy controls. Likewise, haplotype frequencies of both genes were similar across groups. No associations were observed when patients were stratified by clinical features, including renal involvement, age at onset, or articular/gastrointestinal symptoms.
Conclusion: Our findings do not support a major role for the NFKB1 or NFKBIA variants studied in IgAV susceptibility or severity. These results suggest that if NF-κB signaling contributes to IgAV pathogenesis, it likely involves other biological mechanisms.
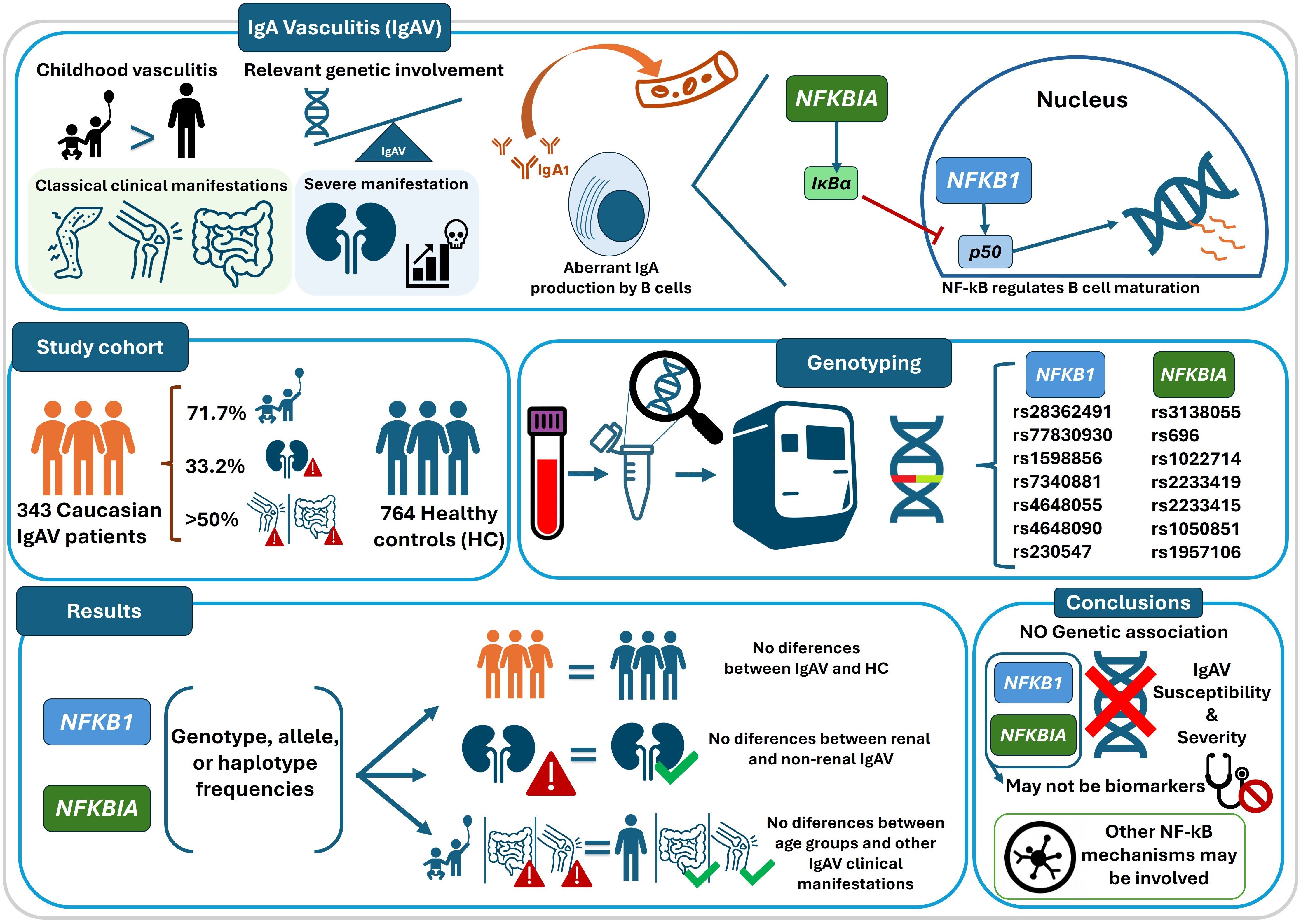
Graphical Abstract. Graphical summary of the study design, genotyping analyses, and main findings on NFKB1 and NFKBIA polymorphisms in IgA vasculitis. Some icons in this figure were adapted from Flaticon (www.flaticon.com).
Introduction
Immunoglobulin A vasculitis (IgAV) is an inflammatory disease characterized by the deposition of immune complexes of aberrantly glycosylated IgA1 in the walls of small blood vessels, leading to inflammatory responses and tissue injury (1–3). IgAV occurs more frequently in children and is the most common systemic vasculitis in the pediatric population (4, 5), although adults can also be affected (6, 7). The most common clinical manifestations of IgAV are the classic triad of palpable purpura, arthralgia/arthritis, and gastrointestinal (GI) symptoms (8–10). However, the most life-threatening complication is the development of renal manifestations (2, 3, 11). In this regard, adult patients have poorer outcomes than children, as they are more likely to experience chronic renal damage and progress to end-stage renal disease (3, 6, 12). Moreover, the severity of renal involvement does not always correlate with the initial presentation of the disease (2, 10, 13). Accurate diagnosis of IgAV and confirmation of nephritis require skin and renal biopsy, which are highly invasive procedures. Consequently, diagnosis is often delayed, and early detection of nephritis is difficult, leading to poorer patient outcomes (14). Identifying reliable disease biomarkers could therefore provide a minimally invasive complementary tool to support earlier and more accurate diagnosis, as well as to enable the detection and treatment of severe nephritis cases in the initial stages of renal injury.
Although the pathogenesis of IgAV remains largely unknown, it has been described as a B cell–mediated disease (2). Multiple studies, including our previous work, have highlighted the importance of genetic factors in its pathogenesis (3, 9, 11). Notably, our earlier studies indicated that polymorphisms in B cell activation and signaling genes—such as BAFF, APRIL, BAFFR, CD40, BLK, and BANK1—do not appear to play a significant role in IgAV development (15, 16). However, given the established involvement of B cells in the disease, it is reasonable to hypothesize that other genes regulating B cell function may contribute to IgAV pathogenesis.
In this context, nuclear factor kappa B (NF-κB) is a known regulator of multiple physiological processes and plays a central role in the activation, response, and survival of mature B cells, as well as in the regulation of class switching that leads to IgA production (17–22). Specifically, NF-κB1 (p105), encoded by the NFKB1 gene located on chromosome 4q24, is one of the principal members of the NF-κB family of proteins (23). Following proteolytic processing, NF-κB1 generates p50, the central component of the canonical NF-κB activation pathway (18). The activity of p50 is regulated in the cytoplasm by NF-κB inhibitor alpha (IκBα), encoded by the NFKBIA gene located on chromosome 14q13 (24). NF-κB has been implicated in the pathogenesis of several pediatric autoimmune diseases. In IgAV, tumor necrosis factor–like weak inducer of apoptosis activates NF-κB through IκBα phosphorylation, while studies in juvenile systemic lupus erythematosus and juvenile systemic sclerosis have reported elevated NF-κB levels and reduced peroxisome proliferator-activated receptor gamma expression, further supporting the role of NF-κB–mediated proinflammatory signaling in childhood-onset autoimmunity (25, 26). Interestingly, genetic variants in both NFKB1 and NFKBIA have been proposed as risk loci for multiple immune-mediated diseases (24, 27–31) and, in particular, for other systemic vasculitides such as Behçet’s disease (32). However, the role of these genes in the pathogenesis of IgAV remains to be elucidated.
Considering these precedents, the present study aims to elucidate, for the first time, whether polymorphisms in NFKB1 and NFBKIA are associated with susceptibility to IgAV or with specific clinical and demographic manifestations of the disease, and to explore their potential value as candidate biomarkers in a large, well-characterized cohort of IgAV patients and controls.
