 Sarina C. Lowe
Sarina C. Lowe Madelaine Leitman
Madelaine Leitman Noah Yan
Noah Yan Alexander Hoffmann
Alexander Hoffmann- 1Department of Microbiology, Immunology and Molecular Genetics, University of California, Los Angeles, Los Angeles, CA, United States
- 2Vatche and Tamar Manoukian Division of Digestive Diseases, University of California, Los Angeles, Los Angeles, CA, United States
- 3Department of Medicine, University of California, Los Angeles, Los Angeles, CA, United States
- 4Institute for Quantitative and Computational Biosciences, University of California, Los Angeles, Los Angeles, CA, United States
Macrophages are key innate immune cells responsible for initiating and coordinating immune responses. A major determinant of macrophage function is their tissue-context-dependent polarization state, regulated by the cytokines present in the tissue microenvironment. Yet, the capability of characterizing macrophage polarization states in clinical studies remains limited. Here, we used a defined set of cytokines to polarize human PBMC-derived macrophages and determine their transcriptional signatures and stimulus responsiveness. The resultant atlas of transcriptional signatures for human macrophage polarization states was applied to a dataset of intestinal biopsies from Crohn’s disease patients and healthy controls. Our analysis identified a dominant population of IFNγ-polarized macrophages in areas of active Crohn’s intestinal inflammation and a loss of wound healing IL-4-, IL-10- and IL-13-polarized macrophages. This study demonstrates that in vitro datasets of macrophages in defined conditions can be leveraged to interpret the functionality of cells transcriptional profiled in clinical studies.
Introduction
Macrophages serve diverse roles within the immune system, ranging from the acute response to pathogens to wound healing and maintenance of homeostasis. Tissue microenvironmental cytokines may cause macrophage polarization, resulting in changes in their functional repertoire. These adaptations affect macrophage morphology, signaling dynamics, gene expression, and cytokine production (1–7). This is particularly relevant in the intestine, where most macrophages originate from circulating monocytes (8) that extravasate into the microenvironment of the intestinal lamina propria, where they have a lifespan of 3–5 weeks (9).
In Crohn’s disease, macrophages are preferentially enriched at sites of active inflammation and are a key source of proinflammatory cytokines that contribute to disease pathogenesis (4, 10). Although many of the macrophage-derived cytokines (i.e. TNF, IL-12, IL-23) are therapeutic targets, the repertoire of macrophage polarization states that drive cytokine production in Crohn’s disease is unknown.
Single-cell RNA sequencing (scRNA-seq) enables multi-dimensional interrogation of cell states. Applied to in vitro-cultured macrophages exposed to specific cytokines, it reveals transcriptional profiles that define polarization states. This reference atlas can then be leveraged to analyze in vivo patient-derived scRNA-seq data, enabling characterization of macrophage populations in Crohn’s disease. Prior studies of macrophage subtypes in Crohn’s disease have been limited to categorization as ‘inflammatory’ and ‘noninflammatory’ based on biomarker expression, or gene expression clusters with little clinical or biologic significance (11). Here we produced an extensive scRNA-seq dataset of in-vitro-polarized macrophages that we leveraged to interpret in vivo scRNA-seq data produced from Crohn’s disease clinical samples. As a result, we identified distinct macrophage subsets that are more or less abundant in active Crohn’s disease lesions and regions of healing.
Methods
In vitro macrophage culture, polarization, and stimulation
Human blood from a deidentified donor was obtained from the UCLA CFAR Centralized Laboratory Support Core. Peripheral blood mononuclear cells (PBMCs) were isolated from blood by Ficoll density centrifugation followed by CD14+ bead selection for monocytes. The monocytes were then plated on 6-well tissue culture plates at a density of 5 x 105 cells/well, and cultured in RPMI media supplemented with 10% ES fetal bovine serum and 20 ng/ml human macrophage colony-stimulating factor. Media was replaced 4 days after isolation, and after 6 days the cells were polarized for 24 hours with either 100 U/mL IFN-β (PBL Assay Science, 11415-1), 20 ng/mL IFN-γ (Peprotech, 300-02), 50 ng/mL IL-10 (R&D, 217-IL), 40 ng/mL IL-13 (R&D, 213-ILB), 20 ng/mL IL-4 (R&D, 204-IL), or left naïve. After 24 hours of polarization, the cells were stimulated for 2 hours with LPS 10 ng/ml (Sigma-Aldrich, B5:055), TNF 100 μg/ml (R&D, 210-TA), or left unstimulated.
