 Jérémie Lebeurre1*
Jérémie Lebeurre1* Sandrine Dahyot2
Sandrine Dahyot2 Seydina Diene3
Seydina Diene3 Amandine Paulay1Marion Aubourg4
Amandine Paulay1Marion Aubourg4 Xavier Argemi5
Xavier Argemi5 Jean-Christophe Giard4Isabelle Tournier6
Jean-Christophe Giard4Isabelle Tournier6 Patrice François3Martine Pestel-Caron2
Patrice François3Martine Pestel-Caron2- 1UNIROUEN, GRAM EA2656, Normandie Université, Rouen, France
- 2UNIROUEN, GRAM EA2656, Rouen University Hospital, Normandie Université, Rouen, France
- 3Genomic Research Laboratory, Service of Infectious Diseases, Geneva University Hospitals, Geneva, Switzerland
- 4EA4655 U2RM (Équipe Antibio-Résistance), Université de Caen Basse-Normandie, Caen, France
- 5CHRU de Strasbourg, VBP EA7290, Fédération de Médecine Translationnelle de Strasbourg (FMTS), Institut de Bactériologie, Université de Strasbourg, Strasbourg, France
- 6UNIROUEN, Inserm U1245, Normandy Centre for Genomic and Personalized Medicine, Rouen University Hospital, Normandie Université, Rouen, France
Staphylococcus lugdunensis is a commensal bacterium of human skin that has emerged as a virulent Coagulase-Negative Staphylococcus in both community-acquired and healthcare associated infections. Genotyping methods have shown a clonal population structure of this pathogen but failed to identify hypervirulent lineages. Here, complete genomes of three pathogenic and three carriage S. lugdunensis strains were obtained by Single-Molecule sequencing (PacBio) and compared to 15 complete genomes available in GenBank database. The aim was to identify (i) genetic determinants specific to pathogenic or carriage strains or specific to clonal complexes (CCs) defined by MultiLocus Sequence Typing, and (ii) antibiotic resistance genes and new putative virulence factors encoded or not by mobile genetic elements (MGE). Comparative genomic analysis did not show a strict correlation between gene content and the ability of the six strains to cause infections in humans and in a Galleria mellonella infection model. However, this study identified new MGEs (five prophages, two genomic islands and one plasmid) and genetic variations of some putative virulence-associated loci, especially in CC3 strains. For a clonal population, high variability and eight CC-dependent genetic organizations were observed for the ess locus, which encodes a putative type VII secretion system (T7SS) homologous to that of S. aureus. Further phenotypic and functional studies are needed to characterize this particular CC3 and to evaluate the role of T7SS in the virulence of S. lugdunensis.
Introduction
Staphylococcus lugdunensis is a Coagulase-Negative Staphylococcal species (CoNS) that is a part of the normal human skin flora. However, unlike other CoNS, S. lugdunensis exhibits a high degree of virulence similar in some respects to Staphylococcus aureus (Frank et al., 2008).
Staphylococcus lugdunensis is responsible for infections ranging from minor skin and soft tissue infections (Bocher et al., 2009; Arias et al., 2010) to invasive diseases such as infective endocarditis (Non and Santos, 2017), bone and joint infections (Douiri et al., 2016; Lourtet-Hascoët et al., 2016; Argemi et al., 2017a), prosthetic device-infections (Anguera, 2005; Shah et al., 2010) and vascular catheter-related infections (Ebright et al., 2004), both in healthy host and immune-compromised patients. One of the characteristics of S. lugdunensis infections is the development of deep-seated abscess lesions that cannot be cleared without surgical intervention associated with effective antibiotic therapy (Arias et al., 2010). Interestingly, few virulence factors have been described to date to explain the high ability of S. lugdunensis to cause acute and suppurative infections, and particularly tissue damage and elevated mortality observed for infections such as infective endocarditis (Anguera, 2005; Sabe et al., 2014).
Several molecular typing methods have been described to search for a potential link between within-species genetic variations and pathogenic potential or virulence. Multi-Locus Sequence Typing (MLST) has shown a clonal population structure of this pathogen (Chassain et al., 2012), and has allowed the description of seven Clonal Complexes (CCs)1 but failed to identify hypervirulent lineages or clusters specific to carriage strains. Multi-Virulence Locus Sequence Typing (Didi et al., 2014) as well as Multiple Locus Variable Number Tandem Repeat Analysis (MLVA) and Tandem Repeat Sequence Typing (TRST) recently developed, have confirmed this absence of link between clustering and clinical settings (Dahyot et al., 2018).
Whole-genome sequencing (WGS) has become a powerful tool in microbiology allowing the sequencing of hundreds of S. aureus genomes but to date only 15 complete genomes of S. lugdunensis are available at the National Center for Biotechnology Information (NCBI). The only two studies comparing S. lugdunensis whole genomes revealed that the genome (i) is closer to that of S. aureus than other CoNS, (ii) harbors mobile genetic elements (MGE) such as prophages and plasmids, but (iii) displays a closed pan-genome and several systems (restriction-modification, toxin/antitoxin and CRISPR/Cas systems) to prevent horizontal gene transfer (Argemi et al., 2017b, 2018).
In this context, we first sequenced the whole genome of three pathogenic and three carriage strains, previously characterized by MLST (Didi et al., 2014) and collected from four geographic origins in France and Sweden. These whole-genome sequences were compared with data from 15 genomes available in the GenBank database to identify genetic variations specific to pathogenic or carriage strains or to CCs, antibiotic resistance genes and new putative virulence factors carried or not by MGE.
Comparative genomic analysis did not show correlation between gene content and the ability of the six strains to cause infections in humans and in a Galleria mellonella infection model. It showed CC-dependent genetic variations of some putative virulence-associated loci among 21 genomes analyzed. Surprisingly for a highly clonal population, a great diversity was observed for the ess locus, which putatively encodes a T7SS homologous to that of S. aureus in which this multiprotein complex plays a key role in pathogenesis. Indeed, it promotes chronically long-term persistence infections with abscesses, lesions and lethal outcomes in a mouse model (Burts et al., 2005, 2008).
Materials and Methods
G. mellonella Infection Experiments
Infection of G. mellonella larvae with S. lugdunensis was performed as previously described (Michaux et al., 2011). Briefly, using a syringe pump (KD Scientific, Holliston, MA, United States), larvae (about 0.3 g and 3 cm in length) were infected subcutaneously with washed cells of S. lugdunensis from an overnight culture in Brain Heart Infusion, with 3(±2) × 106 CFU per larva administered in 10 μl of sterile saline buffer. In each test, 10 insects were infected, and experiments were repeated at least four times. Larval killing was then monitored at 1-, 2- and 3-days post-infection. Sterile saline buffer was also tested under the same conditions as control. Results were analyzed using one-way analysis of variance with a Bonferroni correction with free “R” software. For all comparisons, a p value <0.05 was considered significant.
Pacific Biosciences Sequencing
The complete genome of three pathogenic and three carriage strains of S. lugdunensis previously characterized by MLST and Multi-Virulence Locus Sequence Typing (Chassain et al., 2012; Didi et al., 2014) was sequenced using Single Molecule sequencing Pacific Biosciences technology (Gupta, 2008). Strains SL13, SL29 and SL55 were, respectively, isolated from infective endocarditis, vascular prosthesis infection and skin and soft tissue infection. Carriage strains were isolated from groin (SL118 and 122) and toe (SL117) (van der Mee-Marquet et al., 2003; Bieber and Kahlmeter, 2010). Four strains belonged to CC1 (SL13, SL29, SL117, SL122) and 2 to CC6, as defined by MLST (Chassain et al., 2012; Table 1).
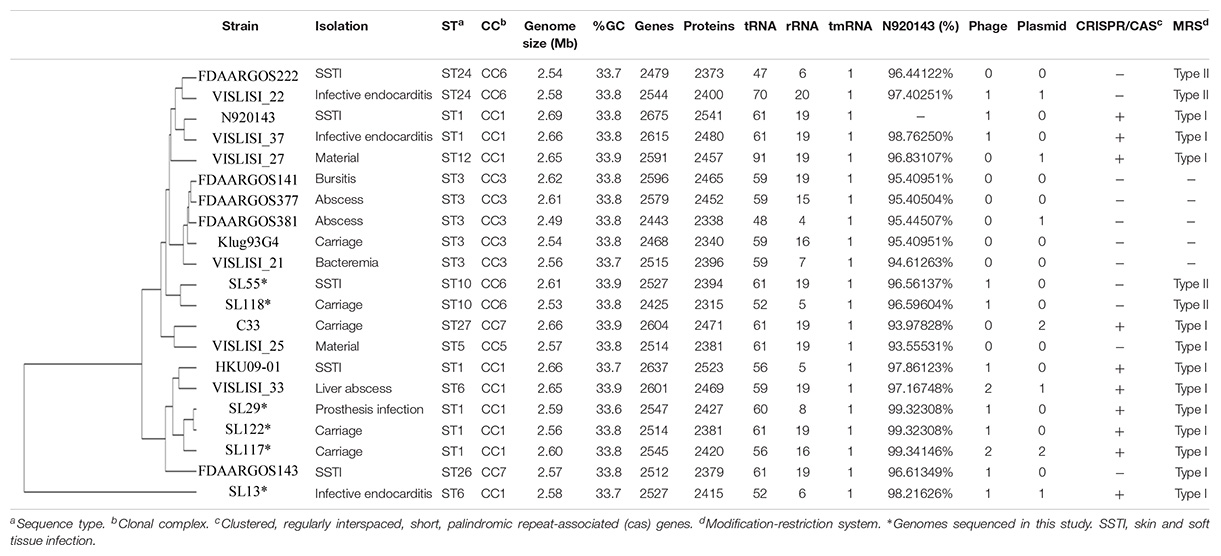
Table 1. Whole genome sequence content of the 6 S. lugdunensis strains sequenced with Pacific Biosciences technology and the 15 S. lugdunensis genomes available on NCBI.
High molecular weight genomic DNA was purified from 20-ml exponential-phase cultures by using genomic-tip-500g kit (Qiagen) following manufacturer’s instructions. DNA was precipitated in isopropanol, extensively washed with cold 75% ethanol and in 1 × Tris-EDTA buffer. Purity and quantity were assessed by using NanoDrop and Qubit assays, respectively, before size evaluation with a Tape Station (Agilent). DNA was sequenced on a Pacific Biosciences RS system at the Lausanne Genomic Technologies Facility, Center for Integrative Genomics. Using P6-C4 chemistry, libraries of approximately 8–20 kb were generated. Analyses of reads were carried out by using SMRT pipe version 2.3.0 software (Pacific Bioscience). The quality value (QV) threshold for motif calling was set at 30.
Computational Analysis
In addition to the 6 genomes sequenced by PacBio, 15 complete genomes published or available were retrieved from GenBank for comparative analyses (Supplementary Table S1). All complete genomes were annotated using the software Prokka v1.13 (Seemann, 2014). Genomes were compared using orthologsorter2 to generate ortholog clustering sequences among the overall 21 strains and to define the pan-genome, the core-, and the accessory-genome of S. lugdunensis. Genomes were also compared according to the clinical context (infection vs. carriage) and the CC of strains. A similarity matrix of complete genomes and a phylogenetic tree were created based on the neighbor-joining method with the software BioNumerics v7.6. Sequence Types (STs) and CCs defined by MLST were identified by extracting sequences of genes aroE, dat, ddl, gmk, ldh, recA and yqiL (Chassain et al., 2012). STs and CCs were obtained by submission of gene sequences into the international MLST database3. The presence of genes encoding restriction enzymes was predicted by comparison and Blastn with datasets of the Restriction Enzyme Database (Roberts et al., 2013).
Integrated prophage regions were identified and annotated by using the online PHAge Search Tool Enhanced Release (PHASTER) (Arndt et al., 2016)4 with default parameters for the six genomes sequenced by PacBio, the complete genome sequences of the 5 FDAARGOS strains and the Klug93-G4 strain, available on NCBI, and for which no phage has been described to date. PHASTER provided the region length and position of the prophages on the chromosome, their GC content and the most common related phages. Known prophage sequences ΦSL1 (Heilbronner et al., 2011), ΦSL2, ΦSL3, ΦSL4 and ΦSL5 (Argemi et al., 2017b) were compared with prophage regions identified and Mauve alignments were used to confirm PHASTER results.
Plasmid identification was done by using Prokka v1.13 for annotation of extra-chromosomic contigs obtained by PacBio sequencing and was compared to plasmid sequences retrieved from the NCBI database (pVISLISI_1, pVISLISI_2, pVISLISI_3, pVISLISI_4 pVISLISI_5 (Argemi et al., 2017b) and to unnamed plasmid from the FDAARGOS381 strain). Nucleotide sequence similarities were investigated by Blastn to identify the closest related plasmid.