Methods
Study population
This study, conducted between 2012 and 2024, included a large multicenter cohort of 343 unrelated Spanish patients diagnosed with IgAV who met the criteria established by Michel et al. (33) and the American College of Rheumatology classification criteria for IgAV (34). Patients were recruited from nine Spanish hospitals: Hospital Universitario Marqués de Valdecilla (Santander), Hospital Universitario Clínico San Cecilio (Granada), Hospital Universitario de Bellvitge (Barcelona), Hospital Universitario Central de Asturias (Oviedo), Hospital Universitario Severo Ochoa (Leganés), Hospital Universitario de La Princesa (Madrid), Hospital Universitario Virgen del Rocío (Seville), Hospital General de Jerez (Jerez de la Frontera), and Hospital Universitario de Basurto (Bilbao). Clinical and demographic data for all participants were collected. The final sample size represents all patients who met the inclusion criteria and were available during the recruitment period across the nine participating centers. Patients were considered to have renal manifestations if they presented hematuria, proteinuria, nephrotic syndrome, or any combination thereof at any time during the disease course and if renal sequelae (persistent renal involvement) were documented at their last follow-up. Articular manifestations were defined by the presence of arthralgia and/or arthritis, and gastrointestinal (GI) manifestations by the occurrence of bowel angina, GI bleeding, or both. For subsequent analyses, IgAV patients were stratified according to age at disease onset (children ≤20 years; adults >20 years) and the presence or absence of renal, articular, and GI manifestations.
In addition, 764 unrelated, ethnically matched individuals without a history of cutaneous vasculitis or other autoimmune diseases were included as healthy controls. These controls were recruited from Hospital Universitario Marqués de Valdecilla (Santander, Spain) and the National DNA Bank Repository (Salamanca, Spain). All patients with IgAV and healthy controls provided written informed consent. The study procedures followed approved guidelines, regulations, and ethical standards in accordance with the principles of the Declaration of Helsinki. The protocol was approved by the Institutional Review Board or Ethics Committee for Clinical Research of Cantabria, Spain.
Single nucleotide polymorphism selection
For this study, the NFKB1 −94 ins/del ATTG (rs28362491) single nucleotide polymorphism (SNP), previously associated with other immune-mediated diseases (28, 30), was selected. In addition, comprehensive tagging of NFKB1 and NFKBIA was performed using data from the 1000 Genomes Project (http://www.internationalgenome.org/) and the Haploview v4.2 software, considering an r2 threshold set at 0.8 and a minimum minor allele frequency (MAF) of 0.10. As a result, six tag SNPs located in NFKB1 (rs77830930, rs1598856, rs7340881, rs4648055, rs4648090, and rs230547) and seven tag NFKBIA SNPs (rs1957106, rs1050851, rs2233415, rs2233419, rs1022714, rs696, and rs3138055) were selected, covering most of the variability in both genes.
Genotyping
Genomic deoxyribonucleic acid (DNA) was extracted from peripheral blood samples of all individuals included in this study using the REALPURE “SSS” kit (RBME04, REAL, Durviz S.L., Spain), based on a salt precipitation method involving selective protein precipitation with a high-salt solution and DNA recovery by alcohol precipitation. All individuals were genotyped for the 14 genetic variants mentioned above using TaqMan genotyping probes (C_61632788_30 for rs28362491, C_102965997_10 for rs77830930, C:_3066480_10 for rs1598856, C:_3066465_10 for rs7340881, C:_3066440_10 for rs4648055, C:32331236_20 for rs4648090, C: 3066430_10 for rs230547, C:_2797261_1_ for rs1957106, C:_2797260_1_ for rs1050851, C::170225_20 for rs2233415, C::165701_10 for rs2233419, C:_7581384_10 for rs1022714, C::145669_30 for rs696, and C:27468306_10 for rs3138055) by real-time polymerase chain reaction (qPCR) in a QuantStudio™ 7 Flex qPCR system (Applied Biosystems, Foster City, CA, USA), according to the manufacturer’s recommended conditions. The corresponding context sequence of each Applied Biosystems TaqMan assay used in this study is provided in Supplementary Table S1.
Negative controls and duplicate samples were included to validate genotyping accuracy.
The NFKB1 and NFKBIA genotypes were analyzed to verify conformity with Hardy–Weinberg equilibrium (HWE).
Statistical analysis
An association analysis comparing the NFKB1 and NFKBIA genotype and allele frequencies between IgAV patients and healthy controls, and among IgAV patients stratified according to the presence of renal manifestations, age at disease onset, and the presence or absence of articular and GI manifestations, was performed using nonparametric tests (chi-squared test or Fisher’s exact test when expected values were <5). Analysis of allelic combinations (haplotypes) was conducted for the seven NFKB1 SNPs and the seven NFKBIA SNPs using Haploview v4.2 software. Haplotype frequencies were calculated, and those with a frequency greater than 5% were selected and compared between the groups mentioned above using the chi-squared test. The strength of all associations was estimated using odds ratios (ORs) and 95% confidence intervals (CIs).
The two-tailed p-values obtained from all statistical analyses were corrected for a false discovery rate (FDR) of 0.05 using the Benjamini–Hochberg procedure. FDR-corrected p-values less than 0.05 were considered statistically significant.
All statistical analyses were performed using STATA statistical software 12 (Stata Corp., College Station, TX, USA). Statistical power calculations were performed with QUANTO v1.2.4 under a log-additive model, using our study design (343 cases and 764 controls, α = 0.05, two-sided). Power was estimated at different ORs and minor allele frequencies (MAFs), considering the tagging threshold of MAF = 0.10 as well as more common variants (MAF ≥ 0.20).
Results
Patient characteristics
Among the 343 patients with IgAV, 71.7% (246) were children (≤20 years), 33.2% (114) developed renal manifestations, 55.1% (189) presented articular involvement, and 51.0% (175) showed GI complications. Supplementary Table S2 summarizes the main demographic characteristics and clinical manifestations of the IgAV patients.
Genotyping results
A genotyping success rate above 98% was achieved for all NFKB1 and NFKBIA polymorphisms. All polymorphisms in NFKB1 and NFKBIA were in Hardy–Weinberg equilibrium (HWE) in healthy controls (p > 0.05 in all cases).
The genotype and allele frequencies of all polymorphisms studied in NFKB1 and NFKBIA among healthy controls were consistent with those reported in the 1000 Genomes Project (https://www.internationalgenome.org/) for European populations.
For our sample size, statistical power was 66% for OR = 1.4 and 82% for OR = 1.5 when MAF = 0.10. For more common variants (MAF ≥ 0.20), power exceeded 80% to detect OR ≥ 1.4.
Association analysis of NFKB1 and NFKBIA polymorphisms between IgAV patients and controls
To determine the influence of NFKB1 and NFKBIA on IgAV susceptibility, we compared the genetic frequencies of these genes between IgAV patients and healthy controls.
In this respect, comparison of the genotype and allele frequencies of the seven NFKB1 and seven NFKBIA polymorphisms yielded no statistically significant differences between IgAV patients and healthy individuals (Tables 1, 2).
