 Marisa Haenni1*
Marisa Haenni1* Pierre Châtre1Céline Dupieux-Chabert2,3Véronique Métayer1
Pierre Châtre1Céline Dupieux-Chabert2,3Véronique Métayer1 Michèle Bes2,3
Michèle Bes2,3 Jean-Yves Madec1
Jean-Yves Madec1 Frédéric Laurent2,3,4
Frédéric Laurent2,3,4- 1Unité Antibiorésistance et Virulence Bactériennes, French Agency for Food, Environmental and Occupational Health Safety, Claude Bernard University Lyon 1, Lyon, France
- 2Bacteriology Department, Institut for Infectious Agents, French National Reference Centre for Staphylococci, Hospices Civils de Lyon, Lyon, France
- 3INSERM U1111, CNRS, International Center for Infectiology Research, UMR5308, École Normale Supérieure de Lyon, Claude Bernard University Lyon 1, Lyon, France
- 4Institut for Pharmaceutical and Biological Sciences of Lyon, Lyon, France
Methicillin-resistant Staphylococcus aureus (MRSA) has been reported as a worldwide pathogen in humans and animals including companion animals, i.e., cats, dogs, and horses. France lacked a comprehensive nationwide study describing the molecular features of MRSA circulating among companion animals over a large period of time. Here is reported the characterization of 130 non-duplicate clinical MRSA isolates collected from those three animal species from 2010 to 2015 through the French national Resapath network. Characterization of isolates was performed using phenotypic (antimicrobial susceptibility tests) and molecular (DNA arrays, spa-typing) methods. A horse-specific epidemiology was observed in France with the large dissemination of a unique clone, the CC398 clone harboring a Staphylococcal chromosomal cassette mec (SCCmec) type IV and spa-type t011. It was even the unique clone collected in 2015 whereas the clone CC8 USA500 (SCCmec type IV), classically described in horses, was present until 2014. Contrarily, cats and dogs were mainly infected by human-related MRSA isolates, i.e., clones usually reported in human infections, thus mirroring the human epidemiology in hospitals in France. Isolates belonging to the CC398 clone (SCCmec type IV or V) were also identified in 21.4% of dogs’ and 26.5% of cats’ MRSA isolates. In order to differentiate human-related from CC398 MRSA, tetracycline-resistance [or tet(M) detection] could be useful since this resistance is scarce in human-related strains but constant in CC398 MRSA isolates. In all, our data give a nationwide epidemiological picture of MRSA in companion animals over a 5-year period in France, adding further epidemiological information on the contribution of those animal species to a major public health issue. Considering the wide dissemination of CC398 MRSA isolates and the fact that 11/64 (17.2%) of them presented the Immune Evasion Cluster which enhances CC398 capacities to colonize humans, a specific attention should be paid in the coming years to determine the risk associated to the transmission in people in frequent contacts with companion animals. Our data also show that the prevalence of MRSA has likely decreased in cats, dogs, and horses between 2012 and 2015 in France. This trend should be monitored in the years to come.
Introduction
The role of animals in the transmission of methicillin-resistant Staphylococcus aureus (MRSA), has been extensively documented in the case of the livestock-associated MRSA (LA-MRSA) complex clonal (CC)398 in pigs and people in contact (Aires-de-Sousa, 2017). Whereas pig farming is restricted to specific areas and human communities, companion animals (including cats, dogs, and horses) are geographically wide spread and mostly not related to occupational activities. It has been estimated that 19 millions of cats and dogs live in households in France and that 750,000 equines (horses and ponies) are owned by stud farms or families for horse riding. Deciphering the routes of transmission of MRSA in companion animals is an ongoing issue. Even though all three animal species are in frequent and close contacts with humans, the epidemiology of MRSA is quite divergent, with cats and dogs being frequently colonized or infected by human clones and horses showing a species-specific MRSA distribution.
Several studies showed that MRSA clones circulating in cats and dogs are similar to the ones identified in humans and belong mostly to hospital-acquired clones (HA-MRSA) (Loeffler and Lloyd, 2010). This is clinically important since HA-MRSA isolates usually carry more virulence genes [such as those coding for enterotoxins or the toxic shock syndrome toxin (TSST)], and also more resistance genes than MRSA originating from animals (Mutters et al., 2016; Ballhausen et al., 2017). The only exception is the tet(M) gene, which is highly prevalent in LA-MRSA and MRSA isolates of animal origin but rare in HA-MRSA. This human-related epidemiology suggests that humans may be the source of MRSA isolated in cats and dogs, although those animals may act as a secondary reservoir capable of human re-infections in specific contexts (Harrison et al., 2014; Bierowiec et al., 2016). Reports on MRSA in cats and dogs from England, Germany, Switzerland, and Portugal described the predominance of the sequence type (ST)22 clone, also known as EMRSA-15 or Barnim clone, which is prevalent in human patients in the same countries (Strommenger et al., 2006; Grundmann et al., 2010; Harrison et al., 2014; Vincze et al., 2014; Couto et al., 2016; Wipf and Perreten, 2016). The CC398 LA-MRSA has also been sporadically reported in cats and dogs, but always as a minor lineage (Haenni et al., 2012; Vincze et al., 2014; Wipf and Perreten, 2016).
Methicillin-resistant Staphylococcus aureus epidemiology in horses presents two distinct time phases. Until the early 2000s, a majority of the reported strains in North America and Europe belonged to the clonal complex (CC)8, with a predominance of the CC8-IV USA500 clone, and clustered into the ST254 and ST8 (Weese et al., 2005a; Moodley et al., 2006; Cuny et al., 2008; Walther et al., 2009; van Balen et al., 2014). Most of these CC8 strains likely originated from humans, and subsequently adapted to horses and disseminated in the equine host, independent to the human epidemiology (Weese and van Duijkeren, 2010). After 2004, the emergence and dissemination of a CC398 MRSA clone [Staphylococcal chromosomal cassette mec (SCCmec) IV, spa-type t011] has been reported, from potential direct or indirect livestock origin (Cuny et al., 2008; Van den Eede et al., 2009; van Duijkeren et al., 2010), and this clone is now massively prevalent in horses worldwide. A CC398-IV-t011 MRSA sub-lineage – named clade C – has also been described, which most probably encompasses most of the CC398 MRSA that have already been described in hospitalized horses and veterinarians in contact (Abdelbary et al., 2014). This clade C is presenting a specific single-nucleotide polymorphism (SNP) consisting in a synonymous substitution in the SNP au309-2. It represents nearly 90% of the MRSA isolated from equine wound infections (Vincze et al., 2014) in Germany. A recent study shows it present since 2010 in France (Guerin et al., 2017). An important characteristic of the CC398 isolates is the potential presence of the Immune Evasion Cluster (IEC), composed of combined virulence factors, such as staphylococcal complement inhibitor (scn), chemotaxis inhibitory protein (chp), staphylokinase (sak), and specific staphylococcal enterotoxin genes such as sea and sep (Hau et al., 2015). This bacteriophage-encoded cluster is a marker of re-adaptation to the human host and enhances its human-to-human transmission capacity.