In-vitro-polarized macrophage scRNA-seq and analysis
Cells were lifted and the samples were multiplexed using the BD Human Single-Cell Multiplexing Kit (BD, 633781) and loaded onto the cartridge following the manufacturer’s instructions (BD Rhapsody #210967). Targeted mRNA amplification was performed utilizing the Immune Response Panel Hs primer panel (BD, 633750), with subsequent library preparation. The pooled library was sequenced with paired-end 100 bp reads on an Illumina NovaSeq X Plus. The raw FASTQ files and reference for the Immune Response Panel were run through the BD Rhapsody Sequence Analysis Pipeline which includes quality filtering with read overlap detection, trimming and filtering, followed by identification of the cell barcode and unique molecular identifier (UMI), alignment to the human genome, and sample determination from the multiplexed sample tags, resulting in an output of adjusted molecule counts.
In vivo terminal ileum scRNA-seq data processing and annotation
Published single-cell RNA sequencing (scRNA-seq) data was obtained from terminal ileal (TI) biopsies of Crohn’s disease patients (12). Immune cells were separated from stromal and epithelial cells based on annotations from the original study (12) and prepared for downstream analysis with log-normalization and standardization of expression. As macrophage subsets identified by marker genes had no clear clinical or physiologic relevance, we combined subsets into an all-encompassing macrophage group. We constructed a UMAP using principal component analysis (PCA) reduction and labeled cell types by manual annotation of the top 20 differentially expressed genes for each cluster of immune cells. Macrophages were identified by high expression of canonical macrophage genes (including C1QA, C1QB, C1QC, CD14, and FCN1). The immune cell clusters were relabeled by manual annotation, and a macrophage-only object was subsequently processed independently.
Integration of in vivo and in vitro macrophage data
To integrate the in vivo TI macrophage data composed of 28, 923 genes with the in-vitro-polarized macrophage data with 344 genes, both datasets were standardized by retaining only the 325 overlapping genes for downstream analysis (13). With the in vivo TI macrophage dataset as the reference, Seurat’s anchor-based integration framework was used with the in-vitro-polarized dataset as the query. Integration was performed using Seurat’s SCTransform-based normalization to mitigate technical noise. Each cell was then projected into a common space by applying an anchor-derived transformation (14). This anchor strategy identifies pairs of transcriptionally similar cells across the datasets, allowing the two datasets to be aligned in a common low-dimensional space. The in-vitro-polarized macrophages were assigned to an in vivo TI unsupervised cluster (1, 2, or 3), mapping the in-vitro-polarized cells’ PCA profile onto the reference in vivo TI macrophage’s PCA space. The most transcriptionally similar in vivo TI cluster based on a mutual nearest-neighbor classification (14) (k=30) was assigned to each in-vitro-polarized macrophage cell profile using the highest similarity score.
Results
We developed a workflow to better characterize the macrophage cell states found in Crohn’s disease (Figure 1). We first systematically profiled in-vitro-polarized human macrophages exposed to defined cytokine polarization conditions and identified polarization gene signatures. Applying these to clinical samples enabled identification of specific intestinal macrophage subpopulations in Crohn’s patients.

Figure 1. Overview of an experimental and analytic workflow to better characterize macrophage states in clinical scRNA-seq datasets. Workflow used to generate an in vitro atlas of macrophage polarization states and apply it to functionally annotate an in vivo Crohn’s clinical dataset. Peripheral blood mononuclear cells (PBMCs) were isolated from human blood and differentiated into macrophages. After six days, cells were polarized with IFNγ, IFNβ, IL-4, IL-10 or IL-13, left unstimulated or stimulated with LPS or TNF, and processed for scRNA-seq using the BD Rhapsody platform. In vivo intestinal immune cell data from healthy controls and Crohn’s disease patients [Kong et al. (12)] were re-analyzed, and macrophages were identified by annotation of cluster-specific differentially expressed genes. Using 325 shared genes and a mutual nearest neighbors approach to identify anchors, the in vivo and in vitro datasets were integrated into a shared space, enabling characterization of the polarizating state of macrophages present in clinical samples. Created in BioRender. Lowe, S. (2025) https://BioRender.com/2hr0tbu.
