 María E. Pachón-Ibáñez1*
María E. Pachón-Ibáñez1* Gema Labrador-Herrera1Tania Cebrero-Cangueiro1Caridad Díaz2
Gema Labrador-Herrera1Tania Cebrero-Cangueiro1Caridad Díaz2 Younes Smani1
Younes Smani1 José P. del Palacio2
José P. del Palacio2 Jesús Rodríguez-Baño3,4Alvaro Pascual3,5Jerónimo Pachón1,4
Jesús Rodríguez-Baño3,4Alvaro Pascual3,5Jerónimo Pachón1,4 M. Carmen Conejo5
M. Carmen Conejo5- 1Clinical Unit of Infectious Diseases, Microbiology, and Preventive Medicine, Institute of Biomedicine of Seville, University Hospital Virgen del Rocío/CSIC/University of Seville, Seville, Spain
- 2Fundacion Centro de Excelencia en Investigación de Medicamentos Innovadores en Andalucía, MEDINA Foundation, Granada, Spain
- 3Clinical Unit of Infectious Diseases, Microbiology, and Preventive Medicine, Institute of Biomedicine of Seville, University Hospital Virgen de Macarena/CSIC/University of Seville, Seville, Spain
- 4Department of Medicine, University of Seville, Seville, Spain
- 5Department of Microbiology, University of Seville, Seville, Spain
Despite the relevance of carbapenemase-producing Klebsiella pneumoniae (CP-Kp) infections there are a scarce number of studies to evaluate in vivo the efficacy of combinations therapies. The bactericidal activity of colistin, rifampin, and its combination was studied (time–kill curves) against four clonally unrelated clinical isolates of CP-Kp, producing VIM-1, VIM-1 plus DHA-1(acquired AmpC β-lactamase), OXA-48 plus CTX-M-15 (extended spectrum β-lactamase) and KPC-3, respectively, with colistin MICs of 0.5, 64, 0.5, and 32 mg/L, respectively. The efficacies of antimicrobials in monotherapy and in combination were tested in a murine peritoneal sepsis model, against all the CP-Kp. Their efficacies were tested in the pneumonia model against the OXA-48 plus CTX-M-15 producers. The development of colistin-resistance was analyzed for the colistin-susceptible strains in vitro and in vivo. In vitro, colistin plus rifampin was synergistic against all the strains at 24 h. In vivo, compared to the controls, rifampin alone reduced tissue bacterial concentrations against VIM-1 and OXA-48 plus CTX-M-15 strains; CMS plus rifampin reduced tissue bacterial concentrations of these two CP-Kp and of the KPC-3 strain. Rifampin and the combination increased the survival against the KPC-3 strain; in the pneumonia model, the combination also improved the survival. No resistant mutants appeared with the combination. In conclusion, CMS plus rifampin had a low and heterogeneous efficacy in the treatment of severe peritoneal sepsis model due to CP-Kp producing different carbapenemases, increasing survival only against the KPC-3 strain. The combination showed efficacy in the less severe pneumonia model. The combination prevented in vitro and in vivo the development of colistin resistant mutants.
Introduction
Carbapenem-resistant Klebsiella pneumoniae strains are spreading worldwide, representing an urgent threat to public health, as stressed by the Center for Disease Control and Prevention (CDC) of United States, the European Centre for Disease Prevention and Control (ECDC) and the World Health Organization (WHO). The rapid spread, mostly in hospital settings, is transforming many common health care-associated complications into infections that are sometimes untreatable with the currently available antimicrobials (Nordmann et al., 2012).
The carbapenem resistance in K. pneumoniae is mainly due to the production of acquired carbapenemases (Falagas et al., 2014). The most important carbapenemases found in this species may belong to the Ambler classes A (mainly KPC), B (the most frequent are VIM and IMP) and D (OXA-48-like enzymes). Invasive infections by isolates producing VIM and KPC are associated with high death rates (Tzouvelekis et al., 2012). The information about infections caused by OXA-48-producers is scarce, mostly because of its difficult identification (Nordmann et al., 2011; Canton et al., 2012; Tzouvelekis et al., 2012). Nevertheless, OXA-48 is the most frequent carbapenemase produced by Enterobacteriaceae isolated in many European countries (Canton et al., 2012; Palacios-Baena et al., 2016; De Laveleye et al., 2017).
Although the hydrolysis spectrum of these enzymes may vary, they hydrolyze most beta-lactams, including carbapenems. Moreover, carbapenemase producers often show co-resistance to other antimicrobial agents, leaving very few treatment options, such as tigecycline, colistin, and some aminoglycosides (de Oliveira et al., 2015). So, in an attempt to improve the poor clinical efficacy of the available drugs, combination therapy is often used as definitive therapy for infections caused by carbapenemase-producing K. pneumoniae (CP-Kp). To date, recommendations are based on few retrospective clinical studies and in vitro studies (Tzouvelekis et al., 2012). In addition, a scarce number of in vivo studies have assessed the efficacy of antimicrobial combinations against CP-Kp (Tzouvelekis et al., 2012). Clinical studies have reported favorable outcomes for patients treated with combinations of colistin and a carbapenem, tigecycline, fosfomycin, or an aminoglycoside (Michalopoulos et al., 2010; Lee and Burgess, 2012; Munoz-Price et al., 2013; Daikos et al., 2014). However, little data exists on which combination therapy is superior.
Several studies have reported synergistic activity of colistin and rifampin against colistin-resistant and colistin-susceptible KPC-producing K. pneumoniae clinical strains, using the checkerboard method. Similarly, studies based on time-kill experiments detected synergy with this combination against colistin-resistant KPC or NDM producers clinical strains, but no synergistic effect against VIM producers. To our knowledge, there is no data regarding the efficacy of this combination against K. pneumoniae producing OXA-48, nor in vivo studies to validate the in vitro results previously mentioned (Elemam et al., 2010; Tascini et al., 2013; Nastro et al., 2014; Tangden et al., 2014). Thus, the aim of this study was to evaluate the in vitro and in vivo efficacy of CMS plus rifampin against CP-Kp clinical strains producing different carbapenemases.
Materials and Methods
Bacterial Strains, Beta-lactamase Characterization and Molecular Typing
Four genetically unrelated clinical isolates of CP-Kp were studied: Kp07, a VIM-1 ST 1603 clone producer (Miro et al., 2013); Kp21, which co-produced VIM-1 and the acquired AmpC type beta-lactamase DHA-1 ST 11 clone (Miro et al., 2013); Kp28, co-producing OXA-48 ST11 clone and the extended spectrum beta-lactamase (ESBL) CTX-M-15 (Oteo et al., 2015); and Kp29, co-producing KPC-3 ST512 clone with the broad spectrum beta-lactamases TEM-1 and SHV-11 (Lopez-Cerero et al., 2014), thereinafter VIM-1, VIM-1/DHA-1, OXA-48 plus CTX-M-15, and KPC-3 producers, respectively. Identification of these isolates was confirmed by a Microflex LT-MALDI Biotyper mass spectrometer (Bruker Daltonics GmbH, Bremen, Germany). The presence of carbapenemase genes, and genes coding for other beta-lactamases was confirmed by PCR and sequencing as described previously. The absence of genetic relation among the isolates was confirmed by PFGE analysis of chromosomal restriction fragments obtained after XbaI cleavage following the criteria of Tenover et al. (1995). Two of the strains, KPC-3 and VIM-1 (DHA-1) were multidrug-resistant while the other two were not. The antibiotic susceptibility profiles are included in the Supplementary Information.