Methicillin-resistant Staphylococcus aureus colonizing or infecting companion animals have become of public health concern since transmissions between animals and humans have been documented many times (Weese et al., 2005b; Walther et al., 2009; van Duijkeren et al., 2010; Bergstrom et al., 2012; Van den Eede et al., 2013). In France, MRSA clones involved in human infections have been reported in specific studies in cats and dogs (Haenni et al., 2012) and recently in horses (Guerin et al., 2017). However, there is still no nationwide comprehensive molecular information on MRSA circulating in cats, dogs, and horses in France, and this was investigated and clarified here over a 5-year period through the French national network for surveillance of resistant bacteria in animals (Resapath1). This study was lined up with the 5-year French action plan EcoAntibio set up by the Ministry of Agriculture, Agri-Food and Forestry2 in 2011. Its first and recently achieved goal was to reduce animal exposure to antibiotics by 25%. The data presented here are of major importance for risk-assessment studies on MRSA transmission in a One-Health perspective.
Materials and Methods
Bacterial Strains
From December 2010 to April 2015, antimicrobial susceptibility test (AST) data of all coagulase-positive staphylococci isolated from horses, cats, and dogs (Supplementary Table S1) were collected through the Resapath network. The Resapath is a network of 78 veterinary laboratories that are transmitting on a voluntary basis all their AST data as well as the strains of interest to the ANSES (French agency for Food, Environmental and Occupational Health Safety). Only clinical isolates were considered in the frame of this study since the Resapath network does not encompass bacteria colonizing animals. All cefoxitin-intermediate or cefoxitin-resistant isolates, as tested by laboratories member of Resapath using the disk diffusion method (see below), were sent to the ANSES laboratory in Lyon for further characterization. After confirmation of the phenotype by disk diffusion, two PCRs were systematically performed: a triplex PCR targeting the 16S rRNA, mecA and nuc genes to detect the presence of MRSA (Maes et al., 2002), and a simplex PCR using mecC-specific primers to detect non-mecA MRSA (Garcia-Alvarez et al., 2011). Species identification was then further assessed using the PCR-RFLP designed by Blaiotta et al. (2010) to differentiate S. aureus from S. pseudintermedius, which is the most frequently encountered coagulase-positive Staphylococcus in dogs.
Susceptibility Testing
Antimicrobial susceptibility test was confirmed by the ANSES laboratory using the disk diffusion method and interpreted according to the guidelines of the Antibiogram Committee of the French Society for Microbiology3. S. aureus ATCC 25923 was used as quality control. In addition, 16 antibiotics of veterinary and/or human interest were tested: penicillin G, cefoxitin, cefovecine, kanamycin, gentamicin, tobramycin, tetracycline, erythromycin, spiramycin, lincomycin, chloramphenicol, florfenicol, fusidic acid, enrofloxacin, vancomycin, teicoplanin (Mast Diagnostics, Amiens, France).
Molecular Typing
The presence of the mecA/mecC gene was systematically confirmed by PCR (Maes et al., 2002; Garcia-Alvarez et al., 2011). Specific detection of the CC398 by PCR was also systematically performed (Stegger et al., 2011). This PCR is based on the detection of a specific sau1-hsdS1 gene (responsible for the restriction modification specificity), which is conserved among the CC398 isolates but differs from other MRSA clones. The spa-types were determined and assigned as previously described4 (Koreen et al., 2004).
DNA Microarray
All mecA/mecC-positive strains were characterized using microarray-based assay (S. aureus Genotyping, Identibac – Alere) allowing detection of virulence and resistance genes, SCCmec types, and assignment to clones and/or MLST clonal complexes. Protocols, target genes for virulence and resistance, databases, typing information, and interpretation management were available at the following internet address: https://alere-technologies.com/fileadmin/Media/Downloads/op/10620/Manuals/05_16_04_0001_V05_Manual_S.aureus_Genotyping_Kit_2_0.pdf.
Results
Epidemiological Data and Antibiotic Resistance of MRSA in Cats, Dogs, and Horses
A total of 130 consecutive non-duplicate clinical MRSA isolates were collected from 43 different French districts: 68 isolates from horses, 34 from cats, and 28 from dogs (Supplementary Table S1). Since 2012, the total number of MRSA collected has significantly decreased in all three animal species (Figure 1), despite the fact that the total number of coagulase-positive isolates collected (susceptible and resistant to methicillin) followed either a steady or increasing trend.
