 Maximiliano Venegas1
Maximiliano Venegas1 Salvador Barahona2
Salvador Barahona2 Ana María González1Dionisia Sepúlveda2
Ana María González1Dionisia Sepúlveda2 Gustavo E. Zúñiga3Marcelo Baeza1,2Víctor Cifuentes1,2
Gustavo E. Zúñiga3Marcelo Baeza1,2Víctor Cifuentes1,2 Jennifer Alcaíno1,2*
Jennifer Alcaíno1,2*- 1Departamento de Ciencias Ecológicas, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 2Centro de Biotecnología, Facultad de Ciencias, Universidad de Chile, Santiago, Chile
- 3Departamento de Biología, Facultad de Química y Biología, CEDENNA, Universidad de Santiago de Chile, Santiago, Chile
Xanthophyllomyces dendrorhous synthesizes astaxanthin, a carotenoid used in aquaculture. Astaxanthin is synthesized from metabolites of the mevalonate pathway, which are also precursors for sterols biosynthesis. The interruption of the CYP61 gene, which is involved in the synthesis of ergosterol (mutant CBS.cyp61–), resulted in a phenotype that overproduces carotenoids due to the activation of the SREBP pathway. In this work, we constructed other mutants of ergosterol biosynthesis in this yeast to evaluate whether they have the same phenotype as mutant CBS.cyp61–. By bioinformatic analysis, the ERG3 and ERG4 genes of X. dendrorhous were identified, and each gene was deleted in the wild-type strain. Mutants CBS.Δerg3 and CBS.Δerg4 did not produce ergosterol; CBS.Δerg3 primarily accumulated episterol, and CBS.Δerg4 primarily accumulated ergosta-5,7,22,24(28)-tetraenol. The transcription levels of the HMGS gene of the mevalonate pathway were evaluated by RT-qPCR, which showed a slight increase in CBS.Δerg4, but the transcription levels were still 10-fold lower than in strain CBS.cyp61–. Both CBS.Δerg3 and CBS.Δerg4 did not overproduce carotenoids, even though they do not produce ergosterol. Thus, the results of this study indicate that the absence of ergosterol does not activate the SREBP pathway in X. dendrorhous, but rather it depends on other alterations in sterol composition.
Introduction
Xanthophyllomyces dendrorhous is a basidiomycete yeast that produces carotenoids, which are natural liposoluble pigments of 40 carbon atoms that exhibit shades ranging from yellow to red (Golubev, 1995) and have antioxidant properties attributed to their structure (Guerin et al., 2003). Among carotenoids, astaxanthin is of biotechnological interest due to its antioxidant capacity, which is the reason why it is currently used in the nutraceutical and cosmetic industries (Johnson, 2003; Higuera-Ciapara et al., 2006). Astaxanthin also has an important economic interest, since it is used in aquaculture as a food supplement.
In X. dendrorhous, the biosynthesis of astaxanthin begins with the condensation of dimethylallyl-pyrophosphate (DMAPP, C5) and isopentenyl-pyrophosphate (IPP, C5) (Figure 1), which are synthesized through the mevalonate (MVA) pathway, producing geranyl-pyrophosphate (GPP, C10). Then, a second IPP molecule is added, giving rise to farnesyl-pyrophosphate (FPP, C15), which is a substrate for both sterol and carotenoid synthesis. In sterols synthesis, two FPP molecules are condensed, giving squalene, and in the synthesis of carotenoids, FPP is joined to a third molecule of IPP, giving geranylgeranyl-pyrophosphate (GGPP, C20). The condensation of two molecules of GGPP gives phytoene (the first carotenoid of the synthesis pathway), which is transformed into lycopene through four desaturation steps. The cyclization of both ends of lycopene produces β-carotene, and finally, the addition of a keto and a hydroxyl to both terminal rings of β-carotene gives astaxanthin (Loto et al., 2012).
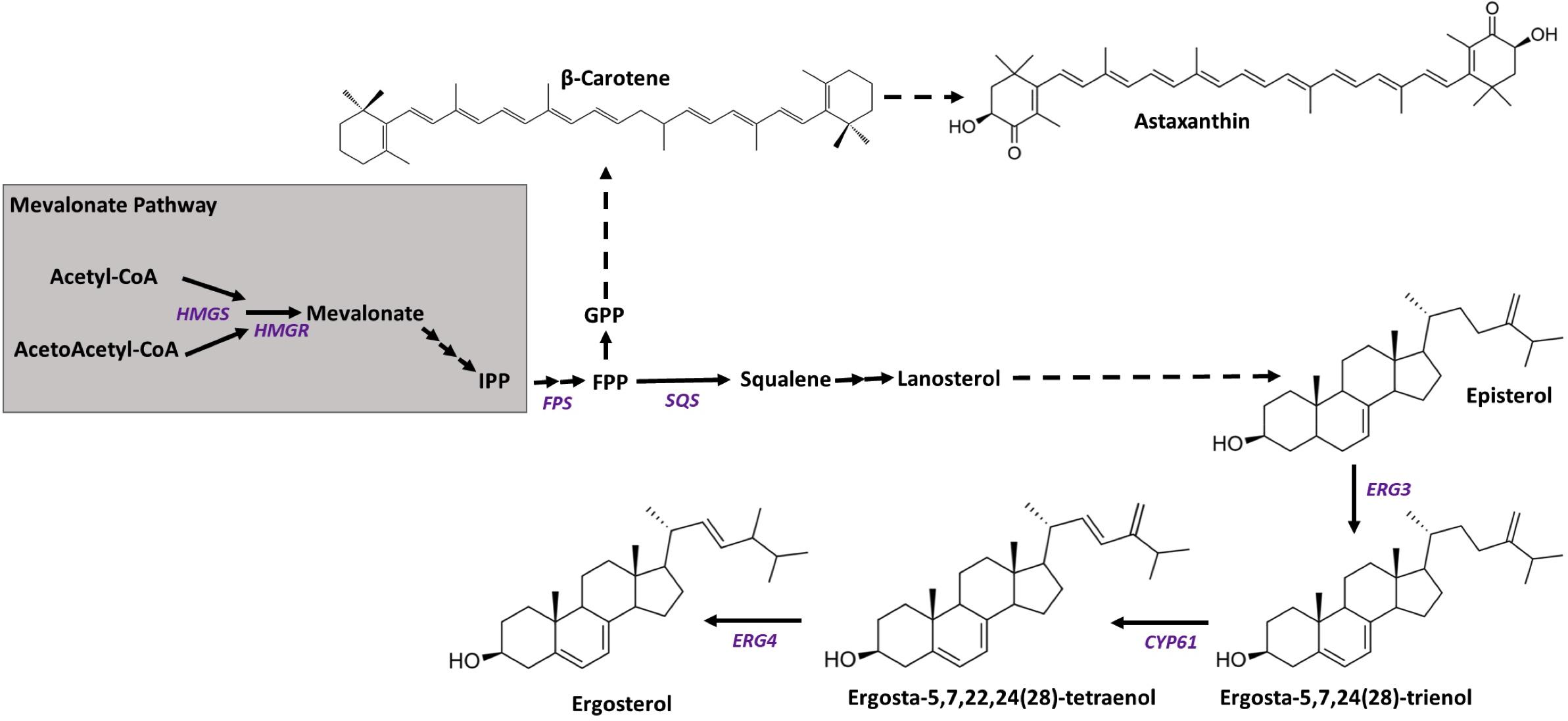
