 Carla Rodrigues
Carla Rodrigues Jan Bavlovič2,3
Jan Bavlovič2,3 Luísa Peixe
Luísa Peixe Ângela Novais
Ângela Novais- 1UCIBIO/REQUIMTE, Laboratório de Microbiologia, Faculdade de Farmácia, Universidade do Porto, Porto, Portugal
- 2Faculty of Pharmacy in Hradec Králové, Charles University, Prague, Czech Republic
- 3Faculty of Military Health Sciences, University of Defense, Brno, Czech Republic
- 4FP-ENAS/CEBIMED, Faculdade de Ciências da Saúde, Universidade Fernando Pessoa, Porto, Portugal
- 5Botelho Moniz Análises Clínicas, Santo Tirso, Portugal
KPC-3-producing bacteria are endemic in many countries but only recently became apparent their wide distribution in different Portuguese hospitals. The aim of this study is to characterize genetic backgrounds associated with blaKPC−3 among Klebsiella pneumoniae isolates recently identified on non-hospitalized patients in Portugal. Twenty KPC-producing K. pneumoniae identified between October 2014 and November 2015 in three different community laboratories were characterized. Isolates were mainly from patients from long-term care facilities (n = 11) or nursing homes (n = 6), most of them (75%) previously hospitalized in different Portuguese hospitals. Standard methods were used for bacterial identification and antibiotic susceptibility testing. Carbapenemase production was assessed by the Blue-Carba test, and identification of bla genes was performed by PCR and sequencing. Epidemiological features of KPC-producing K. pneumoniae included population structure (XbaI-PFGE, MLST and wzi sequencing), genetic context (mapping of Tn4401), and plasmid (replicon typing, S1-PFGE, and hybridization) analysis. All K. pneumoniae isolates produced KPC-3, with two MDR K. pneumoniae epidemic clones representing 75% of the isolates, namely ST147 (wzi64/K14.64, February–November 2015) and ST15 (two lineages exhibiting capsular types wzi19/K19 or wzi93/K60, July-November 2015). Other sporadic clones were detected: ST231 (n = 3; wzi104), ST348 (n = 1; wzi94) and ST109 (n = 1, wzi22/K22.37). blaKPC−3 was identified within Tn4401d in all isolates, located in most cases (80%) on cointegrated plasmids (repAFIA+repAFII+oriColE1;105-250 kb) or in 50 kb IncN plasmids. In conclusion, this study highlights a polyclonal structure of KPC-3-producing K. pneumoniae and the predominance of the ST147 clone among non-hospitalized patients in Portugal, linked to platforms still unnoticed in Europe (blaKPC−3-Tn4401d-IncFIA) or firstly reported (blaKPC−3-Tn4401d-IncN). This scenario underlines the recent penetration of successful mobile genetic elements in previously circulating MDR K. pneumoniae lineages (mainly ST147 and ST15) in Portugal, rather than the importation of the global lineages from clonal group 258.
Introduction
In the last years, carbapenem-resistant Enterobacteriaceae have spread globally, being responsible for high rates of morbidity and mortality among healthcare-associated infections, mainly due to the depletion of effective therapeutic options (WHO, 2014; Albiger et al., 2015, http://www.cdc.gov/drugresistance/threat-report-2013/). After the first strain identified in 1996 in a North Carolina hospital (USA; Yigit et al., 2001), Klebsiella pneumoniae carbapenemases (KPCs) have exploded worldwide predominantly among K. pneumoniae isolates (Munoz-Price et al., 2013; Chen et al., 2014b). To date, 23 KPC variants (KPC-2 to KPC-24) have been described (http://www.lahey.org/Studies/other.asp#table1), being KPC-2 and KPC-3 the most widespread variants with variable geographic distribution (Munoz-Price et al., 2013; Nordmann and Poirel, 2014). While in some countries (USA, Colombia, Italy, and Israel) both KPC-2- and KPC-3-producing bacteria are endemic, in others (Argentina, Brazil, Greece, Poland, and China) KPC-2 producers are predominant (Munoz-Price et al., 2013; Albiger et al., 2015).
The blaKPC genes are commonly located on Tn4401, a 10 kb Tn3-like transposon delimited by two 39-bp imperfect inverted repeat sequences harboring blaKPC, transposase and resolvase genes, and insertion sequences ISKpn7 (upstream blaKPC) and ISKpn6 (downstream blaKPC; Chen et al., 2014b). It is recognized as a highly active transposon enhancing the spread of blaKPC genes to different plasmid scaffolds (Cuzon et al., 2011). To date, six Tn4401 isoforms have been described with variable deletions between ISKpn7 and blaKPC providing different promoter regions to the gene (a, −99 bp; b, no deletion; c, −215 bp; d, −68 bp; e, −255 bp; g, equal to isoform c but with one single nucleotide mutation on P2 promotor), and consequently different expression levels of the blaKPC gene (Naas et al., 2012; Chmelnitsky et al., 2014). Besides its genetic environment, other factors are known to have greatly contributed to the spread of KPC producers in many countries, leading to an increasing challenge in the design of effective infection control measures. First, the introduction and subsequent expansion of blaKPC−2 and blaKPC−3 on multidrug resistant (MDR) K. pneumoniae lineages from clonal group (CG) 258 [sequence types (ST) 11, 258, 512] (Munoz-Price et al., 2013; Chen et al., 2014b), followed in a few countries (e.g., Israel, Italy, Colombia) by subsequent dispersion to other clonal backgrounds (Baraniak et al., 2015; Bonura et al., 2015; Ocampo et al., 2015). Second, the acquisition of blaKPC by plasmids from different incompatibility groups (IncFIIK2, IncFIA, IncI2, IncN, IncX3, ColE), favored a quick intra- and inter-species dissemination (Chen et al., 2014b).