Virulence factors were identified using the Virulence Factors database (VFDB)5 (Chen et al., 2016) and tBlastn searches were performed to compare the presence or absence of known virulence genes of staphylococci. Comparisons of agr and cap hypervariable regions were performed by extracting and aligning agr and cap loci with MUSCLE. Allelic agr type-I (agrSl-type I, accession number: AF173933) and type-II (agrSl-type II, accession number: AF346728) described by Dufour et al. (2002) were used as reference. The hypervariable regions (capH to capK) from S. aureus cap8 and cap5 loci were downloaded from the NCBI database. Phylogenetic trees were constructed using the maximum likelihood method implemented in the PhyML program v3.0.
Resistance genes were identified by using ResFinder v3.16 (Zankari et al., 2012). A cutoff of 80% minimum length and 80% identity were used to search for resistance genes in the 21 genomes and plasmids of S. lugdunensis.
Comparisons of individual sequences of the ess locus were performed on the 21 genomes of S. lugdunensis by using Blastn and Blastp against the NCBI database, and were facilitated by using the Artemis Comparison Tool (Carver et al., 2005). T7SS genes and proteins were identified from available genomes of HO 5096 0412 (Accession number HE861097) (Holden et al., 2013) and RJ-ST398 reference strains (Accession number AM990992) (Wang et al., 2016). Pfam database was also used to identify protein domains.
Results
G. mellonella Infection Experiments
In order to evaluate and compare the virulence of 6 S. lugdunensis strains (SL13, SL29, SL55, SL117, SL118 and SL122) under the same conditions in vivo, a G. mellonella model of infection was used.
As shown in Figure 1, carriage isolates (SL117, SL118, and SL122) as well as the pathogenic isolate SL29 (from vascular prosthesis infection) revealed moderate virulence in this model of infection. After 72 h, 31–87% of the larvae infected by these four isolates were still alive. However, they could not be clustered because statistical analysis displayed significant differences between them (p values <0.05) (Table 2). In contrast, isolates SL13 (from infective endocarditis) and SL55 (from a skin and soft tissue infection) were significantly more virulent than the others since only 4 and 8% of the infected caterpillars survived 3-days post infection, respectively (Figure 1 and Table 2). No mortality was observed for the larvae into which sterile saline buffer was injected (data not shown). Of note, no differences were observed between strains in terms of colony morphology, hemolysis (β-hemolysis on blood agar) and growth characteristics under standard laboratory conditions.
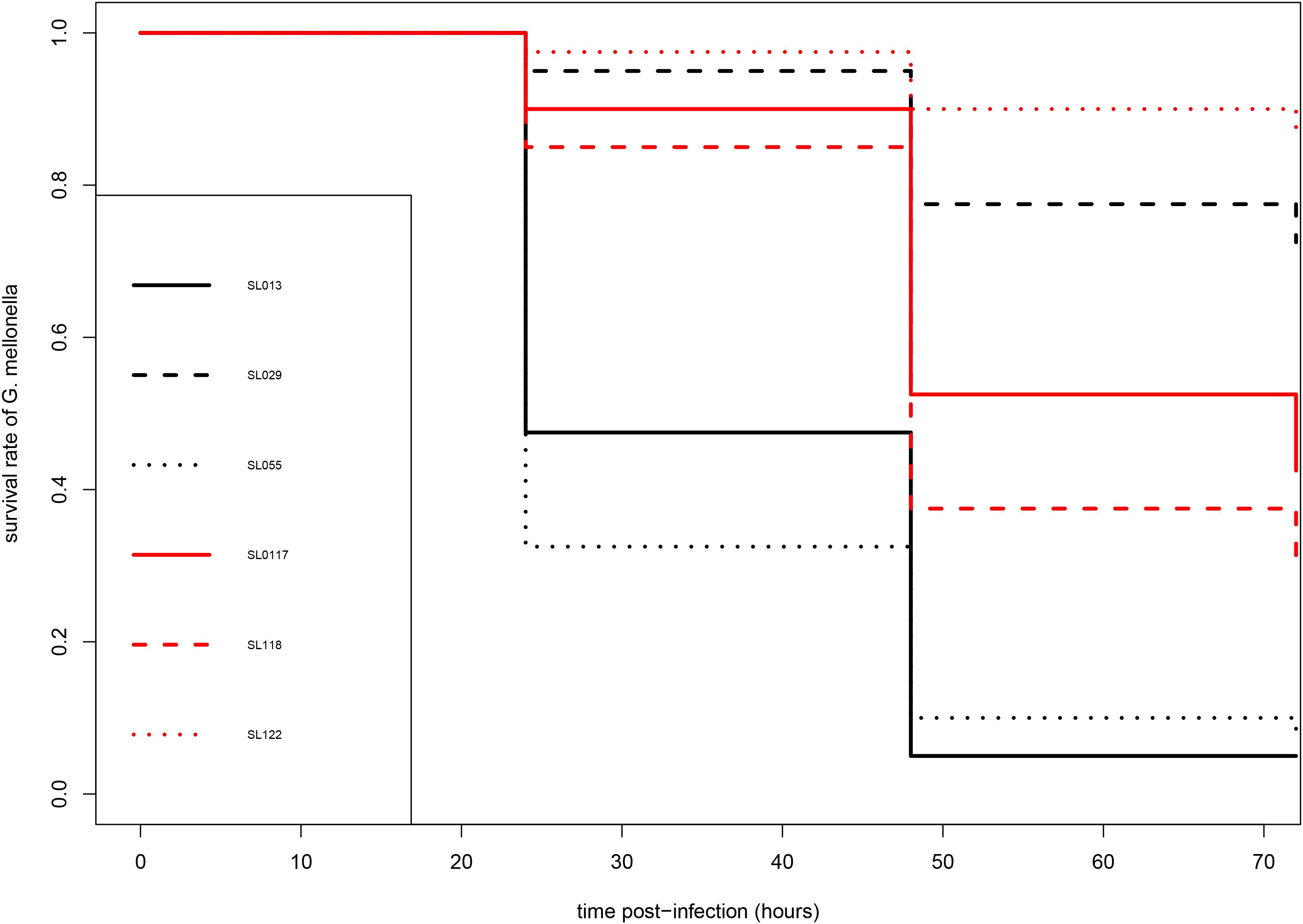
Figure 1. Kaplan-Meier survival curves of infected larvae of G. mellonella after infection by S. lugdunensis strains. Pathogenic (SL13, SL29, SL55) and carriage (SL117, SL118, SL122) are shown in black and red, respectively.
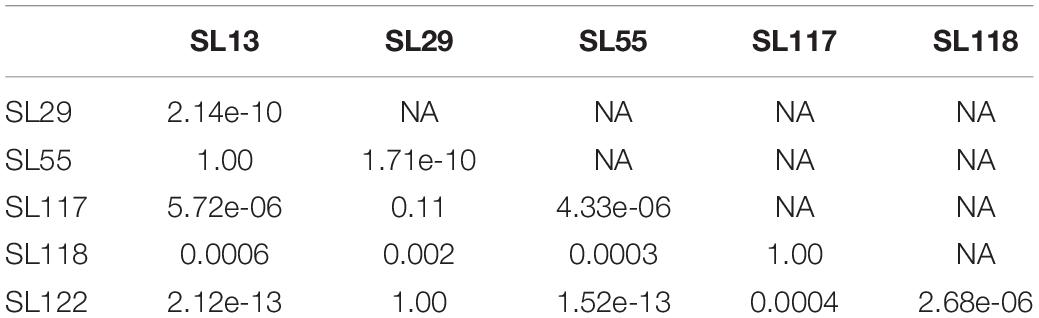
Table 2. Pair test statistical comparison (p values) of the virulence of 6 strains of S. lugdunensis in a Galleria mellonella infection model.
Pacific Biosciences Genome Sequences and Functional Genome Annotation
PacBio sequencing of 6 S. lugdunensis strains resulted in full genome recovery on a single contig with sequence length ranging from 2.53 (SL118) to 2.61 Mb (SL55). Between 67457 (SL55) and 79450 (SL118) post-filter polymerase reads were generated with an average read length ranging from 13.7 (SL55) to 14.8 kb (SL13). The N50 read length was comprised between 19.2 and 22.7 kb, and the average coverage between 272× and 367×. In addition to the contigs corresponding to genomes, 0 (SL29 and SL118) to 3 (SL117) extra-chromosomic contigs were generated.
The complete genome sequences of 15 different S. lugdunensis strains available in the NCBI database (Supplementary Table S1) were compared to the six genomes sequenced by PacBio. The size of these 21 complete genomes ranged between 2.53 (SL118) and 2.69 Mb (N920143). All strains presented similar %GC-content (33.6–33.9%). The number of genes automatically predicted by Prokka software ranged from 2443 (FDAARGOS381) to 2675 (N920143) genes encoding between 2338 and 2541 proteins. Genomic data for the 21 strains are shown in Table 1.
Pan-, Core- and Accessory-Genome Analysis
Pan-genome analysis led to identification of the total gene repertoire of the species and sets of core and accessory genes available in each S. lugdunensis genome. The pan-genome of the 21 S. lugdunensis strains was found to be composed of a total repertoire of 3077 genes. Of these, 2185 genes (71.01%) were part of the core-genome, and 892 genes (28.99%) were part of the accessory genome. Based on this analysis, no gene retrieved from the accessory genome belonged only to pathogenic or only to carriage strains. In the same way, no gene was specifically associated with strains isolated from a deep infection such as infectious endocarditis, deep abscess, or device-associated infection.
However, CC-dependent genetic variations were observed. Accessory-genome analysis of CC1 strains revealed that 18 genes were unique to these nine strains. Among them, nine genes were clustered, regularly interspaced, short, palindromic repeat (CRISPR) and loci-associated corresponding to type III-A proteins: Cas1, Cas2, Cas10, Csm2, Csm3, Csm4, Csm5, Csm6, and Cas6. The cas locus from CC1 strains showed a sequence homology of 76% with that of S. epidermidis RP62a strain. In all CC1 strains, except strains SL29 and SL122, a potential frameshift is located in the gene encoding the Csm6 protein and probably results in a non-functional CRISPR/Cas system. The remaining nine genes unique to CC1 strains were carried by a transposon and encode arsenical resistance proteins.
Eight genes were unique to CC6 strains. Six of them encode hypothetical proteins but two encode a type II restriction-modification system (RMS). Indeed, we identified a C-5 cytosine-specific DNA methylase and a Type-2 restriction enzyme Sau3AI sharing 82% identity with RMS type II from S. haemolyticus JCSJ1435 strain. Only 2 genes were found specific to CC7 strains: one encoding a DUF600 protein and one encoding a hypothetical protein. No gene was identified as specific to CC3 strains.
Mobile Genetic Elements
Five new intact prophage regions named ΦSL6 to ΦSL10 were identified by PHASTER analysis for 12 of the 21 strains (Table 3). Prophage regions ΦSL6, ΦSL7 and ΦSL8 were identified in CC1 strains (SL13, SL29, SL117, and SL122) while prophage regions ΦSL9 and ΦSL10 were found in CC6 (SL55 and SL118) and CC7 (FDAARGOS143) strains, respectively. The total length of these prophages ranged from 46.6 kb (ΦSL9) to 49.9 kb (ΦSL7); they encoded 26–69 proteins. Among these proteins, no virulence factors or antibiotic resistance-associated proteins were identified. However, we identified a putative bifunctional autolysin, a cell wall hydrolase lytN precursor, a collagen triple-helix protein and a LexA repressor which might be involved in the remodeling of the bacterial cell wall.