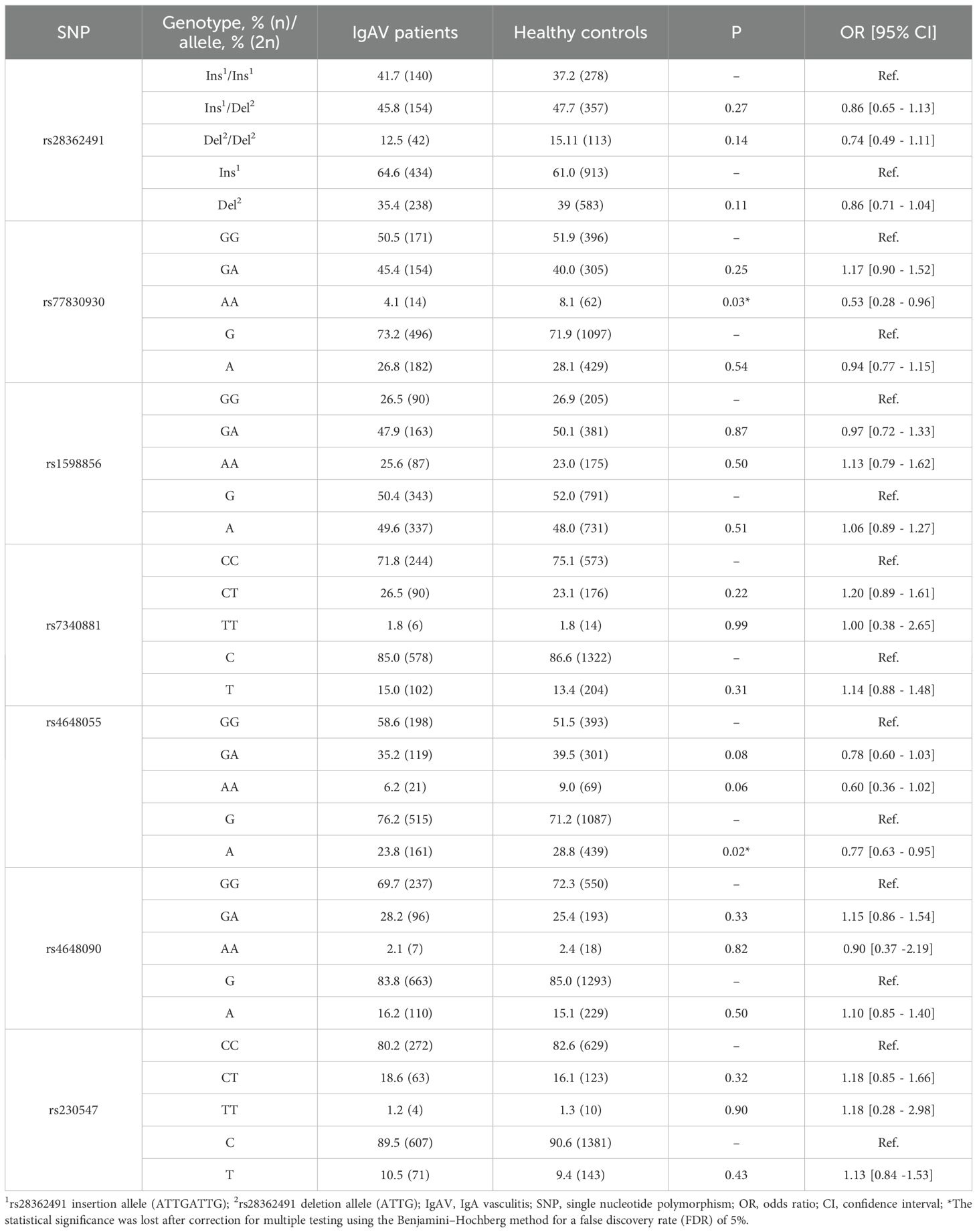
Table 1. Differences in genotype and allele frequencies of NFKB1 between IgAV patients and healthy controls.
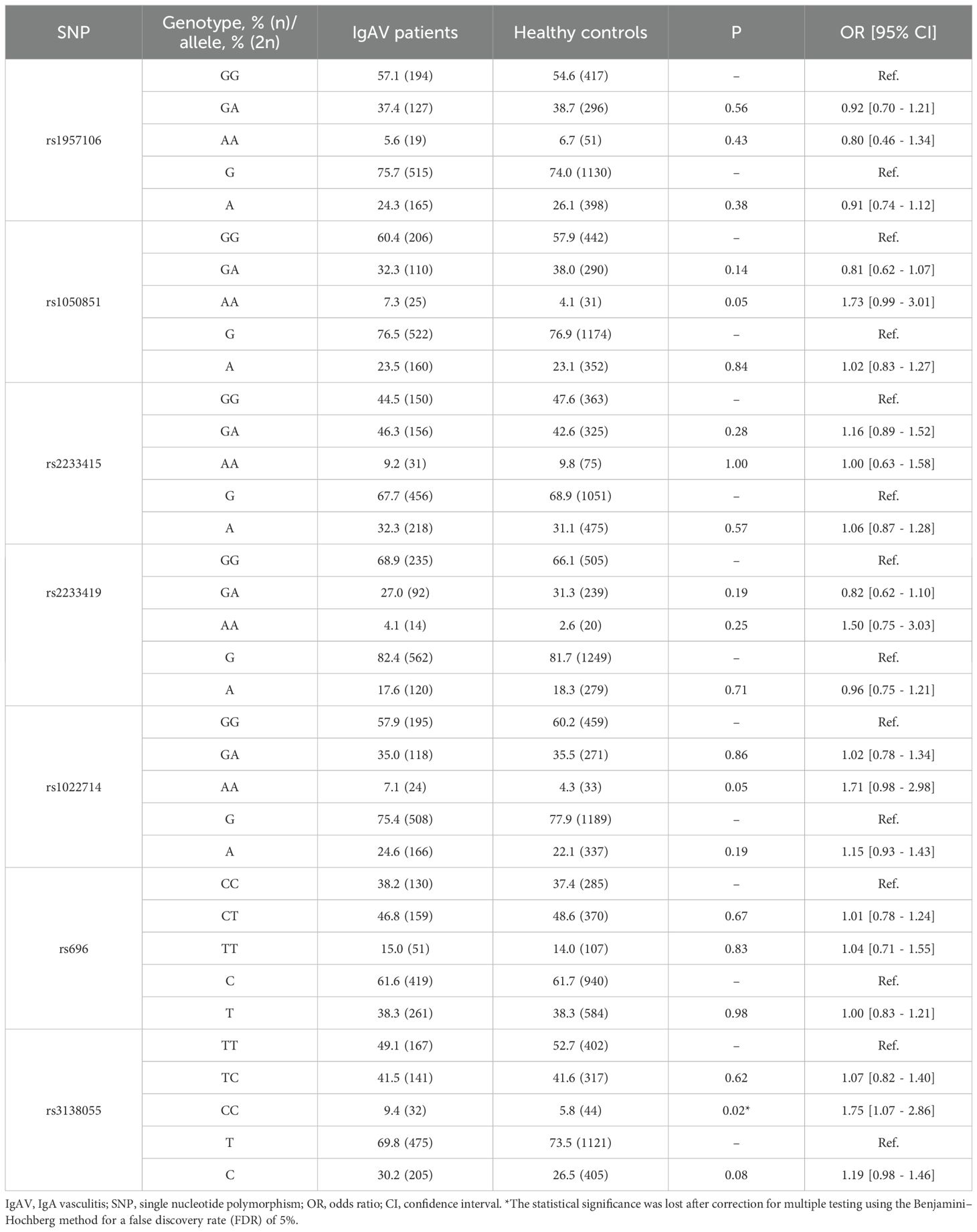
Table 2. Differences in genotype and allele frequencies of NFKBIA between IgAV patients and healthy controls.
Furthermore, no statistically significant differences were observed between the haplotype frequencies of NFKB1 and NFKBIA in IgAV patients and healthy controls (Table 3).