In Portugal, KPC-2 was identified only in an environmental Escherichia coli isolate in 2010 (Poirel et al., 2012), while KPC-3 producers were first detected in 2009 in a central hospital (Machado et al., 2010). However, only recently became evident the widespread distribution of KPC-3 among K. pneumoniae isolates in different Portuguese hospitals (Silva et al., unpublished data; Manageiro et al., 2015). In this study, we aim to trace the landscape of KPC-3-producing K. pneumoniae isolates recently identified outside hospital boundaries in Portugal by detailed characterization of clonal and plasmid genetic backgrounds.
Materials and Methods
Bacterial Isolates and Epidemiological Data
Thirty K. pneumoniae isolates showing reduced susceptibility to carbapenems were identified between October 2014 and November 2015 in three different community laboratories in the North of Portugal, one of them receiving samples from all over the country. Twenty of them were identified as KPC producers and further characterized in this study. They were detected in urine samples (n = 19) or sputum (n = 1) of patients between 61 and 89 years old (mean age = 83; 16 females, 4 males; Table 1). Most of these patients were institutionalized in long-term care facilities (LTCFs) [n = 11; 55%; six different LTCFs (A–F)] or nursing homes (NH) [n = 6; 30%; five different NH (A–E)], while some were identified in ambulatory (n = 3; 15%; Table 1). Most of them (n = 13; 65%) had been hospitalized in the previous month in different hospitals from the North or Centre of Portugal, although in three cases no hospitalization or older hospitalization events (4–9 months) were detected (Table 1). Travel history abroad was discarded for 60% of the patients (n = 12/20), or considered improbable for the remaining patients due to their clinical conditions (impaired mobilization and chronic underlying diseases).
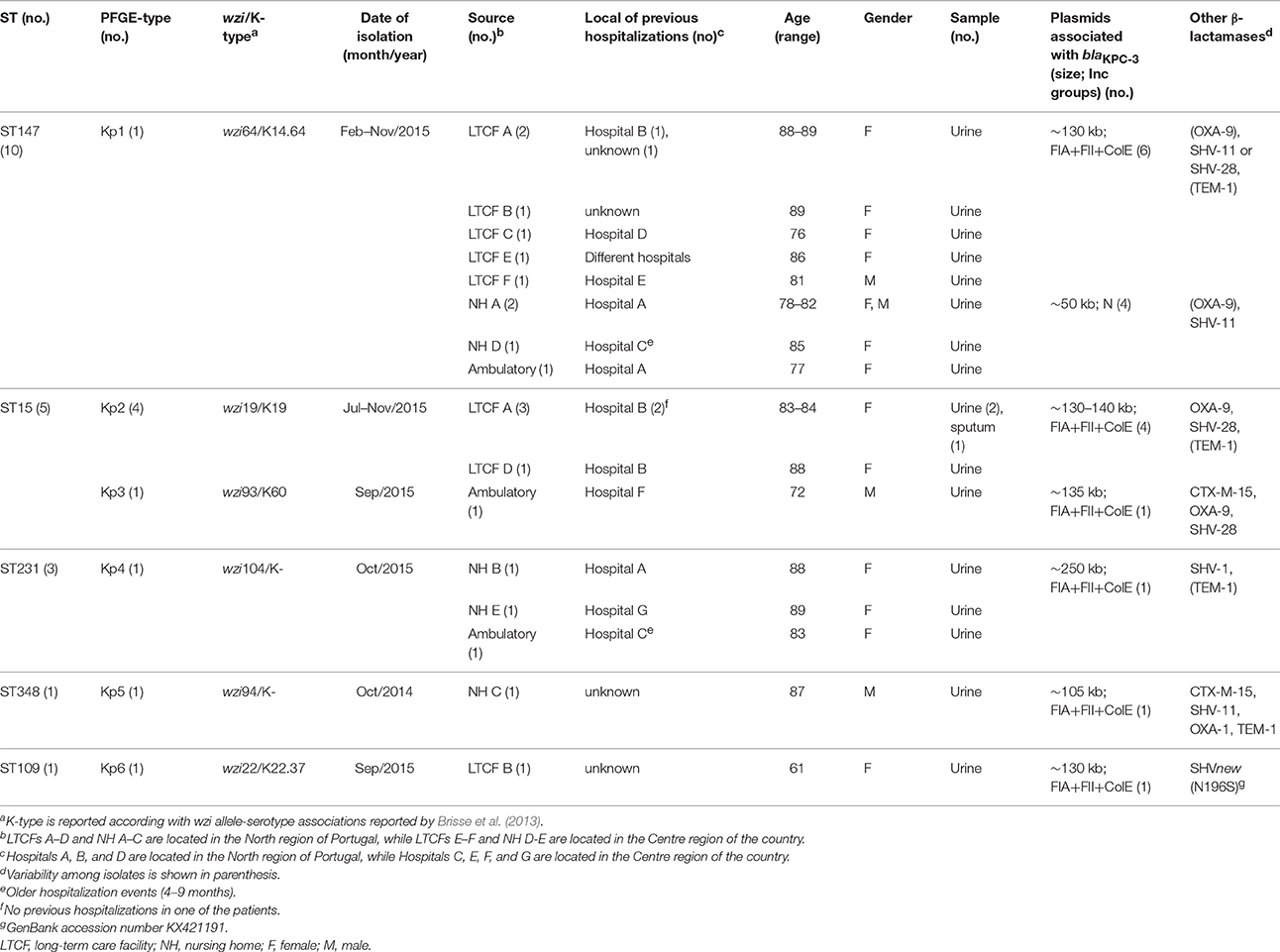
Table 1. Epidemiological data of K. pneumoniae isolates carrying blaKPC−3-Tn4401d identified in non-hospitalized patients in Portugal (October 2014–November 2015).