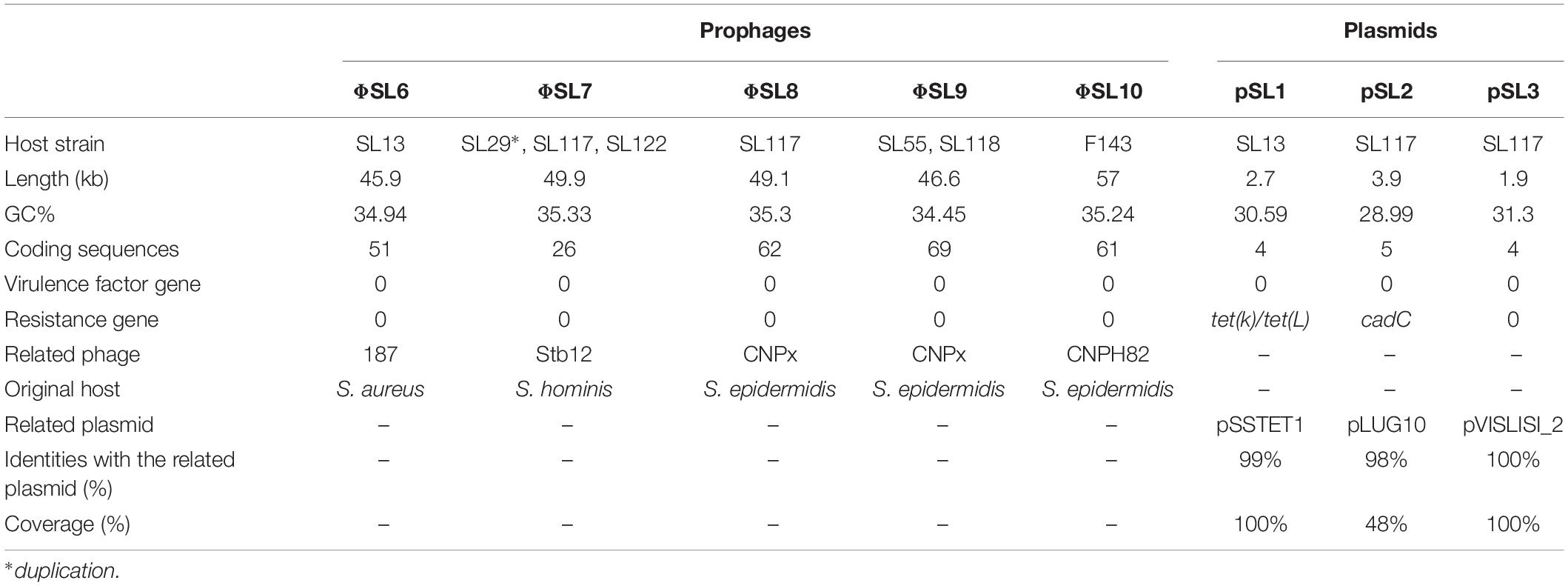
Table 3. Prophage regions and plasmids identified among 21 genomes of S. lugdunensis.
Functional annotation by using Prokka on additional contigs led to identification of three plasmids in two of the six newly sequenced genomes: pSL1 in strain SL13, and pSL2 and pSL3 in strain SL117 (Table 3). The total length of the plasmids identified ranged from 1.9 (pSL3) to 3.9 kb (pSL1); they encode between 4 and 5 proteins. Plasmids pSL2 and pSL3 respectively correspond to cadmium resistance-associated plasmid pLUG10 (Poitevin-Later et al., 1992) and plasmid pVISLISI_2 (Argemi et al., 2017b) previously described. Of note, no virulence factor was carried on these plasmids but tetracycline and cadmium resistance genes were found.
In addition, we identified a region of 13 kb length restricted to CC1 strains SL13 and VISLISI_33. Around 5 kb of this region shared a sequence homology of 94% with the pathogenicity islands SaPITokyo11212 and SaPIivm60 of S. aureus strains Tokyo11212 and IVM60, respectively. This region encodes an integrase, an excisionase, a primase and a terminase. Other interesting genes were carried by this putative pathogenicity island as those encoding Ear penicillin binding protein, FhuD ferrichrome ABC transporter and a protein homologous to part of the S. aureus Pathogenicity Island (SAPI) domain.
Resistome
Several antibiotic resistance-associated genes were identified by using the web tool ResFinder v3.0 (Table 4). A gene homologous (83% identity) to the fosB gene of Bacillus cereus AS4-12 was identified in the chromosome of all S. lugdunensis strains. A penicillin resistance-associated gene, blaZ was also identified. This gene was carried by a transposon and found in 7 of the 21 S. lugdunensis strains. The blaZ gene showed a homology ranging from 92 to 100% with the blaZ gene carried by the plasmid pSAP106A of S. epidermidis. Otherwise, annotation of the extra-chromosomal contig of SL13 strain led to identification of a tetK tetracycline resistance gene which shared 99% identity with tetK gene from the plasmid pT181 of S. aureus. Of note, no gene of methicillin resistance, within a composite staphylococcal chromosome cassette mec element, was found.
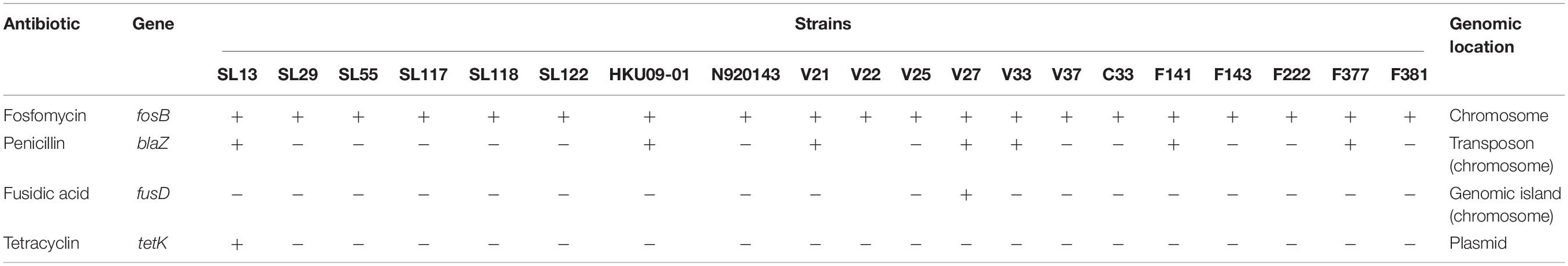
Table 4. Antibiotic resistance-associated genes identified among 21 genomes of S. lugdunensis and their genomic location.
Finally, we also identified a fusidic acid resistance gene located in a genomic island of 16 kb, only in strain VISLISI_27. This gene had 91% homology with the fusB gene carried by a genomic island (SaRIfusB) of the S. aureus CS6-EEFIC strain. In addition, in this genomic island, we identified a gene encoding a putative virulence factor, vapE (for virulence-associated protein E). vapE gene was also found in a genomic island of 40 kb in the genome of strain HKU09-01 with an identity of 85%. However, Blastn comparison of these two genomic islands showed 80% identity for only 3 kb, suggesting that strains VISLISI_27 and HKU09-01 acquired these regions independently and from different sources.
Virulome
Analysis of virulence gene composition based on the Virulence Factor Database (VFDB) showed a number of putative virulence factors that were detected in most of the S. lugdunensis isolates. To improve the identification of notable features, tblastn searches of well-characterized staphylococcal virulence factors and transcriptional regulators were performed (Table 5). No gene duplication was detected, except in isd locus for one strain (HKU09-01) and in sst locus for 20 of 21 strains.
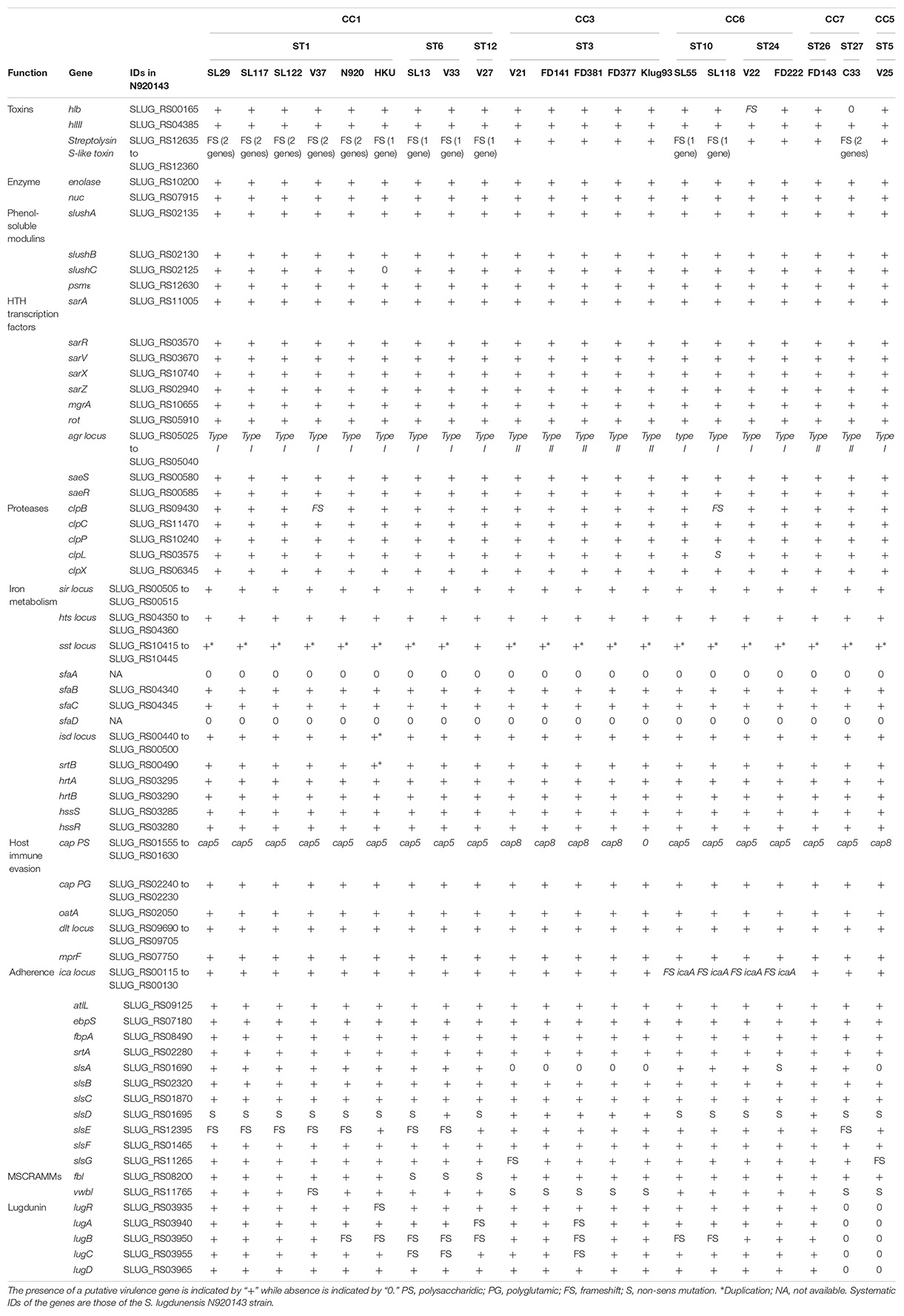
Table 5. Comparison of virulence factors among 21 S. lugdunensis strains according to clonal complexes (CCs).
The accessory gene regulator (agr) locus (agrA, agrB, agrC and agrD) was found in all S. lugdunensis strains. Phylogenetic analysis based on alignment of the hypervariable agr region (agrB to agrC) led to identification of 2 genetic groups, agrSl-types I and II (Supplementary Figure S1A). Of note, all CC3 isolates were clustered with allele agrSl-type II whereas all other strains were closer to allele agrSl-type I.
In a region of 16 kb in the genome of the 21 S. lugdunensis strains, 16 genes homologous to those encoding polysaccharidic capsule type 5 or 8 in S. aureus were detected (Supplementary Figure S1B). In S. lugdunensis, only the central region composed of capH, capI, capJ and capK genes, was found different between strains; the 12 other genes of the cap locus were conserved. Based on phylogenetic analysis, the cap locus of CC3 and VISLISI_25 strains (CC5) was closer to the cap8 locus of S. aureus while other strains were related to the cap5 locus.
In the genome of S. lugdunensis, up to seven sls genes (slsA to slsG) were identified. Comparative analysis of the predicted S. lugdunensis surface (Sls) proteins revealed important diversity between the 21 S. lugdunensis strains. The size of SlsA, varying from 1634 to 5228 amino acids, correlated with the number of IgG-like domain ranging from 8 to 45. Sequence comparison of SlsA proteins using Blastp showed 51–52% identity with the Bhp protein of S. epidermidis RP62a and 63–64% with a cell wall associated biofilm Bhp/SesD protein from Staphylococcus caprae JMUB590. Pfam analysis of these two staphylococcal proteins also showed the presence of a similar IgG-like domain, with 18 (Bhp) and 50 (Bhp/SesD) repeats. Interestingly, slsA gene was absent from the genomes of all CC3 isolates and from FDAARGOS377 isolates (CC5), and a nonsense mutation was detected in the slsA gene of the isolate FDAARGOS222 (CC6). Furthermore, in the slsD gene, a nonsense mutation located upstream of the region coding the LPXTG motif was found in 14 of the 21 S. lugdunensis strains but was not present in CC3 strains or the VISLISI_33 strain (CC1). Sequence comparison of SlsD proteins using Blastp showed an identity of 40 and 32% with SdrF and SdrG adhesins from S. epidermidis, respectively. The size of the SlsE predicted protein sequences varied from 2818 to 4166 amino acids among the 21 S. lugdunensis strains. Interestingly, a frameshift was located in the slsE gene, upstream of the region encoding the LPXTG motif and was found in all CC1 strains except the strain HKU09-01. The size of the SlsG predicted protein sequences, varying from 1604 to 2286 amino acids, correlated with the Rib repeat number ranging from 7 to 21. A frameshift was located within Rib repeats for VISLISI_21 and VISLISI_25 isolates. On the other hand, SlsB, SlsC and SlsF proteins were highly conserved (99–100% of identity) in the 21 S. lugdunensis strains.