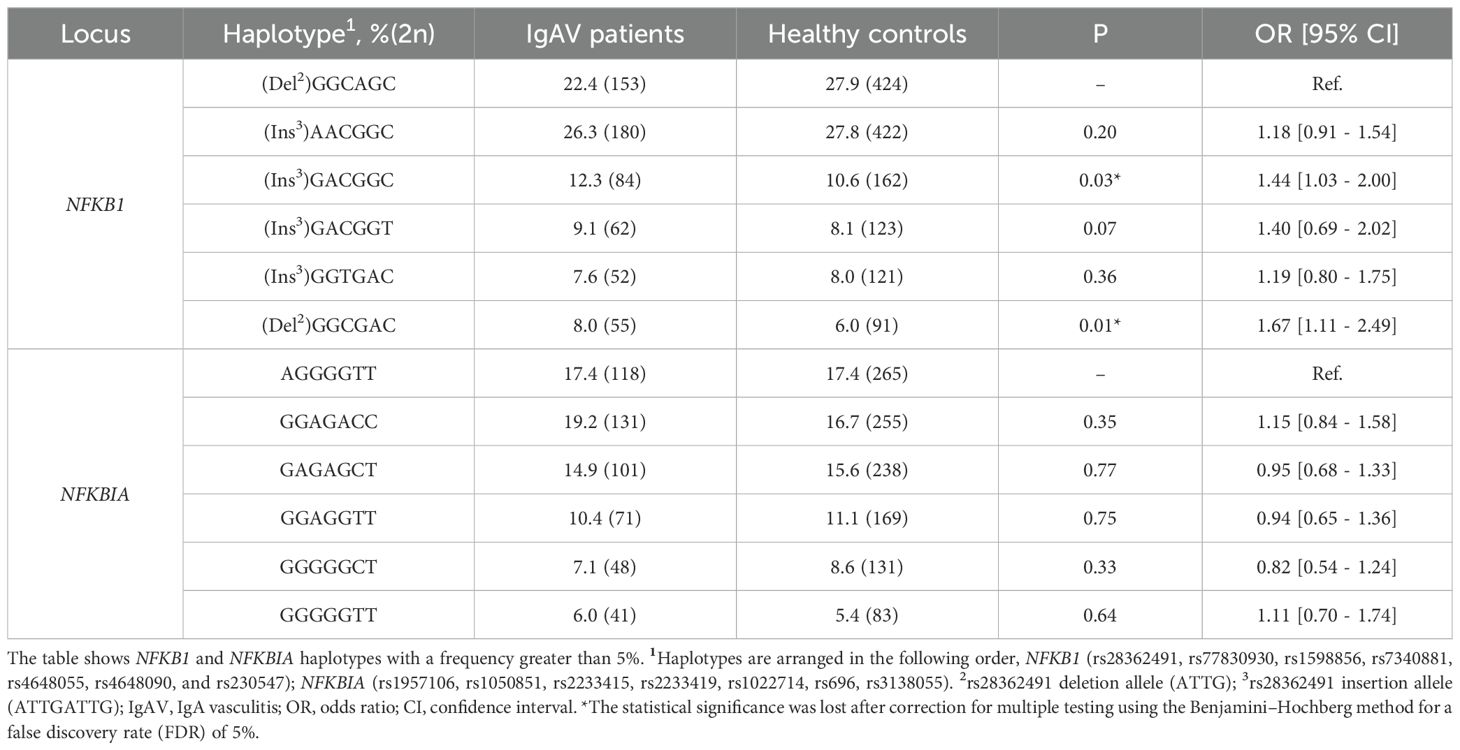
Table 3. Differences in haplotype frequencies of NFKB1 and NFKBIA between IgAV patients and healthy controls.
Stratified analysis of NFKB1 and NFKBIA polymorphisms according to renal manifestations in IgAV
Potential differences in genotype, allele, and haplotype frequencies of NFKB1 and NFKBIA between IgAV patients stratified by disease severity were subsequently investigated. Because renal manifestations represent the most severe complication of IgAV, patients were stratified according to the presence or absence of nephritis.
Our analysis showed that IgAV patients who developed renal manifestations exhibited no statistically significant differences in the genotype and allele frequencies of NFKB1 and NFKBIA compared with those without this complication (Tables 4, 5). Likewise, similar results were obtained when comparing the haplotype frequencies of NFKB1 and NFKBIA between IgAV patients with and without nephritis (Table 6).
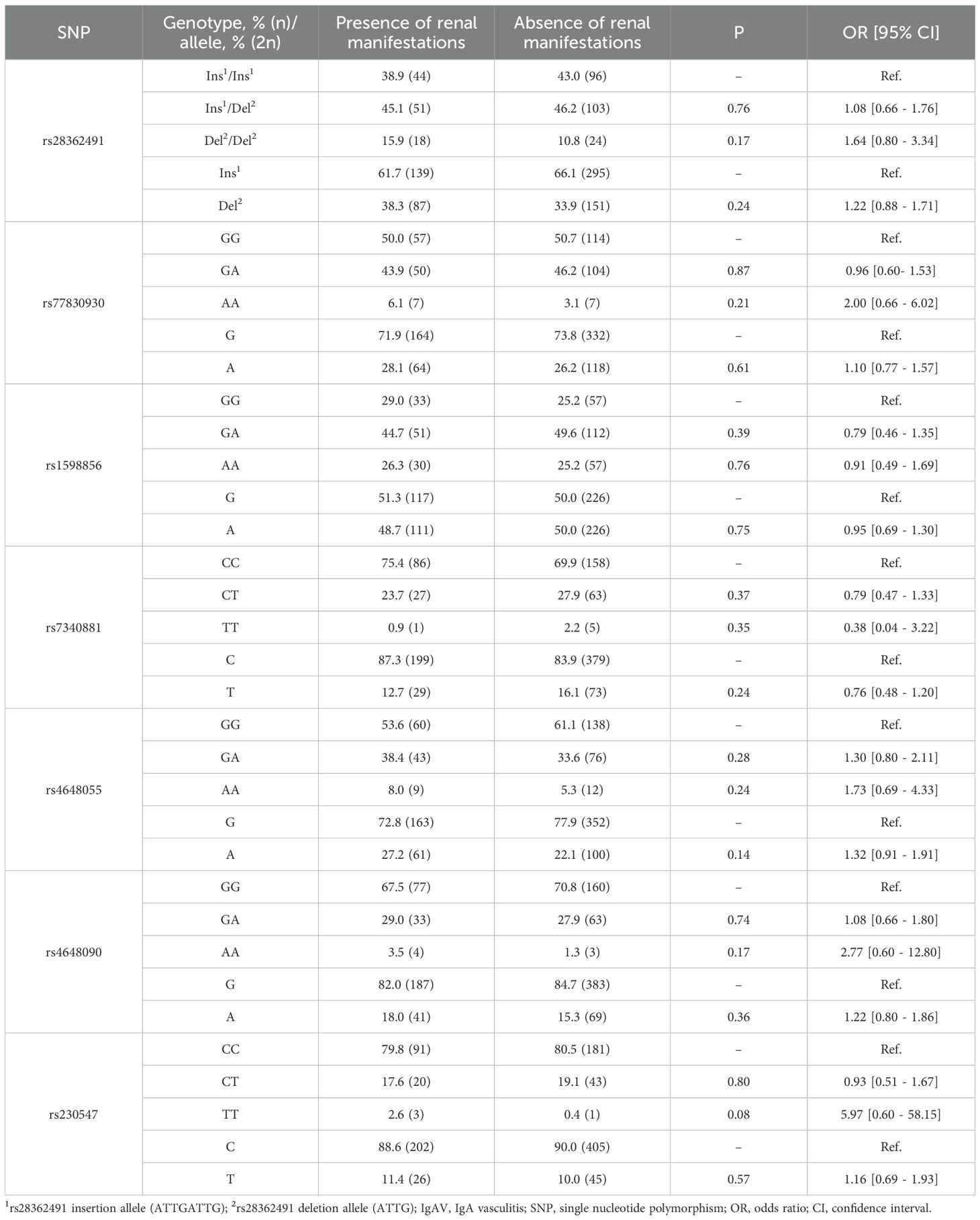
Table 4. Differences in genotype and allele frequencies of NFKB1 in IgAV patients stratified by the presence or absence of renal manifestations.