Antibiotic Resistance Phenotypes and Genotypes
Bacterial identification and preliminary antibiotic susceptibility testing were performed by Vitek II system (BioMérieux, Marcy l'Étoile, France). Confirmatory and additional tests for β-lactams (amoxicillin-clavulanic acid, mecillinam, cefoxitin, extended-spectrum cephalosporins, aztreonam, carbapenems), aminoglycosides (amikacin, gentamicin, netilmicin, tobramycin), fluoroquinolones (ciprofloxacin), folate pathway inhibitors (trimethoprim, trimethoprim-sulfamethoxazole), chloramphenicol, fosfomycin, and colistin were assessed by standard disc diffusion (Oxoid Ltd., Basingstoke, United Kingdom), agar dilution (for fosfomycin; in the presence of glucose-6-phosphate at 25 mg/L), broth microdilution (for colistin) or E-test (for carbapenems) (Liofilchem, Italy) methods according to EUCAST (www.eucast.org).
Production of carbapenemases was assessed by the Blue-Carba test (Pires et al., 2013), and identification of carbapenemases (blaNDM, blaVIM, blaIMP, blaKPC,blaOXA−48),or other bla genes (blaCTX−M, blaSHV, blaTEM, blaOXA) was performed by PCR and sequencing (Curiao et al., 2010; Bogaerts et al., 2013; Rodrigues et al., 2014).
Population Structure Analysis
Population structure characterization included XbaI-Pulsed-Field Gel Electrophoresis (PFGE) (electrophoresis conditions: 5–20 s for 4 h and 25–50 s for 18 h, 14°C, 6 V/cm2), and multi-locus sequence typing (MLST) (http://bigsdb.web.pasteur.fr/klebsiella/primers_used.html) in representative isolates, as described (Rodrigues et al., 2014). Molecular capsule typing was performed by PCR and sequencing of wzi gene (Brisse et al., 2013).
Characterization of the Genetic Environment and Location of blaKPC Genes
The genetic context of blaKPC−3 was investigated by PCR and sequencing targeting Tn4401 conserved sequences (Chen et al., 2014a). Location of bla (blaKPC−3, blaCTX−M−15) genes and plasmid characterization were assessed by S1- and I-CeuI-PFGE and identification of replication genes by PCR, sequencing and hybridization (Carattoli et al., 2005; García-Fernández et al., 2009; Villa et al., 2010; Chen et al., 2014a; Rodrigues et al., 2014).
Results
Carbapanemase Production and Variable Antibiotic Resistance Phenotypes
All isolates produced KPC-3 and demonstrated resistance or intermediate phenotypes to ertapenem (MIC = 1 to 16 mg/L), and susceptible, intermediate or resistance phenotypes to imipenem (MIC = 2 to 16 mg/L) and meropenem (MIC = 1 to 8 mg/L), with colonies growing within the inhibition zone of all carbapenems tested, a hetero-resistance phenotype usually observed for KPC producers (Nordmann et al., 2009; Table 2). Although for some isolates the MIC values for imipenem and meropenem were interpreted as susceptible by the clinical breakpoints defined by EUCAST, in all cases they were above the epidemiological cut-off values (ECOFFs) defined for K. pneumoniae (http://www.eucast.org/mic_distributions_and_ecoffs/; Table 2).
