 Patryk Krauze
Patryk Krauze Horst Kämpf2
Horst Kämpf2 Fabian Horn
Fabian Horn Dirk Wagner
Dirk Wagner Mashal Alawi
Mashal Alawi- 1GFZ German Research Centre for Geosciences, Section 5.3 Geomicrobiology, Potsdam, Germany
- 2GFZ German Research Centre for Geosciences, Section 3.2 Organic Geochemistry, Potsdam, Germany
- 3Hydroisotop GmbH, Schweitenkirchen, Germany
- 4Institute for Earth and Environmental Sciences, University of Potsdam, Potsdam, Germany
The Cheb Basin (NW Bohemia, Czech Republic) is a shallow, neogene intracontinental basin. It is a non-volcanic region which features frequent earthquake swarms and large-scale diffuse degassing of mantle-derived CO2 at the surface that occurs in the form of CO2-rich mineral springs and wet and dry mofettes. So far, the influence of CO2 degassing onto the microbial communities has been studied for soil environments, but not for aquatic systems. We hypothesized, that deep-trenching CO2 conduits interconnect the subsurface with the surface. This admixture of deep thermal fluids should be reflected in geochemical parameters and in the microbial community compositions. In the present study four mineral water springs and two wet mofettes were investigated through an interdisciplinary survey. The waters were acidic and differed in terms of organic carbon and anion/cation concentrations. Element geochemical and isotope analyses of fluid components were used to verify the origin of the fluids. Prokaryotic communities were characterized through quantitative PCR and Illumina 16S rRNA gene sequencing. Putative chemolithotrophic, anaerobic and microaerophilic organisms connected to sulfur (e.g., Sulfuricurvum, Sulfurimonas) and iron (e.g., Gallionella, Sideroxydans) cycling shaped the core community. Additionally, CO2-influenced waters form an ecosystem containing many taxa that are usually found in marine or terrestrial subsurface ecosystems. Multivariate statistics highlighted the influence of environmental parameters such as pH, Fe2+ concentration and conductivity on species distribution. The hydrochemical and microbiological survey introduces a new perspective on mofettes. Our results support that mofettes are either analogs or rather windows into the deep biosphere and furthermore enable access to deeply buried paleo-sediments.
Introduction
Mofettes are cold (≤30°C) natural exhalations of magmatogene carbon dioxide, which migrates through the lithospheric upper mantle and crust through surface water and soil to the atmosphere (Pfanz, 2008; Kämpf et al., 2013). Such CO2 vents are located both in volcanically active and in non-volcanic, but seismically active regions (Evans et al., 2002; Lan et al., 2007). The cold exhalations of mantle-derived carbon dioxide occur as “wet mofettes” (CO2 runs/bubbles through surface water) and “dry mofettes” (CO2 migrates through subsurface sediments and soil; Kämpf et al., 2013). Both types show diffuse and channelized CO2 degassing. Mofettes are local degassing phenomena, but also integral parts of larger Diffuse Degassing Structures (DDS) on the scale of up to a few square kilometers, controlled by the fluid migration inside of fault zones (e.g., Girault and Perrier, 2014; Nickschick et al., 2015) or in volcano-hydrothermal areas (e.g., Chiodini et al., 2008; Girault et al., 2014). Recently, mofettes were used as model ecosystems for studying the response of soil microbiota to potential CO2 leakage from underground carbon capture and storage systems (e.g., Krüger et al., 2009, 2011; Frerichs et al., 2013).
At the surface, the dry mofettes show a characteristic vegetation which is at some sites strongly declined or even completely missing. Dry mofettes draw attention as an extreme habitat in ecological research investigating the response of plants (Vodnik et al., 2002), arbuscular mycorrhizal fungi (Maček et al., 2011) or soil archaea and bacteria (Šibanc et al., 2014) to elevated CO2 concentrations. Generally, a shift to anaerobic/microaerophilic and acidophilic community compositions has been reported for mofette soils compared to respective control sites. Bacterial community richness, evenness and diversity decreased with increasing CO2 flux (Sáenz de Miera et al., 2014). The abundance of methanogenic archaea and sulfate reducing bacteria, which could use geogenic CO2 for assimilatory biosynthesis, increased toward a CO2 vent core (Oppermann et al., 2010; Beulig et al., 2015). Furthermore, increasing abundances of putative anaerobes related to the Chloroflexi and Firmicutes phyla have been reported (Frerichs et al., 2013; Sáenz de Miera et al., 2014). Besides methanogens, ammonia oxidizing archaea related to the Thaumarchaeota were abundant at a CO2 vent core (Frerichs et al., 2013). Šibanc et al. (2014) reported oxygen concentration, soil pH and total nitrogen to be the strongest parameters shaping the archaeal community structure, while the bacterial composition is mainly shaped by the oxygen concentration of soil pore air. In addition to these studies on dry mofette soils, metagenomic analyses on deeply sourced CO2-saturated fluids from Crystal Geyser (Utah, USA) illuminated the potential influence of high CO2 concentrations on aquatic prokaryotic communities. This environment was dominated by populations capable of chemolithotrophy (“marine” and “freshwater” iron-oxidizing bacteria, sulfur-oxidizers and Thiobacillus-like Hydrogenophilales) and showed a broad diversity of uncultured species (Emerson et al., 2016). In the same system, Probst et al. (2017) observed autotrophic capabilities in organisms which account for over 70% of the community. Regarding microbial carbon fixation, high CO2 concentrations in the subsurface select for the Wood-Ljungdahl pathway and the Calvin-Benson-Bassham cycle, and for form II RuBisCOs, which are most likely adaptations to anaerobic and high CO2 conditions.
For the first time, this study presents a comprehensive insight into the microbial community structure of mineral and mofette waters influenced by active, strongly degassing CO2 conduits. Communities of aquatic bacteria and archaea were characterized using high-throughput sequencing of the 16S rRNA gene and quantitative PCR. To get indications for the origin of the waters in terms of depth and to check whether indications for microbial activity can be found, a chemical characterization of the waters using ICP-OES (cations), ion chromatography (anions) and isotope investigations using mass spectrometry (water phase: 2HH2O, 18OH2O and 34SSO4, 18OSO4 and gas phase: methane, ethane and propane: 2H, 13C) were performed. Additionally, literature data of gas chemistry and isotope characteristics of the CO2-dominated gas (13CCO2, 3He/4He), based on multi-year investigations of the investigated sites, were used. With this interdisciplinary approach, we aimed (I) to retrieve a detailed geochemically characterization of the waters and the free, CO2 dominated gas phase, and to determine their origin, (II) to get a better understanding of the abundance and composition of aquatic microbial communities facing high CO2 partial pressures, (III) to identify key organisms and related metabolic pathways and (IV) to determine major community-shaping environmental factors.
Geological Background
The Cheb Basin (NW Bohemia, Czech Republic) is a shallow, neogene intracontinental basin that has formed since the Cenozoic at the intersection of the E-NE trending Eger Rift and the N-S trending Regensburg-Leipzig-Rostock seismoactive zone (Bankwitz et al., 2003; Fischer et al., 2014). The NW-trending pre-Neogene Mariánské Lázne Fault (MLF) forms the eastern boundary of the Cheb Basin and is marked by a 50–100 m high escarpment. The area is a non-volcanic region which features frequent earthquake swarms up to ML 4.5 (hypocenter depths range between 6.5 and 11 km with some clusters down to 13 km; Fischer et al., 2014; Hainzl et al., 2016) and large-scale diffuse degassing of mantle-derived carbon dioxide at the surface that occurs in the form of CO2-rich mineral springs and wet and dry mofettes (Weinlich et al., 1998, 1999; Kämpf et al., 2013; Nickschick et al., 2015). Most of the earthquake activity (about 90% of the total seismic moment) is concentrated at the Nový Kostel focal zone (NKFZ), located at the intersection between the N-S trending Počatky-Plesná fault zone (PPZ) and the MLF (Fischer and Horálek, 2003). South of the Nový Kostel focal zone (NKFZ), the PPZ is characterized by intense CO2-degassing (mofette fields).
The migrating gas consists of up to >99 vol% CO2 and can contain traces of hydrogen, helium, argon, methane, oxygen or nitrogen (Bräuer et al., 2011, 2014). This migration of gas results in substantially changed soil gases and localized soil hypoxia (Kämpf et al., 2013; Nickschick et al., 2015). Carbon isotope signatures up to −70‰ of methane, a minor component of the CO2 dominated upstreaming gases, indicate an interaction between geological, geophysical and microbial driven processes in the deep subsurface (Bräuer et al., 2005, 2007). Mofettes share distinct geochemical features due to the exposure to elevated CO2 partial pressures. Acidification is a common feature which is reflected by low to very low pH values (3.5–4, Beaubien et al., 2008; 3.5–4.7, Rennert et al., 2011) and is negatively correlated to increasing CO2 concentration in soil gas or pore water (Mehlhorn et al., 2014; Rennert and Pfanz, 2016). Lowered pH and decreased redox potentials have been shown to increase metal mobilization in a mofette which could influence soil nutrient availability (Mehlhorn et al., 2014, 2016). Another common feature is the accumulation of organic matter (OM) in mofette soils (e.g., Beaubien et al., 2008; Beulig et al., 2015), which is a result of the assimilation of substantial quantities of geogenic CO2 via primary production by plants and subsurface carbon fixation (Oppermann et al., 2010; Beulig et al., 2016). A restricted degradation of OM by microorganisms under an increased CO2 atmosphere was indicated by the C/N ratio, the occurrence of undegraded plant material and high organic carbon contents (Rennert et al., 2011). Recently, Beulig et al. (2016) suspected that the permanent exclusion of meso-/macroscopic eukaryotes and related physiological capacities is the reason for the restricted degradation rather than an impaired biochemical potential of microorganisms.
Methods
Site Description and Sampling
All studied mofettes and mineral water springs are located in the Cheb Basin (Figures 1, 2). The sampling sites included two wet mofettes (“Bublák C” and “Bublák NW,” Figure 1 and Table 1), which were situated in swampy woodlands of the flood plain of the Plesná river. In contrast, the investigated mineral waters had a deeper origin, which is reflected in the respective hydrochemical properties. These included three waters, which had a shallow subsurface origin and were obtained from the Mostek u Križovatky pramen (“U Mostku”, Figure 1 and Table 1), a mineral water spring close to the Plesná river (“Plesná”, Figure 1 and Table 1) and a freshwater spring (“Kopanina”, Figure 1 and Table 1) close to a brook. The fourth water has a deeper origin and was obtained from the Císarský pramen at the Soos area (“Soos”, Figure 1 and Table 1). Sampling (water and gas) took place in March, April, October and December 2014 to capture a possible seasonality. The samples collected in October 2014 (at 29.10.2014) were used for detailed microbial studies. For microbiological investigations at each site, a 5 L amber stained laboratory glass bottle was filled with water either from a pond or directly from a wellhead. To show the distinctness of the aquatic microbial communities of CO2-influenced waters, related soils need to be investigated. Thereby, the top five centimeters directly next to a wet mofette (Bublák C), a mineral water spring (Plesná) and sediment from the Císarský pramen well (Soos) were sampled. Additionally, several smaller volumes of water were sampled for the analysis of element hydrochemistry and isotope (18O, 2H, 34S) hydrochemistry. Depending on filter clogging, varying volumes (2.5–5.5 l) of water were filtered in replicate using 0.2 μm cellulose-acetate filters (Sartorius AG, Germany) and stored at −20°C till DNA extraction. The gas was sampled using glass vessels with two stopcocks for isotope analysis of methane (13C, 2H), ethane and propane (13C). The vessels were filled with spring water, which was replaced by the free gas bubbling out of the water in the glass vessel.
