 Peter Møller
Peter Møller Marie B. Lund
Marie B. Lund Andreas Schramm
Andreas Schramm- 1Section for Microbiology, Department of Bioscience, Aarhus University, Aarhus, Denmark
- 2Aarhus Institute of Advanced Studies, Aarhus University, Aarhus, Denmark
Nephridial (excretory organ) symbionts are widespread in lumbricid earthworms and the complexity of the nephridial symbiont communities varies greatly between earthworm species. The two most common symbionts are the well-described Verminephrobacter and less well-known Flexibacter-like bacteria. Verminephrobacter are present in almost all lumbricid earthworms, they are species-specific, vertically transmitted, and have presumably been associated with their hosts since the origin of lumbricids. Flexibacter-like symbionts have been reported from about half the investigated earthworms; they are also vertically transmitted. To investigate the evolution of this tri-partite symbiosis, phylogenies for 18 lumbricid earthworm species were constructed based on two mitochondrial genes, NADH dehydrogenase subunit 2 (ND2) and cytochrome c oxidase subunit I (COI), and compared to their symbiont phylogenies based on RNA polymerase subunit B (rpoB) and 16S rRNA genes. The two nephridial symbionts showed markedly different evolutionary histories with their hosts. For Verminephrobacter, clear signs of long-term host-symbiont co-evolution with rare host switching events confirmed its ancient association with lumbricid earthworms, likely dating back to their last common ancestor about 100 million years (MY) ago. In contrast, phylogenies for the Flexibacter-like symbionts suggested an ability to switch to new hosts, to which they adapted and subsequently became species-specific. Putative co-speciation events were only observed with closely related host species; on that basis, this secondary symbiosis was estimated to be minimum 45 MY old. Based on the monophyletic clustering of the Flexibacter-like symbionts, the low 16S rRNA gene sequence similarity to the nearest described species (<92%) and environmental sequences (<94.2%), and the specific habitat in the earthworm nephridia, we propose a new candidate genus for this group, Candidatus Nephrothrix.
Introduction
Earthworms of the family Lumbricidae have long been known to harbor extracellular symbiotic bacteria in their nephridia (Knop, 1926). The nephridia are the worm's excretory system involved in excretion of nitrogenous waste and osmoregulation (Edwards and Bohlen, 1996). They are found in pairs in every segment, and each nephridium is coiled into three loops, where the symbionts reside in the ampulla part of the second loop (Figure 1A). Complexity and composition of the symbiont communities vary between worm species; few species host exclusively the betaproteobacterial Verminephrobacter, while most lumbricids have mixed nephridial communities (Davidson et al., 2010, 2013; Lund et al., 2010a). Verminephrobacter have been studied in detail during the past decade (for a recent review see Lund et al., 2014); these symbionts are species-specific (Schramm et al., 2003; Lund et al., 2010a), beneficial (Lund et al., 2010b), vertically transmitted via the cocoon (Davidson and Stahl, 2006, 2008), and presumably co-evolved with their host (Lund et al., 2010a).
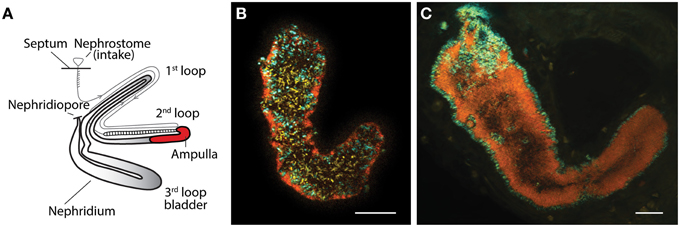
Figure 1. (A) Schematic outline of the structure of a single nephridium showing the three major loops and the ampulla (red) where the symbionts reside (modified from Schramm et al., 2003). (B,C) FISH images of ampullas from two earthworm species after triple hybridization with the general bacterial probe EUB338,-II,-III (yellow), a Verminephrobacter-specific probe LSB145 (cyan/green), and a probe specific for Flexibacter-like symbionts FLX-226 (red/orange). (B) Al. chlorotica showing the Flexibacter-like symbionts in close proximity to the ampulla epithelium and Verminephrobacter in the lumen. Notice the large amount of unidentified bacteria in the lumen. (C) E. andrei with Verminephrobacter lining the epithelium and the Flexibacter-like symbionts in the lumen. Scale bar is 20 μm.
In contrast, little is known about the other nephridial symbionts that represent at least 27 different taxa from six phyla and occur inconsistently across earthworm species (Davidson et al., 2013). Most information is available for a Flexibacter-like symbiont (phylum Bacteroidetes), which was first discovered in the compost worm Eisenia fetida and shown to be vertically transmitted via the cocoon, together with Verminephrobacter and a member of the Microbacteriaceae (Davidson et al., 2010). The Flexibacter-like symbionts even seem to colonize the ampulla of developing embryos before the arrival of Verminephrobacter (Davidson et al., 2010; Dulla et al., 2012). In a global survey, Flexibacter-like 16S rRNA gene sequences were detected in about half of the lumbricid earthworms analyzed and in four other earthworm families (Davidson et al., 2013); since sequences from the same host families grouped together, the authors suggested host specificity of the Flexibacter-like symbionts. None of the other detected symbionts appeared to be widely present in earthworms (Davidson et al., 2013).
Based on their presence in multiple earthworm species, putative host specificity (Davidson et al., 2013), and vertical transmission (Davidson et al., 2010), we hypothesized that Flexibacter-like symbionts had a long evolutionary history with their earthworm hosts and with Verminephrobacter. The objectives of this study were (i) to investigate the putative host specificity of Flexibacter-like symbionts in more detail, and (ii) to elucidate the evolution of the tripartite symbiosis between earthworms, Verminephrobacter, and Flexibacter-like symbionts.
Material and Methods
Collection and Preparation of Samples
A total of 18 earthworm species, 1–16 individuals of each (Supplementary Table S1), were collected in Denmark and Germany and identified by analytical keys (Sims and Gerard, 1985; Andersen, 1997). Specimens were cleaned with water, killed in 50% ethanol, and cut open along the dorsal line. Under a dissection microscope, the gut was carefully removed without disrupting it. For nucleic acid extraction, nephridia were collected by forceps from the anterior half of the gut-free worm and stored at −20°C. For fluorescence in situ hybridization (FISH), the gut-free body wall with attached nephridia was cut into smaller pieces (1–3 cm), fixed in 4% (w/v) paraformaldehyde for 2–3 h, washed twice in 1 × phosphate buffered saline (PBS), and stored in 70% ethanol/PBS at −20°C.