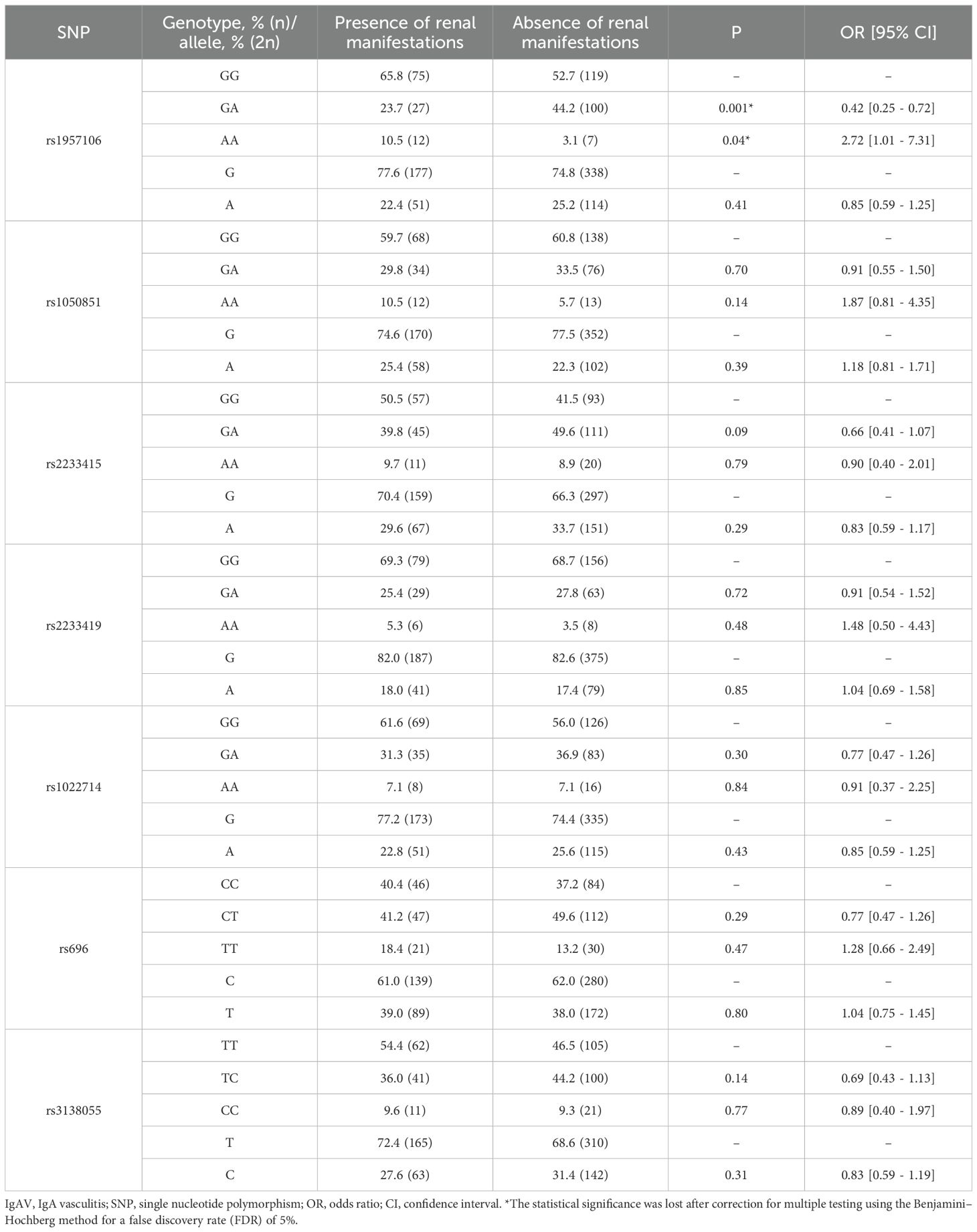
Table 5. Differences in genotype and allele frequencies of NFKBIA in IgAV patients stratified by the presence or absence of renal manifestations.
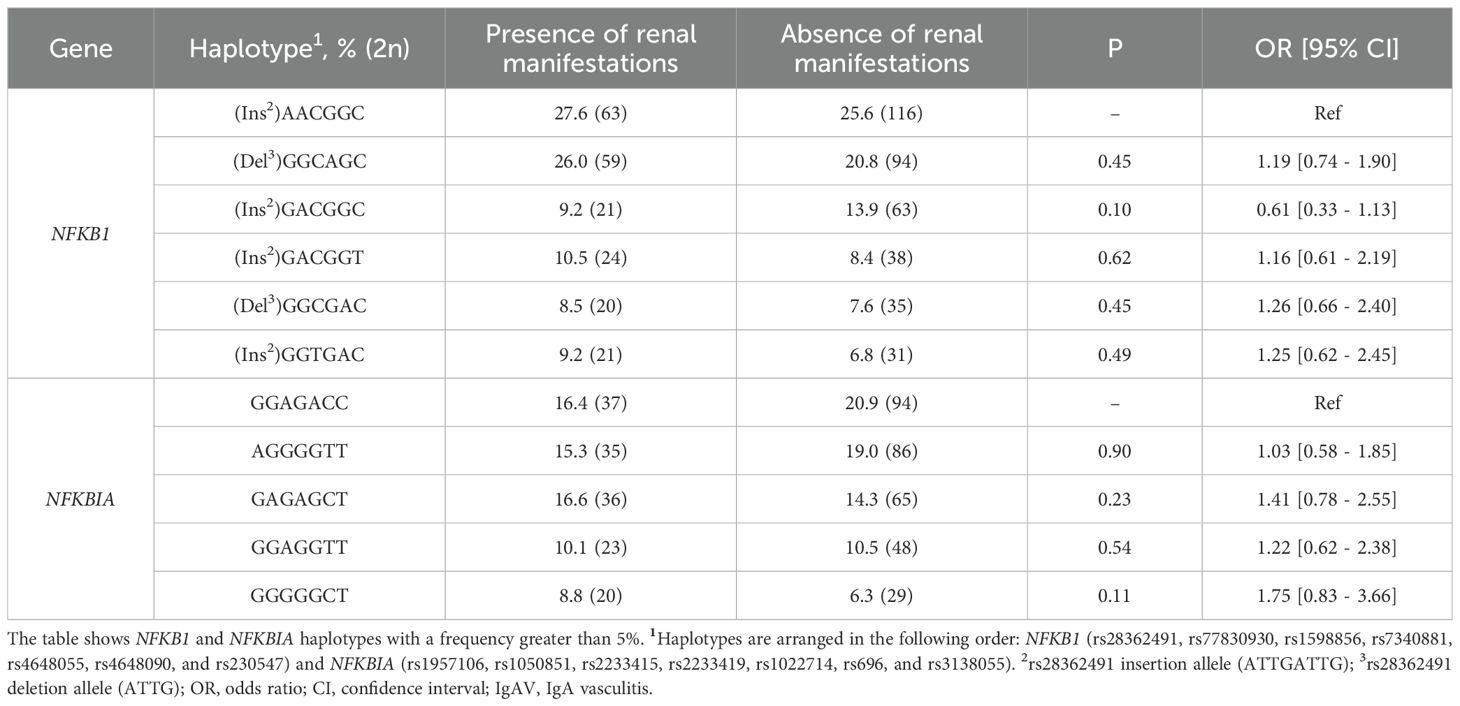
Table 6. Differences in haplotype frequencies of NFKB1 and NFKBIA between IgAV patients stratified by the presence renal manifestations.
Stratified analysis of NFKB1 and NFKBIA polymorphisms by age at onset, articular, and GI manifestations in IgAV
Finally, we investigated whether NFKB1 and NFKBIA polymorphisms were related to age at disease onset and the occurrence of articular and GI symptoms.
No statistically significant differences in the genotype, allele, or haplotype frequencies of NFKB1 and NFKBIA were found between IgAV patients who developed the disease as children (≤20 years) and those who developed it as adults (Supplementary Tables S3-S5).
Likewise, similar genotype and allele frequencies of NFKB1 and NFKBIA were observed in IgAV patients who developed articular or GI manifestations compared with those who did not (Supplementary Tables S3, S4). In addition, haplotype analysis did not reveal any further differences in haplotype frequencies between these patient groups (Supplementary Table S5).