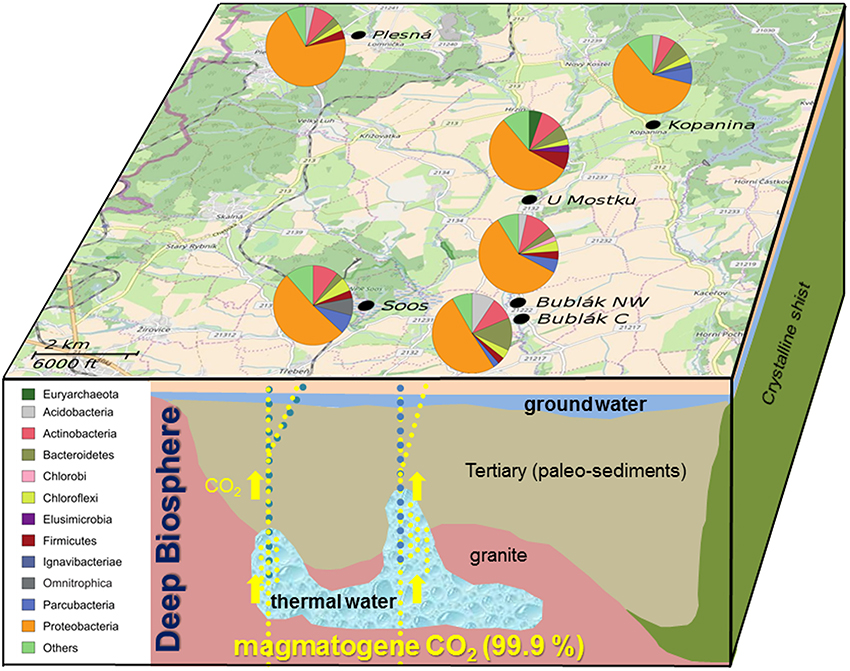
Figure 1. Pathway of magmatogene CO2, mixing with deep thermal water, paleo-sediment and ground water. Location of sampling sites and relative abundances of phyla determined by Illumina MiSeq sequencing of the 16S rRNA gene in different CO2 affected waters from the Cheb Basin, NW Bohemia. Only phyla with an abundance of at least 2% at a given site are shown. Map provided by © OpenStreetMap-Mitwirkende.

Figure 2. The six investigated sites. (a) Bublák C; (b) Bublák NW; (c) Kopanina; (d) Plesná; (e) Soos; (f) U Mostku.
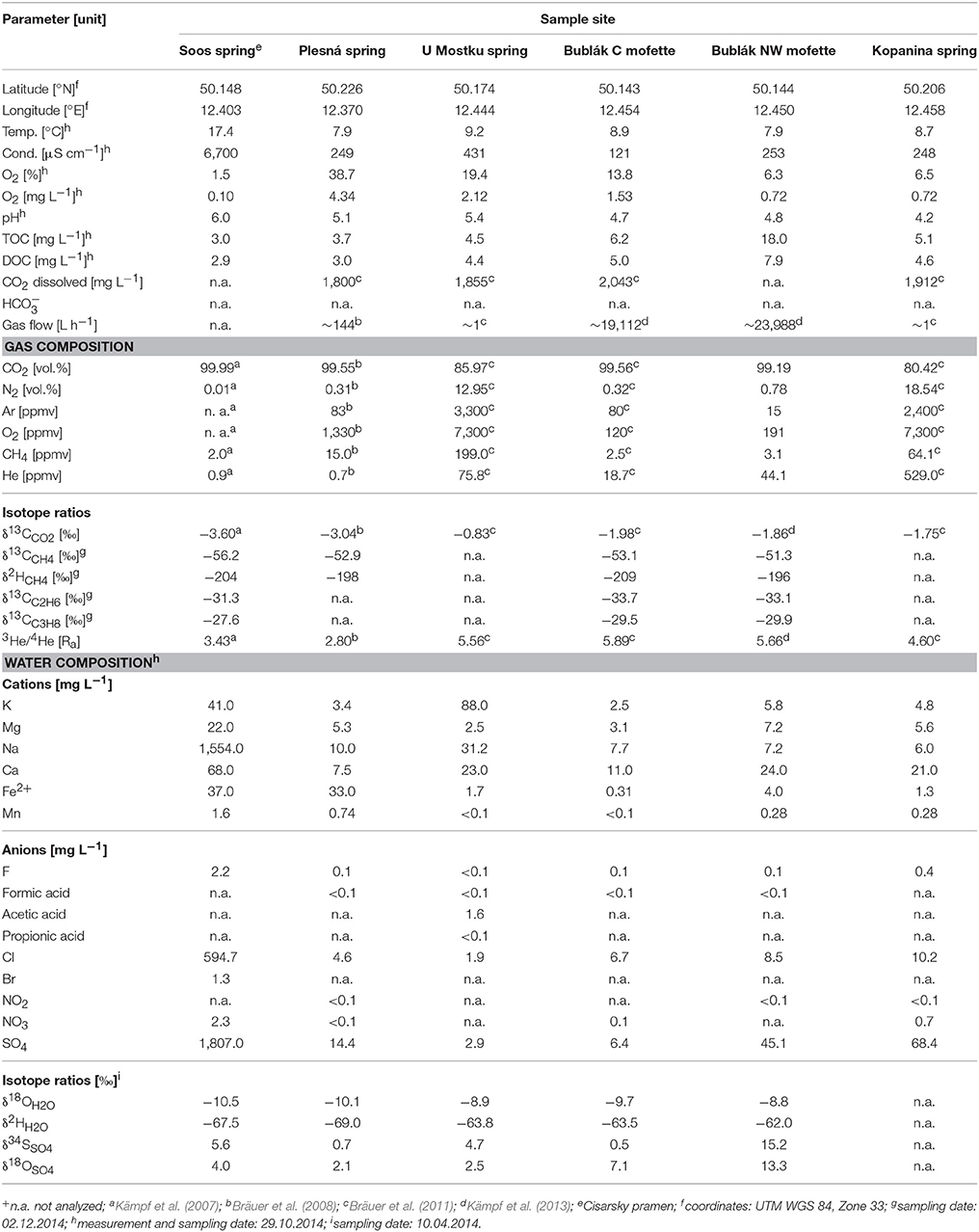
Table 1. Water/gas chemistry and related isotopic analysis data of mofette and mineral waters from the Cheb Basin, NW Bohemia.
Geochemical and Isotope Analysis of Waters
Electrical conductivity (EC), pH, O2 and water temperature were directly measured in the field (Multi 3420 digital analyzer, WTW GmbH Weilheim). To investigate the main cations (K+, Mg2+, Na+, Ca2+, Fe2+, and Mn2+), water samples were filtered (<0.2 μm) and acidified to pH <2 with HNO3 for storage in 50 ml PP bottles at 4°C. Analysis took place at the GFZ German Research Centre for Geosciences in Potsdam, Germany by ICP-OES (VarianVista-MPX). Standards (plasma standard solution, Johnson Matthey Company), containing all of the investigated compounds, were measured in different concentrations every day.
The standard deviation of sample and standard quantification is <2% for Mg2+, Ca2+, Fe2+, and Mn2+, <3% for Na+ and <4% for K+ and was determined by at least three measurements. For the investigation of anions, the water samples were filled in 1 L glass bottles in the field. The samples were analyzed at the GFZ German Research Centre for Geosciences by ion chromatography (IC) with conductivity detection (ICS 3000, Dionex) to determine the content of F−, , NO2−, NO3−, Br−, Cl−, and and organic acids (formate, acetate, propionate). Standards containing all of the investigated compounds were measured in different concentrations every day. The standard deviation of sample and standard quantification is below 10% (determined by at least two measurements).
For sulfate isotope analyses, dissolved sulfate stored in 0.5 up to 1 L PP bottles (depending on the quantity of dissolved sulfate) was precipitated using BaCl2·2H2O. The precipitated BaSO4 was collected by filtration through nitrocellulose membranes, washed to remove residual BaCl2 and dried at 50°C. Sulfur isotopic compositions were measured after conversion of BaSO4 to SO2 using an elemental analyzer (continuous flow flash combustion technique) coupled with an isotope ratio mass spectrometer (Delta S, ThermoFinnigan, Bremen, Germany) at the stable isotope laboratory of the Helmholtz Centre for Environmental Research–UFZ, Germany.
Analytical errors of the measurement of more than ±0.3‰ and results are reported in delta notation (δ34S) as part per thousand (‰) deviation relative to the Vienna Cañon Diablo Troilite (VCDT) standard. Oxygen isotope analysis of sulfate was performed using a delta plus XL mass spectrometer (ThermoFinnigan, Bremen, Germany) with an analytical precision of more than ±0.5‰. Results of oxygen isotope measurements are expressed in delta notation (δ18OSO4) as part per thousand (‰) deviation relative to Vienna Standard Mean Ocean Water (VSMOW). For normalization of the δ34S and δ18OSO4 data, the IAEA-distributed reference material NBS 127 (BaSO4) was used. The assigned values were +20.3‰ (VCDT) δ34S and +8.6‰ (VSMOW) for δ18OSO4.
Measurements of stable isotopes of δ18O and δ2H in the water samples were performed at the stable isotope laboratory of the UFZ in Halle/Saale, Germany using a laser-based analyzer (L1102-I, Picarro Inc.). This instrument has an analytical precision of 0.5‰ for δ2H and 0.1‰ for δ18O. For normalization of the δ18O water data, the IAEA-distributed reference materials VSMOW and SLAP were used.
Isotope Analysis of Gases
The carbon isotope ratios of CO2, CH4, and higher gaseous hydrocarbons and hydrogen isotope ratios of CH4 were measured at the stable isotope laboratory of Hydroisotop GmbH Schweitenkirchen using a GC-IRMS equipped with a purge and trap device. The line consists of PTA−3000 Purge and Trap autosampler (IMT Germany), a Trace GC Ultra gas chromatograph (Thermo Scientific) with Hayesep Q separation column (VICI) and helium as carrier gas, and Delta V isotope ratio mass spectrometer–IRMS (Thermo Scientific). The Isodat 3 software was used to evaluate the signals. Results are reported in δ values relative to International standards: V-PDB for carbon and V-SMOW for hydrogen. The instrumental error of δ13C in CO2, CH4 and higher hydrocarbons is ±0.5‰ and that of δ2H is ±5‰.
Nucleic Acids Extraction
Genomic DNA of the sampled waters was extracted from filters using the PowerWater® DNA Isolation Kit (MO BIO Laboratories Inc., USA) according to the manufacturer's specifications with minor changes to the protocol. Once 100 μl of elution buffer was added, the samples were incubated at 55°C for 5 min before the final centrifugation step. The extracted DNA was stored at −20°C.
Genomic DNA from 0.5 mg of the sampled sediments was extracted using the PowerSoil® DNA Isolation Kit (MO BIO Laboratories Inc., USA).
These DNA preparations were used as a template for the quantification by quantitative PCR (qPCR) and next-generation sequencing.
