Abstract
The recent worldwide discovery of plant growth-promoting (PGP) Kosakonia radicincitans in a large variety of crop plants suggests that this species confers significant influence on plants, both in terms of yield increase and product quality improvement. We provide a comparative genome analysis which helps to unravel the genetic basis for K. radicincitans' motility, competitiveness and plant growth-promoting capacities. We discovered that K. radicincitans carries multiple copies of complex gene clusters, among them two flagellar systems and three type VI secretion systems (T6SSs). We speculate that host invasion may be facilitated by different flagella, and bacterial competitor suppression by effector proteins ejected via T6SSs. We found a large plasmid in K. radicincitans DSM 16656T, the species type strain, that confers the potential to exploit plant-derived carbon sources. We propose that multiple copies of complex gene clusters in K. radicincitans are metabolically expensive but provide competitive advantage over other bacterial strains in nutrient-rich environments. The comparison of the DSM 16656T genome to genomes of other genera of enteric plant growth-promoting bacteria (PGPB) exhibits traits unique to DSM 16656T and K. radicincitans, respectively, and traits shared between genera. We used the output of the in silico analysis for predicting the purpose of genomic features unique to K. radicincitans and performed microarray, PhyloChip, and microscopical analyses to gain deeper insight into the interaction of DSM 16656T, plants and associated microbiota. The comparative genome analysis will facilitate the future search for promising candidates of PGPB for sustainable crop production.
Introduction
Among the approaches to achieve food security and the sustainable handling of resources, one of the most promising is considered the tailored application of microorganism-based inoculants in agriculture, with an expected compound annual growth rate of 9% during 2015–2020 on a global scale (http://www.researchandmarkets.com/reports/3786558). Plant-associated microorganisms have proven to protect their hosts from pathogens and herbivores, thus providing efficient supplements of, or alternatives to pesticides, while other microorganisms were shown to increase nutrient availability, thus promoting plant growth and yield. Over the past two decades effort has been made to explore the association between so-called plant growth-promoting bacteria (PGPB) and non-leguminous plants for producing agricultural inoculants made from bacteria. Being the three most produced cereals of the world and accounting for approximately two-thirds of human food consumption, maize, rice and wheat are of particular interest for the application of PGPB. In contrast to symbiotic, root nodule-colonizing rhizobia (= plant development-enhancing strains of family Rhizobiaceae) and filamentous Actinobacteria from genus Frankia, many enteric PGPB, particularly strains from genera Enterobacter, Klebsiella and Kosakonia, are able to colonize a wide range of hosts. The identification of PGPB is challenging and time-consuming. Reducing the entire microbiota of plants to a few promising candidates with high PGP potential requires numerous experiments on beneficial traits. In 1986 we isolated nearly a thousand bacterial strains from the rhizosphere and phyllosphere of winter wheat (Triticum aestivum cv. “Alcedo”) and selected putatively plant growth-promoting strains. One of them possessed stable and high plant growth-promoting as well as yield-promoting activity (Ruppel, 1988). The bacterial strain was explored as the new species Enterobacter radicincitans, strain D5/23T (Kampfer et al., 2005), today deposited as type strain at the Leibniz Institute DSMZ-German Collection of Microorganisms and Cell Cultures under DSM 16656T. In 2013, the year after we published a draft genome of E. radicincitans (Witzel et al., 2012), Enterobacter was taxonomically divided into five genera, one of which was named Kosakonia (Brady et al., 2013). In the course of this reclassification E. radicincitans was renamed Kosakonia radicincitans.
Remus (Remus et al., 2000) found that inoculating K. radicincitans into its native host, the wheat cv. “Alcedo,” resulted in higher grain yields, indicating that even plants naturally hosting PGPB can show better performance when particular microorganisms are enriched. However, K. radicincitans does not only positively affect cereals but also vegetables, for instance of the family Brassicaceae, e.g., Brassica oleracea (Ruppel et al., 2006), Raphanus sativus (Berger et al., 2015) and Solanaceae, e.g., Solanum lycopersicum (Berger et al., 2013). The PGP capacity of DSM 16656T and other Kosakonia strains was experimentally demonstrated. Distinct increases in growth, yield and product quality by K. radicincitans DSM 16656T have been confirmed in glasshouse experiments as well as in field trials (Schreiner et al., 2009; Berger et al., 2017, 2018) and highlight the potential of this strain for different cultivation management systems.
PGP Kosakonia strains have been found to naturally colonize plants in nearly all major clades of angiosperms. In contrast to root nodule-inducing bacteria (Frankia and rhizobia), Kosakonia is not restricted to a subgroup of rosids, but does also inhabit asterids and monocots. Among the natural hosts of K. radicincitans are many crops of global economic relevance. Most notably are the staple crops maize, rice, wheat and sweet potato, the multipurpose crop sugarcane, the fiber crop cotton, and the biofuel plant Jatropha. However, peanut, pineapple, tomato and the beverage plant yerba mate are also among the domesticated plants that host Kosakonia (for references see Supplementary Table S1). Most of these Kosakonia strains were shown to promote plant growth. To date, full genomes or whole genome shotgun (WGS) sequences of 20 strains of Kosakonia have been made available at NCBI (although two of them are also referred to as Enterobacter): K. radicincitans (DSM 16656T, YD4, GXGL-4A, UMEnt01/12), K. oryzae (Ola-51, KO348, D4, CGMCC 1.7012), K. sacchari (SP1, BO-1, CGMCC 1.12102), K. arachidis (Ah-143), K. pseudosacchari (JM-387), K. oryziphila (REICA_142), K. oryzendophytica (REICA_082, LMG 26432), K. cowanii (888-76, DSM 18146), Enterobacter sp. (R4-368, FY-07).
The recently sequenced and published genomes of Kosakonia spp. provided the opportunity to perform comparative genome analyses among Kosakonia and between main groups of enteric PGPB. For this purpose we re-sequenced the genome of DSM 16656T, the type strain of K. radicincitans, using the SMRT (single molecule real time) technology of Pacific Biosciences (PacBio) and applied it as reference. One aim of this study was to evaluate the usefulness of comparative genome analyses for in silico prediction of promising bacterial features. Very recently, a comprehensive comparative genome analysis by Levy and co-workers pursued the same goal and found that plant-associated bacteria from across the bacterial kingdom share particular genomic features (Levy et al., 2018).
Here, we present a comprehensive overview of the enteric bacterium Kosakonia radicincitans, its phylogenetic relationships and hosts based on the literature. We provide the complete genome sequence and annotation for type strain K. radicincitans DSM 16656T. Using comparative genomics we were able to unravel the genetic basis for the high PGP competence, motility and competitiveness of this strain. Based on genomic features unique to K. radicincitans we developed hypotheses that we tested by in vivo experiments applying transcriptomic, bacterial community composition, and microscopical analyses.
Materials and methods
Details such as primer sequences, links to webpages and applied computer programs are provided in the Supplementary File.
Pacbio library preparation and sequencing
SMRTbell™ template library was prepared according to the instructions from Pacific Biosciences, following the Procedure & Checklist − 20 kb Template Preparation using BluePippin™ Size-Selection System. Briefly, for preparation of 15 kb libraries 8 μg genomic DNA libraries was sheared using g-tubes™ from Covaris, according to the manufacturer's instructions. DNA was end-repaired and ligated overnight to hairpin adapters applying components from the DNA/Polymerase Binding Kit P6 from Pacific Biosciences. Reactions were carried out according to the manufacturer's instructions. BluePippin™ Size-Selection to 7 kb was performed according to the manufacturer's instructions (Sage Science). Conditions for annealing of sequencing primers and binding of polymerase to purified SMRTbell™ template were assessed with the Calculator in RS Remote, Pacific Biosciences. SMRT sequencing was carried out on the PacBio RSII (Pacific Biosciences) taking one 240-min movie for each SMRT cell. In total 1 SMRT cell was run. 98,548 reads with a mean read length of 12,899 bp were obtained.
Genome assembly, error correction, and annotation
SMRT Cell data was assembled using the “RS_HGAP_Assembly.3” protocol included in SMRT Portal version 2.3.0 using default parameters. The assembly revealed a circular chromosome and two plasmids. Validity of the assembly was checked using the “RS_Bridgemapper.1” protocol. Each genome was error-corrected by a mapping of Illumina reads onto finished genomes using BWA (Li and Durbin, 2009) with subsequent variant and consensus calling using VarScan (Koboldt et al., 2012). A consensus concordance of QV60 could be confirmed for the genome. Finally, an annotation was generated using the software tool for rapid prokaryotic genome annotation (PROKKA) version 1.8 (Seemann, 2014). The genome was deposited in NCBI GenBank under the accession numbers CP018016.1 (chromosome), CP018017.1 (large plasmid), CP018018.1 (small plasmid).
Gene annotation and functional annotation
Genome annotation was derived from PROKKA, RNAmmer and ARAGORN. Functional annotation was received from the SEED-based “Rapid Annotations using Subsystems Technology” (RAST) tool from the RAST server and pathway classification from “KEGG Orthology And Links Annotation” tool (BlastKOALA).
Phylogenetic analysis
16S rRNA gene sequences and metadata of published K. radicincitans strains, closely related Kosakonia species and other genera of enteric PGPB were collected from NCBI databases. The 16S rRNA gene sequences originating from the species whole genome sequence were aligned with MUSCLE and the resulting tree was inferred based on the Maximum Likelihood approach considering the Jukes Cantor model of evolution. An initial tree was obtained by the Neighbor Joining algorithm. Bootstraps values indicated as numbers in percentage close to the nodes represent the confidence level of a clade by resampling the dataset 1,000 times. All algorithms were executed with Geneious (8.1.9). Likewise, a protein tree was built on concatenated amino acid sequences of the phylogenetic marker genes atpD, gyrB, infB, and recA taking the BLOSUM62 substitution matrix into account. According to the phylogenetic distances and the pairwise identity, received by the BLAST results distinct phylogenetic groups could be determined.
Genome comparison
The nucleotide and amino acid sequences for the whole genomes were retrieved from NCBI. We compared the DSM 16656T genome sequence to 31 closely related and three distantly related, fully sequenced and annotated genomes of the following bacteria: K. radicincitans group (Ola 51T, GXGL-4A, YD4, UMEnt01/12, REICA_142), K. sacchari group (SP1T, R4-368, KO348, BO-1), K. variicola group (DX120E, DSM 15968, 342, D5A), Enterobacter group (REICA_082, FY-07, SBP-8, P101, DC1, DC3, DC4, ENHKU01, SST3, GS1, 638, UCD-UG_FMILLET), Citrobacter (FDAARGOS_122, FDAARGOS_156, FDAARGOS_164, FDAARGOS_165, Y19, CAV1321), Pseudomonas fluorescens (F113), Rhizobium leguminosarum bv. viciae (3841), and Bacillus velezensis (FZB42). To obtain conserved genomic regions a MAUVE (version 2.4.0) alignment (http://darlinglab.org/mauve/mauve.html) was generated for each phylogenetic group: K. radicincitans (KORA), K. sacchari (KOSA), Enterobacter (ENTERO) and Klebsiella variicola (KLEVA) with DSM 16656T as reference. To detect orthologous genes and gene clusters an in-house pipeline was applied. In its first step all proteins were aligned to each other using BLASTp+ by excluding hits with an identity lower 35%, a coverage lower 60% and an e-value cut-off with 1e−5, and considering the reciprocal best hit algorithm (RBH) and scoring, as well as the conserved regions. In a second step the orthologous genes/clusters were weighted for their affiliation to a conserved region defined by MAUVE, as mentioned above. In a third step the resulting clusters were divided into the different phylogenetic group clusters for core genome assignment accordingly to their binary absence/presence pattern. To expose differences in specific functional genes or gene clusters occurrences their copy number (for a single gene) or total gene content was extracted from the RBH-table and verified by manual revision of all considered genomes. Finally for visualization SVG files were generated with BRIG (Version 0.95, http://brig.sourceforge.net/) and in-house scripts.
For core genome analysis we chose three representative strains per group [(i) KORA (DSM 16656T, YD4, Ola 51), (ii) KOSA (KO348, SP1, BO-1), (iii) ENTERO (DC1, SST3, P101), and (iv) KLEVA (342, DX120E, DSM 15968)] according to the following criteria: (i) endophytic, plant growth-promoting and/or diazotrophic (N2-fixing), (ii) isolated from crop plants, (iii) collected in different countries, and (iv) high genetic identity among representatives of each group (Supplementary Table S1).
Root exudate collection
Tomato plants, Solanum lycopersicum cv. “Vanessa” (HILD) were cultivated in a sand: vermiculite mixture (v: v, 1:1) for 4 weeks. For exudate collection the substrate was gently removed by washing the roots. Then, the tomato plants were kept in sterile bi-distilled water for 1 h, followed by a 4 h collection period in bi-distilled water. The root exudate-containing water was two step-vacuum filtered (through 0.45 and 0.2 μm sterile filters) and then frozen and freeze-dried for further analysis. The C/N ratio was measured using a CNS Vario EL analyzer (Elementar).
Customized microarray: cultivation of bacteria, microarray design, analysis and quality control
K. radicincitans DSM 16656T was grown in 20 mL shaking culture at 28°C and 190 rpm for 24 h in mineral medium (Gerhardt et al., 1994) supplemented either with sucrose and glucose or with tomato root exudates (filter sterilized), both adjusted to 5 g C L−1. After 24 h bacteria were pelleted and frozen in liquid nitrogen. RNA was extracted from the pellet using innuPREP RNA Mini Kit (Analytik Jena) and quantified by electrophoresis using an Agilent 2100 Bioanalyzer®. Four replicates of each bacterial treatment were fragmented, labeled and hybridized to the DSM 16656T custom array. Using the web based eArray application (Agilent Technologies) we designed a microarray on an 8 × 60 k formatted slide using 5,512 protein coding genes of K. radicincitans DSM 16656T, which could be assigned unambiguously to PacBio sequences. For each gene 10 specific oligonucleotide probe sequences were designed for hypervariable regions and spotted along with 1,319 internal Agilent quality controls. Raw signal intensity values were determined using Agilent standard protocol for Affymetrix Microarray Analyzer from ATLAS-Biolabs. Probe level values (Agilent “gProcessedSignal”) were preprocessed using Quantile normalization from R-package preprocessCore and log transformed. The Agilent “IfPosAndSig” values 1 or 0 were used as detection-threshold. In order to estimate the quality of the used samples, the Pearson correlation between all sample pairs was calculated and used as distance measurement. Using Pearson correlation together with “Ward” linkage method in the hierarchical clustering one sample was detected to be an outlier and removed. Follow-up analyses were performed on the seven remaining samples. In order to determine genomic regions of K. radicincitans DSM 16656T differentially expressed (up- or down-regulated), a genome-wide sliding window calculation was performed considering a window size of 15 consecutive genes.
