 Jun Gong
Jun Gong Yao Qing1,2
Yao Qing1,2 Xiaoli Zhang
Xiaoli Zhang- 1Laboratory of Microbial Ecology and Matter Cycles, Yantai Institute of Coastal Zone Research, Chinese Academy of Sciences, Yantai, China
- 2School of Life Science, South China Normal University, Guangzhou, China
Protistan bacterivory, a microbial process involving ingestion and digestion, is ecologically important in the microbial loop in aquatic and terrestrial ecosystems. While bacterial resistance to protistan ingestion has been relatively well understood, little is known about protistan digestion in which some ingested bacteria could not be digested in cells of major protistan grazers in the natural environment. Here we report the phylogenetic identities of digestion-resistant bacteria (DRB) that could survive starvation and form relatively stable associations with 11 marine and one freshwater ciliate species. Using clone library and sequencing of 16S rRNA genes, we found that the protistan predators could host a high diversity of DRB, most of which represented novel bacterial taxa that have not been cultivated. The localization inside host cells, quantity, and viability of these bacteria were checked using fluorescence in situ hybridization. The DRB were affiliated with Actinobacteria, Bacteroidetes, Firmicutes, Parcubacteria (OD1), Planctomycetes, and Proteobacteria, with Gammaproteobacteria and Alphaproteobacteria being the most frequently occurring classes. The dominance of Gamma- and Alphaproteobacteria corresponds well to a previous study of Global Ocean Sampling metagenomic data showing the widespread types of bacterial type VI and IV secretion systems (T6SS and T4SS) in these two taxa, suggesting a putatively significant role of secretion systems in promoting marine protist-bacteria associations. In the DRB assemblages, opportunistic bacteria such as Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae often presented with high proportions, indicating these bacteria could evade protistan grazing thus persist and accumulate in the community, which, however, contrasts with their well-known rarity in nature. This begs the question whether viral lysis is significant in killing these indigestible bacteria in microbial communities. Taken together, our study on the identity of DRB sheds new light on microbial interactions and generates further hypotheses including the potential importance of bacterial protein secretion systems in structuring bacterial community composition and functioning of “microbial black box” in aquatic environments.
Introduction
Protistan grazing on bacteria is one of the most important ecological processes in microbial food webs that channel carbon and energy to higher trophic levels and regenerate nutrients (Azam et al., 1983). Typically, heterotrophic nanoflagellates (HNFs) are the primary grazers of bacteria, and ciliates can be significant bacterivores in eutrophic habitats (Sherr and Sherr, 2002). In the long evolutionary history of the interplay between bacterial preys and protistan predators, bacteria have seemingly developed many strategies to survive protistan grazing. These include: changes in cell size and filamentation, formation of aggregates, microcolonies and biofilms, increases of swimming speed, and chemical resistance to ingestion (for reviews see Jürgens and Güde, 1994; Hahn and Höfle, 2001; Jürgens and Matz, 2002; Matz and Kjelleberg, 2005; Pernthaler, 2005; Montagnes et al., 2008). It has been hypothesized the existence and development of predation-resistant bacteria may decrease of carbon and energy transfers in the microbial loop and limit nutrient regenerations (Jürgens and Güde, 1994).
Bacterial resistance to digestion represents another important means to survive protistan predation (Jürgens and Güde, 1994; Jousset, 2012). For example, certain Synechococcus and actinobacterial strains could not be digested by nanoflagellates (Boenigk et al., 2001; Zwirglmaier et al., 2009; Apple et al., 2011; Šimek et al., 2013). It was suggested that the presence of protective S-layer in the cell wall could protect Synechococcus cells from enzymatic degradation in food vacuoles of the ciliate Tetrahymena (Koval, 1993). Freshwater isolates of Janthinobacterium lividum and Chromobacterium violaceum could kill the nanoflagellate grazers by releasing a toxin (Matz et al., 2004). The pathogenic bacterium Campylobacter jejuni remained viable after ingestion for 5 h by a freshwater ciliate Colpoda sp. (First et al., 2012). Many bacterial strains (e.g., Legionella, Listeria, Vibrio, and Salmonella) could persist inside Acantbamoeba and Tetrahymena cells, which might have given rise to intracellular symbionts, parasites, and pathogens (Barker and Brown, 1994; Greub and Raoult, 2004; Brandl et al., 2005; Matz and Kjelleberg, 2005). However, these studies have mostly tested the digestibility of selected bacterial strains of pathogenicity and/or from freshwater environments. What has not been investigated much, so far, is the diversity and composition of the bacterial assemblages that are resistant to digestion by major protistan bacterivores in complicated microbial communities of aquatic systems.
Recently, we have investigated a range of ciliate species for identities of putatively DRB. Ciliates were chosen primarily because of their large cell size, which allowed to be easily manipulated at a single-cell level to minimize the chance of bacterial contaminations. Previously, we reported a new intracellular bacterial species belonging to the phylum Parcubacteria (the candidate division OD1) in a starved freshwater ciliate Paramecium bursaria (Gong et al., 2014). Here, we extend this line of research by identifying some DRB (and endosymbionts) in 11 marine and 1 freshwater ciliate species, with which we hope to provide a broad view of the diversity of bacterial populations that might have escaped from protistan digestion. The unveiled taxonomic affiliations of DRB enable us to link to enormous microbiological, genetic and ecological knowledge bearing on these bacterial names, which lays a basis to a better understanding of associations and interactions between bacteria and protists in marine microbial food webs, and to generate new ecological hypotheses.
Materials and Methods
Organisms, Source, and Culture Conditions
Thirteen strains of 12 (11 marine and 1 freshwater) ciliate species belonging to four classes, Spirotrichea, Oligohymenophorea, Heterotrichea, and Prostomatea, were investigated (Table 1). Ten free-living species were kindly provided by Prof. Weibo Song’s lab at Ocean University of China (OUC), Qingdao, or directly sampled from aquatic environments by the authors of this work. Two endosymbiotic ciliates, Boveria labialis and Urceolaria urechi, were isolated from the sea cucumber (Apostichopus japonicas) and the Chinese penis fish (Urechis unicinctus), respectively. The host animals were purchased from local seafood markets in Yantai. Seven cultured strains were maintained in Petri dishes at 18°C for several days, with water from the sampling sites and several rice grains to enrich bacteria for food (Table 1). All ciliates were observed in vivo for living features (Figure 1), and identified according to the taxonomic reference (Song et al., 2009).
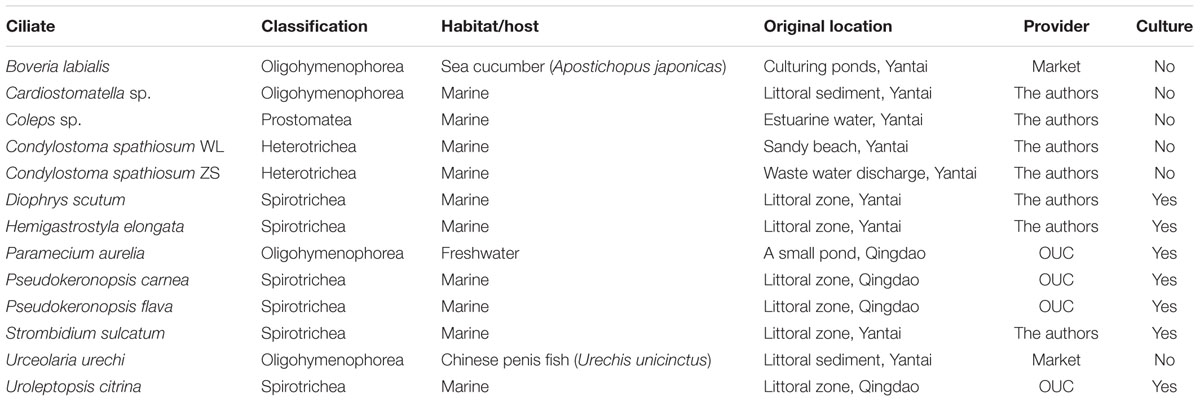
TABLE 1. A summary of ciliate species investigated in this study.
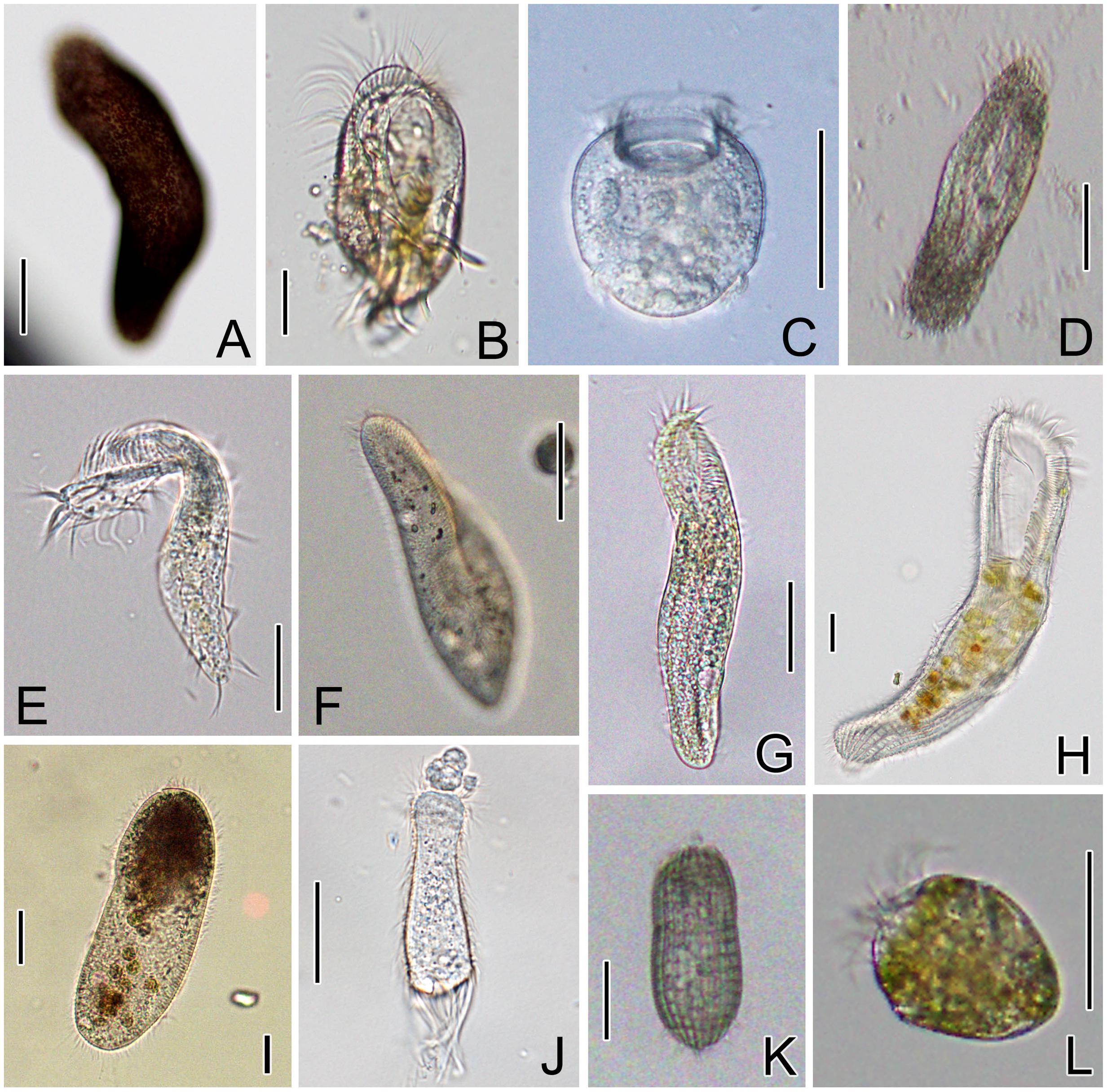
FIGURE 1. Morphology of ciliate species in vivo. (A) Pseudokeronopsis carnea; (B) Diophrys scutum; (C) Urceolaria urechi; (D) Hemigastrostyla elongata; (E) Pseudokeronopsis flava; (F) Paramecium aurelia; (G) Uroleptopsis citrina; (H) Condylostoma spathiosum; (I) Cardiostomatella sp.; (J) Boveria labialis; (K) Coleps sp.; (L) Strombidium sulcatum. Scale bars = 50 μm.