Quantification of Bacterial 16S rRNA Genes and Functional Genes
Quantitative polymerase chain reaction was used to quantify total bacterial abundances and the functional genes of sulfate reducing bacteria and methanogenic archaea. All qPCR essays were performed in triplicates on a CFX96 Real-time thermal cycler (Bio-Rad Laboratories Inc., USA) and contained 12.5 μl iTaq™ Universal SYBR® Green Supermix (ThermoFisher Scientific Inc., USA), 8.5–10.5 μl PCR water, each 0.5 μl of forward and reverse primer (20 μM) and 1–3 μl template. The setup was optimized for each target regarding the cycler program, used volumes and dilution factors of the samples. The quantification of the bacterial 16S rRNA gene was based on the primers 331F (5′-TCCTACGGGAGGCAG-CAGT-3′) and 797R (5′-GGACTACCAGGGTATCTAATCCTGTT-3′) (Nadkarni et al., 2002). After an initial denaturing phase of 5 min at 98°C, the cycler included 40 cycles of 5 s at 98°C, 20 s at 57°C and 60 s at 72°C plus the plate read. The quantification of sulfate reducers was based on the primers dsr2060F (5′-CAACATCGTYCAYACCCAGGG-3′) (Geets et al., 2006) and dsr4R (5′-GTGTAGCAGTTACCGCA-3′) (Wagner et al., 1998) targeting the dissimilatory sulfite reductase β-subunit (dsrB) gene and included an initial denaturing for 10 min at 95°C, followed by 40 cycles of 30 s at 95°C, 60 s at 60°C, 60 s at 72°C and 3 s at 80°C plus the plate read. The quantification of methanogenic archaea was based on the primers mlas-F (5′-GGTGGTGTMGGDTTCACMCARTA-3′) and mcrA-R (5′-CGTTCATBGCGTAGTTVGGRTAGT-3′) (Steinberg and Regan, 2009) targeting the methyl coenzyme M reductase (mcrA) gene and included an initial denaturing for 3 min at 95°C, followed by 40 cycles of 5 s at 95°C, 20 s at 58.5°C, 30 s at 72°C and 3 s at 80°C plus the plate read. All cycling programs included a melting curve from 60 to 95°C with 0.5°C steps per plate read. The analysis of quantification data was performed with the CFX Manager™ Software (Bio-Rad Laboratories Inc., USA).
Illumina MiSeq Sequencing
Unique combinations of tagged 515F (5′-GTGCCAGCMGCCGCGGTAA-3′) and 806R (5′-GGACTACHVGGGTWTCTAAT-3′) (Caporaso et al., 2011) primers were assigned to each sample. The samples were processed in duplicates and pooled afterwards to reduce PCR variability. Additionally, technical replicates for the sequencing of each sample were produced to reduce sequencing variability. The PCR was performed on a T100™ Thermal Cycler (Bio-Rad Laboratories Inc., USA) in 25 μl reactions, containing 12.5 μl iTaq™ Universal SYBR® Green Supermix (ThermoFisher Scientific Inc., USA), 8.75 μl PCR water, each 0.625 μl of forward and reverse primer (20 μM) and 2.5 μl genomic DNA using following cycler program: Initial denaturing step for 3 min at 95°C followed by 10 cycles of 1 min at 94°C, 1 min at 53°C (−0.2°C/cycle) and 1 min at 72°C, followed by 20 cycles of 1 min at 94°C, 1 min at 50°C and 1 min at 72°C, followed by a final extension step for 10 min at 72°C. All samples were pooled by adding an equal amount of DNA (60 ng DNA per sample). Subsequently, a purification of the PCR product pool was achieved by using the Hi Yield Gel/PCR DNA Fragment Extraction. The Illumina MiSeq sequencing was performed by EuroFins Scientific SE, Luxembourg.
Bioinformatics and Statistical Analysis
Sequencing was performed on an Illumina MiSeq (2 × 250 bp). Reads were demultiplexed using CutAdapt (options: e0.1; trim-n; Martin, 2011). Read pairs were merged using PEAR (options: Q25; p10−4; o20; Zhang et al., 2014). QIIME (version 1.9.1) (Caporaso et al., 2010) was employed for microbiome analysis. USEARCH (Edgar, 2010) was used for the detection and removal of chimeric sequences. The SILVA database (version 128) (DeSantis et al., 2006) was utilized for open-reference OTU clustering (97% sequence similarity) and taxonomic assignments. Singletons and OTUs assigned to chloroplasts were removed. For the processing and visualization of the obtained OTU table, R, CANOCO 5 (Šmilauer and Lepš, 2014) and PAST3 (Hammer et al., 2001) software was used. For alpha diversity analyses, the data were rarefied to 61,042 reads per sample. Alpha diversity was estimated using the Shannon's H index, Faith's Phylogenetic diversity and Pielou's evenness. Beta diversity was calculated using weighted UniFrac distance. Similarity Percentages (SIMPER) analyses were used to determine the main drivers of dissimilarity of community composition. Sequencing data were submitted to the European Nucleotide Archive (http://www.ebi.ac.uk/ena) under accession number PRJEB20063.
Results
Characterization of Mofette and Mineral Waters
Distinct differences in hydrochemical properties were measured in water samples from different locations (Table 1). The waters were acidic with pH values ranging from 4.2 to 6.0 and showed low contents of dissolved oxygen of 0.10–4.34 mg L−1. The deep thermal water (Soos) differed from all other waters featuring a relative high temperature of 17.4°C and conductivity of 6,700 μS cm−1, indicating its deeper subsurface origin. Additionally the concentration of several ions, for instance magnesium, calcium, sodium, manganese, chloride and sulfate, was very high compared to the other waters. The mineral water springs (Soos, Plesná) showed especially high concentrations of Fe2+ at 37 and 33 mg L−1 respectively. The total organic carbon (TOC) and dissolved organic carbon (DOC) concentrations varied greatly: Lowest TOC values could be observed in the mineral water springs (Soos: 3.0 mg L−1, Plesná: 3.7 mg L−1) and highest in the mofette waters (Bublák C: 6.2 mg L−1, Bublák NW: 18.0 mg L−1). A similar trend was detected regarding the DOC concentrations. Mineral water springs showed lowest (Soos: 2.9 mg L−1, Plesná: 3.0 mg L−1) and mofette waters highest (Bublák C: 5.0 mg L−1, Bublák NW: 7.9 mg L−1) amounts of DOC. Furthermore, only the shallow subsurface water (U Mostku) showed detectable amounts of acetic acid (1.6 mg L−1). The concentrations of other organic acids (e.g., formic acid, propionic acid) were under the detection limit. Nitrite was not detectable. The nitrate concentration was overall low, and the deep mineral water (Soos) showed the highest concentration with 2.3 mg L−1.
The δ2H vs. δ18O values of sampled waters showed similar values (δ2H from −69 to −62.0‰ SMOW and δ18O from −8.8 to −10.5‰ SMOW). The δ18O vs. δ34S values of the dissolved sulfate of the waters showed large differences in the values (δ18O from 2.1 to 13.3‰ and δ34S from 0.5 to 15.2‰). In most cases, the sulfate content amounts to <100 mg L−1, except for Soos (1,807.0 mg L−1).
Chemical and Isotopic Composition of Free Gas Samples
The chemical composition (in % by vol.) and the isotopic composition (3He/4He ratio as R/Ra, Ra: atmospheric 3He/4He ratio and δ13CCO2 in ‰-V-PDB) of gas samples collected from the gas of bubbling waters are reported in Table 1. The gas compositions of the mineral springs Soos, Plesná and both Bublák mofettes were dominated by CO2 (>99%), while the springs U Mostku and Kopanina showed lower CO2 concentrations (<90%).
The δ13CCO2 values ranged between −3.6‰ and −0.83‰ relative to the V-PDB standard (Kämpf et al., 2007, 2013; Bräuer et al., 2008, 2011). The highest 3He/4He ratios were obtained along the PPZ (Bublák C: 5.89 Ra, Bublák NW: 5.66 Ra and U Mostku: 5.56 Ra), whereas at Kopanina, located at the MLF the 3He/4He ratio is lower (4.6 Ra). The lower portions of mantle-derived helium were shown outside of these fault zones at the Soos mineral spring (3.43 Ra) and the lowest on the periphery of the degassing center of the Cheb Basin at the Plesná mineral spring (2.8 Ra). The δ13CCH4 values ranged between −56.2 and −51.3‰ relative to the V-PDB standard and the δ2HCH4 values ranged between −209 and −196‰ relative to the V-SMOW standard.
Abundances of Microorganisms
Clear differences in 16S rRNA gene abundances were detected between waters from surface and deep locations (Table 2). Abundances ranged from 8.8 × 105 copies L−1 (Soos) to 1.7 × 1010 copies L−1 (Bublák NW), while in most cases values between 108 and 109 copies L−1 could be observed. The dsrB gene was successfully quantified in each water except for the deep mineral water (Soos). The abundance of this gene ranged between 1.9 × 105 copies L−1 (Plesná) and 2.5 × 107 copies L−1 (Bublák NW), following a similar trend as the 16S rRNA gene abundances. The quantification of the mcrA gene only succeeded for both Bublák mofettes (Bublák C; Bublák NW) and the shallow groundwater (U Mostku). The abundances were comparatively low with 6.0 × 105 genes L−1 (Bublák NW), 1.6 × 105 genes L−1 (Bublák C) and 8.8 × 104 (U Mostku). McrA gene abundances of the remaining waters were under the limit of detection (~104 genes L−1).
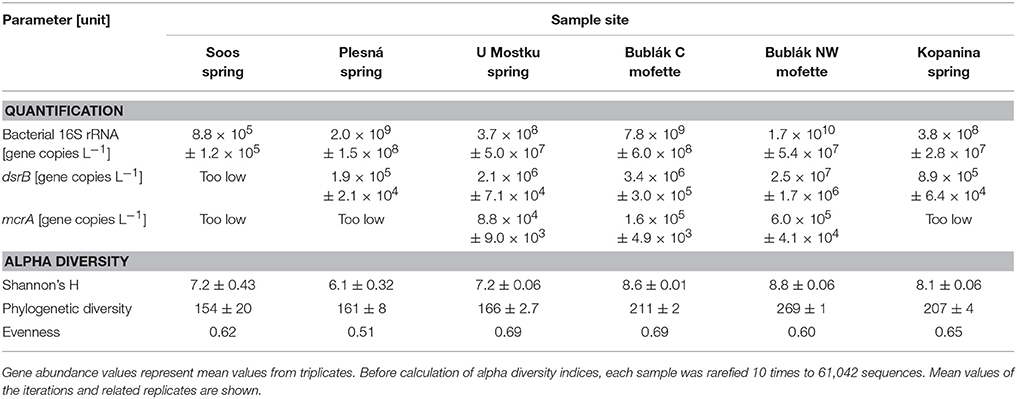
Table 2. Abundances of bacterial 16S rRNA, dsrB and mcrA genes revealed by quantitative PCR and calculated alpha diversity values based on the OTUs in the investigated CO2 affected mofette and mineral waters from the Cheb Basin, NW Bohemia.
Diversity and Microbial Community Composition
In total, 1.63 million reads were obtained after merging the forward and reverse reads and demultiplexing. After quality filtering and deletion of chimeric sequences, 1.46 million high quality reads remained in the sample set. The amount of reads per sample ranged from 68,784 to 477,069 with a mean value of 129,181 (Table S1). Hereby, rarefaction analyses showed that no sample exhibited a conspicuous increase of its Shannon's H index after including more than 10,182 sequences (Figure S1, Table S2). All samples have been sufficiently covered by sequencing, since an increasing number of reads per sample does not bias diversity. As indicated by boxplots analyses (Figure S2), the replicates of a sample showed only a small distance to each other, whereby larger distances were observed between different sites. A total of 25,909 OTUs were calculated. Before analysis, we removed chloroplast-related OTUs (3.11% of total read counts) and OTUs that could not be assigned to any domain (5.55% of total read counts) from the data set. After taxonomic classification, 1,399 putative genera were obtained.