Fluorescence In Situ Hybridization (FISH)
The localization of Verminephrobacter and Flexibacter-like bacteria in the nephridia was confirmed in two earthworm species; Eisenia andrei and Allolobophora chlorotica, using FISH. Hybridization was done at 35% formamide for 2.5 h directly on fixed pieces of body wall. The probes: LSB145-CY5 (Schweitzer et al., 2001), targeting some Acidovorax spp. and all Verminephrobacter spp., FLX226-CY3 (this study), targeting all Flexibacter-like symbionts, and EUB338,-II,-III-FAM (Daims et al., 1999), targeting all bacteria, were used in combination (Supplementary Table S1). The optimal formamide concentration of 35% for the probe FLX226 was determined using mathFISH (Yilmaz et al., 2011). The hybridizations were carried out according to published protocols (Pernthaler et al., 2001). Following hybridization and washing, the samples were counterstained with 4′,6-diamidino-2-phenylindole (DAPI; 1 μg mL−1) for 10 min on ice, rinsed thoroughly with water, and immersed in a 3:1 mixture of Vectashield (Vector Laboratories, Inc., Burlingame, CA) and Citifluor (Citifluor Ltd., London, UK). Single nephridia were dissected from the body wall and mounted on a glass slide prior to epifluorescence microscopy. Images were captured on an Axio Vert 200M epifluorescent light microscope, fitted with an Apotome for optical sectioning and an AxioCam MRm camera, all controlled by the AxioVision software (v4.8.1.0— all by Carl Zeiss, Jena, Germany).
Nucleic Acid Extraction, PCR, Cloning and Sequencing
DNA was extracted from the dissected nephridia using the DNeasy Blood & Tissue kit (Qiagen, Valencia, CA, USA) following the protocol for animal tissue. DNA extracts were used for PCR amplification of both host and symbiont genes. All PCR reactions were run with the Taq master mix (VWR, Herlev, Denmark) containing 1.5 mM MgCl2, 0.2 mM of each dNTP, 0.1 units/μL VWR Taq polymerase, inert red dye, and 0.2 μM of each primer. Primers and thermal cycling protocols are summarized in Supplementary Table S2.
Two mitochondrially encoded genes, NADH dehydrogenase subunit II (ND2) and cytochrome c oxidase subunit I (COI), were targeted. These host genes are located adjacent to each other on the mitochondrial chromosome and were therefore amplified as one long fragment (ND2-COI) using the newly designed primers Lum-ND2-322F or Lum-ND2-370F, and Lum-COI-723R. Some COI sequences had been obtained in a previous study, in which cases only ND2 was amplified using the reverse primer Lum-COI-14R (Lund et al., 2010a, Supplementary Table S2). PCR fragments were purified (GenElute™ PCR Clean-Up kit, Sigma) and directly sequenced by Sanger sequencing (Macrogen, Korea) using the same primers as in the PCR reaction.
Earthworms were screened for the presence of Flexibacter-like symbiont 16S rRNA genes by specific PCR yielding a 174 bp-fragment (primers Flexi-145F and CF319aR, Supplementary Table S2). From the samples that tested positive, a longer fragment (primers Flexi-145F and 1492R, 1347 bp, Supplementary Table S2) was amplified, cloned, and sequenced. To obtain higher resolution in the phylogenetic trees, primers were designed for rpoB from Verminephrobacter (referred to as VrpoB; primers VrpoB-43F and VrpoB-1430R) and Flexibacter-like symbionts (referred to as FrpoB; primers FrpoB-1905F and FrpoB3213R, Supplementary Table S2). Primers were designed based on available rpoB sequences from the closest related organisms (Flexibacter flexilis for FrpoB and V. eiseniae EF2-01 and V. aporrectodeae At4 for VrpoB). rpoB was amplified, cloned using pGEM®-T easy vector system (Promega™) or TOPO® TA Cloning kit (Invitrogen™), and sequenced at GATC (Germany).
All obtained sequences have been submitted to GenBank under accession numbers KP420532-KP420720 and KM058238-KM058572 (Supplementary Table S1).
Phylogenetic and Statistical Analyses
The newly obtained Flexibacter-like 16S rRNA gene sequences were aligned using the on-line SINA aligner (Pruesse et al., 2012) and imported into an existing ARB database (SSURef_111_SILVA_NR, (Ludwig et al., 2004; Pruesse et al., 2007). The ARB database was updated by identifying the closest relatives of the Flexibacter-like 16S rRNA gene sequences in GenBank using BLAST and importing them to the database if not already present. All published Verminephrobacter 16S rRNA gene sequences were also imported into the database. For constructing Flexibacter-like and Verminephrobacter 16S rRNA gene trees, a broad selection of related sequences from cultured organisms and environmental samples was included and the trees were calculated by Bayesian Inference using the software MrBayes (v3.2.3) (Ronquist et al., 2012). The substitution model was set to GTR + I + Γ and 60% majority rule consensus trees were calculated based on four simultaneous runs with six chains each and a sample frequency of 500. After 2,000,000 generations the potential scale reduction factors (PSRF) showed 1.000 indicating stationarity (Ronquist et al., 2012). The first 25% of the data was discarded as burn-in (Figure 2, Supplemental Figure S1).