DNA Extraction, Clone Libraries, and Sequencing
Ciliate cells were transferred into autoclaved seawater (sterilized double distil water for the freshwater species Paramecium aurelia) with a micropipette. In order to minimize contaminations, cells were washed for three to five times to remove microorganisms attaching cilia and cell surface. The ciliates were then maintained in the sterilized water for 12 to 24 h, allowing the starving hosts to digest the ingested bacteria as much as possible. After starvation, the remaining individuals were washed again. Three to five individuals were transferred to a PCR tube with a minimum volume of water for DNA extraction, and up to 20 individuals were mounted onto slides for subsequent fluorescence in situ hybridization (FISH) assays.
Genomic DNA extraction was performed as previously described (Gong et al., 2014). Bacterial 16S rRNA genes were PCR amplified with primer set 8F (5′- AGAGTTTGATCCTGGC TCAG -3′) and 1492R (5′-GGTTACCTTGTTACGACTT-3′), or with 8F and 1392R (5′- ACGGGCGGTGTGTAC -3′) (Lane, 1991). The PCR reaction solution (25 μl) contained 1 μl of 10 μM primers, 1 μl extracted DNA solution, 2.5 μl dNTP mix (0.2 mM of each) and 0.625 units of DreamTaq DNA polymerase and 2.5 μl 10X DreamTaq buffer with MgCl2 at a concentration of 20 mM (Thermo Scientific, USA). All PCR reactions were performed in a Biometra thermal cycler with the following program: an initial denaturation 94°C for 3 min, followed by 34 cycles of 94°C for 1 min, annealing (at 50°C for primers 8F/1492R, and 52°C for primers 8F/1392R) for 1 min, and 72°C for 1 min, with a final extension step of 72°C for 10 min. The amplified PCR products were purified with a gel purification kit (Tiangen Biotech, China), ligated into pTZ57R/T vector using InsTAclone PCR Clone Kit (Thermo Scientific) and transformed into competent cells of Escherichia coli DH5α. The clones containing the DNA inserts were randomly selected. These positive clones were either pre-screened using restricted fragment length polymorphism (RFLP) analysis with two (Taq and HhaI, or Taq and MspI) or one restricted enzyme (MspI) (FastDigest, Thermo Scientific, USA), or directly sent for sequencing on an ABI 377 automated sequencer (Sangon, Shanghai, China). A total of 13 clone libraries of bacterial 16S rRNA genes were constructed for the 13 ciliate strains.
Phylogenetic Analysis
The newly obtained 16S rRNA gene sequences were first aligned using MAFFT v.7 (Katoh and Standley, 2013). Chimeric sequences were identified using Bellerophon (Huber et al., 2004), and then removed for the subsequent analyses. The remaining sequences were subjected to BLAST against GenBank, and to ribosomal database project (RDP) databases for classification (Cole et al., 2009). Closely related sequences were retrieved from GenBank and aligned with these newly obtained. The compiled sequences were then aligned using SINA (ARB-Silva) with default settings (Pruesse et al., 2012) and manually modified, resulting in a final alignment of 1,365 positions. Maximum likelihood (ML) trees were constructed with FastTree V.2 program by default settings (Price et al., 2010), under a GTR+CAT model. The resulted ML tree were further organized and revised by Interactive Tree of Life (iTOL)1.
For some sequences that assigned into unclassified Gammaproteobacteria, Rickettsiales, and Flavobacteria by the RDP classifier, both ML and Bayesian inference (BI) analyses were carried out to further resolve their taxonomic ranks. PhyML program was used for building a ML tree under a best-fit GTR+G+I model. BI analyses were performed with MrBayes 3.1.2 (Ronquist and Huelsenbeck, 2003). Markov chain Monte Carlo (MCMC) simulations were run with two sets of four chains using the default settings, with a sampling frequency of 0.01. Convergence of the chain length was confirmed from the standard deviation of split frequencies (<0.01). 1,000,000 or 2,000,000 generations were run for these datasets. Twenty-five percent of generations were discarded as burn-in in each analysis. To characterize the “species”-level composition and variations of the DRB assemblages, operational taxonomic units (OTUs) were defined at a cutoff of 97% sequence similarity and analyzed using the Mothur program (Schloss et al., 2009).
To explore the beta diversity of DRB among ciliate specimens, a Bray–Curtis similarity matrix was calculated based on the relative abundance of different families, and visualized with the Clustering method using the software PRIMER 6 (PRIMER-E, UK). Differences in assemblage structure among samples were statistically tested using analysis of similarity (ANOSIM) (Clarke and Gorley, 2006), to examine the possible effect of habitat (marine vs. freshwater), class-level taxonomy (four classes), life style (free-living vs. symbiotic), and sampling method (environmental isolate vs. laboratory culture) of the hosts.
Probe Design and Fluorescence In Situ Hybridization
A 16S rRNA-targeted oligonucleotide probe targeting the genus Aestuariibacter, which included the most common DRB phylotypes in this study, was designed as previously described (Gong et al., 2014). In brief, conserved 16S rRNA regions of Aestuariibacter species were identified based on the rRNA alignment of a range of species. Several short fragments (length of 16–20 nucleotides) in these regions were then selected and evaluated using PROBE MATCH of RDP release 10 (Cole et al., 2009). A web tool, mathFISH, was used for assessing sensitivity and specificity, and the optimum formamide concentration (40%) for mismatch discrimination optimizing (Yilmaz et al., 2011). The newly designed probe was named ALT658, with the sequence 5′-TTCCACTCCCCTCTCCAA-3′.
A number of ciliates examined in this study were subjected to FISH with a mixture of universal eubacterial probes, EUB338, II and III (Amann et al., 1995; Daims et al., 1999). The non-sense probe NON338, complementary to EUB338, was used as a negative control for the hybridization protocol (Manz et al., 1992). In the case of detection of Aestuariibacter phylotypes in ciliate hosts, FISH with the probe ALT658 was performed separately to reveal the quantity and location of the bacteria. All probes in this study were labeled with Cy3 at the 5′ end.
Whole-cell hybridization was according to (Fried et al., 2002). Cells were fixed with Bouin’s solution (50%, final concentration). Cell suspensions were dropped onto microscopic slides (SuperFrost Plus) and air dried at room temperature. The slides were put away in a black box and stored at 4°C. Before FISH assay, the slides were washed in distilled water three times for 10 min and then progressively dehydrated via an ethanol gradient (30, 50, 80, and 100%). Slides were incubated at 46°C for 3 h in hybridization buffer, which contained 20 mM Tris-HCl (pH 8.0), 0.9 M NaCl, 0.01% sodium dodecyl sulphate (SDS), 30% (40% for the probe ALT658) formamide and the relevent fluorescent probe (5 ng μl-1 final concentration). After hybridization, slides were washed for 15 min at 48°C with wash buffer [20 mM Tris-HCl (pH 8.0), 450 mM NaCl, 0.01% SDS], and then rinsed with chilled Milli-Q water. Slides mounted with anti-fade mounting medium (Beyotime, China) and DAPI (50 ng ml-1) were observed under an epifluorescence microscope (Olympus BX51, Japan) with green-light excitation (wavelength 505 to 560 nm) for Cy3 and UV excitation (wavelength 330 to 385 nm) for DAPI signals. A SPOT RT3 digital camera (SPOT Imaging Solutions, Sterling Heights, USA) was used for visualization.
Sequence Accession Numbers
The bacterial 16S rRNA gene sequences newly obtained in this study have been deposited in the GenBank database under the accession numbers KU524761 – KU524878.
Results
Diversity and Composition of Digestion-Resistant Bacteria (DRB)
A total of 118 bacterial 16S rRNA gene sequences were obtained from clone libraries constructed for DRB in 12 ciliate species. Classification via RDP classifier pipeline showed that the digestion-resistant bacteria (DRB) were highly diverse. There were 40 OTUs, which were affiliated with 13 identified families of seven bacterial phyla/candidate divisions, Actinobacteria, Bacteroidetes, Firmicutes, GN02, Planctomycetes, Proteobacteria (including four classes: Alpha-, Beta-, Gamma-, and Epsilonproteobacteria), and Parcubacteria (OD1) (Figure 2; Table 2).
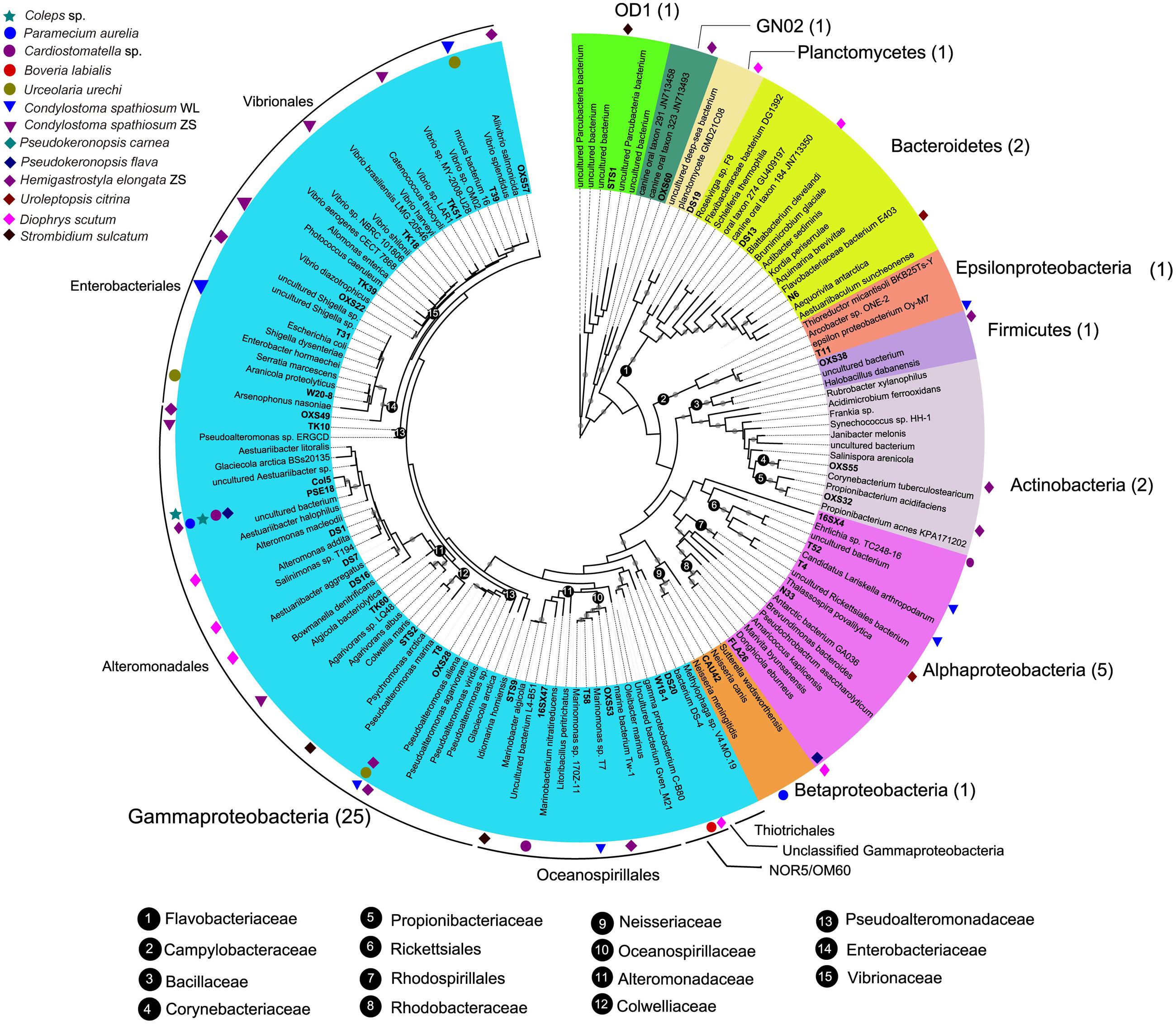
FIGURE 2. A maximum likelihood (ML) tree of 16S rRNA genes showing the phylogeny of digestion-resistant bacteria (DRB) detected in 12 species of ciliated protists. Representative sequences of 40 bacterial OTUs were used to build the tree under a GTR+CAT model. Numbers in parentheses indicate the number of bacterial OTUs affiliated with the phyla/candidate divisions. Note that OTUs of Gammaproteobacteria (Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae) and Alphaproteobacteria represent the most abundant digestion-resistant bacterial groups.