Microbiome phylochip analysis
K. radicincitans DSM 16656T was grown, washed and diluted as described in Ruppel et al. (2006). Seeds of tomato, S. lycopersicum cv. “Vanessa” were either exposed to 108 cells of DSM 16656T or to pure buffer solution (0.05 M NaCl = control), then germinated and again inoculated, this time by spraying 108 cells onto the leaves of each plant at seedling (2-leaf) stage. Pure buffer was sprayed on controls. Plants were grown in non-sterile quartz sand (grain size 0.5–1.0 mm). Two weeks after spraying, leaves and roots were collected separately in liquid nitrogen, freeze-dried and total DNA was extracted using DNeasy Plant MiniKit (Qiagen). Colonization with K. radicincitans was quantified using a K. radicincitans-specific TaqMan™ probe labeled with 6-FAM and a black hole quencher at the 3′-end was used in quantitative real-time PCR using the plant TEF gene as a reference gene as described in Ruppel et al. (2006). The absolute bacterial cell quantification was conducted using the target gene calibration curve ranging from 10 to 109 copy numbers per μL. In high quality DNA the bacterial 16S rRNA genes were amplified. Sixteen samples (four replicates of root and shoot non-inoculated and inoculated by K. radicincitans) were moved forward for hybridization at Second Genome, Inc. PhyloChip Control Mix™ was added to each amplified product. Labeled bacterial products were fragmented, biotin labeled and hybridized to the PhyloChip™ Array, version G3 at Second Genome, Inc. Each scan was captured using standard Affymetrix software (GeneChip® Microarray Analysis Suite). Data analysis was performed using Second Genome's PhyCA-Stats™ Analysis Software (http://www.secondgenome.com/platform/services/phylochip/).
CLSM and TEM microscopy
S. lycopersicum cv. “Micro-Tom” were raised as in vitro cultures. Young seedlings were inoculated with GFP-labeled K. radicincitans as described in Witzel et al. (2017). Bacterial root colonization was recorded with a Zeiss LSM 510 META laser scanning confocal microscope (Carl Zeiss Jena GmbH). Bacterial eGFP fluorescence signals were captured using argon laser excitation at 488 nm (BP505-550 180 filter, Plan Apo 63/1.4 oil lens), and root images were captured using bright field settings.
Conventional negative contrast staining was used for electron microscopic investigations. Cells of K. radicincitans DSM 16656T were taken from semi solid (0.5%) agar and placed in 0.05 M saline solution. One drop of this bacterial solution was applied to Pioloform-carbon-coated, 400-mesh copper grids (Plano GmbH) for 10 min, fixed with 2.5% aqueous glutaraldehyde solution for 1 min, stained with 2.5% uranyl acetate solution for 1 min, and examined by transmission electron microscopy using a JEM-1400 Plus (JEOL) at an acceleration voltage of 120 kV.
Results
The genome annotation of DSM 16656T revealed one chromosome, two plasmids and a high percentage of genes involved in plant growth-promotion
Kosakonia radicincitans type strain DSM 16656T is an enteric bacterium that was shown to promote plant growth (Supplementary Table S2). Exhibiting two scaffolds and no plasmid sequence the preexisting draft genome of DSM 16656T was incomplete. Here we provide the fully sequenced and annotated genome, one chromosome (5,817,639 bp) and two plasmids (290,108 bp and 14,706 bp) of DSM 16656T (GenBank: CP018016.1, CP018017.1, CP018018.1) generated by combined PacBio and Illumina HiSeq sequencing. Apart from a rather small fraction of 6.3% (= 370 genes) biological functions could be assigned to all remaining of the 5,839 coding sequences (CDS) of DSM 16656T combining results from BlastKOALA (KEGG), Interpro (GO, protein domains) and PROKKA (note that there are only 5,827 genes and 5,712 CDS in the NCBI annotation; (Supplementary Table S3). Almost one third (32.9% = 1,800 genes) of all annotated CDS of DSM 16656T are involved in environmental information processing, 288 genes (= 5.3%) code for secretion systems and 214 genes (= 3.9%) for proteins involved in cell motility. A category summarizing PGP genes does not exist in the applied functional classification tools (RAST from SEED and BlastKOALA from KEGG), but summing up the CDS allocated to PGP we found 243 genes (= 4.4% of the genome) of K. radicincitans DSM 16656T (Table 1). Throughout the article we compare functional classification and genomic composition of K. radicincitans to three other groups of enteric PGPB in order to distinguish shared features from unique features of the K. radicincitans species and the type strain DSM 16656. The following paragraph describes how these groups have been defined.
Table 1
| Symbol | Traits | Genes |
|---|---|---|
| a | Nitrogen fixation and metabolism | amtB; anfA, anfD, anfF*, anfGH, anfK, anfO; narB, narG(2), narHIJKL, narX; nasR, nasD; nifAB, nifDEF, nifHIJKLMN, nifQ, nifSTUVWX, nifZ; nirB, nirD; nrtABC |
| b | Phosphate solubilization | phoAB, phoC(2) phoE, phoH, phoR, phoU; pqqE-like; pstABC, pstS(2) |
| c | IAA production | ipdC |
| d | GABA production | gabD(3), puuE |
| e | ACC deaminase | dcyD |
| f | Acetoin and butanodiol synthersis | als; budAB, budC(2); poxB |
| g | H2S production | cysCD, cysG(2), cysHI, cysJ(2), cysN |
| h | Trehalose metabolism | sugBC; otsAB; treA(2), treB, treRS, treYZ |
| i | Catalase | katB, katE, katG, katN |
| j | Peroxidases | Peroxidase gene homolog (34) |
| k | Superoxide dismutase | sodABC |
| l | Quorum sensing | lsrB(2), IsrD, IsrFG, IsrK; luxS(2) |
| m | Heat shock proteins | dnaJ, dnaK(2); groE |
| n | Cold shock proteins | cspA(5), cspE, cspG, cspJ |
| o | Glycine-betaine production | opuAB, opuAC, opuCA(2), opuCB(4), opuCC(2); osmY(2); proV(2), proWX |
| p | Effective rhizosphere colonizer | xerC(6) |
| q | Siderophore production | acrB(3); besA; btr(3); efeB, efeO, efeU; entA(2), entBC, entEF, entH, entS(2); exbB(3), exbD(3); feoABC; fepA(2), fepB, fepDE, fepG; fes; fhuA(3), fhuB(3), fhuC(3), fhuD(3), fhuEF; fptA; mbtH-like; mdtA(2), mdtB(2), mdtC(2); mexA(2), mexB; tonB; yusV(2) |
| r | Pyocin | prtN |
| s | Phenazone production | yddE(2) |
| t | 4-hydroxybenzoate production | ubiC |
| u | Chitinase production | chiA1(2), Chitinase gene homolog(2) |
Genes of Kosakonia radicincitans DSM 16656T involved in plant growth-promotion according to literature on homologous genes of other bacteria.
Gene within the anf gene cluster encoding a flavin nucleotide-binding protein similar to flavotoxin-encoding nifF.
Phylogenetic studies allowed a preselection of other enteric PGPB for comparative analyses
Multiple copies of 16S rRNA genes were found in fully sequenced K. radicincitans and closely related K. oryzae strains: DSM 16656T (seven copies), Ola 51T (five copies), GXGL-4A (seven copies), YD4 (seven copies), UMEnt01/12 (two copies). The 16S rRNA “main” (or consensus) gene sequence—represented by the most identical or very similar copies (Supplementary Figure S1A)—was chosen for phylogenetic analyses. A phylogeny of Kosakonia and closely related strains was based on these main sequences; only for K. radicincitans DSM 16656T all copies have been considered (Figure 1A). Our phylogenetic analyses of 16S rRNA gene sequences based on 70 bacterial strains reveal the presence of two distinct clades of PGP Kosakonia. The first (Figure 1A, depicted in red) comprises K. radicincitans, K. oryzae, K. oryziphila, and K. arachidis, the second (Figure 1A, depicted in yellow) comprises K. sacchari, K. oryzae, and K. pseudosacchari. Note that Kosakonia strains named K. oryzae do not form a monophyletic group. A third Kosakonia clade comprises Enterobacter sp. FY-07 and E. oryzendophyticus REICA 082, more recently named Kosakonia oryzendophytica. Kosakonia cowanii is outgroup to these three Kosakonia groups (Figure 1A). A more comprehensive 16S rRNA gene tree based on 195 bacterial strains (Supplementary Figure S2) and a phylogeny based on concatenated amino acid sequences of reference genes atpD, gyrB, infB, and recA (Supplementary Figure S3) show similar affiliations. In the following analyses we focus on the two large Kosakonia subgroups that were shown to contain many PGPB, and we refer to them as the K. radicincitans group (Figure 1, KORA clade, red) and the K. sacchari group (Figure 1, KOSA clade, yellow). Sharing >97% 16S rRNA gene sequence identity with K. radicincitans, strains of closely related Klebsiella variicola and Enterobacter sp. (both family Enterobacteriaceae) were also included in the analysis. The selected strains were shown to be naturally associated with crop plants (Figure 1C) and capable of promoting plant growth. Here we refer to the Klebsiella variicola group (Figure 1, KLEVA clade, blue) and the Enterobacter group (Figure 1, ENTERO clade, green). A detailed list of Kosakonia strains (including accession, reference, location and 16S rRNA gene sequence identity to DSM 16656T) and the selected Enterobacter and Klebsiella strains is provided in Supplementary Table S1. Additionally, data on a few Citrobacter strains of particular interest (s. below) is given in this table.
Figure 1
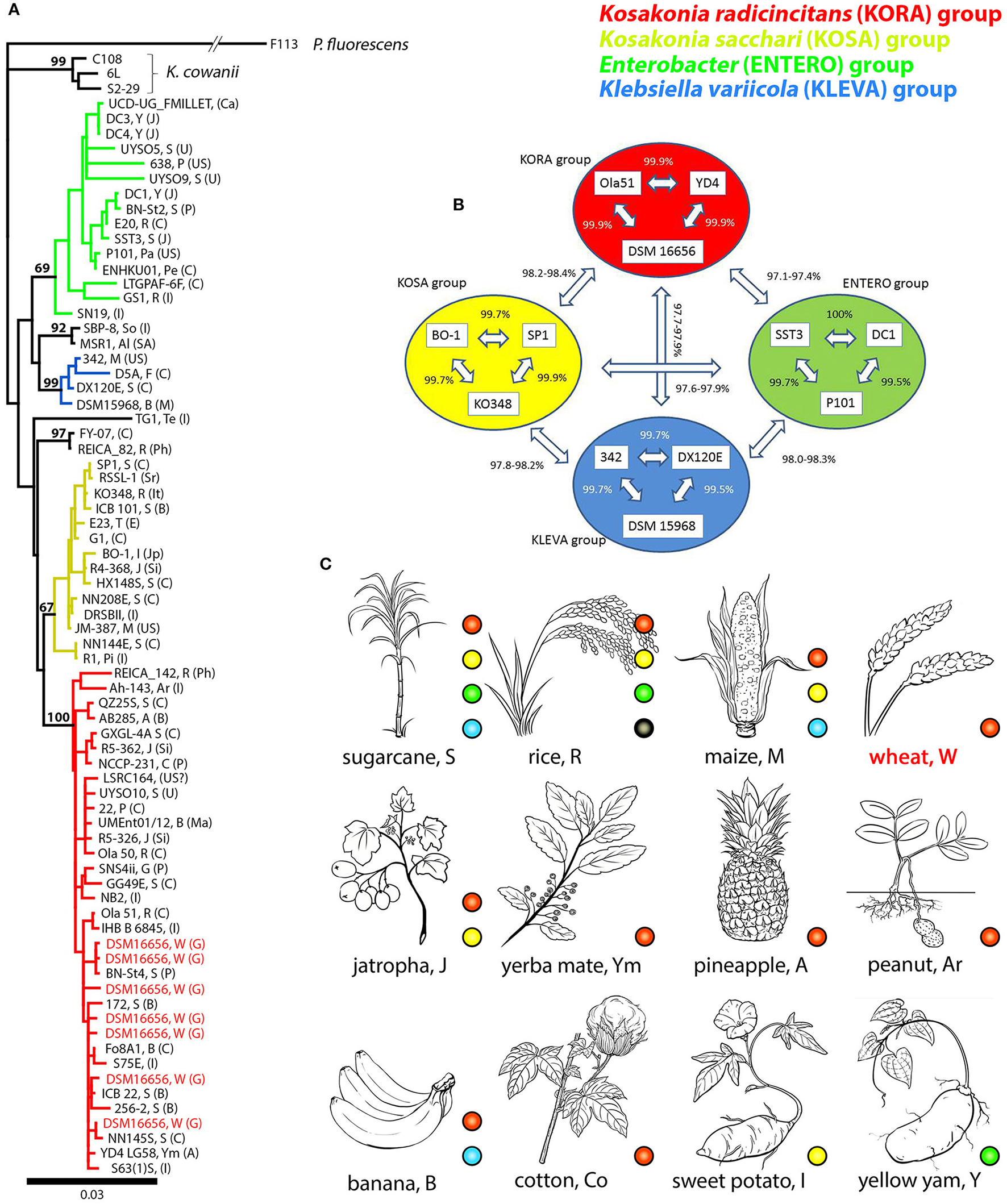
Phylogeny, selection of reference strains and plant hosts of PGP Kosakonia and closely related bacterial species; Four major groups are considered and depicted in different colors: red, Kosakonia radicincitans (KORA) group; yellow, Kosakonia sacchari (KOSA) group; green, Enterobacter (ENTERO) group; blue, Klebsiella variicola (KLEVA) group. Capital letters in (A,C) refer to plant hosts (Supplementary Table S1). (A) 16S rRNA gene tree of Kosakonia strains and a selection of Enterobacter and Klebsiella strains closely related to Kosakonia. Outgroup is Pseudomonas fluorescens. Three representatives of putatively pathogenic Kosakonia cowanii are shown. All 16S rRNA gene copies of DSM16656T (= type strain of Kosakonia radicincitans) are considered, but only a single (the most prominent) copy of other strains. Capital letters in brackets represent sampling site countries (Supplementary Table S1). (B) Selection of 12 bacterial strains from gene tree shown in (A); (C) Important crop plants hosting enteric PGPB depicted in (A); only hosts of published strains are shown. Black dots and branches represent SBP-8, MSR1, TG1, and REICA-082, the strains nested in between the four depicted major groups of Kosakonia/Enterobacter/Klebsiella.
Functional classification of four subgroups of enteric PGPB show large fraction of Kosakonia radicincitans genes involved in motility and chemotaxis
Using RAST-annotated genomes we studied the functional classification of DSM 16656T and other enteric PGPB (Figure 2). We compared the four robust groups of enteric PGPB, prior determined by 16S rRNA gene sequence analyses. We chose three representative strains per group [(i) KORA (DSM 16656T, YD4, Ola 51), (ii) KOSA (KO348, SP1, BO-1), (iii) ENTERO (DC1, SST3, P101), and (iv) KLEVA (342, DX120E, DSM 15968)]. 16S rRNA gene identities among selected representatives of each clade are 99.5–100% and between clades 97.1–98.4% (Figure 1B). Although the overall pattern of functional classification is very similar in DSM 16656T and the four investigated groups of enteric PGPB (Figures 2A,B), there is one striking difference (highlighted in yellow). The KORA group has more genes for motility and chemotaxis than the three other groups, particularly more than the KLEVA group (Figure 2C). All three K. radicincitans strains analyzed have an equally high number of genes involved in motility and chemotaxis, and all three Klebsiella strains an equally low number of these genes, but members of the KORA group and the ENTERO group show higher variability in the number of genes allocated to motility and chemotaxis (as visualized by error bars). A detailed genome map of DSM 16656T is provided in Supplementary Figure S4, and the number of genes per functional category for all representative strains is given in Supplementary Figure S5.