The alpha diversity of the samples was calculated using Shannon's H index (Table 2). The alpha diversity of the water obtained from a surface mineral spring (Plesná) showed the lowest microbial diversity, followed by both subsurface waters (Soos; U Mostku). Generally the diversity of the surface waters (Bublák C, Bublák NW, and Kopanina) was higher. The differences between surface and subsurface waters are similarly observable in the Faith's PD index (Table 2). None of the considered environmental parameters showed a significant correlation with the calculated alpha diversity indices.
To access beta diversity and therefore the differences in microbial community composition in the mofette and mineral waters, weighted UniFrac distance was calculated. The clustering of the samples in the corresponding PCoA (Figure 3) showed that the microbial communities found in subsurface waters (U Mostku, Soos) differed from the surface waters (Plesná, Bublák C, Bublák NW, and Kopanina). SIMPER test revealed Sulfuricurvum, Gallionella, Comamonadaceae, Omnitrophica, Denitratisoma, and Sulfurimonas to be the main drivers of dissimilarity (>1.5% contribution) between surface and subsurface waters (Table S3). Microbial communities of related sediments differed from the respective waters. Abundances of different phylotypes in CO2 affected waters were analyzed on phylum and, depending on the possibility of an assignment, on the lowest identifiable taxonomic level. Generally, the phylum Proteobacteria was very abundant across all mofette and mineral waters (Figure 1). However, the abundances of Proteobacteria-related families and genera and other phyla differed substantially (Figures 1, 4, Table S4). In the following part, the most abundant groups in the investigated waters are presented.
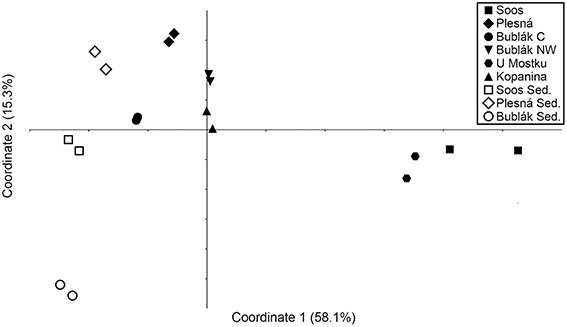
Figure 3. Principal coordinates analysis (PCoA) plot derived from weighted UniFrac distance between the investigated microbial communities. Axis 1 and Axis 2 explain 73.4% of the variance.
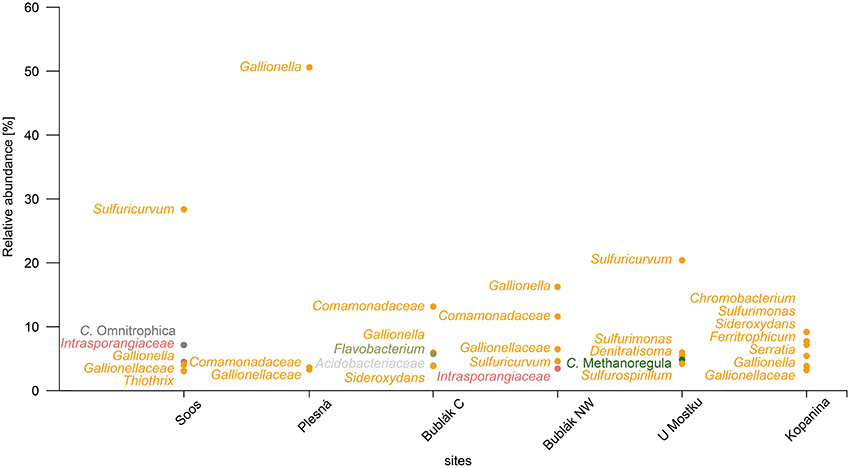
Figure 4. Dominant groups at the lowest assignable taxonomic level in the microbial communities of CO2 affected waters. Labels are shown only for groups with average abundances over 3%. Groups of taxa belonging to the same phylum are colored according to Figure 1.
At the phylum level, the microbial community of the deep mineral waters in Soos was composed of Proteobacteria (51.6%), Actinobacteria (10.29%), Parcubacteria (7.84%), Omnitrophica (7.15%), and Chloroflexi (5.61%). In comparison to the other investigated waters, a large fraction of unassignable sequences was found. Most abundant genera belonged to the phylum Proteobacteria, namely Sulfuricurvum (28.37%), Gallionella (4.03%), Thiothrix (3.05%), and Sulfurimonas (2.86%). A similar profile was observed for the shallow groundwater (U Mostku): Proteobacteria (55.95%), Actinobacteria (9.02%), Firmicutes (7.09%), and Bacteroidetes (5.96%) represented the most abundant phyla, while the most abundant genera consisted of Sulfuricurvum (20.39%), Sulfurimonas (5.96%), Denitratisoma (5.47%), and Sulfurospirillum (4.21%). Unique for this water was the occurrence of a larger fraction related to archaeal taxa, namely the phylum Euryarchaeota (5.29%) and the corresponding genus Methanoregula (4.85%). In contrast to these two waters, microbial communities of the remaining CO2 affected waters showed less OTUs related to the sulfur cycling (e.g., Sulfuricurvum or Sulfurimonas). The community of the mineral spring water (Plesná) was dominated by Proteobacteria (70.08%) and particularly by Gallionella-related OTUs (50.59%). OTUs related to Sideroxydans (1.35%) and Bacteriovorax (1.67%) were significantly less abundant. The communities of the mofette waters (Bublák C; Bublák NW) shared several features in terms of abundant phylotypes. Besides, a large portion of Proteobacteria-related OTUs (50.9%; 58.53%), smaller abundances of Bacteroidetes (13.27%; 3.8%), Acidobacteria (9.38%; 3.4%) and Actinobacteria (9.25%; 10.49%) were present in both habitats. Similar to some other waters, smaller fractions of Gallionella-related OTUs (5.94%; 16.23%), Sideroxydans (3.85%; 1.78%), Sulfuricurvum (1.33%; 4.61%) and Sulfurimonas (2.47%; 1.04%) were observed. The occurrence of greater fractions of Flavobacteriaceae and especially OTUs related to Flavobacterium (5.52%) was a unique feature of the mofette water at Bublák C. On a phylum level, the community structure of the surface water (Kopanina) was similar to the other surface waters, with high abundances of Proteobacteria (60.28%), Bacteroidetes (6.84%), Actinobacteria (6.57%), and Parcubacteria (6.71%). However, on a deeper taxonomic level, several differences compared to all other waters were observed. The most abundant genera were Chromobacterium (9.18%), Sulfurimonas (7.73%), Sideroxydans (7.18%), and Ferritrophicum (5.44%).
Overall, the investigated waters shared 707 out of 1,399 assignable genera. This shared fraction (generalists) comprised 50.5% of all taxa and 98.9% of the total read counts in the data set (Figure 5). On the other hand, unique and site specific genera (specialists) were rare and made up 126 assignable genera (9% of all taxa) and 0.05% of the total read counts. Based on the total read counts in the data set, most abundant common taxa were the iron oxidizers Gallionella (13.71%), another Gallionellaceae-related taxon (3.06%), Sideroxydans (2.59%), the sulfur oxidizers Sulfuricurvum (9.46%) and Sulfurimonas (3.43%) and taxa related to Comamonadaceae (5.47%) and Intrasporangiaceae (2.81%). None “specialist” taxa showed relative abundances above 0.03%.
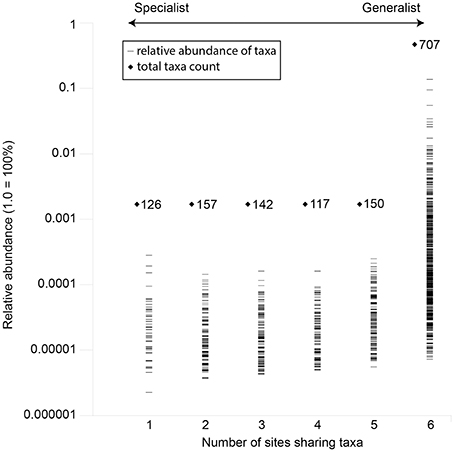
Figure 5. Occurrence and mean abundance of generalists and site-specific specialists across the investigated mofette sites. Unassigned taxa were not considered.
In all investigated waters more than 50 obligate marine taxa, such as Nitrospinaceae, Maritimimonas, Oceanobacillus, Marinobacter, Marinilabiaceae, Pseudohongiella, Acidimicrobiaceae (marine group), Desulfatiferula, Dehalococcoidia (GIF3, GIF9, MSBL5, Napoli-4B) and Thaumarchaeota (marine group) were identified (Table S4). The deep mineral waters of Soos showed the largest fraction of these marine taxa (rel. abundance 2.22%), whereby the relative abundance at the other sites was smaller (0.09–0.28%).
Correlation between Environmental Parameters and the Microbial Communities
The relationship between microbial community structure and the environment was examined by canonical correlation analysis (Figure 6). The pH value (23.9%), Fe2+ concentration (18.5%), and conductivity (18.2%) were the optimal subset of environmental parameters to explain the community structure of the investigated waters (all p-values < 0.05). Thereby, the strength of influence of given parameters on the sites differed greatly. The community structures from both U Mostku and Soos correlate positively with pH and conductivity. The Fe2+ concentration showed a strong positive correlation with the community structure of the mineral spring water (Plesná). On the contrary, a moderate negative correlation with the communities from Bublák C and Bublák NW was detected. Both Fe2+ concentration and pH had a strong negative correlation with the microbial composition from the Kopanina water. Correlation coefficients for the most abundant taxa and all measured environmental parameters were calculated. Thereby, Sideroxydans showed a strongly negative (p = 0.03, R2 = 0.69) and Sulfuricurvum a strongly positive (p = 0.03, R2 = 0.71) correlation with the pH value.
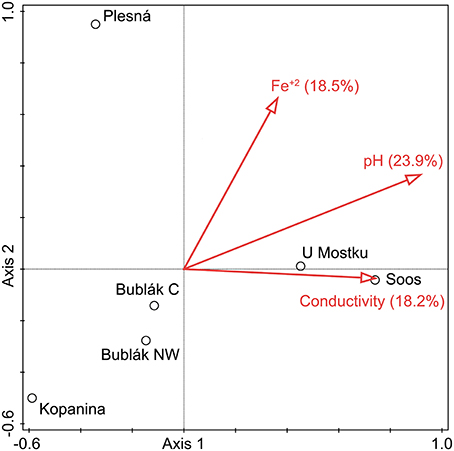
Figure 6. Canonical correlation analysis of the microbial composition on OTU level and environmental parameters. If the Bonferroni corrected padj was <0.05, a given parameter was included. Conductivity, pH and Fe2+ concentration explained 60.6% of the compositional variation among the investigated waters. Circles indicate sampling locations.
To examine possible correlations of gene abundances to the environmental parameters, the Pearson correlation coefficient was calculated. A strong positive correlation between dissolved organic carbon and the abundance of 16S rRNA genes (p = <0.02; R2 = 0.79) was observed. Due to low abundances, this calculation could not be performed for dsrB and mcrA genes.