Of note, notable diversity was observed in lug locus encoding a peptide antibiotic (lugdunin). Of the 21 strains, 8 contained a frameshift in one to 2 genes of the locus while the latter was absent in 2 strains.
Description of the CC-Dependent Genetic Diversity of the S. lugdunensis ess Locus
Comparative genomic analysis showed variations in the size of the ess locus of the 21 S. lugdunensis isolates. Indeed, ess locus size ranged from 22134 bp (HKU09-01) to 38834 bp (SL29 and SL122). This difference was the result of an important polymorphism identified in CC1 strains. For a better understanding of these variations, we divided the ess locus into 6 distinct modules, each based on gene content and relative conservation (Figure 2). Eight genetic organizations were observed.
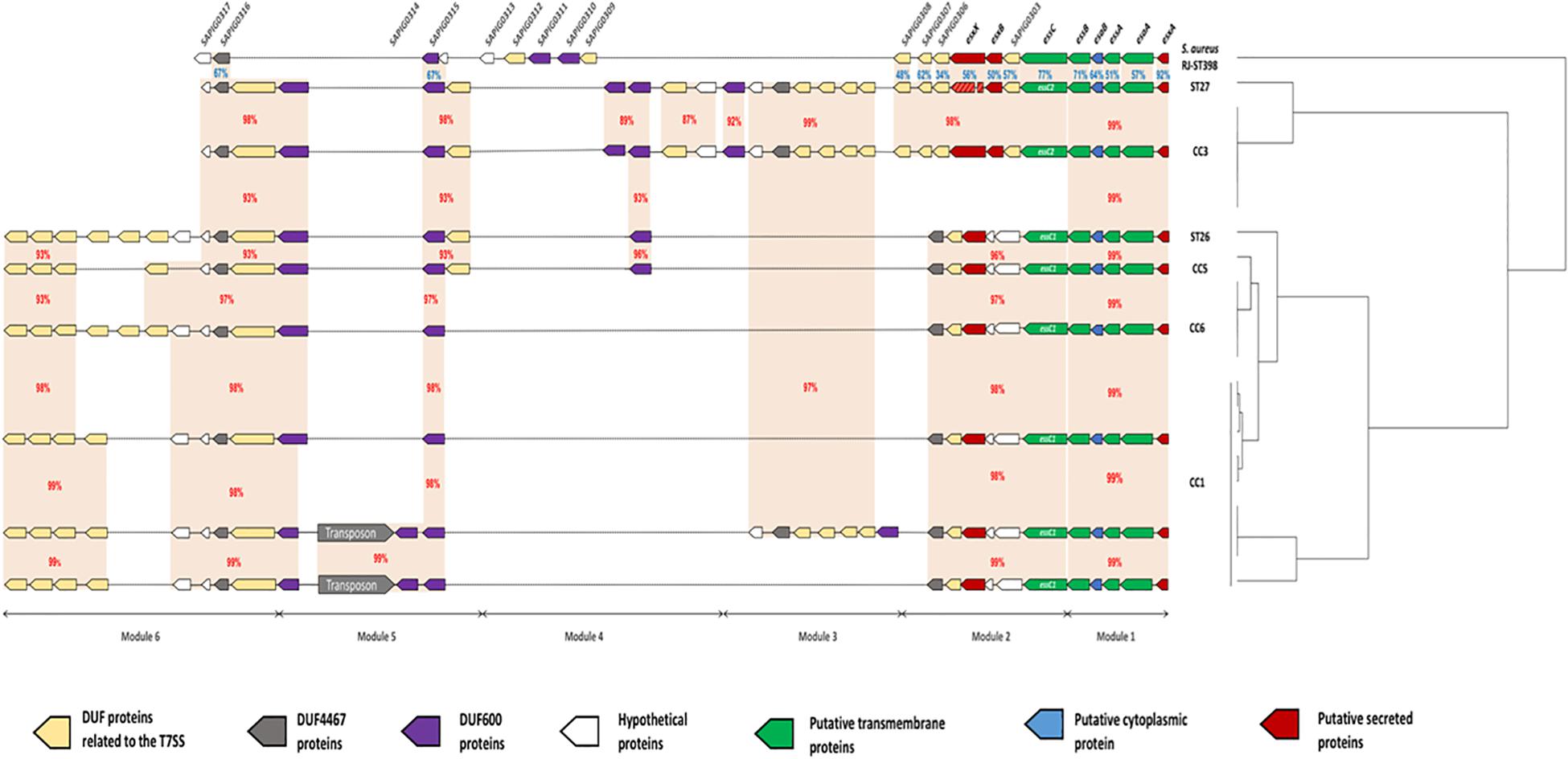
Figure 2. Schematic representation of 8 genetic organizations of the ess locus from genomes of 21 strains of S. lugdunensis. For each strain, clonal complexes (CC) are indicated to the right. The ess locus was divided into 6 modules depending on gene-based content and relative conservation defined with the Artemis comparison tool. Module 1 contained 5 genes homologous of the core components (esxA to essB) of S. aureus T7SS. Module 2 included the essC gene, of which there are 2 variants, each associated with a distinct cluster of 5–6 genes. Modules 3, 4 and 6 contained a complex arrangement of coding sequences that encode a variable number of uncharacterized proteins. Module 5 is composed of 2 distinct genetic organizations: one of them is composed of a transposon associated with 3 coding sequences encoding hypothetical DUF600 proteins whereas the other one is comprised of only 2 genes. Orange areas highlight conserved coding sequences between strains with the percentage of amino acid identity in red between S. lugdunensis strains and in blue with S. aureus strain RJ-ST398.
Module 1 contained five genes present in all sequences analyzed and homologous to genes encoding proteins of T7SS in S. aureus: EsxA, EsaA, EssA, EsaB and EssB. This module was highly conserved among all S. lugdunensis strains, with slight modifications in the amino acid sequences of these five proteins (99–100% identity).
Module 2 was variable between S. lugdunensis CCs. Indeed, 2 distinct organizations of the ess locus were identified: one shared by all CC3 strains and by one CC7 strain (C33), and the other one exhibited by other CC strains (CC1, CC5 and CC6). The first important variation concerned the essC gene for which 2 essC sequence variants (essC1 and essC2) were observed. The 5′ region of the essC gene was conserved within all S. lugdunensis isolates (3762 bp) whereas the 3′ region (around 681 bp) differed according to CCs. The essC1 variant, identified in 15 strains, encodes a putative protein of 1476 residues whereas the essC2 variant encodes a putative protein of 1481 amino acids. This latter was only identified in 5 CC3 strains and in one CC7 strain (C33). Of note, S. lugdunensis N920143 contained a frameshift in the gene encoding the membrane-associated protein EssC. Otherwise, each essC gene variant was associated with its own cluster of genes. essC1 gene was associated with five genes highly conserved among 15 S. lugdunensis isolates while essC2 gene variant was associated with six genes conserved for six isolates. Conversely, these six genes shared no identity with the five genes associated with the essC1 variant (Figure 3).
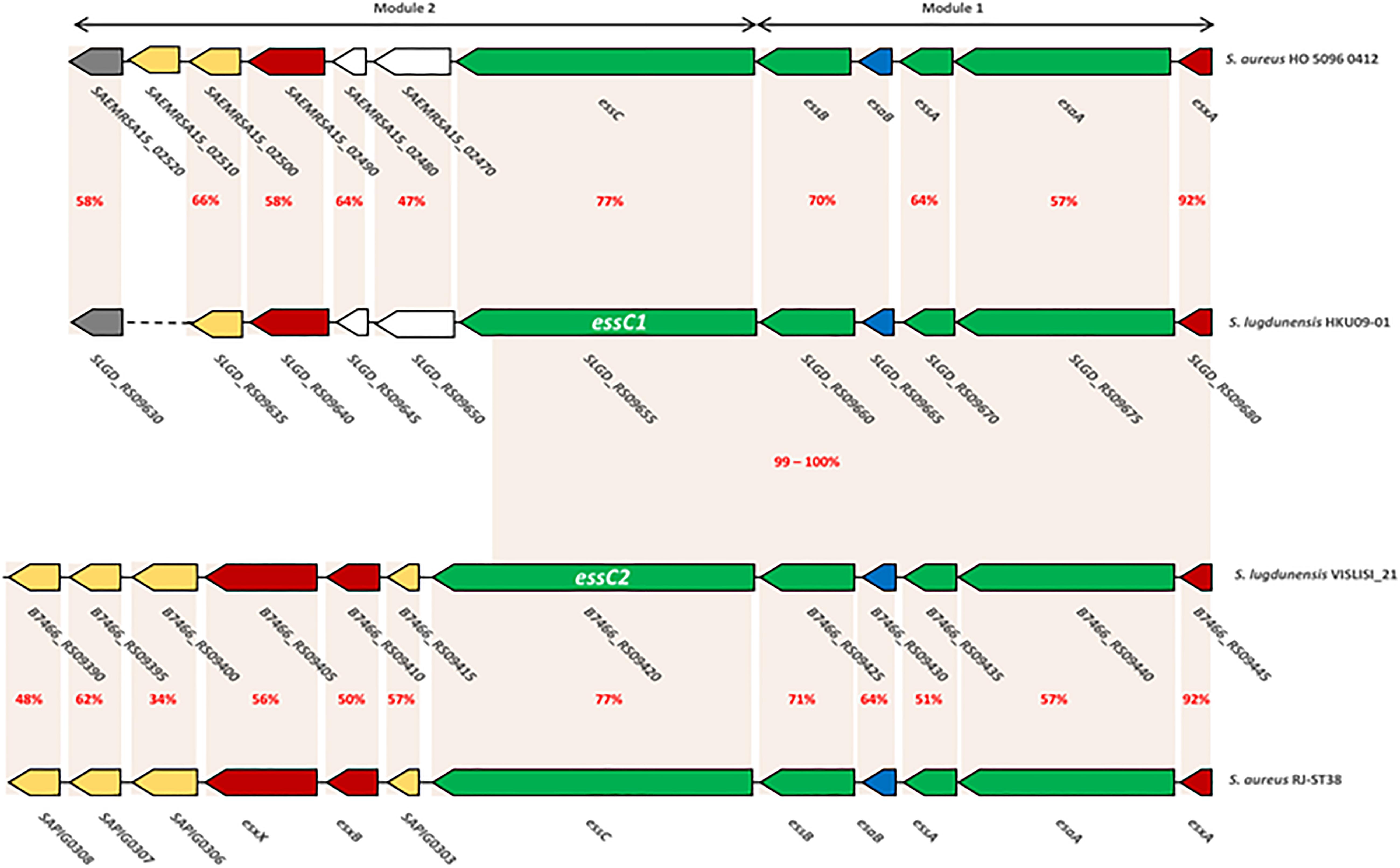
Figure 3. Comparison of ess modules 1 and 2 of S. aureus HO 5096 0412 and RJ-ST398 strains with HKU09-01 and VISLISI_21 S. lugdunensis strains. Gene-based content and relative conservation defined with the Artemis comparison tool. Orange areas highlight conserved coding sequences between strains with the percentage of amino acid identity in red.
In modules 3, 4 and 6, a complex arrangement of sequences encoding a variable number of uncharacterized proteins was observed. In particular, an important number of genes which encode proteins containing Domains of Unknown Function (DUFs) were identified. Among them, DUF600 and some others (DUF5079, DUF5080, DUF5020, DUF5084 and DUF5085) are suspected to be involved in T7SS on the basis of homology with characterized T7SS systems. The number of genes encoding DUF600 proteins varied from 2 in CC6 strains to 5 in CC3 strains while the number of genes encoding the other DUF proteins ranged from 6 to 11 among all strains (Figure 2).
In module 5, a transposon was found in 4 of the 9 CC1 isolates, associated with genes encoding hypothetical DUF600 proteins (Figure 2).