Figure 2
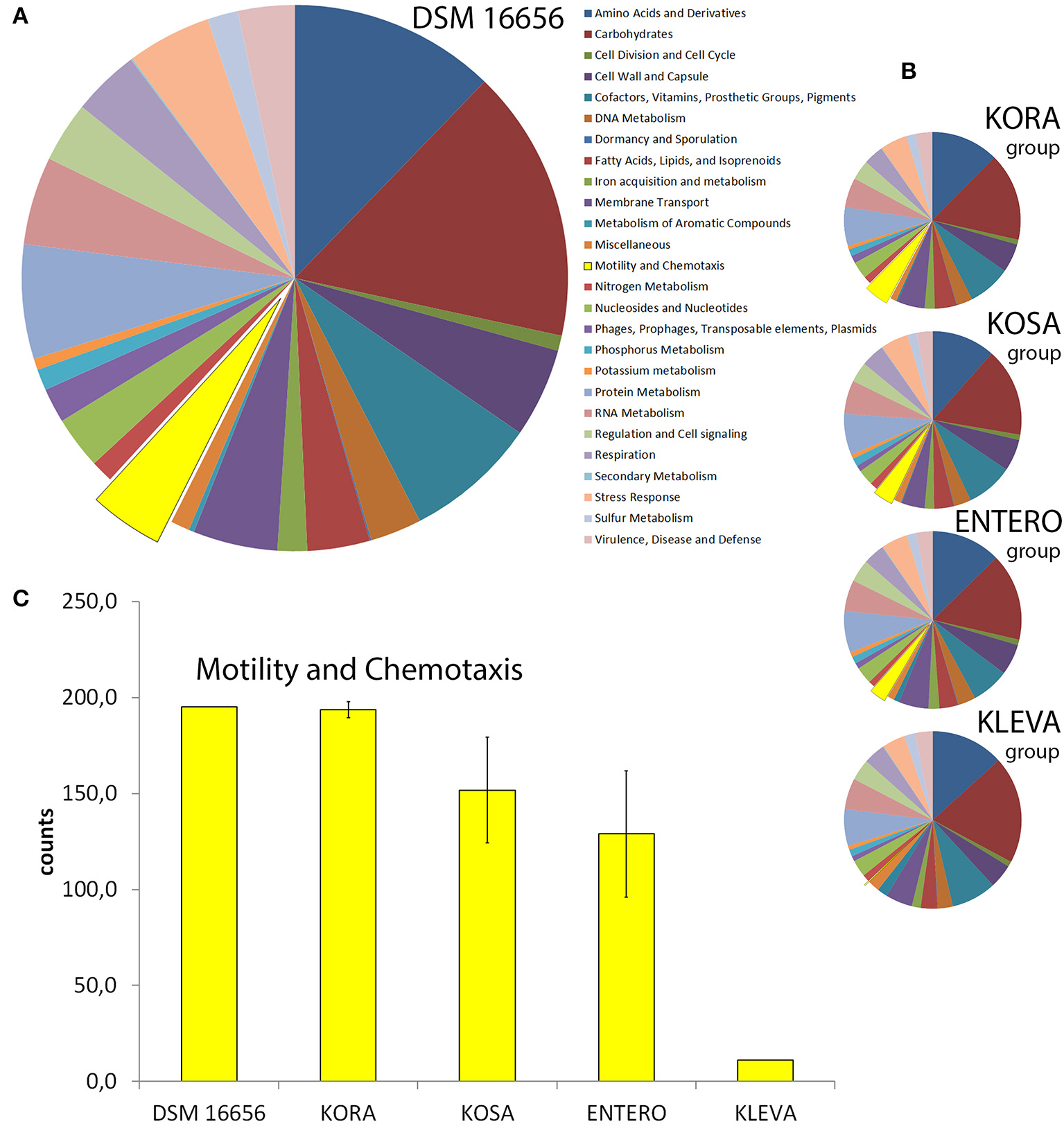
Functional classification of DSM 16656T, Kosakonia radicincitans (KORA group), K. sacchari (KOSA group), Enterobacter spp. (ENTERO group) and Klebsiella variicola (KLEVA group) by SEED using RAST-annotated genomes. (A) DSM 16656, (B) each pie chart represents the average of a group of three bacterial strains (KORA, KOSA, ENTERO, and KLEVA group). The size of a section corresponds to the number of counts for the functional category; a single gene may have multiple counts when present in multiple sub-categories; motility and chemotaxis are depicted in yellow. (C) Extraction from (A) and (B); error bars show standard deviation of the three strains.
Comparison of core genomes simplifies the search for Kosakonia-specific characteristics as well as features shared between enteric PGPB
Considering the same 12 strains that were used for the abovementioned functional classification we determined the core genome for each group of three enteric PGP strains (KORA, KOSA, ENTERO, KLEVA). We compared these core genomes and the genome of a Rhizobium outgroup (R. leguminosarum bv. viciae 3841) to DSM 16656T (Figure 3). The purpose of this fast and easy comparison was to simplify the search for (i) genomic features that are shared between DSM 16656T and closely related enteric PGPB, (ii) genomic features that are unique to Kosakonia radicincitans, and (iii) genomic features that are unique to DSM 16656T. Although K. radicincitans strains from Germany (DSM 16656T), China (Ola 51T) and Argentina (YD4) share 99.9% of their 16S rRNA gene sequence (Figure 1B), genomes of these strains exhibited significant rearrangements, with approximately half of the gene order conserved, as depicted in circle C3 of Figure 3. White areas in Figure 3 ranging centripetally from circle C3 to circle C12 represent genomic regions of DSM 16656T lacking in all four core genomes and Rhizobium. Seven of these regions are mobile genetic elements (MGEs), of which six have been identified as phage regions by PHAST software (Zhou et al., 2011; light blue numbers in Figure 3A), and another region is dominated by genes defining integrative conjugative elements (ICEs). One of the MGEs, phage region 2, consisting of 21 phage and six hypothetical genes, is different since occurring in both Kosakonia core genomes (KORA and KOSA), and its gene order is conserved among the three strains of K. radicincitans (Figure 3A, circles C3 and C5), which may indicate an important function.
Figure 3
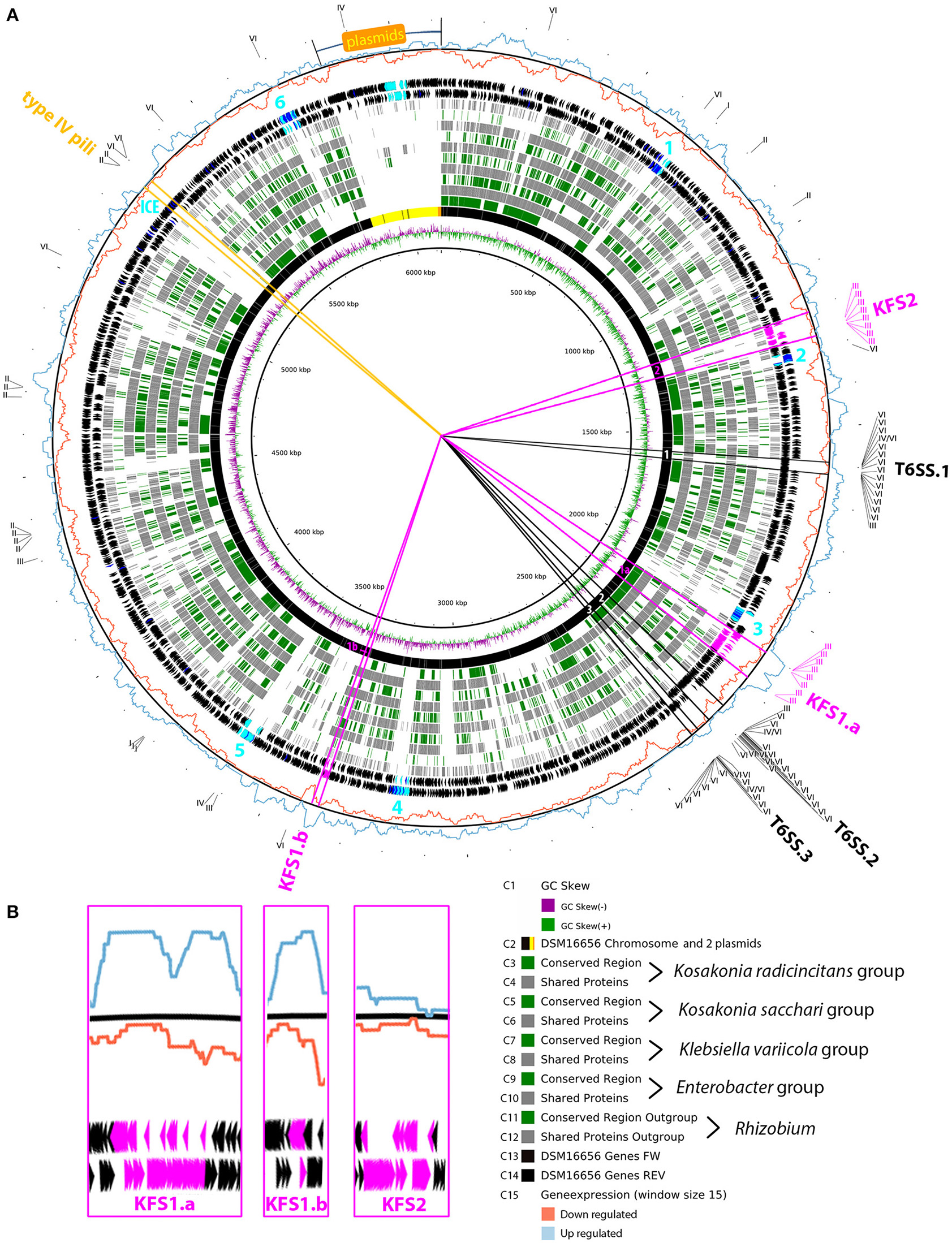
Core genome comparison. (A) Comparison of the Kosakonia radicincitans DSM 16656T genome (including plasmid sequences highlighted by yellow/orange area in circle C2) to core genomes of other enteric PGPB (KORA, KOSA, ENTERO and KLEVA group). Gray lines and boxes in circles C4, C6, C8, C10, C12 point to the presence of genes (according to NCBI BLASTp searches) shared among the three strains of a core genome; green lines and boxes in circles C3, C5, C7, C9, C11 represent collinear units of nucleotide sequences determined by MAUVE alignments, i.e., conserved gene clusters in which the order of genes is equal to DSM 16656T. Light blue genes depicted in circles C13 and C14 belong to six phage regions detected by PHAST software, and are highlighted by light blue numbers 1–6; dark blue genes have phage-related functions according to our own annotation pipeline. The ICE region is an integrative conjugative element. Pink lines demarcate genes of flagellar systems (KFS1 and KFS2), black lines genes of type VI secretion systems (T6SSs), and orange lines genes of type IV pili. Due to the homology of the flagellar basal body and the transmembrane export complex of injectisomes, eight motility genes per flagellar system were annotated to type three secretion systems (T3SSs). The outermost circle shows the differential expression of the transcriptome of DSM 16656T when exposed to tomato root exudates, captured by a sliding window of 15 consecutive genes; blue graph, up-regulated genes; red graph, down-regulated genes. (B) Magnification of flagellar system genes depicted in (A) highlighting the different expression of KFS1 and KFS2.
Species-specific genomic features: K. radicincitans has two flagellar systems and three type VI secretion systems
The genome of DSM 16656T carries genes of five types of secretion systems (I, II, III, IV, and VI), particularly of type III (T3SS) and VI (T6SS), indicating that K. radicincitans has evolved an arsenal of gene systems for environmental interaction. Whereas, T6SSs encode injectisomes, which export effector proteins to the extracellular milieu, eukaryotic cells or other bacteria, genes allocated to T3SS are not necessarily coding for injectisomes, but may contribute to flagella biosynthesis. In K. radicincitans DSM 16656T genes of T3SS cluster together with chemotaxis genes and all other genes required for flagella biosynthesis, suggesting that this strain uses “T3SS” genes for motility purposes. Genomic regions encoding important secretion systems are highlighted in Figure 3. The core genome comparison nicely illustrates that the KORA group — but none of the other groups — possess the second flagellar system (KFS2, Kosakonia Flagellar System 2). KFS2 of strain DSM 16656T is a compact gene cluster comprising — apart from five genes — only essential flagellar genes; the other flagellar system (KFS1) is divided into two gene clusters, KFS1.a and KFS1.b, with 1,148 genes in between. The comparison of core genomes (Figure 3A) shows that most genes of KFS1.a and KFS1.b are shared between the KORA, the KOSA and the ENTERO group, but missing in the KLEVA group, an observation that matches common knowledge that Klebsiella is a non-motile bacterium.
The order of the 71 genes constituting KFS1.a is highly conserved in K. radicincitans, but the order of 55 genes constituting KFS2 varies between strains of this species (Figure 3A, circle C3) probably indicating that KFS2 is rapidly evolving in K. radicincitans. The conserved Flg22 motif of the flagellin molecule, and particularly its C-terminal Flg15 segment, may induce MAMP (microbe-associated molecular pattern) triggered immunity. The Flg15 peptide sequence of K. radicincitans KFS1 and E. coli are identical and differ from Pseudomonas aeruginosa in two of the four C-terminal residues (Supplementary Figure S6). The Flg15 peptide sequence of K. radicincitans KFS2 has an additional AA substitution at the very last position of the C terminus.
Next to KFS2 there is another duplicated secretion system in the KORA group, which is missing in other enteric bacteria: an additional type VI secretion system (T6SS.3). The two T6SS clusters commonly found in Enterobacter, also occur in K. radicincitans, here called T6SS.1 and T6SS.2. However, the T6SS.3 of the KORA group is new to enteric bacteria. Genes of T6SS.3 are completely missing in the KOSA, ENTERO and KLEVA group and also in the Rhizobium outgroup. The order of T6SS.3 genes in the KORA group is highly conserved (Figure 3).
Pairwise genome comparisons confirmed assumptions based on core genome analysis
The comparison of core genomes of PGPB was helpful for rapidly distinguishing genes that DSM16656T shares with different groups of enteric bacteria and genes putatively unique to the KORA group, but whether these candidate genes are indeed restricted to K. radicincitans had to be confirmed by a thorough analysis applying a larger number of bacterial genomes (Figure 4A). We compared the DSM 16656T genome sequence to 31 closely related and three distantly related, fully sequenced and annotated bacterial genomes listed in the Material and Method section. From hundreds of Klebsiella and Enterobacter strains available at NCBI the given ones were chosen due to their endophytic lifestyle and their PGP potential as described in literature (Supplementary Table S1).