Antimicrobials
For the in vitro assays, antimicrobials, colistin sulfate salt and rifampin, were used as standard laboratory powders (Sigma-Aldrich, Madrid, Spain). For in vivo experiments, clinical formulations were used: colistimethate sodium (CMS) (Genéricos Españoles S.A., Madrid, Spain) and rifampin (Sanofi-Aventis, Madrid, Spain).
In Vitro Studies
Antimicrobial Susceptibility Testing
MICs of antibiotics were determined by broth microdilution as recommended by the Clinical and Laboratory Standards Institute [CLSI] (2012), using Mueller Hinton broth II (MHB) (Becton Dickinson & Co., Sparks, MD, United States) and agar dilution method for fosfomycin. MIC results were interpreted according to the European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2016)1 breakpoints. Studies were performed in triplicate.
Time-Kill Curves
The concentrations of colistin used for susceptible strains corresponded to the MIC value obtained by microdilution, whereas the concentration used for resistant strains was 2 mg/L, the susceptibility breakpoint recommended by European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2016). Rifampin was used at a fixed concentration of 2 mg/L. Experiments were carried out with a starting inoculum of 1 × 106 cfu/mL and the antibiotics alone or in combination. Tubes were incubated at 37°C, with shaking, and samples were taken at 0, 1, 3, 6, and 24 h, serially diluted and plated using a spiral platter (Eddy Jet, IUL S.A., Barcelona, Spain). Bacterial colonies were counted using an automatic counter (Flash & Go, IUL S.A., Barcelona, Spain) (Pournaras et al., 2011; Souli et al., 2011). Experiments were performed three times on separate occasions. Synergy was defined as a decrease ≥ 2 log10 cfu/mL for the antimicrobial combination compared with the most active single agent. Bactericidal activities of single antibiotics or combination were defined as a decrease ≥ 3 log10 cfu/mL from the starting inoculum (Souli et al., 2009). Studies were performed in triplicate.
Animals
Immunocompetent C57BL/6 female mice weighing approximately 20 g (Production and Experimentation Animal Center, University of Seville, Seville, Spain) were used; they had a sanitary status of murine pathogen free and were assessed for genetic authenticity. Mice were housed in an individually ventilated cage system under specific pathogen-free conditions, and water and food supplied ad libitum. This study was carried out following the recommendations in the Guide for the Care and Use of Laboratory Animals (Souli et al., 2011). Experiments were approved by the Committee on the Ethics of Animal Experiments of the University Hospital Virgen Macarena, Seville, Spain (CI 1961). All procedures were performed under sodium thiopental (B. Braun Medical S.A., Spain) anesthesia, and all efforts were made to minimize suffering.
Pharmacokinetic/Pharmacodynamic Analysis
Serum antibiotic concentrations were determined in healthy mice after a single intraperitoneal (ip) administration of CMS (20 mg/kg) or rifampin (25 mg/kg). In sets of three anesthetized mice, blood samples from the periorbital plexus were obtained at different time points after the administration of CMS and rifampin. Blood samples were immediately centrifuged (4500 rpm, 15 min at 4°C), and serum samples stored at -80°C until the analysis. Both serum free and total antibiotic concentrations were measured using HPLC-tandem mass spectrometry (LC-MS/MS) (Waters et al., 2008; Gobin et al., 2010). Free fractions of the drugs were calculated as described previously (Waters et al., 2008; Gonzalez et al., 2013; Cheah et al., 2015).
Maximum concentration of drug in serum (Cmax), elimination half-life (t1/2), area under the concentration-time curve from 0 to 24 h (AUC0-24), free AUC0-24 (fAUC0-24), AUC0-24/MIC and fAUC0-24/MIC ratios were calculated using the PKSOLVER program (Zhang et al., 2010). The pharmacodynamic targets to assess the efficacy were fAUC0-24/MIC and AUC0-24/MIC for CMS and rifampin, respectively.
Experimental Models
Peritoneal Sepsis Model
A previously described murine peritoneal sepsis model was used (Parra Millan et al., 2016). Briefly, groups of un-anesthetized C57BL/6 mice were infected by ip injection of 0.5 mL of the Minimal Lethal Dose (MLD) of each strain, corresponding to (log10 cfu/mL): 8.97, 8.94 9.35, and 8.38 for VIM-1, VIM-1/DHA-1, OXA-48 plus CTX-M-15, and KPC-3 strains, respectively. Treatments were initiated 4 h post-inoculation. For each one of the strains, mice were randomly included into four different therapeutic groups: (1) controls (untreated), (2) CMS, 20 mg/kg/8 h/ip, (3) rifampin, 25 mg/kg/6 h/ip, and (4) CMS plus rifampin, using the same dosing schedule as in monotherapy. The sample size for the combination groups and CMS monotherapy for the colistin-susceptible strains (VIM-1 and OXA-48 plus CTX-M-15) was 15 mice; nevertheless, for the monotherapies in case of resistance to colistin (VIM-1/DHA-1 and KPC-3) and for rifampin, the sample size was 10 to accomplish the 3Rs (Replacement, Reduction and Refinement) rules for performing animal research. Afterward, mice were treated and monitored for 72 h. The antimicrobial dosages were based on the PK/PD data, and their proven efficacy, alone and in combination, in previous experimental murine models of infection (Pachon-Ibanez et al., 2010). Samples were extracted and processed immediately after the death of mice; the survivor mice were sacrificed (sodium thiopental) at the 72 h. Aseptic thoracotomies were carried out, and blood samples were obtained for qualitative blood cultures; results were expressed as positive (≥1 cfu present in the plate) or negative. Spleens were aseptically extracted, weighed, and homogenized in sterile saline (Stomacher 80; Tekmar Co., Cincinnati, OH, United States) before quantitative cultures (log10 cfu/g) in Columbia agar with 5% sheep blood plates.
Pneumonia Model
A previously characterized murine pneumonia model was used (Pachon-Ibanez et al., 2010; Docobo-Perez et al., 2012; Parra Millan et al., 2016) with the OXA-48 plus CTX-M-15 strain, as OXA-48 is the most prevalent carbapenemase in Spain nowadays (Palacios-Baena et al., 2016). Anesthetized mice were infected intratracheally, using 50 μl of a final inoculum of 8.18 log10 cfu/mL. Therapies were initiated 4 h post-infection and treatment groups and dosages were the same as in the peritoneal sepsis model. Mice were treated and monitored over 72 h. After death or sacrifice of the mice at 72 h, blood and lungs samples were aseptically obtained and processed as detailed above.