To prolife single macrophage polarization states we cultured PBMC-derived macrophages and polarized them with individual cytokines from a larger panel. We used an established sc-RNAseq targeted gene platform with a select panel of immune response genes that show stimulus-specific expression and assess the functional state of macrophages (15). The in-vitro-polarized macrophages showed distinct transcriptional profiles after 24 hours of polarization with IFNγ, IFNβ, IL-4, IL-10 and IL-13 (Figure 2A). The expression of known M1 and M2 macrophage marker genes (Supplementary Figure S1) showed heterogeneity across the transcriptional profiles of the 6 polarization conditions (naïve and 5 cytokines), showing the need for more robust classification of macrophage polarization beyond the M1 and M2 dichotomy.

Figure 2. In vitro polarization and stimulation of human PBMC-derived macrophages define an atlas of macrophage polarization states. (A) UMAP of PBMC-derived day 6 macrophages polarized for 24 hours without acute stimulation. The samples are colored by polarization condition with naïve in green, IL-13 in yellow, IL-10 in orange, IFNγ in blue, IFNβ in purple and IL-4 in red. (B) UMAP of polarized macrophages subsequently stimulated for 3 hours with 10ng/ml LPS. (C) UMAP of polarized macrophages subsequently stimulated for 3 hours with 100μg/ml LPS. (A–C) are all mapped in the same UMAP space. (D–F) Violin plots of TNF, IL1β and CXCL10 expression in polarized macrophages stimulated with LPS. (G–I) Violin plots of TNF, IL1β and CXCL10 expression in polarized macrophages stimulated with TNF. Each dot represents an individual cell while the density curve depicts the distribution of numeric data. Counts were normalized by regularized negative binomial regression.
To assess functional differences between these polarization states, the macrophages were subsequently stimulated with either lipopolysaccharide (LPS) (Figure 2B) or tumor necrosis factor (TNF) (Figure 2C). The stimulated macrophages showed differences in expression of key inflammatory cytokines and chemokines when normalized by regularized negative binomial regression. In response to LPS stimulation, IFNγ- and IFNβ-polarized macrophages showed increased TNF expression compared to naïve cells, while the IL-10-polarized macrophages displayed reduced TNF expression (Figure 2D). However, this polarization effect on LPS-responsive gene expression was not uniform across all cytokines and chemokines. IL1β expression showed minimal variation between polarization conditions (Figure 2E), whereas CXCL10 expression differed markedly, with IFNγ- and IFNβ-polarized macrophages exhibiting much higher levels of expression than other conditions (Figure 2F). Notably, these gene-specific polarization effects differed between LPS and TNF stimulation. With TNF stimulation, there was little difference in subsequent TNF expression between polarization conditions (Figure 2G), but an increase in IL1β expression in the naïve and IL-10 polarized macrophages compared to the other conditions (Figure 2H), while CXCL10 was not expressed in any condition but the IFNγ and IFNβ polarized (Figure 2I). By assessing inflammatory gene expression following stimulation, we identified functional differences in macrophages across polarization conditions.
After establishing an atlas of polarized macrophage transcriptional states, we leveraged it to better characterize the macrophage populations present in Crohn’s disease. Terminal ileum data was available from 27 Crohn’s patients and 9 healthy donors (Figures 3A, B), with a higher percentage of macrophages in the Crohn’s samples, predominantly in areas of active inflammation (Figure 3C). Macrophage-specific data from healthy donors (N = 9), Crohn’s noninflamed biopsies (N = 23), and Crohn’s inflamed biopsies (N = 8) (Figure 3D) revealed notable differences in macrophage subgroup distribution. Cluster 2 macrophages were dominant in biopsies from healthy donors, while cluster 1 macrophages were dominant in Crohn’s noninflamed biopsies, and cluster 3 in Crohn’s inflamed biopsies (Figure 3E). The same workflow was applied to colonic macrophages from the published dataset; however, the number of macrophages, particularly from Crohn’s inflamed donors, was too low to reliably assign polarization states (Supplementary Figure S2). For instance, Cluster 3, contained only 271 cells (Supplementary Figure S2D). This limitation was not due to a reduction in the proportion of macrophages within Crohn’s disease colon samples, which was comparable to that in healthy donors (Supplementary Figure S2C). Rather, there were fewer colonic samples from Crohn’s inflamed donors (N = 5) compared with Crohn’s non-inflamed (N = 17) and healthy (N = 16) samples.