Figure 1. Scheme of the mevalonate pathway and the synthesis of carotenoids and sterols. Arrows represent catalytic steps with the corresponding enzyme-encoding gene studied in this work. Gene names are written in purple, and their GenBank accession numbers are HMGS (XDEN_03265), HMGR (GenBank: AJ884949), ERG3 (GenBank: MN930922), ERG5/CYP61 (GenBank: JX183236), and ERG4 (GenBank: MN930923). The final steps of ergosterol biosynthesis correspond to steps described in S. cerevisiae steroid biosynthesis at the KEGG database (sce00100).
Sterols regulate biological processes and contribute to the cell membrane structure, where they are considered membrane stabilizers (Zhang and Rao, 2010). While cholesterol is the main sterol in mammals, ergosterol plays a key role in fungi. Plants generally have more complex sterol compositions (Schaller, 2003). In the yeast Saccharomyces cerevisiae, the biosynthesis of ergosterol has been extensively studied, and the last three steps are catalyzed by enzymes encoded by the ERG3, ERG5, and ERG4 genes. The ERG5 gene is also referred to as CYP61, since the gene product belongs to the superfamily of hemoproteins known as cytochrome P450s (P450s). P450s are monooxygenases that are present in all domains of life, and their function is related to oxidative metabolism (Degtyarenko and Archakov, 1993). Two P450s, Cyp51 and Cyp61, are crucial in the synthesis of ergosterol; for this reason, clinical studies against fungal infections have attempted to develop compounds that target P450 enzymes (Parker et al., 2014).
In S. cerevisiae, the ERG3 gene encodes the C-5 sterol desaturase [EC:1.14.19.20], which catalyzes the conversion of episterol to ergosta-5,7,24(28)-trienol using NADPH as a cofactor, a reaction that involves the cis removal of hydrogens located in the α position of carbons 5 and 6 of episterol coupled to the reduction of an oxygen molecule. It has been described that the erg3– mutation in Candida glabrata affects yeast growth, resulting in a phenotype that is not capable of producing ergosterol, thereby accumulating other intermediary sterols (Geber et al., 1995). It has also been described that the ERG3 gene is essential for growth in S. cerevisiae when cultured with fermentable carbon sources, such as glycerol and ethanol (Smith and Parks, 1993). On the other hand, the ERG4 gene encodes a C-24 sterol reductase [EC:1.3.1.71] that participates in the last step of ergosterol biosynthesis and converts ergosta-5,7,22,24(28)-tetraenol into ergosterol using NADPH as a cofactor. The S. cerevisiae erg4– mutant does not produce ergosterol and accumulates the precursor ergosta-5,7,22,24(28)-tetraen-3β-ol. Under the tested conditions, the erg4– mutation did not affect cell viability, although it reduced the yeast resistance to different stress conditions and increased its sensitivity to multiple chemicals, thereby reducing its competitive fitness (Zweytick et al., 2000).
In X. dendrorhous, two genes controlling the synthesis of ergosterol have been functionally described: CYP51/ERG11 (Leiva et al., 2015) and CYP61/ERG5 (Loto et al., 2012), both of which encode P450 enzymes (Córdova et al., 2017). The CYP61 gene has been characterized in S. cerevisiae as a non-essential gene (Skaggs et al., 1996). Interestingly, the cyp61– mutation, in addition to blocking ergosterol biosynthesis, enhanced the production of carotenoids and sterols in different X. dendrorhous strains (Loto et al., 2012). Similarly, astaxanthin-overproducing Phaffia rhodozyma [the anamorphic state of X. dendrorhous (Golubev, 1995)] mutants obtained by random mutagenesis showed a lower ergosterol content compared to the original strains (Miao et al., 2010, 2011).
