 Kaixin Shi1†
Kaixin Shi1† Jincheng Song
Jincheng Song Li Li
Li Li- 1Department of Hematology, The Second Hospital of Dalian Medical University, Dalian, China
- 2Department of Oncology, The Second Hospital of Dalian Medical University, Dalian, China
The role of lipid metabolism in cancer and immune regulation has received significant attention in recent years. Reprogramming of lipid metabolism is one of the key hallmarks of cancer and plays a critical role in cancer progression by supporting the rapid proliferation, survival, and metastasis of tumor cells. Importantly, beyond its well-established functions in cancer cells, lipid metabolism dynamically regulates the functions of various immune cells within the TME (e.g., T cells, natural killer cells, macrophages), thereby molding antitumor immune responses. This review combines the contemporary awareness of the reciprocal interactions between lipid metabolism and the TME. We start with a simple overview of key lipid metabolic pathways in cancer cells, followed by an in-depth exploration of the way lipid uptake, synthesis, and oxidation influence the fate and role of tumor-infiltrating immune. We also appraise the translational potential of targeting lipid metabolism and propose that combining inhibitors of key metabolic enzymes, for example fatty acid synthase or acetyl-CoA carboxylase, with immunotherapy can not only effectively alleviate immunosuppression but also overcome immunosuppression. Finally, we spotlight the remaining knowledge gaps and put forward future research priorities and potential. Intervening in lipid metabolic interactions represents a promising prospect for developing the novel cancer treatment strategies.
1 Introduction
Cancer is the second most common cause of death worldwide and is projected to be the leading cause of death by 2060 (1). In recent years, there has been a significant increase in the interest of abnormal metabolism in cancer cells, and an increased focus on the increasingly important role of lipids in tumorigenesis and cancer progression. In order to survive and divide under unfavorable conditions, such as hypoxia and nutrient-poor conditions, cancer cells re-model the surrounding immune and stromal cells, and consequently build a supportive TME. Within this TME, multiple components interact to provide energy for the tumor and regulate tumor progression through multiple signaling pathways (2). Cancer cells exhibit uncontrolled proliferation and have a high demand for energy. Although angiogenesis is enhanced in the TME, the tumor vasculature cannot fully provide the necessary demands of cancer cells. Therefore, cancer cells reprogram their metabolism and hyperactivate lipid metabolism to support their proliferation.
Growing evidence has revealed that lipid metabolism is generally enhanced at multiple stages of cancer development. Cancer cells contain elevated lipid levels, such as increased uptake of exogenous lipids and lipoproteins, as well as over-activated de novo lipid synthesis. These events directly promote the malignant transformation and progression of tumor cells, as well as the accumulation of abnormal lipids in the TME (3). For instance, the expression of fatty acid synthase (FASN) is upregulated in TNBC, causing lipid accumulation. In addition, lipid metabolism also provides energy for tumor cells. For example, the products of (FAO), such as reduced nicotinamide adenine dinucleotide, flavin adenine dinucleotide, reduced nicotinamide adenine dinucleotide phosphate and adenosine triphosphate (ATP), are important energy sources for tumor cells under hypoxic conditions (4).
In the past few years, lipids have been shown not only as an alternative energy source to bridge energy deficits but also as essential components in the synthesis of biomembranes, substrates for biomass and as activators of highly complex signaling pathways regulating cancer cell growth and migration (5). Such characteristics have been described in several types of cancer such as breast, colorectal and ovarian cancer and are highly correlated with a poor prognosis (6). In addition, lipid metabolism also influences the function of immune cells within the TME, which interact with cancer cells to form a complex network of metabolic crosstalk that critically regulate anti-tumor immunity and ultimately determine tumor progression. There are considerable clinical gaps in translating the understanding of these mechanistic insights into therapies. One of the main challenges is the heterogeneity of lipid metabolic dependencies between patients and cancer types, which limits the identification of universal therapies. The other major challenge is that the regulation between lipid metabolism and function of immune cells is a poorly studied mechanism for the development of new therapies, especially for the overcoming resistance to conventional chemotherapy and immunotherapy. In this review, we perform an in-depth discussion of the mechanistic interplay between lipid metabolic reprogramming in cancer cells and the immune microenvironment of the TME. We critically discuss how the modulation of these highly complicated interconnected pathways represents a promising therapeutic strategy to overcome treatment resistance and improve cancer outcomes. By discussing the interplay between cancer cells and the TME, we fill the existing clinical gaps and uncover unique mechanistic insights that provide novel research directions for cancer immunotherapy. We provide a solid theoretical basis for developing personalized metabolic therapies by determining key metabolic nodes that can be exploited based on the specific tumor and immune context. These new findings reveal the translational potential of targeting lipid metabolism to overcome therapeutic resistance and improve prognostic outcomes in cancer patients. Thus, providing a new perspective for clinical application.
2 Lipid metabolism
Lipid metabolism encompasses the processes of lipid synthesis, catabolism, transport, and regulation (7). The regulation of lipid metabolism maintains cellular homeostasis. In the interest of understanding the metabolic communications of the TME, we summarize the most salient alterations of lipid metabolism in cancer cells (Figure 1).
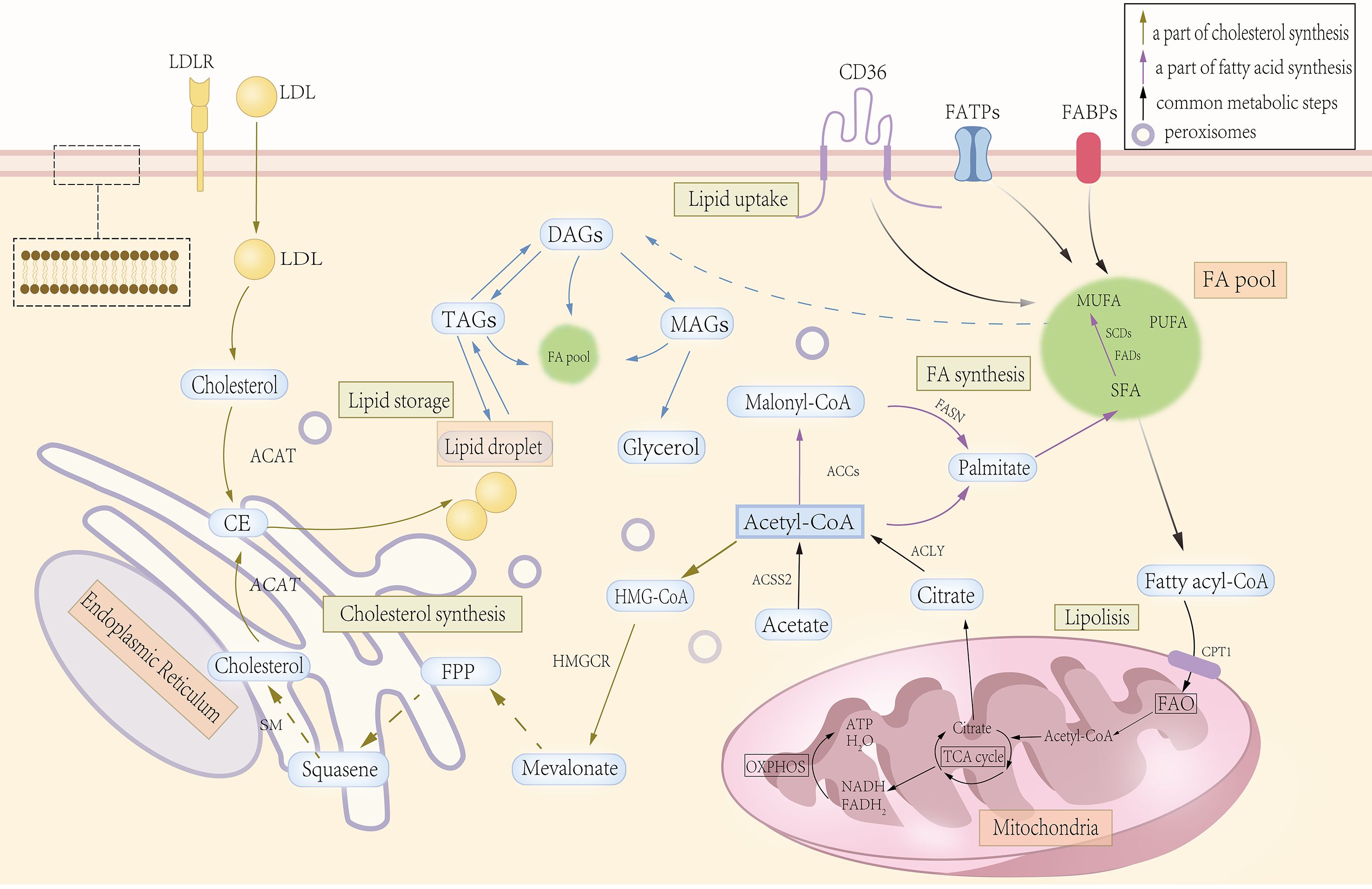
Figure 1. Significant steps in lipid metabolism. Lipid absorption is mainly accomplished through specific channels and transport mechanisms in the intestinal epithelial cells. Lipid synthesis begins with acetyl-CoA, which gradually extends the carbon chain through the fatty acid synthase complex to form fatty acids, which are then esterified with glycerol-3-phosphate to form triglycerides or phospholipids, while cholesterol is synthesized from acetyl-CoA via the mevalonate pathway. Lipid catabolism and oxidation begin with the hydrolysis of triglycerides in adipose tissue to glycerol and free fatty acids by lipase, and fatty acids enter the cell and are catabolized in the mitochondria through β-oxidation to acetyl-CoA, which enters the tricarboxylic acid cycle for complete oxidative energy supply. Inside the cell, lipids (e.g., triglycerides and cholesterol esters) are synthesized on the endoplasmic reticulum (ER). As lipids continue to accumulate, they eventually form separate lipid droplets, which are used for energy storage as well as to regulate lipid metabolism and signal transduction.
2.1 Fatty acids in cancer
2.1.1 Uptake
Cancer cells increase the uptake of exogenous lipids by upregulating the expression of FA transporters, such as CD36, and FABPs (FA-binding proteins) (8). High CD36 expression has been correlated with poor prognosis in breast, ovarian, gastric, and prostate cancer patients (9). Interestingly, different FABP isoforms have opposing functions: A-FABP drives the growth of breast cancer, while E-FABP enhances anti-tumor immunity (10, 11). However, the strong functional antagonism between pro-tumorigenic A-FABP and anti-tumorigenic E-FABP poses a major obstacle in the translational application of these two isoforms, calling for highly specific targeting strategies. The first challenge that the TME poses to metabolic oncology is also the biggest, namely, how does the TME decide which FABP isoform wins the battle? Future studies should focus on identifying the upstream signals and downstream effectors that confer this context-dependency to avoid further immunosuppressive or tumor-promoting effects from the TME. This challenge also offers a promising opportunity in metabolic oncology.
2.1.2 Synthesis
De novo lipogenesis is the most important source of fatty acids (FAs). Studies indicate that approximately 95% of FAs are synthesized endogenously by cancer cells despite the availability of extracellular FAs. Its principal substrate, acetyl-CoA, is produced from citrate by ATP citrate lyase (ACLY) or from acetate by acetyl-CoA synthetase (ACSS2).
2.1.2.1 Acetyl-CoA-producing enzymes: ACLY and ACSS
Expression or activity of ACLY is associated with tumor progression in glioblastoma, colorectal, breast, lung, and hepatocellular carcinomas and promotes cancer cell proliferation and stemness. Consequently, ACLY has emerged as a promising therapeutic target. However, ACLY inhibition induced immunosuppression in immunocompetent mice by promoting the production of immunosuppressive polyunsaturated fatty acid peroxidation products, activating the cGAS-STING pathway, and upregulating PD-L1, which drives immune evasion and metastasis (12). In human cancers, reduced ACLY expression correlates with cGAS-STING activation and decreased T-cell infiltration. Therefore, since ACLY inhibition may carry an immunosuppressive effect, ACLY inhibitor monotherapy for cancers in preclinical models and clinical trials needs careful consideration and evaluation (12). This example clearly demonstrates that one cannot think of attacking a key metabolic enzyme in isolation of its dramatic impact on the immune landscape and renders it a “combination” approach from the beginning. Obviously, the anti-PD-1/PD-L1 combination therapy could have blocked this immunosuppressive signal and enabled the activated T cells to mediate their cytotoxic activities and thus, overcame the immunosuppression emerged from ACLY inhibitor treatment to obtain a synergistic antitumor effect. This suggests that inhibition of ACLY with immune checkpoint blockage may be a sustained anticancer strategy.
Acetyl-CoA synthetase (ACSS): ACSS1 and ACSS3 are expressed in mitochondria, whereas ACSS2 is expressed in the cytoplasm and nucleus (13). Previous studies have concentrated on ACSS2, which is upregulated in cancer cells under stress conditions, such as nutrient deprivation and hypoxia. Depletion/reduction of ACSS2 has been reported to impair the growth of multiple cancer types, including breast, prostate, liver and glioblastomas (13–15). This highlights the critical importance of ACSS2 in tumor growth and thus justifies its use as an anticancer agent. In addition, ACSS2 is involved in promoting the use of acetic acid for lipid synthesis and the growth of tumors (14, 16). ACSS2 deletion also protects cells and mice from fibrosis development, suggesting its potential as a therapeutic target for renal diseases (17, 18). Knockdown of ACSS1 reduces hepatocellular carcinoma cell viability and suppresses melanoma tumor growth (18–20). ACSS3 inhibits prostate cancer progression by increasing endoplasmic reticulum stress-mediated apoptosis through downregulation of the lipid droplet-associated protein PLIN3 and reverses drug resistance by reducing lipid droplet deposition in tumors (21).