Discussion
Natural CO2 conduits offer a unique opportunity to investigate adaptation of microbial communities to extremely elevated CO2 concentrations and its side-effects. Likewise, insight into geo-bio interactions and matter cycling in a habitat interconnected with the deep subsurface can be gained. Several studies focused on hydrochemical and geophysical features of wet mofettes (Pfanz, 2008; Kämpf et al., 2013), but biological processes were so far not considered. Most knowledge on environmental microbial communities in habitats with elevated CO2 concentrations were either gained from dry mofettes or soils (Krüger et al., 2009, 2011; Beulig et al., 2016), Carbon Capture and Storage (CCS) sites (Morozova et al., 2010; Wandrey et al., 2011a,b; Pellizzari et al., 2016) and deep subsurface fluids (Emerson et al., 2016; Probst et al., 2017).
The present interdisciplinary survey provides insight into the hydrochemistry and complex microbial community structure and ecology of six CO2 influenced mofette and mineral waters of the Cheb Basin. The microbiological survey introduces a new perspective on mofettes. Community analyses revealed that mofettes are either analogs or rather windows into the deep biosphere and furthermore enable access to deeply buried paleo-sediments.
The Core Community of Wet Mofettes in the Cheb Basin
Hydrochemical analyses revealed that the investigated sites do not only differ in terms of physico-chemical properties (pH, conductivity, temperature, ion composition, DOC) but also with respect to their water origin (δ2H: δ18O, δ18O: δ34S) (Table 1). Waters of Plesná, U Mostku, and Bublák NW are shaped through meteoric origin whereby Soos and Bublák C waters bear higher proportions of deep ground water. Most likely the differences between both Bublák sites can be explained by the rate of degassing and admixing of shallow groundwater.
With respect to the differing origins and hydrochemistry of the investigated waters, it is remarkable that a core microbial community can be defined which made up the majority of the observed taxa. In total 50.5% of all taxa (98.9% of all reads) occurred at all sites. The microbial communities were mainly composed of putative microaerophilic, anaerobic lithoautotrophic organisms, capable to fix the provided magmatogene CO2 and gain energy by oxidation or reduction of inorganic substrates such as iron or sulfur. The simultaneous occurrence of organotrophic species can be linked to the admixture from surrounding soil. Potential phototrophic organisms (Table S4) played only a role at the ponds from unprotected springs (Bublák and Plesná). DOC data from the ponds in Bublák and Plesná, where foliage builds large benthic and littoral layers, led to the assumption that microbial degradation of complex organic carbon is restricted under anaerobic or microaerophilic conditions. This degradation inhibiting effect of CO2 coupled to anoxia and low pH was also reflected in the low amounts of organic acids. Due to these conditions, the up streaming CO2 was already considered as primary carbon source for microbial metabolism at mofette soils (Beulig et al., 2016).
The genera Gallionella (13.71%), another Gallionellaceae-related taxon (3.06%), Sideroxydans (2.59%), the sulfur oxidizers Sulfuricurvum (9.46%), and Sulfurimonas (3.43%) were identified as generalists occurring at all sites in high relative abundances. At the same time, the SIMPER analysis revealed that the abundances of these taxa determine the differences between the microbial communities of the surface and subsurface waters (Table S3). Similar taxonomic and physiological groups (e.g., Gallionellales, sulfur oxidizers) were dominant in deep, CO2 affected subsurface fluids (Emerson et al., 2016). Therefore, microbial iron and sulfur oxidation might play an important role in CO2 influenced aquatic subsurface systems. In contrast, only a few specialists such as Hadesarchaea were site-specific. The identification of such a copious core community (98.3–99.62% of total counts at a given site) could indicate connectivity through a deep aquifer between the investigated sites, despite the observed different geochemical properties of the waters and the distances of more than 15 km. Accordingly, multivariate statistics showed that the differing, site-specific environmental parameters (e.g., Fe+2 concentration, conductivity) did not determine the occurrence of most taxa, but rather shifted the community composition and relative abundances of certain organisms (Figure 6).
Quantity and Diversity of Microorganisms with Respect to Environmental Parameters
Our results indicate that a high CO2 partial pressure in aquatic habitats does not necessarily cause low microbial abundances, since the autochthonous microorganisms are well adapted to these conditions. The observed abundances for 16S rRNA genes (qPCR, Table 2) in the investigated waters are in compliance with studies in similar, but not CO2 affected habitats, for instance ground water or water from granite systems (Griebler and Lueders, 2009 and references therein). The differences in microbial abundances between the investigated waters could be explained by the varying amount of allochthonous influx of substrates and organisms. Comparative analyses of the water phases and the surrounding sediments indicated a rather different microbial community (Figure 3). The same argument explains the higher microbial diversity in surface waters compared to the subsurface originated waters, namely U Mostku and Soos.
The influence of elevated CO2 concentrations coupled with changes of pH as well as high concentrations of iron mainly shaped the community structure and abundances of specific taxa (Figures 1, 4). In addition, the Fe2+ concentration showed a strong negative correlation with the diversity. In the case of the investigated waters, higher Fe2+ concentrations led to a decreased diversity and an increased abundance of iron oxidizing bacteria, especially of Gallionella-related organisms. The only exception to this observation was the water from Soos. Even though the Fe2+ concentration was on par with the one measured in the water from Plesná, the very low amount of oxygen and high salinity inhibits the growth of Gallionella-related organisms (McBeth et al., 2013). Generally, decreased bacterial community richness, evenness and diversity have been reported for dry mofette sites (Sáenz de Miera et al., 2014). Our results indicate that an active fluid flow, driven by the uprising CO2, provides better substrate supply and enables a higher microbial diversity in comparison to dry mofette sites.
In order to interpret the ecological function of the detected microbial communities, we assigned the functional potential of single OTUs based on a literature review. Despite the fact that this approach cannot guarantee completeness and accuracy, we can observe interesting patterns in respect to the major matter cycles. The occurrence of iron-oxidizing bacteria was a common feature across all investigated waters. They were especially abundant in surface waters (Plesná, Bublák Central, Bublák NW, Kopanina). Besides smaller quantities of Sideroxydans- and Ferritrophicum-related organisms, Gallionella was the most abundant genus related to iron cycling. Organisms affiliated to the genus Gallionella are chemolithoautotrophic, microaerophilic iron-oxidizers, which increase their biomass production with increased carbon dioxide concentrations (Hallbeck et al., 1993). Gallionella-related organisms are common in various freshwater habitats where ferruginous water comes in contact with oxygen, for instance spring water or groundwater. Their habitat needs a low redox potential, a pH 6–7.6, low oxygen concentration (0.1–1 mg L−1), CO2 > 20 mg L−1 and high amounts of Fe+2 (5–25 mg L−1) (Hanert, 1975). Further, it is known that Gallionella stimulates the formation of iron oxides at the early stage of clogging (Wang et al., 2014), thereby affecting injection or re-injection rates of wells. Interestingly, the most acidic surface water (Kopanina) showed no predominance of Gallionella, but a shift to Sideroxydans- and especially acidophilic Ferritrophicum-related organisms. Metagenomic studies on deeply sourced CO2-enriched fluids showed that the metabolic capability of nitrate/nitrite reduction could be present in Gallionellales-related organisms (Emerson et al., 2016), increasing the fitness in a microaerophilic/anaerobic environment. CO2 affected surface waters could represent oxic-anoxic transition zones, which provide beneficial conditions for the growth of iron oxidizing bacteria. Both the acidification of the respective water (pH 4.2–6), and the low oxygen concentrations, due to a high CO2 partial pressure, promote microbial iron oxidation in such a habitat. This is reflected in the high abundances of iron oxidation related genera in the studied surface waters, which made up to approximately 20% of the microbial communities.
Another common feature of the investigated waters is the occurrence of sulfur cycle related organisms. Especially the waters from U Mostku and Soos showed high abundances of microorganisms involved in sulfur cycling. These results coincide with the isotopic ratio of the dissolved sulfate of the waters, which indicate bacterial oxidation of sulfur and reduced sulfur compounds to sulfate. The largest fraction of sulfur cycling related taxa is associated with sulfur oxidation. The genus Sulfuricurvum was highly abundant, making up approximately 20% of the microbial communities from the subsurface waters (U Mostku, Soos). Sulfuricurvum-related organisms are described as anaerobic/microaerophilic and sulfur-oxidizing chemolithoautotrophs, which can use a variety of electron acceptors (e.g., oxygen, nitrate) and electron donors (e.g., elemental sulfur, sulfide, thiosulfate) (Kodama and Watanabe, 2004) and are widely distributed in subsurface habitats (e.g., Engel et al., 2003). Besides sulfur oxidation related organisms, sulfate reducing bacteria (SRB) were present in CO2 influenced mofette and mineral waters, which made up to 0.5–3.8% of the total microbial community. Sulfate-reducing bacteria obtain energy by coupling the oxidation of organic compounds (e.g., alcohols, organic acids) or H2 to the reduction of sulfate, generating hydrogen sulfide. The largest fraction of SRB was observed in the water containing the highest concentrations of acetic acid (U Mostku). It is assumable, that the limited availability of low molecular organic acids (e.g., acetic acid) due to a restricted degradation of organic material under anoxic conditions could inhibit the growth of SRB. At least 24 different taxa related to sulfate reduction were found. The most abundant genus Desulfosporosinus is, according to several studies (e.g., Senko et al., 2009; Sánchez-Andrea et al., 2015), associated with acidic environments. Strictly anaerobic sulfate reduction was also observed at dry mofettes, which most probably was enabled by the absence of oxygen and the presence of organic carbon and geogenic carbon dioxide and hydrogen (Beaubien et al., 2008; Frerichs et al., 2013). We conclude that both, wet and dry mofettes represent, depending on the availability of low molecular weight organic acids, suitable habitats for sulfate reducing bacteria.
Regarding the nitrogen cycle, several pathways were indicated by the taxonomic analyses. In total, 17 bacterial and archaeal taxa (2% of all reads) implicated in nitrification were found. Among the ammonia-oxidizing taxa, Nitrosomonadaceae and interestingly the archaeon “Candidatus Nitrosotalea” were most abundant. Recently, Lehtovirta-Morley et al. (2016) could show that the obligate acidophilic ammonia oxidizer “Candidatus Nitrosotalea devanaterra” contains genes encoding both a predicted high-affinity substrate acquisition system and potential pH homeostasis mechanisms absent in neutrophilic species. Therefore, previously proposed mechanisms used by ammonia-oxidizing bacteria for growth at low pH are not essential for archaeal ammonia oxidation in acidic environments. Low abundances of taxa involved in the anaerobic oxidation of ammonia (ANAMMOX) were also found. Besides “Candidatus Anammoximicrobium,” also “Candidatus Brocadiaceae” was detected in the Kopanina and U Mostku waters. Whereby ubiquitous distributed nitrite-oxidizing bacterial (NOB) genera, such as Nitrospira and Nitrobacter, were found in low relative abundances, “Candidatus Nitrotoga”-like bacteria (Alawi et al., 2007) were dominating the NOB community. Again, most likely the pH is the crucial environmental parameter controlling the distribution pattern of the microbial community. Community analyses of nitrifying biofilms revealed a coexistence of Nitrospira and “Candidatus Nitrotoga,” and it is hypothesized that a slightly acidic pH in combination with lower temperatures favors the growth of the latter (Alawi et al., 2009; Hüpeden et al., 2016). We conclude that “Candidatus Nitrotoga” is not only from high importance in permafrost regions, rivers or wastewater plants, but in general in microaerophilic habitats facing mean temperatures below 18°C.