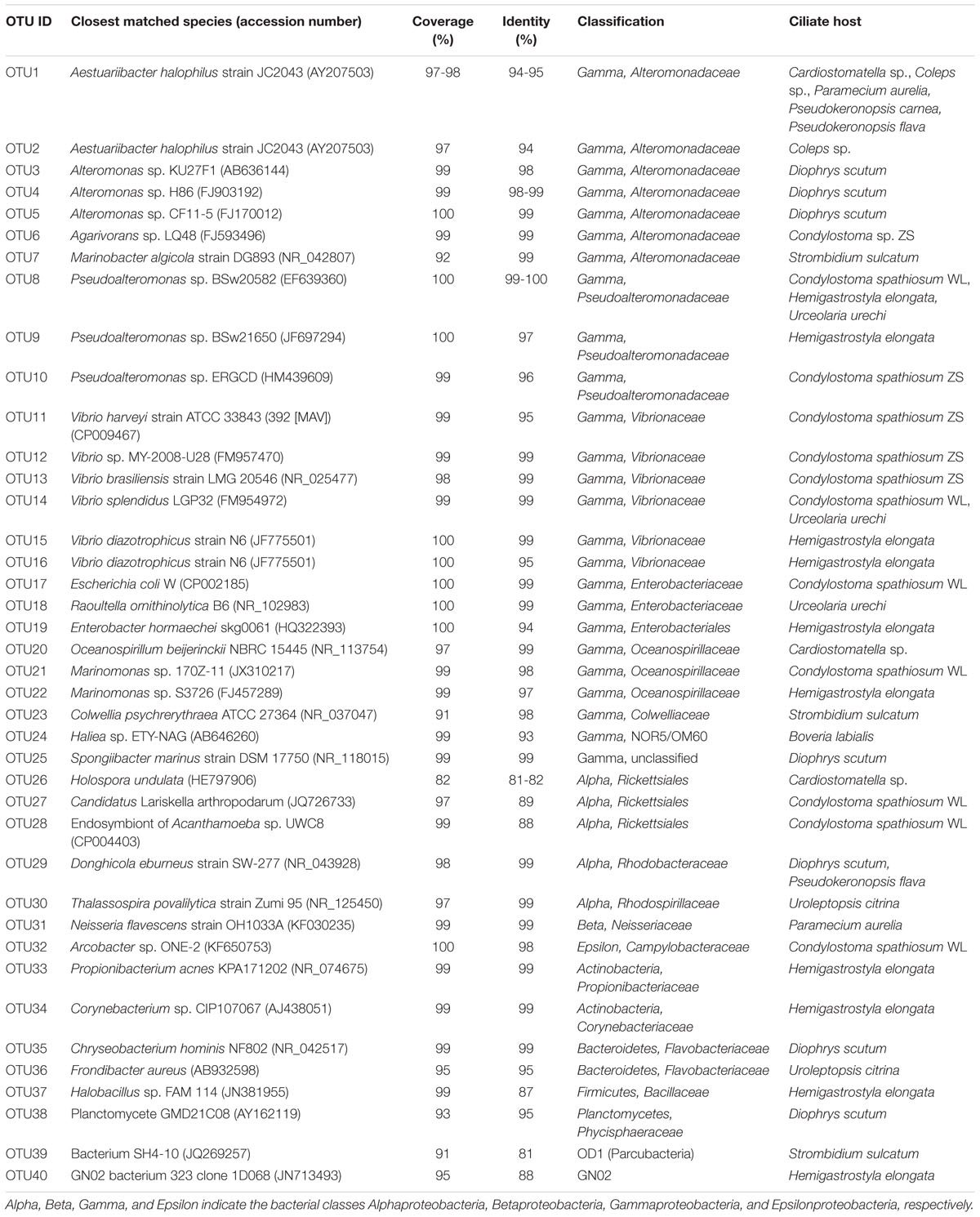
TABLE 2. The closest matches in the Genbank by BLASTing 16S rRNA genes and ribosomal database project (RDP) classification of bacteria associated with ciliate host species subjected to starvation.
Gammaproteobacteria
Overall, Gammaproteobacteria phylotypes dominated the DRB assemblages. About 63% (25) observed OTUs of the DRB were affiliated with this class (Figure 2; Table 2). These included Alteromonadaceae (7 OTUs), Pseudoalteromonadaceae (3 OTUs), Vibrionaceae (6 OTUs), Enterobacteriaceae (3 OTUs), Oceanospirillaceae (3 OTUs), Colwelliaceae (1 OTU), the NOR5/OM60 group (1 OTU) and an unclassified OTU within the class.
Alteromonadaceae phylotypes (OTU1–OTU7) were observed in eight ciliate strains, being the most frequently occurred family in the DRB assemblages (Figures 2 and 3; Table 2). A single OTU of this family (OTU1), which exhibited 94–95% similarities to Aestuariibacter halophilus strain JC2043 in 16S rRNA gene sequences, was detected coincidently in five ciliate species, with higher proportions in Coleps sp. (100%), Pseudokeronopsis carnea (100%), Paramecium aurelia (96%), and Pseudokeronopsis flava (90%) than in Cardiostomatella sp. (10%). Other phylotypes of this family were closely related to Alteromonas sp. (98–99% similarities) and Marinobacter algicola (99%), and found with high proportions in the DRB assemblages of Diophrys scutum (78%) and Strombidium sulcatum (52%), respectively. The Agarivorans phylotype occurred only in Condylostoma spathiosum strain ZS with a proportion of 10% (Figures 2 and 3; Table 2).
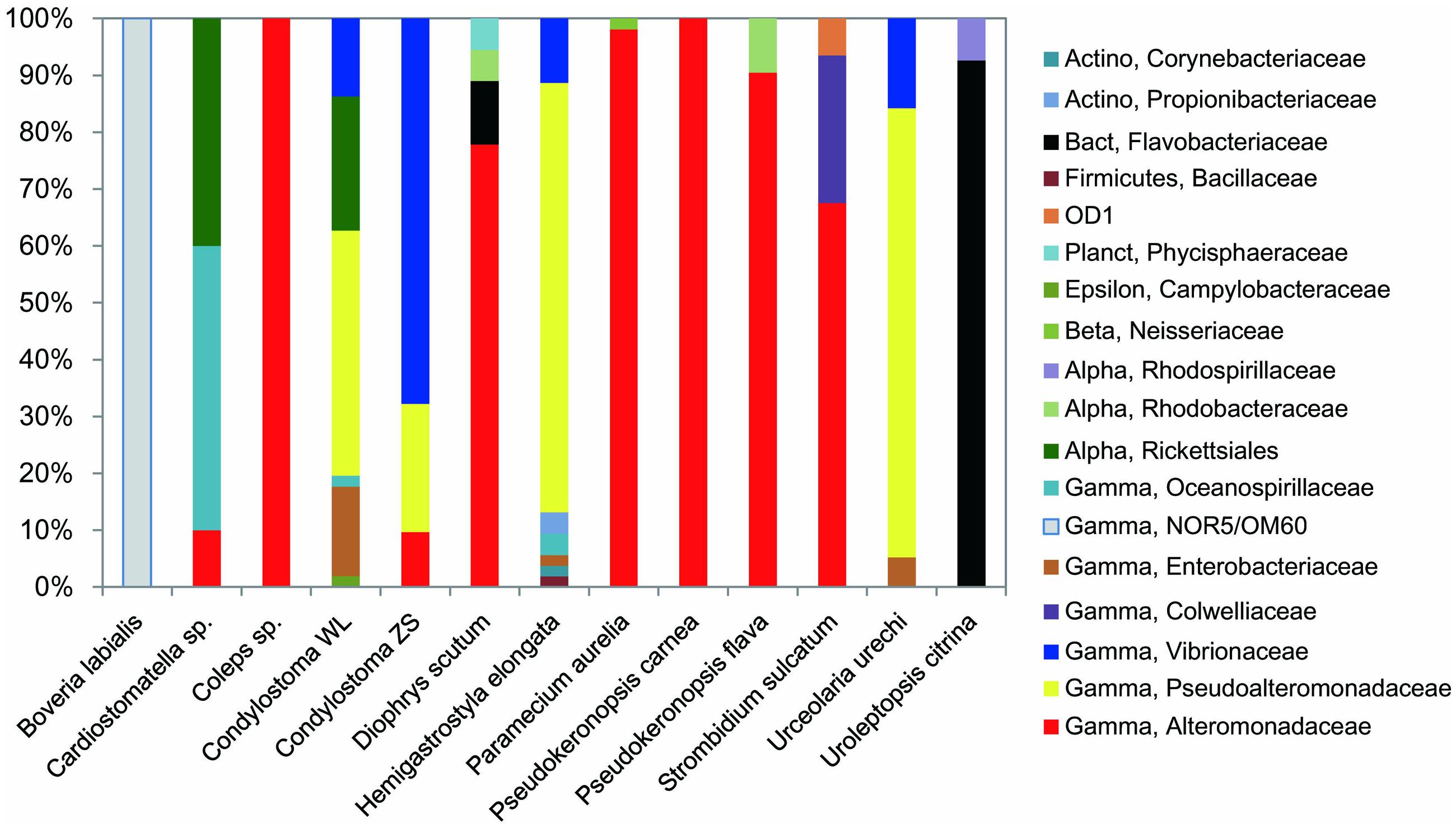
FIGURE 3. Relative abundances of major taxa in digestion-resistant bacterial assemblages in 12 species (13 strains) of ciliates. Actinobacteria (Actino), Alphaproteobacteria (Alpha), Bacteroidetes (Bact), Betaproteobacteria (Beta), Gammaproteobacteria (Gamma), Epsilonproteobacteria (Epsilon), Firmicutes, Parcubacteria (OD1), and Planctomycetes (Planct) were detected. Taxa are classified at the family level unless it is not supported (<a threshold of 80%) by ribosomal database project (RDP) classifier pipeline. Note that the families Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae are the most frequently occurred and generally account for a large proportion in the assemblages.
Pseudoalteromonadaceae phylotypes (OTU8–OTU10) in the DRB assemblages showed 95–100% sequence similarities with members of Pseudoalteromonas. These phylotypes occurred with high proportions in the DRB assemblages of Urceolaria urechi (79%) and Hemigastrostyla elongata (68%), but lower in the two strains of Condylostoma spathiosum (43 and 23%) (Figures 2 and 3; Table 2).
Vibrionaceae phylotypes (OTU11–OTU16) were frequently detected among the DRB assemblages of three ciliate species (four strains) (Figure 3). These phylotypes shared 95–99% sequence similarities with vibrios (Table 2). The highest proportion (68%) of Vibrionaceae was recorded in Condylostoma spathiosum strain ZS. However, much lower proportions of vibrios were observed in the Condylostoma strain WL (14%), Hemigastrostyla elongata (10%), and Urceolaria urechi (16%) (Table 2).