2.1.2.2 FA biosynthetic enzymes: ACC, FASN, and SCD
Acetyl-CoA carboxylase (ACC): Mammalian ACC has two tissue-specific isoforms: ACC1 and ACC2. ACC1 is highly expressed in various human cancers, including breast, gastric, liver, and prostate cancers, correlating with reduced patient survival (22, 23). ACC2 is highly expressed in laryngeal cancer, and its expression levels are positively correlated with advanced clinical stages and reduced 5-year survival (24). Under nutrient-rich conditions, prolyl hydroxylase domain protein 3 hydroxylates and activates ACC2, inhibiting FAO. In acute myeloid leukemia (AML), reduced prolyl hydroxylase domain protein 3 expression promotes FAO, driving AML cell proliferation and disease progression (25).
Fatty acid synthase (FASN): FASN expression is upregulated in early-stage lung, prostate, and breast cancers (26–28), with further increases observed as cancer progresses (29, 30). This indicates that cancer cells express enhanced de novo lipid synthesis at an early stage of tumorigenesis and is associated with cancer recurrence and poor survival (31). Inhibition of FASN can decrease FA synthesis, increase the accumulation of malonyl-CoA and inhibit CPT1-mediated FAO, induce cell cycle arrest and apoptosis (31, 32). Moreover, FASN inhibitors can block palmitoylation and plasma membrane- and mitochondria-associated EGFR activation and enhance cancer cell sensitivity to EGFR inhibitors by promoting EGFR ubiquitination, which further results in attenuating tumor growth (33, 34).
Stearoyl-CoA desaturase (SCD): SCD catalyzes the conversion of saturated fatty acids (SFAs) to monounsaturated fatty acids (MUFAs) and converts stearoyl-CoA (C18:0) and palmitoyl-CoA (C16:0) to oleoyl-CoA (C18:1) and palmitoyl-CoA (C16:1), respectively, with the introduction of double bonds, which increase the level of FAs unsaturation and serve as an important energy source for tumor cells. SCD is highly expressed in several human cancers, such as lung, breast, colorectal, renal, prostate and hepatocellular carcinomas (35–37), and is associated with poor survival (38). Downregulating SCD expression can induce cell cycle arrest and apoptosis and inhibit the proliferation of lung cancer cells (39). SCD inhibitors are particularly effective in blocking cancer cell proliferation, as their effects are linked to AMPK activation, which phosphorylates ACC and inhibits FA synthesis (40). This prevents excessive saturated FA accumulation while blocking desaturation.
2.1.3 Catabolism and FAO
It has been demonstrated that lipolysis plays a critical role in lipid-mediated signaling processes. Lipids released from low-density lipoprotein (LDL), such as certain FAs, are not only vital sources of energy but also signaling molecules, directly regulate cellular signaling pathways and gene transcription. For example, FAs or FA derivatives can bind to and activate members of the nuclear receptor family of transcription factors, such as peroxisome proliferator-activated receptors (PPARs), which regulate gene expression and influence lipid metabolism and inflammatory genes (41).
FAO plays a critical role in cell proliferation and is closely associated with ATP production and nicotinamide adenine dinucleotide homeostasis, both of which are essential for mitochondrial function and cell survival (Figure 2). High FAO activity has been reported in several cancers, such as triple-negative breast cancer and gliomas (4, 42). I In glioblastoma cells, the inhibition of FAO leads to a decrease in the levels of NADPH and an increase in ROS levels, which promote apoptosis (43). However, FAO has dual functions: it maintains cell survival under stress conditions but promotes cytotoxicity when excessive. This suggests the need for strict regulation of cancer cells and highlights an attractive target for cancer therapy because strict regulation rather than complete inhibition of FAO is required. Thus, maintaining a balance between the production of ATP and nicotinamide adenine dinucleotide phosphate through FAO inhibition prevents lipotoxicity, and dynamic regulation of FAO can inhibit the proliferation and survival of tumor cells. Consistently, key enzymes and transporters involved in FAO are often upregulated in various human cancers. In breast cancer, mammary adipocyte-derived leptin upregulates STAT3 and induces the expression of CPT1B and enhances FAO activity in cancer stem cells, promoting stemness and chemotherapeutic resistance (44). The transcriptional regulators, such as PPARs and AMP-activated protein kinase, can activate the expression of transcriptional regulators to promote FAO (45).
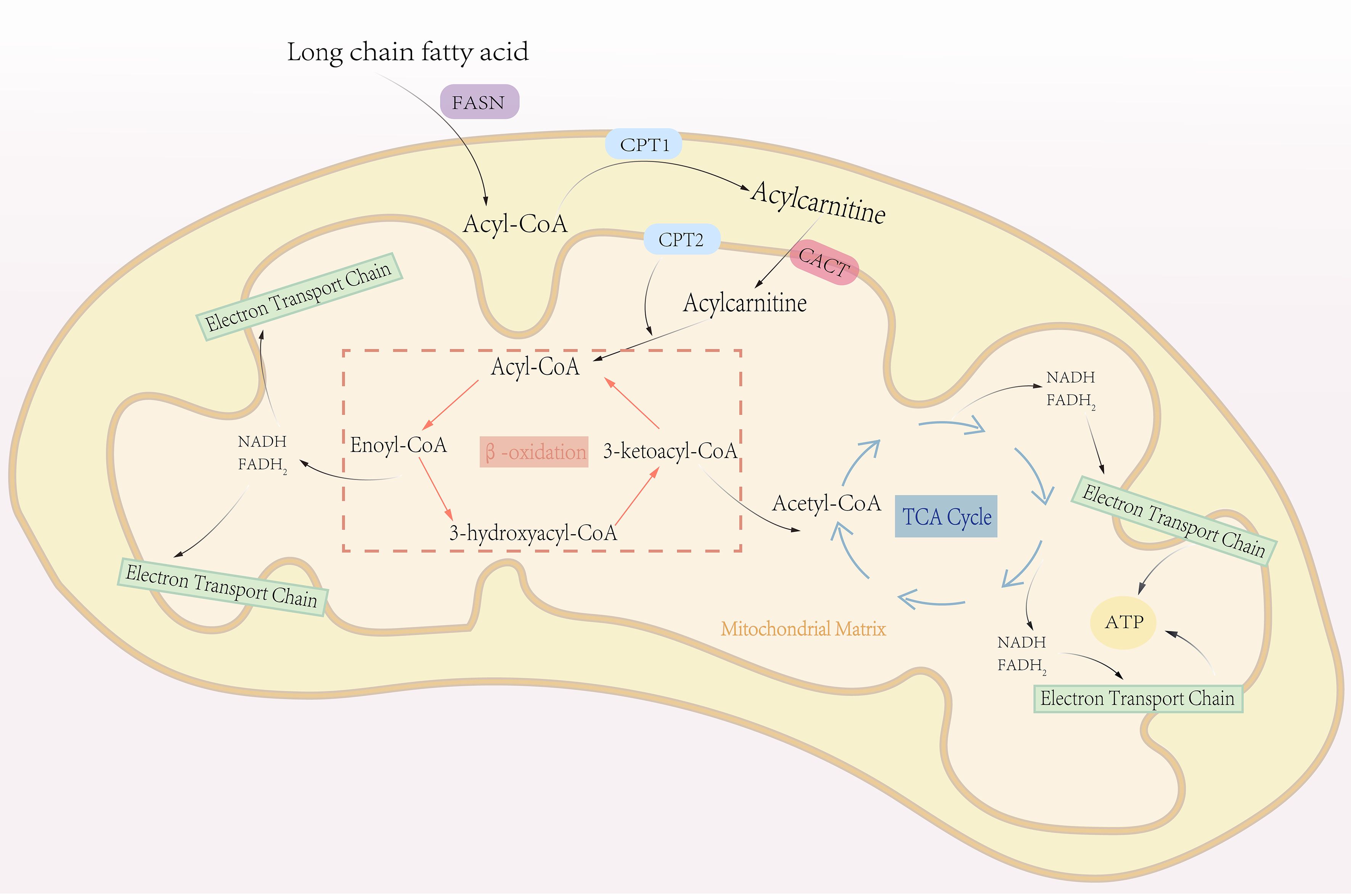
Figure 2. The process of fatty acid oxidation. Fatty acid oxidation, also known as β-oxidation, is a central metabolic pathway that breaks down fatty acids to generate energy. This process primarily occurs in the mitochondrial matrix. Triglycerides stored in adipose tissue are hydrolyzed into glycerol and free fatty acids, which are then activated by combining with CoA to form fatty acyl-CoA. Fatty acyl-CoA is transported into the mitochondrial matrix via the carnitine shuttle system. Within the mitochondrial matrix, fatty acyl-CoA undergoes β-oxidation, a cyclic process that sequentially removes two-carbon units in the form of acetyl-CoA. Each cycle of β-oxidation generates one molecule of NADH and one molecule of FADH2. The acetyl-CoA produced enters the TCA cycle, where it is fully oxidized to produce ATP. Simultaneously, NADH and FADH2 donate electrons to the electron transport chain, driving oxidative phosphorylation to generate additional ATP. This process repeats iteratively until the fatty acid is completely broken down into acetyl-CoA.
2.1.4 Storage
Lipid droplets (LDs) are important organelles involved in lipid and energy storage. It is well accepted that LDs are derived from endoplasmic reticulum (46). LDs are involved in energy storage and serve as the main source of precursors for cell membrane phospholipids and cholesterol (47). LDs have a dynamic life cycle in cells and their size, intracellular location, and lipid and protein composition can change rapidly in response to various environmental conditions and cell states (48). Enzymes and proteins involved in lipid metabolism localize to the surface of LDs and regulate lipid homeostasis by maintaining a balance between lipid storage and catabolism (49). LDs also promote lipid catabolism through the action of neutral lipases, such as ATGL and HSL. These enzymes mediate the breakdown of triglycerides to release large amounts of free FAs that can be transported to mitochondria for oxidation to produce ATP to meet cell energy demands (50, 51).
2.2 Cholesterol in cancer
Cholesterol participates in the composition of biological membranes, maintaining the fluidity of lipid bilayers, forming lipid rafts, and solubilizing other lipids to coordinate the activation of many signal transduction pathways Altered cholesterol metabolism facilitates the proliferation of tumor cells and induces the apoptosis of tumor cells by regulating the oncogenic pathway PI3K/AKT/mTOR, and it induces CD8+ T cell exhaustion to create an immunosuppressive TME (52–54). Cholesterol is regulated by two pathways: dietary intake and de novo synthesis through the mevalonate pathway. Transcription factors involved in the regulation of these pathways include SREBPs and LXRs.
2.2.1 Uptake
Cancer cells have high demands for cholesterol and enhance uptake to meet these demands by overexpressing LDLR. This has been demonstrated in hepatocellular, breast, and prostate cancers, and high expression of LDLR correlates with poor prognosis (55–57). Deletion of LDLR prevents the growth of tumors in HER2+ breast cancer and PDAC models hyperlipidemic mice (58). In prostate cancer cells, deletion of PTEN leads to the activation of PI3K/AKT, and this greatly enhances the uptake of exogenous LDL; this process is essential for the growth of tumors (59). Abnormalities in lipid metabolism can cause lipotoxicity, inducing oxidative stress and causing a large increase in ROS. Progressive oxidative stress can induce the oxidative modification of intracellular LDL to ox-LDL. Ox-LDL can bind to scavenger receptors, such as LOX-1, an oxidized LDL receptor-1, and CD36 (60), causing mutations that induce inflammation, cell proliferation, and metastasis. Based on the experimental evidence above, empirical studies have demonstrated the pro-tumorigenic effects of these molecules. Zettler et al. reported that ox-LDL upregulates cyclin expression, which leads to an increase in cell proliferation (61). Similarly, another study reported that treating ovarian cancer cells with ox-LDL enhances their proliferative capacity (62). Furthermore, oxidative stress can accelerate DNA damage in cancer cells, which leads to the malignant transformation of cells and carcinogenesis (63).
Due to the complexity of the mechanisms underlying the role of LDL and its oxidized forms in cancer, little is known about how LDL, through its receptors, regulates the reprogramming of lipid metabolism in cancer cells, and how it activates signaling pathways within cancer cells through their receptors, leading to oxidative stress and inflammatory responses, and how it regulates the function of immune cells, which is important for exploring its potential as a target for inducing oxidative stress and inflammatory responses, and for developing novel therapeutic interventions or combined treatment regimens, which is crucial for fully exploring its potential as a target.
2.2.2 Synthesis
2.2.2.1 Cholesterol biosynthetic enzymes: HMGCR and SM
3-Hydroxy-3-methylglutaryl-CoA reductase (HMGCR). HMGCR is the rate-limiting enzyme in cholesterol biosynthesis, overexpressed in gastric, glioblastoma and prostate cancers, and its upregulation facilitates cancer cell growth and migration, while its knockdown attenuates tumorigenesis. Therefore, HMGCR inhibition may provide a promising therapeutic approach for solid tumors, hematologic malignancies and drug-resistant cancers (64–66). However, the risk reduction associated with the two polymorphisms might be affected by confounding factors or biases inherent to observational studies, which limit the possibility of establishing a causal relationship. Therefore, further mechanistic studies and prospective clinical trials are required to confirm whether HMGCR inhibition per se confers a direct anti-cancer effect or is only associated with a lower underlying risk.