In Vitro and in Vivo Selection of Colistin-Resistant and Rifampin-Resistant Mutants
Up to 10 colonies, of colistin-susceptible strains exposed during 24 h to colistin at the MIC, alone or in combination with rifampin, in time-kill curves, and from controls (non-exposed to the antimicrobials), were sub-cultured twice in antimicrobial free medium and frozen at -80°C until MIC testing.
In the experimental murine pneumonia model, after processing the lungs as described above, the remaining homogenized tissue was vortexed and centrifuged, and the pellet was resuspended in 2 mL of sterile saline. All the volume was spread on agar and incubated for 48 h at 37°C. A maximum of ten colonies recovered from each lung were selected, sub cultured in antibiotic free medium twice and frozen at -80°C until MIC testing.
MICs determinations of colistin and rifampin were carried out in triplicate.
Statistical Analysis
Mortality and bacteremia rates are expressed as percentages and bacterial tissue concentrations (log10 cfu/g) as means ±SD. The two-tailed Fisher’s test, analysis of variance (ANOVA), and the Dunnet and Tukey post hoc tests were used. A P < 0.05 was considered significant. The SPSS v22.0 statistical package was used (SPSS Inc).
Results
In Vitro Results
Antimicrobial Susceptibility Testing
MICs of each antibiotic for the four clinical isolates are shown in Table 1. Two strains were resistant to colistin (VIM-1/DHA-1 and KPC-3 producers).
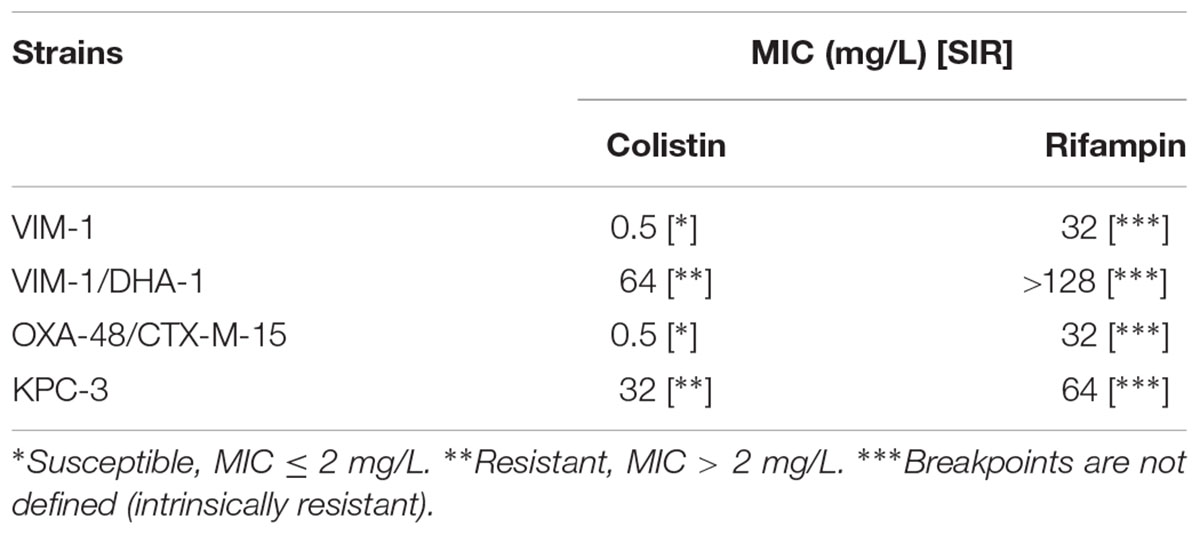
TABLE 1. MICs of colistin and rifampin for the four carbapenemase-producing K. pneumoniae clinical strains.
Time-Kill Curves
The results are shown in Figure 1. Colistin was bactericidal at 3 h against the VIM-1 producer, but an important regrowth at 24 h. In addition, colistin reduced bacterial concentration (2.5 log10 decrease in cfu/mL) of the OXA-48 plus CTX-M-15-producing isolate, again with an important regrowth at 24 h. Colistin did not display any bactericidal activity against the colistin-resistant isolates (VIM-1/DHA-1 and KPC-3 producers). Rifampin did not show bactericidal activity against any strain. The combination was synergistic against all the strains at 24 h, but achieving only a bacteriostatic effect against the VIM-1, VIM-1/DHA-1, and OXA-48 plus CTX-M-15 isolates; on the contrary, the combination was bactericidal against the KPC-3 isolate.
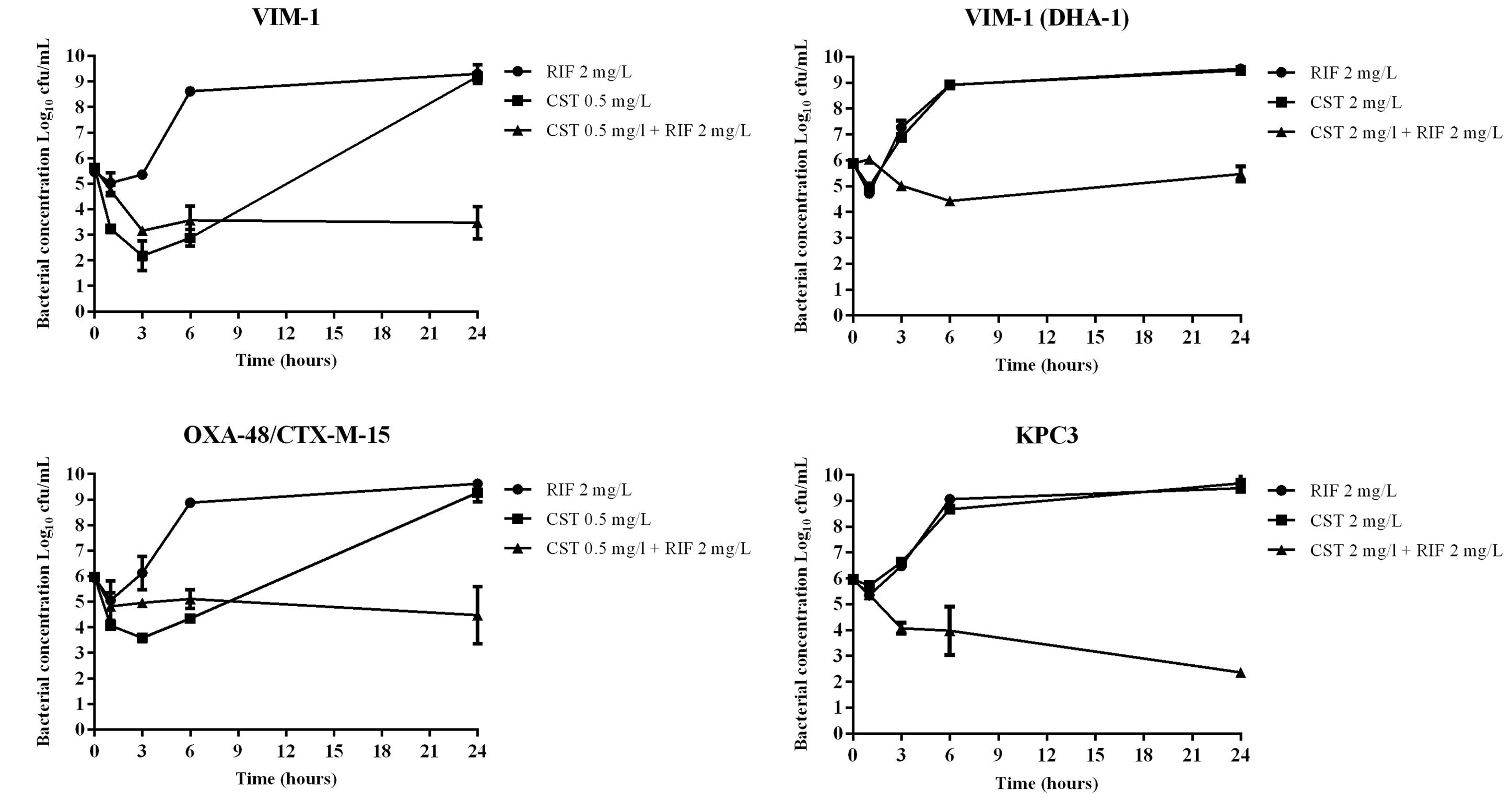
FIGURE 1. Time-kill curves for colistin (CST) and rifampin (RIF) alone and in combination against four clinical strains of carbapenemase producing strains (VIM-1, VIM-1/DHA-1, OXA-48/CTX-M-15 and KPC3). The CST concentration used for colistin-susceptible strains corresponded to the value of their MICs (colistin 0.5 mg/L for VIM-1 and OXA-48/CTX-M-15); for colistin-resistant isolates the concentration of CST was that corresponding to the susceptibility breakpoint recommended by EUCAST (colistin 2 mg/L for VIM-1/DHA-1 and KPC-3). The RIF concentration used for all strains was 2 mg/L. RIF, filled circles; CST, filled squares; combination of both antimicrobials, filled triangles.