In addition to iron, sulfur and nitrogen cycling microorganisms, archaea related to methane cycling were found. These methanogenic archaea showed, in relation to the whole community, low abundances. Only the shallow subsurface water (U Mostku) had major portions in methanogenic archaea (Methanoregula) in relation to the total community. Methanoregula is an acidophilic methanogen, which utilizes hydrogen and CO2 but no organic compound for methanogenesis (Brauer et al., 2011), and is widely distributed in different environments across the globe (Wen et al., 2017; Yang et al., 2017). Furthermore, acidophilic Methanoregulaceae-related microorganisms have been observed before at dry mofette sites (Beulig et al., 2015). Methanoregula depends on acetate for growth, which was mainly present in the shallow subsurface water U Mostku (1.6 mg L−1; Table 1). Therefore, we hypothesize that Methanoregulaceae play an important role in carbon cycling in the anoxic environment of wet and dry mofettes. In contrast, Beaubien et al. (2008) observed an absence of hydrogen-dependent methanogenesis at mofette sites and suggested this pathway is inhibited in high CO2 environments due to a lack of hydrogen. The abundance of this genus militates against a putative inhibition of hydrogen dependent methanogenesis in such a habitat. Despite the abundance of methanogenic archaea, observed methane fluxes were very low and the isotopic data indicated a thermogenic origin of the methane. It should be considered that a mixture of methane formed biogenically near the surface (δ13C ≈ −80‰) and highly 13C-enriched methane originating from the upper mantle (13C ≈ −15‰; e.g., Etiope and Sherwood Lollar, 2013) could result in the measured δ13C-methane values (Bräuer et al., 2005). Therefore, microbial methane production cannot be completely ruled out, especially because methanogens were detected in each of the investigated mofette waters.
Interconnection of Mofette Waters with Paleo-Sediment and the Deep Biosphere
Our results, based on hydrochemical and isotopic analyses as well as Illumina 16S rRNA gene sequencing, not only provide first evidence that the surface waters are interconnected via the up streaming fluids with deep subsurface paleo-sediment but in addition they are linked to the deep biosphere.
Hadesarchaea (formerly South-African Gold Mine Miscellaneous Euryarchaeal Group, SAGMEG) were solely found in the Soos mineral water (0.4%). This group is described as metabolically versatile and shares several physiological mechanisms with strict anaerobic Euryarchaeota (Takai et al., 2001; Parkes et al., 2005; Biddle et al., 2006). They are prominent members of the deep subsurface biosphere and occur both in terrestrial and marine environments, including hot springs. Based on genomic reconstructions, it is assumed that Hadesarchaea are mediating key geochemical processes which allow them to successfully inhabit the deep subsurface (Baker et al., 2016). They show metabolic characteristics, such as CO and H2 oxidation (or H2 production), with potential coupling to nitrite reduction to ammonia (DNRA). Hadesarchaea possess central carbon metabolic (C1 pathway) genes, which may be used for carbon fixation (Baker et al., 2016) and thereby fit to the conditions in the mineral water of Soos.
Besides of Euryarchaeota involved in methane cycling, we identified Bathyarchaeota (formerly Miscellaneous Crenarchaeota Group 1, 6, 7/17 and 15). These organisms have been found in marine sediments and deep aquifer waters and seem to be capable of acetogenesis and methane metabolism (Kubo et al., 2012; Evans et al., 2015; He et al., 2016). Sequences of this group not only have been found in the mineral fluid with a deep origin (Soos, 0.82%), but also in smaller quantities in the CO2 affected surface waters. In the extreme environment of active CO2 conduits, these organisms could represent another player involved in anaerobic carbon cycling. Furthermore, this finding is, in addition to the occurrence of Hadesarchaea, another indicator for the connection between surface and deep subsurface habitats—or reflects deep biosphere conditions in a surface habitat.
The carbon/hydrogen isotope ratios of methane of the investigated sites indicated a thermal gas, similar to the methane of the high saline brine analyzed in the German Continental Deep Drilling Project (KTB) pilot borehole, which is located approximately 50–80 km away from the investigated mofette region (Faber, 1995; Möller et al., 1997; Lippmann et al., 2005). Both, the methane of the KTB borehole and the methane of the free gases from mofettes of the Cheb Basin could be linked to the same or similar marine-influenced Permo-Carboniferous sedimentary basin.
The finding, that the mofette waters are potentially interconnected to deeply buried marine paleo-sediment, is supported by the observation of more than 50 taxa known to be obligate “marine” organisms, which were to our best knowledge, so far not observed in shallow terrestrial Central European ecosystems. The highly saline deep waters might serve sufficient conditions for the microorganisms to survive and proliferate; however, this explanation does not provide an adequate answer of their origin. Combining the results of the microbiological and isotopic water/gas analyses, the saline fluids are most likely allochthonous and migrate from the sediments of marine origin of one of the Permo-Carboniferous basins into the vicinity, for instance the Weiden Basin, located approximately 50–80 km SW of the Cheb Basin. Originally, this hypothesis was formulated for the highly saline basement brines of the KTB-pilot hole (Faber, 1995; Möller et al., 1997, 2005; Lippmann et al., 2005). Möller et al. (2005) assumed that during the Upper Cretaceous uplift of the adjacent basement (>2,000 m according to Wagner et al., 1997) the formation waters from Triassic to Carboniferous strata penetrated along the Franconian lineament NE- wards into metamorphic sequences of the crystalline basement of the Bohemian Massif. Infiltration of the brine might have occurred about 70 Ma concurrent with the uplift of the Bohemian Massif.
The microbial community of the investigated waters consists of a mixture of species which originate from the deep subsurface (saline thermal waters and paleo-sediments) and the surface (shallow groundwater aquifers filled with meteoric water and surrounding sediments). The emanating CO2-rich fluids cross and interconnect these habitats. Our analysis of the carbon/hydrogen isotope ratios of methane linked the origin of the fluids to a marine-influenced sedimentary basin. Therefore, marine species might originate from deep saline aquifers or marine sediments from the Weiden Basin. These marine species possibly persisted over millions of years in great depth and are transported to the surface via the emanating waters. These observations strengthen our assumption that wet mofettes are not only partly analogs but moreover windows to the deep biosphere and paleo-sediment.
However, it cannot be ruled out that the particular geochemical setting of the fluids enable marine species, introduced through meteoric waters, to proliferate. In 2016, a drilling campaign was conducted at one of the mofette fields in the Cheb Basin (Bussert et al., 2017). The 108.5 m deep drilling into a CO2 conduit will provide further insights into microbial processes and the origin of these microorganisms.
Conclusion
The present interdisciplinary survey on wet mofettes improves our understanding of microbial life under strongly elevated CO2 concentrations. The results from hydrochemical analyses, from isotopic signatures of key elements and from the 16S rRNA gene profiling unveil a new perspective on mofette research. The results indicate that mofette waters in the Cheb Basin are connected with the deep subsurface, specifically paleo-sediments and the deep biosphere. Therefore, mofettes provide access to deeply buried geo-bio-archives. Further on, it has to be considered that the mofette waters are at least in one direction interconnected. This argument is strengthened by the observation that among all investigated sites a large fraction of generalists and only a very small fraction of site specific specialists was found. Moreover, the described community in surface waters from mofettes of central Europe is in large proportions similar to the deep biosphere of geysers and marine thermal vents, such as black smokers. Mainly the influence of elevated CO2 concentrations coupled with changes of pH as well as varying concentrations of iron shaped the community structures and abundances of specific taxa. The gained knowledge and especially the community shift to taxa well adapted to low pH might be useful regarding geo-engineered systems (e.g., geothermal energy, wastewater treatment, drinking water reservoirs, regional spas and carbon or hydrogen subsurface storage facilities).
Author Contributions
PK performed DNA extractions and prepared genetic analyses and gene quantifications, and contributed writing the present manuscript. HK sampled during the field campaign, performed geochemical analyses and contributed writing the manuscript. FH performed bioinformatic analyses of the sequence data set and contributed writing the manuscript. QL was involved interpreting the results of the community study and designed figures. AV performed isotopic analyses and interpreted the results. DW provided important financial and technical support and supported the writing of the manuscript. MA designed the study and supervised DNA extractions and genetic analyses and led the writing of the present manuscript. All authors have taken part in the manuscript revisions and agreed with its scientific content.