Enterobacteriaceae phylotypes (OTU17–OTU19) were detected with relatively lower proportions in the DRB assemblages of Condylostoma spathiosum strain WL (16%), Urceolaria urechi (5%), and Hemigastrostyla elongata (2%) (Figure 3). BLAST against GenBank indicated that these phylotypes were closely related to Escherichia coli, Enterobacter hormaechei, or Raoultella ornithinolytica, with sequence similarities of 94 and 99%. Hemigastrostyla elongata hosted the OTU19, which was affiliated with the order Enterobacteriales (Table 2).
Three OTUs of Oceanospirillaceae (OTU20–OTU22) likely representing species of Oceanospirillum and Marinomonas were detected in three ciliate species with variable proportions: Cardiostomatella sp. (50%), Condylostoma spathiosum WL (2%), and Hemigastrostyla elongata (3%) (Figure 3; Table 2). A Colwelliaceae OTU closely related to Colwellia psychrerythraea (98% sequence similarity) accounted for 20% in the DRB assemblage of Strombidium sulcatum (Figure 3; Table 2).
Ribosomal database project pipeline assigned two OTUs (OTU24 and OTU25) into the class Gammaproteobacteria, but lower rank classification could not be resolved. The OTU24 were only observed in Boveria labialis, showing a sequence identity of 93% to Haliea sp. ETY-NAG, an ethylene-assimilating marine bacterium (Suzuki et al., 2012). Nevertheless, our phylogenetic analyses further demonstrated that OTU24 was affiliated to an uncultured gammaproteobacterial group called NOR5/OM60, and closely related to the members of subclade NOR5–12 (see Figure 4), of which sequences are mainly from deep-sea samples and cultured members are known as aerobic anoxygenic phototrophs that need organic substrates like carboxylic acids, oligopeptides, or fatty acids for growth (Fuchs et al., 2007; Yan et al., 2009). OTU25 detected in Diophrys scutum ciliates could represent a strain of Spongiibacter marinus (99% similarity) (Table 2), a gammaproteobacterium originally isolated from a marine sponge (Graeber et al., 2008).
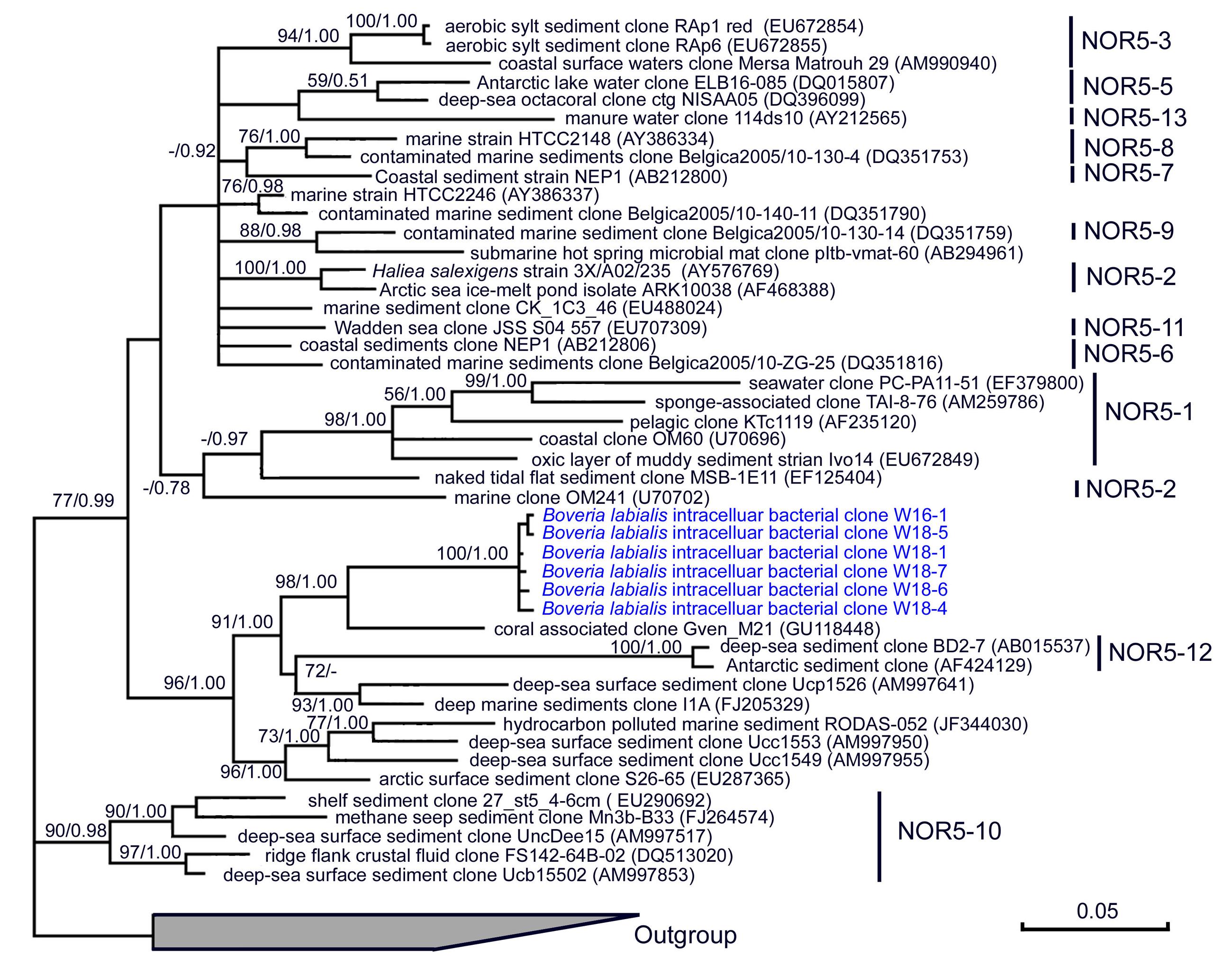
FIGURE 4. A consensus tree based on 16S rRNA genes showing the phylogenetic position (class Gammaproteobacteria, group NOR5/OM60) of a bacterium detected in the macronucleus and micronucleus of ciliate Boveria labialis, an endosymbiotic protist from sea cucumbers. The trees were constructed using ML (under GTR+G+I model) and Bayesian algorithms. Bootstrap values lower than 50% and posterior probability lower than 0.5 were not shown.
Alphaproteobacteria
Five out of the 40 OTUs (13%) of DRB found in this study were affiliated to the class Alphaproteobacteria. The Donghicola eburneus-like OTU29 (family Rhodobacteraceae) occurred in the two ciliates Pseudokeronopsis flava and Diophrys scutum with minor proportions (<10%) (Figure 3; Table 2). The OTU (OTU46) of the family Rhodospirillaceae was Thalassospira povalilytica-like (99% similarity), occurred only in the ciliate Uroleptopsis citrina (Table 2).
Three alphaproteobacterial OTUs (OTU26–OTU28) were assigned with the order Rickettsiales by RDP classifier (Table 2). Phylogenetic analyses showed that the two OTUs (OTU27 and OTU28) detected in Condylostoma spathiosum WL were closely related to a symbiont of Acanthamoeba sp. (88% similarity) and Candidatus Lariskella arthropodarum (89% similarity), and placed well within Midichloriaceae (ML 100%, BI 0.98) (see Figure 5A), one of three families of the order Rickettsiales (Ferla et al., 2013). The remaining OTU (OTU26) sharing 81–82% sequence similarities with Holospora undulata represented one of the basal lineages to Rickettsiales and Holosporales (Figure 5A). OTU26 occurred only in the ciliate Cardiostomatella sp. with a moderate proportion (40%).
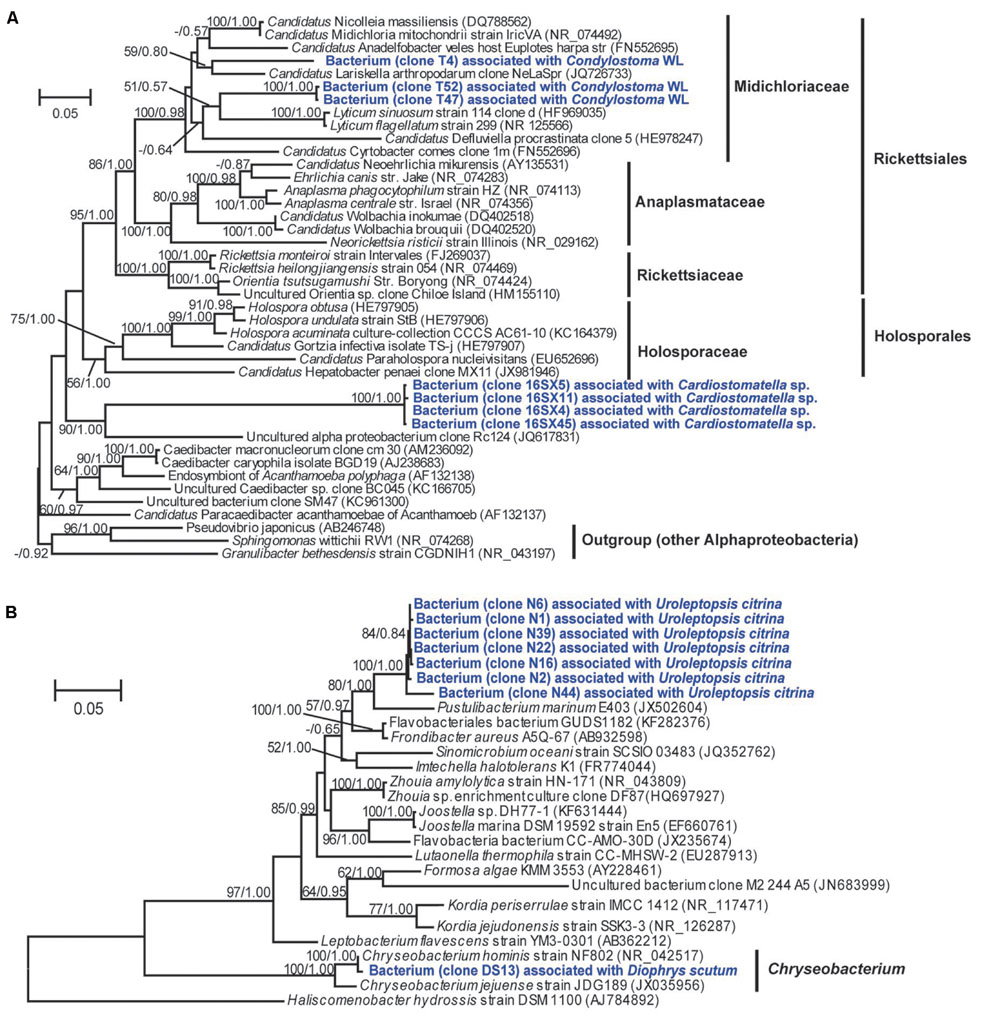
FIGURE 5. Consensus trees based on 16S rRNA genes showing the phylogenetic positions of digestion-resistant bacterial phylotypes affiliated with the order Rickettsiales (A) and the family Flavobacteriaceae (B). The trees were constructed using ML (under GTR+G+I model) and Bayesian algorithms. Bootstrap values lower than 50% and posterior probability lower than 0.5 were not shown.
Betaproteobacteria and Epsilonproteobacteria
Phylotypes of these two classes were rarely detected in the DRB assemblages. The freshwater ciliate Paramecium aurelia hosted a bacterial phylotype (OTU31) of Neisseria flavescens (99% similarity), a member of the family Neisseriaceae. OTU32, an epsilonproteobacterial phylotype detected once Condylostoma spathiosum WL, appeared to be a member of the genus Arcobacter (98%), family Campylobacteraceae (Table 2).
Actinobacteria
Two actinobacterial OTUs (OTU33 and OTU34) were classified into two families, Corynebacteriaceae and Propionibacteriaceae, which all were recovered from the hypotrich ciliate Hemigastrostyla elongata (Table 2). These sequences resemble most to these of Propionibacterium acnes and Corynebacterium acnes (99%) and Propionibacterium sp. (99%), respectively. Nevertheless, the proportion of Actinobacteria in the DRB assemblages was much lower (3%) (Figure 3).