Discussion
NF-κB activity is critical for the activation and survival of B lymphocytes (20–22). NFKB1 and NFKBIA, genes of the canonical NF-κB activation pathway, have been proposed as risk loci for many immune-mediated diseases (24, 27–31). Despite the major role that B cells play in IgAV, the involvement of these genes in this condition is still unknown. Accordingly, we evaluated, for the first time, the implication of NFKB1 and NFKBIA in the pathogenesis of IgAV. For this purpose, we examined most of the variability of NFKB1 and NFKBIA in a large and well-characterized series of 343 Caucasian IgAV patients.
Our results showed no significant differences in genotype, allele, or haplotype frequencies between IgAV patients and healthy individuals, suggesting that neither NFKB1 nor NFKBIA is associated with susceptibility to IgAV. In this regard, there are no previous studies on the involvement of NFKB1 and NFKBIA in IgAV, which emphasizes the importance of our findings. In line with our data, a study in a Spanish cohort with giant cell arteritis (35), a large-vessel vasculitis (36), found no association between the NFKB1 polymorphism −94 ins/del ATTG (rs28362491) and disease susceptibility. Accordingly, we could hypothesize that the evaluation of polymorphisms in these genes does not appear to be useful for identifying these systemic vasculitides in Caucasian populations in clinical practice. Our results show, for the first time, that NFKB1 and NFKBIA may not constitute suitable biomarkers for IgAV susceptibility.
In the context of IgAV severity, we found no association between NFKB1 and NFKBIA polymorphisms and the presence of renal manifestations, suggesting that these genes may not be associated with the risk of developing nephritis in IgAV. No previous studies have examined the implication of genes in the NF-κB1 pathway in IgA-mediated renal inflammatory diseases in Caucasian populations. Consequently, a novel finding has emerged from our study, revealing that NFKB1 and NFKBIA may not be useful in clinical practice for predicting renal damage in IgAV.
Regarding the implication of NFKB1 and NFKBIA in other demographic and clinical manifestations of IgAV, we observed no relationship between polymorphisms in these genes and the age at disease onset or the development of articular manifestations. Concerning the latter, and in agreement with our findings, no influence of NFKB1 and NFKBIA on the pathogenesis of rheumatoid arthritis—the prototypical articular inflammatory disease—has been reported in Spanish patients (37, 38). Furthermore, no involvement of NFKB1 and NFKBIA polymorphisms in the development of GI complications was detected in our cohort of IgAV patients. In this context, GI symptoms of IgAV can be difficult to distinguish from those of Crohn’s disease (CD) and ulcerative colitis (UC) (39–41). Interestingly, NFKB1 −94 ins/del ATTG (rs28362491) and NFKBIA rs696 have previously been linked to an increased risk of CD and UC, respectively (28, 42, 43). Therefore, and supported by our findings, polymorphisms in NFKB1 and NFKBIA may serve as potential biomarkers for differentiating IgAV cases presenting only GI manifestations from CD and UC.
Our findings suggest that polymorphisms in the canonical NF-κB pathway genes NFKB1 and NFKBIA are unlikely to exert a major influence on IgAV susceptibility or its clinical presentation. By using a tagging strategy, we captured most of the common variability within these loci, which strengthens the robustness of our results. While our study design and sample size provide a solid foundation, several diagnostic considerations should be acknowledged. Although IgAV was classified according to established criteria, histopathological confirmation by skin or renal biopsy was not uniformly available, as these procedures are invasive and not routinely performed in all patients. These diagnostic limitations should be considered when interpreting our results, although the large sample size and use of standardized criteria mitigate this concern. Interestingly, previous work from our group assessing other canonical NF-κB regulators (IL33, IL1R1, VAV3, and CARD9) (16, 44) also found no association with IgAV, further supporting a lack of contribution from the variants studied in this pathway to the disease. Nevertheless, the absence of genetic associations does not exclude the possibility that NF-κB signaling contributes to IgAV pathogenesis through mechanisms such as regulatory or epigenetic alterations or post-translational modifications.
Future studies integrating epigenomic, transcriptomic, and functional approaches will be necessary to fully delineate the contribution of NF-κB signaling to IgAV pathogenesis. Moreover, given the central role of NF-κB in immune regulation, the involvement of genes within the noncanonical pathway remains an important area for further exploration. Likewise, studies including non-Caucasian cohorts would be of interest to confirm and broaden the applicability of our findings across different ethnic backgrounds.
Conclusions
In conclusion, in a large and well-characterized Caucasian cohort of IgAV patients representing the full clinical spectrum of the disease, our results do not support an association between common NFKB1 and NFKBIA polymorphisms and IgAV susceptibility or severity, thereby contributing to refining the genetic landscape of this vasculitis.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Ethics statement
The studies involving humans were approved by the Institutional Review Board or Ethics Committee for Clinical Research of Cantabria, Spain. The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants' legal guardians/next of kin.
Author contributions
JB-L: Data curation, Investigation, Visualization, Formal Analysis, Software, Writing – review & editing, Methodology, Writing – original draft. MS-G: Investigation, Methodology, Writing – review & editing, Visualization, Formal Analysis, Writing – original draft, Data curation. MRG: Writing – review & editing, Writing – original draft, Data curation. MTL: Data curation, Writing – original draft, Writing – review & editing. AP: Data curation, Writing – review & editing, Writing – original draft. LG: Writing – review & editing, Writing – original draft, Data curation. RG: Writing – original draft, Data curation, Writing – review & editing. LM-P: Writing – review & editing, Writing – original draft, Data curation. JN: Writing – original draft, Data curation, Writing – review & editing. BS-P: Writing – original draft, Writing – review & editing, Data curation. RR: Writing – review & editing, Writing – original draft, Data curation. JC-R: Writing – original draft, Writing – review & editing, Data curation. LC-M: Writing – review & editing, Data curation, Writing – original draft. PC: Data curation, Writing – original draft, Writing – review & editing. JP: Writing – original draft, Data curation, Writing – review & editing. MRV: Writing – original draft, Data curation, Writing – review & editing. DD: Writing – review & editing, Writing – original draft, Data curation. PQ: Writing – review & editing, Writing – original draft, Data curation. EV: Data curation, Writing – review & editing, Writing – original draft. ER: Writing – review & editing, Writing – original draft, Data curation. MLL: Writing – review & editing, Writing – original draft, Data curation. JB-M: Writing – review & editing, Writing – original draft, Data curation. EG-A: Writing – original draft, Data curation, Writing – review & editing. JO-V: Data curation, Writing – review & editing, Writing – original draft. RB: Writing – review & editing, Writing – original draft, Conceptualization, Project administration. VP-C: Project administration, Supervision, Writing – review & editing, Methodology, Writing – original draft, Investigation. RL-M: Supervision, Methodology, Conceptualization, Writing – review & editing, Funding acquisition, Writing – original draft, Project administration.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This research was funded by European Union FEDER funds and “Fondo de Investigaciones Sanitarias” from “Instituto de Salud Carlos III” (ISCIII, Health Ministry, Spain), grant numbers PI21/00042 and PI24/00382. JCB-L. is a recipient of a PFIS program fellowship from the ISCIII, co-funded by the European Social Fund (‘Investing in your future’), grant number FI22/00020. VP-C received funding from IDIVAL, grant numbers NVAL23/02 and INNVAL24/10. RL-M is a recipient of a Miguel Servet type II program fellowship from the ISCIII, co-funded by ESF (“Investing in your future”), grant number CPII21/00004.