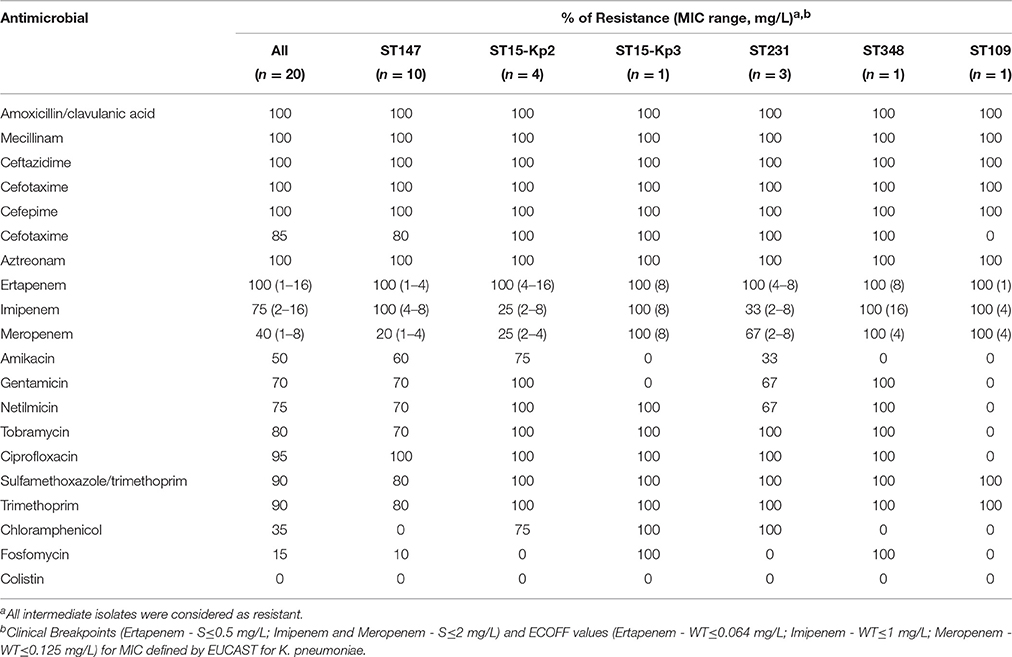
Table 2. Antimicrobial resistance patterns of KPC-3-producing K. pneumoniae clones.
All isolates were defined as multidrug resistant (MDR) in accordance with the definition of MDR for Enterobacteriaceae (non-susceptible to ≥1 agent in ≥3 antimicrobial categories; Magiorakos et al., 2012), although some of them exhibited a less extensive resistance profile to non-β-lactams, being resistant only to ciprofloxacin (Table 2). All isolates were susceptible to colistin (MIC = 0.25–2 mg/L; Table 2).
KPC-3 was Identified Among Locally Circulating K. pneumoniae Clones
KPC-3-producing K. pneumoniae isolates were assigned to six different PFGE-types (arbitrarily designated as Kp1 to Kp6), each one of them linked to a specific capsular type (Table 1). Most isolates belonged to ST147 carrying wzi64 (K14.64; n = 10 Kp1, 50%; detected between February to November 2015) and produced additionally SHV-11 (n = 9) or SHV-28 (n = 1), OXA-9 (n = 6) and/or TEM-1 (n = 1; Table 1). Some (n = 4) of these patients had recently been hospitalized in hospitals (A and D) where KPC-3-producing ST147 isolates exhibiting the same PFGE-type were detected (data not shown, Silva et al., unpublished data). ST15 was also frequent (n = 5, 25%), with two different clones being identified (n = 4 Kp2 carrying wzi19/K19; and n = 1 Kp3 carrying wzi93/K60) between July and November 2015. These isolates co-produced OXA-9 and SHV-28. All ST15-Kp2 were identified in patients from the same LTCF or region, and in one case there was no previous hospitalization (Table 2). ST231 isolates carrying wzi104 (n = 3 Kp4; October 2015) co-produced SHV-1 and TEM-1 (n = 1), and were identified in patients for which no epidemiological link could be established. Sporadic clones such as ST348 carrying wzi94 (n = 1 Kp5; October 2014) or ST109 carrying wzi22 (K22.37; n = 1 Kp6; September 2015) producing other β-lactamases were also detected (Table 1).
IncFIA and IncN Plasmids Involved in the Dissemination of blaKPC−3-Tn4401d
In all isolates, the blaKPC−3 was identified between ISKpn7 (upstream) and ISKpn6 (downstream), in a structure previously described as Tn4401 isoform d, known to have a 68bp deletion between ISKpn7 and blaKPC gene (Chen et al., 2014b). Isolates showed a variable number of plasmids (2–5 plasmids) with different sizes (40–500 kb), frequently from IncFIIK and IncR families. In most of the cases (n = 16/20 isolates from different clones), blaKPC−3 was located within cointegrated plasmids (105 to 250 kb) carrying repAFIA(100% identity with that of pBK30661 plasmid, GenBank accession number KF954759), repAFII (100% identity with that of pBK30683 plasmid, GenBank accession number KF954760), and oriColE1 (100% identity with ori p15 gene pKBuS13 plasmid, GenBank accession number KM076933). In the remaining isolates (n = 4/20 isolates belonging to ST147), blaKPC−3 was identified in a ca. 50 kb IncN plasmid [repN showing 100% identity with that of pKPC_FCF/3SP plasmid (defined by pMLST as repN allele 7; ST15), GenBank accession number CP004367] (Table 1). The two isolates for which a less extended resistance profile was observed (Table 2) carried blaKPC−3 within IncN plasmids and no additional IncF plasmids were observed. The blaCTX−M−15 (when present) was variably located in a ca. 200 kb-IncFIIK7 (ST348) or in a ca. 60 kb-IncR (ST15-Kp3) plasmid type.