Recent works revealed that the carotenoid and sterol overproducer phenotype in X. dendrorhous cyp61– mutants depends on the sterol regulatory element-binding protein (SREBP) pathway (Gutiérrez et al., 2019; Gómez et al., 2020). SREBPs are a family of transcription factors that regulate the expression of genes controlling the synthesis of sterols and fatty acids. These factors are synthesized as inactive precursor proteins anchored in the endoplasmic reticulum membrane that, depending on the cellular lipid demand, are activated through proteolytic processing (Bien and Espenshade, 2010). An X. dendrorhous SREBP homolog encoded by the SRE1 gene was recently described and shown to be involved in the regulation of carotenoid and sterol synthesis in this yeast (Gutiérrez et al., 2019). Unlike in the wild-type strain, Sre1 is in its active state in the cyp61– mutant when cultured under standard laboratory conditions (Gómez et al., 2020). The above results demonstrate that the cyp61– mutant provides the conditions that meet the activation of Sre1, which is responsible for the overproduction of carotenoids and sterols in this strain. In support of this statement, the SRE1 gene deletion in the cyp61– mutant reverted the carotenoid- and sterol-overproducing phenotype, reaching levels equivalent to those produced by the wild-type strain. Conversely, the replacement of the SRE1 gene in the wild-type strain by the gene version that only encodes the active form of Sre1 favors the production of carotenoids and sterols at levels similar to those produced by the cyp61– mutant (Gutiérrez et al., 2019). This background suggests that the absence of ergosterol in the cyp61– mutants triggers Sre1 activation in X. dendrorhous. Then, to move forward to understand what triggers Sre1 activation in X. dendrorhous, ergosterol mutants of genes other than CYP61 were constructed and analyzed in this work. The genes ERG3 and ERG4 were selected, as these genes encode enzymes catalyzing the previous and the following step of the one that is catalyzed by the CYP61 gene product, and because the mutation of these genes in other yeasts were viable and blocked the synthesis of ergosterol.
Materials and Methods
Microorganisms, Plasmids and Culture Conditions
The microorganisms and plasmids used or created in this work are listed in Table 1. The CBS 6938 strain of X. dendrorhous was used as the wild-type strain, and E. coli DH5α was used as a host for plasmid propagation. X. dendrorhous strains were cultured at 22°C with constant agitation at 170 rpm in YM medium (0.3% yeast extract, 0.3% malt extract and 0.5% peptone) supplemented with 1% glucose. Yeast transformant selection was performed on YM-agar 1.5% plates supplemented with 20 μg/ml hygromycin B and/or 35 μg/ml zeocin (Sigma, Saint Louis, MI, United States). E. coli strains were cultured with constant agitation at 37°C in LB medium (1% tryptone, 0.5% yeast extract and 0.5% NaCl) supplemented with 100 μg/ml ampicillin to select strains carrying plasmids and 40 μl of a 2% solution of X-gal (5-bromo-4-chloro-3-indolyl-β-D-galactopyranoside) for recombinant clone selection (Sambrook and Russell, 2001, #31893). The recombinant clones carrying plasmids with the hygromycin B or zeocin resistance cassette were identified by direct colony PCR with specific primers for each cassette.
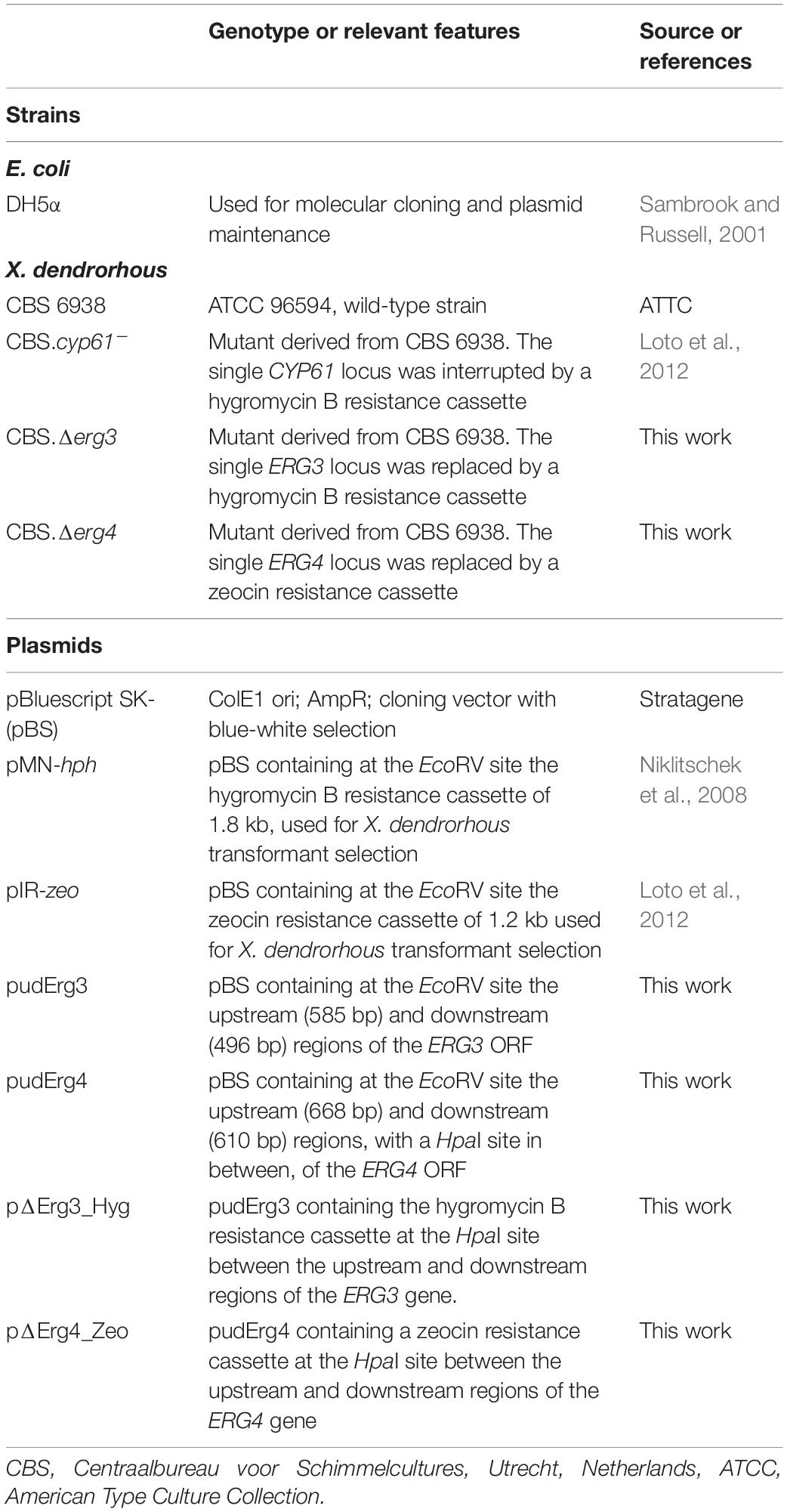
Table 1. Strains and plasmids used and built in this work.