Funding
This study was supported by the Deutsche Forschungsgemeinschaft (DFG) within the framework of the priority program 1006 “International Continental Drilling Program” (ICDP) by a grant to MA (AL 1898/1) and HK (KA 902/16).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank K. Knöller (Helmholtz Centre for Environmental Research–UFZ, Department Catchment Hydrology) for the 18O, 2H, 34S ratio measurements on water, J. Tesar (Laborunion CZ, Františkový Lázne) for the gas composition measurements. Special thanks to Oliver Burckhardt (GFZ German Research Centre for Geosciences) for his valuable help during the sampling campaigns and support in the lab, and Sabine Tonn and Kristin Günther (both GFZ German Research Centre for Geosciences) for hydrochemical measurements.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02446/full#supplementary-material
References
Alawi, M., Lipski, A., Sanders, T., and Spieck, E. (2007). Cultivation of a novel cold-adapted nitrite oxidizing betaproteobacterium from the Siberian Arctic. ISME J. 1, 256–264. doi: 10.1038/ismej.2007.34
Alawi, M., Off, S., Kaya, M., and Spieck, E. (2009). Temperature influences the population structure of nitrite-oxidizing bacteria in activated sludge. Environ. Microbiol. Rep. 1, 184–190. doi: 10.1111/j.1758-2229.2009.00029.x
Baker, B. J., Saw, J. H., Lind, A. E., Lazar, C. S., Hinrichs, K.-U., Teske, A. P., et al. (2016). Genomic inference of the metabolism of cosmopolitan subsurface Archaea, Hadesarchaea. Nat. Microbiol. 1:16002. doi: 10.1038/nmicrobiol.2016.2
Bankwitz, P., Schneider, G., Kämpf, H., and Bankwitz, E. (2003). Structural characteristics of epicentral areas in Central Europe: study case Cheb Basin (Czech Republic). J. Geodyn. 35, 5–32. doi: 10.1016/S0264-3707(02)00051-0
Beaubien, S. E., Ciotoli, G., Coombs, P., Dictor, M. C., Krüger, M., Lombardi, S., et al. (2008). The impact of a naturally occurring CO2 gas vent on the shallow ecosystem and soil chemistry of a Mediterranean pasture (Latera, Italy). Int. J. Greenhouse Gas Control 2, 373–387. doi: 10.1016/j.ijggc.2008.03.005
Beulig, F., Heuer, V. B., Akob, D. M., Viehweger, B., Elvert, M., Herrmann, M., et al. (2015). Carbon flow from volcanic CO2 into soil microbial communities of a wetland mofette. ISME J. 9, 746–759. doi: 10.1038/ismej.2014.148
Beulig, F., Urich, T., Nowak, M., Trumbore, S. E., Gleixner, G., Gilfillan, G. D., et al. (2016). Altered carbon turnover processes and microbiomes in soils under long-term extremely high CO2 exposure. Nat. Microbiol. 1:15025. doi: 10.1038/nmicrobiol.2015.25
Biddle, J. F., Lipp, J. S., Lever, M. A., Lloyd, K. G., Sørensen, K. B., Anderson, R., et al. (2006). Heterotrophic Archaea dominate sedimentary subsurface ecosystems off Peru. Proc. Natl. Acad. Sci. U.S.A. 103, 3846–3851. doi: 10.1073/pnas.0600035103
Bräuer, K. H., Kämpf, U., and Koch Strauch, G. (2011). Monthly monitoring of gas and isotope compositions in the free gas phase at degassing locations close to the Nový Kostel focal zone in the western Eger Rift, Czech Republic. Chem. Geol. 290, 163–176. doi: 10.1016/j.chemgeo.2011.09.012
Bräuer, K. H., Kämpf, U., Koch, S., Niedermann, S., and Strauch, G. (2007). Seismically induced changes of the fluid signature detected by a multi-isotope approach (He, CO2, CH4, N-2) at the Wettinquelle, Bad Brambach (central Europe). J. Geophys. Res. 112:B04307. doi: 10.1029/2006JB004404
Bräuer, K., Kämpf, H., and Strauch, G. (2014). Seismically triggered anomalies in the isotope signatures of mantle-derived gases detected at degassing sites along two neighboring faults in NW Bohemi, central Europe. J. Geophys. Res. Solid Earth 119, 5613–5632. doi: 10.1002/2014JB011044a
Bräuer, K., Kämpf, H., Faber, E., Koch, U., Nitzsche, H.-M., and Strauch, G. (2005). Seismically triggered microbial methane production relating to the Vogtland-NW Bohemia earthquake swarm period 2000, Central Europe. Geochem. J. 39, 441–450. doi: 10.2343/geochemj.39.441
Bräuer, K., Kämpf, H., Niedermann, S., Strauch, G., and Tesar, J. (2008). Natural laboratory NW Bohemia: comprehensive fluid studies between 1992 and 2005 used to trace geodynamic processes. Geochem. Geophys. Geosys. 9, 4, 1–30. doi: 10.1029/2007GC001921
Brauer, S. L., Cadillo-Quiroz, H., Ward, R. J., Yavitt, J. B., and Zinder, S. H. (2011). Methanoregula boonei gen. nov., sp. nov., an acidiphilic methanogen isolated from an acidic peat bog. Int. J. System. Evol. Microbiol. 61, 45–52. doi: 10.1099/ijs.0.021782-0
Bussert, R., Kämpf, H., Flechsig, C., Hesse, K., Nickschick, T., Liu, Q., et al. (2017). Drilling into an active mofette: pilot-hole study of the impact of CO2-rich mantle-derived fluids on the geo–bio interaction in the western Eger Rift (Czech Republic). Sci. Dril. 23, 13–27. doi: 10.5194/sd-23-13-2017
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Caporaso, J. G., Lauber, C. L., Walters, W. A., Berg-Lyons, D., Lozupone, C. A., Turnbaugh, P. J., et al. (2011). Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U.S.A. 108, 4516–4522. doi: 10.1073/pnas.1000080107
Chiodini, G., Caliro, S., Cardellini, C., Avino, R., Granieri, D., and Schmidt, A. (2008). Carbon isotopic composition of soil CO2 efflux, a powerful method to discriminate different sources feeding soil CO2 degassing in volcanic-hydrothermal areas. Earth Planet. Sci. Lett. 274, 372–379. doi: 10.1016/j.epsl.2008.07.051
DeSantis, T. Z., Hugenholtz, P., Larsen, N., Rojas, M., Brodie, E. L., Keller, K., et al. (2006). Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 72, 5069–5072. doi: 10.1128/AEM.03006-05
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Emerson, J. B., Thomas, B. C., Alvarez, W., and Banfield, J. F. (2016). Metagenomic analysis of a high carbon dioxide subsurface microbial community populated by chemolithoautotrophs and bacteria and archaea from candidate phyla. Environ. Microbiol. 18, 1686–1703. doi: 10.1111/1462-2920.12817
Engel, A. S., Lee, N., Porter, M. L., Stern, L. A., Bennett, P. C., and Wagner, M. (2003). Filamentous “Epsilonproteobacteria” Dominate Microbial Mats from Sulfidic Cave Springs. Appl. Environ. Microbiol. 69, 5503–5511. doi: 10.1128/AEM.69.9.5503-5511.2003
Etiope, G., and Sherwood Lollar, B. (2013). Abiothic methane on earth. Rev. Geophys. 51, 276–299. doi: 10.1002/rog.20011
Evans, P. N., Parks, D. H., Chadwick, G. L., Robbins, S. J., Orphan, V. J., Golding, S. D., et al. (2015). Methane metabolism in the archaeal phylum Bathyarchaeota revealed by genome-centric metagenomics. Science 350, 434–438. doi: 10.1126/science.aac7745
Evans, W. C., Sorey, M. L., Cook, A. C., Kennedy, B. M., and Shuster, E. M. (2002). Tracing and quantifying magmatic carbon discharge in cold groundwaters: lessons learned from Mammoth Mountain, USA. J. Volcanol. Geother. Res. 114, 291–312. doi: 10.1016/S0377-0273(01)00268-2
Faber, E. (1995). Origin of hydrocarbon gases in the pump-test of the KTB pilot well. Sci. Drill. 5, 123–128.
Fischer, T., and Horálek, J. (2003). Space-time distribution of earthquake swarms in the principal focal zone of the NWBohemia/Vogtland seismoactive region. J. Geodyn. 35, 125–144. doi: 10.1016/S0264-3707(02)00058-3
Fischer, T., Horálek, J., Hrubcová, P., Vavryčuk, V., Bräuer, K., and Kämpf, H. (2014). Intra-continental earthquake swarms in West-Bohemia and Vogtland: a review. Tectonophysics 611, 1–27. doi: 10.1016/j.tecto.2013.11.001
Frerichs, J., Oppermann, B. I., Gwosdz, S., Möller, I., Herrmann, M., and Krüger, M. (2013). Microbial community changes at a terrestrial volcanic CO2 vent induced by soil acidification and anaerobic microhabitats within the soil column. FEMS Microbiol. Ecol. 84, 60–74. doi: 10.1111/1574-6941.12040
Geets, J., Borremans, B., Diels, L., Springael, D., Vangronsveld, J., van der Lelie, D., et al. (2006). DsrB gene-based DGGE for community and diversity surveys of sulfate-reducing bacteria. J. Microbiol. Methods 66, 194–205. doi: 10.1016/j.mimet.2005.11.002
Girault, F., and Perrier, F. (2014). The Syabru-Bensi hydrothermal system in central Nepal: 2. Modeling and significance of the radon signature. J. Geophys. Res. Solid Earth 119, 4056–4089. doi: 10.1002/2013JB010302
Girault, F., Perrier, F., Crockett, R., Bhattarai, M., Koirala, B. P., France- Lanord, C., et al. (2014). The Syabru-Bensi hydrothermal system in central Nepal: 1. Characterization of carbon dioxide and radon fluxes. J. Geophys. Res. Solid Earth 119, 4017–4055. doi: 10.1002/2013JB010301
Griebler, C., and Lueders, T. (2009). Microbial biodiversity in groundwater ecosystems. Freshw. Biol. 54, 649–677. doi: 10.1111/j.1365-2427.2008.02013.x
Hainzl, S., Fischer, T., Cermáková, H., Bachura, M., and Vlček, J. (2016). Aftershocks triggered by fluid intrusion: evidence for the aftershock sequence occurred 2014 in West Bohemia/Vogtland. J. Geophys. Res. Solid Earth 121, 2575–2590. doi: 10.1002/2015JB012582
Hallbeck, L., Staahl, F., and Pedersen, K. (1993). Phytogeny and phenotypic characterization of the stalk-forming and iron-oxidizing bacterium Gallionella ferruginea. Microbiology 139, 1531–1535.
Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electr. 4:9.
Hanert, H. (1975). Entwicklung, Physiologie und Ökologie des Eisenbacteriums Gallionella Ferruginea Ehrenberg. Professorial dissertation. Naturwissenschaftliche Fakultät der Technischen Universität Braunschweig, 1–104.
He, Y., Li, M., Perumal, V., Feng, X., Fang, J., Xie, J., et al. (2016). Genomic and enzymatic evidence for acetogenesis among multiple lineages of the archaeal phylum Bathyarchaeota widespread in marine sediments. Nat. Microbiol. 1:16035. doi: 10.1038/nmicrobiol.2016.35
Hüpeden, J., Wegen, S., Off, S., Lücker, S., Bedarf, Y., Daims, H., et al. (2016). Relative Abundance of Nitrotoga spp. in a Biofilter of a Cold-Freshwater Aquaculture Plant Appears To Be Stimulated by Slightly Acidic pH. Appl. Environ. Microbiol. 82, 1838–1845. doi: 10.1128/AEM.03163-15
Kämpf, H., Bräuer, K., Schumann, J., Hahne, K., and Strauch, G. (2013). CO2 discharge in an active, non-volcanic continental rift area (Czech Republic): characterisation (δ13C, 3He/4He) and quantification of diffuse and vent CO2 emissions. Chem. Geol. 339, 71–83. doi: 10.1016/j.chemgeo.2012.08.005
Kämpf, H., Geissler, W. H., and Bräuer, K. (2007). “Combined gas-geochemical and receiver function studies, of the Vogtland/NW-Bohemia intraplate mantle degassing field Central Europe,” in Mantle Plumes—A Multidisciplinary Approach, eds J. R. R. Ritter and U. R. Christiansen (Berlin; Heidelberg; New York, NY: Springer-Verlag), 127–158.