Discussion
In this study, we highlight a polyclonal structure of KPC-3 producing K. pneumoniae isolates among patients outside hospital boundaries in Portugal consistent with nosocomial acquisition, and unveil novel or uncovered plasmid backbones carrying blaKPC−3 in Europe.
The first clinical cases of KPC-3 producers in Portugal were detected in 2009 in a pediatric unit of a hospital from the Lisbon and Tagus Valley region and involved 2 K. pneumoniae belonging to ST11 (Machado et al., 2010). Months later and until 2011, an outbreak involving 41 KPC-3-producing isolates, most of them (n = 29) assigned to ST14, was reported (Calisto et al., 2012). More recently, a nationwide study reported 22 K. pneumoniae producing KPC-3 (mainly ST11, ST14, ST15, and ST147 clones) in several hospitals between 2010 and 2013 but plasmid backgrounds had been poorly characterized (Manageiro et al., 2015). However, the situation concerning carbapenemase-producing Enterobacteriaceae in Portugal was only recently recognized in the EuSCAPE survey, where our country appeared in level 2b (sporadic hospital outbreaks) mainly due to the expansion of KPC producers (Albiger et al., 2015). The data presented in this study strengthens an ongoing dissemination of KPC-3 producers in Portugal, where the identification of these bacteria among such a high diversity of healthcare institutions other than hospitals might potentiate their impact for both hospital and community settings. It is thus advisable a reinforcement of infection control measures, surveillance, and tracking of isolates resistant to carbapenems in clinical institutions, and a coordinated action between clinicians, epidemiologists and national reference laboratories for guidance and harmonization of protocols (Albiger et al., 2015).
We observed a polyclonal structure of KPC-3-producing K. pneumoniae isolates, where most of the clones identified (ST15, ST147, ST348) exhibited the same PFGE-pattern as CTX-M-15 (ST348, ST15-Kp2, and -Kp3) or SHV-12 (ST147) producers previously involved in hospital- and community-acquired infections at least in the North region of Portugal (2010–2012; Rodrigues et al., unpublished data; Rodrigues et al., 2014). These “high-risk clones” have been linked to the worldwide expansion of different ESBL (CTX-M-15 and different SHV-types) and carbapenemases (KPC, VIM, NDM, OXA-48-like), including in Portugal (Rodrigues et al., 2014). This scenario suggests recent acquisition of blaKPC−3 by MDR K. pneumoniae genetic lineages that were already circulating in Portugal (ST15, ST147, ST231, ST348), a situation observed less frequently than the amplification of CG258 lineages (Baraniak et al., 2015; Bonura et al., 2015; Ocampo et al., 2015).
The ST147 clone [clonal group (CG) 147] exhibiting capsular type K14.K64 was identified in patients from diverse LTCFs and NHs for a long period of time and seems to be the predominant lineage among KPC-3-producing K. pneumoniae in different healthcare settings (Silva et al., unpublished data; Manageiro et al., 2015). In fact, identical KPC-3-producing ST147 isolates were recently involved in outbreaks in hospitals where some of the patients had been previously hospitalized. Although, nosocomial acquisition is the most probable origin for most KPC-3 producers identified in patients included in this study, it is of notice that in three cases no obvious hospitalization link could be established. Indeed, considering the frequent displacement of these patients between institutions (integrated network of LTCFs in Portugal) and hospitals (we had only access to the last hospitalization event) and that intestinal colonization might be persistent in time (Feldman et al., 2013), we cannot completely discard cross transmission events in the units analyzed.
The identification of two distinct ST15 (CG15) lineages in this study (ST15-K19 and ST15-K60) is in line with recent studies based on wzi-capsule typing unveiling the circulation of distinct lineages within this CG, that might have differences in their relative occurrence, geographical, or niche distribution and/or host susceptibility (Bialek-Davenet et al., 2014; Holt et al., 2015; Rodrigues et al., 2015; Zhou et al., 2015, http://bigsdb.web.pasteur.fr/klebsiella/klebsiella.html). The ST231 K. pneumoniae clone (CG231) had already been linked to GES-5 plus SHV-12 production in Portugal (Manageiro et al., 2015) and its association with community invasive infections (sepsis, lethal pneumonia, or meningitis), and high content in virulence and antimicrobial resistance genes have recently been highlighted (Holt et al., 2015). The ST109 clone is rarely reported (http://bigsdb.web.pasteur.fr/klebsiella/klebsiella.html) and it is described for the first time in Portugal. However, it belongs to the CG17, associated with the expansion of CTX-M-15 and different carbapenemases worldwide (Rodrigues et al., 2014; Holt et al., 2015).