Squalene monooxygenase (SM). SM is the second rate-limiting enzyme in cholesterol biosynthesis, located downstream of HMGCR, and is also regulated by SREBP2 (67). Its stability is regulated by its substrate and lipid interactions: squalene prevents its ubiquitination and subsequent degradation by MARCH6, while unsaturated FAs inhibit its degradation (68, 69). Defective SM expression was observed in anaplastic lymphoma kinase (ALK)-positive anaplastic large-cell lymphoma cells, leading to cholesterol dysregulation and dependence of tumor growth on LDL receptor (LDLR)-mediated cholesterol uptake (70).
2.2.3 Efflux and storage
ATP-binding cassette (ABC) transporters, including ABCA1 and ABCG1, mediate cholesterol efflux to maintain homeostasis. In tumor-associated macrophages (TAMs), ABCA1/ABCG1-driven efflux potentiates IL-4/STAT6 signaling, enforcing an M2-like, pro-tumorigenic phenotype. Genetic ablation of these transporters reverses TAM-mediated immunosuppression and impairs angiogenesis (71, 72). This positions cholesterol efflux inhibition as a promising strategy to reprogram TAMs. LXRs and their downstream target genes, such as the ABC family, enzymes involved in lipid metabolism, and extracellular lipid receptors, regulate cholesterol transport and downregulate genes related to lipid uptake and synthesis. These mechanisms may contribute to the formation of an immunosuppressive TME. LXRs promote lipogenesis and the transcription of ABCA1 and ABCG1 (73). To prevent cytotoxicity from excessive free cholesterol accumulation, acyl-CoA cholesterol acyltransferase (ACAT) converts cholesterol into cholesteryl esters, which are stored in LDs (74). Targeting this storage mechanism may offer another avenue to disrupt cholesterol homeostasis in cancer cells.
3 Crosstalk between lipid metabolism and tumor immune response in the TME
Despite the harsh conditions of the TME established by tumor cells, immune cells have adapted by utilizing lipids as fuel to support anti-tumor immune responses. The major immune cells involved in these metabolic cross-talks are T cells, tumor-associated macrophages, dendritic cells, neutrophils, natural killer cells and myeloid derived suppressor cells. The response of immune cells to lipid metabolic changes is not uniformly induced but depends on the involvement of a multitude of factors. Factors such as the specific lipid species, type of immune cell and the dynamic TME can determine whether a pro- or anti-tumor response is elicited. As a result, the interpretations drawn from these findings are often context-dependent and appear contradictory (Figure 3). This inherent contextual dependency poses a major challenge to translate these findings into effective therapies and calls for the development of individualized context-dependent strategies targeting lipid metabolism in immunotherapy. In this section, we will critically review how distinct lipid metabolic programs not only nourish but also instruct the polarized functional maturation of various immune cells, generating a metabolic landscape that can either resists or responds to immunotherapy. The inherent context-dependency of these interactions is the main challenge of translating these findings into effective universal therapies.
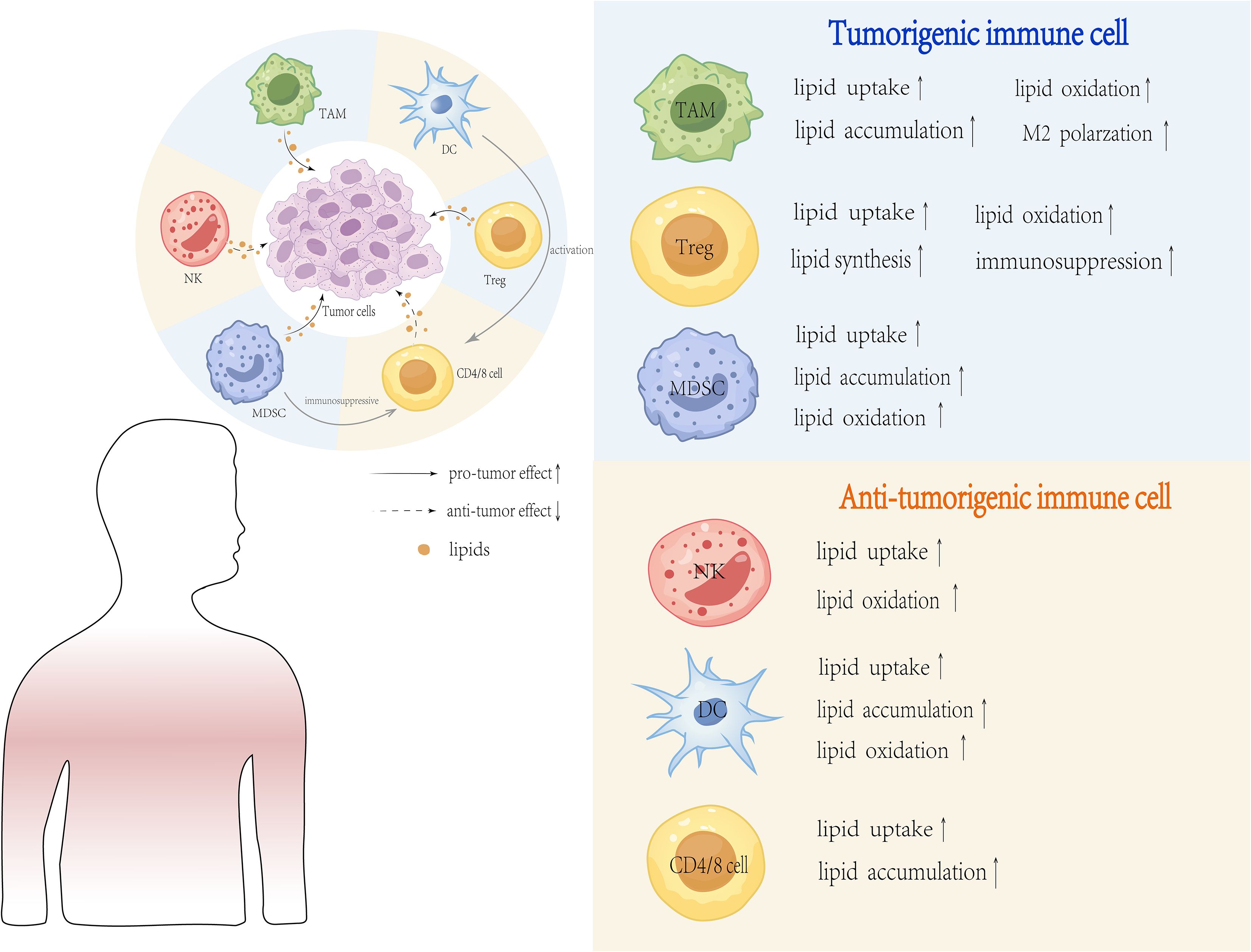
Figure 3. Major Lipid Metabolic Changes in the Tumor Microenvironment. Lipid metabolic changes in the TME are a hallmark of tumor cells adapting to rapid proliferation and survival. These changes include increased lipid uptake and synthesis, lipid storage and utilization, enhanced lipid oxidation, reprogramming of lipid metabolism, lipid-mediated signaling and lipid metabolism in tumor-associated immune cells. These metabolic adaptations enable tumor cells and immune cells to thrive in the harsh TME, facilitating tumor progression and immune evasion.
3.1 T cells in tumor immunity
T cells are an essential component of the adaptive immune response that provides protection against pathogens and tumors. Naïve T cells can proliferate and differentiate into effector cells upon antigen and co-stimulatory signals as well as cytokines, which are involved in the process of tumor development and progression. CD8+ T cells are believed to be the effector cells of adaptive anti-tumor immunity that can differentiate into cytotoxic T lymphocytes (CTLs) and eliminate tumor cells in an MHC-dependent manner (75). CD4+ T cells include pro-inflammatory T-helper 1 (Th1) cells, immunosuppressive T-helper 2 (Th2) cells, T-helper 17 (Th17) cells and regulatory T-cells (Tregs) that have pro- or anti-inflammatory effects on tumors (76).
Immune signaling pathways, such as phosphoinositide 3-kinase (PI3K)-Akt and mechanistic target of rapamycin (mTOR), regulate T cell lipid metabolism. Elevated levels of cholesterol and FAs in the TME can alter T cell function. T cell receptor (TCR) stimulation activates PI3K-Akt and mTOR signaling, inducing FA and mevalonate synthesis (77, 78). Inhibition of ACC1 reduces Th17 cell differentiation (79) while enhancing memory CD4+ T cell formation (80). ACC1 is also essential for antigen-specific CD8+ T cell accumulation during bacterial infection and Treg suppressive function in the TME (81).
CD36-mediated uptake of lipids, such as long-chain fatty acids and oxidized low-density lipoproteins (ox-LDL), promotes Treg function but can lead to lipid peroxidation, p38-mitogen-activated protein kinase (MAPK) activation, or ferroptosis in CD8+ T cells, impairing their function (82–84). Increased FAs in the TME can activate CD8+ T cells, promoting lipid metabolism and maintaining effector function through peroxisome proliferator-activated receptor alpha (PPARα) signaling (82). Furthermore, enhancing FA catabolism in tumor-infiltrating CD8+ T cells boosts their anti-tumor activity. Lipid depletion in CD8+ T cells significantly impairs their proliferation and cytotoxicity, leading to functional exhaustion characterized by reduced interferon (IFN)-γ production and increased programmed cell death protein-1 (PD-1) expression (85, 86).
Cholesterol and its biosynthetic intermediates also have a profound impact on T cell function in the TME. Cholesterol induces CD8+ T cell exhaustion by inducing endoplasmic reticulum stress and promoting uncontrolled tumor growth (87). LDL, rich in cholesterol, is uptaken by LDL receptor (LDLR). LDL depletion or LDLR deficiency impair CD8+ T cell function and anti-tumor activity (88). These observations provide strong rationale for targeting lipid pathways in T cells.
3.2 Tumor-associated macrophages
TAMs are one of the most highly infiltrating immune cells in the TME. TAMs interact with cancer cells and promote tumor growth and progression. Reprogramming of lipid metabolism is a hallmark of TAMs, and lipid metabolites secreted by TAMs promote or inhibit tumor growth. The initial steps of this reprogramming are initiated by a combination of extrinsic cues. These include soluble factors in the TME, such as cytokines (IL-4, IL-13, M-CSF), hypoxia, and uptake of exosomes from tumors. These different signals converge to activate some key transcription factors, including PPARγ and STAT6, and further induce transcriptional upregulation of genes involved in lipid uptake and de novo lipogenesis.
TAMs can be further polarized into classically activated (M1) macrophages and alternatively activated (M2) macrophages. M1 macrophages participate in anti-tumor immune responses by secreting pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), and inducible nitric oxide synthase (iNOS). In contrast, M2 macrophages are characterized by an anti-inflammatory phenotype and secrete pro-tumor factors, such as arginase 1 (ARG1) (89). Peroxisome proliferator-activated receptors (PPARs) are involved in almost all aspects of FAs metabolism (37) and have been shown to regulate TAM polarization toward the M2 phenotype through multiple signaling pathways, promoting tumor cell proliferation, angiogenesis, and immunosuppression (90, 91). Both PPARγ agonists and inhibitors exhibit anti-tumor activities. PPARγ inhibitors attenuate the pro-inflammatory effects of M1 TAMs and suppress the secretion of pro-tumorigenic cytokines by M2 macrophages, further inhibiting tumor progression (92, 93).
The phenotype of TAMs, expressed as pro- or anti-tumorigenic functions, is directly determined by their underlying lipid metabolic state. Upregulation of critical lipid metabolism genes in TAMs by tumor cells induce lipid accumulation and M2 polarization, which further promote tumor progression.
Reprogramming of cholesterol metabolism regulates TAM activity and recruitment by inducing M2 polarization and tumor progression. Currently, research on cholesterol metabolism in TAMs is focused on the cholesterol efflux pathway. For example, ABCA1/ABCG1-mediated cholesterol efflux is an active driver of M2-like polarization through its enhancement of the IL-4/STAT6 signaling (72). Targeting this pathway to reprogram TAMs appears very attractive; however, this strategy raises a serious safety concern. Systemic inhibition of the reverse cholesterol transport may induce atherosclerosis at the expense of an on-target, off-tumor toxicity. Therefore, safer alternatives are needed for future efforts. We believe future efforts should focus on developing TAM-specific delivery systems or targeting context-specific lipid dependencies (94). Lipid metabolism in TAMs is associated with immunosuppression and chemotherapy resistance. For example, in a gastric cancer mouse model, lipid accumulation in TAMs upregulates the expression of phosphatidylinositol 3-kinase gamma (PI3K-γ), promoting M2 polarization. This decreases phagocytosis and upregulates programmed death ligand 1 (PD-L1) expression in TAMs, attenuating the anti-tumor T-cell response and enhancing immunosuppression (95). TAMs show high plasticity, and targeting lipid metabolism to induce M1 polarization may improve the efficacy of tumor immunotherapy and reduce chemoresistance (96). Considering the important role of lipid metabolism in TAMs, several studies have focused on targeting lipid metabolism in TAMs or inducing their conversion into anti-tumor phenotypes. However, lipid metabolism in TAMs has not been systematically investigated, and little is known about how its initial events are initiated and how it exerts dual roles in cancer. Therefore, further investigation is required to elucidate the mechanisms underlying lipid metabolism in TAMs and to provide a theoretical basis for developing effective therapeutic strategies.