Figure 4
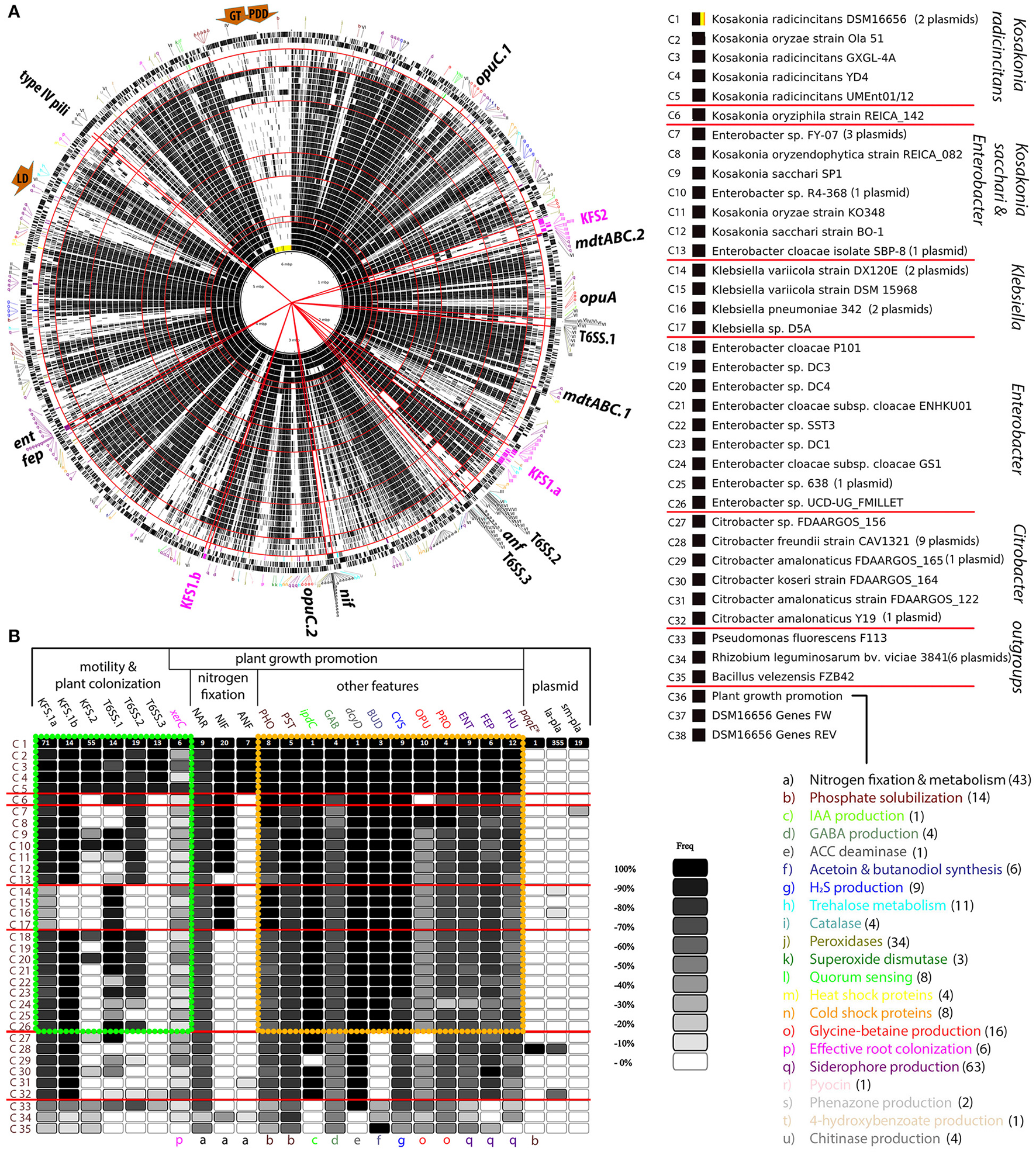
Genome comparison of Kosakonia radicincitans DSM 16656T to 31 other strains of enteric bacteria and three strains of non-enteric bacteria. (A) Comparative genome map including plasmid sequences (highlighted by yellow and orange area in circle C1). Circles C1 to C35 depict each one bacterial strain; black lines within a circle represent genes. The order of strains is given by 16S rRNA gene sequence identities to DSM 16656T, descending from highest to lowest. Circle C36 depicts genes supposed to be involved in plant growth-promotion (PGP); lowercase letters in outermost circle refer to particular PGP functions (Table 1). Red circular lines separate phylogenetic groups. Red straight lines demarcate gene clusters of particular interest, such as PGP genes, type VI secretion systems, type IV pili, and flagellar systems. (B) Selection from (A) depicting important loci and gene clusters with significant impact on plant growth and plant colonization. Each box represents a gene count per locus for a particular strain compared to DSM 16656T. A locus may consist of 1–71 coding sequences in DSM 16656T or entire plasmids with up to 355 coding sequences (la-pla, large plasmid; sm-pla, small plasmid); black boxes represent the maximum count (100%) and white boxes the minimum count (< 10%) of genes found in DSM 16656T, gray shadings represent percentages in between. The exact number of genes per box is given in Supplementary Table S5. The yellow rectangle frames some of the most significant PGP genes of Kosakonia, Enterobacter and Klebsiella, the green rectangle frames genes involved in motility and plant colonization. Dark brown arrows, exploitation of unusual carbon sources (LD, lichenan degradation; GT, glycerol transformation; PDD, 1,2-propanediol degradation).
The genome comparison of the 35 individual strains of bacteria confirms assumptions based on the core genome analysis: T6SS.3 appears to be restricted to taxa of the KORA group; all five strains of this group included in the genome comparison share the corresponding genes (Figure 4B). BLASTp searches to NCBI databases support this result, yielding very low gene sequence similarities to DSM 16656T of strains other than K. radicincitans (average identity for all T6SS.3 genes < 68.5%, Supplementary Table S4).
Similar to the K. radicincitans-specific flagellar system (KFS2), T6SS.3 is reduced to the essential core component genes required for T6SS biosynthesis, regulation and functioning (Supplementary Figure S7). The only exception is a gene encoding a T6SS accessory protein of unknown function, here called tagK1. While K. radicincitans strains share 95% amino acid sequence identity of tagK1, the next best hit outside this species shares only 55% sequence identity with DSM 16656T (Supplementary Table S4). The genomic neighborhood of K. radicincitans T6SS.3 harbors genes for multiple antibiotic resistance (marR), multidrug resistance (mdtN, emrB), and resistance to oxidative stress (marR, katG; Supplementary Figure S7). Remarkably, Pseudomonas fluorescens F113 shares more T6SS.3 genes with K. radicincitans than any Kosakonia sacchari, Klebsiella, or Enterobacter strain included in our comparative genome analysis (Figure 4B).
Just like T6SS.3, KFS2 is shared by all members of the KORA group but in contrast to T6SS.3 it is not absent from all the other groups of enteric bacteria. Two Enterobacter strains (P101 and DC4) and one strain from the K. sacchari group (R4-368) possess KFS2; K. oryzae SP1 has half of KFS2 (Figure 4).
Shared genomic features: K. radicincitans and other species of enteric PGPB share the bulk of genes involved in plant growth-promotion
Apart from flagellar systems and type VI secretion systems our analysis focused on genes reported to be involved in PGP: most notably genes for atmospheric nitrogen fixation, siderophore production, phosphate solubilization and glycine-betaine production. The corresponding genes are listed in Table 1 and depicted by lowercase letters in Figure 4 and Supplementary Figure S4. Important regions of the K. radicincitans DSM 16656T genome are indicated by straight red lines and arrows (Figure 4A). The exact numbers of homologous genes illustrated in Figure 4B are given in Supplementary Table S5.
The bulk of genes encoding PGP traits are shared between Kosakonia (both large groups), Enterobacter and Klebsiella (framed by a yellow rectangle in Figure 4B), but a significant fraction of genes required for atmospheric nitrogen fixation and for protecting host plants from osmotic stresses via glycine-betaine (GB) production is unique to K. radicincitans or shared with only a subset of other strains. GB production requires the uptake of GB precursor compounds via osmoprotectant uptake systems (Opu). There are three opu-gene clusters in K. radicincitans (Figure 4): opuC.1 is shared with the K. sacchari group and the Klebsiella group but missing in most strains of Enterobacter and Citrobacter, opuC.2 in contrast is shared with Enterobacter and Citrobacter but missing in the K. sacchari and the Klebsiella group, and opuA is restricted to K. radicincitans and Rhizobium.
Although atmospheric nitrogen fixation is an extremely energy-consuming process, K. radicincitans has two gene clusters for this purpose, the nif regulon and anf operon. The nif regulon is shared with taxa from the K. sacchari group, the K. variicola group and Rhizobium, but missing in taxa from the Enterobacter group (Figure 4). The anf operon is not found in any other species included in the analysis but is known from distantly related taxa.
Next to atmospheric nitrogen fixation, phosphate solubilizing activity is supposed to be one of the major advantages provided by PGPB. Most genes for phosphorus assimilation (pho, pst) and siderophore production/transport are shared among enteric PGPB (Figure 4). However, while K. radicincitans shares the largest gene cluster for siderophore production (ent/fep) and an mdtABC operon (mdtABC.1) with almost all other strains included in the analysis, other genes for siderophore production are unique to K. radicincitans, such as a duplicated mdtABC operon (mdtABC.2) next to KFS.2 (Figure 4).
Taken together, K. radicincitans DSM 16656T is well equipped with genes attributed to PGP and differs from many other enteric bacteria predominantly by the presence of its gene clusters (nif and particularly anf) for atmospheric nitrogen fixation.
Strain-specific genomic features of K. radicincitans DSM 16656T: most large plasmid genes are shared with Citrobacter but absent from other Kosakonia strains
Our new genome sequencing approach of K. radicincitans DSM 16656T revealed the presence of one chromosome and two plasmids (yellow and orange highlighted area in Figures 3A, 4A; for simplification purposes plasmids were included as part of the circular genome map). The comparative genomics approach showed that DSM 16656T shares its small plasmid with the closely related Enterobacter sp. FY-07, a cellulose producing strain from China, and its large plasmid with the distantly related Citrobacter freundii CAV1321 (USA) and Citrobacter amalonaticus Y19 (South Korea; Figures 4B, 5). Genes of CAV1321 and Y19 appeared also as best hits when blasting large plasmid genes of DSM 16656T to NCBI databases. The small plasmid of DSM 16656T carries 19 genes, the large plasmid 359 genes, of which 355 are coding sequences (Supplementary Table S3). A notable fraction (38%) of large plasmid genes are unique to the large plasmid, i.e., missing on the chromosome and the small plasmid of DSM 16656T. Among the 57 unique genes of the large plasmid that could be assigned to KEGG pathways 39% refer to microbial metabolism in diverse environments, indicating that the large plasmid increases the capability of DSM 16656T to cope with different habitats (Supplementary Table S6). Remarkably, the large plasmid is missing from all other fully sequenced Kosakonia genomes, and from representative Klebsiella and Enterobacter strains included in the analysis (Figure 4) suggesting that the plasmid may have been transferred from Citrobacter to DSM 16656T. 145 of the 355 coding sequences of the large plasmid of DSM 16656T are shared exclusively with Citrobacter and missing in other Kosakonia strains. The contribution of the presumably Citrobacter-derived plasmid to the PGP potential of DSM 16656T appears to be low as measured by the usual PGP repertoire (Table 1), since all genes allocated to PGP occur on the chromosome, except a single (pqqE-like) gene from the large plasmid (Figure 4B).
Figure 5
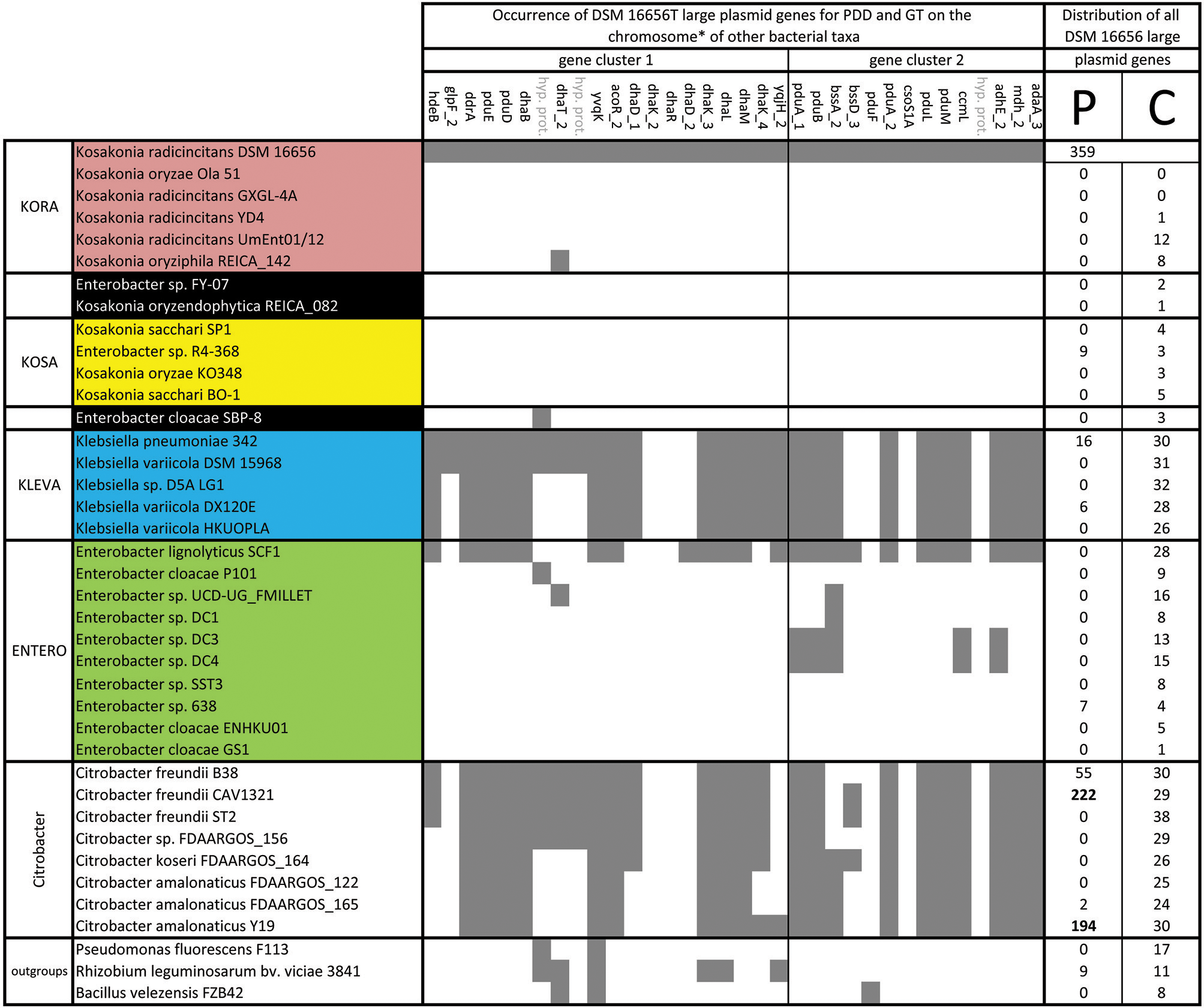
Occurrence and distribution of large plasmid genes of K. radicincitans DSM 16656T in chromosomes and plasmids of other bacterial taxa; GT, glycerol transformation; PDD, 1,2-propanediol degradation; P, plasmid; C, chromosome; *none of these PDD and GT genes occur on plasmids in the other investigated taxa.