Pharmacokinetics and Pharmacodynamics
Pharmacokinetic parameters of each antimicrobial are shown in Table 2. Pharmacodynamics profiles are shown in Tables 3, 4.
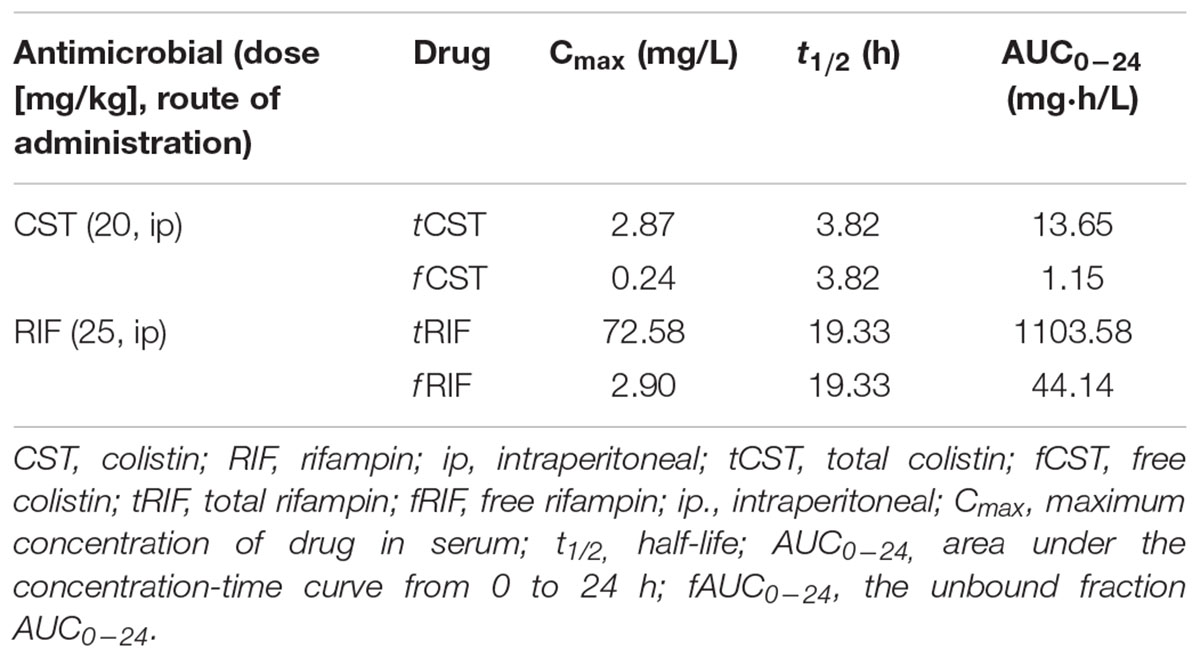
TABLE 2. Pharmacokinetic profiles of CMS and rifampin in mice serum.
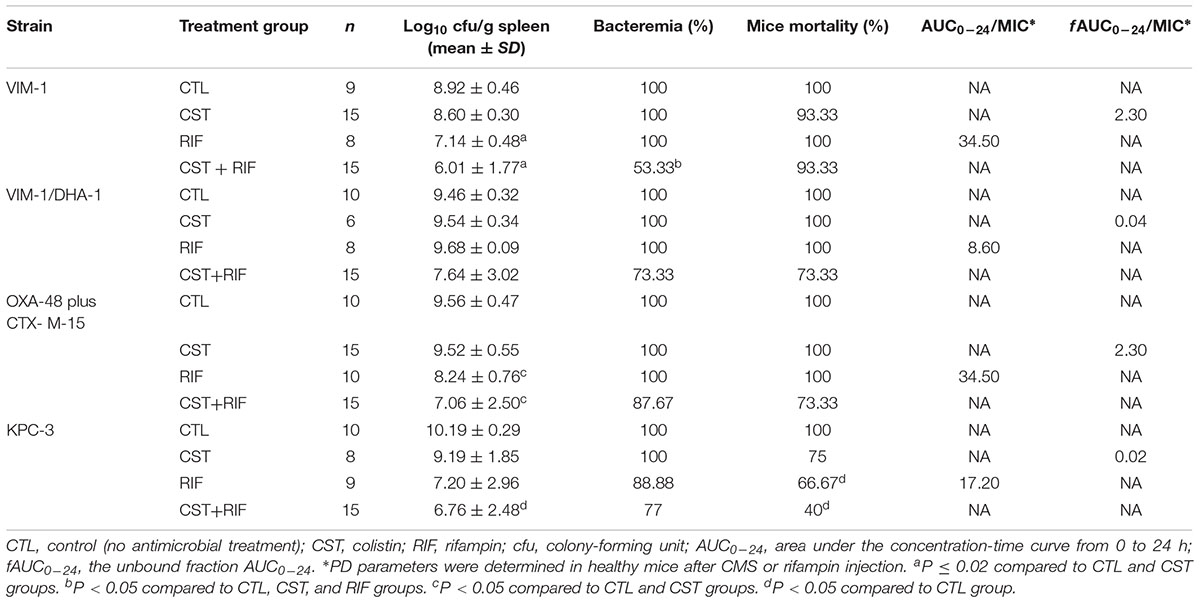
TABLE 3. In vivo efficacy and pharmacodynamics of CMS and rifampin, alone and in combination, for the experimental peritoneal sepsis model.