Bioinformatic Analyses and Nucleic Acid Extraction
The ERG3 and ERG4 genes of X. dendrorhous were identified by local tBLASTn search over the genomic and transcriptomic databases from the X. dendrorhous strain UCD 67-385 (Baeza et al., 2015) with Geneious R11 using related sequences obtained from the GenBank database as queries. The exon-intron structure of each gene was manually cured by comparing genomic and transcriptomic data. Both gene sequences were uploaded to the GenBank database under the following accession numbers MN930922 and MN930923 for ERG3 and ERG4, respectively. The analysis of the promoter regions of the HMGS, ERG3 and ERG4 genes was carried out with the JASPAR1 platform using the TRANSFAC database (Mathelier et al., 2016). The transcriptional factor SREBF2 was used as a consensus binding site. Protein sequence analyses were performed with programs available at http://www.ebi.ac.uk/interpro (Jones et al., 2014), https://embnet.vital-it.ch/software/TMPRED_form.html (Hofmann and Stoffel, 1993), and https://psort.hgc.jp/form2.html (Nakai and Horton, 1999).
Plasmid DNA was extracted from E. coli using the GeneJET Plasmid Miniprep Kit (Thermo Fisher, Waltham, MA, United States), and DNA sequencing was performed at Macrogen, Inc (Seoul, South Korea). The extraction of genomic DNA and total RNA from yeast X. dendrorhous was performed by mechanical rupture of a cell pellet obtained from a culture in liquid medium as described previously (Gómez et al., 2020, #70812). The obtained genomic DNA and RNA were stored at −20 and −80°C, respectively, until use.
Polymerase Chain Reaction (PCR), Reverse Transcription (RT) and Quantitative PCR (qPCR)
All oligonucleotides designed and used in this work were synthesized by Integrated DNA Technologies (Coralville, IA, United States) and are listed in Supplementary Table S1.
To amplify the upstream and downstream fragments of the ERG3 and ERG4 genes, PCR analyses were performed using Pfu DNA polymerase (Agilent Technologies, Santa Clara, CA, United States). For E. coli recombinant clone selection and X. dendrorhous transformant confirmation, Taq DNA polymerase (Centro de Biotecnología, Facultad de Ciencias, Universidad de Chile, Chile) was used. PCR reactions were performed in a final volume of 25 μl using an Applied Biosystem 2720 thermocycler with the following program for standard PCR: initial denaturation at 95°C for 3 min (or 5 min in case of colony PCR or amplification with Pfu DNA polymerase), 35 cycles of denaturation at 94°C for 30 s, alignment of primers at 55°C for 30 s, and elongation at 72°C for 3 to 4 min followed by a final elongation step at 72°C for 10 min, and finally, the reaction was maintained at 4°C until analysis was performed.
Overlap extension PCR (OE-PCR) (Urban et al., 1997) was used to join the upstream and downstream DNA fragments of both studied genes, and the reaction was performed in a final volume of 25 μl with 1 U of Pfu DNA polymerase, PCR buffer 1X, each dNTP at 200 μM and 100 ng of each DNA fragment. The following program was used: initial denaturation at 94°C for 1 min, then 10 cycles of denaturation at 94°C for 30 s, DNA alignment at 55°C for 45 s, elongation at 72°C for 90 s, then a final elongation step at 72°C for 10 min, and finally the reaction was kept at 4°C until the next step. Then, the obtained reaction mixture was amplified by a standard PCR analysis with Pfu DNA polymerase and the same forward and reverse primers used to amplify the upstream and downstream DNA fragments, respectively.
RT reactions were performed in a final volume of 20 μl. First, 5 μg of total RNA in 11 μl of sterile water was mixed with 1 μl of oligo-dT (25 μM) and 1 μl of dNTPs (at 10 mM each) and incubated at 65°C for 5 min. Then, 1 μl of the enzyme Maxima Reverse Transcriptase (Thermo Fischer, Waltham, MA, United States), 4 μl of RT buffer 5X and 2 μl of DTT (0.1 M) were added. The mixture was incubated at 37°C for 52 min and then left at 70°C for 15 min, enabling it to cool at 4°C.
Real-time qPCR reactions were performed in a Mx3000P real-time PCR system (Agilent, Santa Clara, CA, United States) using Fast EvaGreen qPCR Master Mix (Bio-Rad Laboratories, Hercules, CA, United States). Samples were prepared in a final volume of 20 μl containing 1 μl of cDNA, 0.25 μM of each primer and 10 μl of kit reaction mixture. The efficiency of the primers used was greater than 95%, as determined by standard curves with a correlation coefficient R2 ≥ 0.99. The obtained Ct (threshold cycle) values were normalized with respect to the corresponding value of the X. dendrorhous β-actin gene [GenBank: X89898.1] and expressed as fold change with respect to the control (Livak and Schmittgen, 2001).