Figure 3. Immune cell compositions of terminal ileal samples using established marker genes. (A) UMAP of terminal ileum immune cells from healthy donors (N = 9) and Crohn’s patients (N = 27), from both noninflamed and inflamed regions. Immune cells were separated from stromal and epithelial cells based on annotations from the original study (Kong et al) and clustered in an unsupervised manner. The numbers delineate each individual cluster. (B) Immune cell types manually annotated based on the top 20 differentially expressed genes for each unsupervised cluster, with some clusters consolidated into broader cell type categories. The macrophages are shown in pink. (C) Percentage of immune cell types (of total immune cells) stratified by donor types (healthy in blue, Crohn’s noninflamed in purple, and Crohn’s inflamed in pink). (D) Unsupervised clustering of the macrophage subset, separated by donor type. Cluster 1 is marked in blue, cluster 2 in green, and cluster 3 in orange. (E) Distribution of macrophages assigned to each of the three clusters (colors as in D), separated by donor type.
The in-vitro-polarized macrophage data (Figure 4A) was integrated with the in vivo Crohn’s disease terminal ileum dataset (Figure 4B) using a shared set of 325 genes. After integration, the polarization labels from the in vitro macrophages were projected onto the in vivo dataset using a weighted nearest-neighbor (k=30) classification (Figure 4C), where each in vitro macrophage was assigned the label of its most transcriptionally similar in vivo cell based on the highest similarity score (Figure 4D).
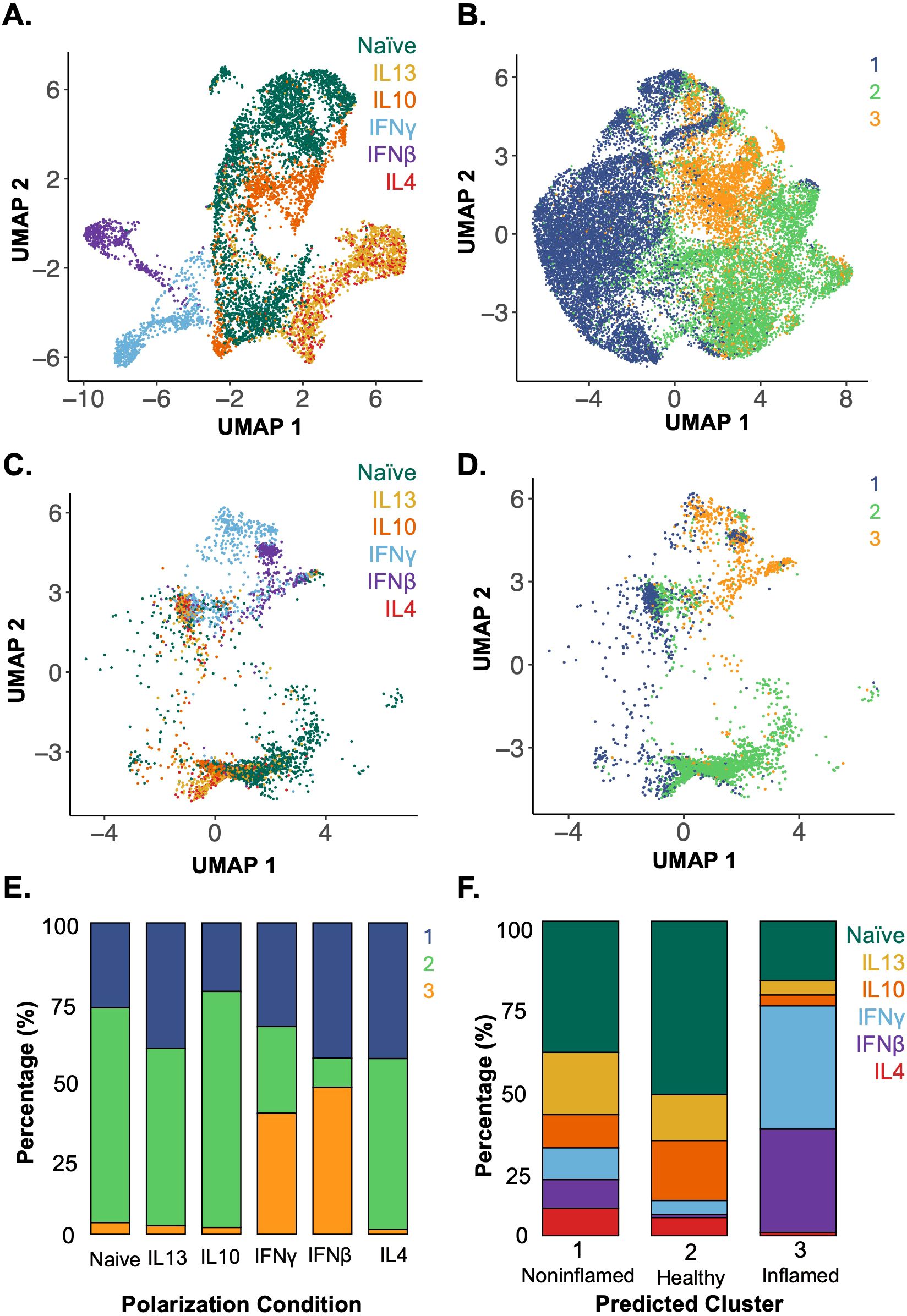
Figure 4. Applying a model trained on in-vitro-polarized macrophage datasets identifies the polarization states of in vivo terminal ileal macrophages in clinical scRNA-seq data. (A) UMAP of in-vitro-polarized macrophage scRNA-seq data using the shared set of 325 genes labeled by polarization condition (colors as in Figure 2). (B) UMAP of in vivo terminal ileal macrophages labeled by unsupervised clusters from Figure 3D after reducing the 28, 923 gene dataset to the 325 genes shared with the in vitro atlas. (C) In-vitro-polarized macrophages integrated with the terminal ileal dataset using SCTransform