TABLE 4. In vivo efficacy and pharmacodynamics of CMS and rifampin, alone and in combination, for the experimental pneumonia model.
In Vivo Results: Peritoneal Sepsis Model
The efficacies of the antimicrobials are shown in Table 3. Mortality, bacterial clearance on spleen and bacteremia, are analyzed immediately after the death of mice or at the end of the experiment (72 h of treatment).
Mortality
Mortality in all control groups (non-treated) was 100% within the first 24 h post-infection. Rifampin and its combination with CMS reduced mortality in animals infected with the KPC-3 strain to a 66.67% and a 40%, respectively, but not CMS alone. There was no reduction of mortality with any treatment when the peritoneal sepsis was produced by the other strains.
Bacterial Clearance From Spleen
Rifampin alone improved significantly the clearance of bacteria from spleen (CFU/g of tissue) compared with the control and the CMS groups in mice infected with either of the two colistin-susceptible strains producing VIM-1 (7.14 ± 0.48 vs. 8.92 ± 0.46 and 7.14 ± 0.48 vs. 8.60 ± 0.30, respectively) or OXA-48 plus CTX-M-15 (8.24 ± 0.76 vs. 9.56 ± 0.47, and 8.24 ± 0.76 vs. 9.52 ± 0.55, respectively). CMS plus rifampin reduced the bacterial concentration compared with the controls for both colistin-susceptible strains, producing VIM-1 or OXA-48 plus CTX-M-15 and for the colistin-resistant strain producing KPC-3, (6.01 ± 1.77 vs. 8.92 ± 0.46, 7.06 ± 2.50 vs. 9.56 ± 0.47, and 6.76 ± 2.48 vs. 10.19 ± 0.29, respectively). The combination was also better than CMS alone for both colistin-susceptible strains (6.01 ± 1.77 vs. 8.60 ± 0.30, 7.06 ± 2.50 vs. 9.52 ± 0.55, respectively).
Bacteremia
CMS plus rifampin showed the best effect among all the treatment groups in sterilizing blood cultures of mice infected with any of the four strains. Nevertheless, this reduction was only significant in mice infected with the VIM-1 producer (53.33% vs. 100%).
Pneumonia Model Results
Table 4 summarizes the results for the pneumonia model.
Mortality
The severity of this model was lower, with a mortality of 40% at the end of the experiment (72 h) using the MLD in the control group. Only the combination therapy decreased significantly mortality compared with the control group (6.26% vs. 40%; P < 0.05).
Bacterial Clearance From Lungs
Rifampin alone and its combination with CMS decreased significantly bacterial lung concentration compared with the control (3.07 ± 1.63 vs. 8.05 ± 2.36, and 2.99 ± 1.36 vs. 8.05 ± 2.36, respectively).
Bacteremia
None of the antimicrobial treatments achieved a significant reduction of the bacteremia in comparison with the control group.
In Vitro and in Vivo Selection of Colistin-Resistant Mutants and Rifampin-Resistant Mutants
Colistin MICs of colonies recovered from susceptible strains (producing VIM-1 or OXA-48 plus CTX-M-15) after exposure at the MIC during 24 h in time-kill curves increased from 0.5 mg/L to > 8 mg/L. No mutants resistant to colistin were detected when these strains were exposed to the combination of colistin plus rifampin.
The MIC of rifampin for the colonies recovered from the lungs of mice challenged with OXA-48 plus CTX-M-15 producer and treated with rifampin alone increased from 16 to > 256 mg/L (16-folds) and the MIC of colistin from the colonies recovered from the lungs of mice treateted with CMS as monotherapy increased from 1 to > 32 mg/L (32-folds). Nevertheless, the MICs of rifampin and colistin remained unchanged for those recovered from the control and the combination group.
Discussion
The results of this study show that CMS alone has no significant efficacy in terms of bacterial clearance either from tissue and blood or in the survival rate in the peritoneal sepsis model, even in animals infected with strains susceptible to this antimicrobial. To investigate whether these disappointing results were due to a model effect, additional experiments were made in a less severe murine pneumonia model using the strain producing OXA-48 plus CTX-M-15. Once again, the bacterial lung and blood concentrations as well as mortality were no different from those of the control group. We believe that these results are related with the development of colistin resistance during the CMS monotherapy found in the pneumonia model.
The CMS dosage used in this study has been proven to be effective in several murine experimental studies using clinical multidrug-resistant (MDR) Acinetobacter baumannii strains susceptible to colistin (MIC = 0.5 mg/L) (Pachon-Ibanez et al., 2010; Docobo-Perez et al., 2012; Parra Millan et al., 2016). Nevertheless, when this CMS dosage was used in a murine pneumonia model by a NDM-1-producing K. pneumoniae clinical strain, CMS monotherapy was not effective as we found in the present study (Docobo-Perez et al., 2012). Similar results were reported by de Oliveira et al. (2015) in a retrospective clinical study, observing suboptimal efficacy of polymyxins in the treatment of KPC-producing Enterobacteriaceae infections, even in combination with imipenem or meropenem.
One possible explanation to the lack of efficacy found with CMS alone, could be that the PD parameter predictive of colistin efficacy, fAUC0-24/MIC ratio (Cheah et al., 2015), has been described in infections caused by other Gram-negative bacilli, such as Pseudomonas aeruginosa and A. baumannii. So, the present results would suggest that the described fAUC0-24/MIC values to achieve various magnitudes of bacterial killing in these Gram-negative bacilli are not the appropriate for the treatment of CP-Kp infections, even for isolates with the same MIC values. However, in these studies the end-point to define the fAUC0-24/MIC values predicting colistin efficacy was to achieve a 2 log10 decrease in bacterial concentrations in lungs or thigh, but not to evaluate the mice survival rates.
Colistimethate sodium monotherapy did not reduce significantly the bacterial lung and blood results or the survival in the less severe pneumonia model by the strain producing OXA-48 plus CTX-M-15. However, it is noteworthy that CMS achieved a bacterial lung decrease of 2.91 log10 cfu/g, a value defined as optimal for P. aeruginosa and A. baumannii (Cheah et al., 2015), although the wide SD precluded the significance of these data. In addition, the results in the pneumonia model demonstrate that the severity of the chosen animal model is important when studying the efficacy of antimicrobial treatments. Thus, new colistin fAUC0-24/MIC values have to be defined for CP-Kp and, in our opinion, for the treatment of other Gram-negative bacilli infections.