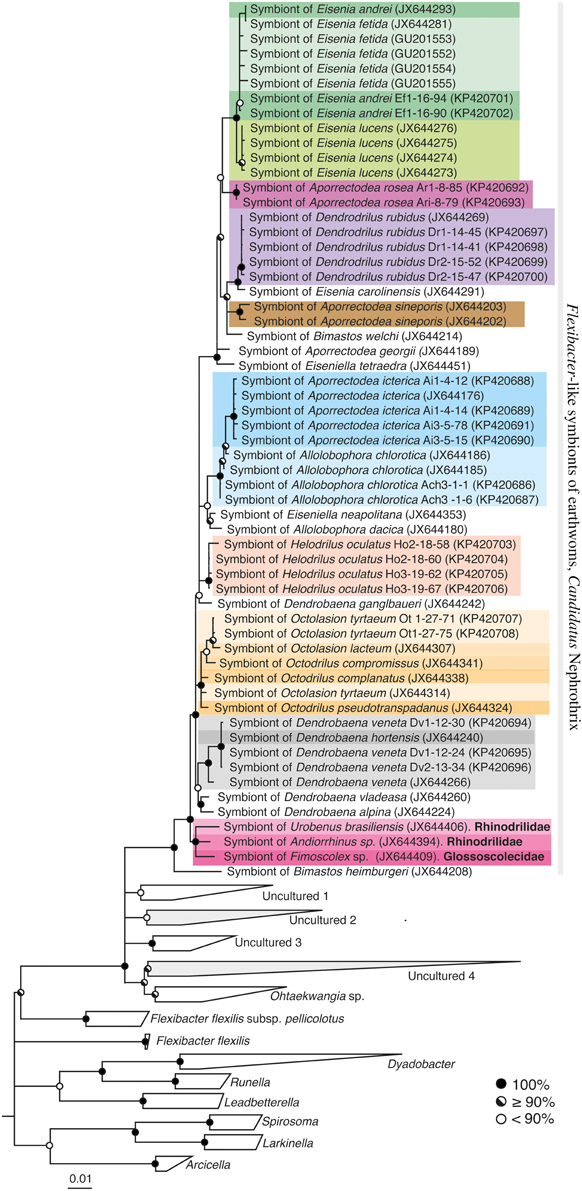
Figure 2. 16S rRNA gene sequence phylogeny of Flexibacter-like symbionts of earthworms and close relatives within the Cytophagaceae (Bacteroidetes). 60% majority rule consensus tree from Bayesian Inference. Circles on nodes show posterior probabilities according to legend. The Flexibacter-like symbionts of earthworms form a monophyletic group. If nothing else is indicated the earthworm species belongs to the family Lumbricidae. Gray clades contain other sequences detected in the nephridia of non-lumbricid earthworm families (Davidson et al., 2013): “Uncultured 2” contains three sequences from Megascolecidae species; Arctiostrotus sp. (JX644412), Terisswalkerius covacevihae (JX644411), T. erici (JX644410). “Uncultured 4” contains sequences from Andiorrhinus sp. (Pontoscolecidae, JX644395).
The host (ND2 and COI) and rpoB sequences were assembled and aligned in Geneious v. 5.6.7 (Biomatters LTD. Drummond et al., 2011) using the translation alignment algorithm for coding sequences. To determine an appropriate model of evolutionary nucleotide substitution for the individual target genes, the alignments where analyzed in jModelTest (Guindon and Gascuel, 2003; v2.1.5—Darriba et al., 2012). For each of the four target genes, the appropriate model was chosen based on the Akaike Information Criterion (AIC) (Akaike, 1974). The best fit substitution models were GTR + I + Γ for ND2, HKY + I + Γ for COI, GTR + Γ for VrpoB, and GTR + I + Γ for FrpoB. Phylogenetic trees were calculated by Bayesian Inference using MrBayes (v3.2.3) (Ronquist et al., 2012). The host genes (ND2 and COI) were concatenated and divided into six partitions; one for each codon position in both genes. The three partitions from each gene were set to evolve under the substitution models estimated with jModelTest. Both rpoB genes, VrpoB and FrpoB, were divided into three partitions (one for each codon position) and substitution models were chosen according to jModelTest. Majority rule consensus trees were calculated, based on two simultaneous runs with four chains each, and a sample frequency of 500. After 2,000,000 generations the PSRF showed 1.000 indicating stationarity (Ronquist et al., 2012). The first 500,000 generations were discarded as burn-in and the remainder were used to calculate 60% majority rule consensus trees (Figure 3).
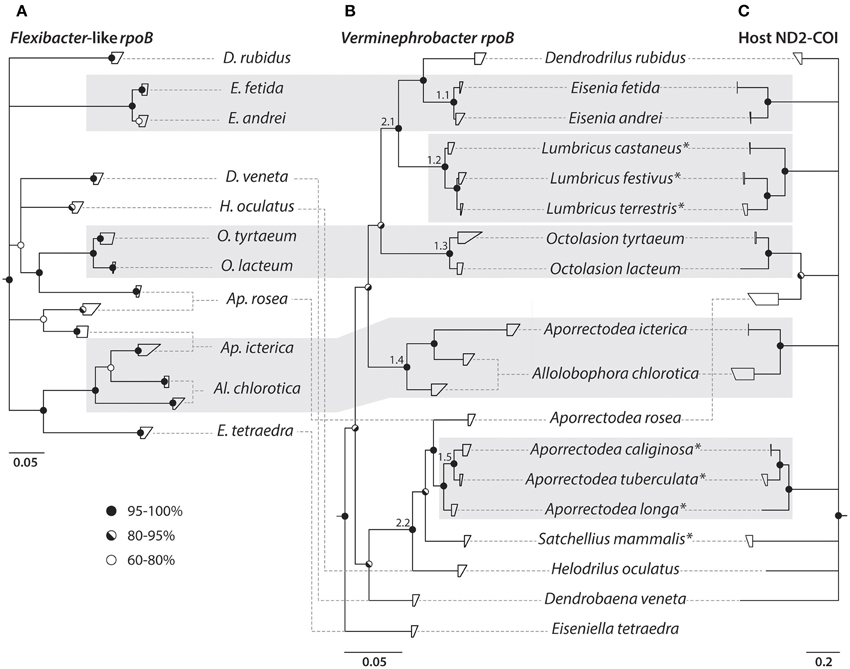
Figure 3. Phylogenetic trees of earthworms and their Verminephrobacter and Flexibacter-like symbionts. Nucleotide based rpoB phylogenies of (A) Flexibacter-like symbionts and (B) Verminephrobacter. (C) Earthworm phylogeny based on ND2-COI nucleotide sequences. Clades showing signs of co-diversification are highlighted by shaded boxes. All trees are majority rules (60%) consensus trees from Bayesian inference. Circles on nodes show posterior probabilities according to legend. Host species that do not harbor the Flexibacter-like symbiont are marked by an asterisk. Small numbers on branches in (B) are discussed in the text.