3.3 Dendritic cells
DCs and NK cells are critically important for successful anti-tumor immune responses. However, multiple immunosuppressive factors can exist in the TME to impair their function and promote a pro-tumorigenic phenotype (97, 98). DCs are innate immune cells that can be grouped into conventional DCs (cDCs), plasmacytoid DCs (pDCs), and monocyte-derived DCs (moDCs) depending on their respective developmental pathways (99). The main role of DCs is to capture and process antigen, present them to the T lymphocytes, and thus, induce adaptive immune responses. Upon maturation and stimulation of DCs, naïve CD4+ and CD8+ T cells differentiate into different types of effector T cells with distinct functions (100, 101). In addition, DCs can activate other immune cells of the innate immune system, such as NK cells and macrophages (102), DCs connect the innate and adaptive arms of the immune system and are essential for the induction of immune responses.
Recent reports have implicated lipid metabolism as an essential regulator of DC development, maturation, and function (103). Excessive lipid accumulation in DCs is a major cause of DC dysfunction. In a clinical study on lung cancer, Arai et al. observed that the fluorescence intensity of lipid accumulation in DCs was significantly higher in patients with stage III/IV lung cancer compared with the controls. The accumulated lipids were mainly TGs, and this dysfunction impaired DC function, contributing to tumor immune escape (104, 105). In a mouse model of ovarian cancer, up-regulation of FASN expression in tumor cells resulted in excessive lipid accumulation, which inhibited the activation of T cells infiltrating the tumor by DCs (106). Kratchmarov et al. found that the inhibition of FAO with the CPT1 inhibitor etomoxir did not affect the overall development of cDCs or pDCs, although it significantly modified their subset frequencies, with an increase in cDC2 and a decrease in cDC1 (107, 108). These findings suggest that the subsets of DCs is determined by immune signals and microenvironment, and metabolic reprogramming allows them to function in different microenvironments.
Interestingly, FAO, regulated by signaling through peroxisome proliferator-activated receptors (PPARs), AMP-activated protein kinase (AMPK), mechanistic target of rapamycin (mTOR), signal transducer and activator of transcription 3 (STAT3), and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), is critical for DC differentiation. For example, PPARs, key transcription factors regulating FAO, are significantly upregulated in granulocyte-macrophage colony-stimulating factor (GM-CSF)- and interleukin-4 (IL-4)-induced moDCs and are essential for their generation (109, 110). AMPK phosphorylation upregulates FAO, inducing an immunosuppressive phenotype in tolerant DCs (111), and the activation of the JAK2-STAT3 signaling pathway has been shown to inhibit dendritic cell (DC) maturation (112). These mechanisms highlight the intricate relationship between lipid metabolism and DC function within the TME. Future investigations should therefore prioritize elucidating how the PPAR, AMPK, mTOR, STAT3, and PGC-1α signaling networks dynamically interact to regulate FAO, thereby orchestrating the metabolic reprogramming that enables distinct DC subsets to adapt and function within the diverse and challenging niches of the TME.
3.4 Natural killer cells
NK cells are cytotoxic lymphocytes of the innate immune system, capable of killing tumor cells and virus-infected cells. They exhibit direct effector functions against cellular targets and play a role in generating, forming, and maintaining multicellular immune responses. Glucose metabolism is critical for key NK cell functions, including cytokine production, receptor-mediated activation, ligand expression, degranulation, and the regulation of glycolysis, oxidative phosphorylation (OXPHOS), and adenosine triphosphate (ATP) production (113, 114).
Although NK cells are major players in the innate elimination of viral, bacterial, and tumor cells, research on lipid metabolism in NK cells and tumor-associated NK (TANK) cells remains limited. Nevertheless, the importance and clinical relevance of the crosstalk between lipid metabolism, NK/TANK cells, and tumors have been clearly demonstrated. A growing body of evidence suggests that the TME produces soluble modulators that inhibit NK cell maturation, proliferation, and effector functions. These immunosuppressive factors may directly affect NK cells or indirectly influence them by inducing the production of additional immunosuppressive molecules by other immune cells, such as regulatory T cells (Tregs), antigen-presenting cells (APCs), and myeloid-derived suppressor cells (MDSCs). Metabolism is a critical component of NK cell immunobiology and a key determinant of their response to immunotherapy in tumors.
Therefore, in the future, we should focus on exploring the specific mechanisms by which lipid metabolism regulates NK cell function dynamically and the effects of lipid metabolic heterogeneity in the TME on NK cell function. That is, we should use patient samples from clinical trials for multi-omics profiling in subsequent studies. By integrating single-cell transcriptomics and lipidomics with multi-omics technologies, we can delineate the lipid metabolic signatures underlying NK cell dysfunction and identify potential targets for therapy. Using clinical trials and multi-omics technologies will be crucial for the further development of NK cell-based therapies for cancer with improved therapeutic effect and translational value.
3.5 Tumor-associated neutrophils
Neutrophils are differentiated from hematopoietic stem cells in the bone marrow and further mature in the bloodstream. In addition to being fighters against pathogenic infections, neutrophils also serve as coordinators that regulate, target, and activate inflammatory and adaptive immune responses. Neutrophils can express various cytokines and immunosuppressive molecules and also interact with other effector cells, such as B cells, DCs, macrophages, and T cells. Because of their functional plasticity, neutrophils can either kill tumor cells or promote tumor growth. Lipid metabolism regulates the immune function of neutrophils by affecting their activation, migration and effector functions.
In 2009, Friedlander et al. first proposed the classification of neutrophils into anti-tumor (N1) and pro-tumor (N2) subsets (115). Neutrophil activation and function depend on FAs utilization and oxidation. Free FAs enter the FAO pathway and undergo oxidative phosphorylation (OXPHOS) to produce adenosine triphosphate (ATP), which supports neutrophil differentiation, maturation, and function. In the early stages of tumorigenesis, TANs are predominantly of the N1 phenotype, exerting anti-tumor functions through the activation of interleukin-18 (IL-18) and the secretion of interferon-beta (IFN-β). However, as tumors progress, TANs often shift to the N2 phenotype. N2-like TANs promote tumor progression by regulating cancer cell proliferation, angiogenesis, and metastasis. They also inhibit T cell responses and disrupt T cell function and polarization by generating reactive oxygen species (ROS) and overexpressing programmed death ligand 1 (PD-L1) and arginase-1 (116, 117).
Peroxisome proliferator-activated receptor gamma (PPARγ) is a key transcription factor that regulates adipogenesis and stimulates lipid droplet formation. PPARγ ligands not only promote adipogenesis but also induce neutrophil differentiation (118). Similarly, during the differentiation of hematopoietic progenitor cells into neutrophils in the bone marrow, all-trans retinoic acid (ATRA) and granulocyte colony-stimulating factor (G-CSF) stimulate the accumulation of lipid droplets (LDs) in neutrophils via PPARγ (119). Emerging evidence highlights the critical role of neutrophils in tissue damage and repair processes, mediated by lipid metabolism-driven inflammatory responses in chronic inflammatory diseases (120). Consequently, therapeutic strategies aimed at modulating neutrophil lipid metabolism—such as targeting key metabolic pathways or developing precision-based pharmacological agents—hold significant promise. These strategies not only can improve the neutrophil immune functions, but also provide new approaches for cancer and inflammatory diseases therapy.
3.6 Myeloid-derived suppressor cells
MDSCs are generated from immature myeloid cells and can be divided into two subsets: polymorphonuclear MDSCs (PMN-MDSCs) and monocytic MDSCs (M-MDSCs). MDSCs are closely involved in tumor progression, metastasis, and therapy resistance via the promotion of Treg proliferation and the secretion of immunosuppressive molecules, including arginase 1 (Arg1), inducible nitric oxide synthase (iNOS), reactive oxygen species (ROS), and prostaglandin E2 (PGE2). Tumor-derived exosomes can exert their onco-suppressive effect by activating and expanding MDSCs through the transfer of microRNAs (miRNAs), interleukins (ILs), transforming growth factor-beta (TGF-β), and PGE2 (121, 122).
Tumor-derived exosomes can exert their onco-suppressive effect by activating and expanding MDSCs through the transfer of microRNAs (miRNAs), interleukins (ILs), transforming growth factor-beta (TGF-β), and PGE2 (121, 122).
Fatty acid transporter protein 2 (FATP2) is responsible for lipid accumulation in PMN-MDSCs. The lipid content is essential for the expression of pro-inflammatory genes and suppression of CD8+ T cells. Pharmacological inhibition of FATP2 can attenuate lipid accumulation, ROS production, and immunosuppressive activity, and further inhibit tumor growth (123). Tumor-infiltrating PMN-MDSCs increase FAs uptake and activate FAO. Inhibition of FAO blocks their immunosuppressive pathways and reduces the production of inhibitory cytokine (124). The serine/threonine kinase PIM1 regulates lipid oxidation through peroxisome proliferator-activated receptor gamma (PPARγ)-mediated activity. Enhanced PPARγ expression rescues the metabolic and functional defects in Pim1-/- PMN-MDSCs (125), underscoring the critical role of lipid metabolism in MDSC-mediated immunosuppression.
3.7 Cancer-associated fibroblasts
In addition to immune cells, CAFs are a major stromal component of the TME. CAFs exhibit remarkable heterogeneity and plasticity, playing a crucial role in the TME formation, tumor metabolic reprogramming, proliferation, invasion, and immune modulation. In the TME, CAFs are activated by signaling pathways involving transforming growth factor-beta (TGF-β) and lysophosphatidic acid (LPA) (126, 127). They synthesize and secrete lipids and biologically active lipid signaling molecules. When biologically active lipids, such as lysophosphatidylcholine (LPC), are overexpressed in CAFs, they can be released into the TME and taken up by tumor cells, promoting tumor proliferation and migration through intracellular lipid metabolic reprogramming (128, 129).
CAFs also secrete cytokines or engage in direct cell-to-cell interactions, promoting malignant phenotypes in tumor cells, enhancing therapeutic resistance, and modulating the activity of other tumor-infiltrating immune cells (130). In response to the TME, CAFs reprogram their lipid metabolism by upregulating key enzymes such as FASN and SCD, thereby increasing lipid synthesis, storage, and secretion (128, 131).
4 Targeting lipid metabolism in cancer therapy
4.1 Immunotherapy and the challenge of resistance
Chemotherapy and immunotherapy represent the cornerstone treatments for metastatic cancer. While standard chemotherapeutic agents include antibiotics, plant-derived compounds, and platinum-based drugs, the most widely utilized immunotherapeutic agents are immune checkpoint inhibitors, such as monoclonal antibodies targeting PD-1, PD-L1, and CTLA-4. The CTLA-4 pathway primarily plays a role in the initial phase of the activation of the immune response. When T cells are activated in the lymph nodes, CTLA-4 competes with the co-stimulatory molecule CD28 for interaction with its ligands on antigen-presenting cells. Since CTLA-4 delivers an inhibitory signal to the T cell, it can serve to suppress the early activation and proliferation of the T cell (132). Tumor cells can use this mechanism to suppress initial activation of the immune system (133). In contrast, the PD-1 pathway primarily plays a role in the effector phase of the immune response, mainly in peripheral tissues and the TME. Under normal conditions, engagement of PD-1 on normal cells by its ligands on the cell surface represents a protective mechanism, or self-tolerance, that prevents T cell-mediated damage to the host (134–136). Many tumor cells express high levels of PD-L1. When Tumor-infiltrating T cells express PD-1 and encounter PD-L1 on tumor cells, they receive a powerful inhibitory signal and become “exhausted.” An exhausted T cell is characterized by its weakened ability to induce cytotoxicity and its failure to kill tumor cells, which leads to tumor immune escape (134–136).
In contrast, clinically, inhibitors targeting the CTLA-4 pathway and PD-1/PD-L1 inhibitors exert their clinical effects by antagonizing the respective receptor and unleashing T cells from their inhibitory states, which restores anti-tumor immunity. Their clinical benefits in NSCLC have been validated and they have been incorporated into clinical practice (134–136). However, among patients, there is still a high degree of heterogeneity in responses to these therapies. One of the major sources of this heterogeneity is the reprogramming of metabolism in the TME, particularly the dysregulation of lipid metabolism, which has been shown to play a critical role in mediating resistance to immunotherapeutic agents.
4.2 Mechanisms of lipid metabolism-mediated immunosuppression
Reprogrammed lipid metabolism in the TME represents one of the key mechanisms underlying the immune evasion and resistance to checkpoint inhibitor therapy. This immunosuppression involves the following mechanisms: First, metabolic competition. Cancer cells overexpressing FAs transporters, such as CD36, scavenge extracellular lipids extensively, resulting in cancer cell nutrient deprivation and leading to functional exhaustion of T cells (83, 137). Second, active immunosuppression. Notably, some specific lipid mediators are involved in immunosuppression, including oxysterols and PGE2. These substances are generated by tumor cells and immunosuppressive cells, including Tregs and M2-type TAMs. On one hand, they directly impair the cytotoxicity of CD8+ T cells (138, 139). On the other hand, they upregulate inhibitory receptors, such as PD-1 and TIM-3. These two mechanisms collectively induce the exhausted T cell phenotype (140, 141). Therefore, targeting key nodes in lipid metabolism may conversely enhance T cell function and render cancer immunotherapy more sensitive.