anchors and mapped onto a shared UMAP, labeled by polarization condition. (D) Assignment of in-vitro-polarized macrophages to in vivo clusters 1–3 using a weighted mutual nearest neighbor approach. (E) Proportion of cells from each polarization condition assigned to each in vivo macrophage cluster by mutual nearest-neighbor classification. (F) Assignment of polarization states to macrophages found in vivo which were grouped by unsupervised clustering (Figure 3D) and are predominantly associated with noninflamed (cluster 1), healthy (cluster 2) and inflamed (cluster 3) tissues (Figure 3E).
When examining the different macrophage polarization states and their assigned in vivo clusters, the majority of naïve, IL-4, IL-10 and IL-13 in-vitro-polarized macrophages aligned to cluster 2, the cluster most highly represented in biopsies from healthy donors. IFNβ- and IFNγ-polarized macrophages aligned with cluster 3, which best represented macrophages from areas of active Crohn’s disease (Figure 4E). This integrated data and cluster assignments were used to examine the polarization states of the in vivo terminal ileum macrophage populations (Figure 4F). Macrophages in cluster 1 were distributed throughout the range of polarization conditions with the largest portion of cells aligning to the naïve and IL-13-polarized states. Cluster 2 was enriched in cells aligning to the naïve condition and under-enriched in IFN-polarized cells. Lastly, in cluster 3, which was highly enriched in macrophages from inflamed Crohn’s subjects, the majority of macrophages aligned to the IFNβ- and IFNγ-polarized conditions. These analyses were able to characterize the polarization state of macrophages within biopsies based on an atlas of in-vitro-defined polarized macrophages.
Discussion
Culturing macrophages with defined cytokines produced distinct populations of polarized macrophages. Not only did the different macrophage polarization states exhibit varying baseline transcriptomic profiles, but their gene expression in response to stimuli, both a pathogen associated molecular pattern (PAMP), lipopolysaccharide (LPS), and a proinflammatory cytokine, tumor necrosis factor (TNF), differed, highlighting the impact of polarization on macrophage function.