A regular Mantel test was used to examine how well the host and symbiont phylogenies correspond to each other by testing the correlation between host (ND2-COI) and the corresponding symbiont rpoB distance matrices (host vs. FrpoB and host vs. VrpoB). For constructing the distance matrices highly similar sequences were grouped following the terminal nodes in Figure 3 and the between-group mean distance was calculated in MEGA6 (Tamura et al., 2013). Both Mantel tests were performed in R (R Core Team, 2013) as implemented in the package vegan (Oksanen et al., 2015) using the Pearson correlation method with 1000 permutations.
Results
Fluorescence In Situ Hybridization (FISH)
The presence of Verminephrobacter and Flexibacter-like bacteria was confirmed in Al. chlorotica and E. andrei (Figures 1B,C, respectively). In Al. chlorotica the Flexibacter-like symbionts line the lumen wall and Verminephrobacter is sparsely scattered throughout the lumen. The lumen is dominated by unidentified bacteria targeted by the general bacterial probe (EUB338,-II,-III). In E. andrei the Verminephrobacter lines the lumen wall, as typically found in other earthworm species (Davidson et al., 2010; Lund et al., 2014), and the Flexibacter-like symbionts fill up the lumen like cotton wool where individual cells are indistinguishable.
Phylogeny of Flexibacter-like Earthworm Symbionts
Flexibacter-like symbionts were detected in 11 of the 18 earthworm species investigated in this study; thus in total, 26 of the 51 lumbricid earthworm species ever analyzed scored positive for Flexibacter-like symbionts (Table 1). The 16S rRNA gene sequences of Flexibacter-like symbionts were also found in two sister groups to the Lumbricidae; one Glossoscolecidae (Fimoscolex sp.) and two Pontoscolecidae (Andiorrhinus sp. and Urobenus brasiliensis) (Davidson et al., 2013). The phylogeny of the Flexibacter-like symbionts was constructed using both 16S rRNA gene sequences (Figure 2) and rpoB sequences (Figure 3A). The 16S rRNA gene sequences were monophyletic within the Cytophagaceae and had low sequence similarities (90.6–92%) to the closest cultured relative; Ohtaekwangia koreensis. They were 91.5–94.2% similar to the closest environmental sequence identified by BLAST. The low intragroup genetic distance of only 4.8% makes the variation within the Flexibacter-like 16S rRNA gene sequences too low to reliably reconstruct the symbiont phylogeny.

Table 1. Summary of earthworm species that have been investigated in this and previous studies for the presence/absence of Verminephrobacter and Ca. Nephrothrix.
The rpoB-based tree has low resolution of the basal branches but the Flexibacter-like symbionts from different host species can clearly be distinguished and are indeed species-specific (Figure 3A). However, Aporrectodea rosea, Al. chlorotica, and Ap. icterica each carry two distinct symbiont types that are not monophyletic with each other (Figures 3A, 4A). The Flexibacter-like symbionts of Ap. rosea form one group clustering with Octolasion and the other with Ap. icterica. For the Ap. icterica symbionts, one group clusters with Ap. rosea and the other with Al. chlorotica. In Al. chlorotica both groups cluster with Ap. icterica, but together they are not monophyletic. In both Ap. icterica and Al. chlorotica the two variants of Flexibacter-like symbionts coexist in the same individual worms (Figure 4A); this was found in two individuals of each species. For Ap. rosea, however, all of the sequences from one of the symbiont clusters originate from worms of the same geographic origin (Figure 4A).
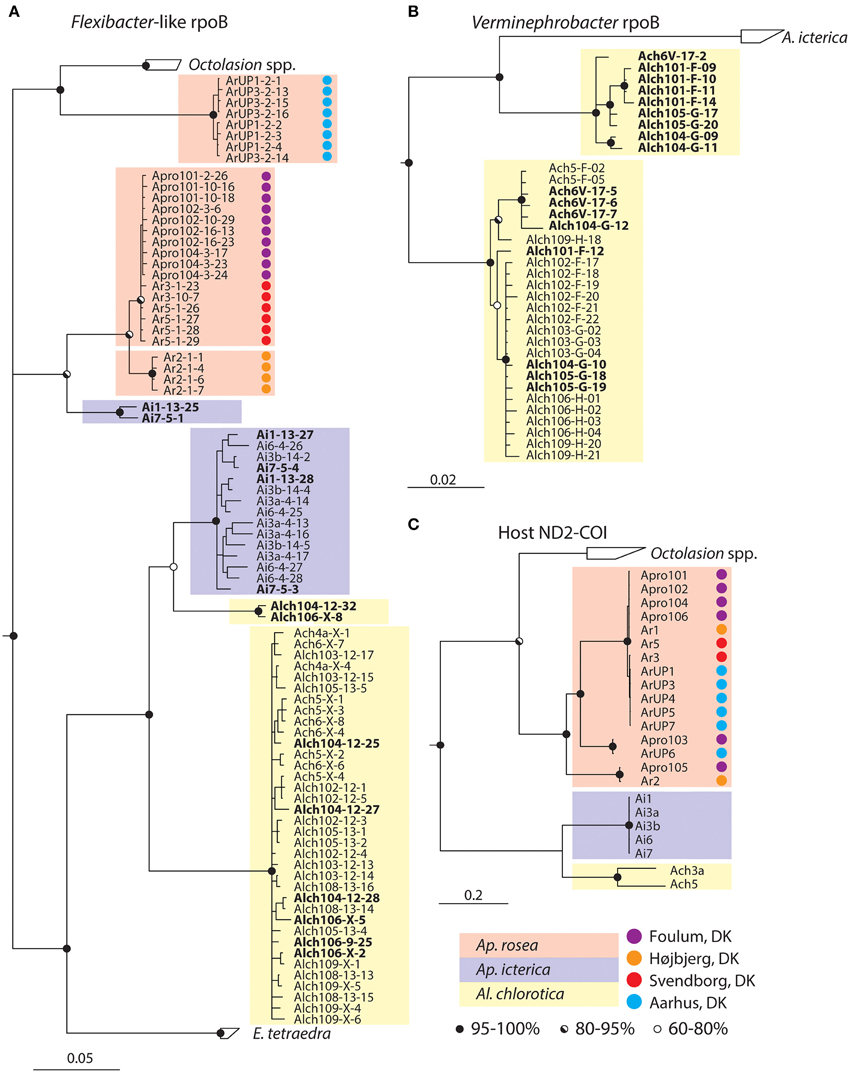
Figure 4. Details of selected clades from trees in Figure 3. (A) Flexibacter-like symbionts, (B) Verminephrobacter, and (C) earthworms. Different earthworm species are highlighted with colored boxes according to legend. Names in bold highlight clones from the same individual host that are associated with two different clades. The first part of the clone name indicates host individual. Colored circles denote the geographic origin of host individuals according to legend. All trees are majority rule (60%) consensus trees from Bayesian inference. Circles on nodes are posterior probabilities according to legend.