Here, the comparison focuses on modules 1 and 2 which are the only ones to have a strong homology with those of S. aureus (48–92% identity). This was performed on the one hand between the genomes of S. aureus HO 5096 0412 and S. lugdunensis HKU09-01 strains, and on the other hand between the genomes of S. aureus RJ-ST398 and S. lugdunensis VISLISI_21 strains. These 4 strains were chosen to be representative of the diversity of the locus in both species (Figure 3).
Comparative analysis of module 1 showed that S. lugdunensis genes SLGD_RS09680 (HKU09-01) and B7466_RS09445 (VISLISI_21) which encode a putative secreted protein shared 92% identity with the EsxA protein of S. aureus strains. This putative protein of 97 amino acids contains a WXG100 domain and a conserved C-terminal hydrophobic pattern for the formation of specific α-helical surface. Moreover, several putative transmembrane proteins in S. lugdunensis HKU09-01 and VISLISI_21 isolates showed homologies (51–64% identity) with the transmembrane proteins EsaA, EssA and EssB of S. aureus strains (Tables 6, 7). The EsaB cytoplasmic protein of S. aureus strains exhibited 64% identity with proteins SLGD_RS09065 (HKU09-01) and B7466_RS09420 (VISLISI_21) which contained a YukD family domain. Thus, functional analysis of T7SS core components using pfam database allowed the prediction and identification of the same protein domains for S. aureus and S. lugdunensis.
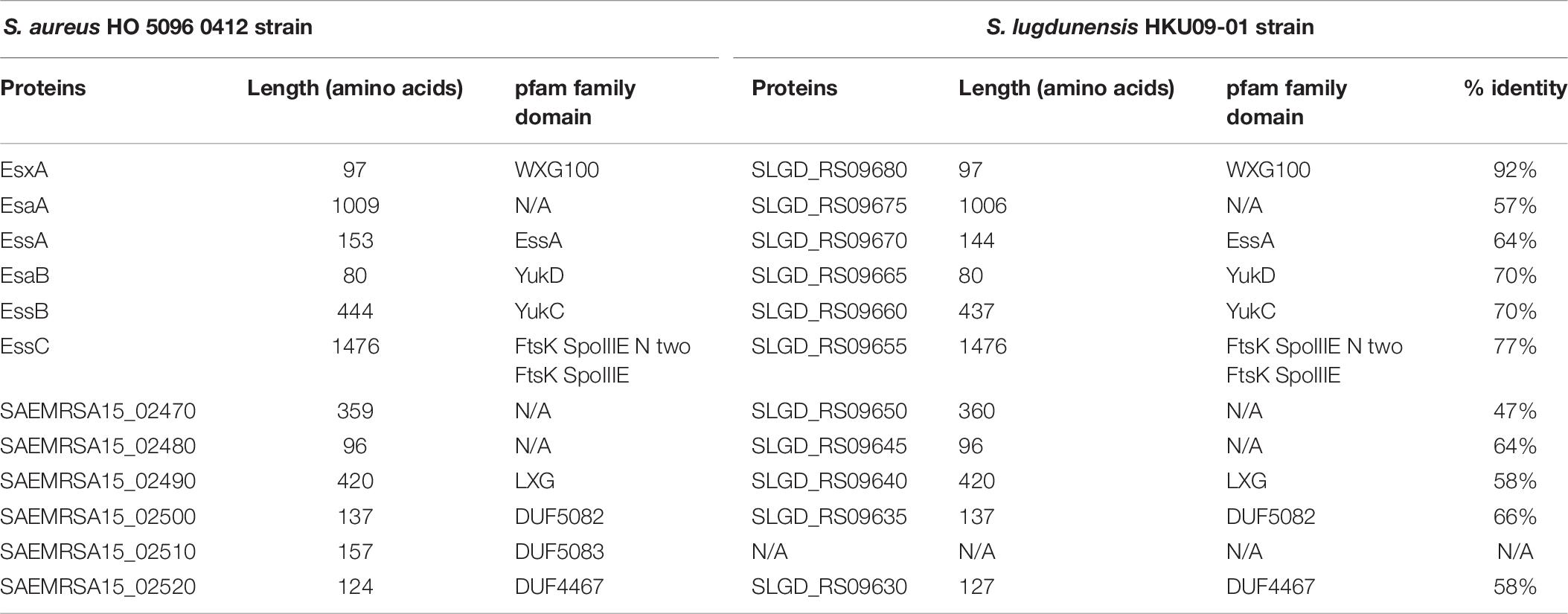
Table 6. Comparison of proteins encoded by ess modules 1 and 2 of S. aureus HO 5096 0412 with HKU09-01 S. lugdunensis strains.
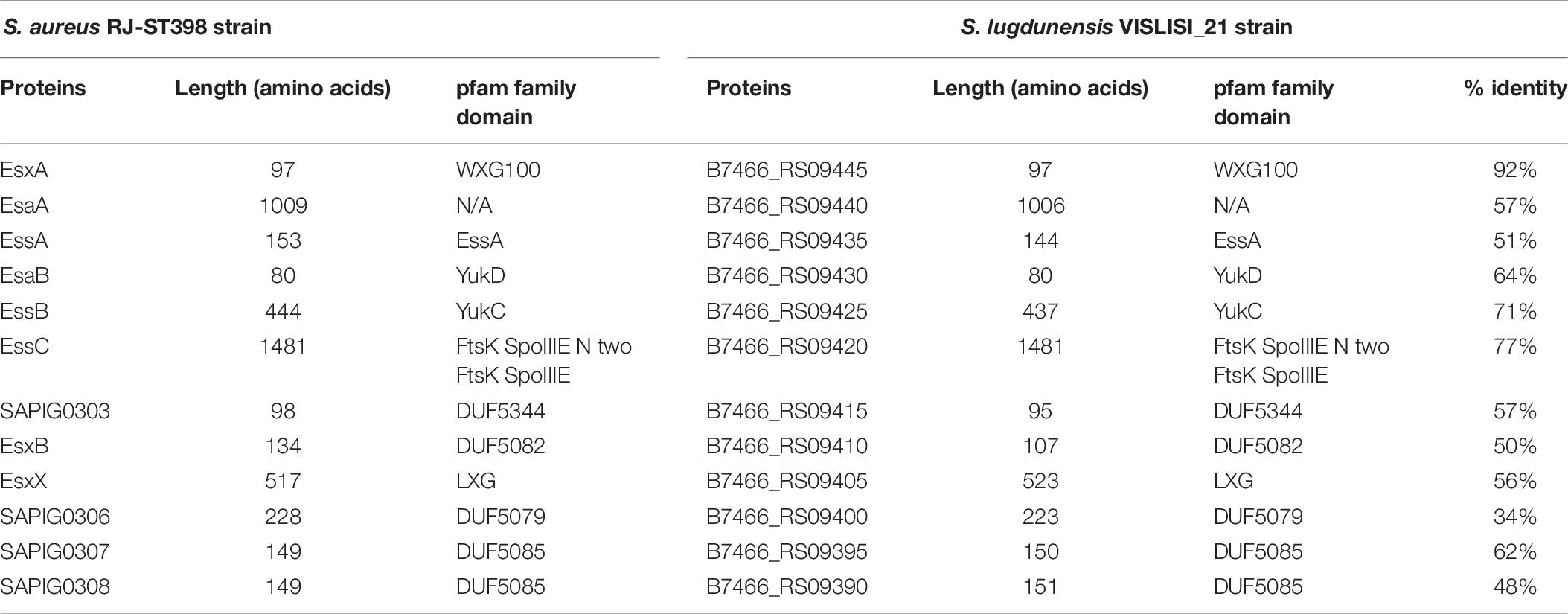
Table 7. Comparison of proteins encoded by ess modules 1 and 2 of S. aureus RJ-ST398 strains with VISLISI_21 S. lugdunensis strains.
Then, pairwise comparison of module 2 showed homologous genes and comparable organization (Figure 3 and Tables 6, 7). First, the essC1 and essC2 variants of S. lugdunensis strains encode proteins sharing 77% identity with the EssC proteins of S. aureus strains. The putative EssC proteins of S. lugdunensis contain a N-terminal FtsK-SpoIIIE domain for DNA transportation and 2 domains of the SpoIIIE-FtsK-like ATPase family, like those of S. aureus. Second, in both S. aureus and S. lugdunensis, variants of the essC gene are associated with their own gene cluster. Among putative encoded proteins, there are proteins with a N-terminal LXG domain or a central WXG motif (Tables 6, 7). For example, the putative proteins B7466_RS09410 and B7466_RS09405 of S. lugdunensis VISLISI 21 have 50% and 56% identity with the EsxB and EsxX proteins of S. aureus RJ-ST398, respectively.
Discussion
Staphylococcus lugdunensis is associated with a wide range of diseases including skin and soft tissue infections, infective endocarditis and bone and joint infections (Frank et al., 2008). This organism is considered as an opportunistic pathogen which can cause infections with unusual severity compared to other CoNS. This particular virulence is not currently explained by the rare virulence factors described so far, and no typing method (Chassain et al., 2012; Didi et al., 2014) has yet been able to identify invasive clonal complexes as described for Neisseria meningitidis (Bratcher et al., 2012) or Streptococcus pneumoniae (Dagerhamn et al., 2008).
The advent of high throughput sequencing technology offers new insights into the identification of determinants associated with bacterial virulence. Before this study, only 15 complete genomes of S. lugdunensis were available in GenBank and no comparison between pathogenic and carriage strains had been conducted. In this context, we sequenced the genome of 6 new strains (3 pathogenic and 3 carriage strains) whose virulence was characterized in a G. mellonella infection model.
This model has been applied to characterize the virulence and pathogenesis of a wide range of microorganisms (Kavanagh and Reeves, 2004; Ramarao et al., 2012) but has never been implemented for S. lugdunensis. Interestingly, our results suggest that this model could be useful for assessing the virulence of S. lugdunensis in vivo as the SSTI-associated strain (SL55) and the infective endocarditis strain (SL13) were significantly more lethal than the four other isolates. Although insects lack an adaptative immune response, their innate immune response still retains remarkable similarities with the immune response in vertebrates; this allowed to obtain relevant information about the infection process (Kavanagh and Reeves, 2004). In addition, infected larvae can be incubated at 37°C, which favors the study of human pathogenic bacteria (Kavanagh and Reeves, 2004). Of course, these preliminary results must be confirmed in a larger collection of isolates. Although it will not replace mammalian models, the larvae of G. mellonella provide a fast and cost-effective alternative to collect initial data.
With the addition of six new S. lugdunensis genomes, this study has raised the total number of complete genomes available for comparative purposes from 15 to 21. This result has increased the genetic diversity of the genomic data for this species as these strains were collected from one Swedish and 3 different French cities, whilst 7 of the 15 genomes in GenBank originated from a unique French location (Argemi et al., 2018).
Here, we have characterized the pan-, core- and accessory-genome of the species, particularly according to CCs defined by MLST. Furthermore, we have described the first genomic comparison of carriage and pathogenic strains representative of the major lineages of S. lugdunensis to explore whether virulence can be linked to specific genes or genetic variations.
Presence of MGE Despite a Closed Pan-Genome
Comparative genomic analysis favored a closed pan-genome, as suggested by Heaps’ law of extrapolation applied by Argemi et al. (2018). The S. lugdunensis pan-genome appeared in fact to be poorly expanding with the inclusion of six new complete genomes reaching a total of only 3077 genes. On the contrary, S. aureus and S. epidermidis pan-genomes reached more than 3800 genes, with a constant increase when adding new genomes (Argemi et al., 2018). This should be confirmed by the analysis of human S. lugdunensis strains from other continents but also of environmental and animal origin.
In this context, it is not surprising that a large number of S. lugdunensis strains have barriers to horizontal gene transfer like CRISPR/Cas system or RMS (Argemi et al., 2018). Our study shows that 48% of strains possess a CRISPR/Cas system. These findings confirm the particular status of this species among CoNS. Indeed, Rossi et al. (2017) showed that only 9% (i.e., 3/34) and 3% (i.e., 1/32) of S. epidermidis and Staphylococcus haemolyticus strains respectively presented characteristic CRISPR/Cas systems. Of note, the CRISPR/Cas system was only found in CC1 strains. This high prevalence may be explained by the fact that most of the sequenced genomes belonged to CC1 strains (9/21, i.e., 43%). This CC is currently over-represented in NCBI in comparison with other CCs (CC3 [5/21, i.e., 24%], CC5 [1/21, i.e., 4%], CC6 [4/21, i.e., 19%] and CC7 [2/21, i.e., 10%]) in relation to its global distribution (Yeh et al., 2016; Dahyot et al., 2019). It would be interesting to sequence the whole genome of CC2 and CC4 strains, as none are available in the GenBank database.