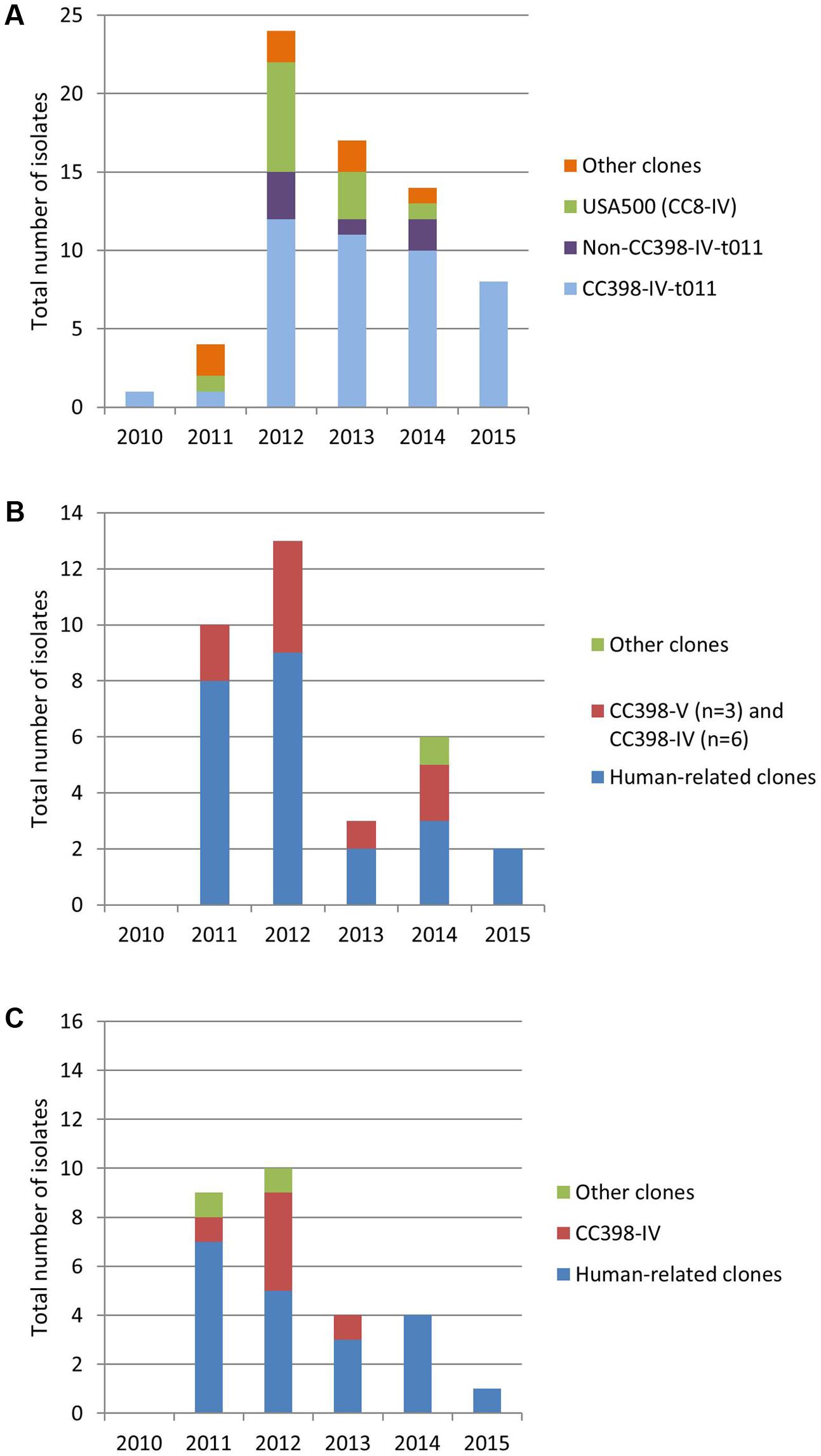
FIGURE 1. Distribution of clones per year in horses (A), cats (B), and dogs (C).
None of the MRSA infected animals presented any epidemiological links since they all originated from different cities, departments or regions, and none of the cats or dogs came from a kennel. MRSA were mostly isolated from skin and soft tissue infections (SSTI) [horse, n = 32 (32/68, 47.1%); dog, n = 11 (11/28, 39.3%); cat, n = 9 (9/34, 26.5%)], while MRSA from reproductive tract infections were also highly prevalent in horses (22/68, 32.4%) (Supplementary Table S2). Isolates globally presented high levels of resistance to several antibiotics, particularly to aminoglycosides (50.0% to gentamicin, 70.0% to kanamycin, 67.7% to tobramycin), tetracyclines (60.0%) and enrofloxacin (48.50%) (Table 1). Major differences were however observed between the CC398 and human-related isolates, with resistances to tetracyclines, phenicols, and aminoglycosides being more prevalent in CC398 isolates, while resistances to macrolides-lincosamides, fusidic acid, and enrofloxacin were more prevalent in human-related isolates (Table 1).
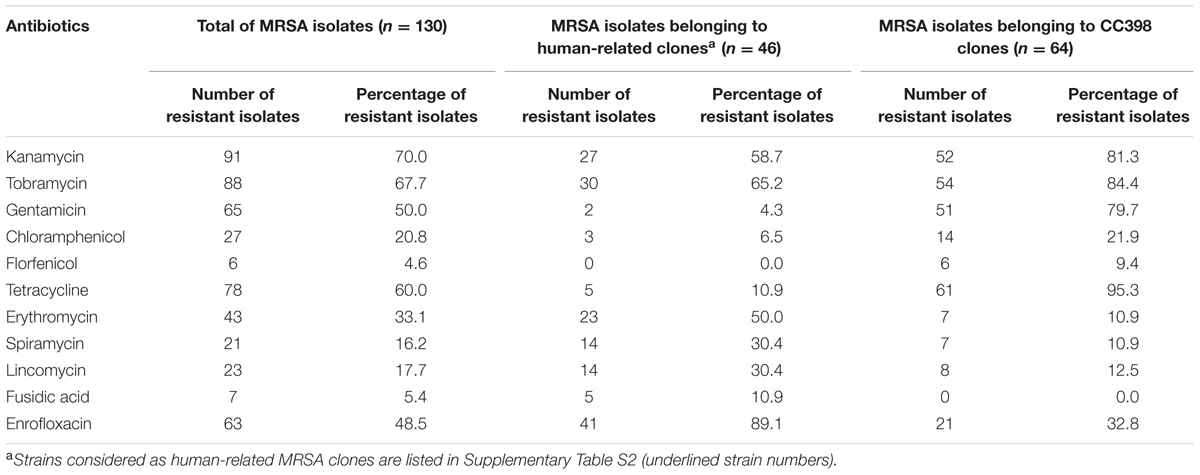
TABLE 1. Resistance to non-beta-lactam antibiotics among MRSA isolated from companion animals.