Phylogeny of Verminephrobacter Symbionts
Two separate Verminephrobacter phylogenies were constructed using rpoB (Figure 3B) and 16S rRNA gene sequences (Supplementary Figure S1). All 16S rRNA gene sequences were retrieved from GenBank. The rpoB-based phylogeny has a better resolution of closely related strains, like Eisenia sp. and Lumbricus sp., which cannot be resolved using 16S rRNA gene sequences (Supplementary Figure S1). However, the rpoB-based tree has low posterior probabilities (down to 80%) toward the basal nodes of the tree (Figure 3B). There are large discrepancies in the basal branching pattern between the two phylogenies, but the terminal clades labeled 1.1–1.5 are found in both trees. The nodes labeled 2.1 and 2.2 are also found in both trees but the branching patterns within these two clades differ in the two phylogenies.
Comparison of Host and Symbiont Phylogenies
The earthworm phylogeny is based on concatenated ND2 and COI sequences (Figure 3C). Even though the combined PCR fragment is about 1400 bp, only 921 bp of high quality sequence could be retrieved after sequencing. Five clusters could be resolved with high (94–100%) posterior probabilities but the basal branching of the tree was not resolved, which is a common problem in earthworm phylogenies (reviewed by Chang and James, 2011). It was not possible to amplify ND2-COI from Eiseniella tetraedra.
All of the resolved groups in the earthworm phylogeny are reflected in the Verminephrobacter rpoB and 16S rRNA gene trees, with A. rosea as an obvious exception (Figures 3B,C, Supplementary Figure S1). As for the Flexibacter-like symbionts, the Verminephrobacter symbionts in Al. chlorotica formed two distinct, coexisting clades, one of which grouped with the symbionts of Ap. icterica (Figures 3B, 4B). The Mantel test showed a strong positive relationship (r = 0.5065; P < 0.001) between host and Verminephrobacter genetic distances based on ND2-COI and rpoB, respectively.
Due to low resolution of both the host and the rpoB-based tree of Flexibacter-like symbionts it is not possible to evaluate the degree of congruency between the two (Figures 3A,C). However, for closely related earthworm species there are signs of co-speciation, i.e., Eisenia fetida and E. andrei (and E. lucens, Figure 1), Octolasion tyrtaeum and O. lacteum. Also, leaving aside the second symbiont clades (see above), the Flexibacter-like symbionts, Verminephrobacter, and the hosts, all have the same clustering of Al. chlorotica and Ap. icterica (Figure 3). These signs of co-speciation are also reflected in the Mantel test which showed a moderate positive relationship (r = 0.3753; P < 0.01) between host and Flexibacter-like genetic distances based on ND2-COI and rpoB, respectively. Meanwhile, there are clear signs of host switching in the three worm species all having two distinct Flexibacter-like symbiont groups (Ap. rosea, Ap. icterica, and Al. chlorotica, Figure 3A).
When comparing the rpoB-based phylogenies of Verminephrobacter and the Flexibacter-like symbionts (Figures 3A,B) there are clear signs of incongruences in the deeper branching patterns.
Discussion
Flexibacter-like Symbiont Phylogeny and Proposal of the Candidate Genus Nephrothrix
The Flexibacter-like symbionts of lumbricid earthworms clearly form their own monophyletic cluster based both on 16S rRNA (Figure 2) and rpoB (Figure 3A) gene sequences. The monophyletic group is very coherent and distantly related to other known organisms; based on 16S rRNA gene sequences they have an intragroup genetic distance of only 4.8% and a low similarity (91.5–94.2%) to other known sequences. Flexibacter-like symbionts are widespread in the Lumbricidae, where they are present in about 50% of the investigated worms (Table 1; Davidson et al., 2013); absence was confirmed in eight species using specific PCR (Table 1). The Flexibacter-like symbionts were also found in one member of the Glossoscolecidae and two of Pontoscolecidae (Table 1, Davidson et al., 2013), both sister families to Lumbricidae (James and Davidson, 2012). These Flexibacter-like symbionts of non-lumbricid worms form their own monophyletic clade toward the base of the group (the most basal sequence is from Bimastos heimburgeri, Lumbricidae, Figure 2). The presence of Flexibacter-like symbionts in the nephridia of these worms has not been confirmed with FISH. Davidson et al. (2013) also reported the presence of Flexibacter-like bacteria in other earthworm families; however, their 16S rRNA gene sequences group outside the above described monophyletic cluster (Figure 2). Apparently, they represent different evolutionary lineages and are therefore not considered part of the novel symbiont genus proposed below (Figure 2).
For the monophyletic Flexibacter-like symbionts of lumbricid earthworms, the low similarity (91.5–94.2%) to other known 16S rRNA gene sequences, combined with their unique habitat in earthworm nephridia and their conspicuous morphology, supports the establishment of a novel genus. In spite of numerous attempts, the symbionts could not be cultivated yet (Marie Lund, unpublished; Flávia Viana, personal communication); they can, however, be easily identified in situ by the newly developed FISH probe (FLX226, Supplementary Table S2). In accordance with the Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology (Stackebrandt et al., 2002), we therefore propose the establishment of a candidate genus and the name “Candidatus Nephrothrix” [Ne.phro.thrix. Gr. n. nephros, kidney; Gr. fem. n. thrix, hair; N. L. fem. n. Candidatus Nephrothrix, kidney (associated) hair], which reflects the specific habitat (earthworm nephridia) and the slender, hair-like morphology of these symbionts. As a first representative of the candidate genus, we suggest the specific symbiont of Eisenia fetida to be named “Ca. N. davidsonii” in honor of Seana Davidson, the scientist who first discovered the Flexibacter-like symbionts in earthworms.