The large plasmid of DSM 16656T carries many genes for exploiting plant-derived carbon sources
Although 14% of all KEGG identified genes of the large plasmid encode ABC transporters for nutrient uptake (e.g., mineral and carbohydrate transporters), this percentage is low compared to chromosomal genes, where 28% code for ABC transporters (Supplementary Table S6). Consistently, a good portion of the ABC transporter spectrum is covered by the chromosome of DSM 16656T and only a single additional gene (troA) is contributed by the plasmids. However, the fraction of genes involved in propanoate and glycerolipid metabolism is 5-fold higher in the large plasmid (Supplementary Table S6) indicating that plasmid genes may play a significant role in exploiting plant-derived carbon sources: the presence of pdu genes (pduABDEFLMPQ) for propanediol degradation (PDD) and dha genes (dhaBDKLMT) for glycerol transformation (GT) on the large plasmid (Figure 4) suggests that DSM 16656T has the potential to anaerobically degrade rhamnose and fucose in plant cell walls and glycerol in plant lipids. Genes for PDD and GT are shared between DSM 16656T, Citrobacter and Klebsiella (Figure 4A), but are missing from other Kosakonia strains and Enterobacter (with the exception of E. lignolyticus SCF1; Figure 5). Although these genes constitute a prominent feature in Citrobacter and Klebsiella, they are not located on plasmids of the investigated strains but on chromosomes (Figure 5).
DSM 16656T carries further genes for exploiting plant-derived carbon sources on the chromosome
Particular genes on the chromosome have further increased the potential of DSM 16656T to exploit plant-derived carbon sources. Genes for degradation of lichenan, a beta-1,3-1,4-glucan from cell walls of lichen and—in a slightly different form—from barley and oat, may be attributed to horizontal gene transfer from Bacillus to Kosakonia. Three of the genes required for lichenan degradation (licABR) are shared between Bacillus subtilis FZB42 and two Kosakonia strains, DSM 16656T and UMEnt01/12, but are missing in other Kosakonia, Enterobacter, Klebsiella, and Citrobacter strains.
The two flagellar systems of K. radicincitans DSM 16656T are operating in different environments
We assumed that the existence of two distinct flagellar systems (KFS1 and KFS2) in K. radicincitans reflects different purposes, i.e., the individual operation of each flagellar system in different environments. To test this hypothesis we designed a DSM 16656T-specific microarray and exposed bacterial cells from liquid culture to root exudates (C/N ratio of 9.39) of tomato, S. lycopersicum cv. “Vanessa”. The tomato cultivar “Vanessa” was shown to respond to DSM 16656T by increased growth and yield (Berger et al., 2013, 2017). In this experiment we simulated the condition, in which the flagellum for swimming toward host would be required. Determining the number of differentially expressed genes in a sliding window of 15 consecutive coding sequences (CDS) throughout the genome we searched for highly enriched sets of differentially expressed genes, depicted in the outermost circle of Figure 3. Applying a p-value of 0.05 and a FC threshold of 1.5, a fraction of 52% of motility and chemotaxis genes (111 of 214 CDS) was differentially expressed. However, while nearly the same number of all differentially expressed CDS was up- (1,197) and down-regulated (1,135), most of the differentially expressed chemotaxis and motility genes, namely 86%, were up-regulated (83 genes up vs. 13 genes down), clearly suggesting that DSM 16656T responded by increased motility to root exudate exposure (Supplementary Table S3). Most importantly, we found the expression of genes of KFS1 but not of KFS2 to be differentially expressed (Figure 3B). Although being separated by more than a thousand genes, expression of both clusters of KFS1 was up-regulated (KFS1.a up to 22.3-fold, KFS1.b up to 29.4-fold). In contrast, expression of genes encoding the master regulator of KFS2 was significantly down-regulated (flhC 23.9-fold and flhD 26.8-fold). Hence, we concluded that KFS1 but not KFS2 is required for bacterial swimming toward the host. The active down-regulation of the KFS2 master regulator suggests that KFS2 biosynthesis has to be avoided in the tested environment.
K. radicincitans DSM 16656T alters the bacterial community composition of plants
To test our second hypothesis that the three distinct type VI secretion systems (T6SS.1-3) enable K. radicincitans to exert significant influence on its environment, e.g., by suppressing other bacteria competing for the same ecological niche, we performed a PhyloChip analysis. Two weeks after inoculating K. radicincitans DSM 16656T into leaves and roots of young tomato plants the bacterial and archaeal community composition was significantly affected (Figure 6). The presence of DSM 16656T was confirmed by qPCR; four days after inoculation the average concentration of DSM 16656T was 2.44E+07 cells per g fresh weight of leaf tissue and 2.88E+06 cells per g fresh weight of root tissue (Supplementary Figure S8). In total 1,220 operational taxonomic units (OTUs) could be detected, of which 78 (belonging to 33 families) showed significantly altered abundance in leaves (Figure 6A), and 122 (belonging to 76 families) showed significantly altered abundance in roots (Figure 6B). Performing Welch's t-test the number of significantly affected OTUs increased to 153 in leaves (13%) and 194 in roots (16%). Most of these OTUs displayed increased abundance in both leaf and root samples when inoculated with K. radicincitans, but the Betaproteobacteria clearly differed from the overall picture showing decreased abundance in leaves and roots. Interestingly, Alphaproteobacteria showed a decreased abundance in roots, but increased abundance in leaves.
Figure 6
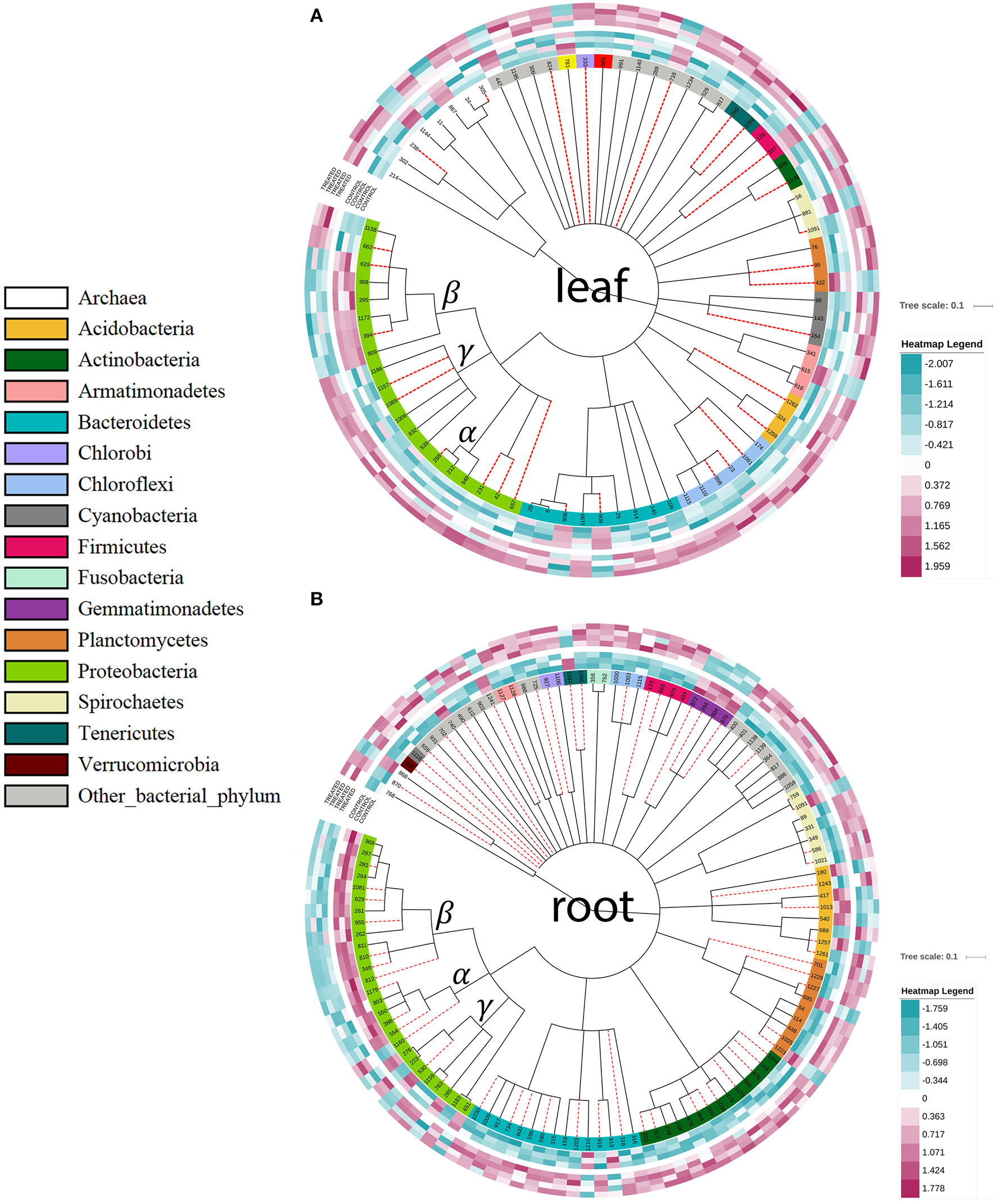
PhyloChip analysis of the bacterial community composition of tomato plants 14 dpi with K. radicincitans DSM 16656T. Operational taxonomic units (OTUs) with significantly changed abundance values; outermost circles represent the four replicates of tomato plants treated with K. radicincitans; inner circles represent the controls; purple colored boxes represent increased OTU abundance and blue colored boxes decreased OTU abundance. (A) leaf samples; (B) root samples. Red dashed lines, unclassified taxa; α, β, and γ, subgroups of Proteobacteria.
K. radicincitans DSM 16656T employs the same colonization pattern in different plant species
To determine the motility and plant colonization potential of DSM 16656T we performed microscopical analyses (Figure 7). Visualizing flagella of bacterial cells by transmission electron microscopy (TEM), K. radicincitans DSM 16656T was found to be peritrichously flagellated with flagella approximately 10 μm long (Figure 7B). For elucidating the places of plant colonization, we generated green fluorescent protein (eGFP) expressing mutants of DSM 16656T and studied zones of bacterial agglomeration on and inside roots of tomato (asterids group) and Arabidopsis thaliana (rosids group). DSM 16656T was able to colonize both plant species in the same manner: being initially attracted by root hairs (Figure 7C), it entered the plant by cracks surrounding newly emerging lateral roots (Figure 7D), and eventually colonized the interior of individual root parenchyma cells (Figure 7E, Supplementary Videos 1, 2). Although complete colonization of plant parenchyma cells was infrequently observed, it could nonetheless be documented in both the distantly related plant species. Strong expression of eGFP indicated that the bacteria were still metabolically active, i.e., not destructively affected by plant defense.
Figure 7
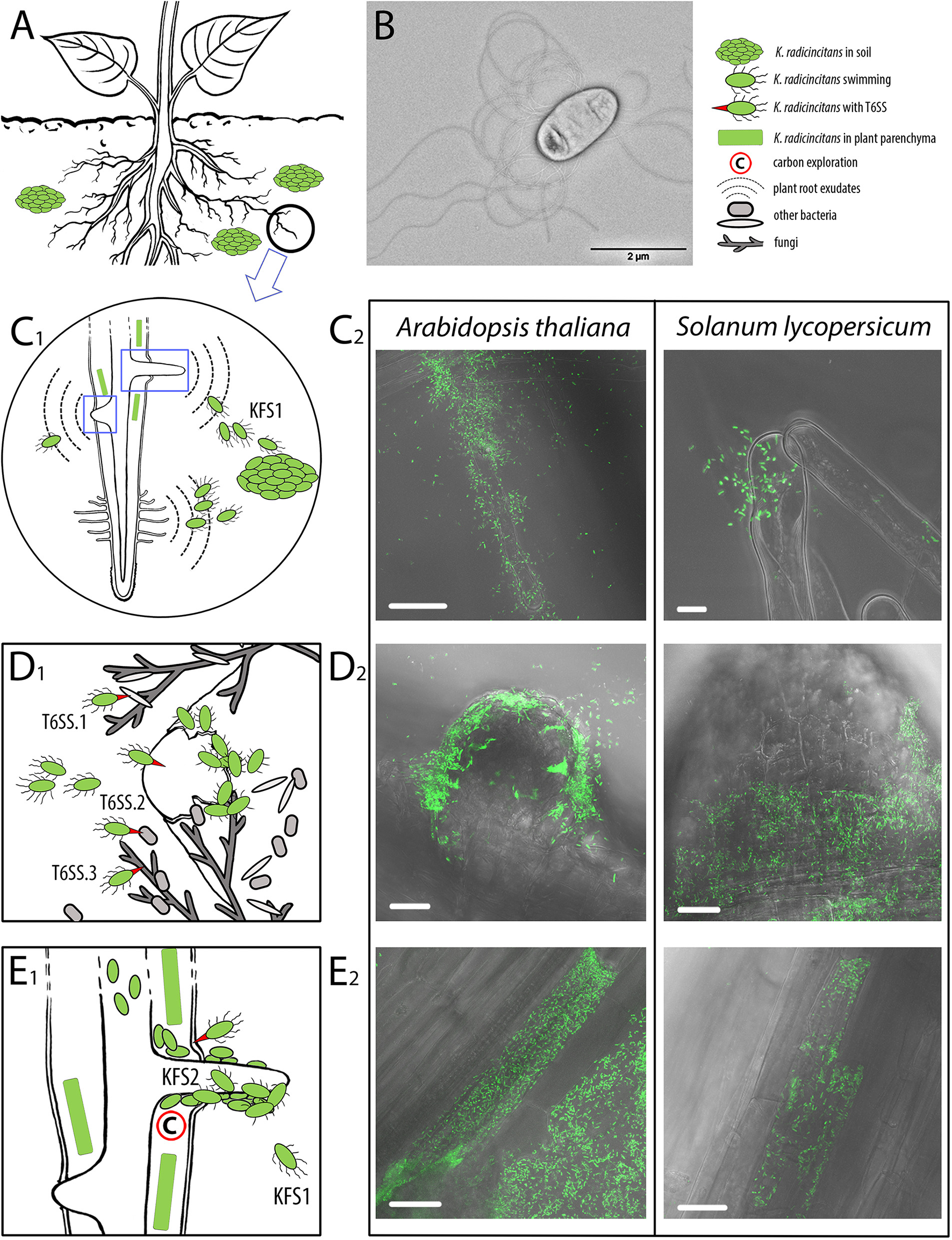
Plant root colonization scenario of Kosakonia radicincitans and supporting micrographs. (A) Supposed survival of Kosakonia radicincitans aggregates in the soil. (B) Transmission electron micrograph of peritrichously flagellated cell of K. radicincitans DSM 16656T cultivated on semi-solid agar for 24 h. (C1), (D1), and (E1). Schematic drawings illustrating the colonization steps of K. radicincitans. (C2), (D2), and (E2). Confocal laser scanning micrographs of (eGFP) expressing DSM 16656T colonizing roots of Arabidopsis thaliana and Solanum lycopersicum 6 dpi. (C) Plant roots exudate organic compounds that attract K. radicincitans toward root hairs and lateral roots. K. radicincitans has two flagellar systems (KFS1 and KFS2); when exposed to root exudates KFS1 of K. radicincitans enables high swimming motility and possibly attachment to plant surface. (D)K. radicincitans is particularly attracted by newly emerging lateral roots and colonizes intercellular spaces of root parenchyma via cracks around lateral roots; three distinct type VI secretion systems (T6SSs) help to interact with both plant host (evasion of plant immunity) and associated microbiota. (E) Migration into plant parenchyma cells supported by T6SSs injecting effector proteins. The scenario of K. radicincitans cells switching from KFS1 to KFS2 when attaching to plant surface and colonizing the roots is speculative; the purpose of KFS2 has not been determined yet. Note that proportions of plant, fungi and bacteria are not to scale. Green rectangle, root parenchyma cell densely colonized by K. radicincitans. White scale bar 20 μm.