Plasmid Construction and X. dendrorhous Transformation
The ERG3 and ERG4 genes were replaced by an antibiotic resistance cassette in the X. dendrorhous genome by homologous recombination using plasmids pΔErg3_Hyg and pΔErg4_Zeo (Table 1). Plasmids were constructed by joining the upstream (“UP”) and downstream (“DOWN”) regions of each gene of interest by OE-PCR, introducing an HpaI recognition site between them, according to the primer design (primers 5, 6, 15, and 16, Supplementary Table S1). Both regions were PCR-amplified from genomic DNA from X. dendrorhous strain CBS 6938, and the OE-PCR product was inserted at the EcoRV site of plasmid pBluescript SK to obtain plasmids pudErg3 and pudErg4. Then, each plasmid was digested with HpaI to insert either a hygromycin B or zeocin resistance cassette, which were obtained by EcoRV digestion of plasmids pMN-hph or pIR-zeo (Table 1), respectively.
Xanthophyllomyces dendrorhous was transformed by electroporation using a GenePulser XcellTM (BioRad, Hercules, CA, United States). Electrocompetent cells were prepared from cultures in YM medium at the exponential phase of growth at an OD600 nm of 4–5 (Adrio and Veiga, 1995). Electroporation was performed using 6 μl of transforming DNA (10 μg) employing a 2 mm cuvette and the following conditions: 450 V, 125 μF and 600 Ω. After the pulse, 1 ml of YM was added to the cells, which were incubated for 4 h at 22°C before seeding in YM-agar 1.5% plates with the respective antibiotics to select the transformants.
Phenotypic Analyses of the Mutant Strains
Phenotypic analysis was performed using at least three biological replicas of each strain, including the mutant strains CBS.Δerg3 and CBS.Δerg4 and strains CBS.cyp61- and CBS 6938 as controls. Strains were cultivated in 100 ml of YM in 250 ml Erlenmeyer flasks at 22°C with constant agitation. Yeast growth was evaluated by the OD600 nm of the culture, which was measured in a UV-vis V-630 spectrophotometer from JASCO. After 120 h of culture (stationary phase of growth), samples were taken to extract carotenoids, ergosterol and total RNA. This time point was selected as major differences in the transcript levels of gene HMGR of the MVA pathway, which is a known Sre1 gene target in X. dendrorhous (Gutiérrez et al., 2019), were observed when strain CBS.cyp61– was compared to the wild-type (Loto et al., 2012).
Extracted carotenoids and sterols were quantified spectrophotometrically and normalized to the dry weight of the yeast. Carotenoids were extracted from cell pellets from 20 ml culture samples disrupted using glass beads and acetone (An et al., 1989) and quantified at 465 nm using an absorption coefficient of A1% = 2,100. Sterols were extracted according to Shang et al. (2006). In brief, cell pellets from 5 ml culture samples were saponified in 16 ml of 60% ethanol and 4 g of KOH at 80°C for 2 h. Then, after the mixture was cooled, 5 ml of petroleum ether was added, vortexed for 10 s and centrifuged for 5 min. The supernatant was recovered, and this last step was repeated. Total sterols were quantified at 280 nm using an absorption coefficient of A1% = 11,500.
Carotenoid and sterol compositions were evaluated by reverse-phase liquid chromatography (RP-HPLC) using a LiChrospher RP18 125-4 C-18 column (Merck Millipore, Billerica, MA United States). Carotenoid and sterol samples were run using acetonitrile: methanol: isopropanol (85: 10: 5, v/v/v) and methanol: water (97: 3, v/v), respectively, as the mobile phase with a 1 ml/min flux under isocratic conditions, and the elution spectra were recovered using a diode array detector. Sterols and carotenoids were visualized at the 280 and 474 nm channels, respectively, and compared to standards according to retention time and absorption spectra. The ergosterol standard was acquired from Sigma (Saint Louis, MI, United States).
Sterol identification was performed by a 1100 HPLC Series system equipped with a G1322A degasser and G1311A binary pump. For separation, a reversed-phased Zorbax SB-C18 analytical column (100 mm × 3.0 mm i.d., 5 μm particles) fitted with precolumn Zorbax SB-C18 was used. The mobile phase was prepared from methanol and acetonitrile 30: 70 (v/v). The flow rate was 0.6 ml/min, and 20 μl was injected. All solvents used were filtered through 0.5 ml Sartorius filters and degassed with ultrasounds. The flow rate was set to 0.5 ml/min, and the column temperature was set at 40°C. The mass spectra were acquired in positive mode using electrospray ionization, and analysis of all analytes was carried out in MRM mode. The other operating parameters were as follows: nebulizer gas flow, 3 L/min; drying gas flow, 15 L/min; desolvation line (DL) temperature, 250°C; and heat block temperature, 400°C. All chromatographic data were processed using ChemStation (v A09.03) software and Data Analysis (v 5.3).
Results
Isolation and Sequence Analysis of the ERG3 and ERG4 Genes From X. dendrorhous
The ERG3 and ERG4 genes from X. dendrorhous were identified by BLAST search over a genomic and transcriptomic databases from the yeast using homologous sequences from other organisms as queries. Once genes were identified, the exon-intron structure of each gene was manually cured by comparing the genomic and transcriptomic data. The ERG3 gene contains 6 exons of 554, 68, 102, 44, 105, and 114 bp, and the ERG4 gene has 15 exons of 202, 107, 97, 276, 134, 133, 4, 93, 35, 151, 16, 17, 55, 103, and 35 bp. From the coding sequence, it was deduced that the ERG3 gene encodes a polypeptide of 328 amino acids with an estimated molecular weight of 38.1 kDa and an isoelectric point (PI) of 7.88. In addition, the ERG3 polypeptide has 26 positively charged and 25 negatively charged residues. The ERG4 gene encodes a polypeptide of 485 amino acids. This polypeptide is different from the one deposited in the NCBI database, given that our CDS prediction contemplates the CDS of sterol reductase/beta-sheet receptor (CED83197.1) and C-24 sterol reductase (CED83198.1), with an estimated molecular weight of 55.1 kDa and a PI of 9.14. It has 31 negatively charged residues and 43 positively charged residues.