Ca. Nephrothrix Switches Earthworm Host
It can not be concluded if Ca. Nephrothrix is an ancient co-evolving symbiont of earthworms due to the poor basal resolution of both phylogenies (Figures 3A,C). There are some indications of co-evolution (Figures 3A,C), which is supported by the Mantel test showing a moderate positive correlation between the Ca. Nephrothrix and host distance matrices. However, there are also clear examples of host switching where distinct Ca. Nephrothrix types are found in the same host species (Figure 3A). Also, the monophyletic group of non-lumbricid derived 16S rRNA gene sequences toward the base of the Ca. Nephrothrix cluster (Figure 2) indicates a host switch because the most basal sequence in the group is from a lumbricid worm. Whether the first host of Ca. Nephrothrix was a lumbricid or belonged to one of the sister families is unknown but given the basal grouping of the sequences from Glossoscolecidae and Pontoscolecidae it is possible that Ca. Nephrothrix originated in a non-lumbricid worm.
After each host switch, Ca. Nephrothrix apparently became species-specific, and during two more recent, separate host speciation events (in Eisenia and in Octolasion) Ca. Nephrothrix possibly co-diversified with its host (Figure 3A, Supplementary Figure S2); in which case Ca. Nephrothrix was already present in the ancestors of Eisenia and Octolasion. Alternatively, Eisenia sp. and Octolasion sp. speciated before colonization and picked up closely related Ca. Nephrothrix at a later point. This would result in the same pattern of apparent co-speciation.
Interestingly, in three earthworm species (Ap. rosea, Ap. icterica, and Al. chlorotica) two distinct Ca. Nephrothrix rpoB variants are found (Figure 4A). In Ap. rosea, all of the sequences in one cluster originate from worms collected at the same site, thus possibly representing a local host-switching event. In Ap. icterica and Al. chlorotica, on the other hand, the two rpoB variants coexist in the same worm individuals. The rpoB gene is generally found to be a single copy gene (Case et al., 2007), and we therefore assume that the gene variants represent two different Ca. Nephrothrix populations. In both cases of co-existing Ca. Nephrothrix populations, the symbionts from one of the populations are much more common than from the other (Figure 4A). When considering the tree topology, it is likely that the dominant populations represent the original Ca. Nephrothrix symbionts, which have co-diversified with their hosts during the speciation event between Ap. icterica and Al. chlorotica. The secondary Ca. Nephrothrix populations may then originate from recent host-switching events.
The co-existing symbiont populations in the same earthworm species or individuals show that Ca. Nephrothrix can switch between earthworm hosts. After each host-switching event, they became species-specific again, indicating that they adapted to the new host environment. Host switching can only occur if Ca. Nephrothrix is sometimes horizontally transmitted, thus the vertical transmission previously demonstrated in E. fetida (Davidson et al., 2010) must be leaky in at least some earthworm species. It is not known if the greatest barrier for host switching is (i) lack of opportunity, i.e., worms rarely encounter non-native symbionts, or (ii) species-specificity where non-native symbionts are excluded because they do not respond correctly to host signals. It is furthermore unknown if the horizontal transmission occurs by direct contact between the different worm species or if there is an environmental reservoir of Ca. Nephrothrix.
Distribution of Verminephrobacter and Co-speciation with its Earthworm Host
Verminephrobacter are widespread in the Lumbricid earthworms, where they are present in about 80% of the investigated species (Table 1, Lund et al., 2010a; Davidson et al., 2013). However, most investigations have been based on bacterial 16S rRNA gene clone libraries rather than Verminephrobacter–specific FISH or PCR assays; thus if the symbionts were not detected it could mean that they were overlooked rather than truly absent. The absence of Verminephrobacter has only been confirmed by specific FISH in three worm species; Dendrobaena octaedra, D. attemsi, and D. byblica (Table 1, Lund et al., 2010a; Davidson et al., 2013). Since Verminephrobacter was generally not detected in Dendrobaena species, it is likely indeed absent from this genus.
Davidson et al. (2013) also reported Verminephrobacter 16S rRNA gene sequences in two worms from the earthworm family Microchaetidae. These two sequences were presented in a tree with one large unresolved group of Acidovorax and Verminephrobacter. In contrast, our analyses clearly established Verminephrobacter from Lumbricidae as monophyletic genus with high confidence (Supplementary Figure S1), and thus confirmed earlier phylogenies (Schramm et al., 2003; Pinel et al., 2008; Lund et al., 2010a). In addition, the two sequences from Microchaetidae clustered with the genus Acidovorax and therefore do not belong within Verminephrobacter.
The poor basal resolution of both host and Verminephrobacter trees (Figures 3B,C, Supplemental Figure S1) prevents a direct proof of co-diversification of the ancient symbiotic partners. However, clear congruence of the five clades that could be resolved in both host and Verminephrobacter trees does support the hypothesis of co-evolution along with the Mantel test showing a strong positive relationship between the host and Verminephrobacter distance matrices. Meanwhile, the Ap. rosea symbiont is a clear example of host switching.
Another putative example of host switching between closely related worms is Verminephrobacter in Al. chlorotica: Interestingly, four Al. chlorotica individuals from two geographic locations (Figure 4C, Supplementary Table S1) have two coexisting rpoB gene variants. These gene variants are likely to originate from two distinct Verminephrobacter populations since rpoB is generally a single copy gene (Case et al., 2007). One of the rpoB variants groups with Ap. icterica and could therefore have originated from this species. This is the only reported example of distinct Verminephrobacter types coexisting in the same host. FISH analysis shows that Verminephrobacter only comprises a small fraction of the nephridial community in Al. chlorotica (Figure 1B, (Lund et al., 2010a)) and that Verminephrobacter is located in the lumen, rather than being associated with the ampulla wall as usual (Figures 1B,C, Davidson et al., 2010; Lund et al., 2014). Potentially, Al. chlorotica is in the process of losing its Verminephrobacter symbiont; signals involved in symbiont specificity may already be weakened, thus allowing the colonization by a non-native Verminephrobacter.