By applying a model based on in vitro single-cell macrophage polarization data to in vivo single-cell data from Crohn’s patients, this analysis revealed macrophages from inflamed Crohn’s samples have a transcriptional program closest to proinflammatory IFN-polarized macrophages at the expense of anti-inflammatory IL-4-, IL-10- and IL-13-polarized macrophages. The results support the notion of the cytokine microenvironment as a driver for distinct macrophage subsets in Crohn’s disease. IFNγ is largely produced by innate lymphoid cells (ILCs), NK cells, and activated T cells, and is elevated in the mucosa of patients with Crohn’s disease (16) along with animal models of colitis (17). From a macrophage standpoint, the IFNγ environment is significant because it is known to polarize towards a proinflammatory macrophage phenotype (18), which exhibits enhanced inflammatory responses with increased proinflammatory cytokine production (19). The development of IFNγ-polarized macrophages in response to an inflammatory environment further promotes intestinal inflammation as these macrophages are primed to produce more proinflammatory cytokines, activating and recruiting further immune effector cells.
In contrast, the cluster of macrophages from noninflamed segments (cluster 1) exhibited an even distribution of macrophage polarization states, and particularly more IL-4-, IL-10- and IL-13-polarized macrophages. These samples were from patients with Crohn’s disease, but with inactive disease, or no evidence of active inflammation in the biopsied portion of terminal ileum. The distribution of polarization states skewing towards more IL-4-, IL-10- and IL-13-polarized macrophages is consistent with the pivotal role of anti-inflammatory macrophages in wound healing and resolution of inflammation (4).
The findings highlight the potential pathogenic role of IFN-polarized macrophages in active Crohn’s intestinal inflammation, and the possible healing role of anti-inflammatory IL-4-, IL-10- and IL-13-polarized macrophages. This could impact therapy by identifying patients with a predominant interferon macrophage signature who may benefit from JAK-STAT inhibition, which is a therapeutic option for Crohn’s disease. Recent work in ulcerative colitis (UC) demonstrated the JAK inhibitor tofacitinib exerts distinct effects on macrophages dependent on their cytokine environment. Tofacitinib more effectively suppressed inflammation in IFNγ-exposed macrophages, mirroring clinical observations that patients with an IFN/JAK-STAT signature respond favorably to treatment with tofacitinib. In contrast, the JAK inhibitor was not as effective in patients with high baseline IL-10 activity, hypothesizing that blockade of the anti-inflammatory cytokine IL-10 removes regulation of the pro-inflammatory response (20). Together, this study and our findings highlight macrophage polarization states as a promising precision medicine tool to guide IBD therapy selection. They also suggest potential for combination strategies that modulate macrophage polarization—through additional drugs or cytokines to steer macrophages and optimize response to our existing treatments.
This work pioneers the generation of an in vitro defined macrophage polarization atlas characterized by transcriptomes and applying them to better characterize macrophage populations in Crohn’s disease. Prior investigations of macrophages in Crohn’s disease relied on markers of ‘inflammatory’ M1 or ‘anti-inflammatory’ M2 macrophages. The present study is an initial proof of concept that single-cell RNA sequencing data of in vitro cytokine-specific polarization of macrophages can inform in vivo macrophage polarization states with potential clinical impact. The approach provides a robust characterization of macrophage states that identifies the key cytokines shaping the macrophage population in Crohn’s disease. These results highlight opportunities to possibly treat Crohn’s disease by altering the inflamed microenvironment, inhibiting the formation of pathogenic proinflammatory macrophages and shifting the population to immune suppressing anti-inflammatory macrophages. This work also identifies the need for a larger atlas of macrophage states, to include multiple donors to understand and account for inter-donor variation and intra-donor heterogeneity. This atlas can begin with transcriptomes and extend to cell type-specific functions – to advance our understanding of disease pathogenesis and inform treatment.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/geo/, GSE297735.