Discussion
Phylogenetic and comparative genome analyses of Kosakonia
Our phylogenetic analyses confirm results of other working groups that there are two main clades of Kosakonia (Brady et al., 2013; Gu et al., 2014). Our comparative genome and functional classification analyses exhibit clade-specific features beyond 16S rRNA gene sequences and “housekeeping” genes. Two outstanding genomic features of the KORA (= Kosakonia radicincitans) clade conserved among strains of this species but missing in most other PGPB are an additional flagellar system (KFS2) and an additional type VI secretion system (T6SS.3). We considered the consistency of these two gene clusters in K. radicincitans to reflect important functions and asked the three following questions: (1) Why has K. radicincitans two flagellar systems? (2) Does K. radicincitans affect the natural microbiota of plants (possibly via T6SSs)? (3) How and where is K. radicincitans colonizing the plant?
Genomic features of PGPB adaptation to plants
The PGP capability of bacteria is supposed to be conferred by genes involved in e.g., nitrogen fixation, mineral nutrient solubilization or phytohormone production. Although some of the underlying gene clusters have been studied in detail, the purpose of others, such as the anf operon, is still puzzling (Yang et al., 2014). On the other hand can some capabilities of bacteria not be explained by the presence or absence of genes per se. The ability to solubilize rock-phosphate or calcium-phosphate has been attributed to the pqq operon, which is missing in DSM 16656T, with the exception of a pqqE-like gene on the large plasmid (Figure 4B). While the missing pqq operon would suggest a limited ability of this strain to solubilize rock phosphates, recent experiments have proven that DSM 16656T is capable of solubilizing an entire range of phosphates such as Ca3(PO4)2, Fe3(PO4)2, AlPO4 and even hydroxyapatite (Supplementary Figure S9). It will be interesting to see whether other genes have adopted the role of missing pqq genes in DSM 16656T and whether they form a functional unit together with pqqE from the large plasmid.
In order to tap their PGP potential bacteria need to be in contact with the plant. For plant-associated bacteria the rapid colonization of the plant is crucial for establishing a metabolically active population, i.e., a population of sufficient “quorum sensing density” (Hartmann et al., 2014). The requirement of bacterial chemotactic motility for establishing a bacterial-plant association is well-known and was shown long ago (Vande Broek et al., 1998). Many bacterial strains of the family Enterobacteriaceae were reported to be (i) rapidly moving (Wolf et al., 2015) and (ii) plant growth-promoting (Lin et al., 2012), tempting us to speculate that soil-borne enteric PGPB may rely on exceptional motility in order to approach and colonize plants before other bacteria do. Colonization of the host is preceded by swimming toward host attractants (root exudates) using powerful flagella. Arriving at the plant surface some bacteria use pili to promote attachment to specific host cell receptors and apply secretion systems to inject effector proteins into the host in order to evade host immunity. Once the niche at the host plant has been reached the colonizing bacteria must compete with other microorganisms for this rare habitat. Possessing three T6SSs, we suspect K. radicincitans has an appropriate answer to many different environmental cues. Two general observations on the distribution of type VI secretion systems (T6SSs) suggest that the species-wide occurrence of three T6SSs in K. radicincitans is extraordinary: (i) T6SSs occur in only one quarter of bacteria (Bingle et al., 2008) and (ii) only 7% of T6SS-containing bacteria possess three or more T6SSs (Bernal et al., 2018). T6SSs have long been considered virulence factors of pathogenic bacteria, but next to evasion of host immunity they have meanwhile been associated with interbacterial competition for niches and survival in harmful environments (Wang et al., 2015).
“The wide distribution of T6SS clusters in plant-associated bacteria suggests that this molecular weapon is important for optimal fitness during plant colonization be it to cope with the plant response as much as with the resident microbiota” (Bernal et al., 2018). Based on genetic architecture and phylogeny T6SSs have been divided into five groups (Boyer et al., 2009), and searching for correlations between groups of T6SS and similar ecological niches Boyer and co-workers referred to group 4 as the group of “plant associated bacteria.” However, Bernal and co-workers found it more reasonable that the role of a specific T6SS is not defined by the group affiliation but primarily driven by the function of the effectors that the system secretes. Many effector-encoding genes are found in close proximity to vgrG, hcp, or paar genes and effectors are often fused to components of the Hcp–VgrG–PAAR structure; and VgrG, Hcp and PAAR-domain containing proteins (e.g., Rhs) may represent toxic compounds themselves (Cianfanelli et al., 2016). T6SS.3 of K. radicincitans possesses vgrG, hcp and paar genes (Supplementary Figure S7) indicating its large potential to eject effectors. Additional work will be required for detecting cargo effectors that are not fused with any component of the secretion machinery and, in some cases, are not genetically linked with any other T6SS genes (Cianfanelli et al., 2016). “The presence of non-T6SS-related elements encoded within or near T6SS clusters is an important factor to take into account” (Bernal et al., 2018). Interestingly, the genetic neighborhood of K. radicincitans' T6SS.3 harbors genes for multiple antibiotic resistance (marR), multidrug resistance (mdtN, emrB), and oxidative stress mitigation (marR, katG) suggesting that T6SS.3 provides fitness advantages in both polymicrobial and plant environments. The order of T6SS.3 genes in the KORA group is highly conserved (Figure 3) suggesting that T6SS.3 is of rather ancient origin in this species and may confer an important function to K. radicincitans.
Using the plant growth-promoting endophytic strain Enterobacter cloacae ENHKU01, Liu and co-workers performed antagonistic assays and found significant effects against pathogenic microbes within the environmental niche of ENHKU01, such as Ralstonia solanacearum and a wide range of fungal species (Liu et al., 2013). The authors hypothesized that E. cloacae strains possessing more than one distinct T6SS, such as ENHKU01, are likely to have fitness advantages in a broader range of habitats. Considering (i) that K. radicincitans possesses both T6SSs of ENHKU01 and another distinct T6SS (Supplementary Figure S7) and (ii) that T6SS likely provides fitness and colonization advantages to phytobacteria in planta (Bernal et al., 2018) we speculate that K. radicincitans benefits from its three T6SSs while colonizing its host.
Multiple flagellar systems and multiple secretion systems may help to avoid pattern triggered immunity of plants
Microorganisms are known to trigger plant immunity via microbe-associated molecular patterns (MAMPs). Pattern-recognition receptors (PRRs) of plants recognize MAMPs and induce pattern-triggered immunity (PTI) that minimizes microbial invasion. In order to evade plant immunity microorganism respond in different ways. Given a mutualistic symbiosis, endophyte and host may co-evolve toward perception and signaling without triggering each other's immunity. This may also involve masking or non-synthesis of MAMPs such as chitin (Becker et al., 2016). In contrast, any plant-microbe interaction not based on long-term co-evolution is likely followed by defense upon microbe perception, initially yielding slower plant growth. However, K. radicincitans DSM 16656T promotes growth of a wide range of plant species shortly (two weeks) after inoculation, suggesting an ability to circumvent or rapidly overcome perception by the plant and to avoid MAMP triggered immunity.
Investigating Kosakonia radicincitans strain YD4, Bergottini (2015) was the first to describe two “flagellar biosynthesis operons” in this species, but the dimension and rareness of the two flagellar systems among enteric bacteria was not determined. We found that all strains of K. radicincitans but only few strains of other enteric bacteria have two flagellar systems, here called KFS1 and KFS2 (with “K” referring to Kosakonia). KFS1 and KFS2 differ in gene cluster composition and in the amino acid sequence of their flagellin protein. The C-terminal end of the conserved Flg15 motif of the flagellin molecule is of high importance for perception by the plant. While synthetic peptides that lack the two amino acid residues present at the C terminus proved completely inactive as elicitors in A. thaliana (Bauer et al., 2001), in tomato abrupt loss of elicitor activity occurred for peptides lacking four amino acid residues (Felix et al., 1999). The substitution of two of the four last amino acid residues, which occurs in E. coli and K. radicincitans (Supplementary Figure S6), was shown to reduce the elicitor activity of Flg15 to < 1/3 when compared to Flg15 of Pseudomonas aeruginosa (Meindl et al., 2000). The substitution of the very last amino acid of Flg15 in KFS2 of K. radicincitans may further reduce its elicitor activity. A putative switch from KFS1 expression to KFS2 expression in the vicinity of plants would help K. radicincitans to avoid PTI of plants. However, expression of genes encoding pathogenesis-related (PR) proteins was induced in Arabidopsis thaliana treated with DSM 16656T (Brock et al., 2013) suggesting that DSM 16656T was recognized by the plant. Thus, we speculate, that effector proteins ejected by the three T6SSs of K. radicincitans are involved in interbacterial competition and plant immunity suppression likewise.
K. radicincitans inoculation affects the microbiota of plants
Assuming a correlation between secretion systems, motility and low host specificity one may expect Kosakonia to strongly compete with the natural microbiota of plants for available niches. K. radicincitans DSM 16656T was shown to possess high competitiveness and plant colonizing ability when applied to Triticum aestivum, its native host (Ruppel et al., 1992). Six weeks after inoculation wheat roots and shoots were equally colonized by about 106 cells per mL root or shoot sap, irrespective of the inoculum density, i.e., 102, 104, 106, or 108 cells per plant. Even the non-host plant Brassica oleracea was densely colonized with K. radicincitans accounting for 10–16% of the total plant bacterial community 14 days after inoculation (Ruppel et al., 2006). Our PhyloChip analysis (Figure 6) showed that DSM 16656T has a massive impact on the bacterial community composition of tomato. Whether Kosakonia's competitor suppression is simply caused by niche exclusion via rapid plant colonization or by a more sophisticated approach including antibiotic compounds needs to be determined. However, type VI secretion systems are known for directly targeting eukaryotic cells, and also for interbacterial interactions in polymicrobial environments (Records, 2011; Ho et al., 2014; Alteri and Mobley, 2016; Cianfanelli et al., 2016). Microbiota analyses on a range of plants (other than tomato) will be required to see whether an inoculation of K. radicincitans shifts plant bacterial communities always in the same direction, irrespectively of the plant genotype, and whether fungi are also affected. Documenting the altered microbiota of plants during a time series will help to determine whether differences in the plant microbiota are compensated over time, or can be carried via seeds to the next plant generation, thus having a massive impact on the holobiont.
Dual flagellar systems may allow increased motility in different environments
Bacteria are supposed to benefit from multiple flagellar systems due to increased motility in different environments (McCarter, 2004, 2005; Bubendorfer et al., 2014). Confronted by an antagonistic microorganism high motility may e.g., help to escape from destructive competition. However, in general dual flagellar systems were reported to be required for distinct modes of migration in different abiotic environments. For instance it was shown that some bacteria have primary and secondary flagellar systems for swimming and swarming (McCarter, 2004, 2005). According to the repertoire of motility genes, Kosakonia having two entire flagellar systems, Enterobacter having one and Klebsiella having only a fraction of one (Figure 2C), the three genera may differ in their potential to swim and swarm. Our studies have shown that the first flagellar system of DSM 16656T (KFS1.a + KFS1.b) is expressed in liquid culture and upregulated in response to root exudates (Figure 3) indicating that KFS1 is used for swimming toward host plants; but the purpose of KFS2 has to be determined yet. Kosakonia oryzae Ola 51T is closely related to K. radicincitans DSM 16656T (Figure 1), possesses both flagellar systems (Figure 4) and like DSM 16656T was found to be peritrichously flagellated (Li et al., 2017), but whether both flagellar systems contribute to formation of peritrichously inserted flagella still has to be investigated.
In addition to earlier reports, recent analyses have indicated that duplicated flagellar systems may be involved in biotic interaction. Contributing to competition for nodulation the secondary flagellar systems of Bradyrhizobium diazoefficiens (Alphaproteobacteria) is involved in plant colonization (Mongiardini et al., 2017). Levy and co-workers found that additional flagellum-like gene clusters are distributed within Burkholderiales (Betaproteobacteria) among plant- and soil-associated strains and concluded that this flagellar structure variant evolved in the plant environment (Levy et al., 2018). Moreover, the authors discovered that genomes of plant-associated bacteria encode more carbohydrate metabolism functions than related non-plant-associated genomes do. Interestingly, there are only five genes inserted into KFS2 of DSM 16656T that do not directly belong to flagellar biosynthesis, functioning, and chemotaxis, and four of them encode proteins involved in carbohydrate metabolism. Among these four genes is axe1-6A that encodes a highly efficient carbohydrate esterase for degrading matrix polysaccharides (hemicellulose) in plant cell walls (Kabel et al., 2011). Our finding that there is a duplicated mdtABC operon (mdtABC.2) adjacent to KFS2, which is restricted to strains that possess KFS2, suggests a functional link of both gene clusters. Next to export of the siderophore enterobactin (Horiyama and Nishino, 2014) the MdtABC efflux pump was found to be involved in resistance to plant antimicrobials, such as flavonoids and tannin, and MdtABC-deficient mutants of Erwinia amylovora showed reduced ability to multiply in apple rootstock (Pletzer and Weingart, 2014). Although Erwinia amylovora is a plant pathogen, it is obvious that beneficial plant-associated bacteria would equally benefit from resistance to antimicrobial plant compounds. Assuming that mdtABC.2 and axe1-6A of DSM 16656T are involved in plant interaction it is tempting to speculate that KFS2 contributes to the plant-associated lifestyle of K. radicincitans.
DSM 16656T exhibits an endophytic, non-damaging lifestyle
Genes involved in degradation of the major cell wall components lignin and cellulose, next to chitin the most common organic compounds on earth, appear to be missing from K. radicincitans. Lignin is an aromatic polymer and as such a potential carbon source for enteric bacteria such as Klebsiella (Diaz et al., 2001), a genus that carries up to 10 times as many genes for metabolism of aromatic compounds as Kosakonia (Supplementary Figure S5). Nonetheless, the absence of lignin and cellulose degrading enzymes in K. radicincitans may refer to its endophytic, non-damaging lifestyle, which we infer from our observation that K. radicincitans has never caused any detrimental effect on host plants despite massive colonization. Interestingly, DSM 16656T carries two chromosomal regions that contain several genes for cellulose synthesis (bcsABCEZ, acsABCD, yhjDEHUT), possibly important for biofilm formation and plant colonization (Römling and Galperin, 2015). These gene clusters are shared with a few other enteric bacteria, among them Enterobacter sp. FY-07 which was confirmed to produce bacterial cellulose (Ji et al., 2016).