Bacteroidetes
There were two bacteroidetes OTUs (OTU35, OTU36), all of which were affiliated to the family Flavobacteriaceae. OTU35 was associated with Diophrys scutum, representing the species Chryseobacterium hominis (99% similarity) (Figure 5B; Table 2). OTU36 was closely related to Frondibacter aureus (95% similarity) and appeared with a high proportion (93%) in the ciliate Uroleptopsis citrina (Table 2).
Firmicutes, Planctomycetes, Parcubacteria (OD1), and GN02
Digestion-resistant bacteria affiliated with these phyla/candidate divisions were generally minor in the assemblages. Hemigastrostyla elongata hosted phylotypes of Bacillaceae (Firmicutes) (OTU37) and the candidate division GN02 (OTU40), which shared low sequence identities with Halobacillus sp. (87%) and a GN02 bacterium (88%), respectively (Table 2; Figure 3). Also, a phylotype (OTU38) of Phycisphaeraceae (Planctomycetes) was found in Diophrys scutum, showing a sequence identity of 95% with a cultured planctomycete (Table 2). One of bacterial phylotypes from the ciliate Strombidium sulcatum belonged to Parcubacteria (OD1), of which many sequences await to be further designated to lower taxonomic ranks. This DRB phylotype showed a closest match (90% identity) with an environmental sequence from a trichloroethylene-contaminated aquifer (Table 2), but had no signature sequence fragment of OD1-p, a clade previously recognized within the candidate division (Gong et al., 2014).
Beta Diversity of DRB
Since few OTUs were in common among DRB assemblages of ciliate species, we calculated their beta diversity based on the relative abundance variations of phylotypes at the family level. At the level of 10% similarity, the 13 DRB assemblages were clustered into four groups (Figure 6). The DRB turnover appeared to be high and did not follow a specific pattern with respective to the taxonomic affiliation (ANOSIM, R = -0.045, P = 0.578), habitat (R = -0.24, P = 0.923), or the sampling source (R = 0.106, P = 0.133). However, compared with the free-living species, the two symbiotic ciliates (Boveria labialis and Urceolaria urechi) hosted significantly different DRB assemblages (R = 0.44, P = 0.026).
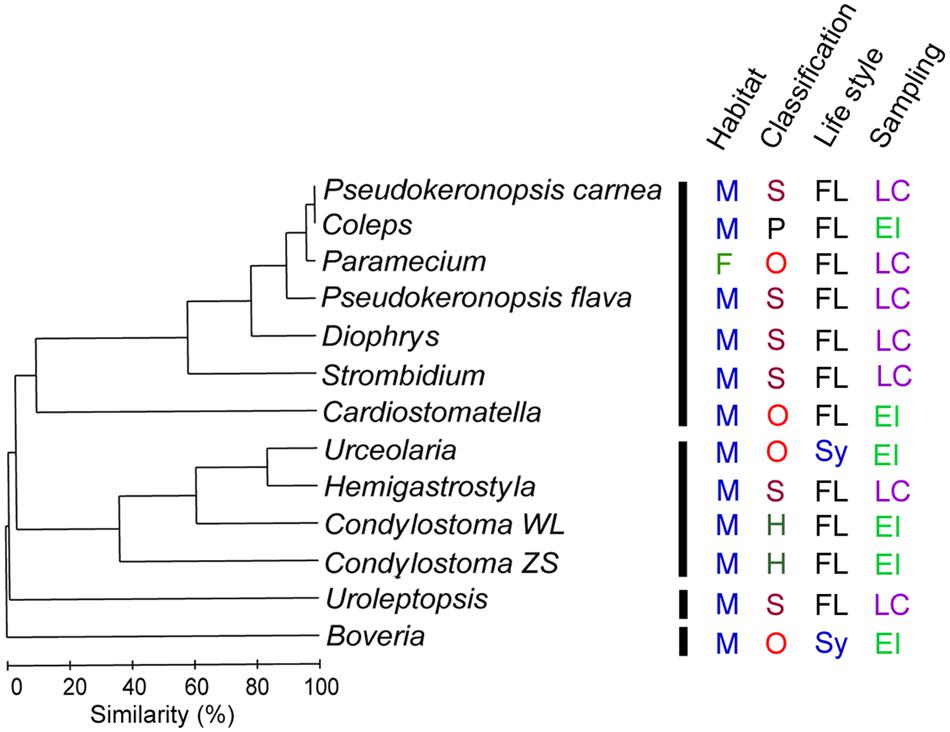
FIGURE 6. Clustering of digestion-resistant bacterial (DRB) assemblages among 13 ciliate specimens. Based on the distribution of each bacterial families detected, the assemblages can be divided into four groups at a similarity level of 10%. This grouping pattern of DRB composition hardly reflects differences between marine (M) and freshwater (F) habitats, class-level affiliations (Heterotrichea, H; Oligohymenophorea, O; Prostomatea, P; and Spirotrichia, S), and sampling sources (environmental isolation, EI and laboratory culture, LC) of the hosts. Nevertheless, two symbiotic (Sy) ciliates appeared to have significantly different DRB compared with those free-living (FL) species (ANOSIM, P < 0.05).
Fluorescence In Situ Hybridization
Searching through Probe Match function of RDP showed that the newly designed probe ALT658 completely matched 4273 sequences in the database, among which 4223 sequences (98.8%) were affiliated with the family Alteromonadaceae (13694 sequences curated in RDP), particularly the three genera of the family: Aestuariibacter (407/524), Alteromonas (2715/4114), and Glaciecola (621/759) (accessed on January 3, 2016). This indicates that the probe ALT658 is of high specificity and targets all Alteromonadaceae bacterial taxa recovered in this study, though not all known members of Alteromonadaceae could be matched.
Fluorescence in situ hybridization with eubacterial probes (EUB338, II, and III) and the Alteromonadaceae-specific probe ALT658 revealed that most of the targeted bacteria presented in the cytoplasm of ciliate cells (Figure 7). In Uroleptopsis citrina and Pseudokeronopsis flava ciliates, red fluorescence of Cy3-labeled probes was relatively stronger, mostly presented as aggregates and positioned at the anterior portion of the host cells, indicating the DRB were active (Figures 7A–C,M–O). However, in other ciliate species such as Paramecium aurelia and two strains of Condylostoma spathiosum, the positive signal was relatively weak, diffuse and/or irregularly distributed in the ciliate cytoplasm (Figures 7G–L,S–X). In all cases, the negative controls with the non-sense probe NON338 were applied, and no Cy3-labeled signal was observed (see Figure 7W); no obvious food vacuoles were observed in starved and fixed specimens either.
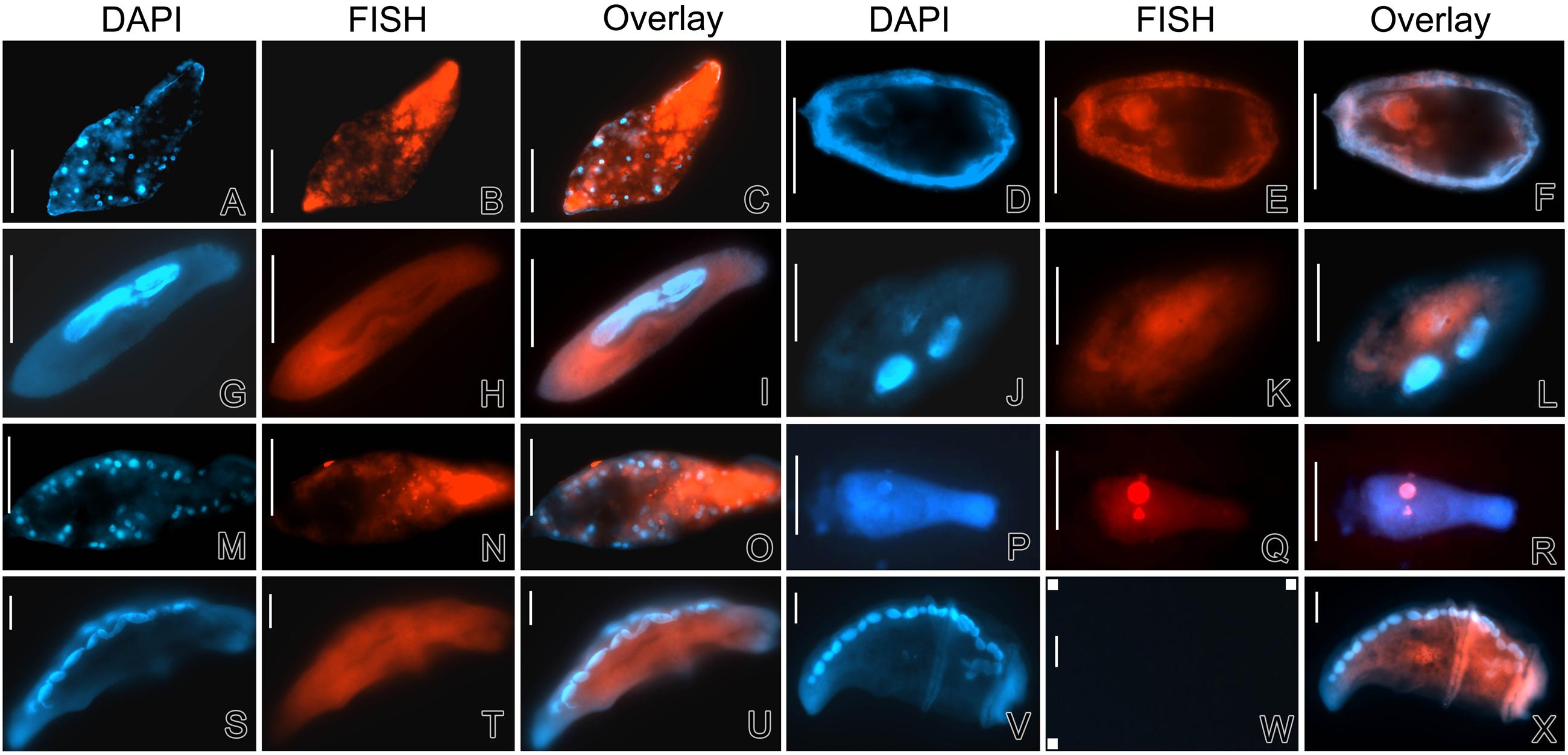
FIGURE 7. Detection of DRB in ciliates using fluorescence in situ hybridization (FISH) and DAPI staining. (A–C) Pseudokeronopsis flava; (D–F) Coleps sp.; (G–I) Paramecium aurelia; (J–L) Hemigastrostyla elongata; (M–O) Uroleptopsis citrina; (P–R) Boveria labialis; (S–U) Condylostoma spathiosum strain WL; (V–X) Condylostoma spathiosum strain ZS. The specimens were DAPI-stained and excited with UV, focusing on the nuclear apparatus (A,D,G,J,M,P,S, and V). FISH was performed with Cy3-labeled probe ALT658 targeting members of the family Alteromonadaceae in ciliates Coleps sp., Uroleptopsis citrina, and Paramecium aurelia; other specimens were examined with universal eubacterial probes (EUB338, II and III). (W) Shows an example of negative controls by using the non-sense probe Cy3-NON338 in the FISH protocol, in which neither Cy3 fluorescence nor autofluorescence could be observed. Scale bars = 50 μm.
There were positive FISH signatures inside the cell and on the surface of the ciliate Coleps sp. (Figures 7D–F), indicating some of the associated bacterial individuals were indeed intracellular, but others might be contamination or of ectosymbiontic nature. Nevertheless, all these bacteria (OTU1 and OTU2) associating with the Coleps species appeared to be closely related to Aestuariibacter. In the endosymbiotic ciliate Boveria labialis, the bacteria (belonging to Gammaproteobacteria, NOR5/OM60 group; see the results mentioned above) were located in both the macronucleus and the micronucleus (Figures 7P–R).