An Ancient and a Young Symbiont
The age of the two symbiont groups can be estimated by comparison with the age of the earthworm hosts. In a recently published Lumbricidae phylogeny, the family was estimated to originate during the Cretaceous about 110 MYA (95% confidence interval; 100–120 MYA) and the split to the most basal worm species with Verminephrobacter (Allolobophora dacia) happened 95 MYA (95% confidence interval 90–100 MYA) (Domínguez et al., 2015). Thus, Verminephrobacter has likely been associated with lumbricid earthworms for about 100 MY. Since Ca. Nephrothrix has apparently switched hosts it is not possible to date the origin of this symbiosis. However, the putative recent co-speciation events with Octolasion sp. and Eisenia sp. facilitate a minimum estimate of the age of the group. According to Domínguez et al. (2015), O. tyrtaeum and O. lacteum diverged during the Paleogene about 40 million years ago (MYA, 95% confidence interval; 15–70 MYA), while Eisenia lucens diverged from E. fetida/E. andrei about 45 MYA (95% confidence interval; 35–55 MYA). Thus, Ca. Nephrothrix has likely been associated with the Lumbricidae for a minimum of 45 MY. Furthermore, the intragroup genetic distance in the 16S rRNA gene in Verminephrobacter (7.3%) is higher than in Ca. Nephrothrix (4.8%), supporting that the genus Verminephrobacter is older than Ca. Nephrothrix.
Conclusion and Perspective
In the tripartite symbiosis between earthworms, Verminephrobacter and Ca. Nephrothrix, the two bacterial partners have followed markedly different evolutionary trajectories. Symbionts from the novel candidate genus, Ca. Nephrothrix, are found in approximately half of all lumbricid earthworm species, they have apparently switched hosts, and subsequently adapted and become species-specific. In contrast, Verminephrobacter show signs of long-term cospeciation with their host, they are present in almost all lumbricid earthworms, and have rare host switching events. Verminephrobacter is thus likely to have originated in the last common ancestor of lumbricid earthworms 100 MYA (Lund et al., 2010a; Domínguez et al., 2015, whereas Ca. Nephrothrix is much younger with an estimated age of ≥45 MY. This scenario still allows for co-occurrence of the two symbionts in a range of earthworm species for at least 45 MY, and thus plenty of opportunity for interspecies genetic exchange as predicted by Kjeldsen et al. (2012) to explain Verminephrobacter genome evolution. In support of this possibility, natural competence and transformation of Verminephrobacter within earthworm cocoons has recently been demonstrated (Davidson et al., 2014). Whether Ca. Nephrothrix has left its genetic trace in Verminephrobacter and vice versa remains to be shown.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We wish to thank Susanne Nielsen for invaluable expert help in the laboratory, and Michael Selorm Avumegah for collecting earthworms. Funding for this research was provided by The Aarhus Institute of Advanced Studies (AIAS) to ML, The Danish Council for Independent Research | Natural Sciences (FNU), and the EU Marie Curie Initial Training Network “SYMBIOMICS.”
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2015.00529/abstract
References
Akaike, H. (1974). A new look at the statistical model identification. IEEE Trans. Autom. control. AC-19, 716–723. doi: 10.1109/TAC.1974.1100705
Case, R. J., Boucher, Y., Dahllöf, I., Holmström, C., Doolittle, W. F., and Kjelleberg, S. (2007). Use of 16S rRNA and rpoB genes as molecular markers for microbial ecology studies. Appl. Environ. Microbiol. 73, 278–288. doi: 10.1128/AEM.01177-06
Chang, C. H., and James, S. (2011). A critique of earthworm molecular phylogenetics. Pedobiologia (Jena). 54, S3–S9. doi: 10.1016/j.pedobi.2011.07.015
Daims, H., Brühl, A., Amann, R., Schleifer, K. H., and Wagner, M. (1999). The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 22, 434–444. doi: 10.1016/S0723-2020(99)80053-8
Darriba, D., Taboada, G. L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9, 772. doi: 10.1038/nmeth.2109
Davidson, S. K., Dulla, G. F., Go, R. A., Stahl, D. A., and Pinel, N. (2014). Earthworm symbiont Verminephrobacter eiseniae mediates natural transformation within host egg capsules using type IV pili. Front. Microbiol. 5:546. doi: 10.3389/fmicb.2014.00546
Davidson, S. K., Powell, R., and James, S. (2013). A global survey of the bacteria within earthworm nephridia. Mol. Phylogenet. Evol. 67, 188–200. doi: 10.1016/j.ympev.2012.12.005
Davidson, S. K., Powell, R. J., and Stahl, D. A. (2010). Transmission of a bacterial consortium in Eisenia fetida egg capsules. Environ. Microbiol. 12, 2277–2288. doi: 10.1111/j.1462-2920.2010.02231.x
Davidson, S. K., and Stahl, D. A. (2008). Selective recruitment of bacteria during embryogenesis of an earthworm. ISME J. 2, 510–518. doi: 10.1038/ismej.2008.16
Davidson, S., and Stahl, D. A. (2006). Transmission of nephridial bacteria of the earthworm Eisenia fetida. Appl. Environ. Microbiol. 72, 769–775. doi: 10.1128/AEM.72.1.769-775.2006
Domínguez, J., Aira, M., Breinholt, J. W., Stojanovic, M., James, S. W., and Pérez-Losada, M. (2015). Underground evolution: new roots for the old tree of lumbricid earthworms. Mol. Phylogenet. Evol. 83, 7–19. doi: 10.1016/j.ympev.2014.10.024
Drummond, A., Ashton, B., Buxton, S., Cheung, M., Cooper, A., Duran, C., et al. (2011). Geneious. Available online at: http://www.geneious.com/
Dulla, G. F. J., Go, R. A., Stahl, D. A., and Davidson, S. K. (2012). Verminephrobacter eiseniae type IV pili and flagella are required to colonize earthworm nephridia. ISME J. 6, 1166–1175. doi: 10.1038/ismej.2011.183
Edwards, C. A., and Bohlen, P. J. (1996). Biology and Ecology of Earthworms. London: Chapman & Hall.