Ethics statement
Human blood was obtained from a deidentified donor from the UCLA CFAR Centralized Laboratory Support Core in accordance with UCLA IRB 11-000443. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from The CFAR Virology Core maintains Institutional Review Board approval (UCLA IRB 11-000443) to obtain buffy coat blood products, or Trima filters, from the UCLA Blood and Platelet Center. The UCLA Blood and Platelet Center provides Trima filters from platelet units drawn from individuals who have consented to donate platelets to the UCLA Blood and Platelet Center and have also consented to donate their blood cells for research purposes. The UCLA Virology Core also maintains IRB approval to obtain fresh peripheral blood from a pool of healthy donors recruited by the Core. A separate consent form is maintained for those donors. Written informed consent for participation was not required from the participants or the participants' legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
SL: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Supervision, Visualization, Writing – original draft, Writing – review & editing. ML: Formal Analysis, Visualization, Writing – review & editing, Investigation, Methodology, Writing – original draft. NY: Investigation, Writing – review & editing. AH: Funding acquisition, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. SCL was supported by NIDDK T32DK007180. AH was supported by NIH R01AI173214.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1707719/full#supplementary-material
Supplementary Figure 1 | M1 and M2 macrophage marker gene expression in polarized unstimulated macrophages. (A) UMAP of PBMC-derived day 6 macrophages polarized for 24 hours without acute stimulation. The samples are colored by polarization condition with naïve in green, IL-13 in yellow, IL-10 in orange, IFNγ in blue, IFNβ in purple and IL-4 in red. (This figure is the same as Figure 2A but shown here for reference). (B–H) Expression of M1 marker genes amongst the unstimulated in vitro polarized macrophages. (I–L) Expression of M2 marker genes amongst the unstimulated in vitro polarized macrophages.
Supplementary Figure 2 | Applying the data driven model of macrophage polarization states to colonic macrophages from Crohn’s patients. (A) UMAP of colonic immune cells from healthy donors and Crohn’s patients, from both noninflamed and inflamed regions. Immune cells were separated from stromal and epithelial cells based on annotations from the original study (Kong et al) and clustered in an unsupervised manner. The numbers delineate each individual cluster. (B) Immune cell types manually annotated based on the top 20 differentially expressed genes for each unsupervised cluster, with some clusters consolidated into broader cell type categories. The macrophage clusters are highlighted in pink. (C) Fraction of macrophages among immune cells from Crohns vs healthy donors in the colon vs terminal ileum. (D) Unsupervised clustering of the macrophage subset, separated by donor type. Cluster 1 is marked in blue, cluster 2 in yellow, and cluster 3 in red. (E) Distribution of macrophages assigned to each of the three clusters, separated by donor type. (F) In-vitro-polarized macrophages integrated with the colonic macrophage dataset using SCTransform anchors and mapped onto a shared UMAP, labeled by polarization condition. (G) Assignment of in-vitro-polarized macrophages to in vivo clusters using a weighted mutual nearest neighbor approach. (H) Proportion of cells from each polarization condition assigned to each in vivo macrophage cluster by mutual nearest-neighbor classification. (I) Assignment of polarization states to macrophages found in vivo which were grouped by unsupervised clustering (Supplementary Figure S2C) and are predominantly associated with noninflamed (cluster 1) and healthy (cluster 2). None were assigned to cluster 3.
References
1. Mosser DM and Edwards JP. Exploring the full spectrum of macrophage activation. Nat Rev Immunol. (2008) 8:958–69. doi: 10.1038/nri2448
2. Wynn TA, Chawla A, and Pollard JW. Macrophage biology in development, homeostasis and disease. Nature. (2013) 496:445–55. doi: 10.1038/nature12034
3. Murray PJ. Macrophage polarization. Annu Rev Physiol. (2017) 79:541–66. doi: 10.1146/annurev-physiol-022516-034339
4. Bain CC and Mowat AM. Macrophages in intestinal homeostasis and inflammation. Immunol Rev. (2014) 260:102–17. doi: 10.1111/imr.12192