The subcellular localization of each protein was predicted using the PSORT II prediction tool. Both proteins are predicted to associate with the endoplasmic reticulum with 34.8% for Erg3p in the K-nn test and 55.6% for Erg4p, which is consistent with their potential function in the biosynthesis of ergosterol at the endoplasmic reticulum (Nishino, 1981). The number of possible transmembrane segments in each of them was also predicted by using the TMHMM Server v. 2.0, where it was observed that Erg3p has 3 transmembrane segments (Figure 2A), while Erg4p has 9 (Figure 2B). Additionally, through the InterPro platform, the presence of conserved domains for both proteins were predicted in silico. Erg3p contains 1 domain of the fatty acid hydroxylase superfamily (IPR006694), while for Erg4p, two conserved domains belonging to the family of sterol reductase proteins were observed (IPR018083).

Figure 2. Sequence analysis of the X. dendrorhous Erg3 and Erg4 predicted proteins. (A) Representation of the deduced Erg3 protein and sequence alignment of the sterol desaturase/sphingolipid hydroxylase, fatty acid hydroxylase superfamily (FA_hydroxylase; COG3000) domain (residues 174 to 299 of the X. dendrorhous Erg3 protein). (B) Representation of the deduced Erg4 protein and sequence alignment of the two identified sterol reductase (Sterol_Reduct_1 and 2; PS01017 and PS01018) domains (residues 218 to 233 and 449 to 472 of the X. dendrorhous Erg4 protein). Predicted transmembrane segments are represented in blue boxes. The UniProt accession numbers (Erg3; Erg4) of sequences used in the alignments are Aspergillus niger (A0A100II78_ASPNG; A0A254U2Z2_ASPNG), Aspergillus fumigatus (Q2VWQ7_ASPFM; Q4WKA5_ASPFU), Cryptococcus neoformans (Q5KNX1_CRYNJ; Q6U7Q6_CRYNH), Schizosaccharomyces pombe (ERG31_SCHPO; STS1_SCHPO), Laccaria bicolor (B0DA59_LACBS; B0CTW1_LACBS) and Saccharomyces cerevisiae (P32353; P25340). Alignments are colored by percentage identity as follows: black 100% identity, gray>80% identity, light gray>60% identity and white<60% identity.
To evaluate the functionality of both identified genes, they were independently replaced by an antibiotic resistance module in the X. dendrorhous genome. To this end, recombination modules were constructed and inserted at the EcoRV site of plasmid pBluescript II SK- to obtain plasmids pCBS-Δerg3 and pCBS-Δerg4, respectively (Table 1). The ERG3 recombination module contained 585 bp upstream and 538 bp downstream of the gene flanking a hygromycin B resistance cassette, while the ERG4 recombination module contained 610 bp upstream and 579 bp downstream of the gene and a zeocin resistance cassette between them.