There is a strong link between Citrobacter, Kosakonia and other enteric PGPB
Genes shared between DSM 16656T and Citrobacter, presumably resulting from horizontal gene transfer, have increased the adaptive potential of DSM 16656T. There is an integrative conjugative element (ICE) on the chromosome comprising 109 genes which is shared between DSM 16656T and Citrobacter amalonaticus Y19, but missing for the most part in the other 33 bacterial genomes we analyzed (Figure 4A). The ICE contains a gene cluster (pilLM/bfpB/pilO/epsE/tcpE/pilS) for type IV pili formation, bacterial appendages that are involved in adhesion to host cells and biofilm formation. These pili or fimbriae are filamentous structures like flagella, but smaller, and are thought to enable gliding or twitching motility, a synchronized and flagella-independent form of bacterial movement over moist surfaces, important in rapid host colonization (Mattick, 2002).
The observation that most genes of the large plasmid of DSM 16656T are shared with Citrobacter and missing in other Kosakonia strains is further evidence for the exchange of genomic material between both genera. The large genome-wide homology of Enterobacter sp. 638, an endophyte of poplar from USA, to Citrobacter koseri ATCC BAA-895 (Taghavi et al., 2010) suggest regular horizontal gene transfer between Citrobacter and other groups of enteric bacteria. The same accounts to the fact that Enterobacter sp. 638 was nested within a Citrobacter clade in our comprehensive 16S rRNA gene tree (Supplementary Figure S2). Some Citrobacter strains were found to be plant endophytes with plant protecting capacities and PGP potential (Wang et al., 2006; Mousa et al., 2015). Hence, they do likely share habitats with endophytic bacteria of genus Kosakonia and Enterobacter and may synergistically interact while exhibiting a beneficial impact on plants.
The large plasmid may enhance the plant-associated lifestyle of DSM 16656T
Some bacteria are able to produce 1,2-propanediol (1,2-PD) by fermenting rhamnose and fucose, common sugars in plant cell walls (Badia et al., 1985), and the genome composition of DSM 16656T indicates that K. radicincitans is among these bacteria. However, 1,2-PD itself is a valuable carbon source and can be degraded by DSM 16656T. Intermediate products of 1,2-PD degradation are mutagens and need to be detoxified by coenzyme B12 in bacterial microcompartments (Sampson and Bobik, 2008). Consistently, genes for coenzyme B12 metabolism are provided by the large plasmid and the chromosome. Ongoing work has to clarify whether this anaerobic degradation of plant compartments is involved in establishing a microaerobic endophytic niche for N2-fixation or simply in decomposing dead plant material. 1,2-PD degradation is likely to provide a selective advantage in the anaerobic depths of soils (Sampson and Bobik, 2008), and “Enterobacter lignolyticus” SCF1, isolated from anaerobic tropical forest soils (DeAngelis et al., 2011), was the only Enterobacter strain found to share the bulk of pdu genes with DSM 16656T (Figure 5). However, intermediate and final products of PDD and GT may also be involved in better plant performance, both the volatile organic compounds 1,2-propanediol and 1,3-propanediol were reported to promote plant growth (Nakajima et al., 2005; Tahir et al., 2017). Taken together, the genomic composition of K. radicincitans underpins its tight interaction with plants, but the large plasmid of DSM 16656T appears to have further enhanced the plant-associated lifestyle of this particular strain.
Multiple rRNA operons may allow responsiveness to diverse environments
Next to pili, flagella and secretion systems, another bacterial component has been reported to increase the fitness of bacteria: rRNA operons (rrn), of which DSM 16656T possesses seven copies (Supplementary Figure S1B). The persistence of multiple rrn copies has been considered to reflect ecological strategies of bacteria. Klappenbach and co-workers discovered that soil bacteria grew rapidly on nutritionally complex medium and tolerated herbicide 2,4-dichlorophenoxyacetic acid (2,4-D) when rrn copy number was high (Klappenbach et al., 2000). The maximum reproductive rate of bacteria doubles with a doubling of rrn copy number, and the efficiency of carbon use is inversely related to maximal growth rate and rrn copy number (Roller et al., 2016). These reports suggest that bacteria with a high rrn copy number must exploit many nutrient sources but are rewarded by fast growth.
Bacterial N2-fixation promotes plant growth, which in turn provides larger habitats to endophytic bacteria
The most striking difference between Kosakonia, Klebsiella, and Enterobacter in terms of PGP traits is the ability to fix atmospheric nitrogen: Kosakonia has two gene clusters for N2-fixation, Klebsiella has one and Enterobacter has none. Whereas, nifHDK of K. radicincitans encodes a component of the iron–molybdenum (FeMo) nitrogenase, which is found in all diazotrophs, its anfHDGK encodes for a component of the iron-only (Fe) nitrogenase found secondarily in only some diazotrophs (Ekandjo et al., 2017). In conformity with two nitrogenase gene clusters there are also two genetic components for regulatory cascades controlling nif transcription in K. radicincitans. Diazotrophic Gammaproteobacteria and rhizobia differ in the first level of the cascade, a two-component-system comprising the genes ntrBC and fixJL, respectively (Dixon and Kahn, 2004). While Klebsiella carries the ntrBC system and rhizobia the fixJL system, Kosakonia has both of them. Unlike ntrBC, fixJL is responsive to oxygen (Dixon and Kahn, 2004) guaranteeing that nif transcription only happens in a microaerobic habitat. Legume-rhizobia symbiosis is thought to start when particular plant root exudates bind to NodD, the transcriptional regulator of bacterial nodulation (Mus et al., 2016). Although not inducing nodule organogenesis, K. radicincitans DSM 16656T carries two nodD genes and each of them is spatially close to either of the two-component systems controlling nif transcription. Hence, the presence of fixJL and nodD genes suggests that N2-fixation of DSM 16656T is affected by plants and O2 concentration. Ekandjo and co-workers could not detect significant contribution of the Fe nitrogenase to biological dinitrogen assimilation under pure bacterial culture experimental conditions but speculated that having both nitrogenases might enable K. radicincitans to fix atmospheric nitrogen in different environments, however fixation of atmospheric nitrogen in planta was not studied (Ekandjo et al., 2017). Our microscopic analyses of DSM 16656T colonizing root parenchyma cells of plants (Figure 7) suggest that enteric bacteria may be capable of generating their own microaerobic niche for atmospheric nitrogen fixation. A combination of microscopic and functional analyses is required in order to determine whether plant parenchyma cells are the location of Kosakonia's N2-fixation.
Costs and benefits of multiple gene clusters
Next to atmospheric nitrogen fixation other metabolically expensive gene clusters, such as dual flagellar systems, triple T6SSs (Figures 3, 4), and multiple rRNA operons (Supplementary Figure S1B) suggest that K. radicincitans DSM 16656T encounters nutrient-rich environments, where high metabolic costs are not detrimental but outweighed by benefits. Addressing the need for high amounts of energy, K. radicincitans DSM 16656T exhibits a genetic composition well-suited for exploiting a large variety of carbon sources. Pathways involved in oxidative phosphorylation are the only pathways shared between the chromosome and both plasmids of K. radicincitans DSM 16656T, probably indicating that generating sufficient energy is pivotal for this strain. Being a generalist plant colonizer, K. radicincitans must be highly adaptable to plant immunity systems in order to achieve host colonization. In this context, it will be interesting to see whether Kosakonia from the nutrient-poor soil of sundew (Drosera burmannii; Supplementary Table S1) also exhibits low host specificity and multiple copies of the gene clusters mentioned above.
Summary and outlook
In this article we provide comprehensive knowledge about the newly emerging PGP species Kosakonia radicincitans and reveal duplicated species-specific gene clusters that have not yet been described in other bacteria. The competence of K. radicincitans to colonize and exert growth-promoting effects in a wide range of plants is intriguing and might be facilitated by genes contributing to high motility (two flagellar systems) and high competitiveness (three T6SSs). These genes now need to be functionally analyzed. Studies on the PTI capacities of K. radicincitans knock-out mutants, in which single T6SSs are deleted, will be required to answer the question, whether T6SSs found in K. radicincitans DSM 16656T are involved in plant immunity suppression, in interbacterial competition, or both. Likewise, experiments on KFS knock-out mutants will be required to reveal the purpose of KFS2. More research on K. radicincitans strains other than DSM 16656T is required in order to unravel the plant-colonizing and growth-promoting capacity of the whole species. Given that K. radicincitans exhibits unique, complex, and intra-specific consistent gene clusters this species has the potential to become a model organism for studies on multiple modes of motility and bacteria-host interactions.
Garcia-Fraile and co-workers stated that the use of “Enterobacteria” as biofertilizers involves a risk to humans and plants (Garcia-Fraile et al., 2012). However, the fact that natural hosts of Kosakonia belong to the most consumed crop plants in the world indicates that in general Kosakonia's potential for causing plant and human diseases must be low. Although being closely related to pathogenic strains the potential of presumably beneficial bacteria has to be determined since the percentage of highly efficient PGPB among cultivable endophytic bacteria is low. We discovered a rate of 0.5% when we tested a thousand isolates from the surface-sterilized phyllosphere of winter wheat (Ruppel, 1988). This emphasizes the need for the synergistic use of bioinformatics for in silico prediction and affirmative in vivo experiments in order to distinguish promising candidates from really effective PGPB and also from possibly pathogenic strains. The comprehensive genome comparisons by Levy and co-workers found that plant-associated bacteria from across the bacterial kingdom share gene clusters for nodulation, nitrogen fixation, chemotaxis, flagellum biosynthesis and type VI secretion systems (Levy et al., 2018). Independently, we found the same gene clusters multiplied in K. radicincitans DSM 16656T, while searching for unique genomic features of an extraordinary successful colonizer and growth promoter of plants.
A database assigning PGP traits, e.g., a gene ontology (GO) term “plant growth-promotion,” would enable pre-selection of putatively beneficial microorganisms from a list of all isolated and fully sequenced ones. Deciphering the PGP potential of microorganisms by in silico analyses would significantly facilitate the future search for promising candidates for application in sustainable crop production.
Statements
Author contributions
MB designed research and wrote the manuscript. SP and MB performed in silico analyses. YB generated eGFP mutants of DSM 16656T and performed CLSM analyses. BeB performed in vivo experiments and generated material for microarray and PhyloChip analyses. MD processed microarray data. JO, BoB, and CS sequenced the genome of DSM 16656T by SMRT and Illumina sequencing. JR performed TEM analyses, GT assessed phosphate solubilization activity. SR initiated microarray and PhyloChip analyses. MB, SP, YB, BeB, and SR analyzed the data. All authors contributed to writing the manuscript.
Funding
MB received funding from DIP (grant 742266/1) and BeB from BMBF (grant FKZ 031A282). The funders had no role in study design, data collection and interpretation.
Acknowledgments
We thank Simone Severitt and Nicole Mrotzek for excellent technical assistance in PacBio and Illumina sequencing of K. radicincitans DSM 16656T and Birgit Wernitz for technical assistance in bacteria-plant interaction experiments. The 16S rRNA gene sequence of isolate RSSL-1 was provided by Dagmar Woebken, David Seki, and Hannes Schmidt, Department of Microbiology and Ecosystem Science, University of Vienna. We thank Ellen Lagendjik, Leiden University, Netherlands, for providing the eGFP plasmid pMP4655.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01997/full#supplementary-material
References
1
Alteri C. J. Mobley H. L. (2016). The versatile type VI secretion system. Microbiol. Spectrum 4, 10.1128/microbiolspec.VMBF-0026-2015
2
Badia J. Ros J. Aguilar J. (1985). Fermentation mechanism of fucose and rhamnose in Salmonella thyphimurium and Klebsiella pneumoniae. J. Bacteriol.161, 435–437.
3
Bauer Z. Gómez-Gómez L. Boller T. Felix G. (2001). Sensitivity of different ecotypes and mutants ofarabidopsis thaliana toward the bacterial elicitor flagellin correlates with the presence of receptor-binding sites. J. Biol. Chem.276, 45669–45676. 10.1074/jbc.M102390200
4
Becker M. Becker Y. Green K. Scott B. (2016). The endophytic symbiont Epichloe festucae establishes an epiphyllous net on the surface of Lolium perenne leaves by development of an expressorium, an appressorium-like leaf exit structure. New Phytol.211, 240–254. 10.1111/nph.13931
5
Berger B. Baldermann S. Ruppel S. (2017). The plant growth-promoting bacterium Kosakonia radicincitans improves fruit yield and quality of Solanum lycopersicum. J. Sci. Food Agric.97, 4865–4871. 10.1002/jsfa.8357
6
Berger B. Brock A. K. Ruppel S. (2013). Nitrogen supply influences plant growth and transcriptional responses induced by Enterobacter radicincitans in Solanum lycopersicum. Plant Soil370, 641–652. 10.1007/s11104-013-1633-0
7
Berger B. Patz S. Ruppel S. Dietel K. Faetke S. Junge H. et al . (2018). Successful formulation and application of plant growth-promoting Kosakonia radicincitans in maize cultivation. BioMed Res. Int.2018:8. 10.1155/2018/6439481
8
Berger B. Wiesner M. Brock A. K. Schreiner M. Ruppel S. (2015). K. radicincitans, a beneficial bacteria that promotes radish growth under field conditions. Agron. Sustain. Dev.35, 1521–1528. 10.1007/s13593-015-0324-z
9
Bergottini V. M. (2015). Assessing the Role of Native Plant Growth Promoting Rhizobacteria as Bio-Inoculants for Yerba Mate (Ilex paraguariensis). PhD thesis. University of Neuchatel, Neuchatel, Switzerland.