The blaKPC−3 was linked to Tn4401d isoform in all characterized KPC-3-producing Enterobacteriaceae from Portugal (this study; Manageiro et al., 2015). In this study, we show that in most cases (80%) Tn4401d-blaKPC−3 was located within cointegrated FIA, FII, and ColE1 plasmids (105–250 kb; Table 1) corroborating the strong association between Tn4401d-blaKPC−3 with IncFIA plasmids pointed out previously in large collections from the USA (Chen et al., 2014a; Deleo et al., 2014; Bowers et al., 2015; Chavda et al., 2015). We detected FIA and FII replicons identical to those of pBK30683 plasmid (GenBank accession number KF954760) plus an additional oriColE1 gene identical to that of the ColE1 plasmid pKBuS13 (GenBank accession number KM076933), supporting the role of these mobilizable plasmids in the assembly of MDR plasmids (Chen et al., 2014a; Garbari et al., 2015). These and other (IncFIA plus IncA/C2 or IncFIA plus IncX3) cointegration forms seem to play an important role in the intra- and inter-species spread of carbapenem resistance genes (Chen et al., 2014a,b; Chavda et al., 2015). To the best of our knowledge, we unveil for the first time a cointegrate IncFIA platform carrying Tn4401d-blaKPC−3 in Europe, characterized and highly represented by far only in isolates from the USA (mainly among non-ST258 and non-K. pneumoniae isolates; Chen et al., 2014a). In the remaining isolates (20%, 4 ST147), blaKPC−3-Tn4401d was located on ca. 50 kb IncN plasmids, an association primarily described in this study.
In conclusion, this study highlights a polyclonal structure among KPC-3 producers identified in geographically dispersed non-hospitalized patients in Portugal, not always linked to nosocomial acquisition, a situation that deserves close monitoring due to its high clinical or epidemiological impact. In all cases, a common platform (blaKPC−3-Tn4401d) was identified in plasmids still unnoticed in Europe (blaKPC−3-Tn4401d-IncFIA) or firstly reported here (blaKPC−3-Tn4401d-IncN). Their identification in previously circulating MDR K. pneumoniae lineages in our area (ST147, ST15, ST231, ST348) underlines the recent penetration of successful mobile genetic elements in locally circulating clonal backgrounds, rather than the importation of the most common global lineages from CG258.
Author Contributions
CR and JB performed the experiments and contributed with the acquisition of molecular data. CR and ÂN wrote the article and performed analysis and interpretation of molecular data. EM and JA contributed with epidemiological data and revision of the manuscript. ÂN and LP contributed with the design of the study and final revision of the manuscript. All authors read and approved the final version of the manuscript.
Funding
This work received financial support from the European Union (FEDER funds) through Programa Operacional Factores de Competitividade – COMPETE and National Funds (FCT, Fundação para a Ciência e Tecnologia) (UID/Multi/04378/2013). ÂN and CR were supported by fellowships (grant number SFRH/BPD/104927/2014 and SFRH/BD/84341/2012, respectively) from FCT through Programa Operacional Capital Humano (POCH).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Carlos Mendes, Dr. Alexandra Gomes, and Dr. Bibiana Marques (Laboratório de Análises Clínicas Doutor Carlos Torres), Dr. Rui Campainha and Dr. Sofia Moniz (Botelho Moniz Análises Clínicas - BMAC), and Dr. Ricardo Silva (Santa Casa da Misericórdia, Vila do Conde) for providing the strains and the clinical/epidemiological data included in this study.
References
Albiger, B., Glasner, C., Struelens, M. J., Grundmann, H., Monnet, D. L. and European Survey of Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) working group (2015). Carbapenemase-producing Enterobacteriaceae in Europe: assessment by national experts from 38 countries, May 2015. Euro Surveill. 20:30062. doi: 10.2807/1560-7917.ES.2015.20.45.30062
Baraniak, A., Izdebski, R., Fiett, J., Herda, M., Derde, L. P. G., Bonten, M. J. M., et al. (2015). KPC-like carbapenemase-producing Enterobacteriaceae colonizing patients in Europe and Israel. Antimicrob. Agents Chemother. 60, 1912–1917. doi: 10.1128/AAC.02756-15
Bialek-Davenet, S., Criscuolo, A., Ailloud, F., Passet, V., Jones, L., Delannoy-Vieillard, A.-S., et al. (2014). Genomic definition of hypervirulent and multidrug-resistant Klebsiella pneumoniae clonal groups. Emerg. Infect. Dis. 20, 1812–1820. doi: 10.3201/eid2011.140206
Bogaerts, P., Rezende de Castro, R., de Mendonça, R., Huang, T.-D., Denis, O., and Glupczynski, Y. (2013). Validation of carbapenemase and extended-spectrum β-lactamase multiplex endpoint PCR assays according to ISO 15189. J. Antimicrob. CheMother. 68, 1576–1582. doi: 10.1093/jac/dkt065
Bonura, C., Giuffrè, M., Aleo, A., Fasciana, T., Di Bernardo, F., Stampone, T., et al. (2015). An update of the evolving epidemic of blaKPC carrying Klebsiella pneumoniae in Sicily, Italy, 2014: emergence of multiple Non-ST258 clones. PLoS ONE 10:e0132936. doi: 10.1371/journal.pone.0132936
Bowers, J. R., Kitchel, B., Driebe, E. M., MacCannell, D. R., Roe, C., Lemmer, D., et al. (2015). Genomic analysis of the emergence and rapid global dissemination of the clonal group 258 Klebsiella pneumoniae pandemic. PLoS ONE 10:e0133727. doi: 10.1371/journal.pone.0133727
Brisse, S., Passet, V., Haugaard, A. B., Babosan, A., Kassis-Chikhani, N., Struve, C., et al. (2013). wzi Gene sequencing, a rapid method for determination of capsular type for Klebsiella strains. J. Clin. Microbiol. 51, 4073–4078. doi: 10.1128/JCM.01924-13
Calisto, F., Caneiras, C., Serqueira, S., Lito, L., Melo-Cristino, J., and Duarte, A. (2012). Klebsiella pneumoniae producing carbapenemase KPC-3 identifed in hospital wards. Rev. Port. Doenças Infecc. 8, 127–134.