Once plasmids pCBS-Δerg3 and pCBS-Δerg4 were obtained, the wild-type strain CBS 6938 was transformed to replace the respective gene by the antibiotic resistance module through homologous recombination. In this way, three transformants resistant to hygromycin B and four transformants resistant to zeocin were obtained for the ERG3 and ERG4 gene mutations, respectively. To confirm the gene replacement events on the obtained transformants, their genomic DNA was extracted, and PCR analyses were performed using an informative set of primers (Figure 3). For the following analysis, a mutant of each gene was randomly selected.
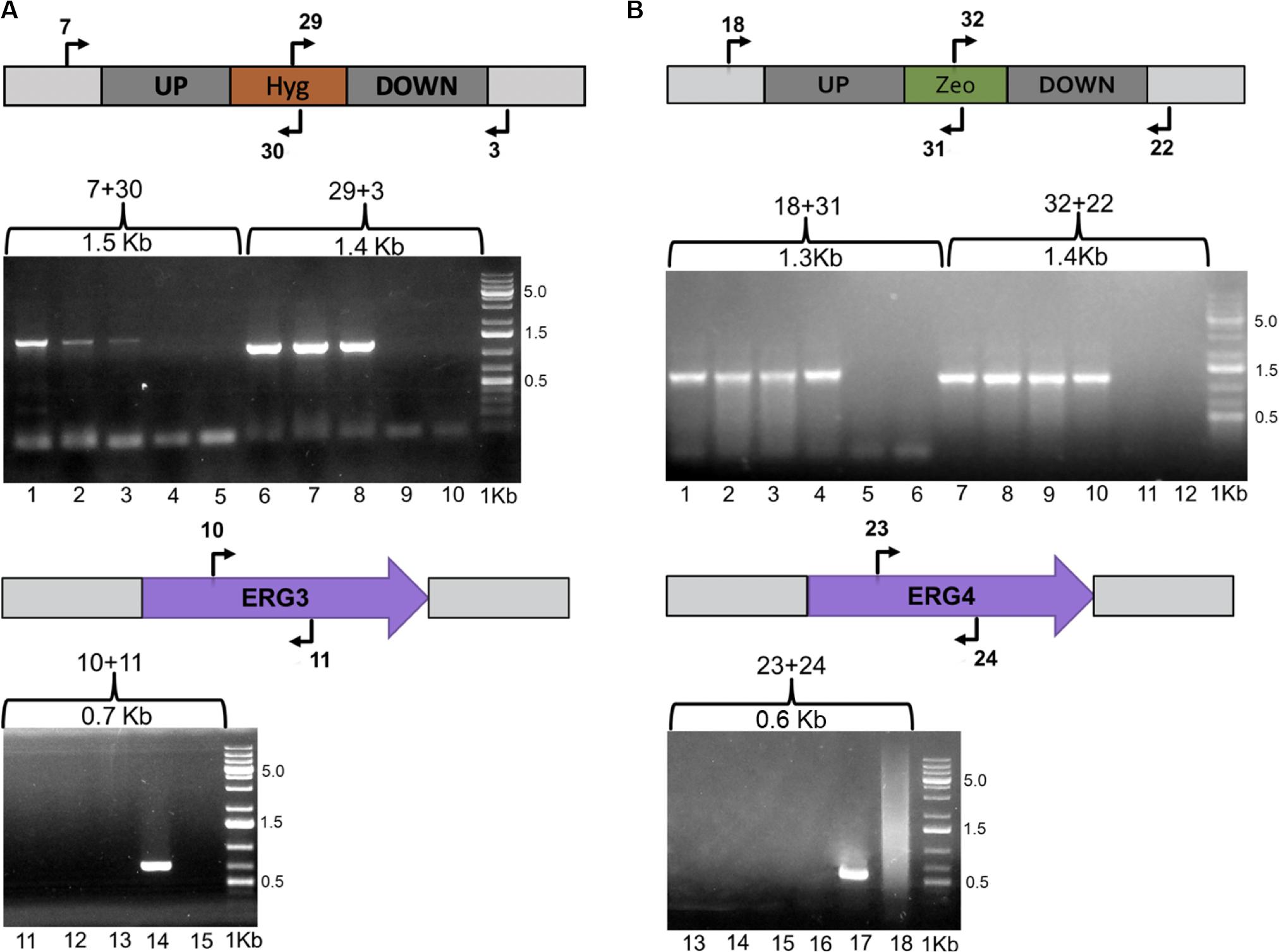
Figure 3. Evaluation of the Δerg3 and Δerg4 gene mutations. The X. dendrorhous ERG3 and ERG4 genes were replaced by hygromycin B and zeocin resistance modules through homologous recombination, respectively, which was confirmed by PCR analysis. Schemes including the primers used in the analysis (Supplementary Table S1) and the expected PCR-product size are indicated in the figure. (A) The replacement of the ERG3 locus by the hygromycin B resistance module was confirmed with primer pairs 7 + 30 and 29 + 3, and the presence of the ERG3 gene was confirmed with primer pair 10 + 11. Lanes 1, 2, 3, 6, 7, 8, 11, 12, and 13 correspond to the analysis of the three transformants obtained; lanes 4, 9 and 14 to the parental strain CBS 6938, and lanes 5, 10, and 15 to PCR negative controls. (B) The replacement of the ERG4 locus by the zeocin resistance module was confirmed with primer pairs 18 + 31 and 32 + 22, and the presence of the ERG4 gene was confirmed with the primer pair 23 + 24. Lanes 1, 2, 3, 4, 7, 8, 9, 10, 13, 14, 15, and 16 correspond to the analysis of three transformants; lanes 5, 11 and 17 correspond to the parental strain CBS 6938, and lanes 6, 12 and 18 correspond to PCR negative controls. 1 Kb Plus was used as molecular weight marker 0.5, 1.5, and 5.0 corresponding to the molecular weight sizes in Kb.
Evaluation of the Phenotype of the Mutant Strains: Production of Ergosterol and Carotenoids
Four strains were analyzed and compared: the wild-type strain CBS 6938 and the mutant strains CBS.Δerg3, CBS.Δerg4 and CBS.cyp61– (Figure 4A). Only strain CBS.Δerg4 showed a different growth curve, reaching a lower OD at the stationary phase of growth (Supplementary Figure S1).
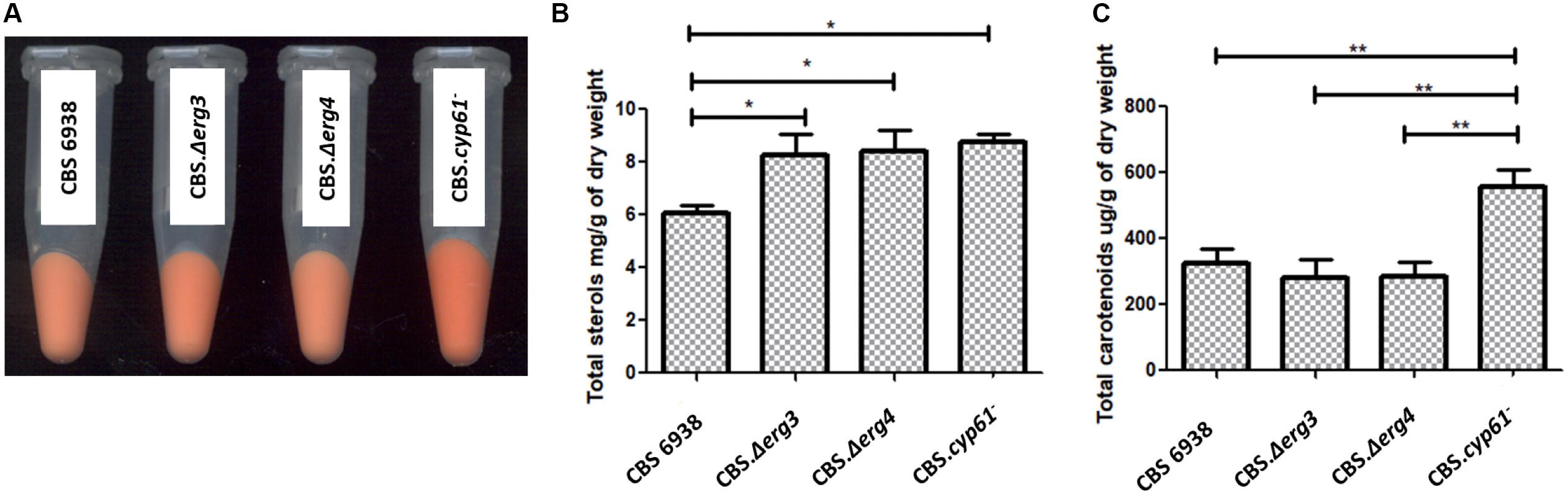
Figure 4. Phenotypic analysis, total sterols and carotenoids between strains CBS 6938, CBS.Δerg3, CBS.Δerg4 and CBS.cyp61–. (A) Cell pellets of strains CBS 6938 (parental, wild-type strain), CBS.Δerg3, CBS.Δerg4, and CBS.cyp61– obtained from cultures grown on semisolid YM medium incubated at 22°C for 4 days. (B) Sterols and (C) carotenoids were extracted at the stationary phase of growth cultures (120 h of culture) and quantified at 280 nm and 474 nm, respectively. Values correspond to the average of 3 biological replicas, and the error bars correspond to the standard deviation. Data were normalized with respect to the dry weight of the yeast. *p < 0.05 and **p < 0.01, Student’s t-test.