Despite the presence of CRISPR/Cas immunity systems and several type I and type II RMS, most (15/21, i.e., 71%) of the S. lugdunensis strains have acquired MGE probably from other staphylococcal species by horizontal gene transfer. This was particularly observed for CC1 strains. This latter could be explained by a frameshift within the csm6 gene that might impact the initial role of the CRISPR/Cas system. Indeed, in S. epidermidis, the protein Csm6 was found essential for preventing plasmid transformation (Hatoum-Aslan et al., 2014; Jiang et al., 2016). Further investigations in a larger collection of S. lugdunensis strains and functional characterization are needed to understand the ability of CRISPR/Cas to prevent MGE acquisition. Another hypothesis would be that MGE may have been present before acquisition of the CRISPR/Cas region. Enriching the database with genomes of ancestral strains would help to resolve this issue.
On the contrary, while CC3 strains do not exhibit CRISPR/Cas systems or RMS, only one plasmid was detected within their genome. This can be related to the great difficulties encountered in genetically manipulating S. lugdunensis. The rare mutant strains published were only constructed after the introduction of plasmids by protoplast transformation (Szabados et al., 2011; Marlinghaus et al., 2012) or by electroporation of Escherichia coli strains expressing hsdM/hsdS RMS genes of S. lugdunensis. Interestingly, despite a new electroporation protocol, Heilbronner et al. (2013) highlighted that CC3 and CC4 isolates were harder to transform than CC1, CC2 and CC5 strains. This suggests that other mechanisms may play the role of barrier against the acquisition of MGE in S. lugdunensis. Bacteria can display physical barriers such as extracellular polysaccharidic substances, including capsules to prevent phage infection (Wilkinson and Holmes, 1979; Labrie et al., 2010). Thus, cap8 could be involved in the synthesis of capsule in S. lugdunensis CC3 strains and act as a protective barrier.
S. lugdunensis exhibits a highly conserved antibiotic susceptibility to most anti-staphylococcal antibiotics, which may be related to these different barriers against the integration of foreign DNA (Higaki et al., 1999; McHardy et al., 2017). Indeed, only one plasmid carrying an antibiotic resistance gene (tetK) was identified in a newly sequenced genome. Fosfomycin, penicillin and fusidic acid resistance-associated genes were identified with a chromosomic location. Fosfomycin resistance-associated gene was highly conserved among the 21 strains while the distribution of penicillin and fusidic acid resistance genes was variable between isolates. This finding is in accordance with the results of McHardy et al. (2017). who showed that the blaZ gene was detected in 39.1% of oxacillin-susceptible S. lugdunensis isolates. The presence of the blaZ gene was not correlated with CCs.
In this study, no specific virulence determinant was identified in S. lugdunensis pathogenic isolates compared to carriage strains. Nevertheless, two genomic islands were only identified in three pathogenic strains. One genomic island of 16 kb carries fusidic acid resistance fusB gene and vapE gene which might contribute to bacterial adaptation and virulence (Novick et al., 2010; Chen et al., 2013). A second one of 13 kb, partially homologous to the pathogenicity islands of S. aureus (SaPI) (Novick and Subedi, 2007), was found in S. lugdunensis strains SL13 (causing infective endocarditis) and VISLISI_33 (causing liver abscess). In S. aureus, SaPIs play an important role in genomic evolution, leading to an increase in its ability to cause severe infections by acquisition of numerous virulence determinants such as pore-forming toxins and superantigens (Malachowa and DeLeo, 2010; Novick and Ram, 2017). In S. lugdunensis, no enterotoxin, superantigen or toxic shock syndrome toxin-1 was found in this region. However, an Ear penicillin binding protein and a FhuD ferrichrome ABC transporter were carried by this potential pathogenicity island. S. aureus Ear protein may act as a superantigenic exoprotein which could play a role in virulence (Singh et al., 2017). Otherwise, the ferric hydroxamate uptake D (FhuD) protein could be involved in iron acquisition by S. lugdunensis as it can promote ferrisiderophore import in S. aureus (Beasley et al., 2009) and facilitate heme uptake in blood and serum (Malachowa et al., 2011). Therefore, identification of these genes in the genomes of S. lugdunensis suggests that genomic island, other than MGE as bacteriophage or plasmids, could also contribute to the acquisition of antibiotic resistance and virulence genes in this species even if the phenomenon is probably rare (Argemi et al., 2018).
CC-Dependent Genetic Variations of Putative Virulence-Associated loci
Based on genomic comparison using VFDB, we identified CC-dependent polymorphisms in several loci (agr, cap, sls and ess) putatively associated with S. lugdunensis virulence.
Our study shows that all CC3 strains belonged to the agrSl-type II whereas all other strains were associated with the allele agrSl-type I. However, no link has been shown between S. lugdunensis agr polymorphisms (agrSl-types) and specific clinical infection entities. Many studies have attempted to associate agr types with one or more of the biological characteristics of staphylococcal species. Tseng et al. (2015) thus showed that 7 of 11 isolates collected from bacteremia belonging to agrSl-type II presented higher hemolytic and protease activities than 4 agrSl-type I isolates. In S. aureus, agr groups have been shown to vary by clonal lineage distribution, antibiotic resistance profile, biofilm formation, expression of virulence factors, and autoinducing peptide structures (Holtfreter et al., 2007; Tan et al., 2018). In addition, Jarraud et al. (2000) have shown that S. aureus endocarditis strains were mainly associated with phylogenetic group AF2 (agr groups I or II) while phylogenetic group AF1 (agr group IV) strains are mainly responsible for generalized exfoliative syndromes and bullous impetigo. Similarly, agr-specificity groups in S. epidermidis are also associated with specific clinical infections (Hellmark et al., 2013; Harris et al., 2017). The potential impact of this CC-agrSl-type association on the regulation of virulence could be explored by a functional analysis of the agr locus in S. lugdunensis.
Interestingly, CC3 strains are also distinguished from all other strains by the allele of the cap gene (cap8). Except for the study of Lambe et al. (1994) that showed that S. lugdunensis possessed glycocalyx, no work has yet described the prevalence, composition, diversity and role of the capsule in this species. In S. aureus, several studies have shown that serotype 5 or 8 capsular isolates account for approximately 80% of isolates recovered from humans, the remaining isolates being not encapsulated or non-typeable (Arbeit et al., 1984; Roghmann et al., 2005; Cocchiaro et al., 2006). Like S. aureus, the genes specifying capsular polysaccharides in S. lugdunensis are encoded by a highly conserved cap locus including 4 genes, capHIJK, which may specify chemical diversity among serotypes (O’Riordan and Lee, 2004).
Heilbronner et al. (2011) described 7 genes encoding “S. lugdunensis surface proteins” named SlsA to SlsG within the genome of S. lugdunensis N920143 strain. Here, we have shown that several Sls proteins were absent or potentially non-functional in some S. lugdunensis strains according to CC. These CC-dependent differences could result in variations in the biofilm-forming and adhesion capacities between isolates. Indeed, SlsA contains putative protein domains and structural organization homologous to a known cell wall-associated biofilm Bhp protein (Post et al., 2017), which has been shown to promote primary attachment to abiotic surfaces as well as intercellular adhesion during biofilm formation (Cucarella et al., 2001; Bowden et al., 2005). In addition, the SlsD protein has putative structural similarity with members of the Sdr adhesin family such as SdrF and SdrG, crucial for the attachment of S. epidermidis to surfaces coated with human fibrinogen or type I collagen (Bowden et al., 2005; Arrecubieta et al., 2007). A functional characterization of these Sls proteins could be helpful to explore whether they are involved in the ability of S. lugdunensis to bind medical devices or biotic surfaces, such as host tissues.
On the other hand, significant genetic variations independent of CCs were detected in the lug locus. The impact of this diversity on the production and activity of lugdunine as well as on the structure of the microbiota of ecological niches of S. lugdunensis remains to be demonstrated (Zipperer et al., 2016).
High Genetic Diversity of the ess/Type VII Locus, a Potential Key Virulence Determinant
This study provides, to the best of our knowledge, the first description of the CC-dependent genetic organization of the ess locus from S. lugdunensis. Initially mentioned for one strain of S. lugdunensis (Warne et al., 2016), this locus has shown considerable diversity for a clonal species (eight genetic CC-dependent organizations among only 21 strains). Homologous genes of those encoding the six core components of S. aureus T7SS were identified among the 21 genomes: the four membrane-associated proteins (EsaA, EssA, EssB, and EssC), a soluble cytosolic protein (EsaB) and the Early Secreted Antigenic Target-6 kDa (ESAT-6, also termed EsxA) (Warne et al., 2016; Unnikrishnan et al., 2017). Membrane and cytoplasmic proteins have been demonstrated to be necessary to export effector proteins across the bacterial membrane during host infection (Unnikrishnan et al., 2017), as proteins from the superfamily WXG100 (Burts et al., 2005) or LXG domain-containing proteins (Dai et al., 2017).
Whereas S. aureus T7SS can secrete two WXG100 proteins (EsxA, EsxB), only one member of the WXG100 protein superfamily, EsxA, has been identified in S. lugdunensis. As for S. aureus EsxA, the putative S. lugdunensis EsxA protein contains conserved C-terminal hydrophobic patterns HxxxD/ExxhxxxH (where H stands for highly conserved hydrophobic, h for less conserved hydrophobic residues, x for any amino acid and D/E for either aspartic or glutamic acids, respectively). This domain could constitute a key component of T7SS-substrate recognition (Poulsen et al., 2014).
In addition, S. lugdunensis possesses a gene encoding a LXG domain-containing protein which is found in a group of polymorphic toxins predicted to use the T7SS pathway (Zhang et al., 2011; Whitney et al., 2017). Thereby, these data highlight the possible extracellular translocation of these proteins. In S. aureus, proteins with domain LXG are involved in immune evasion and virulence as the protein EsxX (Dai et al., 2017) or in bacterial competition as the EsaD nuclease (Anderson et al., 2013; Cao et al., 2016). Of note, no gene homologous to the esaD gene encoding the anti-bacterial toxin with a C-terminal nuclease domain was identified within the genome of S. lugdunensis strains.
A great diversity in the genetic organization of the ess locus was described by Warne et al. (2016) in 153 S. aureus isolates belonging to different CCs. They showed that the S. aureus essC gene exhibited 4 sequence variants, each associated with a specific cluster of 4–6 different genes. In S. aureus, the essC4 variant (homologous to the S. lugdunensis essC1 variant) was only found in CC22 isolates whereas the essC2 variant (homologous to the S. lugdunensis essC2 variant) was identified in CC15 and ST398 strains. In S. lugdunensis, a strong association between CCs and essC variants was also observed. These results suggest that S. lugdunensis strains may have acquired these regions independently and from difference sources.
Moreover, genetic diversity was observed downstream of modules 1 and 2, based on (i) a variable number of genes encoding uncharacterized proteins, (ii) the presence of a cluster of 7 genes specific to CC1 and CC3, (iii) the insertion of a transposon in CC1 strains, and (iv) the variable copy number of genes encoding DUF600 proteins. The mechanisms of occurrence of these inter- and intra-CC variations are unclear and need further investigations in a larger collection of S. lugdunensis strains, and especially in strains belonging to CC2 and CC4 which could present different organizations of this locus.
Thus, we describe here homologs of a large number of genes encoding T7SS in S. lugdunensis which shares a high number of clinical features with S. aureus, including ability to form abscesses. S. lugdunensis is the sole CoNS to possess all the components required for functional Ess machinery. T7SS could play a key role in abscess development and hence in the currently poorly understood virulence of this bacterial species.
Taken together, these results support the “closed” state of the pan-genome of S. lugdunensis, but identification of MGE carrying putative virulence and resistance genes suggests some possible horizontal gene transfers between S. lugdunensis and other staphylococcal species. No specific variation or virulence determinant was associated with the pathogenic strains, but the distribution of potential genomic islands constitutes an interesting option to explore in a larger collection of strains. Here, we identified several CC-dependent variations, especially for CC3 strains which showed specific variations in several loci potentially associated with virulence. Further phenotypic and functional studies are needed to fully characterize this particular CC and to evaluate the role of T7SS in the virulence of S. lugdunensis. Finally, whole genome sequencing of CC2 and CC4 strains is needed for a complete picture of the genetic diversity of S. lugdunensis.
Data Availability Statement
The datasets generated for this study can be found in the GenBank database, accession: CP041722, CP041723, CP041724, CP041725, CP041726, CP041727.