The activity of rifampin in monotherapy in the peritoneal sepsis model was also limited, reducing only bacterial spleen concentration compared to control and CMS groups, infected with VIM-1 or OXA-48 plus CTX-M-15 strains, for which an AUC0-24 h/MIC = 34.5, predictor of efficacy was achieved. In the case of animals infected with the KPC-3 strain, rifampin did not decrease significantly the bacterial load in spleen or blood, but reduced the mortality. However, the survival mice conditions suggested that if the experiment were longer than 72 h, all animals included in this treatment group would have died shortly after. Moreover, a huge increase in MIC of rifampin was observed after rifampin in vivo monotherapy. In the less severe pneumonia model by OXA-48 plus CTX-M-15 strain, rifampin monotherapy increased significantly the bacterial clearing from lungs and blood, and also increased the mice survival. The efficacy of rifampin against other Gram-negative bacteria has been showed in experimental pneumonia models in mice (Wolff et al., 1999; Montero et al., 2004; Pachon-Ibanez et al., 2010, 2011). Nevertheless, rifampin alone cannot be an alternative due to the generation of rifampin-resistant mutants (Pachon-Ibanez et al., 2006).
Colistimethate sodium in combination with rifampin demonstrated in vitro a synergistic effect against the four strains, but the effect was bactericidal only against KPC-3 strain. These results are in accordance with other studies showing synergy of this combination against KPC-producing K. pneumoniae strains. Nastro et al. (2014) reported a synergistic effect at 24 h against 27 colistin-resistant KPC-2-producing K. pneumoniae clinical strains. Similarly, Elemam et al. (2010) found synergy with polymyxin B plus rifampin against 12 KPC-producing K. pneumoniae clinical strains. Tascini et al. (2013) also reported that CMS plus rifampin exhibited synergy against 13 colistin-resistant KPC-producing K. pneumoniae clinical strains, moreover being bactericidal against the 62%. On the contrary, in the case of K. pneumoniae producing VIM-1 the results of the present study differ from those by Tangden et al. (2014), where CMS plus rifampin did not exhibit synergy against two colistin-susceptible VIM-1-producing K. pneumoniae clinical strains.
With regard to the in vivo efficacy of CMS plus rifampin in the experimental murine peritoneal sepsis model by the colistin-susceptible VIM-1, the combination was better than the control and CMS groups taking into account the bacterial clearance from spleen and blood. When using the colistin-susceptible OXA-48 plus CTX-M-15 strain, the combination only reduced the bacterial spleen concentration. For these two colistin-susceptible strains, CMS and rifampin achieved the pharmacodynamic values described as optimal, fAUC0-24/MIC and AUC0-24/MIC, respectively (Jayaram et al., 2003; Gumbo et al., 2007; Landersdorfer et al., 2017). Nevertheless, is worth mentioning that the efficacy of the combination for both colistin-susceptible strains was not optimal, if we consider that mortality rates remained 93.3 and 73.3%, respectively.
Against the colistin-resistant KPC-3 strain, this combination reduced the bacterial spleen concentration and mortality, in accordance with the in vitro synergy studies. Nastro et al. (2014) reported a favorable outcome in five patients with colistin-resistant KPC-2-producing K. pneumoniae infections when treated with a combination of CMS plus rifampin. Against the other colistin-resistant strain, producing VIM-1/DHA-1, this combination was not efficacious, in contrast with the synergy observed in the time-kill studies.
In the pneumonia model caused by the OXA-48 plus CTX-M-15 strain, the activity of CMS plus rifampin was similar to that of rifampin in monotherapy, decreasing the bacterial lung concentration and increasing the mice survival compared with the control group. This combination has been successfully used against other Gram-negative bacilli (Pachon-Ibanez et al., 2010). In this pneumonia model, the strain of K. pneumoniae developed colistin resistance and a considerable increase in the MIC of rifampin when used as monotherapies, which was prevented with the combination. This prevention of colistin resistant mutant with this combination have been reported previously as in the in vitro study published by Rodriguez et al. (2010) in which they proved that the association of colistin plus rifampin was synergistic against heteroresistant A. baumannii isolates and prevented the development of colistin-resistant mutants (Rodriguez et al., 2010).
Limitations of the study have to be done, first that even though we wanted to evaluate the in vitro and in vivo effect of colistin plus rifampin combination against clonally unrelated clinical isolates of K. pneumoniae producing other carbapenemases, the number of the tested strains is low. Moreover, the less severe pneumonia model was only performed with one of the strains, and we believe it will be very interesting to do it with all of the strains.
In summary, the results obtained suggest that CMS plus rifampin has a low and heterogeneous efficacy in the treatment of severe infections, such as a peritoneal sepsis infection, caused by different CP-Kp strains, increasing only the mice survival in the infection caused by the KPC-3 strain. CMS plus rifampin combination prevents in vivo the development of mutants resistant to colistin. Because of the lack of available alternatives for these colistin-resistant KPC-3-producing strains, the combination of CMS and rifampin might be considered for further evaluation. Moreover, this combination showed efficacy in the less severe pneumonia model by the OXA-48 plus CTX-M-15 strain. Finally, an optimal PD index value for CMS efficacy needs to be defined for K. pneumoniae. Overall, the results of the present study suggest that the efficacy of CMS plus rifampin depends on the class of carbapenemase produced by K. pneumoniae and on the severity of the infection.
Author Contributions
MP-I has planned and coordinated the experiments, analyzed the results, and written the manuscript. GL-H and TC-C had performed the in vitro and in vivo experiments. CD and JPP had performed the antibiotic concentrations studies by HPLC-tandem mass spectrometry (LC-MS/MS). YS had reviewed the manuscript and the experiments. JR-B, AP, and JP had reviewed the manuscript and the experiments. MCC obtained the funds to perform the studies and wrote the project, contributed to the performance of the in vitro experiment, and reviewed the results and the manuscript.
Funding
This study was supported by the Consejería de Salud of the Junta de Andalucía (PI-0622-2012) and supported by Plan Nacional de I+D+i and Instituto de Salud Carlos III, Subdirección General de Redes y Centros de Investigación Cooperativa, Ministerio de Economía y Competitividad, Spanish Network for Research in Infectious Diseases (REIPI RD12/0015/0001) – co-financed by European Development Regional Fund “A way to achieve Europe” ERDF. The MEDINA authors disclosed the receipt of financial support from Fundación MEDINA, a public-private partnership of Merck Sharp & Dohme de España S.A./Universidad de Granada/Junta de Andalucía.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00912/full#supplementary-material
Footnotes
References
Canton, R., Akova, M., Carmeli, Y., Giske, C. G., Glupczynski, Y., Gniadkowski, M., et al. (2012). Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 18, 413–431. doi: 10.1111/j.1469-0691.2012.03821.x
Cheah, S. E., Wang, J., Nguyen, V. T., Turnidge, J. D., Li, J., and Nation, R. L. (2015). New pharmacokinetic/pharmacodynamic studies of systemically administered colistin against Pseudomonas aeruginosa and Acinetobacter baumannii in mouse thigh and lung infection models: smaller response in lung infection. J. Antimicrob. Chemother. 70, 3291–3297. doi: 10.1093/jac/dkv267
Clinical and Laboratory Standards Institute [CLSI] (2012). Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard—Ninth Edition. CLSI Document M07-A9. Wayne, PA: Clinical and Laboratory Standards Institute.