Distribution of MRSA Clones in Horses
The most prevalent MRSA clone in horses was the CC398 MRSA clone presenting a SCCmec type IV (hereafter named CC398-IV), represented in forty-nine MRSA isolates (49/68, 72.1%) (Figures 1, 2 and Supplementary Table S2). The spa-type t011 was identified in the majority of the isolates (n = 42/49, 85.7%), while t899 (n = 2), t779 (n = 2), t108 (n = 1), t1451 (n = 1) and one non-typeable isolate were also detected. Conversely to cats and dogs, only two isolates displayed enterotoxin genes [i.e., sek/seq (n = 1) and sea (n = 1)]. The IEC was present in 12.2% (n = 6, including 5 type B and 1 type D) of the 49 CC398 isolates. Tetracycline resistance, which has been strongly associated with CC398 strains from livestock so far (Price et al., 2012), was systematically detected and the tet(M) gene identified in 48 of the 49 isolates. The aacA-aphD gene conferring resistance to gentamicin, kanamycin/amikacin, and tobramycin was also frequently identified (46/49, 93.9%) and correlated with phenotypic resistance (Supplementary Table S2). The other resistance genes were scarce, with the sporadic occurrence of genes conferring resistances to macrolides/lincosamides/streptogramins B [erm(A), n = 3], streptogramin A [vga(A), n = 2], trimethoprim (dfrS1, n = 2), fosfomycin (fosB, n = 1), chloramphenicol (cat, n = 1), tetracyclines [tet(K), n = 2], or quaternary ammonium compounds (qacC, n = 1).
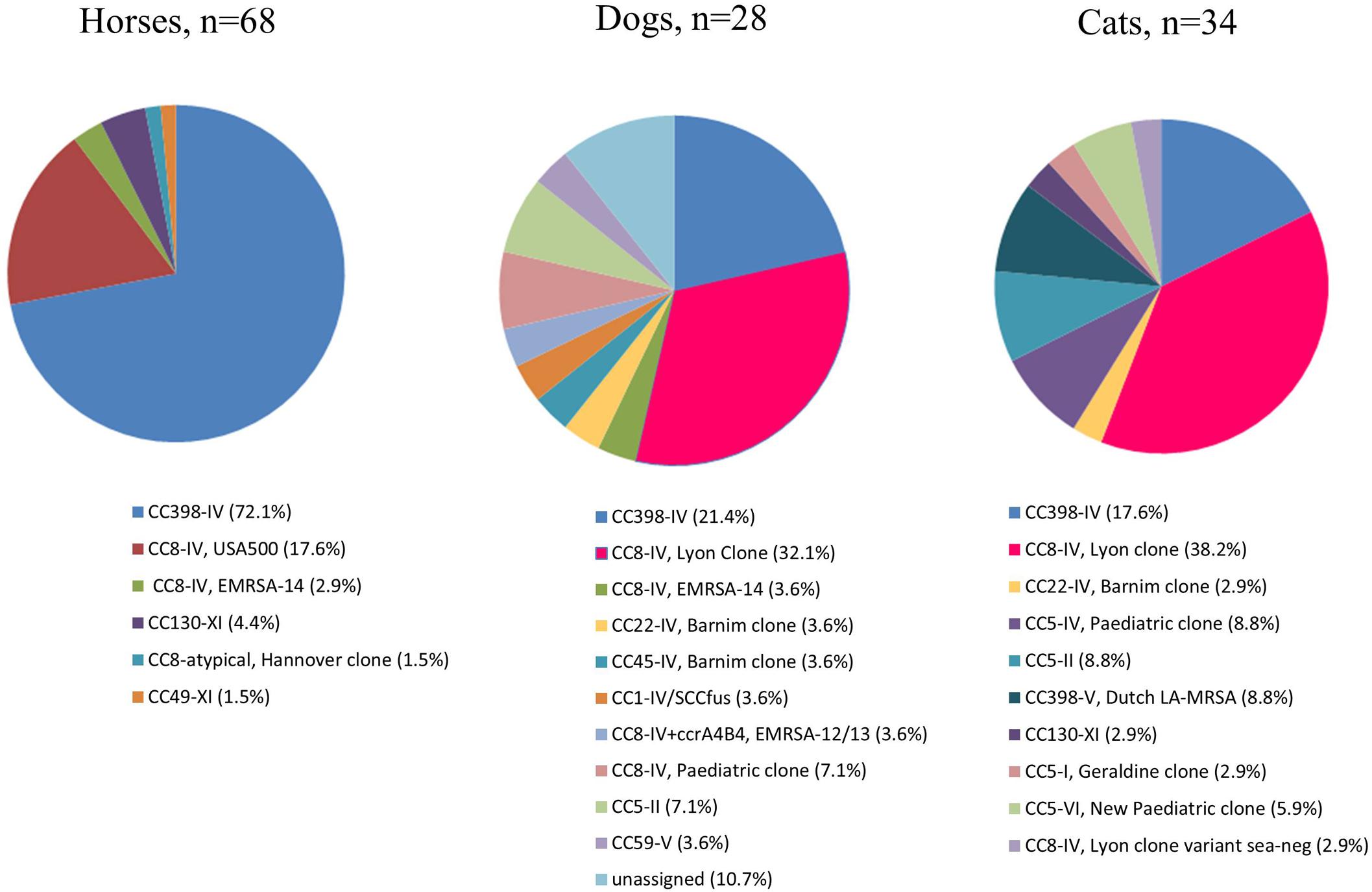
FIGURE 2. Schematic representation of all clones associated to horses, cats, and dogs. In the dog section the “unassigned” clones corresponded to strains that could not be assigned to a specific clone by DNA array. These three strains corresponded to CC-spa types CC30-t012, CC5-t450, and CC8-t2054 (also see Supplementary Table S2). For all strains, associated spa-types can be found in Supplementary Table S2.
Twelve MRSA (17.6%) isolates clustered with the CC8-IV USA500 clone [spa-type t394 (n = 8) and t1952 (n = 1)]. All isolates presented the seb, sek, and seq enterotoxin genes, and the tet(M), aacA/aphD, cat, dfr(S1), and fos(B) resistance genes were detected in all t394 isolates (Supplementary Table S2). None of them presented the IEC cluster that is classically related to human origin/adaptation.