Carattoli, A., Bertini, A., Villa, L., Falbo, V., Hopkins, K. L., and Threlfall, E. J. (2005). Identification of plasmids by PCR-based replicon typing. J. Microbiol. Methods 63, 219–228. doi: 10.1016/j.mimet.2005.03.018
Chavda, K. D., Chen, L., Jacobs, M. R., Rojtman, A. D., Bonomo, R. A., and Kreiswirth, B. N. (2015). Complete sequence of a bla(KPC)-harboring cointegrate plasmid isolated from Escherichia coli. Antimicrob. Agents Chemother. 59, 2956–2959. doi: 10.1128/AAC.00041-15
Chen, L., Chavda, K. D., Melano, R. G., Hong, T., Rojtman, A. D., Jacobs, M. R., et al. (2014a). Molecular survey of the dissemination of two blaKPC-harboring IncFIA plasmids in New Jersey and New York hospitals. Antimicrob. Agents Chemother. 58, 2289–2294. doi: 10.1128/AAC.02749-13
Chen, L., Mathema, B., Chavda, K. D., DeLeo, F. R., Bonomo, R. A., and Kreiswirth, B. N. (2014b). Carbapenemase-producing Klebsiella pneumoniae: molecular and genetic decoding. Trends Microbiol. 22, 686–696. doi: 10.1016/j.tim.2014.09.003
Chmelnitsky, I., Shklyar, M., Leavitt, A., Sadovsky, E., Navon-Venezia, S., Ben Dalak, M., et al. (2014). Mix and match of KPC-2 encoding plasmids in Enterobacteriaceae-comparative genomics. Diagn. Microbiol. Infect. Dis. 79, 255–260. doi: 10.1016/j.diagmicrobio.2014.03.008
Curiao, T., Morosini, M. I., Ruiz-Garbajosa, P., Robustillo, A., Baquero, F., Coque, T. M., et al. (2010). Emergence of blaKPC−3-Tn4401a associated with a pKPN3/4-like plasmid within ST384 and ST388 Klebsiella pneumoniae clones in Spain. J. Antimicrob. Chemother. 65, 1608–1614. doi: 10.1093/jac/dkq174
Cuzon, G., Naas, T., and Nordmann, P. (2011). Functional characterization of Tn4401, a Tn3-based transposon involved in blaKPC gene mobilization. Antimicrob. Agents Chemother. 55, 5370–5373. doi: 10.1128/AAC.05202-11
Deleo, F. R., Chen, L., Porcella, S. F., Martens, C. A., Kobayashi, S. D., Porter, A. R., et al. (2014). Molecular dissection of the evolution of carbapenem-resistant multilocus sequence type 258 Klebsiella pneumoniae. Proc. Natl. Acad. Sci. U.S.A. 111, 4988–4993. doi: 10.1073/pnas.1321364111
Feldman, N., Adler, A., Molshatzki, N., Navon-Venezia, S., Khabra, E., Cohen, D., et al. (2013). Gastrointestinal colonization by KPC-producing Klebsiella pneumoniae following hospital discharge: duration of carriage and risk factors for persistent carriage. Clin. Microbiol. Infect. 19, E190–E196. doi: 10.1111/1469-0691.12099
Garbari, L., Busetti, M., Dolzani, L., Petix, V., Knezevich, A., Bressan, R., et al. (2015). pKBuS13, a KPC-2-encoding plasmid from Klebsiella pneumoniae sequence type 833, carrying Tn4401b inserted into an Xer site-specific recombination locus. Antimicrob. Agents Chemother. 59, 5226–5231. doi: 10.1128/AAC.04543-14
García-Fernández, A., Fortini, D., Veldman, K., Mevius, D., and Carattoli, A. (2009). Characterization of plasmids harbouring qnrS1, qnrB2 and qnrB19 genes in Salmonella. J. Antimicrob. Chemother. 63, 274–281. doi: 10.1093/jac/dkn470
Holt, K. E., Wertheim, H., Zadoks, R. N., Baker, S., Whitehouse, C. A., Dance, D., et al. (2015). Genomic analysis of diversity, population structure, virulence, and antimicrobial resistance in Klebsiella pneumoniae, an urgent threat to public health. Proc. Natl. Acad. Sci. U.S.A. 112, E3574–E3581. doi: 10.1073/pnas.1501049112
Machado, P., Silva, A., Lito, L., Melo-Cristino, J., and Duarte, A. (2010). Emergence of Klebsiella pneumoniae ST11-producing KPC-3 carbapenemase at a Lisbon hospital. [Abstract]. Clin. Microbiol. Infect. 16(Suppl. 2), S28 (O129).