Daikos, G. L., Tsaousi, S., Tzouvelekis, L. S., Anyfantis, I., Psichogiou, M., Argyropoulou, A., et al. (2014). Carbapenemase-producing Klebsiella pneumoniae bloodstream infections: lowering mortality by antibiotic combination schemes and the role of carbapenems. Antimicrob. Agents Chemother. 58, 2322–2328. doi: 10.1128/AAC.02166-13
De Laveleye, M., Huang, T. D., Bogaerts, P., Berhin, C., Bauraing, C., Sacre, P., et al. (2017). Increasing incidence of carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in Belgian hospitals. Eur. J. Clin. Microbiol. Infect. Dis. 36, 139–146. doi: 10.1007/s10096-016-2782-x
de Oliveira, M. S., De Assis, D. B., Freire, M. P., Boas Do Prado, G. V., Machado, A. S., Abdala, E., et al. (2015). Treatment of KPC-producing Enterobacteriaceae: suboptimal efficacy of polymyxins. Clin. Microbiol. Infect. 21, 179.e1–179.e7. doi: 10.1016/j.cmi.2014.07.010
Docobo-Perez, F., Nordmann, P., Dominguez-Herrera, J., Lopez-Rojas, R., Smani, Y., Poirel, L., et al. (2012). Efficacies of colistin and tigecycline in mice with experimental pneumonia due to NDM-1-producing strains of Klebsiella pneumoniae and Escherichia coli. Int. J. Antimicrob. Agents 39, 251–254. doi: 10.1016/j.ijantimicag.2011.10.012
Elemam, A., Rahimian, J., and Doymaz, M. (2010). In vitro evaluation of antibiotic synergy for polymyxin B-resistant carbapenemase-producing Klebsiella pneumoniae. J. Clin. Microbiol. 48, 3558–3562. doi: 10.1128/JCM.01106-10
European Committee on Antimicrobial Susceptibility Testing [EUCAST] (2016). European Society of Clinical Microbiology and Infectious Diseases. Clinical Breakpoints. Bochum: EUCAST.
Falagas, M. E., Tansarli, G. S., Karageorgopoulos, D. E., and Vardakas, K. Z. (2014). Deaths attributable to carbapenem-resistant Enterobacteriaceae infections. Emerg. Infect. Dis. 20, 1170–1175. doi: 10.3201/eid2007.121004
Gobin, P., Lemaitre, F., Marchand, S., Couet, W., and Olivier, J. C. (2010). Assay of colistin and colistin methanesulfonate in plasma and urine by liquid chromatography-tandem mass spectrometry. Antimicrob. Agents Chemother. 54, 1941–1948. doi: 10.1128/AAC.01367-09
Gonzalez, D., Schmidt, S., and Derendorf, H. (2013). Importance of relating efficacy measures to unbound drug concentrations for anti-infective agents. Clin. Microbiol. Rev. 26, 274–288. doi: 10.1128/CMR.00092-12
Gumbo, T., Louie, A., Deziel, M. R., Liu, W., Parsons, L. M., Salfinger, M., et al. (2007). Concentration-dependent Mycobacterium tuberculosis killing and prevention of resistance by rifampin. Antimicrob. Agents Chemother. 51, 3781–3788. doi: 10.1128/AAC.01533-06
Jayaram, R., Gaonkar, S., Kaur, P., Suresh, B. L., Mahesh, B. N., Jayashree, R., et al. (2003). Pharmacokinetics-pharmacodynamics of rifampin in an aerosol infection model of tuberculosis. Antimicrob. Agents Chemother. 47, 2118–2124. doi: 10.1128/AAC.47.7.2118-2124.2003
Landersdorfer, C. B., Nguyen, T. H., Lieu, L. T., Nguyen, G., Bischof, R. J., Meeusen, E. N., et al. (2017). Substantial targeting advantage achieved by pulmonary administration of colistin methanesulfonate in a large-animal model. Antimicrob. Agents Chemother. 61:e01934-16. doi: 10.1128/AAC.01934-16
Lee, G. C., and Burgess, D. S. (2012). Treatment of Klebsiella pneumoniae carbapenemase (KPC) infections: a review of published case series and case reports. Ann. Clin. Microbiol. Antimicrob. 11:32. doi: 10.1186/1476-0711-11-32
Lopez-Cerero, L., Egea, P., Gracia-Ahufinger, I., Gonzalez-Padilla, M., Rodriguez-Lopez, F., Rodriguez-Bano, J., et al. (2014). Characterisation of the first ongoing outbreak due to KPC-3-producing Klebsiella pneumoniae (ST512) in Spain. Int. J. Antimicrob. Agents 44, 538–540. doi: 10.1016/j.ijantimicag.2014.08.006
Michalopoulos, A., Virtzili, S., Rafailidis, P., Chalevelakis, G., Damala, M., and Falagas, M. E. (2010). Intravenous fosfomycin for the treatment of nosocomial infections caused by carbapenem-resistant Klebsiella pneumoniae in critically ill patients: a prospective evaluation. Clin. Microbiol. Infect. 16, 184–186. doi: 10.1111/j.1469-0691.2009.02921.x
Miro, E., Aguero, J., Larrosa, M. N., Fernandez, A., Conejo, M. C., Bou, G., et al. (2013). Prevalence and molecular epidemiology of acquired AmpC β-lactamases and carbapenemases in Enterobacteriaceae isolates from 35 hospitals in Spain. Eur. J. Clin. Microbiol. Infect. Dis. 32, 253–259. doi: 10.1007/s10096-012-1737-0
Montero, A., Ariza, J., Corbella, X., Domenech, A., Cabellos, C., Ayats, J., et al. (2004). Antibiotic combinations for serious infections caused by carbapenem-resistant Acinetobacter baumannii in a mouse pneumonia model. J. Antimicrob. Chemother. 54, 1085–1091. doi: 10.1093/jac/dkh485
Munoz-Price, L. S., Poirel, L., Bonomo, R. A., Schwaber, M. J., Daikos, G. L., Cormican, M., et al. (2013). Clinical epidemiology of the global expansion of Klebsiella pneumoniae carbapenemases. Lancet Infect. Dis. 13, 785–796. doi: 10.1016/S1473-3099(13)70190-7
Nastro, M., Rodriguez, C. H., Monge, R., Zintgraff, J., Neira, L., Rebollo, M., et al. (2014). Activity of the colistin-rifampicin combination against colistin-resistant, carbapenemase-producing Gram-negative bacteria. J. Chemother. 26, 211–216. doi: 10.1179/1973947813Y.0000000136
Nordmann, P., Dortet, L., and Poirel, L. (2012). Carbapenem resistance in Enterobacteriaceae: here is the storm! Trends Mol. Med. 18, 263–272. doi: 10.1016/j.molmed.2012.03.003
Nordmann, P., Naas, T., and Poirel, L. (2011). Global spread of Carbapenemase-producing Enterobacteriaceae. Emerg. Infect. Dis. 17, 1791–1798. doi: 10.3201/eid1710.110655