Resistance to immunotherapy involves the following mechanisms of tumor cell intrinsic factors (impaired antigen presentation, upregulation of immune checkpoint molecules, hyperactivated oncogenic signaling pathways); TME (infiltration of immunosuppressive cells, dysregulated cytokine networks, metabolic dysfunction); and host systemic factors (gut microbiota composition, host immune status). Notably, lipid metabolic reprogramming in the TME participates in T cell exhaustion while promoting the survival and function of immunosuppressive population, including Tregs (142). Clinical practice also shows that patients with favorable response to immunotherapy exhibit different gut microbial species composition, while antibiotics treatment may result in reduced efficiency of immunotherapy (143).
4.3 Therapeutic strategies targeting lipid metabolism
There are major efforts underway towards the development of therapeutic agents targeting lipid metabolism at multiple nodes; many inhibitors of lipid-metabolizing enzymes are highly promising in large preclinical studies (7, 9, 144). Targeting FA Synthesis. FASN inhibitor TVB-2640 is in phase II clinical trials as monotherapy for non-small cell lung cancer with KRAS mutations (NSCLC, KRAS, NCT03808558), in combination with paclitaxel and trastuzumab for triple-negative breast cancer (NCT03179904), and in combination with the anti-angiogenic drug bevacizumab for high-grade astrocytoma (NCT03032484). Preclinical studies have demonstrated that ACC inhibitors ND-646 and ND-654 significantly inhibit the growth of lung tumors in mice and hepatocellular carcinoma (HCC) in rats, respectively. Additionally, the ACC inhibitor ND-630, initially developed for non-alcoholic steatohepatitis, is undergoing phase I trials for cancer treatment (145). Targeting Cholesterol Synthesis. Statins, a class of cholesterol-lowering drugs that selectively inhibit HMGCR, have gained attention for their anti-tumor properties and are being tested in multiple clinical trials as anticancer agents (146). Statins such as pitavastatin, fluvastatin, and simvastatin have been shown to inhibit tumor cell proliferation and angiogenesis while promoting apoptosis (147–149). Statins suppress cancer cell development by blocking the mevalonate pathway and disrupting the transcriptional responses dependent on Yes-associated protein (YAP) and transcriptional coactivator with PDZ-binding motif (TAZ), key regulators of normal organ growth and tumor progression (54). Additionally, statins modulate lipid rafts by lowering cholesterol levels, influencing tumor development. They can also induce autophagy and ferroptosis and alter the TME to exert anti-tumor effects, although the precise mechanisms remain under investigation (150). To preempt the compensatory surge in exogenous cholesterol uptake triggered by synthesis inhibition, a promising strategy is the concurrent disruption of both pathways. Combining statins with LDLR-targeting agents or efflux inhibitors may thus achieve a more durable suppression of tumor cholesterol metabolism.
4.4 Combination therapy: checkpoint inhibitors and metabolic modulators
The strategic combination of immune checkpoint inhibitors with pharmacological modulators of lipid metabolism represents a promising therapeutic frontier. For instance, investigation of the experimental agent MK1775 in lung adenocarcinoma demonstrated its ability to sensitize tumors to PD-1 blockade by reprogramming FAs metabolism and altering interactions between TAMs and CD8+ T cells within the TME (151). Two independent studies found that statins and PD-1 inhibitors were administered in combination in animal models with a synergistic effect, resulting in about 40% tumor regression and reversal of T cell exhaustion (152). Although such synergy seems promising, there are several major challenges to the clinical implementation of combination strategies. Significant metabolic heterogeneity within and between tumors may result in diverse treatment responses. The safety of combination regimens should be rigorously examined, especially the off-target effects of metabolic inhibitors on immune cells. In addition, efficient targeted delivery and maintaining therapeutically effective drug concentrations in tumor tissues are critical for successful treatment.
4.5 Future directions: integrating advanced technologies
Advanced technologies should be used in the future to confront the above challenges.
Lipidomics is an interdisciplinary field characterized by the systematic identification and characterization of the complete lipid repertoire in living organisms. The main application value of lipidomics technologies in the research of oncology is that they can accurately delineate tumor metabolic heterogeneity and can identify specific lipid profiles associated with resistance to immunotherapy (153). Research on hepatocellular carcinoma found that a low abundance of polyunsaturated fatty acids and high abundance of specific sphingomyelins in the TME were associated with CD8+ T cell exhaustion and limited the effectiveness of anti-PD-1 treatment (154). These findings offer a preliminary rationale for the use of lipid molecules as predictive biomarkers and interventional targets.
Metabolic Imaging: Metabolic Imaging refers to a class of non-invasive radiological techniques that can dynamically visualize metabolic process in living subjects. Its most prominent advantage is that it can delineate the metabolic state of tumors in both spatial and temporal dimensions. For example, novel single-tracer, multi-parametric PET imaging technology that can simultaneously evaluate tumor blood flow perfusion and glucose metabolic rate in a single examination enables a powerful methodology to identify metabolically aberrant regions and acts as an robust tool to evaluate early treatment response and predict resistance (155).
Nanomaterials: Nanomaterials are mainly used as intelligent drug delivery vehicles in cancer therapeutics. The most important application of nanomaterials is to achieve targeted delivery and controlled release. Compared with conventional drugs, nanomaterials are intelligent drugs that can overcome the limitations of conventional drugs, such as poor solubility, significant systemic toxicity, and low bioavailability. For example, LNPs have been used to deliver peptide-based drugs targeting SREBP or to co-deliver mIL-12 mRNA with PD-L1 inhibitors. These approaches can cause significant reversal of the immunosuppressive microenvironment and overcoming resistance in models of liver and colon cancer (156, 157).
Thus, lipidomics, metabolic imaging, and nanomaterials are fundamentally transforming the landscape of cancer therapeutic research. Overall, these three technical platforms constitute an integrated R&D chain from target discovery to clinical application. However, this innovative paradigm is accompanied by several challenges, such as technical challenges, bottlenecks for clinical translation, and challenges associated with multidimensional data integration.
5 Summary and prospect
Lipid metabolism has recently emerged as a major area of interest in cancer biology. The high proliferative rate of tumors and nutrient deprivation/hypoxia induced by tumors result in the fact that lipid metabolism has become the most prevailing metabolic process in cancer cells. There is an intricate and complex interplay between lipid metabolism and the TME. Lipid metabolism in cancer cells is not only regulated by intracellular oncogenic signaling, but also by the TME, which is composed of various cell types, cytokines, and growth factors. Meanwhile, abnormal lipid metabolism can modulate the oncogenic signaling pathways in cancer cells and the neighboring cell populations in the TME. Despite the recent advances in the understanding of the effects of lipid metabolism in cancer immunotherapy, there are still many gaps in understanding the dynamic regulatory mechanisms, TME-driven heterogeneity, and clinical translation. We believe that, in the future, investigations should focus on the following questions: deciphering the complex signaling networks that regulate the program of lipid metabolic reprogramming and defining their precise roles in regulating immune responses are fundamental to resolving the currently existing uncertainties in context-dependent regulation and TME-induced heterogeneity. Meanwhile, efforts should be focused on the development of novel therapeutic strategies that specifically target critical nodes in these metabolic pathways. We believe that targeting lipid metabolism will become a new frontier in cancer immunotherapy with transformative insights and novel therapeutic strategies against cancers as research proceeds.
Author contributions
KS: Writing – original draft, Writing – review & editing. BP: Writing – original draft. RZ: Writing – original draft. KL: Writing – review & editing. JS: Writing – review & editing. XW: Writing – review & editing. LL: Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by the National Natural Science Foundation of China funded projects (No. 82100205) and the Project of Liaoning Provincial Department of Education (No. LJ212410161086). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. The authors acknowledge the use of the Generative AI tool DeepSeek (specifically, the DeepSeek Chat model, developed by DeepSeek Company) for the purpose of language polishing and refinement of the manuscript text. The authors are solely responsible for the entire scientific content, accuracy, and integrity of this work.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Mattiuzzi C and Lippi G. Current cancer epidemiology. J Epidemiol Glob Health. (2019) 9:217–22. doi: 10.2991/jegh.k.191008.001
2. Hanahan D and Coussens LM. Accessories to the crime: functions of cells recruited to the tumor microenvironment. Cancer Cell. (2012) 21:309–22. doi: 10.1016/j.ccr.2012.02.022
3. Corn KC, Windham MA, and Rafat M. Lipids in the tumor microenvironment: From cancer progression to treatment. Prog Lipid Res. (2020) 80:101055. doi: 10.1016/j.plipres.2020.101055
4. Carracedo A, Cantley LC, and Pandolfi PP. Cancer metabolism: fatty acid oxidation in the limelight. Nat Rev Cancer. (2013) 13:227–32. doi: 10.1038/nrc3483
5. Wen YA, Xing X, Harris JW, Zaytseva YY, Mitov MI, Napier DL, et al. Adipocytes activate mitochondrial fatty acid oxidation and autophagy to promote tumor growth in colon cancer. Cell Death Dis. (2017) 8:e2593. doi: 10.1038/cddis.2017.21
6. Su P, Wang Q, Bi E, Ma X, Liu L, Yang M, et al. Enhanced lipid accumulation and metabolism are required for the differentiation and activation of tumor-associated macrophages. Cancer Res. (2020) 80:1438–50. doi: 10.1158/0008-5472.CAN-19-2994
7. Röhrig F and Schulze A. The multifaceted roles of fatty acid synthesis in cancer. Nat Rev Cancer. (2016) 16:732–49. doi: 10.1038/nrc.2016.89
8. Su X and Abumrad NA. Cellular fatty acid uptake: a pathway under construction. Trends Endocrinol Metab. (2009) 20:72–7. doi: 10.1016/j.tem.2008.11.001
9. Koundouros N and Poulogiannis G. Reprogramming of fatty acid metabolism in cancer. Br J Cancer. (2020) 122:4–22. doi: 10.1038/s41416-019-0650-z