Among the seven isolates belonging neither to the CC398-IV nor the CC8-IV USA500 clone, three were assigned to the CC8-IV EMRSA-14 (n = 2) or to the CC8 Hannover EMRSA (n = 1) clones. This CC8 Hannover clone belonged to the ST249-t009 and presented an atypical SCCmec element comparable to those described by Monecke et al. (2011). All three strains presented multiple resistance genes, including to tetracyclines (Supplementary Table S2). The last four isolates harbored the mecC gene and have already been reported in the frame of another study (Haenni et al., 2015). They belonged to the CC130-XI-t6220, CC130-XI-t1736, CC130-XI-t11050, and CC49-XI-t208 clones (Supplementary Table S2). These four mecC-positive isolates were the only equine isolates that were not resistant to tetracyclines and presented no other resistance genes.
Distribution of MRSA Clones in Cats
Most isolates belong to the prototypic CC8-IV Lyon clone (13/34, 38.2%) or its sea-negative variant (n = 1). The majority have spa-type t008 (n = 8) (Figure 1 and Supplementary Table S2). In addition to these 14 isolates, another 10 isolates belong to human-related clones: CC5-IV Pediatric (n = 3) and New Pediatric (n = 2) clones, CC5-II (n = 3), CC22-IV Barnim (n = 1), CC5-I Geraldine (n = 1) clones. Among the latter, the IEC (marker of human adaptation) was detected in a vast majority of the isolates (n = 17; 17/24, 70.8%). The IEC type A (sea, sak, scn, chp) was identified in one isolate, the IEC type B (lacking sea) in four isolates, and the IEC type D (lacking chp) in 12/13 isolates from the Lyon clone (van Wamel et al., 2006). Among the human-related clones, only one isolate was resistant to tetracyclines without tet(M) gene.
On the contrary, the tet(M) gene was present in all nine isolates belonging to the CC398 MRSA clone, among which 6 harbored SCCmec type IV (CC398-IV; including four isolates with spa type t011) and three of the SCCmec type V (CC398-V). The IEC cluster was only found in three isolates [type B (n = 1) or type D (n = 2)]. All CC398-V clones presented at least the tet(M) and fexA resistance genes, whereas the CC398-IV clone carried the tet(M) and aacA-aphD genes. The co-occurrence of the tet(M) and tet(K) genes was identified in two spa-type t034 CC398-V. This association, recently shown to increase the fitness of CC398-V strains in the presence of tetracyclines (Larsen et al., 2016), was also identified here in a t1451 CC398-IV isolate originating from a horse (Supplementary Table S2). Finally, one isolate, belonging to the CC130-XI t843 clone, harbored the mecC gene with no associated resistance or enterotoxin genes.
Distribution of MRSA Clones in Dogs
As for cats, nine isolates belonged to the CC8-IV Lyon clone (32.1%) or to other sporadically occurring human-related clones (n = 10) (Figure 1 and Supplementary Table S2). A total of 16 out of the 28 isolates displayed the IEC cluster [type B, n = 6; type D, n = 10 (including the nine Lyon clone isolates)]. Only one isolate did not present any associated enterotoxin gene. Resistance phenotypes to macrolides (10/28, 35.7%), aminoglycosides (15/28, 53.6%), and fosfomycin (18/28, 64.3%) were frequently encountered, whereas only one isolate was resistant to tetracyclines in the absence of the tet(M) gene. The CC398-IV clone was identified in six tetracycline-resistant isolates (21.4%), among which two presented the IEC cluster (type B and type D). The last three isolates belonged to the CC-spa types CC30-t012, CC5-t450, and CC8-t2054 but could not be assigned to any specific clone by the DNA array. Finally, no mecC gene was detected in dogs.
Discussion
This study describes the largest collection of MRSA isolates ever collected in France from companion animals (horses, cats, and dogs). It can be compared with the MRSA collection from these three animal species published by Vincze et al. (2014) in Germany, and also to the data very recently published in a specific study focused on MRSA in horses by Guerin et al. (2017) in France.
In our study, only one MRSA was isolated from horses in 2010, likely due to a poor representation of horses, cats and dogs in the Resapath network at the time (the total number of coagulase-positive staphylococci reported in 2010 was respectively 5 in horses, 11 in cats, 88 in dogs). Since 2011, this gap has been progressively filled in and the obvious decrease in the total number of MRSA collected over years (Figure 1) likely reflects a true decrease in the prevalence of MRSA in horses, cats, and dogs in France. A major reason for this decrease may well be the recent global reduction in the use of antibiotics in the veterinary field in France, in line with the 5-year national action plan EcoAntibio of the Ministry of Agriculture, Agri-Food and Forestry. Indeed, a decrease (from 19.4% to 28.1%) was observed in the global exposure to antibiotics of respectively cats/dogs and horses5. Such a valuable trend will be thoroughly monitored in the near future.
Success in Horses of the CC398-IV MRSA Clone Belonging to the spa-Type t011
The molecular characterization of the horse collected isolates highlights that MRSA identified in these animals are independent to the human MRSA epidemiology. Two major animal-associated clones were identified in France: the CC8-IV USA500 and the CC398-IV-t011 clones. The CC8-IV USA500, which is the predominant clone infecting horses in North America, also recently described as an environmental contaminant in an equine veterinary center (Weese et al., 2005a; van Balen et al., 2014), was detected in France between 2011 and 2013, with a last occurrence in early 2014. This suggests that this clone, uncommon in the human population in France and worldwide, is prone to colonize and/or infect horses worldwide. Yet CC8-IV USA500 seems to have been totally replaced by the horse-specific CC398-IV belonging to the spa-type t011 in France (Figure 1), confirming its international spread and high fitness/adaptation to horses. Between 2013 and 2015, only 7/39 (17.9%) isolates did not belong to the CC398 clone. Of note, four out of these seven isolates displayed the mecC gene. Whether horses may be prone to disseminate mecC genes or not remains questionable. Both in terms of management by veterinarians and with regards to the global burden antimicrobial resistance, the replacement of CC8-IV USA500 by the CC398-IV-t011 clone may be considered good news since CC398 were co-resistant to aminoglycosides and tetracyclines only, whereas USA500 additionally displayed resistances to macrolides-lincosamides, chloramphenicol, sulfonamides, and fosfomycin. Even though these antibiotic molecules are not all licensed for horse treating, any decrease in antibiotic resistance burden is to be noticed since it may also decrease the risk of co-selection of resistant isolates in other animal species or in humans. This is of course only on a resistance perspective, and the differences in virulence capacities (such as biofilm formation or the presence of adhesion factors) will also have to be considered.