Oteo, J., Ortega, A., Bartolome, R., Bou, G., Conejo, C., Fernandez-Martinez, M., et al. (2015). Prospective multicenter study of carbapenemase-producing Enterobacteriaceae from 83 hospitals in Spain reveals high in vitro susceptibility to colistin and meropenem. Antimicrob. Agents Chemother. 59, 3406–3412. doi: 10.1128/AAC.00086-15
Pachon-Ibanez, M. E., Docobo-Perez, F., Jimenez-Mejias, M. E., Ibanez-Martinez, J., Garcia-Curiel, A., Pichardo, C., et al. (2011). Efficacy of rifampin, in monotherapy and in combinations, in an experimental murine pneumonia model caused by panresistant Acinetobacter baumannii strains. Eur. J. Clin. Microbiol. Infect. Dis. 30, 895–901. doi: 10.1007/s10096-011-1173-6
Pachon-Ibanez, M. E., Docobo-Perez, F., Lopez-Rojas, R., Dominguez-Herrera, J., Jimenez-Mejias, M. E., Garcia-Curiel, A., et al. (2010). Efficacy of rifampin and its combinations with imipenem, sulbactam, and colistin in experimental models of infection caused by imipenem-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 54, 1165–1172. doi: 10.1128/AAC.00367-09
Pachon-Ibanez, M. E., Fernandez-Cuenca, F., Docobo-Perez, F., Pachon, J., and Pascual, A. (2006). Prevention of rifampicin resistance in Acinetobacter baumannii in an experimental pneumonia murine model, using rifampicin associated with imipenem or sulbactam. J. Antimicrob. Chemother. 58, 689–692. doi: 10.1093/jac/dkl303
Palacios-Baena, Z. R., Oteo, J., Conejo, C., Larrosa, M. N., Bou, G., Fernandez-Martinez, M., et al. (2016). Comprehensive clinical and epidemiological assessment of colonisation and infection due to carbapenemase-producing Enterobacteriaceae in Spain. J. Infect. 72, 152–160. doi: 10.1016/j.jinf.2015.10.008
Parra Millan, R., Jimenez Mejias, M. E., Sanchez Encinales, V., Ayerbe Algaba, R., Gutierrez Valencia, A., Pachon Ibanez, M. E., et al. (2016). Efficacy of lysophosphatidylcholine in combination with antimicrobial agents against Acinetobacter baumannii in experimental murine peritoneal sepsis and pneumonia models. Antimicrob. Agents Chemother. 60, 4464–4470. doi: 10.1128/AAC.02708-15
Pournaras, S., Vrioni, G., Neou, E., Dendrinos, J., Dimitroulia, E., Poulou, A., et al. (2011). Activity of tigecycline alone and in combination with colistin and meropenem against Klebsiella pneumoniae carbapenemase (KPC)-producing Enterobacteriaceae strains by time-kill assay. Int. J. Antimicrob. Agents 37, 244–247. doi: 10.1016/j.ijantimicag.2010.10.031
Rodriguez, C. H., De Ambrosio, A., Bajuk, M., Spinozzi, M., Nastro, M., Bombicino, K., et al. (2010). In vitro antimicrobials activity against endemic Acinetobacter baumannii multiresistant clones. J. Infect. Dev. Ctries. 4, 164–167. doi: 10.3855/jidc.604
Souli, M., Galani, I., Boukovalas, S., Gourgoulis, M. G., Chryssouli, Z., Kanellakopoulou, K., et al. (2011). In vitro interactions of antimicrobial combinations with fosfomycin against KPC-2-producing Klebsiella pneumoniae and protection of resistance development. Antimicrob. Agents Chemother. 55, 2395–2397. doi: 10.1128/AAC.01086-10
Souli, M., Rekatsina, P. D., Chryssouli, Z., Galani, I., Giamarellou, H., and Kanellakopoulou, K. (2009). Does the activity of the combination of imipenem and colistin in vitro exceed the problem of resistance in metallo-β-lactamase-producing Klebsiella pneumoniae isolates? Antimicrob. Agents Chemother. 53, 2133–2135. doi: 10.1128/AAC.01271-08
Tangden, T., Hickman, R. A., Forsberg, P., Lagerback, P., Giske, C. G., and Cars, O. (2014). Evaluation of double- and triple-antibiotic combinations for VIM- and NDM-producing Klebsiella pneumoniae by in vitro time-kill experiments. Antimicrob. Agents Chemother. 58, 1757–1762. doi: 10.1128/AAC.00741-13
Tascini, C., Tagliaferri, E., Giani, T., Leonildi, A., Flammini, S., Casini, B., et al. (2013). Synergistic activity of colistin plus rifampin against colistin-resistant KPC-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 57, 3990–3993. doi: 10.1128/AAC.00179-13
Tenover, F. C., Arbeit, R. D., Goering, R. V., Mickelsen, P. A., Murray, B. E., Persing, D. H., et al. (1995). Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J. Clin. Microbiol. 33, 2233–2239.
Tzouvelekis, L. S., Markogiannakis, A., Psichogiou, M., Tassios, P. T., and Daikos, G. L. (2012). Carbapenemases in Klebsiella pneumoniae and other Enterobacteriaceae: an evolving crisis of global dimensions. Clin. Microbiol. Rev. 25, 682–707. doi: 10.1128/CMR.05035-11
Waters, N. J., Jones, R., Williams, G., and Sohal, B. (2008). Validation of a rapid equilibrium dialysis approach for the measurement of plasma protein binding. J. Pharm. Sci. 97, 4586–4595. doi: 10.1002/jps.21317
Wolff, M., Joly-Guillou, M. L., Farinotti, R., and Carbon, C. (1999). In vivo efficacies of combinations of beta-lactams, beta-lactamase inhibitors, and rifampin against Acinetobacter baumannii in a mouse pneumonia model. Antimicrob. Agents Chemother. 43, 1406–1411.
Keywords: Klebsiella pneumoniae, animal models, carbapenemase producers, colistin, rifampin
Citation: Pachón-Ibáñez ME, Labrador-Herrera G, Cebrero-Cangueiro T, Díaz C, Smani Y, del Palacio JP, Rodríguez-Baño J, Pascual A, Pachón J and Conejo MC (2018) Efficacy of Colistin and Its Combination With Rifampin in Vitro and in Experimental Models of Infection Caused by Carbapenemase-Producing Clinical Isolates of Klebsiella pneumoniae. Front. Microbiol. 9:912. doi: 10.3389/fmicb.2018.00912
Received: 25 January 2018; Accepted: 19 April 2018;
Published: 15 May 2018.
Edited by:
Noton Kumar Dutta, Johns Hopkins University, United StatesReviewed by:
Maria Bagattini, University of Naples Federico II, ItalyRemy A. Bonnin, Université Paris-Saclay, France
Copyright © 2018 Pachón-Ibáñez, Labrador-Herrera, Cebrero-Cangueiro, Díaz, Smani, del Palacio, Rodríguez-Baño, Pascual, Pachón and Conejo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María E. Pachón-Ibáñez, bXBhY2hvbi1pYmlzQHVzLmVz