10. Hao J, Yan F, Zhang Y, Triplett A, Zhang Y, Schultz DA, et al. Expression of adipocyte/macrophage fatty acid-binding protein in tumor-associated macrophages promotes breast cancer progression. Cancer Res. (2018) 78:2343–55. doi: 10.1158/0008-5472.CAN-17-2465
11. Zhang Y, Sun Y, Rao E, Yan F, Li Q, Zhang Y, et al. Fatty acid-binding protein E-FABP restricts tumor growth by promoting IFN-β responses in tumor-associated macrophages. Cancer Res. (2014) 74:2986–98. doi: 10.1158/0008-5472.CAN-13-2689
12. Xiang W, Lv H, Xing F, Sun X, Ma Y, Wu L, et al. Inhibition of ACLY overcomes cancer immunotherapy resistance via polyunsaturated fatty acids peroxidation and cGAS-STING activation. Sci Adv. (2023) 9:eadi2465. doi: 10.1126/sciadv.adi2465
13. Li X, Yu W, Qian X, Xia Y, Zheng Y, Lee JH, et al. Nucleus-translocated ACSS2 promotes gene transcription for lysosomal biogenesis and autophagy. Mol Cell. (2017) 66:684–97.e9. doi: 10.1016/j.molcel.2017.04.026
14. Schug ZT, Peck B, Jones DT, Zhang Q, Grosskurth S, Alam IS, et al. Acetyl-CoA synthetase 2 promotes acetate utilization and maintains cancer cell growth under metabolic stress. Cancer Cell. (2015) 27:57–71. doi: 10.1016/j.ccell.2014.12.002
15. Comerford SA, Huang Z, Du X, Wang Y, Cai L, Witkiewicz AK, et al. Acetate dependence of tumors. Cell. (2014) 159:1591–602. doi: 10.1016/j.cell.2014.11.020
16. Mashimo T, Pichumani K, Vemireddy V, Hatanpaa KJ, Singh DK, Sirasanagandla S, et al. Acetate is a bioenergetic substrate for human glioblastoma and brain metastases. Cell. (2014) 159:1603–14. doi: 10.1016/j.cell.2014.11.025
17. Mukhi D, Li L, Liu H, Doke T, Kolligundla LP, Ha E, et al. ACSS2 gene variants determine kidney disease risk by controlling de novo lipogenesis in kidney tubules. J Clin Invest. (2023) 134:e172963. doi: 10.1172/JCI172963
18. Lakhter AJ, Hamilton J, Konger RL, Brustovetsky N, Broxmeyer HE, and Naidu SR. Glucose-independent acetate metabolism promotes melanoma cell survival and tumor growth. J Biol Chem. (2016) 291:21869–79. doi: 10.1074/jbc.M115.712166
19. Yun M, Bang SH, Kim JW, Park JY, Kim KS, and Lee JD. The importance of acetyl coenzyme A synthetase for 11C-acetate uptake and cell survival in hepatocellular carcinoma. J Nucl Med. (2009) 50:1222–8. doi: 10.2967/jnumed.109.062703
20. Zhang J, Baddoo M, Han C, Strong MJ, Cvitanovic J, Moroz K, et al. Gene network analysis reveals a novel 22-gene signature of carbon metabolism in hepatocellular carcinoma. Oncotarget. (2016) 7:49232–45. doi: 10.18632/oncotarget.10249
21. Zhou L, Song Z, Hu J, Liu L, Hou Y, Zhang X, et al. ACSS3 represses prostate cancer progression through downregulating lipid droplet-associated protein PLIN3. Theranostics. (2021) 11:841–60. doi: 10.7150/thno.49384
22. Chin K, DeVries S, Fridlyand J, Spellman PT, Roydasgupta R, Kuo WL, et al. Genomic and transcriptional aberrations linked to breast cancer pathophysiologies. Cancer Cell. (2006) 10:529–41. doi: 10.1016/j.ccr.2006.10.009
23. Luo DX, Tong DJ, Rajput S, Wang C, Liao DF, Cao D, et al. Targeting acetyl-CoA carboxylases: small molecular inhibitors and their therapeutic potential. Recent Pat Anticancer Drug Discov. (2012) 7:168–84. doi: 10.2174/157489212799972918
24. Li K, Zhang C, Chen L, Wang P, Fang Y, Zhu J, et al. The role of acetyl-coA carboxylase2 in head and neck squamous cell carcinoma. PeerJ. (2019) 7:e7037. doi: 10.7717/peerj.7037
25. German NJ, Yoon H, Yusuf RZ, Murphy JP, Finley LW, Laurent G, et al. PHD3 loss in cancer enables metabolic reliance on fatty acid oxidation via deactivation of ACC2. Mol Cell. (2016) 63:1006–20. doi: 10.1016/j.molcel.2016.08.014
26. Garrido F, Romero I, Aptsiauri N, and Garcia-Lora AM. Generation of MHC class I diversity in primary tumors and selection of the Malignant phenotype. Int J Cancer. (2016) 138:271–80. doi: 10.1002/ijc.29375
27. Carretero R, Romero JM, Ruiz-Cabello F, Maleno I, Rodriguez F, Camacho FM, et al. Analysis of HLA class I expression in progressing and regressing metastatic melanoma lesions after immunotherapy. Immunogenetics. (2008) 60:439–47. doi: 10.1007/s00251-008-0303-5
28. Seliger B, Maeurer MJ, and Ferrone S. Antigen-processing machinery breakdown and tumor growth. Immunol Today. (2000) 21:455–64. doi: 10.1016/S0167-5699(00)01692-3
29. Nowak AK, Lake RA, Marzo AL, Scott B, Heath WR, Collins EJ, et al. Induction of tumor cell apoptosis in vivo increases tumor antigen cross-presentation, cross-priming rather than cross-tolerizing host tumor-specific CD8 T cells. J Immunol. (2003) 170:4905–13. doi: 10.4049/jimmunol.170.10.4905
30. Sánchez-Paulete AR, Cueto FJ, Martínez-López M, Labiano S, Morales-Kastresana A, Rodríguez-Ruiz ME, et al. Cancer immunotherapy with immunomodulatory anti-CD137 and anti-PD-1 monoclonal antibodies requires BATF3-dependent dendritic cells. Cancer Discov. (2016) 6:71–9. doi: 10.1158/2159-8290.CD-15-0510
31. Menendez JA and Lupu R. Fatty acid synthase and the lipogenic phenotype in cancer pathogenesis. Nat Rev Cancer. (2007) 7:763–77. doi: 10.1038/nrc2222
32. Bandyopadhyay S, Zhan R, Wang Y, Pai SK, Hirota S, Hosobe S, et al. Mechanism of apoptosis induced by the inhibition of fatty acid synthase in breast cancer cells. Cancer Res. (2006) 66:5934–40. doi: 10.1158/0008-5472.CAN-05-3197
33. Ali A, Levantini E, Teo JT, Goggi J, Clohessy JG, Wu CS, et al. Fatty acid synthase mediates EGFR palmitoylation in EGFR mutated non-small cell lung cancer. EMBO Mol Med. (2018) 10:e8313. doi: 10.15252/emmm.201708313
34. Bollu LR, Ren J, Blessing AM, Katreddy RR, Gao G, Xu L, et al. Involvement of de novo synthesized palmitate and mitochondrial EGFR in EGF induced mitochondrial fusion of cancer cells. Cell Cycle. (2014) 13:2415–30. doi: 10.4161/cc.29338
35. Huang GM, Jiang QH, Cai C, Qu M, and Shen W. SCD1 negatively regulates autophagy-induced cell death in human hepatocellular carcinoma through inactivation of the AMPK signaling pathway. Cancer Lett. (2015) 358:180–90. doi: 10.1016/j.canlet.2014.12.036
36. Fritz V, Benfodda Z, Rodier G, Henriquet C, Iborra F, Avancès C, et al. Abrogation of de novo lipogenesis by stearoyl-CoA desaturase 1 inhibition interferes with oncogenic signaling and blocks prostate cancer progression in mice. Mol Cancer Ther. (2010) 9:1740–54. doi: 10.1158/1535-7163.MCT-09-1064
37. Von Roemeling CA, Marlow LA, Wei JJ, Cooper SJ, Caulfield TR, Wu K, et al. Stearoyl-CoA desaturase 1 is a novel molecular therapeutic target for clear cell renal cell carcinoma. Clin Cancer Res. (2013) 19:2368–80. doi: 10.1158/1078-0432.CCR-12-3249
38. Holder AM, Gonzalez-Angulo AM, Chen H, Akcakanat A, Do KA, Fraser Symmans W, et al. High stearoyl-CoA desaturase 1 expression is associated with shorter survival in breast cancer patients. Breast Cancer Res Treat. (2013) 137:319–27. doi: 10.1007/s10549-012-2354-4
39. Scaglia N and Igal RA. Inhibition of Stearoyl-CoA Desaturase 1 expression in human lung adenocarcinoma cells impairs tumorigenesis. Int J Oncol. (2008) 33:839–50. doi: 10.1186/s12943-018-0833-x
40. Scaglia N, Chisholm JW, and Igal RA. Inhibition of stearoylCoA desaturase-1 inactivates acetyl-CoA carboxylase and impairs proliferation in cancer cells: role of AMPK. PLoS One. (2009) 4:e6812. doi: 10.1371/journal.pone.0006812
41. Montaigne D, Butruille L, and Staels B. PPAR control of metabolism and cardiovascular functions. Nat Rev Cardiol. (2021) 18:809–23. doi: 10.1038/s41569-021-00569-6
42. Padanad MS, Konstantinidou G, Venkateswaran N, Melegari M, Rindhe S, Mitsche M, et al. Fatty acid oxidation mediated by acyl-coA synthetase long chain 3 is required for mutant KRAS lung tumorigenesis. Cell Rep. (2016) 16:1614–28. doi: 10.1016/j.celrep.2016.07.009
43. Pike LS, Smift AL, Croteau NJ, Ferrick DA, and Wu M. Inhibition of fatty acid oxidation by etomoxir impairs NADPH production and increases reactive oxygen species resulting in ATP depletion and cell death in human glioblastoma cells. Biochim Biophys Acta. (2011) 1807:726–34. doi: 10.1016/j.bbabio.2010.10.022
44. Wang T, Fahrmann JF, Lee H, Li YJ, Tripathi SC, Yue C, et al. JAK/STAT3-regulated fatty acid β-oxidation is critical for breast cancer stem cell self-renewal and chemoresistance. Cell Metab. (2018) 27:136–50.e5. doi: 10.1016/j.cmet.2017.11.001
45. Poulsen L, Siersbæk M, and Mandrup S. PPARs: fatty acid sensors controlling metabolism. Semin Cell Dev Biol. (2012) 23:631–9. doi: 10.1016/j.semcdb.2012.01.003
46. Ozeki S, Cheng J, Tauchi-Sato K, Hatano N, Taniguchi H, and Fujimoto T. Rab18 localizes to lipid droplets and induces their close apposition to the endoplasmic reticulum-derived membrane. J Cell Sci. (2005) 118:2601–11. doi: 10.1242/jcs.02401
47. Walther TC and Farese RV Jr. Lipid droplets and cellular lipid metabolism. Annu Rev Biochem. (2012) 81:687–714. doi: 10.1146/annurev-biochem-061009-102430
48. Olzmann JA and Carvalho P. Dynamics and functions of lipid droplets. Nat Rev Mol Cell Biol. (2019) 20:137–55. doi: 10.1038/s41580-018-0085-z
49. Martin S. Caveolae, lipid droplets, and adipose tissue biology: pathophysiological aspects. Horm Mol Biol Clin Investig. (2013) 15:11–8. doi: 10.1515/hmbci-2013-0035
50. Zechner R, Zimmermann R, Eichmann TO, Kohlwein SD, Haemmerle G, Lass A, et al. FAT SIGNALS–lipases and lipolysis in lipid metabolism and signaling. Cell Metab. (2012) 15:279–91. doi: 10.1016/j.cmet.2011.12.018
51. Chandak PG, Radovic B, Aflaki E, Kolb D, Buchebner M, Fröhlich E, et al. Efficient phagocytosis requires triacylglycerol hydrolysis by adipose triglyceride lipase. J Biol Chem. (2010) 285:20192–201. doi: 10.1074/jbc.M110.107854
52. Yeganeh B, Wiechec E, Ande SR, Sharma P, Moghadam AR, Post M, et al. Targeting the mevalonate cascade as a new therapeutic approach in heart disease, cancer and pulmonary disease. Pharmacol Ther. (2014) 143:87–110. doi: 10.1016/j.pharmthera.2014.02.007
53. Xiu W, Liu X, Hu K, Zhang Q, and Shi H. The role of cholesterol metabolism in lung cancer. Oncol Res. (2024) 32:1613–21. doi: 10.32604/or.2024.047933
54. Ding X, Zhang W, Li S, and Yang H. The role of cholesterol metabolism in cancer. Am J Cancer Res. (2019) 9:219–27. doi: 10.1038/s41568-019-0166-1
55. Gallagher EJ, Zelenko Z, Neel BA, Antoniou IM, Rajan L, Kase N, et al. Elevated tumor LDLR expression accelerates LDL cholesterol-mediated breast cancer growth in mouse models of hyperlipidemia. Oncogene. (2017) 36:6462–71. doi: 10.1038/onc.2017.247
56. Guillaumond F, Bidaut G, Ouaissi M, Servais S, Gouirand V, Olivares O, et al. Cholesterol uptake disruption, in association with chemotherapy, is a promising combined metabolic therapy for pancreatic adenocarcinoma. Proc Natl Acad Sci U.S.A. (2015) 112:2473–8. doi: 10.1073/pnas.1421601112
57. Zhou T, Zhan J, Fang W, Zhao Y, Yang Y, Hou X, et al. Serum low-density lipoprotein and low-density lipoprotein expression level at diagnosis are favorable prognostic factors in patients with small-cell lung cancer (SCLC). BMC Cancer. (2017) 17:269. doi: 10.1186/s12885-017-3239-z
58. Yue S, Li J, Lee SY, Lee HJ, Shao T, Song B, et al. Cholesteryl ester accumulation induced by PTEN loss and PI3K/AKT activation underlies human prostate cancer aggressiveness. Cell Metab. (2014) 19:393–406. doi: 10.1016/j.cmet.2014.01.019
59. Khaidakov M and Mehta JL. Do atherosclerosis and obesity-associated susceptibility to cancer share causative link to oxLDL and LOX-1? Cardiovasc Drugs Ther. (2011) 25:477–87. doi: 10.1007/s10557-011-6330-8