Magiorakos, A. P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi: 10.1111/j.1469-0691.2011.03570.x
Manageiro, V., Ferreira, E., Almeida, J., Barbosa, S., Simões, C., Bonomo, R. A., et al. (2015). Predominance of KPC-3 in a survey for carbapenemase-producing Enterobacteriaceae in Portugal. Antimicrob. Agents Chemother. 59, 3588–3592. doi: 10.1128/AAC.05065-14
Munoz-Price, L. S., Poirel, L., Bonomo, R. A., Schwaber, M. J., Daikos, G. L., Cormican, M., et al. (2013). Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet. Infect. Dis. 13, 785–796. doi: 10.1016/S1473-3099(13)70190-7
Naas, T., Cuzon, G., Truong, H.-V., and Nordmann, P. (2012). Role of ISKpn7 and deletions in blaKPC gene expression. Antimicrob. Agents Chemother. 56, 4753–4759. doi: 10.1128/AAC.00334-12
Nordmann, P., Cuzon, G., and Naas, T. (2009). The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet. Infect. Dis. 9, 228–236. doi: 10.1016/S1473-3099(09)70054-4
Nordmann, P., and Poirel, L. (2014). The difficult-to-control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clin. Microbiol. Infect. 20, 821–830. doi: 10.1111/1469-0691.12719
Ocampo, A. M., Chen, L., Cienfuegos, A. V., Roncancio, G., Chavda, K. D., Kreiswirth, B. N., et al. (2015). A two-year surveillance in five Colombian Tertiary Care Hospitals reveals high frequency of Non-CG258 clones of carbapenem-resistant Klebsiella pneumoniae with distinct clinical characteristics. Antimicrob. Agents Chemother. 60, 332–342. doi: 10.1128/AAC.01775-15
Pires, J., Novais, Â., and Peixe, L. (2013). Blue-carba, an easy biochemical test for detection of diverse carbapenemase producers directly from bacterial cultures. J. Clin. Microbiol. 51, 4281–4283. doi: 10.1128/JCM.01634-13
Poirel, L., Barbosa-Vasconcelos, A., Simões, R. R., Da Costa, P. M., Liu, W., and Nordmann, P. (2012). Environmental KPC-producing Escherichia coli isolates in Portugal. Antimicrob. Agents Chemother. 56, 1662–1663. doi: 10.1128/AAC.05850-11
Rodrigues, C., Machado, E., Ramos, H., Peixe, L., and Novais, Â. (2014). Expansion of ESBL-producing Klebsiella pneumoniae in hospitalized patients: a successful story of international clones (ST15, ST147, ST336) and epidemic plasmids (IncR, IncFIIK). Int. J. Med. Microbiol. 304, 1100–1108. doi: 10.1016/j.ijmm.2014.08.003
Rodrigues, C., Sousa, C., Machado, E., Novais, Â., and Peixe, L. (2015). “Multidrug resistant (MDR) Klebsiella pneumoniae ST15 isolates: is their genomic plasticity the reason for its success?,” in Abstract retrieved from Abstracts in 25th European Congress of Clinical Microbiology and Infectious Diseases (ECCMID) (Copenhagen), (P0991).
Villa, L., García-Fernández, A., Fortini, D., and Carattoli, A. (2010). Replicon sequence typing of IncF plasmids carrying virulence and resistance determinants. J. Antimicrob. Chemother. 65, 2518–2529. doi: 10.1093/jac/dkq347
WHO (2014). Antimicrobial Resistance: Global Report on Surveillance. Geneva: WHO Press, World Health Organization.
Yigit, H., Queenan, A. M., Anderson, G. J., Domenech-Sanchez, A., Biddle, J. W., Steward, C. D., et al. (2001). Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 45, 1151–61. doi: 10.1128/AAC.45.4.1151-1161.2001
Zhou, K., Lokate, M., Deurenberg, R. H., Tepper, M., Arends, J. P., Raangs, E. G. C., et al. (2015). Use of whole-genome sequencing to trace, control and characterize the regional expansion of extended-spectrum β-lactamase producing ST15 Klebsiella pneumoniae. Sci Rep. 11:6:20840. doi: 10.1038/srep20840
Keywords: multidrug resistance, carbapenemases, international clones, ST15, ST147, cointegrated plasmids, ColE
Citation: Rodrigues C, Bavlovič J, Machado E, Amorim J, Peixe L and Novais  (2016) KPC-3-Producing Klebsiella pneumoniae in Portugal Linked to Previously Circulating Non-CG258 Lineages and Uncommon Genetic Platforms (Tn4401d-IncFIA and Tn4401d-IncN). Front. Microbiol. 7:1000. doi: 10.3389/fmicb.2016.01000
Received: 06 April 2016; Accepted: 13 June 2016;
Published: 28 June 2016.
Edited by:
John W. A. Rossen, University of Groningen, University Medical Center, NetherlandsReviewed by:
Kai Zhou, University Medical Center Groningen, NetherlandsÁkos Tóth, National Center for Epidemiology, Hungary
Copyright © 2016 Rodrigues, Bavlovič, Machado, Amorim, Peixe and Novais. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luísa Peixe, bHBlaXhlQGZmLnVwLnB0;
Ângela Novais, YW5nZWxhc2lsdmFub3ZhaXNAZ21haWwuY29t