Acknowledgments
We would like to thank Dr. Castañeda and Dr. González-Gay for their contribution to previous studies in the IgAV field. In addition, we wish to thank all the subjects for their essential collaboration in this study. We also thank the National DNA Bank Repository (Salamanca) for supplying the control samples.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1692908/full#supplementary-material
Abbreviations
ACR, American College of Rheumatology; APRIL, A Proliferation-Inducing Ligand; BAFF, B-cell Activating Factor; BAFFR, BAFF Receptor; BANK1, B-cell Scaffold Protein with Ankyrin Repeats 1; BLK, B Lymphoid Tyrosine Kinase; CARD9, Caspase Recruitment Domain Family Member 9; CD, Crohn’s Disease; CD40, Cluster of Differentiation 40; CI, Confidence Interval; DNA, Deoxyribonucleic Acid; FDR, False Discovery Rate; GI, Gastrointestinal; HWE, Hardy-Weinberg equilibrium; IgA1, Immunoglobulin A1; IgAV, IgA Vasculitis; IL-33, Interleukin 33; IL1R1, Interleukin 1 Receptor Type 1; IκBα, Inhibitor of Kappa B Alpha; NF-κB, Nuclear Factor-kappa B; NF-κB1, Nuclear Factor-kappa B Subunit 1; NFKB1, Gene Encoding NF-κB1 (p105/p50); NFKBIA, Gene Encoding IκBα; OR, Odds Ratio; qPCR, Quantitative Polymerase Chain Reaction; SNP, Single-Nucleotide Polymorphism; UC, Ulcerative Colitis; VAV3, Vav Guanine Nucleotide Exchange Factor 3.
References
1. Ding L, Chen X, Cheng H, Zhang T, and Li Z. Advances in IgA glycosylation and its correlation with diseases. Front Chem. (2022) 10:974854. doi: 10.3389/fchem.2022.974854
2. Song Y, Huang X, Yu G, Qiao J, Cheng J, Wu J, et al. Pathogenesis of igA vasculitis: an up-to-date review. Front Immunol. (2021) 12:771619. doi: 10.3389/fimmu.2021.771619
3. Xu L, Li Y, and Wu X. IgA vasculitis update: Epidemiology, pathogenesis, and biomarkers. Front Immunol. (2022) 13:921864. doi: 10.3389/fimmu.2022.921864
4. Barut K, Sahin S, and Kasapcopur O. Pediatric vasculitis. Curr Opin Rheumatol. (2016) 28:29–38. doi: 10.1097/BOR.0000000000000236
5. Bayındır Y, Başaran Ö, Bilginer Y, and Özen S. Vasculitis in children. Turkish Arch Pediatr. (2024) 59:517–26. doi: 10.5152/TurkArchPediatr.2024.24181
6. Blanco R, Martínez-Taboada VM, Rodríguez-Valverde V, García-Fuentes M, and González-Gay MA. Henoch-Schönlein purpura in adulthood and childhood: two different expressions of the same syndrome. Arthritis Rheum. (1997) 40:859–64. doi: 10.1002/art.1780400513
7. Calvo-Río V, Loricera J, Mata C, Martín L, Ortiz-Sanjuán F, Alvarez L, et al. Henoch-schönlein purpura in Northern Spain: Clinical spectrum of the disease in 417 patients from a single center. Med (Baltimore). (2014) 93:106–13. doi: 10.1097/MD.0000000000000019
8. Calvo-Río V, Hernández JL, Ortiz-Sanjuán F, Loricera J, Palmou-Fontana N, González-Vela MC, et al. Relapses in patients with Henoch-Schönlein purpura: Analysis of 417 patients from a single center. Med (Baltimore). (2016) 95:e4217. doi: 10.1097/MD.0000000000004217
9. López-Mejías R, Castañeda S, Genre F, Remuzgo-Martínez S, Carmona FD, Llorca J, et al. Genetics of immunoglobulin-A vasculitis (Henoch-Schönlein purpura): An updated review. Autoimmun Rev. (2018) 17:301–15. doi: 10.1016/j.autrev.2017.11.024
10. Hu Y-C, Yang Y-H, and Chiang B-L. Immunoglobulin A vasculitis: The clinical features and pathophysiology. Kaohsiung J Med Sci. (2024) 40:612–20. doi: 10.1002/kjm2.12852
11. Heineke MH, Ballering AV, Jamin A, Ben Mkaddem S, Monteiro RC, and Van Egmond M. New insights in the pathogenesis of immunoglobulin A vasculitis (Henoch-Schönlein purpura). Autoimmun Rev. (2017) 16:1246–53. doi: 10.1016/j.autrev.2017.10.009
12. Audemard-Verger A, Pillebout E, Baldolli A, Gouellec NL, Augusto J-F, Jourde-Chiche N, et al. Impact of aging on phenotype and prognosis in IgA vasculitis. Rheumatol (Oxford). (2021) 60:4245–51. doi: 10.1093/rheumatology/keaa921
13. Sestan M and Jelusic M. Diagnostic and management strategies of igA vasculitis nephritis/henoch-schönlein purpura nephritis in pediatric patients: current perspectives. Pediatr Health Med Ther. (2023) 14:89–98. doi: 10.2147/PHMT.S379862
14. Stanway J, Brown N, Pervez A, Van de Perre E, Tollitt J, Marketos N, et al. IgA vasculitis nephritis-outcomes in adult-onset disease. Rheumatol (Oxford). (2024) 64:690–6. doi: 10.1093/rheumatology/keae030
15. Batista Liz JC, Genre F, Pulito-Cueto V, Remuzgo-Martínez S, Prieto-Peña D, Márquez A, et al. IgA vasculitis: influence of CD40, BLK and BANK1 gene polymorphisms. J Clin Med. (2022) 11:5577. doi: 10.3390/jcm11195577
16. Batista-Liz JC, Calvo-Río V, Sebastián Mora-Gil M, Sevilla-Pérez B, Márquez A, Leonardo MT, et al. Mucosal immune defence gene polymorphisms as relevant players in the pathogenesis of igA vasculitis? Int J Mol Sci. (2023) 24:13063. doi: 10.3390/ijms241713063
17. Sha WC, Liou H-C, Tuomanen EI, and Baltimore D. Targeted disruption of the p50 subunit of NF-KB leads to multifocal defects in immune responses. Cell. (1995) 80:321–30. doi: 10.1016/0092-8674(95)90415-8
18. Li Q and Verma IM. NF-κB regulation in the immune system. Nat Rev Immunol. (2002) 2:725–34. doi: 10.1038/nri910
19. Beinke S and Ley SC. Functions of NF-kappaB1 and NF-kappaB2 in immune cell biology. Biochem J. (2004) 382:393–409. doi: 10.1042/BJ20040544
20. Sasaki Y and Iwai K. Roles of the NF-κB pathway in B-lymphocyte biology. Curr Top Microbiol Immunol. (2015) 393:177–209. doi: 10.1007/82_2015_479
21. Grumont RJ, Rourke IJ, O’Reilly LA, Strasser A, Miyake K, Sha W, et al. B lymphocytes differentially use the Rel and nuclear factor kappaB1 (NF-kappaB1) transcription factors to regulate cell cycle progression and apoptosis in quiescent and mitogen-activated cells. J Exp Med. (1998) 187:663–74. doi: 10.1084/jem.187.5.663
22. Snapper CM, Zelazowski P, Rosas FR, Kehry MR, Tian M, Baltimore D, et al. B cells from p50/NF-kappa B knockout mice have selective defects in proliferation, differentiation, germ-line CH transcription, and Ig class switching. J Immunol. (1996) 156:183–91. doi: 10.4049/jimmunol.156.1.183
23. Zou YF, Wang F, Feng XL, Tao JH, Zhu JM, Pan FM, et al. Association of NFKB1 -94ins/delATTG promoter polymorphism with susceptibility to autoimmune and inflammatory diseases: a meta-analysis. Tissue Antigens. (2011) 77:9–17. doi: 10.1111/j.1399-0039.2010.01559.x
24. Zhang GL, Zou YF, Feng XL, Shi HJ, Du XF, Shao MH, et al. Association of the NFKBIA gene polymorphisms with susceptibility to autoimmune and inflammatory diseases: A meta-analysis. Inflammation Res. (2011) 60:11–8. doi: 10.1007/s00011-010-0216-2