60. Koonen DP, Jacobs RL, Febbraio M, Young ME, Soltys CL, Ong H, et al. Increased hepatic CD36 expression contributes to dyslipidemia associated with diet-induced obesity. Diabetes. (2007) 56:2863–71. doi: 10.2337/db07-0907
61. Zettler ME, Prociuk MA, Austria JA, Massaeli H, Zhong G, and Pierce GN. OxLDL stimulates cell proliferation through a general induction of cell cycle proteins. Am J Physiol Heart Circ Physiol. (2003) 284:H644–53. doi: 10.1152/ajpheart.00494.2001
62. Scoles DR, Xu X, Wang H, Tran H, Taylor-Harding B, Li A, et al. Liver X receptor agonist inhibits proliferation of ovarian carcinoma cells stimulated by oxidized low density lipoprotein. Gynecol Oncol. (2010) 116:109–16. doi: 10.1016/j.ygyno.2009.09.034
63. Hashimoto K, Kataoka N, Mohri S, and Kajiya F. Cadherins in potential link between atherosclerosis and cancer. Int J Cardiol. (2011) 149:397. doi: 10.1016/j.ijcard.2011.03.022
64. Kong Y, Cheng L, Mao F, Zhang Z, Zhang Y, Farah E, et al. Inhibition of cholesterol biosynthesis overcomes enzalutamide resistance in castration-resistant prostate cancer (CRPC). J Biol Chem. (2018) 293:14328–41. doi: 10.1074/jbc.RA118.004442
65. Lee JS, Roberts A, Juarez D, Vo TT, Bhatt S, Herzog LO, et al. Statins enhance efficacy of venetoclax in blood cancers. Sci Transl Med. (2018) 10:eaaq1240. doi: 10.1126/scitranslmed.aaq1240
66. Yang J, Wang L, and Jia R. Role of de novo cholesterol synthesis enzymes in cancer. J Cancer. (2020) 11:1761–7. doi: 10.7150/jca.38598
67. Sharpe LJ and Brown AJ. Controlling cholesterol synthesis beyond 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCR). J Biol Chem. (2013) 288:18707–15. doi: 10.1074/jbc.R113.479808
68. Yoshioka H, Coates HW, Chua NK, Hashimoto Y, Brown AJ, and Ohgane K. A key mammalian cholesterol synthesis enzyme, squalene monooxygenase, is allosterically stabilized by its substrate. Proc Natl Acad Sci U.S.A. (2020) 117:7150–8. doi: 10.1073/pnas.1915923117
69. Stevenson J, Luu W, Kristiana I, and Brown AJ. Squalene mono-oxygenase, a key enzyme in cholesterol synthesis, is stabilized by unsaturated fatty acids. Biochem J. (2014) 461:435–42. doi: 10.1042/BJ20131404
70. Garcia-Bermudez J, Baudrier L, Bayraktar EC, Shen Y, La K, Guarecuco R, et al. Squalene accumulation in cholesterol auxotrophic lymphomas prevents oxidative cell death. Nature. (2019) 567:118–22. doi: 10.1038/s41586-019-0945-5
71. DeNardo DG, Barreto JB, Andreu P, Vasquez L, Tawfik D, Kolhatkar N, et al. CD4(+) T cells regulate pulmonary metastasis of mammary carcinomas by enhancing protumor properties of macrophages. Cancer Cell. (2009) 16:91–102. doi: 10.1016/j.ccr.2009.06.018
72. Goossens P, Rodriguez-Vita J, Etzerodt A, Masse M, Rastoin O, Gouirand V, et al. Membrane cholesterol efflux drives tumor-associated macrophage reprogramming and tumor progression. Cell Metab. (2019) 29:1376–89.e4. doi: 10.1016/j.cmet.2019.02.016
73. Beaven SW and Tontonoz P. Nuclear receptors in lipid metabolism: targeting the heart of dyslipidemia. Annu Rev Med. (2006) 57:313–29. doi: 10.1146/annurev.med.57.121304.131428
74. Zabielska J, Sledzinski T, and Stelmanska E. Acyl-coenzyme A: cholesterol acyltransferase inhibition in cancer treatment. Anticancer Res. (2019) 39:3385–94. doi: 10.21873/anticanres.13482
75. Jiang X, Xu J, Liu M, Xing H, Wang Z, Huang L, et al. Adoptive CD8(+) T cell therapy against cancer:Challenges and opportunities. Cancer Lett. (2019) 462:23–32. doi: 10.1016/j.canlet.2019.07.017
76. Chapman NM and Chi H. Metabolic adaptation of lymphocytes in immunity and disease. Immunity. (2022) 55:14–30. doi: 10.1016/j.immuni.2021.12.012
77. Kidani Y, Elsaesser H, Hock MB, Vergnes L, Williams KJ, Argus JP, et al. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat Immunol. (2013) 14:489–99. doi: 10.1038/ni.2570
78. Su W, Chapman NM, Wei J, Zeng H, Dhungana Y, Shi H, et al. Protein prenylation drives discrete signaling programs for the differentiation and maintenance of effector T(reg) cells. Cell Metab. (2020) 32:996–1011.e7. doi: 10.1016/j.cmet.2020.10.022
79. Endo Y, Asou HK, Matsugae N, Hirahara K, Shinoda K, Tumes DJ, et al. Obesity drives th17 cell differentiation by inducing the lipid metabolic kinase, ACC1. Cell Rep. (2015) 12:1042–55. doi: 10.1016/j.celrep.2015.07.014
80. Endo Y, Onodera A, Obata-Ninomiya K, Koyama-Nasu R, Asou HK, Ito T, et al. ACC1 determines memory potential of individual CD4(+) T cells by regulating de novo fatty acid biosynthesis. Nat Metab. (2019) 1:261–75. doi: 10.1038/s42255-018-0025-4
81. Lim SA, Wei J, Nguyen TM, Shi H, Su W, Palacios G, et al. Lipid signalling enforces functional specialization of T(reg) cells in tumours. Nature. (2021) 591:306–11. doi: 10.1038/s41586-021-03235-6
82. Wang H, Franco F, Tsui YC, Xie X, Trefny MP, Zappasodi R, et al. CD36-mediated metabolic adaptation supports regulatory T cell survival and function in tumors. Nat Immunol. (2020) 21:298–308. doi: 10.1038/s41590-019-0589-5
83. Xu S, Chaudhary O, Rodríguez-Morales P, Sun X, Chen D, Zappasodi R, et al. Uptake of oxidized lipids by the scavenger receptor CD36 promotes lipid peroxidation and dysfunction in CD8(+) T cells in tumors. Immunity. (2021) 54:1561–77.e7. doi: 10.1016/j.immuni.2021.05.003
84. Ma X, Xiao L, Liu L, Ye L, Su P, Bi E, et al. CD36-mediated ferroptosis dampens intratumoral CD8(+) T cell effector function and impairs their antitumor ability. Cell Metab. (2021) 33:1001–12.e5. doi: 10.1016/j.cmet.2021.02.015
85. Hu B, Lin JZ, Yang XB, and Sang XT. Aberrant lipid metabolism in hepatocellular carcinoma cells as well as immune microenvironment: A review. Cell Prolif. (2020) 53:e12772. doi: 10.1111/cpr.12772
86. Angelin A, Gil-De-Gómez L, Dahiya S, Jiao J, Guo L, Levine MH, et al. Foxp3 reprograms T cell metabolism to function in low-glucose, high-lactate environments. Cell Metab. (2017) 25:1282–93.e7. doi: 10.1016/j.cmet.2016.12.018
87. Ma X, Bi E, Lu Y, Su P, Huang C, Liu L, et al. Cholesterol induces CD8(+) T cell exhaustion in the tumor microenvironment. Cell Metab. (2019) 30:143–56.e5. doi: 10.1016/j.cmet.2019.04.002
88. Yuan J, Cai T, Zheng X, Ren Y, Qi J, Lu X, et al. Potentiating CD8(+) T cell antitumor activity by inhibiting PCSK9 to promote LDLR-mediated TCR recycling and signaling. Protein Cell. (2021) 12:240–60. doi: 10.1007/s13238-021-00821-2
89. Liu Y, Xu R, Gu H, Zhang E, Qu J, Cao W, et al. Metabolic reprogramming in macrophage responses. biomark Res. (2021) 9:1. doi: 10.1186/s40364-020-00251-y
90. Schumann T, Adhikary T, Wortmann A, Finkernagel F, Lieber S, Schnitzer E, et al. Deregulation of PPARβ/δ target genes in tumor-associated macrophages by fatty acid ligands in the ovarian cancer microenvironment. Oncotarget. (2015) 6:13416–33. doi: 10.18632/oncotarget.3826
91. Park J, Lee SE, Hur J, Hong EB, Choi JI, Yang JM, et al. M-CSF from cancer cells induces fatty acid synthase and PPARβ/δ Activation in tumor myeloid cells, leading to tumor progression. Cell Rep. (2015) 10:1614–25. doi: 10.1016/j.celrep.2015.02.024
92. Catalano S, Mauro L, Bonofiglio D, Pellegrino M, Qi H, Rizza P, et al. In vivo and in vitro evidence that PPARγ ligands are antagonists of leptin signaling in breast cancer. Am J Pathol. (2011) 179:1030–40. doi: 10.1016/j.ajpath.2011.04.026
93. Bonofiglio D, Qi H, Gabriele S, Catalano S, Aquila S, Belmonte M, et al. Peroxisome proliferator-activated receptor gamma inhibits follicular and anaplastic thyroid carcinoma cells growth by upregulating p21Cip1/WAF1 gene in a Sp1-dependent manner. Endocr Relat Cancer. (2008) 15:545–57. doi: 10.1677/ERC-07-0272
94. King RJ, Singh PK, and Mehla K. The cholesterol pathway: impact on immunity and cancer. Trends Immunol. (2022) 43:78–92. doi: 10.1016/j.it.2021.11.007
95. Luo Q, Zheng N, Jiang L, Wang T, Zhang P, Liu Y, et al. Lipid accumulation in macrophages confers protumorigenic polarization and immunity in gastric cancer. Cancer Sci. (2020) 111:4000–11. doi: 10.1111/cas.14616
96. Zhu L, Yang T, Li L, Sun L, Hou Y, Hu X, et al. TSC1 controls macrophage polarization to prevent inflammatory disease. Nat Commun. (2014) 5:4696. doi: 10.1038/ncomms5696
97. Wang Y, Xiang Y, Xin VW, Wang XW, Peng XC, Liu XQ, et al. Dendritic cell biology and its role in tumor immunotherapy. J Hematol Oncol. (2020) 13:107. doi: 10.1186/s13045-020-00939-6
98. Jhunjhunwala S, Hammer C, and Delamarre L. Antigen presentation in cancer: insights into tumour immunogenicity and immune evasion. Nat Rev Cancer. (2021) 21:298–312. doi: 10.1038/s41568-021-00339-z
99. Wculek SK, Cueto FJ, Mujal AM, Melero I, Krummel MF, and Sancho D. Dendritic cells in cancer immunology and immunotherapy. Nat Rev Immunol. (2020) 20:7–24. doi: 10.1038/s41577-019-0210-z
100. Banchereau J and Steinman RM. Dendritic cells and the control of immunity. Nature. (1998) 392:245–52. doi: 10.1038/32588
101. Hilligan KL and Ronchese F. Antigen presentation by dendritic cells and their instruction of CD4+ T helper cell responses. Cell Mol Immunol. (2020) 17:587–99. doi: 10.1038/s41423-020-0465-0
102. Steinman RM. Decisions about dendritic cells: past, present, and future. Annu Rev Immunol. (2012) 30:1–22. doi: 10.1146/annurev-immunol-100311-102839
103. Ghesquière B, Wong BW, Kuchnio A, and Carmeliet P. Metabolism of stromal and immune cells in health and disease. Nature. (2014) 511:167–76. doi: 10.1038/nature13312
104. Arai R, Soda S, Okutomi T, Morita H, Ohmi F, Funakoshi T, et al. Lipid accumulation in peripheral blood dendritic cells and anticancer immunity in patients with lung cancer. J Immunol Res. (2018) 2018:5708239. doi: 10.1155/2018/5708239
105. O'Neill LA and Pearce EJ. Immunometabolism governs dendritic cell and macrophage function. J Exp Med. (2016) 213:15–23. doi: 10.1084/jem.20151570
106. Jiang L, Fang X, Wang H, Li D, and Wang X. Ovarian cancer-intrinsic fatty acid synthase prevents anti-tumor immunity by disrupting tumor-infiltrating dendritic cells. Front Immunol. (2018) 9:2927. doi: 10.3389/fimmu.2018.02927
107. Kratchmarov R, Viragova S, Kim MJ, Rothman NJ, Liu K, Reizis B, et al. Metabolic control of cell fate bifurcations in a hematopoietic progenitor population. Immunol Cell Biol. (2018) 96:863–71. doi: 10.1111/imcb.12040
108. Shim JK, Choi S, Yoon SJ, Choi RJ, Park J, Lee EH, et al. Etomoxir, a carnitine palmitoyltransferase 1 inhibitor, combined with temozolomide reduces stemness and invasiveness in patient-derived glioblastoma tumorspheres. Cancer Cell Int. (2022) 22:309. doi: 10.1186/s12935-022-02731-7
109. Le Naour F, Hohenkirk L, Grolleau A, Misek DE, Lescure P, Geiger JD, et al. Profiling changes in gene expression during differentiation and maturation of monocyte-derived dendritic cells using both oligonucleotide microarrays and proteomics. J Biol Chem. (2001) 276:17920–31. doi: 10.1074/jbc.M100156200
110. Nencioni A, Grünebach F, Zobywlaski A, Denzlinger C, Brugger W, and Brossart P. Dendritic cell immunogenicity is regulated by peroxisome proliferator-activated receptor gamma. J Immunol. (2002) 169:1228–35. doi: 10.4049/jimmunol.169.3.1228
111. Sun S, Gu Y, Wang J, Chen C, Han S, and Che H. Effects of fatty acid oxidation and its regulation on dendritic cell-mediated immune responses in allergies: an immunometabolism perspective. J Immunol Res. (2021) 2021:7483865. doi: 10.1155/2021/7483865