Discussion
Conceptual and Methodological Considerations
To our knowledge, this study is the first to identify the DRB in ciliates that are isolated directly from microbial communities of lab microcosms or the field. We define DRB as an ecologically functional group of bacteria that are able to evade digestion by bacterivorous protists. To characterize DRB, it may be possible to collect these bacteria ingested and subsequently expelled from the protistan cell, since it has been shown that Salmonella enterica could be packed in vesicles and released by a soil Tetrahymena species (e.g., Brandl et al., 2005). This approach is technically difficult in practice, because such fecal vesicles have not often been observed for other ciliates and there is a high risk of contaminations by environmental bacteria outside the grazer. Alternatively, the bacteria remain inside the predator after starving can be inspected. In this approach, it may be difficult to distinguish these “recently” ingested and these “previously” ingested (i.e., endosymbionts), which both present as the intracellular bacteria. We thus considered both of these groups as DRB because of their similar features in physiology (e.g., indigestibility) and ecological and evolutionary consequences (e.g., adaptation to intracellular life style).
The timespan of starvation may affect our assessment on the richness and composition of DRB assemblages. First et al. (2012) showed that the pathogenic bacterium Campylobacter jejuni remained viable after ingestion for 5 h by a freshwater ciliate Colpoda sp. During the grazing experiments, Tetrahymena cells began expelling vesicles within 1 h after initiation of feeding (Brandl et al., 2005). Starvation for a longer period of time might lead to fewer bacterial species detected in the ciliates (Gong, unpublished data), and a single bacterial species was detected in a paramecium ciliate subjected to starvation for several days (Gong et al., 2014).
The remaining DNA of already digested bacteria might lead to contamination, as DNA-based approaches were applied for detecting DRB in this study. Nevertheless, the DNA of engulfed bacteria that are already digested by host cells of ciliates seems not to be a problem, because these DNA are likely completely hydrolyzed by enzymes in lysosomes during a reasonable period of time. Results on several studies on DNA degradation by lysosomal enzymes are analogous to, and clearly supportive for, this putative intracellular process in protists. Arsenis et al. (1970) found that the calf thymus DNA can be completely degraded into acid-soluble bases by lysosomal extracts of rat liver and Ehrlich ascites tumor cells within 10 h. They also suggested that the hydrolytic action of lysosomes was the highest toward nucleic acids among the various major cellular constituents studied (Arsenis et al., 1970). Another study also found that, when incubated at 37°C, the DNA of apoptotic cells engulfed by macrophages would be degraded by macrophage lysosomal enzymes in a time-dependent manner, and no longer detectable on the gel in a 6-h incubation (Odaka and Mizuochi, 1999). On these bases, we believe the treatment time (12–24 h) was long enough to degrade the DNA of digested bacteria in ciliates, resulting in a minimum probability of this type of contamination. Moreover, this treatment time is not too long to kill the eukaryotic hosts and thus practical in this research.
High Richness and Assemblage Variations (Beta Diversity) of Digestion-Resistant Bacteria in Ciliates
We detected at least one DRB OTU in each strain and on average three OTUs per strain of the 12 ciliate species. The 40 DRB OTUs affiliated with eight phyla and about 14 families were found in this study, suggesting that there is a collectively high phylogenetic diversity of DRB in these bacterivorous protists. Nevertheless, since we used approaches of clone libraries and Sanger sequencing of 16S rRNA genes, which are known to have limited sampling depths to recover rare phylotypes in complicated microbial communities, the OTU-level diversity of DRB in this study is more likely attributed to these most abundant or commonly occurred. Future studies with high-throughput sequencing may reveal higher phylogenetic diversities with more rare phylotypes of various phyla in the DRB assemblages.
Relative to the well-known high diversity and complicated structure of bacterial communities in the natural aquatic environment, the DRB assemblages in ciliates were apparently much simplified. This partly reflects the selective feeding behavior of ciliate grazers, through which some bacterial populations in the community are preferably engulfed. Selective ingestion of bacteria by ciliates depends on bacterial size (Gonzalez et al., 1990; Posch et al., 2001), growing phase (Sherr et al., 1992), ratios of C:P and N:P (Gruber et al., 2009), and motility (Sherr and Sherr, 2002). It is highly likely that only a fraction of ingested bacterial preys could have the ability to survive ciliate digestion, though currently we know little about which groups of ingested bacteria can be completely digested.
Only a few DRB OTUs were shared among the 12 ciliate species and no OTUs were common in these two strains of Condylostoma spathiosum, showing considerable assemblage variations (beta diversity) among ciliate species and even between populations, suggesting there is a high “species”-level diversity of DRB to be discovered. The null hypotheses that there were effects of habitat and isolation source of hosts on the DRB composition at the family level were all rejected. Despite this, we insist that it is premature to rule out the role of environment in structuring the DRB in these protists, as other ecological and physiological factors may not considered in the present study. Boenigk et al. (2001) found that preculture conditions influenced the ingestion and digestion process of HNFs. There are species-specific differences in, and an effect of physiological state on, ciliates’ gazing and digestive ability upon bacteria (Christaki et al., 1998; Weisse, 2002). Also rejected was the hypothesis on the effect of class-level taxonomy of ciliates. However, it should be noted that the two species of Pseudokeronopsis were placed closely to each other in the clustering plot, so were the two Condylostoma strains (Figure 6). This suggests the selection of DRB may be relatively stable at a lower taxonomic level (e.g., genus or species) of the hosts. Our last hypothesis, the DRB composition was different between free-living and endosymbiotic ciliates, was statistically supported. All these imply that the changes of DRB assemblages may be linked with both ecological traits of the host and the environment, but the key factors and their relative importance remain to be revealed.
Novel Diversity and Rare Taxa of Bacteria Associated with Ciliates
Among 40 bacterial OTUs we detected, about 43% (17 OTUs) had a closet match of 16S rRNA gene sequences of identified species available from the GenBank with a similarity < or = 97%, even for some culturable taxa such as Alteromonadaceae, Pseudoalteromonadaceae, Vibrionaceae, and Enterobacteriaceae. These indicate that there are a great number of yet-to-be-described bacteria associating with protists. Furthermore, our previous study demonstrated the association between OD1 bacteria and a freshwater ciliate (Gong et al., 2014). This uncultivated rare bacteria-protist association was observed again in the present study (e.g., OD1 and GN02), highlighting that many of these rare bacteria (also known as “microbial dark matter”) are of digestion-resistant and symbiotic nature in the environment (Brown et al., 2015; Nelson and Stegen, 2015).
Predominance of Gammaproteobacteria and Alphaproteobacteria in the Digestion-Resistant Bacteria Assemblages
Despite of considerable species-level variations, a structuring pattern of DRB at higher taxonomic ranks was evident: in terms of either the number of OTU or the relative abundance, members of Gammaproteobacteria and Alphaproteobacteria were the most abundant in the DRB assemblages. This result may more or less bias to ciliates of marine origin, since we studied more marine than freshwater ciliate species. However, a gammaproteobacterial OTU related to Aestuariibacter halophilus (Alteromonadaceae) comprised of 96% sequences in the clone library of the DRB of the freshwater P. aurelia, which is still supportive of the pattern mentioned above. Our result is in coincidence with two studies demonstrating a predominance of gamma- and alphaproteobacteria in bacterial 16S rRNA gene sequences recovered from a number of heterotrophic and mixotrophic marine protists (mostly flagellates) (Martinez-Garcia et al., 2012), and in the marine ciliate Euplotes focardii (Pucciarelli et al., 2015). Although their experiments were not designed for studying DRB in these protists, the procedures of sample preparation and cell sorting by flow cytometry must have taken some time, which allowed these protistan cells to digest bacteria in their food vacuoles, so that the digestion-resistant populations were more likely retained and detected. As such, the resistance of gamma- and alphaproteobacteria to digestion seems to be applicable for a broad range of marine protistan grazers, which may reflect some biological properties of these two groups of marine origin.
The resistance of marine gamma- and alphaproteobacterial to protistan digestion may be related to their protein secretion systems. The secretion systems in Gram-negative bacteria are often important virulence factors, comprising a diversity of proteinaceous machines to translocate secreted proteins from cell cytosol to the extracellular space or across the host cell membrane. Among the known six general types of these systems in Gram-negative bacteria, only the types III, IV and VI secretion systems (T3SS, T4SS, and T6SS) can deliver proteins further across the plasma membrane of the host in a contact-dependent manner (Tseng et al., 2009). The T4SS are capable of transporting DNA in addition to proteins into many eukaryotic cells and bacteria, suppressing host’s defense mechanisms and facilitating intracellular colonization (Christie and Cascales, 2005). Some bacteria (e.g., Legionella pneumophila) require the T4SS to become resistant to lysosomal degradation (Shintani and Klionsky, 2004). Using the amoeba Dictyostelium discoideum as a model host system, the T6SS was first identified in Vibrio cholera (Pukatzki et al., 2006). The genes encoding T6SS components are widely present in about 25% sequenced genomes of bacteria, mostly of proteobacterial pothogens (e.g., Pseudomonas aeruginosa, Francisella tularensis, and Burkholderia mallei), with the gammaproteobacteria being the most widely representated (Shrivastava and Mande, 2008). According to a metagenomic survey of virulent genes in marine bacteria, among all detected secretion systems, the T6SS and T4SS were the most abundant, and were mostly found in gamma- and alphaproteobacteria, respectively (Persson et al., 2009). Recent studies suggested that T6SS may play an important role in promoting a symbiotic relationship between some bacteria and mammals (Chow and Mazmanian, 2010), and in mediating communication between bacteria and eukaryotic hosts (Jani and Cotter, 2010). Besides T6SS or T4SS, the genomes of several marine Gamma- and Alphaproteobacteria strains possess type III secretion systems (T3SS) (Persson et al., 2009), which has been demonstrated to promote survival of V. parahaemolyticus in the interaction with diverse protists (Matz et al., 2011).
Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae as the Most Frequently Occurring Digestion-Resistant Bacteria in Ciliated Protists
In this study, rice grains were added to enrich bacterial prey to maintain seven ciliated species (Table 1), in which DRB phylotypes affiliated with Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae appeared much more abundant than those in the ciliates obtained directly from field samples. This suggests that the abundance of these digestion-resistant gammaproteobacterial lineages is probably related to the addition of a carbon source. In fact, previous studies demonstrated that the enrichment of dissolved organic matter selected for these Gammaproteobacteria lineages (Allers et al., 2007; Alonso-Saez et al., 2009; McCarren et al., 2010; Kelly et al., 2014). While Gammaproteobacteria lineages might resist grazing by nanoflagellates in the microcosms enriched with glucose by forming filamentation and aggregation (Alonso-Saez et al., 2009), the large particle size seems not a problem for most ciliate species investigated in this study. For example, the cultured hypotrich ciliates (class Spirotrichea, e.g., Diophrys scutum, Hemigastrostyla elongata, Pseudokeronopsis carnea, and Ps. flava) have large cell sizes (150–250 μm in length) and wide oral openings (40–60 μm), which facilitate engulfment of large-sized particles including bacterial filaments, aggregates and dinoflagellate, heterotrophic flagellate, and diatom species. Moreover, when the nanoflagellate grazers are eaten by the ciliates, indigested bacteria in nanoflagellates may retain in ciliate cells. Therefore, the abundance of Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae as DRB in ciliates is likely related to the eutrophic condition during cultivation of ciliates, and may reflect the DRB in the microbial food webs of the studied systems.