10
Bernal P. Llamas M. A. Filloux A. (2018). Type VI secretion systems in plant-associated bacteria. Environ. Microbiol.20, 1–15. 10.1111/1462-2920.13956
11
Bingle L. E. Bailey C. M. Pallen M. J. (2008). Type VI secretion: a beginner's guide. Curr. Opin. Microbiol.11, 3–8. 10.1016/j.mib.2008.01.006
12
Boyer F. Fichant G. Berthod J. Vandenbrouck Y. Attree I. (2009). Dissecting the bacterial type VI secretion system by a genome wide in silico analysis: what can be learned from available microbial genomic resources?BMC Genomics10:104. 10.1186/1471-2164-10-104
13
Brady C. Cleenwerck I. Venter S. Coutinho T. De Vos P. (2013). Taxonomic evaluation of the genus Enterobacter based on multilocus sequence analysis (MLSA): proposal to reclassify E. nimipressuralis and E. amnigenus into Lelliottia gen. nov. as Lelliottia nimipressuralis comb. nov. and Lelliottia amnigena comb. nov., respectively, E. gergoviae and E. pyrinus into Pluralibacter gen. nov. as Pluralibacter gergoviae comb. nov. and Pluralibacter pyrinus comb. nov., respectively, E. cowanii, E. radicincitans, E. oryzae and E. arachidis into Kosakonia gen. nov. as Kosakonia cowanii comb. nov., Kosakonia radicincitans comb. nov., Kosakonia oryzae comb. nov. and Kosakonia arachidis comb. nov., respectively, and E. turicensis, E. helveticus and E. pulveris into Cronobacter as Cronobacter zurichensis nom. nov., Cronobacter helveticus comb. nov. and Cronobacter pulveris comb. nov., respectively, and emended description of the genera Enterobacter and Cronobacter. Syst. Appl. Microbiol.36, 309–319. 10.1016/j.syapm.2013.03.005
14
Brock A. K. Berger B. Mewis I. Ruppel S. (2013). Impact of the PGPB Enterobacter radicincitans DSM 16656 on growth, glucosinolate profile, and immune responses of Arabidopsis thaliana. Microb Ecol.65, 661–670. 10.1007/s00248-012-0146-3
15
Bubendorfer S. Koltai M. Rossmann F. Sourjik V. Thormann K. M. (2014). Secondary bacterial flagellar system improves bacterial spreading by increasing the directional persistence of swimming. Proc. Natl. Acad. Sci. U.S.A.111, 11485–11490. 10.1073/pnas.1405820111
16
Cianfanelli F. R. Monlezun L. Coulthurst S. J. (2016). Aim, load, fire: the type VI secretion system, a bacterial nanoweapon. Trends Microbiol.24, 51–62. 10.1016/j.tim.2015.10.005
17
DeAngelis K. M. D'Haeseleer P. Chivian D. Fortney J. L. Khudyakov J. Simmons B. et al . (2011). Complete genome sequence of “Enterobacter lignolyticus” SCF1. Stand. Genomic Sci.5, 69–85. 10.4056/sigs.2104875
18
Diaz E. Ferrandez A. Prieto M. A. Garcia J. L. (2001). Biodegradation of aromatic compounds by Escherichia coli. Microbiol. Mol. Biol. Rev.65, 523–569. 10.1128/MMBR.65.4.523-569.2001
19
Dixon R. Kahn D. (2004). Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol.2, 621–631. 10.1038/nrmicro954
20
Ekandjo L. K. Ruppel S. Remus R. Witzel K. Patz S. Becker Y. (2017). Site-directed mutagenesis to deactivate two nitrogenase isozymes of Kosakonia radicincitans DSM16656T. Can. J. Microbiol.64, 97–106. 10.1139/cjm-2017-0532
21
Felix G. Duran J. D. Volko S. Boller T. (1999). Plants have a sensitive perception system for the most conserved domain of bacterial flagellin. Plant J.18, 265–276. 10.1046/j.1365-313X.1999.00265.x
22
Garcia-Fraile P. Carro L. Robledo M. Ramírez-Bahena M.-H. Flores-Félix J.-D. Teresa Fernández M. et al . (2012). Rhizobium promotes non-legumes growth and quality in several production steps: towards a biofertilization of edible raw vegetables healthy for humans. PLoS ONE7:e38122. 10.1371/journal.pone.0038122
23
Gerhardt P. Murray R. Krieg N. R. Wood W. A. (1994). Methods for General and Molecular Bacteriology. Washington, D. C.: ASM Press.
24
Gu C. T. Li C. Y. Yang L. J. Huo G. C. (2014). Enterobacter xiangfangensis sp. nov., isolated from Chinese traditional sourdough, and reclassification of Enterobacter sacchari Zhu et al. 2013 as Kosakonia sacchari comb. nov. Int. J. Syst. Evol. Microbiol.64, 2650–2656. 10.1099/ijs.0.064709-0
25
Hartmann A. Rothballer M. Hense B. A. Schröder P. (2014). Bacterial quorum sensing compounds are important modulators of microbe-plant interactions. Front. Plant Sci.5:131. 10.3389/fpls.2014.00131
26
Ho B. T. Dong T. G. Mekalanos J. J. (2014). A view to a kill: the bacterial type VI secretion system. Cell Host Microbe15, 9–21. 10.1016/j.chom.2013.11.008
27
Horiyama T. Nishino K. (2014). AcrB, AcrD, and MdtABC multidrug efflux systems are involved in enterobactin export in Escherichia coli. PLoS ONE9:e108642. 10.1371/journal.pone.0108642
28
Ji K. Wang W. Zeng B. Chen S. Zhao Q. Chen Y. et al . (2016). Bacterial cellulose synthesis mechanism of facultative anaerobe Enterobacter sp. FY-07. Sci. Rep.6:21863. 10.1038/srep21863
29
Kabel M. A. Yeoman C. J. Han Y. J. Dodd D. Abbas C. A. de Bont J. A. et al . (2011). Biochemical characterization and relative expression levels of multiple carbohydrate esterases of the xylanolytic rumen bacterium Prevotella ruminicola 23 grown on an ester-enriched substrate. Appl. Environ. Microbiol.77, 5671–5681. 10.1128/AEM.05321-11
30
Kampfer P. Ruppel S. Remus R. (2005). Enterobacter radicincitans sp. nov., a plant growth promoting species of the family Enterobacteriaceae. Syst. Appl. Microbiol.28, 213–221. 10.1016/j.syapm.2004.12.007
31
Klappenbach J. A. Dunbar J. M. Schmidt T. M. (2000). rRNA operon copy number reflects ecological strategies of bacteria. Appl Environ. Microbiol.66, 1328–1333. 10.1128/AEM.66.4.1328-1333.2000
32
Koboldt D. C. Zhang Q. Larson D. E. Shen D. McLellan M. D. Lin L. et al . (2012). VarScan 2: somatic mutation and copy number alteration discovery in cancer by exome sequencing. Genome Res.22, 568–576. 10.1101/gr.129684.111
33
Levy A. Salas Gonzalez I. Mittelviefhaus M. Clingenpeel S. Herrera Paredes S. Miao J. et al . (2018). Genomic features of bacterial adaptation to plants. Nat. Genet.50, 138–150. 10.1038/s41588-017-0012-9
34
Li H. Durbin R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics25, 1754–1760. 10.1093/bioinformatics/btp324
35
Li Y. Li S. Chen M. Peng G. Tan Z. An Q. (2017). Complete genome sequence of Kosakonia oryzae type strain Ola 51(T). Stand. Genomic Sci.12:28. 10.1186/s40793-017-0240-8
36
Lin L. Li Z. Hu C. Zhang X. Chang S. Yang L. et al . (2012). Plant growth-promoting nitrogen-fixing enterobacteria are in association with sugarcane plants growing in Guangxi, China. Microb Environ.27, 391–398. 10.1264/jsme2.ME11275
37
Liu W. Y. Wong C. F. Chung K. M. Jiang J. W. Leung F. C. (2013). Comparative genome analysis of Enterobacter cloacae. PLoS ONE8:e74487. 10.1371/journal.pone.0074487
38
Mattick J. S. (2002). Type IV pili and twitching motility. Annu. Rev. Microbiol.56, 289–314. 10.1146/annurev.micro.56.012302.160938
39
McCarter L. L. (2004). Dual flagellar systems enable motility under different circumstances. J. Mol. Microbiol. Biotechnol.7, 18–29. 10.1159/000077866
40
McCarter L. L. (2005). Multiple modes of motility: a second flagellar system in Escherichia coli. J. Bacteriol.187, 1207–1209. 10.1128/JB.187.4.1207-1209.2005
41
Meindl T. Boller T. Felix G. (2000). The bacterial elicitor flagellin activates its receptor in tomato cells according to the address–message concept. Plant Cell12, 1783–1794. 10.1105/tpc.12.9.1783
42
Mongiardini E. J. Quelas J. I. Dardis C. Althabegoiti M. J. Lodeiro A. R. (2017). Transcriptional control of the lateral-flagellar genes of Bradyrhizobium diazoefficiens. J. Bacteriol.199, e00253–e00217. 10.1128/JB.00253-17
43
Mousa W. K. Shearer C. R. Limay-Rios V. Zhou T. Raizada M. N. (2015). Bacterial endophytes from wild maize suppress Fusarium graminearum in modern maize and inhibit mycotoxin accumulation. Front. Plant Sci.6:805. 10.3389/fpls.2015.00805
44
Mus F. Crook M. B. Garcia K. Garcia Costas A. Geddes B. A. Kouri E. D. et al . (2016). Symbiotic nitrogen fixation and the challenges to its extension to nonlegumes. Appl. Environ. Microbiol.82, 3698–3710. 10.1128/AEM.01055-16
45
Nakajima E. Iqbal Z. Araya H. Hiradate S. Hamano M. Fujii Y. (2005). Isolation and identification of a plant growth promotive substance from mixture of essential plant oils. Plant Growth Regul.45, 47–51. 10.1007/s10725-004-7087-x
46
Pletzer D. Weingart H. (2014). Characterization and regulation of the resistance-nodulation-cell division-type multidrug efflux pumps MdtABC and MdtUVW from the fire blight pathogen Erwinia amylovora. BMC Microbiol.14:185. 10.1186/1471-2180-14-185
47
Records A. R. (2011). The type VI secretion system: a multipurpose delivery system with a phage-like machinery. Mol. Plant Microb. Interact.24, 751–757. 10.1094/MPMI-11-10-0262
48
Remus R. Ruppel S. Jacob H. J. Hecht-Buchholz C. Merbach W. (2000). Colonization behaviour of two enterobacterial strains on cereals. Biol. Fert. Soils30, 550–557. 10.1007/s003740050035
49
Roller B. R. Stoddard S. F. Schmidt T. M. (2016). Exploiting rRNA operon copy number to investigate bacterial reproductive strategies. Nat. Microbiol.1, 16160. 10.1038/nmicrobiol.2016.160
50
Römling U. Galperin M. Y. (2015). Bacterial cellulose biosynthesis: diversity of operons, subunits, products and functions. Trends Microbiol.23, 545–557. 10.1016/j.tim.2015.05.005
51
Ruppel S. (1988). Isolation of diazotrophic bacteria from the endorhizosphere and rhizoplane of winter wheat in temperate region. Zentralbl Mikrobiol.143, 621–629.
52
Ruppel S. Hechtbuchholz C. Remus R. Ortmann U. Schmelzer R. (1992). Settlement of the diazotrophic, phytoeffective bacterial strain Pantoea agglomerans on and within winter wheat–an investigation using ELISA and transmission electron microscopy. Plant Soil145, 261–273. 10.1007/BF00010355
53
Ruppel S. Ruhlmann J. Merbach W. (2006). Quantification and localization of bacteria in plant tissues using quantitative real-time PCR and online emission fingerprinting. Plant Soil286, 21–35. 10.1007/s11104-006-9023-5
54
Sampson E. M. Bobik T. A. (2008). Microcompartments for B12-dependent 1,2-propanediol degradation provide protection from DNA and cellular damage by a reactive metabolic intermediate. J. Bacteriol.190, 2966–2971. 10.1128/JB.01925-07
55
Schreiner M. Krumbein A. Ruppel S. (2009). Interaction between plants and bacteria: glucosinolates and phyllospheric colonization of cruciferous vegetables by Enterobacter radicincitans DSM 16656. J. Mol. Microbiol. Biotechnol.17, 124–135. 10.1159/000226589
56
Seemann T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics30, 2068–2069. 10.1093/bioinformatics/btu153
57
Taghavi S. van der Lelie D. Hoffman A. Zhang Y. B. Walla M. D. Vangronsveld J. et al . (2010). Genome sequence of the plant growth promoting endophytic bacterium Enterobacter sp. 638. PLoS Genet.6:e1000943. 10.1371/journal.pgen.1000943
58
Tahir H. A. Gu Q. Wu H. Raza W. Hanif A. Wu L. et al . (2017). Plant growth promotion by volatile organic compounds produced by Bacillus subtilis SYST2. Front. Microbiol.8:171. 10.3389/fmicb.2017.00171
59
Vande Broek A. Lambrecht M. Vanderleyden J. (1998). Bacterial chemotactic motility is important for the initiation of wheat root colonization by Azospirillum brasilense. Microbiology144, 2599–2606. 10.1099/00221287-144-9-2599
60
Wang E. T. Tan Z. Y. Guo X. W. Rodríguez-Duran R. Boll G. Martinez-Romero E. (2006). Diverse endophytic bacteria isolated from a leguminous tree Conzattia multiflora grown in Mexico. Arch. Microbiol.186, 251–259. 10.1007/s00203-006-0141-5
61
Wang T. Si M. Song Y. Zhu W. Gao F. Wang Y. et al . (2015). Type VI secretion system transports Zn2+ to combat multiple stresses and host immunity. PLoS Pathog.11:e1005020. 10.1371/journal.ppat.1005020
62
Witzel K. Gwinn-Giglio M. Nadendla S. Shefchek K. Ruppel S. (2012). Genome sequence of Enterobacter radicincitans DSM16656(T), a plant growth-promoting endophyte. J. Bacteriol.194:5469. 10.1128/JB.01193-12
63
Witzel K. Strehmel N. Baldermann S. Neugart S. Becker Y. Becker M. et al . (2017). Arabidopsis thaliana root and root exudate metabolism is altered by the growth-promoting bacterium Kosakonia radicincitans DSM 16656T. Plant Soil419, 557–573. 10.1007/s11104-017-3371-1
64
Wolf A. B. Rudnick M. B. de Boer W. Kowalchuk G. A. (2015). Early colonizers of unoccupied habitats represent a minority of the soil bacterial community. FEMS Microbiol. Ecol.91:fiv024. 10.1093/femsec/fiv024
65
Yang J. Xie X. Wang X. Dixon R. Wang Y. P. (2014). Reconstruction and minimal gene requirements for the alternative iron-only nitrogenase in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A.111, E3718–E3725. 10.1073/pnas.1411185111
66
Zhou Y. Liang Y. Lynch K. H. Dennis J. J. Wishart D. S. (2011). PHAST: a fast phage search tool. Nucleic Acids Res.39, W347–W352. 10.1093/nar/gkr485
Summary
Keywords
Kosakonia radicincitans , Enterobacteriaceae, PGPB, endophytes, comparative genomics, flagellar system, type VI secretion system, plasmid
Citation
Becker M, Patz S, Becker Y, Berger B, Drungowski M, Bunk B, Overmann J, Spröer C, Reetz J, Tchuisseu Tchakounte GV and Ruppel S (2018) Comparative Genomics Reveal a Flagellar System, a Type VI Secretion System and Plant Growth-Promoting Gene Clusters Unique to the Endophytic Bacterium Kosakonia radicincitans. Front. Microbiol. 9:1997. doi: 10.3389/fmicb.2018.01997
Received
25 January 2018
Accepted
08 August 2018
Published
30 August 2018
Volume
9 - 2018
Edited by
Paula García-Fraile, Academy of Sciences of the Czech Republic, Czechia
Reviewed by
Maged M. Saad, King Abdullah University of Science and Technology, Saudi Arabia; Frank O'Neill Aylward, Virginia Tech, United States
Updates
Copyright
© 2018 Becker, Patz, Becker, Berger, Drungowski, Bunk, Overmann, Spröer, Reetz, Tchuisseu Tchakounte and Ruppel.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Matthias Becker becker.matthias@igzev.de
This article was submitted to Microbial Symbioses, a section of the journal Frontiers in Microbiology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.