Author Contributions
JL, SDa, PF, and MP-C designed the study. JL, SDi, and AP analyzed the data. J-CG and MA performed the Galleria mellonella infection experiments. JL, SDa, and MP-C wrote the manuscript. All authors have read and approved the final version of the manuscript.
Funding
This study was supported by an educational grant for JL from the Charles Nicolle Foundation.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors are grateful to Nikki Sabourin-Gibbs (Rouen University Hospital) for her help in editing the manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02479/full#supplementary-material
Footnotes
- ^ http://bigsdb.web.pasteur.fr/staphlugdunensis
- ^ https://git.facom.ufms.br/bioinfo/orthologsorter
- ^ https://bigsdb.pasteur.fr/staphlugdunensis/
- ^ http://phaster.ca
- ^ http://www.mgc.ac.cn/VFs
- ^ https://cge.cbs.dtu.dk/services/ResFinder
References
Anderson, M., Aly, K. A., Chen, Y.-H., and Missiakas, D. (2013). Secretion of atypical protein substrates by the ESAT-6 secretion system of Staphylococcus aureus: ESAT-6 secretion in S. aureus. Mol. Microbiol. 90, 734–743. doi: 10.1111/mmi.12395
Anguera, I. (2005). Staphylococcus lugdunensis infective endocarditis: description of 10 cases and analysis of native valve, prosthetic valve, and pacemaker lead endocarditis clinical profiles. Heart 91:e10. doi: 10.1136/hrt.2004.040659
Arbeit, R. D., Karakawa, W. W., Vann, W. F., and Robbins, J. B. (1984). Predominance of two newly described capsular polysaccharide types among clinical isolates of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2, 85–91. doi: 10.1016/0732-8893(84)90002-6
Argemi, X., Dahyot, S., Lebeurre, J., Hansmann, Y., Ronde Oustau, C., and Prévost, G. (2017a). Staphylococcus lugdunensis small colony variant conversion resulting in chronic prosthetic joint infection. Méd. Mal. Infect. 47, 498–501. doi: 10.1016/j.medmal.2017.05.005
Argemi, X., Martin, V., Loux, V., Dahyot, S., Lebeurre, J., Guffroy, A., et al. (2017b). Whole genome sequencing of 7 strains of Staphylococcus lugdunensis allows identification of mobile genetic elements. Genome Biol. Evol. 9:77. doi: 10.1093/gbe/evx077
Argemi, X., Matelska, D., Ginalski, K., Riegel, P., Hansmann, Y., Bloom, J., et al. (2018). Comparative genomic analysis of Staphylococcus lugdunensis shows a closed pan-genome and multiple barriers to horizontal gene transfer. BMC Genom. 19:621. doi: 10.1186/s12864-018-4978-1
Arias, M., Tena, D., Apellániz, M., Asensio, M. P., Caballero, P., Hernández, C., et al. (2010). Skin and soft tissue infections caused by Staphylococcus lugdunensis: report of 20 cases. Scand. J. Infect. Dis. 42, 879–884. doi: 10.3109/00365548.2010.509332
Arndt, D., Grant, J. R., Marcu, A., Sajed, T., Pon, A., Liang, Y., et al. (2016). PHASTER: a better, faster version of the PHAST phage search tool. Nucleic Acids Res. 44, W16–W21. doi: 10.1093/nar/gkw387
Arrecubieta, C., Lee, M.-H., Macey, A., Foster, T. J., and Lowy, F. D. (2007). SdrF, a Staphylococcus epidermidis surface protein, binds type I collagen. J. Biol. Chem. 282, 18767–18776. doi: 10.1074/jbc.M610940200
Beasley, F. C., Vinés, E. D., Grigg, J. C., Zheng, Q., Liu, S., Lajoie, G. A., et al. (2009). Characterization of staphyloferrin a biosynthetic and transport mutants in Staphylococcus aureus. Mol. Microbiol. 72, 947–963. doi: 10.1111/j.1365-2958.2009.06698.x
Bieber, L., and Kahlmeter, G. (2010). Staphylococcus lugdunensis in several niches of the normal skin flora. Clin. Microbiol. Infect. 16, 385–388. doi: 10.1111/j.1469-0691.2009.02813.x
Bocher, S., Tonning, B., Skov, R. L., and Prag, J. (2009). Staphylococcus lugdunensis, a common cause of skin and soft tissue infections in the community. J. Clin. Microbiol. 47, 946–950. doi: 10.1128/JCM.01024-08
Bowden, M. G., Chen, W., Singvall, J., Xu, Y., Peacock, S. J., Valtulina, V., et al. (2005). Identification and preliminary characterization of cell-wall-anchored proteins of Staphylococcus epidermidis. Microbiol. Read. Engl. 151, 1453–1464. doi: 10.1099/mic.0.27534-0
Bratcher, H. B., Bennett, J. S., and Maiden, M. C. J. (2012). Evolutionary and genomic insights into meningococcal biology. Future Microbiol. 7, 873–885. doi: 10.2217/fmb.12.62
Burts, M. L., DeDent, A. C., and Missiakas, D. M. (2008). EsaC substrate for the ESAT-6 secretion pathway and its role in persistent infections of Staphylococcus aureus. Mol. Microbiol. 69, 736–746. doi: 10.1111/j.1365-2958.2008.06324.x
Burts, M. L., Williams, W. A., DeBord, K., and Missiakas, D. M. (2005). EsxA and EsxB are secreted by an ESAT-6-like system that is required for the pathogenesis of Staphylococcus aureus infections. Proc. Natl. Acad. Sci. U. S. A. 102, 1169–1174. doi: 10.1073/pnas.0405620102
Cao, Z., Casabona, M. G., Kneuper, H., Chalmers, J. D., and Palmer, T. (2016). The type VII secretion system of Staphylococcus aureus secretes a nuclease toxin that targets competitor bacteria. Nat. Microbiol. 2:16183. doi: 10.1038/nmicrobiol.2016.183
Carver, T. J., Rutherford, K. M., Berriman, M., Rajandream, M.-A., Barrell, B. G., and Parkhill, J. (2005). ACT: the artemis comparison tool. Bioinform. Oxf. Engl. 21, 3422–3423. doi: 10.1093/bioinformatics/bti553
Chassain, B., Lemee, L., Didi, J., Thiberge, J.-M., Brisse, S., Pons, J.-L., et al. (2012). Multilocus sequence typing analysis of staphylococcus lugdunensis implies a clonal population structure. J. Clin. Microbiol. 50, 3003–3009. doi: 10.1128/JCM.00988-12
Chen, H.-J., Chang, Y.-C., Tsai, J.-C., Hung, W.-C., Lin, Y.-T., You, S.-J., et al. (2013). New structure of phage-related islands carrying fusb and a virulence gene in fusidic acid-resistant Staphylococcus epidermidis. Antimicrob. Agents Chemother. 57, 5737–5739. doi: 10.1128/AAC.01433-13
Chen, L., Zheng, D., Liu, B., Yang, J., and Jin, Q. (2016). VFDB 2016: hierarchical and refined dataset for big data analysis—10 years on. Nucleic Acids Res. 44, D694–D697. doi: 10.1093/nar/gkv1239
Cocchiaro, J. L., Gomez, M. I., Risley, A., Solinga, R., Sordelli, D. O., and Lee, J. C. (2006). Molecular characterization of the capsule locus from non-typeable Staphylococcus aureus. Mol. Microbiol. 59, 948–960. doi: 10.1111/j.1365-2958.2005.04978.x
Cucarella, C., Solano, C., Valle, J., Amorena, B., Lasa, I., and Penadés, J. R. (2001). Bap, a Staphylococcus aureus surface protein involved in biofilm formation. J. Bacteriol. 183, 2888–2896. doi: 10.1128/JB.183.9.2888-2896.2001
Dagerhamn, J., Blomberg, C., Browall, S., Sjöström, K., Morfeldt, E., and Henriques-Normark, B. (2008). Determination of accessory gene patterns predicts the same relatedness among strains of Streptococcus pneumoniae as sequencing of housekeeping genes does and represents a novel approach in molecular epidemiology. J. Clin. Microbiol. 46, 863–868. doi: 10.1128/JCM.01438-07
Dahyot, S., Lebeurre, J., Argemi, X., François, P., Lemée, L., Prévost, G., et al. (2018). Multiple-locus variable number tandem repeat analysis (MLVA) and tandem repeat sequence typing (TRST), helpful tools for subtyping Staphylococcus lugdunensis. Sci. Rep. 8:11669. doi: 10.1038/s41598-018-30144-y
Dahyot, S., Lebeurre, J., Laumay, F., Argemi, X., Dubos, C., Lemée, L., et al. (2019). fbl-typing of Staphylococcus lugdunensis: a frontline tool for epidemiological studies, but not predictive of fibrinogen binding ability. Front. Microbiol. 10:1109. doi: 10.3389/fmicb.2019.01109
Dai, Y., Wang, Y., Liu, Q., Gao, Q., Lu, H., Meng, H., et al. (2017). A Novel ESAT-6 secretion system-secreted protein esxx of community-associated Staphylococcus aureus lineage ST398 contributes to immune evasion and virulence. Front. Microbiol. 8:819. doi: 10.3389/fmicb.2017.00819
Didi, J., Lemee, L., Gibert, L., Pons, J.-L., and Pestel-Caron, M. (2014). Multi-virulence-locus sequence typing of Staphylococcus lugdunensis generates results consistent with a clonal population structure and is reliable for epidemiological typing. J. Clin. Microbiol. 52, 3624–3632. doi: 10.1128/JCM.01370-14
Douiri, N., Hansmann, Y., Lefebvre, N., Riegel, P., Martin, M., Baldeyrou, M., et al. (2016). Staphylococcus lugdunensis?: a virulent pathogen causing bone and joint infections. Clin. Microbiol. Infect. 22, 747–748. doi: 10.1016/j.cmi.2016.05.031
Dufour, P., Jarraud, S., Vandenesch, F., Greenland, T., Novick, R. P., Bes, M., et al. (2002). High Genetic Variability of the agr locus in Staphylococcus Species. J. Bacteriol. 184, 1180–1186. doi: 10.1128/jb.184.4.1180-1186.2002
Ebright, J. R., Penugonda, N., and Brown, W. (2004). Clinical experience with Staphylococcus lugdunensis bacteremia: a retrospective analysis. Diagn. Microbiol. Infect. Dis. 48, 17–21. doi: 10.1016/j.diagmicrobio.2003.08.008
Frank, K. L., del Pozo, J. L., and Patel, R. (2008). From clinical microbiology to infection pathogenesis: how daring to be different works for Staphylococcus lugdunensis. Clin. Microbiol. Rev. 21, 111–133. doi: 10.1128/CMR.00036-07
Gupta, P. K. (2008). Single-molecule DNA sequencing technologies for future genomics research. Trends Biotechnol. 26, 602–611. doi: 10.1016/j.tibtech.2008.07.003
Harris, L. G., Dudley, E., Rohde, H., Frommelt, L., Siemssen, N., Wilkinson, T. S., et al. (2017). Limitations in the use of PSMγ, agr, RNAIII, and biofilm formation as biomarkers to define invasive Staphylococcus epidermidis from chronic biomedical device-associated infections. Int. J. Med. Microbiol. IJMM 307, 382–387. doi: 10.1016/j.ijmm.2017.08.003
Hatoum-Aslan, A., Maniv, I., Samai, P., and Marraffini, L. A. (2014). Genetic characterization of antiplasmid immunity through a type III-A CRISPR-Cas system. J. Bacteriol. 196, 310–317. doi: 10.1128/JB.01130-13
Heilbronner, S., Hanses, F., Monk, I. R., Speziale, P., and Foster, T. J. (2013). Sortase a promotes virulence in experimental staphylococcus lugdunensis endocarditis. Microbiology 159(Pt 10), 2141–2152. doi: 10.1099/mic.0.070292-0
Heilbronner, S., Holden, M. T. G., van Tonder, A., Geoghegan, J. A., Foster, T. J., Parkhill, J., et al. (2011). Genome sequence of Staphylococcus lugdunensis N920143 allows identification of putative colonization and virulence factors. FEMS Microbiol. Lett. 322, 60–67. doi: 10.1111/j.1574-6968.2011.02339.x
Hellmark, B., Söderquist, B., Unemo, M., and Nilsdotter-Augustinsson, Å (2013). Comparison of Staphylococcus epidermidis isolated from prosthetic joint infections and commensal isolates in regard to antibiotic susceptibility, agr type, biofilm production, and epidemiology. Int. J. Med. Microbiol. IJMM 303, 32–39. doi: 10.1016/j.ijmm.2012.11.001
Higaki, S., Kitagawa, T., Morohashi, M., and Yamagishi, T. (1999). Distribution and antimicrobial susceptibility of coagulase-negative staphylococci from skin lesions. J. Int. Med. Res. 27, 191–195. doi: 10.1177/030006059902700406
Holden, M. T. G., Hsu, L.-Y., Kurt, K., Weinert, L. A., Mather, A. E., Harris, S. R., et al. (2013). A genomic portrait of the emergence, evolution, and global spread of a methicillin-resistant Staphylococcus aureus pandemic. Genome Res. 23, 653–664. doi: 10.1101/gr.147710.112
Holtfreter, S., Grumann, D., Schmudde, M., Nguyen, H. T. T., Eichler, P., Strommenger, B., et al. (2007). Clonal distribution of superantigen genes in clinical Staphylococcus aureus isolates. J. Clin. Microbiol. 45, 2669–2680. doi: 10.1128/JCM.00204-07
Jarraud, S., Lyon, G. J., Figueiredo, A. M. S., Rard, L. G., Vandenesch, F., Etienne, J., et al. (2000). Exfoliatin-producing strains de?ne a fourth agr speci?city group in Staphylococcus aureus. J. Bacteriol 182:6.