Our finding on the indigestibility of Alteromonadaceae, Pseudoalteromonadaceae, and Vibrionaceae is relevant to, and may be accountable for, some previously observed ecological phenomena. Members of these groups are well known for pursuing a “feast-or-famine” growth strategy in marine bacterioplankton, that is, they are able to maintain high ribosome levels during starvation and become rapidly enriched when organic matter are amended, but usually rare in situ (Eilers et al., 2000). In marine confinement experiments, Schäfer et al. (2000) found that Alteromonas-like phylotypes dominated the bacterial assemblages during and after the peak of protistan grazing pressure; they supposed that these populations perhaps have an inedible morphotype to survive protistan grazing. Their suggestion is apparently supported by our finding that these gammaproteobacteria can be ingested but may not be digested by protistan grazers. However, Beardsley et al. (2003) observed that an overproportional decline of Alteromonas, Pseudoalteromonas, and Vibrio during the phase of HNF regrowth. A possible explanation is that the indigestibility of these bacteria might be species- or strain-dependent, as ecotypes of Alteromonas or Pseudoalteromonas could have substantially different gene content and metabolic potentials (Ivars-Martinez et al., 2008; Qin et al., 2011), and it has been shown that the T6SS can be horizontally transferred between some marine vibrios (Salomon et al., 2015). Alternatively, if these phylotypes were indeed indigestible, then their massive mortality after blooming could be due to the viral lysis, which is another important top–down control on bacterial mortality. Both these two hypotheses are needed to be further tested in order to better understand the causes and the consequences of bacterial indigestibility in a microbial loop context.
Activity of Digestion-Resistant Bacteria
The FISH assays targeting bacterial rRNA genes illustrated that at least some of these DRB were still active after starvation. Nevertheless, it should be cautious that there was also diffuse fluorescence of Cy3-probe signals inside some ciliate specimens, indicating non-bacterial-cell bindings of targeted rRNA molecules. This is probably due to the remaining of rRNA of digested bacterial cells. According to Arsenis et al. (1970) who comparatively studied the degradation of DNA and RNA by lysosomal extracts, the complete degradation of RNA was considerably slower (within 60 h) than that of DNA (within 10 h). Considering these analogous situations, we assume that, although morphology of bacterial cells were not recognizable in some intracellular regions, their RNA (including ribosomal RNA) might not be completely degraded during the treatment of ciliate cells for 12 to 24 h, thus gave rise to the diffuse fluorescence after FISH. This may cause some biases on inferring activity of DRB in ciliates. Both diffuse rRNA-targeted Cy3 signals and rDNA-based identity of Alteromonadaceae were obtained from Paramecium aurelia (Figures 7G–I), suggesting the indigestibility of some DRB species might be strain-or phylotypes-dependent. This recalls intra-species variations of genomic contents and inter-species horizontal transfer of genes encoding protein secretion systems (Ivars-Martinez et al., 2008; Qin et al., 2011; Salomon et al., 2015). Further studies using more dedicated tools are needed to reveal their activity and function inside protistan cells and to assess the stain-level capability of digestion resistance.
Conclusion
Effective protistan grazing on bacteria relies on the success of two successive steps, ingestion and digestion, of which the latter has been rarely studied for nanoflagellates and ciliates from an ecological perspective. The causes and consequences of indigestibility of bacteria in the microbial loop remain elusive. For the first time, we explicitly characterized the ingested but not digested bacteria in a range of ciliate species. Our study reveals a snapshot of DRB diversity and structure, in which the number of investigated protists is apparently lower in comparison with the known enormous protistan diversity in nature. Despite this, we found a collectively high phylotype richness and large composition variations of DRB among protistan species, supporting the previous hypothesis that resistance to digestion is a widespread mechanism in natural bacteria (Jürgens and Güde, 1994). Previously, several structural and food quality traits of bacteria have been suggested for bacterial indigestibility. We are the first to find indications of a taxonomical strait of DRB of marine origin and suggest that it may be relevant to the putative symbiosis-promoting secretion systems that exist widely in the genomes of marine bacteria.
The findings of the high indigestibility of several opportunistic and copiotrophic groups not only provide a clue to explain that the mortality of these rapid growing bacteria in marine bacterioplankton, but also raise many further ecological questions. For example, given that there is a high diversity of bacteria capable of escaping protistan predation, why many of these bacterial winners are usually not abundant in the marine bacterioplankton? Does viral lysis play a significant role in killing these DRB? Did we overestimate protistan bacteriovory (feeding/grazing rates) by some traditional approaches, in which short-time uptakes of fluorescent labeled beads or model bacterial strains were estimated, assuming all the ingested fluorescent prey surrogates will be digested? How do the presence and expression of bacterial secretion systems impact the microbial ecology and function? It seems that indigestibility or limited digestibility of bacterial species is among the key factors regulating the interactions among bacteria, protists, and viruses. In the near future, multi-disciplinary approaches may be used to unveil the molecular mechanisms and test the ecological hypotheses concerning DRB in diverse and changing ecosystems.
Author Contributions
JG and YQ conceived and designed the experiments; YQ and SZ performed the experiments; JG, YQ, SZ, RF, LS, XZ, and QZ analyzed data; JG, YQ, SZ, XZ, and QZ contributed to reagents/materials; JG and YQ wrote the paper.
Funding
This work was supported by the Natural Science Foundation of China (Nos. 41176143 and 41522604), the Strategic Priority Research Program of CAS (No. XDA11020702), the Natural Science Foundation for Distinguished Young Scholars of Shandong (No. JQ201210), the Science and Technology Development Program of Yantai (No. 2014ZH073), and the Yantai Double Hundred Talent Plan.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
Thanks are due to two reviewers who provided constructive comments to improve our manuscript.
Footnotes
References
Allers, E., Gómez-Consarnau, L., Pinhassi, J., Gasol, J. M., Šimek, K., and Pernthaler, J. (2007). Response of Alteromonadaceae and Rhodobacteriaceae to glucose and phosphorus manipulation in marine mesocosms. Environ. Microbiol. 9, 2417–2429. doi: 10.1111/j.1462-2920.2007.01360.x
Alonso-Saez, L., Unanue, M., Latatu, A., Azua, I., Ayo, B., Artolozaga, I., et al. (2009). Changes in marine prokaryotic community induced by varying types of dissolved organic matter and subsequent grazing pressure. J. Plankton Res. 31, 1373–1383. doi: 10.1093/plankt/fbp081
Amann, R. I., Ludwig, W., and Schleifer, K.-H. (1995). Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol. Rev. 59, 143–169.
Apple, J. K., Strom, S. L., Palenik, B., and Brahamsha, B. (2011). Variability in protist grazing and growth on different marine Synechococcus isolates. Appl. Environ. Microbiol. 77, 3074–3084. doi: 10.1128/AEM.02241-10
Arsenis, C., Gordon, J. S., and Touster, O. (1970). Degradation of nucleic acids by lysosomal extracts of rat liver and Ehrlich ascites tumor cells. J. Biol. Chem. 245, 205–211.
Azam, F., Fenchel, T., Field, J. G., Gray, J. S., Meyer-Reil, L. A., and Thingstad, F. (1983). The ecological role of water-column microbes in the sea. Mar. Ecol. Prog. Ser. 10, 257–263. doi: 10.3354/meps010257
Barker, J., and Brown, M. R. W. (1994). Trojan horses of the microbial world: protozoa and the survival of bacterial pathogens in the environment. Microbiology 140, 1253–1259. doi: 10.1099/00221287-140-6-1253
Beardsley, C., Pernthaler, J., Wosniok, W., and Amann, R. (2003). Are readily culturable bacteria in coastal North Sea waters suppressed by selective grazing mortality? Appl. Environ. Microbiol. 69, 2624–2630. doi: 10.1128/AEM.69.5.2624-2630.2003
Boenigk, J., Matz, C., Jürgens, K., and Arndt, H. (2001). The influence of preculture conditions and food quality on the ingestion and digestion process of three species of heterotrophic nanoflagellates. Microb. Ecol. 42, 168–176.
Brandl, M. T., Rosenthal, B. M., Haxo, A. F., and Berk, S. G. (2005). Enhanced survival of Salmonella enterica in vesicles released by a soilborne Tetrahymena species. Appl. Environ. Microbiol. 71, 1562–1569. doi: 10.1128/AEM.71.3.1562-1569.2005
Brown, C. T., Hug, L. A., Thomas, B. C., Sharon, I., Castelle, C. J., Singh, A., et al. (2015). Unusual biology across a group comprising more than 15% of domain Bacteria. Nature 523, 208–211. doi: 10.1038/nature14486
Chow, J., and Mazmanian, S. K. (2010). A pathobiont of the microbiota balances host colonization and intestinal inflammation. Cell Host Microbe 7, 265–276. doi: 10.1016/j.chom.2010.03.004
Christaki, U., Dolan, J. R., Pelegri, S., and Rassoulzadegan, F. (1998). Consumption of picoplankton-size particles by marine ciliates: effects of physiological state of the ciliate and particle quality. Limnol. Oceanogr. 43, 458–464. doi: 10.4319/lo.1998.43.3.0458
Christie, P. J., and Cascales, E. (2005). Structural and dynamic properties of bacterial type IV secretion systems (review). Mol. Membr. Biol. 22, 51–61. doi: 10.1080/09687860500063316
Cole, J. R., Wang, Q., Cardenas, E., Fish, J., Chai, B., Farris, R. J., et al. (2009). The ribosomal database project: improved alignments and new tools for rRNA analysis. Nucl. Acids Res. 37, 141–145. doi: 10.1093/nar/gkn879
Daims, H., Brühl, A., Amann, R., Schleifer, K. H., and Wagner, M. (1999). The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 22, 434–444. doi: 10.1016/S0723-2020(99)80053-8
Eilers, H., Pernthaler, J., and Amann, R. (2000). Succession of pelagic marine bacteria during enrichment: a close look at cultivation-induced shifts. Appl. Environ. Microbiol. 66, 4634–4640. doi: 10.1128/AEM.66.11.4634-4640.2000
Ferla, M. P., Thrash, J. C., Giovannoni, S. J., and Patrick, W. M. (2013). New rRNA gene-based phylogenies of the Alphaproteobacteria provide perspective on major groups, mitochondrial ancestry and phylogenetic instability. PLoS ONE 8:e83383. doi: 10.1371/journal.pone.0083383
First, M. R., Park, N. Y., Berrang, M. E., Meinersmann, R. J., Bernhard, J. M., Gast, R. J., et al. (2012). Ciliate ingestion and digestion: flow cytometric measurements and regrowth of a digestion-resistant Campylobacter jejuni. J. Eukaryot. Microbiol. 59, 12–19. doi: 10.1111/j.1550-7408.2011.00589.x
Fried, J., Ludwig, W., Psenner, R., and Schleifer, K. H. (2002). Improvement of ciliate identification and quantification: a new protocol for fluorescence in situ hybridization (FISH) in combination with silver stain techniques. Syst. Appl. Microbiol. 25, 555–571. doi: 10.1078/07232020260517706
Fuchs, B. M., Spring, S., Teeling, H., Quast, C., Wulf, J., Schattenhofer, M., et al. (2007). Characterization of a marine gammaproteobacterium capable of aerobic anoxygenic photosynthesis. Proc. Natl. Acad. Sci. U.S.A. 104, 2891–2896. doi: 10.1073/pnas.0608046104
Gong, J., Qing, Y., Guo, X., and Warren, A. (2014). “Candidatus Sonnebornia yantaiensis,” a member of the candidate division OD1, as intracellular bacteria of the ciliated protist Paramecium bursaria (Ciliophora, Oligohymenophorea). Syst. Appl. Microbiol. 37, 35–41. doi: 10.1016/j.syapm.2013.08.007
Gonzalez, J. M., Sherr, E. B., and Sherr, B. F. (1990). Size-selective grazing on bacteria by natural assemblages of estuarine flagellates and ciliates. Appl. Environ. Microbiol. 56, 583–589.