Considering the nosocomial and zoonotic potential of MRSA isolated from horses, equine veterinarians should pay specific attention to both antibiotic treatments and hygiene measures, to limit MRSA selection and transmission. The rate of MRSA carriage in healthy horses, as well as the potential human-to-animal or animal-to-human transmission in both veterinary clinics and in equestrian centers was studied on certain occasions and certainly deserve further investigation (van Duijkeren et al., 2010; Cuny et al., 2016; Koop, 2016).
Human-Related MRSA Clones in Cats and Dogs
Our data also underline that MRSA isolated from cats and dogs mirror the epidemiology of hospital-acquired human MRSA (Loeffler and Lloyd, 2010). The CC8-IV Lyon clone was the most frequently identified MRSA clone in both animal species, followed by the occurrence of numerous sporadic lineages. This is in accordance with a similar study previously conducted in France on a smaller number of isolates (Haenni et al., 2012). The ST22 Barnim clone is again rare in France despite its high prevalence in animals in surrounding countries (Strommenger et al., 2006; Grundmann et al., 2010; Harrison et al., 2014; Vincze et al., 2014; Couto et al., 2016; Wipf and Perreten, 2016). Its rare occurrence is most likely related to its low prevalence in French hospitals and in the community. The CC398 clone was detected in 21.4% of all dogs’ and 26.5% of all cats’ isolates, which is more frequent that what has been reported in Germany and much more than what has been recently observed in France (2/23, 8.7%) or in other European countries (Haenni et al., 2012; Vincze et al., 2014; Wipf and Perreten, 2016). Since no links could be inferred between animals sampled and pig farming or contact with livestock, the hypothesis of a progressive replacement of existing human MRSA clones in cats and dogs by the CC398 cannot be excluded, as observed in horses,. Data collected are too limited to confirm such an hypothesis and this will deserve to be followed-up, yet CC398 clones disseminated among humans have recently been exemplified in Denmark, not all to be attributed to contact with livestock (Larsen et al., 2015). Interestingly, all CC398 isolates presented the tet(M) gene, whereas all non-CC398 were devoid of the gene. Other tet genes being rare (Supplementary Table S2), tetracycline susceptibility in MRSA from cats and dogs may be used as a marker for human-related isolates, mirroring the fact that tetracycline-resistance [or the tet(M) presence] can be used as a marker for MRSA CC398 in humans living in high pig-farming areas (Lozano et al., 2012).
Conclusion
We show here a divergent epidemiology between cats/dogs and horses, with the first two species being largely colonized by human-related isolates mirroring the human MRSA epidemiology. These strains produce enterotoxins more frequently and display a higher prevalence of the IEC cluster than the animal-associated MRSA isolates colonizing horses. Considering the proximity of humans with their pets and the possibility of human re-infection, prevalence of MRSA in cats and dogs is undoubtedly a public health issue that deserves to be monitored. Conversely, horses are mostly infected by animal-associated isolates that differ from the traditional ST398 LA-MRSA with the particular prevalence of the CC398-IV MRSA clone belonging to the spa-type t011. Cats and dogs are also infected by CC398 MRSA clones, and special attention will have to be paid to the IEC-positive CC398 MRSA isolates, whose prevalence was substantial in our study (11/64, 17.2%). This rate is particularly high compared to the one detected in LA-MRSA from in MRSA from horses in Germany (∼10%) (Cuny et al., 2015). Of note, no IEC-positive CC398 MRSA had been detected from pigs’ isolates in the same study. Re-adaptation to a human host, due to closer contacts between companion animals and humans compared to pigs and farmers, may favor virulence and animal-independent dissemination of such MRSA isolates in humans (Stegger et al., 2013). Taken together, this set of data promote a regular surveillance of MRSA in companion animals in the coming years to assess whether the CC398 would replace human-related MRSA clones in cats and dogs and whether those clones will be able to re-adapt to the human host.
Author Contributions
MH, J-YM, MB, and FL designed the experiments. PC, VM, CD-C, and MB did the experiments. MH and FL analyzed the data. MH and FL drafted the manuscript. J-YM and MB actively contributed to the manuscript’s writing. All authors approved the final version of this manuscript.
Funding
This work was supported by the French Agency for Food, Environmental and Occupational Health Safety (ANSES), Santé Publique France and the Institut National de la Santé Et de la Recherche Médicale (INSERM).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors gratefully thank all peripheral laboratories of the Resapath network involved in the study.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02493/full#supplementary-material
Footnotes
- ^ https://www.resapath.anses.fr/
- ^ http://agriculture.gouv.fr/ecoantibio
- ^ www.sfm-microbiologie.org/
- ^ http://www.spaserver.ridom.de/
- ^ https://www.anses.fr/en/content/monitoring-sales-veterinary-antimicrobials
References
Abdelbary, M. M., Wittenberg, A., Cuny, C., Layer, F., Kurt, K., Wieler, L. H., et al. (2014). Phylogenetic analysis of Staphylococcus aureus CC398 reveals a sub-lineage epidemiologically associated with infections in horses. PLOS ONE 9:e88083. doi: 10.1371/journal.pone.0088083
Aires-de-Sousa, M. (2017). Methicillin-resistant Staphylococcus aureus among animals: current overview. Clin. Microbiol. Infect. 23, 373–380. doi: 10.1016/j.cmi.2016.11.002