Kodama, Y., and Watanabe, K. (2004). Sulfuricurvum kujiense gen. nov., sp. nov., a facultatively anaerobic, chemolithoautotrophic, sulfur-oxidizing bacterium isolated from an underground crude-oil storage cavity. Int. J. Syst. Evol. Microbiol. 54, 2297–2300. doi: 10.1099/ijs.0.63243-0
Krüger, M., Jones, D., Frerichs, J., Oppermann, B. I., West, J., Coombs, P., et al. (2011). Effects of elevated CO2 concentrations on the vegetation and microbial populations at a terrestrial CO2 vent at Laacher See, Germany. Int. J. Greenhouse Gas Control 5, 1093–1098. doi: 10.1016/j.ijggc.2011.05.002
Krüger, M., West, J., Frerichs, J., Oppermann, B., Dictor, M.-C., Jouliand, C., et al. (2009). Ecosystem effects of elevated CO2 concentrations on microbial populations at a terrestrial CO2 vent at Laacher See, Germany. Ener. Proc. 1, 1933–1939. doi: 10.1016/j.egypro.2009.01.252
Kubo, K., Lloyd, K. G., Biddle, J. F., Amann, R., Teske, A., and Knittel, K. (2012). Archaea of the Miscellaneous Crenarchaeotal Group are abundant, diverse and widespread in marine sediments. ISME J. 6, 1949–1965. doi: 10.1038/ismej.2012.37
Lan, T. F., Yang, T. F., Lee, H.-F., Chen, Y.-G., Chen, C.-H., Song, S.-R., et al. (2007). Compositions and flux of soil gas in Liu-Huang-Ku hydrothermal area, northern Taiwan. J. Volcanol. Geother. Res. 165, 32–45. doi: 10.1016/j.jvolgeores.2007.04.015
Lehtovirta-Morley, L. E., Sayavedra-Soto, L. A., Gallois, N., Schouten, S., Stein, L. Y., Prosser, J. I., et al. (2016). Identifying Potential Mechanisms Enabling Acidophily in the Ammonia-Oxidizing Archaeon “Candidatus Nitrosotalea devanaterra.” Appl. Environ. Microbiol. 82, 2608–2619. doi: 10.1128/AEM.04031-15
Lippmann, J., Erzinger, J., Zimmer, M., Schloemer, S., Eichinger, L., and Faber, E. (2005). On the geochemistry of gases and noble gas isotopes (including 222Rn) in deep crustal fluids: the 4000 mKTB-pilot hole fluid production test 2002–03. Geofluids 5, 52–66. doi: 10.1111/j.1468-8123.2004.00108.x
Maček, I., Dumbrell, A. J., Nelson, M., Fitter, A. H., Vodnik, D., and Helgason, T. (2011). Local adaptation to soil hypoxia determines the structure of an arbuscular mycorrhizal fungal community in roots from natural CO2 Springs. Appl. Environ. Microbiol. 77, 4770–4777. doi: 10.1128/AEM.00139-11
Martin, M. (2011). Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 17, 10–12. doi: 10.14806/ej.17.1.200
McBeth, J. M., Fleming, E. J., and Emerson, D. (2013). The transition from freshwater to marine iron-oxidizing bacterial lineages along a salinity gradient on the Sheepscot River, Maine, USA: Estuarine iron mat communities. Environ. Microbiol. Rep. 5, 453–463. doi: 10.1111/1758-2229.12033
Mehlhorn, J., Beulig, F., Küsel, K., and Planer-Friedrich, B. (2014). Carbon dioxide triggered metal(loid) mobilisation in a mofette. Chem. Geol. 382, 54–66. doi: 10.1016/j.chemgeo.2014.05.027
Mehlhorn, J., Byrne, J. M., Kappler, A., and Planer-Friedrich, B. (2016). Time and telperature dependency of carbon dioxide triggered metal(loid) mobilization in soil. Appl. Geochem. 74, 122–137. doi: 10.1016/j.apgeochem.2016.09.007
Möller, P., Weise, S. M., Althaus, E., Bach, W., Behr, H. J., Borchardt, R., et al. (1997). Paleofluids and Recent fluids in the upper continental crust: results from the German Continental Deep Drilling Program (KTB). J. Geophys. Res. Solid Earth 102, 18233–18254.
Möller, P., Woith, H., Dulski, P., Lüders, V., Erzinger, J., Kämpf, H., et al. (2005). Main and trace elements in KTB-VB fluid: composition and hints to its origin. Geofluids 5, 28–41. doi: 10.1111/j.1468-8123.2004.00104.x
Morozova, D., Wandrey, M., Alawi, M., Zimmer, M., Vieth, A., Zettlitzer, M., et al. (2010). Monitoring of the microbial community composition in saline aquifers during CO2 storage by fluorescence in situ hybridisation. Int. J. Greenhouse Gas Control 4, 981–989. doi: 10.1016/j.ijggc.2009.11.014
Nadkarni, M. A., Martin, F. E., Jacques, N. A., and Hunter, N. (2002). Determination of bacterial load by real-time PCR using a broad-range (universal) probe and primers set. Microbiology 148, 257–266. doi: 10.1099/00221287-148-1-257
Nickschick, T., Kämpf, H., Flechsig, C., Mrlina, J., and Heinicke, J. (2015). CO2 degassing in the Hartoušov mofette area, western Eger Rift, imaged by CO2 mapping and geoelectrical and gravity surveys. Int. J. Earth Sci. 104, 2107–2129. doi: 10.1007/s00531-014-1140-4
Oppermann, B. I., Michaelis, W., Blumenberg, M., Frerichs, J., Schulz, H. M., Schippers, A., et al. (2010). Soil microbial community changes as a result of long-term exposure to a natural CO2 vent. Geochim. Cosmochim. Acta 74, 2697–2716. doi: 10.1016/j.gca.2010.02.006
Parkes, R. J., Webster, G., Cragg, B. A., Weightman, A. J., Newberry, C. J., Ferdelman, T. G., et al. (2005). Deep sub-seafloor prokaryotes stimulated at interfaces over geological time. Nature 436, 390–394. doi: 10.1038/nature03796
Pellizzari, L., Morozova, D., Neumann, D., Kasina, M., Klapperer, S., Zettlitzer, M., et al. (2016). Comparison of the microbial community composition of pristine rock cores and technical influenced well fluids from the Ketzin pilot site for CO2 storage. Environ. Earth Sci. 75:1323. doi: 10.1007/s12665-016-6111-6
Probst, A. J., Castelle, C. J., Singh, A., Brown, C. T., Anantharaman, K., Sharon, I., et al. (2017). Genomic resolution of a cold subsurface aquifer community provides metabolic insights for novel microbes adapted to high CO2 concentrations. Environ. Microbiol. 19, 459–474. doi: 10.1111/1462-2920.13362
Rennert, T., and Pfanz, H. (2016). Hypoxic and acidic — Soils on mofette fields. Geoderma 280, 73–81. doi: 10.1016/j.geoderma.2016.06.019
Rennert, T., Eusterhues, K., Pfanz, H., and Totsche, K. U. (2011). Influence of geogenic CO2 on mineral and organic soil constituents on a mofette site in the NW Czech Republic. Eur. J. Soil Sci. 62, 572–580. doi: 10.1111/j.1365-2389.2011.01355.x
Sáenz de Miera, L. E., Arroyo, P., de Luis Calabuig, E., Falagán, J., and Ansola, G. (2014). High-throughput sequencing of 16S RNA genes of soil bacterial communities from a naturally occurring CO2 gas vent. Int. J. Greenhouse Gas Control 29, 176–184. doi: 10.1016/j.ijggc.2014.08.014
Sánchez-Andrea, I., Stams, A. J. M., Hedrich, S., Nancucheo, I., and Johnson, D. B. (2015). Desulfosporosinus acididurans sp. nov.: an acidophilic sulfate-reducing bacterium isolated from acidic sediments. Extremophiles 19, 39–47. doi: 10.1007/s00792-014-0701-6
Senko, J. M., Zhang, G., McDonough, J. T., Bruns, M. A., and Burgos, W. D. (2009). Metal reduction at low pH by a Desulfosporosinus species: implications for the biological treatment of Acidic Mine Drainage. Geomicrobiol. J. 26, 71–82. doi: 10.1080/01490450802660193
Šibanc, N., Dumbrell, A. J., Mandić-Mulec, I., and Maček, I. (2014). Impacts of naturally elevated soil CO2 concentrations on communities of soil archaea and bacteria. Soil Biol. Biochem. 68, 348–356. doi: 10.1016/j.soilbio.2013.10.018
Šmilauer, P., and Lepš, J. (2014). Multivariate Analysis of Ecological Data using CANOCO 5. Cambridge: Cambridge University Press. doi: 10.1017/CBO9781139627061
Steinberg, L. M., and Regan, J. M. (2009). mcrA-Targeted real-time quantitative PCR method to examine methanogen communities. Appl. Environ. Microbiol. 75, 4435–4442. doi: 10.1128/AEM.02858-08
Takai, K., Moser, D. P., DeFlaun, M., Onstott, T. C., and Fredrickson, J. K. (2001). Archaeal diversity in waters from Deep South African Gold Mines. Appl. Environ. Microbiol. 67, 5750–5760. doi: 10.1128/AEM.67.21.5750-5760.2001
Vodnik, D., Pfanz, H., and Macek, I. (2002). Photosynthesis of Cockspur [Echinochloa crus-galli (L.) Beauv.] at Sites of Naturally Elevated CO2 Concentration. Photosynthetica 40, 575–579. doi: 10.1023/A:1024308204086
Wagner, G. A., Coyle, D. A., Duyster, J., Henjes-Kunst, F., Peterek, A., Schröder, B., et al. (1997). Post- Variscan thermal and tectonic evolution of the KTB site and its surroundings. J. Geophys. Res. 102:18221.
Wagner, M., Roger, A. J., Flax, J. L., Brusseau, G. A., and Stahl, D. A. (1998). Phylogeny of dissimilatory sulfite reductases supports an early origin of sulfate respiration. J. Bacteriol. 180, 2975–2982.
Wandrey, M., Fischer, S., Zemke, K., Liebscher, A., Scherf, A.-K., Vieth-Hillebrand, A., et al. (2011a). Monitoring petrophysical, mineralogical, geochemical and microbiological effects of CO2 exposure—Results of long-term experiments under in situ conditions. Ener. Proc. 4, 3644–3650. doi: 10.1016/j.egypro.2011.02.295
Wandrey, M., Pellizari, L., Zettlitzer, M., and Würdemann, H. (2011b). Microbial community and inorganic fluid analysis during CO2 storage within the frame of CO2SINK–Long-term experiments under in situ conditions. Ener. Proc. 4, 3651–3657. doi: 10.1016/j.egypro.2011.02.296
Wang, J., Sickinger, M., Ciobota, V., Herrmann, M., Rasch, H., Rösch, P., et al. (2014). Revealing the microbial community structure of clogging materials in dewatering wells differing in physico-chemical parameters in an open-cast mining area. Water Res. 63, 222–233. doi: 10.1016/j.watres.2014.06.021
Weinlich, F. H., Bräuer, K., Kämpf, H., Strauch, G., Tesar, J., and Weise, S. M. (1999). An active subcontinental mantle volatile system in the western Eger rift, central Europe: gas flux, isotopic (He, C, and N) and compositional fingerprints. Geochim. Cosmochim. Acta 63, 3653–3671.
Weinlich, F. H., Tesar, J., Weise, S., Bräuer, K., and Kämpf, H. (1998). Gas flux distribution in mineral springs and tectonic structure in the western Eger Rift. J. Czech. Geol. Soc. 43, 91–110.
Wen, X., Yang, S., Horn, F., Winkel, M., Wagner, D., and Liebner, S. (2017). Global biogeographic analysis of methanogenic archaea identifies community-shaping environmental factors of natural environments. Front. Microbiol. 8:1339. doi: 10.3389/fmicb.2017.01339
Yang, S., Liebner, S., Winkel, M., Alawi, M., Horn, F., Dörfer, C., et al. (2017). In-depth analysis of core methanogenic communities from high elevation permafrost-affected wetlands. Soil Biol. Biochem. 111, 66–77. doi: 10.1016/j.soilbio.(2017)0.03.007
Keywords: elevated CO2 concentration, microbial ecology, deep biosphere, Eger Rift, paleo-sediment, Sulfuricurvum, Gallionella, Sideroxydans
Citation: Krauze P, Kämpf H, Horn F, Liu Q, Voropaev A, Wagner D and Alawi M (2017) Microbiological and Geochemical Survey of CO2-Dominated Mofette and Mineral Waters of the Cheb Basin, Czech Republic. Front. Microbiol. 8:2446. doi: 10.3389/fmicb.2017.02446
Received: 27 September 2017; Accepted: 24 November 2017;
Published: 11 December 2017.
Edited by:
Mark Alexander Lever, ETH Zurich, SwitzerlandReviewed by:
Nikolai Ravin, Research Center for Biotechnology Russian Academy of Sciences, RussiaMichael J. Wilkins, Ohio State University, United States
Copyright © 2017 Krauze, Kämpf, Horn, Liu, Voropaev, Wagner and Alawi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mashal Alawi, bWFzaGFsLmFsYXdpQGdmei1wb3RzZGFtLmRl