Jiang, W., Samai, P., and Marraffini, L. A. (2016). Degradation of phage transcripts by CRISPR-associated rnases enables type III CRISPR-Cas immunity. Cell 164, 710–721. doi: 10.1016/j.cell.2015.12.053
Kavanagh, K., and Reeves, E. P. (2004). Exploiting the potential of insects for in vivo pathogenicity testing of microbial pathogens. FEMS Microbiol. Rev. 28, 101–112. doi: 10.1016/j.femsre.2003.09.002
Labrie, S. J., Samson, J. E., and Moineau, S. (2010). Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 8, 317–327. doi: 10.1038/nrmicro2315
Lambe, D. W., Jeffery, C., Ferguson, K. P., and Cooper, M. D. (1994). Examination of the glycocalyx of four species of Staphylococcus by transmission electron microscopy and image analysis. Microbios 78, 133–143.
Lourtet-Hascoët, J., Bicart-See, A., Félicé, M. P., Giordano, G., and Bonnet, E. (2016). Staphylococcus lugdunensis, a serious pathogen in periprosthetic joint infections: comparison to Staphylococcus aureus and Staphylococcus epidermidis. Int. J. Infect. Dis. 51, 56–61. doi: 10.1016/j.ijid.2016.08.007
Malachowa, N., and DeLeo, F. R. (2010). Mobile genetic elements of Staphylococcus aureus. Cell. Mol. Life Sci. 67, 3057–3071. doi: 10.1007/s00018-010-0389-4
Malachowa, N., Whitney, A. R., Kobayashi, S. D., Sturdevant, D. E., Kennedy, A. D., Braughton, K. R., et al. (2011). Global changes in Staphylococcus aureus gene expression in human blood. PLoS One 6:e18617. doi: 10.1371/journal.pone.0018617
Marlinghaus, L., Becker, K., Korte, M., Neumann, S., Gatermann, S. G., and Szabados, F. (2012). Construction and characterization of three knockout mutants of the fbl gene in Staphylococcus lugdunensis: characterization of isogenic mutants of fbl. APMIS 120, 108–116. doi: 10.1111/j.1600-0463.2011.02819.x
McHardy, I. H., Veltman, J., Hindler, J., Bruxvoort, K., Carvalho, M. M., and Humphries, R. M. (2017). Clinical and microbiological aspects of β-lactam resistance in Staphylococcus lugdunensis. J. Clin. Microbiol. 55, 585–595. doi: 10.1128/JCM.02092-16
Michaux, C., Sanguinetti, M., Reffuveille, F., Auffray, Y., Posteraro, B., Gilmore, M. S., et al. (2011). SlyA is a transcriptional regulator involved in the virulence of Enterococcus faecalis. Infect. Immun. 79, 2638–2645. doi: 10.1128/IAI.01132-10
Non, L. R., and Santos, C. A. Q. (2017). The occurrence of infective endocarditis with Staphylococcus lugdunensis bacteremia: a retrospective cohort study and systematic review. J. Infect. 74, 179–186. doi: 10.1016/j.jinf.2016.10.003
Novick, R. P., Christie, G. E., and Penadés, J. R. (2010). The phage-related chromosomal islands of gram-positive bacteria. Nat. Rev. Microbiol. 8, 541–551. doi: 10.1038/nrmicro2393
Novick, R. P., and Ram, G. (2017). Staphylococcal pathogenicity islands — movers and shakers in the genomic firmament. Curr. Opin. Microbiol. 38, 197–204. doi: 10.1016/j.mib.2017.08.001
Novick, R. P., and Subedi, A. (2007). “The SaPIs: mobile pathogenicity islands of Staphylococcus,” in Chemical Immunology and Allergy, ed. G. Marone, (Basel: KARGER), 42–57. doi: 10.1159/000100857
O’Riordan, K., and Lee, J. C. (2004). Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev. 17, 218–234. doi: 10.1128/cmr.17.1.218-234.2004
Poitevin-Later, F., Vandenesch, F., Dyke, K., Fleurette, J., and Etienne, J. (1992). Cadmium-resistance plasmid in Staphylococcus lugdunensis. FEMS Microbiol. Lett. 78, 59–63. doi: 10.1016/0378-1097(92)90288-y
Post, V., Harris, L. G., Morgenstern, M., Mageiros, L., Hitchings, M. D., Méric, G., et al. (2017). Comparative genomics study of Staphylococcus epidermidis isolates from orthopedic-device-related infections correlated with patient outcome. J. Clin. Microbiol. 55, 3089–3103. doi: 10.1128/JCM.00881-17
Poulsen, C., Panjikar, S., Holton, S. J., Wilmanns, M., and Song, Y.-H. (2014). WXG100 protein superfamily consists of three subfamilies and exhibits an a-helical c-terminal conserved residue pattern. PLoS One 9:12. doi: 10.1371/journal.pone.0089313
Ramarao, N., Nielsen-Leroux, C., and Lereclus, D. (2012). The insect Galleria mellonella as a powerful infection model to investigate bacterial pathogenesis. J. Vis. Exp. 70:e4392. doi: 10.3791/4392
Roberts, R. J., Carneiro, M. O., and Schatz, M. C. (2013). The advantages of SMRT sequencing. Genome Biol. 14:405. doi: 10.1186/gb-2013-14-6-405
Roghmann, M., Taylor, K. L., Gupte, A., Zhan, M., Johnson, J. A., Cross, A., et al. (2005). Epidemiology of capsular and surface polysaccharide in Staphylococcus aureus infections complicated by bacteraemia. J. Hosp. Infect. 59, 27–32. doi: 10.1016/j.jhin.2004.07.014
Rossi, C. C., Souza-Silva, T., Araújo-Alves, A. V., and Giambiagi-deMarval, M. (2017). CRISPR-cas systems features and the gene-reservoir role of coagulase-negative staphylococci. Front. Microbiol. 8:1545. doi: 10.3389/fmicb.2017.01545
Sabe, M. A., Shrestha, N. K., Gordon, S., and Menon, V. (2014). Staphylococcus lugdunensis?: a rare but destructive cause of coagulase-negative staphylococcus infective endocarditis. Eur. Heart J. Acute Cardiovasc. Care 3, 275–280. doi: 10.1177/2048872614523350
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinforma. Oxf. Engl. 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Shah, N. B., Osmon, D. R., Fadel, H., Patel, R., Kohner, P. C., Steckelberg, J. M., et al. (2010). Laboratory and clinical characteristics of Staphylococcus lugdunensis prosthetic joint infections. J. Clin. Microbiol. 48, 1600–1603. doi: 10.1128/JCM.01769-09
Singh, V. K., Ring, R. P., Aswani, V., Stemper, M. E., Kislow, J., Ye, Z., et al. (2017). Phylogenetic distribution and expression of a penicillin-binding protein homologue, ear and its significance in virulence of Staphylococcus aureus. J. Med. Microbiol. 66, 1811–1821. doi: 10.1099/jmm.0.000630
Szabados, F., Nowotny, Y., Marlinghaus, L., Korte, M., Neumann, S., Kaase, M., et al. (2011). Occurrence of genes of putative fibrinogen binding proteins and hemolysins, as well as of their phenotypic correlates in isolates of S. lugdunensis of different origins. BMC Res. Notes 4:113. doi: 10.1186/1756-0500-4-113
Tan, L., Li, S. R., Jiang, B., Hu, X. M., and Li, S. (2018). Therapeutic targeting of the Staphylococcus aureus accessory gene regulator (agr) system. Front. Microbiol. 9:55. doi: 10.3389/fmicb.2018.00055
Tseng, S.-P., Lin, Y.-T., Tsai, J.-C., Hung, W.-C., Chen, H.-J., Chen, P.-F., et al. (2015). Genotypes and phenotypes of Staphylococcus lugdunensis isolates recovered from bacteremia. J. Microbiol. Immunol. Infect. 48, 397–405. doi: 10.1016/j.jmii.2013.11.006
Unnikrishnan, M., Constantinidou, C., Palmer, T., and Pallen, M. J. (2017). The enigmatic esx proteins: looking beyond mycobacteria. Trends Microbiol. 25, 192–204. doi: 10.1016/j.tim.2016.11.004
van der Mee-Marquet, N., Achard, A., Mereghetti, L., Danton, A., Minier, M., and Quentin, R. (2003). Staphylococcus lugdunensis infections: high frequency of inguinal area carriage. J. Clin. Microbiol. 41, 1404–1409. doi: 10.1128/JCM.41.4.1404-1409.2003
Wang, Y., Hu, M., Liu, Q., Qin, J., Dai, Y., He, L., et al. (2016). Role of the ESAT-6 secretion system in virulence of the emerging community-associated Staphylococcus aureus lineage ST398. Sci. Rep. 6:25163. doi: 10.1038/srep25163
Warne, B., Harkins, C. P., Harris, S. R., Vatsiou, A., Stanley-Wall, N., Parkhill, J., et al. (2016). The Ess/Type VII secretion system of Staphylococcus aureus shows unexpected genetic diversity. BMC Genomics 17:222. doi: 10.1186/s12864-016-2426-7
Whitney, J. C., Peterson, S. B., Kim, J., Pazos, M., Verster, A. J., Radey, M. C., et al. (2017). A broadly distributed toxin family mediates contact-dependent antagonism between gram-positive bacteria. eLife 6:e26938. doi: 10.7554/eLife.26938
Wilkinson, B. J., and Holmes, K. M. (1979). Staphylococcus aureus cell surface: capsule as a barrier to bacteriophage adsorption. Infect Immun. 23, 549–552.
Yeh, C.-F., Chang, S.-C., Cheng, C.-W., Lin, J.-F., Liu, T.-P., and Lu, J.-J. (2016). Clinical features, outcomes, and molecular characteristics of community- and health care-associated Staphylococcus lugdunensis Infections. J. Clin. Microbiol. 54, 2051–2057. doi: 10.1128/JCM.00847-16
Zankari, E., Hasman, H., Cosentino, S., Vestergaard, M., Rasmussen, S., Lund, O., et al. (2012). Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 67, 2640–2644. doi: 10.1093/jac/dks261
Zhang, D., Iyer, L. M., and Aravind, L. (2011). A novel immunity system for bacterial nucleic acid degrading toxins and its recruitment in various eukaryotic and DNA viral systems. Nucleic Acids Res. 39, 4532–4552. doi: 10.1093/nar/gkr036
Keywords: Staphylococcus lugdunensis, genomic comparative analysis, virulence, ess/type VII locus, clonal complexes
Citation: Lebeurre J, Dahyot S, Diene S, Paulay A, Aubourg M, Argemi X, Giard J-C, Tournier I, François P and Pestel-Caron M (2019) Comparative Genome Analysis of Staphylococcus lugdunensis Shows Clonal Complex-Dependent Diversity of the Putative Virulence Factor, ess/Type VII Locus. Front. Microbiol. 10:2479. doi: 10.3389/fmicb.2019.02479
Received: 22 July 2019; Accepted: 15 October 2019;
Published: 31 October 2019.
Edited by:
Frank T. Robb, University of Maryland, Baltimore, United StatesReviewed by:
Timothy J. Foster, Trinity College Dublin, IrelandBen Pascoe, University of Bath, United Kingdom
Simon Heilbronner, University of Tübingen, Germany
Copyright © 2019 Lebeurre, Dahyot, Diene, Paulay, Aubourg, Argemi, Giard, Tournier, François and Pestel-Caron. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jérémie Lebeurre, amVyZW1pZS5sZWJldXJyZUBob3RtYWlsLmZy