5. Singh A, Sen S, Iter M, Adelaja A, Luecke S, Guo X, et al. Stimulus-response signaling dynamics characterize macrophage polarization states. Cell Syst. (2024) 15:563–577.e6. doi: 10.1016/j.cels.2024.05.002
6. Xue J, Schmidt SV, Sander J, Draffehn A, Krebs W, Quester I, et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity. (2014) 40:274–88. doi: 10.1016/j.immuni.2014.01.006
7. Hourani T, Perez-Gonzalez A, Khoshmanesh K, Luwor R, Achuthan AA, Baratchi S, et al. Label-free macrophage phenotype classification using machine learning methods. Sci Rep. (2023) 13:5202. doi: 10.1038/s41598-023-32158-7
8. Yona S, Kim K-W, Wolf Y, Mildner A, Varol D, Breker M, et al. Fate mapping reveals origins and dynamics of monocytes and tissue macrophages under homeostasis. Immunity. (2013) 38:79–91. doi: 10.1016/j.immuni.2012.12.001
9. Jaensson E, Uronen-Hansson H, Pabst O, Eksteen B, Tian J, Coombes JL, et al. Small intestinal CD103+ dendritic cells display unique functional properties that are conserved between mice and humans. J Exp Med. (2008) 205:2139–49. doi: 10.1084/jem.20080414
10. Ogino T, Nishimura J, Barman S, Kayama H, Uematsu S, Okuzaki D, et al. Increased Th17-inducing activity of CD14+ CD163 low myeloid cells in intestinal lamina propria of patients with Crohn’s disease. Gastroenterology. (2013) 145:1380–1391.e1. doi: 10.1053/j.gastro.2013.08.049
11. Chapuy L, Bsat M, Sarkizova S, Rubio M, Therrien A, Wassef E, et al. Two distinct colonic CD14+ subsets characterized by single-cell RNA profiling in Crohn’s disease. Mucosal Immunol. (2019) 12:703–19. doi: 10.1038/s41385-018-0126-0
12. Kong L, Pokatayev V, Lefkovith A, Carter GT, Creasey EA, Krishna C, et al. The landscape of immune dysregulation in Crohn’s disease revealed through single-cell transcriptomic profiling in the ileum and colon. Immunity. (2023) 56:444–458.e5. doi: 10.1016/j.immuni.2023.01.002
13. Hafemeister C and Satija R. Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. (2019) 20:296. doi: 10.1186/s13059-019-1874-1
14. Stuart T, Butler A, Hoffman P, Hafemeister C, Papalexi E, Mauck WM, et al. Comprehensive integration of single-cell data. Cell. (2019) 177:1888–1902.e21. doi: 10.1016/j.cell.2019.05.031
15. Sheu KM, Guru AA, and Hoffmann A. Quantifying stimulus-response specificity to probe the functional state of macrophages. Cell Syst. (2023) 14:180–195.e5. doi: 10.1016/j.cels.2022.12.012
16. Fuss IJ, Neurath M, Boirivant M, Klein JS, de la Motte C, Strong SA, et al. Disparate CD4+ lamina propria (LP) lymphokine secretion profiles in inflammatory bowel disease. Crohn’s disease LP cells manifest increased secretion of IFN-gamma, whereas ulcerative colitis LP cells manifest increased secretion of IL-5. J Immunol Baltim Md. (1996) 1950:157. doi: 10.4049/jimmunol.157.3.1261
17. Ito R, Shin-Ya M, Kishida T, Urano A, Takada R, Sakagami J, et al. Interferon-gamma is causatively involved in experimental inflammatory bowel disease in mice. Clin Exp Immunol. (2006) 146:330–8. doi: 10.1111/j.1365-2249.2006.03214.x
18. Luque-Martin R, Angell DC, Kalxdorf M, Bernard S, Thompson W, Eberl HC, et al. IFN-γ Drives human monocyte differentiation into highly proinflammatory macrophages that resemble a phenotype relevant to psoriasis. J Immunol Baltim Md. (2021) 207:555–68. doi: 10.4049/jimmunol.2001310
19. Kang K, Bachu M, Park SH, Kang K, Bae S, Park-Min K-H, et al. IFN-γ selectively suppresses a subset of TLR4-activated genes and enhancers to potentiate macrophage activation. Nat Commun. (2019) 10:3320. doi: 10.1038/s41467-019-11147-3
Keywords: macrophage polarization, Crohn’s disease, inflammatory bowel disease, scRNAseq, macrophages, mucosal immunology
Citation: Lowe SC, Leitman M, Yan N and Hoffmann A (2025) A data-driven model of macrophage polarization states reveals an IFN macrophage signature in active Crohn’s disease. Front. Immunol. 16:1707719. doi: 10.3389/fimmu.2025.1707719
Received: 17 September 2025; Accepted: 18 November 2025; Revised: 11 November 2025;
Published: 04 December 2025.
Edited by:
Steven O’Reilly, Consultant, Sunderland, United KingdomReviewed by:
Wataru Fujii, Dokkyo Medical University, JapanAndrew Yung Fong Li Yim, Amsterdam University Medical Center (UMC), Netherlands
Copyright © 2025 Lowe, Leitman, Yan and Hoffmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sarina C. Lowe, c2Nsb3dlQG1lZG5ldC51Y2xhLmVkdQ==; Alexander Hoffmann, YWhvZmZtYW5uQGcudWNsYS5lZHU=
†These authors share first authorship