25. Chen T, Guo Z-P, Li M-M, Li J-Y, Jiao X-Y, Zhang Y-H, et al. Tumour necrosis factor-like weak inducer of apoptosis (TWEAK), an important mediator of endothelial inflammation, is associated with the pathogenesis of Henoch-Schonlein purpura. Clin Exp Immunol. (2011) 166:64–71. doi: 10.1111/j.1365-2249.2011.04442.x
26. Durmus S, Sahin S, Adrovic A, Barut K, Gelisgen R, Uzun H, et al. Interplay of NF-κB and PPAR-γ transcription factors in patients with juvenile systemic lupus erythematosus. Lupus Sci Med. (2025) 12:e001263. doi: 10.1136/lupus-2024-001263
27. Liu C, Yan S, Chen H, Wu Z, Li L, Cheng L, et al. Association of GTF2I, NFKB1, and TYK2 regional polymorphisms with systemic sclerosis in a chinese han population. Front Immunol. (2021) 12:640083. doi: 10.3389/fimmu.2021.640083
28. Karban AS, Okazaki T, Panhuysen CIM, Gallegos T, Potter JJ, Bailey-Wilson JE, et al. Functional annotation of a novel NFKB1 promoter polymorphism that increases risk for ulcerative colitis. Hum Mol Genet. (2004) 13:35–45. doi: 10.1093/hmg/ddh008
29. Zhao Q, Sun Y, Fu X, Wang Z, Yu G, Yue Z, et al. Identification of a single nucleotide polymorphism in NFKBIA with different effects on psoriatic arthritis and cutaneous psoriasis in China. Acta Derm Venereol. (2019) 99:84–8. doi: 10.2340/00015555-3027
30. Elkhawaga SY, Gomaa MH, Elsayed MM, and Ebeed AA. NFKB1 promoter -94 insertion/deletion ATTG polymorphism (rs28362491) is associated with severity and disease progression of rheumatoid arthritis through interleukin-6 levels modulation in Egyptian patients. Clin Rheumatol. (2021) 40:2927–37. doi: 10.1007/s10067-021-05584-z
31. López-Mejías R, García-Bermúdez M, González-Juanatey C, Castañeda S, Miranda-Filloy JA, Gómez-Vaquero C, et al. NFKB1-94ATTG ins/del polymorphism (rs28362491) is associated with cardiovascular disease in patients with rheumatoid arthritis. Atherosclerosis. (2012) 224:426–9. doi: 10.1016/j.atherosclerosis.2012.06.008
32. Yenmis G, Oner T, Cam C, Koc A, Kucuk OS, Yakicier MC, et al. Association of NFKB1 and NFKBIA polymorphisms in relation to susceptibility of Behçet’s disease. Scand J Immunol. (2015) 81:81–6. doi: 10.1111/sji.12251
33. Michel BA, Hunder GG, Bloch DA, and Calabrese. Hypersensitivity vasculitis LH. and Henoch-Schönlein purpura: a comparison between the 2 disorders. J Rheumatol. (1992) 19:721–8.
34. Mills JA, Michel BA, Bloch DA, Calabrese LH, Hunder GG, Arend WP, et al. The American College of Rheumatology 1990 criteria for the classification of henoch-schönlein purpura. Arthritis Rheum. (1990) 33:1114–21. doi: 10.1002/art.1780330809
35. Martin J, Perez-Armengol C, Miranda-Filloy JA, Vilchez JR, Lopez-Nevot MA, Garcia-Porrua C, et al. Lack of association of a functional -94ins/delATTG NFKB1 promoter polymorphism with susceptibility and clinical expression of biopsy-proven giant cell arteritis in northwest Spain. J Rheumatol. (2006) 33:285–8.
36. Jennette JC, Falk RJ, Bacon PA, Basu N, Cid MC, Ferrario F, et al. 2012 Revised International Chapel Hill consensus conference nomenclature of vasculitides. Arthritis Rheum. (2013) 65:1–11. doi: 10.1002/art.37715
37. Orozco G, Sánchez E, Collado MD, López-Nevot MÁ, Paco L, García A, et al. Analysis of the functional NFKB1 promoter polymorphism in rheumatoid arthritis and systemic lupus erythematosus. Tissue Antigens. (2005) 65:183–6. doi: 10.1111/j.1399-0039.2005.00341.x
38. Dieguez-Gonzalez R, Akar S, Calaza M, Perez-Pampin E, Costas J, Torres M, et al. Genetic variation in the nuclear factor κB pathway in relation to susceptibility to rheumatoid arthritis. Ann Rheum Dis. (2009) 68:579–83. doi: 10.1136/ard.2007.087304
39. Hatemi I, Hatemi G, and Çelik AF. Systemic vasculitis and the gut. Curr Opin Rheumatol. (2017) 29:33–8. doi: 10.1097/BOR.0000000000000344
40. Desai TS, Jurencak R, Nair A, and Carman N. Atypical immunoglobulin A vasculitis in a pediatric patient with ulcerative colitis. ACG Case Rep J. (2023) 10:e01145. doi: 10.14309/crj.0000000000001145
41. Yavuz A, Yıldız M, Aydın A, Yıldırım AC, Buluş H, and Köklü S. Henoch Schonlein purpura mimicking Crohn’s ileitis. J Crohns Colitis. (2011) 5:271–2. doi: 10.1016/j.crohns.2011.03.002
42. Bank S, Skytt Andersen P, Burisch J, Pedersen N, Roug S, Galsgaard J, et al. Polymorphisms in the inflammatory pathway genes TLR2, TLR4, TLR9, LY96, NFKBIA, NFKB1, TNFA, TNFRSF1A, IL6R, IL10, IL23R, PTPN22, and PPARG are associated with susceptibility of inflammatory bowel disease in a Danish cohort. PLoS One. (2014) 9:e98815. doi: 10.1371/journal.pone.0098815
43. Borm MEA, van Bodegraven AA, Mulder CJJ, Kraal G, and Bouma G. A NFKB1 promoter polymorphism is involved in susceptibility to ulcerative colitis. Int J Immunogenet. (2005) 32:401–5. doi: 10.1111/j.1744-313X.2005.00546.x
Keywords: biomarkers, immunoglobulin A vasculitis (IgAV), NF-kappa B (NF-kB), NFKB1, NFKBIA
Citation: Batista-Liz JC, Sebastián Mora-Gil M, Renuncio García M, Leonardo MT, Peñalba A, Gabrie L, Sánchez RG, Martín-Penagos L, Narvaez J, Sevilla-Pérez B, Ríos Fernández R, Callejas-Rubio JL, Caminal-Montero L, Collado P, Pérez Venegas JJ, Rodríguez Valls MJ, De Árgila D, Quiroga Colina P, Vicente Rabaneda EF, Rubio E, León Luque M, Blanco-Madrigal JM, Galíndez-Agirregoikoa E, Ocejo-Vinyals JG, Blanco R, Pulito-Cueto V and López-Mejías R (2025) The role of NFKB1 and NFKBIA in immunoglobulin A vasculitis. Front. Immunol. 16:1692908. doi: 10.3389/fimmu.2025.1692908
Received: 26 August 2025; Accepted: 10 October 2025;
Published: 24 October 2025.
Edited by:
Ozgur Kasapcopur, Istanbul University-Cerrahpasa, TürkiyeReviewed by:
Vildan Güngörer, University of Health Sciences, TürkiyeSinem Durmus, Izmir Kâtip Çelebi University, Türkiye
Copyright © 2025 Batista-Liz, Sebastián Mora-Gil, Renuncio García, Leonardo, Peñalba, Gabrie, Sánchez, Martín-Penagos, Narvaez, Sevilla-Pérez, Ríos Fernández, Callejas-Rubio, Caminal-Montero, Collado, Pérez Venegas, Rodríguez Valls, De Árgila, Quiroga Colina, Vicente Rabaneda, Rubio, León Luque, Blanco-Madrigal, Galíndez-Agirregoikoa, Ocejo-Vinyals, Blanco, Pulito-Cueto and López-Mejías. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raquel López-Mejías, cmxvcGV6bWVqaWFzNzhAZ21haWwuY29t; Verónica Pulito-Cueto, dmVyb25pY2FfcHVsaXRvX2N1ZXRvQGhvdG1haWwuY29t
†These authors have contributed equally to this work and share first authorship
‡These authors share senior authorship