Ballhausen, B., Kriegeskorte, A., Van Alen, S., Jung, P., Kock, R., Peters, G., et al. (2017). The pathogenicity and host adaptation of livestock-associated MRSA CC398. Vet. Microbiol. 200, 39–45. doi: 10.1016/j.vetmic.2016.05.006
Bergstrom, K., Aspan, A., Landen, A., Johnston, C., and Gronlund-Andersson, U. (2012). The first nosocomial outbreak of methicillin-resistant Staphylococcus aureus in horses in Sweden. Acta Vet. Scand. 54:11. doi: 10.1186/1751-0147-54-11
Bierowiec, K., Ploneczka-Janeczko, K., and Rypula, K. (2016). Is the colonisation of Staphylococcus aureus in pets associated with their close contact with owners? PLOS ONE 11:e0156052. doi: 10.1371/journal.pone.0156052
Blaiotta, G., Fusco, V., Ercolini, D., Pepe, O., and Coppola, S. (2010). Diversity of Staphylococcus species strains based on partial kat (catalase) gene sequences and design of a PCR-restriction fragment length polymorphism assay for identification and differentiation of coagulase-positive species (S. aureus, S. delphini, S. hyicus, S. intermedius, S. pseudintermedius, and S. schleiferi subsp. coagulans). J. Clin. Microbiol. 48, 192–201. doi: 10.1128/JCM.00542-09
Couto, N., Monchique, C., Belas, A., Marques, C., Gama, L. T., and Pomba, C. (2016). Trends and molecular mechanisms of antimicrobial resistance in clinical staphylococci isolated from companion animals over a 16 year period. J. Antimicrob. Chemother. 71, 1479–1487. doi: 10.1093/jac/dkw029
Cuny, C., Abdelbary, M., Layer, F., Werner, G., and Witte, W. (2015). Prevalence of the immune evasion gene cluster in Staphylococcus aureus CC398. Vet. Microbiol. 177, 219–223. doi: 10.1016/j.vetmic.2015.02.031
Cuny, C., Abdelbary, M. M. H., Kock, R., Layer, F., Scheidemann, W., Werner, G., et al. (2016). Methicillin-resistant Staphylococcus aureus from infections in horses in Germany are frequent colonizers of veterinarians but rare among MRSA from infections in humans. One Health 2, 11–17. doi: 10.1016/j.onehlt.2015.11.004
Cuny, C., Strommenger, B., Witte, W., and Stanek, C. (2008). Clusters of infections in horses with MRSA ST1, ST254, and ST398 in a veterinary hospital. Microb. Drug Resist. 14, 307–310. doi: 10.1089/mdr.2008.0845
Garcia-Alvarez, L., Holden, M. T., Lindsay, H., Webb, C. R., Brown, D. F., Curran, M. D., et al. (2011). Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: a descriptive study. Lancet Infect. Dis. 11, 595–603. doi: 10.1016/S1473-3099(11)70126-8
Grundmann, H., Aanensen, D. M., Van Den Wijngaard, C. C., Spratt, B. G., Harmsen, D., Friedrich, A. W., et al. (2010). Geographic distribution of Staphylococcus aureus causing invasive infections in Europe: a molecular-epidemiological analysis. PLOS Med. 7:e1000215. doi: 10.1371/journal.pmed.1000215
Guerin, F., Fines-Guyon, M., Meignen, P., Delente, G., Fondrinier, C., Bourdon, N., et al. (2017). Nationwide molecular epidemiology of methicillin-resistant Staphylococcus aureus responsible for horse infections in France. BMC Microbiol. 17:104. doi: 10.1186/s12866-016-0924-z
Haenni, M., Chatre, P., Dupieux, C., Metayer, V., Maillard, K., Bes, M., et al. (2015). mecC-positive MRSA in horses. J. Antimicrob. Chemother. 70, 3401–3402. doi: 10.1093/jac/dkv278
Haenni, M., Saras, E., Chatre, P., Medaille, C., Bes, M., Madec, J. Y., et al. (2012). A USA300 variant and other human-related methicillin-resistant Staphylococcus aureus strains infecting cats and dogs in France. J. Antimicrob. Chemother. 67, 326–329. doi: 10.1093/jac/dkr499
Harrison, E. M., Weinert, L. A., Holden, M. T., Welch, J. J., Wilson, K., Morgan, F. J., et al. (2014). A shared population of epidemic methicillin-resistant Staphylococcus aureus 15 circulates in humans and companion animals. mBio 5:e00985-13. doi: 10.1128/mBio.00985-13
Hau, S. J., Sun, J., Davies, P. R., Frana, T. S., and Nicholson, T. L. (2015). Comparative prevalence of Immune Evasion Complex genes associated with beta-hemolysin converting bacteriophages in MRSA ST5 isolates from swine, swine facilities, humans with swine contact, and humans with no swine contact. PLOS ONE 10:e0142832. doi: 10.1371/journal.pone.0142832
Koop, G. (2016). MRSA transmission between horses and vets: who’s doing the infecting? Vet. Rec. 178, 471–472. doi: 10.1136/vr.i2531
Koreen, L., Ramaswamy, S. V., Graviss, E. A., Naidich, S., Musser, J. M., and Kreiswirth, B. N. (2004). spa typing method for discriminating among Staphylococcus aureus isolates: implications for use of a single marker to detect genetic micro- and macrovariation. J. Clin. Microbiol. 42, 792–799. doi: 10.1128/JCM.42.2.792-799.2004
Larsen, J., Clasen, J., Hansen, J. E., Paulander, W., Petersen, A., Larsen, A. R., et al. (2016). Copresence of tet(K) and tet(M) in livestock-associated methicillin-resistant Staphylococcus aureus clonal complex 398 is associated with increased fitness during exposure to sublethal concentrations of tetracycline. Antimicrob. Agents Chemother. 60, 4401–4403. doi: 10.1128/AAC.00426-16
Larsen, J., Petersen, A., Sorum, M., Stegger, M., Van Alphen, L., Valentiner-Branth, P., et al. (2015). Meticillin-resistant Staphylococcus aureus CC398 is an increasing cause of disease in people with no livestock contact in Denmark, 1999 to 2011. Euro Surveill. 20:30021. doi: 10.2807/1560-7917.ES.2015.20.37.30021
Loeffler, A., and Lloyd, D. H. (2010). Companion animals: a reservoir for methicillin-resistant Staphylococcus aureus in the community? Epidemiol. Infect. 138, 595–605. doi: 10.1017/S0950268809991476