112. Wang J, Wang J, Hong W, Zhang L, Song L, Shi Q, et al. Optineurin modulates the maturation of dendritic cells to regulate autoimmunity through JAK2-STAT3 signaling. Nat Commun. (2021) 12:6198. doi: 10.1038/s41467-021-26477-4
113. Keating SE, Zaiatz-Bittencourt V, Loftus RM, Keane C, Brennan K, Finlay DK, et al. Metabolic reprogramming supports IFN-γ Production by CD56bright NK cells. J Immunol. (2016) 196:2552–60. doi: 10.4049/jimmunol.1501783
114. Mah AY, Rashidi A, Keppel MP, Saucier N, Moore EK, Alinger JB, et al. Glycolytic requirement for NK cell cytotoxicity and cytomegalovirus control. JCI Insight. (2017) 2:e95128. doi: 10.1172/jci.insight.95128
115. Fridlender ZG, Sun J, Kim S, Kapoor V, Cheng G, Ling L, et al. Polarization of tumor-associated neutrophil phenotype by TGF-beta: "N1" versus "N2" TAN. Cancer Cell. (2009) 16:183–94. doi: 10.1016/j.ccr.2009.06.017
116. Langereis JD, Pickkers P, De Kleijn S, Gerretsen J, De Jonge MI, and Kox M. Spleen-derived IFN-γ induces generation of PD-L1(+)-suppressive neutrophils during endotoxemia. J Leukoc Biol. (2017) 102:1401–9. doi: 10.1189/jlb.3A0217-051RR
117. Marini O, Costa S, Bevilacqua D, Calzetti F, Tamassia N, Spina C, et al. Mature CD10(+) and immature CD10(-) neutrophils present in G-CSF-treated donors display opposite effects on T cells. Blood. (2017) 129:1343–56. doi: 10.1182/blood-2016-04-713206
118. Yasugi E, Horiuchi A, Uemura I, Okuma E, Nakatsu M, Saeki K, et al. Peroxisome proliferator-activated receptor gamma ligands stimulate myeloid differentiation and lipogenensis in human leukemia NB4 cells. Dev Growth Differ. (2006) 48:177–88. doi: 10.1111/j.1440-169X.2006.00855.x
119. Inazawa Y, Nakatsu M, Yasugi E, Saeki K, and Yuo A. Lipid droplet formation in human myeloid NB4 cells stimulated by all trans retinoic acid and granulocyte colony-stimulating factor: possible involvement of peroxisome proliferator-activated receptor gamma. Cell Struct Funct. (2003) 28:487–93. doi: 10.1247/csf.28.487
120. Hsu AY, Huang Q, Pi X, Fu J, Raghunathan K, Ghimire L, et al. Neutrophil-derived vesicles control complement activation to facilitate inflammation resolution. Cell. (2025) 118:1623–41. doi: 10.1016/j.cell.2025.01.021
121. Tian X, Shen H, Li Z, Wang T, and Wang S. Tumor-derived exosomes, myeloid-derived suppressor cells, and tumor microenvironment. J Hematol Oncol. (2019) 12:84. doi: 10.1186/s13045-019-0772-z
122. Ren W, Zhang X, Li W, Feng Q, Feng H, Tong Y, et al. Exosomal miRNA-107 induces myeloid-derived suppressor cell expansion in gastric cancer. Cancer Manag Res. (2019) 11:4023–40. doi: 10.2147/CMAR.S198886
123. Adeshakin AO, Liu W, Adeshakin FO, Afolabi LO, Zhang M, Zhang G, et al. Regulation of ROS in myeloid-derived suppressor cells through targeting fatty acid transport protein 2 enhanced anti-PD-L1 tumor immunotherapy. Cell Immunol. (2021) 362:104286. doi: 10.1016/j.cellimm.2021.104286
124. Hossain F, Al-Khami AA, Wyczechowska D, Hernandez C, Zheng L, Reiss K, et al. Inhibition of fatty acid oxidation modulates immunosuppressive functions of myeloid-derived suppressor cells and enhances cancer therapies. Cancer Immunol Res. (2015) 3:1236–47. doi: 10.1158/2326-6066.CIR-15-0036
125. Xin G, Chen Y, Topchyan P, Kasmani MY, Burns R, Volberding PJ, et al. Targeting PIM1-mediated metabolism in myeloid suppressor cells to treat cancer. Cancer Immunol Res. (2021) 9:454–69. doi: 10.1158/2326-6066.CIR-20-0433
126. Chen X and Song E. Turning foes to friends: targeting cancer-associated fibroblasts. Nat Rev Drug Discov. (2019) 18:99–115. doi: 10.1038/s41573-018-0004-1
127. Kalluri R. The biology and function of fibroblasts in cancer. Nat Rev Cancer. (2016) 16:582–98. doi: 10.1038/nrc.2016.73
128. Gong J, Lin Y, Zhang H, Liu C, Cheng Z, Yang X, et al. Reprogramming of lipid metabolism in cancer-associated fibroblasts potentiates migration of colorectal cancer cells. Cell Death Dis. (2020) 11:267. doi: 10.1038/s41419-020-2434-z
129. Radhakrishnan R, Ha JH, Jayaraman M, Liu J, Moxley KM, Isidoro C, et al. Ovarian cancer cell-derived lysophosphatidic acid induces glycolytic shift and cancer-associated fibroblast-phenotype in normal and peritumoral fibroblasts. Cancer Lett. (2019) 442:464–74. doi: 10.1016/j.canlet.2018.11.023
130. Wang Z, Liu J, Huang H, Ye M, Li X, Wu R, et al. Metastasis-associated fibroblasts: an emerging target for metastatic cancer. biomark Res. (2021) 9:47. doi: 10.1186/s40364-021-00305-9
131. Zhang Y, Gu Z, Wan J, Lou X, Liu S, Wang Y, et al. Stearoyl-CoA Desaturase-1 dependent lipid droplets accumulation in cancer-associated fibroblasts facilitates the progression of lung cancer. Int J Biol Sci. (2022) 18:6114–28. doi: 10.7150/ijbs.74924
132. Burke KP, Chaudhri A, Freeman GJ, and Sharpe AH. The B7:CD28 family and friends: Unraveling coinhibitory interactions. Immunity. (2024) 57:223–44. doi: 10.1016/j.immuni.2024.01.013
133. Ding R, Yu X, Hu Z, Dong Y, Huang H, Zhang Y, et al. Lactate modulates RNA splicing to promote CTLA-4 expression in tumor-infiltrating regulatory T cells. Immunity. (2024) 57:528–40.e6. doi: 10.1016/j.immuni.2024.01.019
134. Hellmann MD, Paz-Ares L, Bernabe Caro R, Zurawski B, Kim SW, Carcereny Costa E, et al. Nivolumab plus ipilimumab in advanced non-small-cell lung cancer. N Engl J Med. (2019) 381:2020–31. doi: 10.1056/NEJMoa1910231
135. Johnson ML, Cho BC, Luft A, Alatorre-Alexander J, Geater SL, Laktionov K, et al. Durvalumab with or without tremelimumab in combination with chemotherapy as first-line therapy for metastatic non-small-cell lung cancer: the phase III POSEIDON study. J Clin Oncol. (2023) 41:1213–27. doi: 10.1200/JCO.22.00975
136. Paz-Ares L, Ciuleanu TE, Cobo M, Schenker M, Zurawski B, Menezes J, et al. First-line nivolumab plus ipilimumab combined with two cycles of chemotherapy in patients with non-small-cell lung cancer (CheckMate 9LA): an international, randomised, open-label, phase 3 trial. Lancet Oncol. (2021) 22:198–211. doi: 10.1016/S1470-2045(20)30641-0
137. Tzeng SF, Yu YR, Park J, Von Renesse J, Hsiao HW, Hsu CH, et al. PLT012, a humanized CD36-blocking antibody, is effective for unleashing antitumor immunity against liver cancer and liver metastasis. Cancer Discov. (2025) 15:1676–96. doi: 10.1158/2159-8290.CD-24-1409
138. Lacher SB, Dörr J, De Almeida GP, Hönninger J, Bayerl F, Hirschberger A, et al. PGE(2) limits effector expansion of tumour-infiltrating stem-like CD8(+) T cells. Nature. (2024) 629:417–25. doi: 10.1038/s41586-024-07254-x
139. Morotti M, Grimm AJ, Hope HC, Arnaud M, Desbuisson M, Rayroux N, et al. PGE(2) inhibits TIL expansion by disrupting IL-2 signalling and mitochondrial function. Nature. (2024) 629:426–34. doi: 10.1038/s41586-024-07352-w
140. Strzyz P. Tying lipid rafts to oncogenic signalling. Nat Rev Mol Cell Biol. (2019) 20:513. doi: 10.1038/s41580-019-0160-0
141. Civita S, Ruglioni M, Mariangeli M, Barachini S, Salvadori T, Cristiani S, et al. Nanoscale engagement of programmed death ligand 1 (PD-L1) in membrane lipid raft domains of cancer cells. FEBS J. (2025). doi: 10.1111/febs.70238
142. Niu C, Wei H, Pan X, Wang Y, Song H, Li C, et al. Foxp3 confers long-term efficacy of chimeric antigen receptor-T cells via metabolic reprogramming. Cell Metab. (2025) 37:1426–41.e7. doi: 10.1016/j.cmet.2025.04.008
143. Wekking D, Silva CAC, Viscò R, Denaro N, Lambertini M, Maccioni A, et al. The interplay between gut microbiota, antibiotics, and immune checkpoint inhibitors in patients with cancer: A narrative review with biological and clinical aspects. Crit Rev Oncol Hematol. (2025) 212:104767. doi: 10.1016/j.critrevonc.2025.104767
144. Ma Y, Temkin SM, Hawkridge AM, Guo C, Wang W, Wang XY, et al. Fatty acid oxidation: An emerging facet of metabolic transformation in cancer. Cancer Lett. (2018) 435:92–100. doi: 10.1016/j.canlet.2018.08.006
145. Lally JSV, Ghoshal S, DePeralta DK, Moaven O, Wei L, Masia R, et al. Inhibition of acetyl-coA carboxylase by phosphorylation or the inhibitor ND-654 suppresses lipogenesis and hepatocellular carcinoma. Cell Metab. (2019) 29:174–82.e5. doi: 10.1016/j.cmet.2018.08.020
146. Mullen PJ, Yu R, Longo J, Archer MC, and Penn LZ. The interplay between cell signalling and the mevalonate pathway in cancer. Nat Rev Cancer. (2016) 16:718–31. doi: 10.1038/nrc.2016.76
147. Kumar N and Mandal CC. Cholesterol-lowering drugs on akt signaling for prevention of tumorigenesis. Front Genet. (2021) 12:724149. doi: 10.3389/fgene.2021.724149
148. Hu T, Shen H, Huang H, Yang Z, Zhou Y, and Zhao G. Cholesterol-lowering drug pitavastatin targets lung cancer and angiogenesis via suppressing prenylation-dependent Ras/Raf/MEK and PI3K/Akt/mTOR signaling. Anticancer Drugs. (2020) 31:377–84. doi: 10.1097/CAD.0000000000000885
149. Hwang KE, Na KS, Park DS, Choi KH, Kim BR, Shim H, et al. Apoptotic induction by simvastatin in human lung cancer A549 cells via Akt signaling dependent down-regulation of survivin. Invest New Drugs. (2011) 29:945–52. doi: 10.1007/s10637-010-9450-2
150. Jiang W, Hu JW, He XR, Jin WL, and He XY. Statins: a repurposed drug to fight cancer. J Exp Clin Cancer Res. (2021) 40:241. doi: 10.1186/s13046-021-02041-2
151. Chen Y, Zhou Y, Ren R, Chen Y, Lei J, and Li Y. Harnessing lipid metabolism modulation for improved immunotherapy outcomes in lung adenocarcinoma. J Immunother Cancer. (2024) 12:e008811. doi: 10.1136/jitc-2024-008811
152. Zhao D, Wu L, and Li Y. Targeting lipid metabolism to enhance cancer immunotherapy. Biochim Biophys Acta Rev Cancer. (2025) 1880:189416. doi: 10.1016/j.bbcan.2025.189416
153. Sun C, Wang A, Zhou Y, Chen P, Wang X, Huang J, et al. Spatially resolved multi-omics highlights cell-specific metabolic remodeling and interactions in gastric cancer. Nat Commun. (2023) 14:2692. doi: 10.1038/s41467-023-38360-5
154. Li Y, Chen Z, Wang D, Du W, Zhu N, Shen X, et al. Transforming acidic coiled-coil-containing protein 3-mediated lipid metabolism reprogramming impairs CD8(+) T-cell cytotoxicity in hepatocellular carcinoma. Signal Transduct Target Ther. (2025) 10:274. doi: 10.1038/s41392-025-02367-9
155. Godbersen GM, Falb P, Klug S, Silberbauer LR, Reed MB, Nics L, et al. Non-invasive assessment of stimulation-specific changes in cerebral glucose metabolism with functional PET. Eur J Nucl Med Mol Imaging. (2024) 51:2283–92. doi: 10.1007/s00259-024-06675-0
156. Luo S, Yang H, Jiang X, Wang Z, He X, Meng Y, et al. Inhibition of tumor lipogenesis and growth by peptide-based targeting of SREBP activation. Adv Sci (Weinh). (2025) 17:e08111. doi: 10.1002/advs.202508111
Keywords: lipid, lipid metabolism, metabolic reprogramming, cancer, tumor microenvironment, immune response
Citation: Shi K, Pan B, Zheng R, Liu K, Song J, Wang X and Li L (2025) Bidirectional regulation of lipid metabolism and the tumor microenvironment: new perspectives from mechanism to therapy. Front. Immunol. 16:1696102. doi: 10.3389/fimmu.2025.1696102
Received: 31 August 2025; Accepted: 29 October 2025;
Published: 11 November 2025.
Edited by:
Subhadeep Roy, Birla Institute of Technology, IndiaReviewed by:
Sanjima Pal, McGill University Health Center, CanadaLakhveer Singh, Gurugram University, India
Copyright © 2025 Shi, Pan, Zheng, Liu, Song, Wang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Li Li, bGlseWRsbXVAaG90bWFpbA==
†These authors have contributed equally to this work