Guindon, S., and Gascuel, O. (2003). A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 52, 696–704. doi: 10.1080/10635150390235520
James, S. W., and Davidson, S. K. (2012). Molecular phylogeny of earthworms (Annelida:Crassiclitellata) based on 28S, 18S and 16S gene sequences. Invert. Systematics. 26, 213. doi: 10.1071/IS11012
Kjeldsen, K. U., Bataillon, T., Pinel, N., De Mita, S., Lund, M. B., Panitz, F., et al. (2012). Purifying selection and molecular adaptation in the genome of Verminephrobacter, the heritable symbiotic bacteria of earthworms. Gen. Biol. Evol. 4, 307–315. doi: 10.1093/gbe/evs014
Knop, J. (1926). Bakterien und bakteroiden bei Oligochaeten. Z. Morph. Ökol Tiere. 6, 588–624. doi: 10.1007/BF00464431
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res. 32, 1363–1371. doi: 10.1093/nar/gkh293
Lund, M. B., Davidson, S. K., Holmstrup, M., James, S., Kjeldsen, K. U., Stahl, D. A., et al. (2010a). Diversity and host specificity of the Verminephrobacter-earthworm symbiosis. Environ. Microbiol. 12, 2142–2151. doi: 10.1111/j.1462-2920.2009.02084.x
Lund, M. B., Holmstrup, M., Lomstein, B. A., Damgaard, C., and Schramm, A. (2010b). Beneficial effect of Verminephrobacter nephridial symbionts on the fitness of the earthworm Aporrectodea tuberculata. Appl. Environ. Microbiol. 76, 4738–4743. doi: 10.1128/AEM.00108-10
Lund, M. B., Kjeldsen, K. U., and Schramm, A. (2014). The earthworm-Verminephrobacter symbiosis: an emerging experimental system to study extracellular symbiosis. Front. Microbiol. 5:128. doi: 10.3389/fmicb.2014.00128
Lund, M. B., Schätzle, S., Schramm, A., and Kjeldsen, K. U. (2012). Verminephrobacter aporrectodeae sp. nov. subsp. tuberculatae and subsp. caliginosae, the specific nephridial symbionts of the earthworms Aporrectodea tuberculata and A. caliginosa. Antonie Van Leeuwenhoek 101, 507–514. doi: 10.1007/s10482-011-9659-x
Oksanen, J. F., Blanchet, G., Kindt, R., Legendre, P., Minchin, P. R., O'Hara, R. B., et al. (2015). vegan: Community Ecology Package. R Package Version 2.2-1. Available online at: http://CRAN.R-project.org/package=vegan
Pernthaler, J., Glöckner, F.-O., Schönhuber, W., and Amann, R. (2001). Fluorescence in situ hybridization (FISH) with rRNA-targeted oligonucleotide probes. Methods Microbiol. 30, 207–226. doi: 10.1016/s0580-9517(01)30046-6
Pinel, N., Davidson, S. K., and Stahl, D. A. (2008). Verminephrobacter eiseniae gen. nov., sp. nov., a nephridial symbiont of the earthworm Eisenia foetida (Savigny). Int. J. Syst. Evol. Microbiol. 58, 2147–2157. doi: 10.1099/ijs.0.65174-0
Pruesse, E., Peplies, J., and Glöckner, F. O. (2012). SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829. doi: 10.1093/bioinformatics/bts,252
Pruesse, E., Quast, C., Knittel, K., and Fuchs, B. (2007). SILVA: a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 35, 7188–7196. doi: 10.1093/nar/gkm864
R Core Team (2013). R: A language and environment for statistical computing. R Foundation for Statistical Computing. (Vienna). Available online at: http://www.R-project.org/
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Schramm, A., Davidson, S. K., Dodsworth, J. A., Drake, H. L., Stahl, D. A., and Dubilier, N. (2003). Acidovorax-like symbionts in the nephridia of earthworms. Environ. Microbiol. 5, 804–809. doi: 10.1046/j.1462-2920.2003.00474.x
Schweitzer, B., Huber, I., Amann, R., Ludwig, W., and Simon, M. (2001). Alpha- and beta-Proteobacteria control the consumption and release of amino acids on lake snow aggregates. Appl. Environ. Microbiol. 67, 632–645. doi: 10.1128/AEM.67.2.632-645.2001
Sims, R. W., and Gerard, B. M. (1985). Earthworms, Synopses of the Brithish Fauna No. 31, Leiden: J. Brill/Dr. W. Backhuys.
Stackebrandt, E., Frederiksen, W., Garrity, G. M., Grimont, P. A. D., Kämpfer, P., Maiden, M. C. J., et al. (2002). Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 52, 1043–1047. doi: 10.1099/ijs.0.02360-0
Tamura, K., Stecher, G., Peterson, D., Filipski, A., and Kumar, S. (2013). MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 30, 2725–2729. doi: 10.1093/molbev/mst197
Keywords: co-evolution, co-speciation, symbiosis, Verminephrobacter, Flexibacter-like, Ca. Nephrothrix, earthworm, nephridia
Citation: Møller P, Lund MB and Schramm A (2015) Evolution of the tripartite symbiosis between earthworms, Verminephrobacter and Flexibacter-like bacteria. Front. Microbiol. 6:529. doi: 10.3389/fmicb.2015.00529
Received: 18 March 2015; Accepted: 13 May 2015;
Published: 27 May 2015.
Edited by:
Shana Goffredi, Occidental College, USAReviewed by:
Nicolás Pinel, Institute for Systems Biology, USAMichael C. Nelson, University of Connecticut, USA
Copyright © 2015 Møller, Lund and Schramm. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marie B. Lund, Aarhus Institute of Advanced Studies, Aarhus University, Høegh Guldbergs Gade 6B, 8000 Aarhus C, Denmark,bWJsdUBhaWFzLmF1LmRr