Lozano, C., Rezusta, A., Gomez, P., Gomez-Sanz, E., Baez, N., Martin-Saco, G., et al. (2012). High prevalence of spa types associated with the clonal lineage CC398 among tetracycline-resistant methicillin-resistant Staphylococcus aureus strains in a Spanish hospital. J. Antimicrob. Chemother. 67, 330–334. doi: 10.1093/jac/dkr497
Maes, N., Magdalena, J., Rottiers, S., De Gheldre, Y., and Struelens, M. J. (2002). Evaluation of a triplex PCR assay to discriminate Staphylococcus aureus from coagulase-negative staphylococci and determine methicillin resistance from blood cultures. J. Clin. Microbiol. 40, 1514–1517. doi: 10.1128/JCM.40.4.1514-1517.2002
Monecke, S., Coombs, G., Shore, A. C., Coleman, D. C., Akpaka, P., Borg, M., et al. (2011). A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLOS ONE 6:e17936. doi: 10.1371/journal.pone.0017936
Moodley, A., Stegger, M., Bagcigil, A. F., Baptiste, K. E., Loeffler, A., Lloyd, D. H., et al. (2006). spa typing of methicillin-resistant Staphylococcus aureus isolated from domestic animals and veterinary staff in the UK and Ireland. J. Antimicrob. Chemother. 58, 1118–1123. doi: 10.1093/jac/dkl394
Mutters, N. T., Bieber, C. P., Hauck, C., Reiner, G., Malek, V., and Frank, U. (2016). Comparison of livestock-associated and health care-associated MRSA-genes, virulence, and resistance. Diagn. Microbiol. Infect. Dis. 86, 417–421. doi: 10.1016/j.diagmicrobio.2016.08.016
Price, L. B., Stegger, M., Hasman, H., Aziz, M., Larsen, J., Andersen, P. S., et al. (2012). Staphylococcus aureus CC398: host adaptation and emergence of methicillin resistance in livestock. mBio 4:e00520-12. doi: 10.1128/mBio.00520-12
Stegger, M., Lindsay, J. A., Moodley, A., Skov, R., Broens, E. M., and Guardabassi, L. (2011). Rapid PCR detection of Staphylococcus aureus clonal complex 398 by targeting the restriction-modification system carrying sau1-hsdS1. J. Clin. Microbiol. 49, 732–734. doi: 10.1128/JCM.01970-10
Stegger, M., Liu, C. M., Larsen, J., Soldanova, K., Aziz, M., Contente-Cuomo, T., et al. (2013). Rapid differentiation between livestock-associated and livestock-independent Staphylococcus aureus CC398 clades. PLOS ONE 8:e79645. doi: 10.1371/journal.pone.0079645
Strommenger, B., Kehrenberg, C., Kettlitz, C., Cuny, C., Verspohl, J., Witte, W., et al. (2006). Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and their relationship to human isolates. J. Antimicrob. Chemother. 57, 461–465. doi: 10.1093/jac/dki471
van Balen, J., Mowery, J., Piraino-Sandoval, M., Nava-Hoet, R. C., Kohn, C., and Hoet, A. E. (2014). Molecular epidemiology of environmental MRSA at an equine teaching hospital: introduction, circulation and maintenance. Vet. Res. 45:31. doi: 10.1186/1297-9716-45-31
Van den Eede, A., Martens, A., Flore, K., Denis, O., Gasthuys, F., Haesebrouck, F., et al. (2013). MRSA carriage in the equine community: an investigation of horse-caretaker couples. Vet. Microbiol. 163, 313–318. doi: 10.1016/j.vetmic.2012.12.038
Van den Eede, A., Martens, A., Lipinska, U., Struelens, M., Deplano, A., Denis, O., et al. (2009). High occurrence of methicillin-resistant Staphylococcus aureus ST398 in equine nasal samples. Vet. Microbiol. 133, 138–144. doi: 10.1016/j.vetmic.2008.06.021
van Duijkeren, E., Moleman, M., Sloet Van Oldruitenborgh-Oosterbaan, M. M., Multem, J., Troelstra, A., Fluit, A. C., et al. (2010). Methicillin-resistant Staphylococcus aureus in horses and horse personnel: an investigation of several outbreaks. Vet. Microbiol. 141, 96–102. doi: 10.1016/j.vetmic.2009.08.009
van Wamel, W. J., Rooijakkers, S. H., Ruyken, M., Van Kessel, K. P., and Van Strijp, J. A. (2006). The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on beta-hemolysin-converting bacteriophages. J. Bacteriol. 188, 1310–1315. doi: 10.1128/JB.188.4.1310-1315.2006
Vincze, S., Stamm, I., Kopp, P. A., Hermes, J., Adlhoch, C., Semmler, T., et al. (2014). Alarming proportions of methicillin-resistant Staphylococcus aureus (MRSA) in wound samples from companion animals, Germany 2010–2012. PLOS ONE 9:e85656. doi: 10.1371/journal.pone.0085656
Walther, B., Monecke, S., Ruscher, C., Friedrich, A. W., Ehricht, R., Slickers, P., et al. (2009). Comparative molecular analysis substantiates zoonotic potential of equine methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 47, 704–710. doi: 10.1128/JCM.01626-08
Weese, J. S., Archambault, M., Willey, B. M., Hearn, P., Kreiswirth, B. N., Said-Salim, B., et al. (2005a). Methicillin-resistant Staphylococcus aureus in horses and horse personnel, 2000-2002. Emerg. Infect. Dis. 11, 430–435.
Weese, J. S., Rousseau, J., Traub-Dargatz, J. L., Willey, B. M., Mcgeer, A. J., and Low, D. E. (2005b). Community-associated methicillin-resistant Staphylococcus aureus in horses and humans who work with horses. J. Am. Vet. Med. Assoc. 226, 580–583.
Weese, J. S., and van Duijkeren, E. (2010). Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary medicine. Vet. Microbiol. 140, 418–429. doi: 10.1016/j.vetmic.2009.01.039
Keywords: MRSA, mecA, clone, horses, cats, dogs, veterinary microbiology
Citation: Haenni M, Châtre P, Dupieux-Chabert C, Métayer V, Bes M, Madec J-Y and Laurent F (2017) Molecular Epidemiology of Methicillin-Resistant Staphylococcus aureus in Horses, Cats, and Dogs Over a 5-Year Period in France. Front. Microbiol. 8:2493. doi: 10.3389/fmicb.2017.02493
Received: 25 August 2017; Accepted: 30 November 2017;
Published: 13 December 2017.
Edited by:
Miklos Fuzi, Semmelweis University, HungaryReviewed by:
Annalisa Pantosti, Istituto Superiore di Sanità, ItalyAnna Shore, Trinity College, Dublin, Ireland
Josep M. Sierra, University of Barcelona, Spain
Copyright © 2017 Haenni, Châtre, Dupieux-Chabert, Métayer, Bes, Madec and Laurent. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marisa Haenni, bWFyaXNhLmhhZW5uaUBhbnNlcy5mcg==