Graeber, I., Kaesler, I., Borchert, M. S., Dieckmann, R., Pape, T., Lurz, R., et al. (2008). Spongiibacter marinus gen. nov., sp. nov., a halophilic marine bacterium isolated from the boreal sponge Haliclona sp. 1. Int. J. Syst. Evol. Microbiol. 58, 585–590. doi: 10.1099/ijs.0.65438-0
Greub, G., and Raoult, D. (2004). Microorganisms resistant to free-living amoebae. Clin. Microbiol. Rev. 17, 413–433. doi: 10.1128/CMR.17.2.413-433.2004
Gruber, D. F., Tuorto, S., and Taghon, G. L. (2009). Growth phase and elemental stoichiometry of bacterial prey influences ciliate grazing selectivity. J. Eukaryot. Microbiol. 56, 466–471. doi: 10.1111/j.1550-7408.2009.00428.x
Hahn, M. W., and Höfle, M. G. (2001). Grazing of protozoa and its effect on populations of aquatic bacteria. FEMS Microbiol. Ecol. 35, 113–121. doi: 10.1111/j.1574-6941.2001.tb00794.x
Huber, T., Faulkner, G., and Hugenholtz, P. (2004). Bellerophon: a program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 20, 2317–2319. doi: 10.1093/bioinformatics/bth226
Ivars-Martinez, E., Martin-Cuadrado, A. B., D’Auria, G., Mira, A., Ferriera, S., Johnson, J., et al. (2008). Comparative genomics of two ecotypes of the marine planktonic copiotroph Alteromonas macleodii suggests alternative lifestyles associated with different kinds of particulate organic matter. ISME J. 2, 1194–1212. doi: 10.1038/ismej.2008.74
Jani, A. J., and Cotter, P. A. (2010). Type VI secretion: not just for pathogenesis anymore. Cell Host Microbe 8, 2–6. doi: 10.1038/ismej.2008.74
Jousset, A. (2012). Ecological and evolutive implications of bacterial defences against predators. Environ. Microbiol. 14, 1830–1843. doi: 10.1111/j.1462-2920.2011.02627.x
Jürgens, K., and Güde, H. (1994). The potential importance of grazing-resistant bacteria in planktonic systems. Mar. Ecol. Prog. Ser. 112, 169–188. doi: 10.3354/meps112169
Jürgens, K., and Matz, C. (2002). Predation as a shaping force for the phenotypic and genotypic composition of planktonic bacteria. Anton. Van Leeuwenhoek. 81, 413–434. doi: 10.1023/A:1020505204959
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kelly, L. W., Williams, G. J., Barott, K. L., Carlson, C. A., Dinsdale, E. A., Edwards, R. A., et al. (2014). Local genomic adaptation of coral reef-associated microbiomes to gradients of natural variability and anthropogenic stressors. Proc. Natl. Acad. Sci. U.S.A. 111, 10227–10232. doi: 10.1073/pnas.1403319111
Koval, S. F. (1993). “Predation on bacteria possessing S-layers,” in Advances in Bacterial Paracrystalline Surface Layers, eds T. J. Beveridge and S. F. Koval (New York, NY: Plenum Publishing Corporation), 85–92. doi: 10.1007/978-1-4757-9032-0_9
Lane, D. J. (1991). “16S/23S rRNA sequencing,” in Nucleic Acid Techniques in Bacterial Systematics, eds E. Stackebrandt and M. Goodfellow (Chichester: John Wiley & Sons), 115–175.
Manz, W., Amann, R., Ludwig, W., Wagner, M., and Schleifer, K. H. (1992). Phylogenetic oligodeoxynucleotide probes for the major subclasses of Proteobacteria: problems and solutions. Syst. Appl. Microbiol. 15, 593–600. doi: 10.1016/S0723-2020(11)80121-9
Martinez-Garcia, M., Brazel, D., Poulton, N. J., Swan, B. K., Gomez, M. L., Masland, D., et al. (2012). Unveiling in situ interactions between marine protists and bacteria through single cell sequencing. ISME J. 6, 703–707. doi: 10.1038/ismej.2011.126
Matz, C., Deines, P., Boenigl, J., Arndt, H., Eberl, L., Kjelleberg, S., et al. (2004). Impact of violacein-producing bacteria on survival and feeding of bacterivorous nanoflagellates. Appl. Environ. Microbiol. 70, 1593–1599. doi: 10.1128/AEM.70.3.1593-1599.2004
Matz, C., and Kjelleberg, S. (2005). Off the hook – how bacteria survive protozoan grazing. Trends Microbiol. 13, 302–307. doi: 10.1016/j.tim.2005.05.009
Matz, C., Nouri, B., McCarter, L., and Martinez-Urtaza, J. (2011). Acquired type III secretion system determines environmental fitness of epidemic Vibrio parahaemolyticus in the interaction with bacterivorous protists. PLoS ONE 6:e20275. doi: 10.1371/journal.pone.0020275
McCarren, J., Becker, J. W., Repeta, D. J., Shi, Y., Young, C. R., Malmstrom, R. R., et al. (2010). Microbial community transcriptomes reveal microbes and metabolic pathways associated with dissolved organic matter turnover in the sea. Proc. Natl. Acad. Sci. U.S.A. 107, 16420–16427. doi: 10.1073/pnas.1010732107
Montagnes, D. J. S., Barbosa, A. B., Boenigk, J., Davidson, K., Jürgens, K., Macek, M., et al. (2008). Selective feeding behaviour of key free-living protists: avenues for continued study. Aquat. Microb. Ecol. 53, 83–98. doi: 10.3354/ame01229
Nelson, W. C., and Stegen, J. C. (2015). The reduced genomes of Parcubacteria (OD1) contain signatures of a symbiotic lifestyle. Front. Microbiol. 6:713. doi: 10.3389/fmicb.2015.00713
Odaka, C., and Mizuochi, T. (1999). Role of macrophage lysosomal enzymes in the degradation of nucleosomes of apoptotic cells. J. Immunol. 163, 5346–5352.
Pernthaler, J. (2005). Predation on prokaryotes in the water column and its ecological implications. Nat. Rev. Microbiol. 3, 537–546. doi: 10.1038/nrmicro1180
Persson, O. P., Pinhassi, J., Riemann, L., Marklund, B.-I., Rhen, M., Normark, S., et al. (2009). High abundance of virulence gene homologues in marine bacteria. Environ. Microbiol. 11, 1348–1357. doi: 10.1111/j.1462-2920.2008.01861.x
Posch, T., Jezbera, J., Vrba, J., Šimek, K., Pernthaler, J., Andreatta, S., et al. (2001). Size selective feeding in Cyclidium glaucoma (Ciliophora, Scuticociliatida) and its effects on bacterial community structure: a study from a continuous cultivation system. Microb. Ecol. 42, 217–227. doi: 10.1007/s002480000114
Price, M. N., Dehal, P. S., and Arkin, A. P. (2010). FastTree 2 – Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 5:e9490. doi: 10.1371/journal.pone.0009490
Pruesse, E., Peplies, J., and Glöckner, F. O. (2012). SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829. doi: 10.1093/bioinformatics/bts252
Pucciarelli, S., Devaraj, R. R., Mancini, A., Ballarini, P., Castelli, M., Schrallhammer, M., et al. (2015). Microbial consortium associated with the Antarctic marine ciliate Euplotes focardii: an investigation from genomic sequences. Microb. Ecol. 70, 484–497. doi: 10.1007/s00248-015-0568-9
Pukatzki, S., Ma, A. T., Sturtevant, D., Krastins, B., Sarracino, D., Nelson, W. C., et al. (2006). Identification of a conserved bacterial protein secretion system in Vibrio cholerae using the Dictyostelium host model system. Proc. Natl Acad. Sci. U.S.A. 103, 1528–1533. doi: 10.1073/pnas.0510322103
Qin, Q. L., Li, Y., Zhang, Y. J., Zhou, Z. M., Zhang, W. X., Chen, X. L., et al. (2011). Comparative genomics reveals a deep-sea sediment-adapted life style of Pseudoalteromonas sp. SM9913. ISME J. 5, 274–284. doi: 10.1038/ismej.2010.103
Ronquist, F., and Huelsenbeck, J. P. (2003). MRBAYES 3: bayesian phylogenetic inference under mixed models. Bioinformatics 19, 1572–1574. doi: 10.1093/bioinformatics/btg180
Salomon, D., Klimko, J. A., Trudgian, D. C., Kinch, L. N., Grishin, N. V., Mirzaei, H., et al. (2015). Type VI secretion system toxins horizontally shared between marine bacteria. PLoS Pathog. 11:e1005128. doi: 10.1371/journal.ppat.1005128
Schäfer, H., Servais, P., and Muyzer, G. (2000). Successional changes in the genetic diversity of a marine bacterial assemblage during confinement. Arch. Microbiol. 173, 138–145. doi: 10.1007/s002039900121
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introducing Mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Sherr, B. F., Sherr, E. B., and McDaniel, J. (1992). Effect of protistan grazing on the frequency of dividing cells in bacterioplankton assemblages. Appl. Environ. Microbiol. 58, 2381–2385.
Sherr, E. B., and Sherr, B. F. (2002). Significance of predation by protists in aquatic microbial food webs. Antonie Van Leeuwenhoek 81, 293–308. doi: 10.1023/A:1020591307260
Shintani, T., and Klionsky, D. J. (2004). Autophagy in health and disease: a double-edged sword. Science 306, 990–995. doi: 10.1126/science.1099993
Shrivastava, S., and Mande, S. S. (2008). Identification and functional characterization of gene components of Type VI secretion system in bacterial genomes. PLoS ONE 3:e2955. doi: 10.1371/journal.pone.0002955
Šimek, K., Kasalický, V., Jezbera, J., Horňák, K., Nedoma, J., Hahn, M. W., et al. (2013). Differential freshwater flagellate community response to bacterial food quality with a focus on Limnohabitans bacteria. ISME J. 7, 1519–1530. doi: 10.1038/ismej.2013.57
Song, W., Warren, A., and Hu, X. (2009). Free-living Ciliates in the Bohai and Yellow Seas, China. Beijing: Science Press.
Suzuki, T., Nakamura, T., and Fuse, H. (2012). Isolation of two novel marine ethylene-assimilating bacteria, Haliea species ETY-M and ETY-NAG, containing particulate methane monooxygenase-like genes. Microbes Environ. 27, 54–60. doi: 10.1264/jsme2.ME11256
Tseng, T.-T., Tyler, B. M., and Setubal, J. C. (2009). Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol. 9:S2. doi: 10.1186/1471-2180-9-S1-S2
Weisse, T. (2002). The significance of inter- and intraspecific variation in bacterivorous and herbivorous protists. Antonie Van Leeuwenhoek 81, 327–341. doi: 10.1023/A:1020547517255
Yan, S., Fuchs, B. M., Lenk, S., Harder, J., Wulf, J., Jiao, N. Z., et al. (2009). Biogeography and phylogeny of the NOR5/OM60 clade of Gammaproteobacteria. Syst. Appl. Microbiol. 32, 124–139. doi: 10.1016/j.syapm.2008.12.001
Yilmaz, L. S., Parnerkar, S., and Noguera, D. R. (2011). mathFISH, a web tool that uses thermodynamics-based mathematical models for in silico evaluation of oligonucleotide probes for fluorescence in situ hybridization. Appl. Environ. Microbiol. 77, 1118–1122. doi: 10.1128/AEM.01733-10
Keywords: bacterial symbiosis, grazing-resistant bacteria, microbial interactions, protein secretion systems, top–down effect
Citation: Gong J, Qing Y, Zou S, Fu R, Su L, Zhang X and Zhang Q (2016) Protist-Bacteria Associations: Gammaproteobacteria and Alphaproteobacteria Are Prevalent as Digestion-Resistant Bacteria in Ciliated Protozoa. Front. Microbiol. 7:498. doi: 10.3389/fmicb.2016.00498
Received: 22 January 2016; Accepted: 27 March 2016;
Published: 11 April 2016.
Edited by:
Dennis A. Bazylinski, University of Nevada, Las Vegas, USACopyright © 2016 Gong, Qing, Zou, Fu, Su, Zhang and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jun Gong, amdvbmdAeWljLmFjLmNu