Considering that the genes ERG3 and ERG4 are involved in the biosynthesis of ergosterol, total sterols were extracted and quantified (Figure 4B). A significant difference was observed in the total amount of sterols produced by the mutant strains CBS.Δerg3 and CBS.Δerg4 with respect to the parental strain CBS 6938; both mutants have a higher sterol content than the wild-type strain. Neither strain CBS.Δerg3 nor CBS.Δerg4 produced ergosterol but rather accumulated other sterols with different retention times (Supplementary Table S2). In addition, to determine the complete profile of sterols in each strain, an LC-MS/MS analysis was performed (Table 2). It was observed that the major compound in strain CBS 6938 corresponds to ergosterol, and it also accumulates other sterols from different levels of the ergosterol synthesis pathway. None of the mutant strains analyzed in this work accumulated ergosterol, and they had a different sterol profile according to each mutation (the affected enzyme). Thus, the main intermediary accumulated in strain CBS.Δerg3 corresponded to episterol. In this case, the affected enzyme converts episterol into ergosta-5,7,24(28)-trienol. The strain CBS.Δerg4 accumulated ergosta-5,7,22,24(28)-tetraenol since the affected enzyme catalyzes the conversion of this molecule to ergosterol. Finally, the CBS.cyp61– strain accumulated ergosta-5,7,24(28)-trienol and episterol as major compounds because the affected enzyme catalyzes the conversion of ergosta-5,7,24(28)-trienol to ergosta-5,7,22,24(28)-tetraenol. In addition, all mutants accumulated upstream sterols, such as lanosterol, zymosterol and fecosterol. In summary, this analysis confirmed that none of the analyzed mutants accumulated ergosterol.
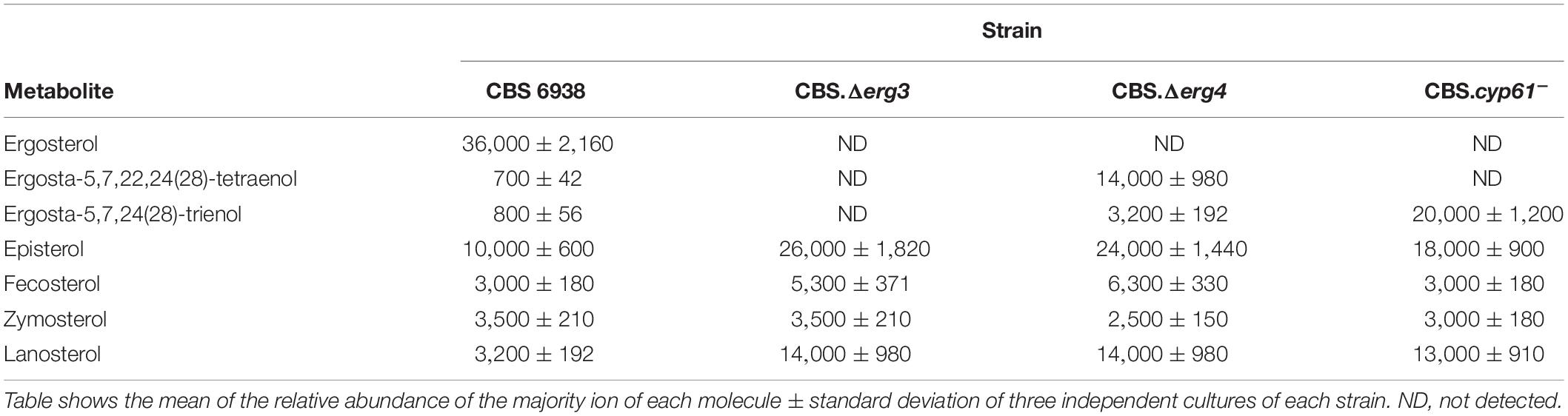
Table 2. Composition (in relative abundance) of sterols of the mutant and parental strains of X. dendrorhous analyzed by HPLC/MS.
Previously, a different color phenotype was observed in cyp61– mutants with respect to their parental wild-type strains (Loto et al., 2012); however, this difference was not observed in the CBS.Δerg3 or the CBS.Δerg4 mutants. To confirm these observations, carotenoids from the analyzed strains were extracted and quantified: only the CBS.cyp61– mutant had a significantly higher carotenoid content than the other strains (Figure 4C). Carotenoid composition was also analyzed, and minimal differences were observed among strains (Table 3). These results indicate that even though the CBS.Δerg3 and CBS.Δerg4 mutants do not produce ergosterol, carotenoid production is not enhanced, as was observed in the X. dendrorhous cyp61– mutants, suggesting that is not the absence of ergosterol what enhances carotenoid production but probably the different sterol composition in this mutant would favor carotenoid production.
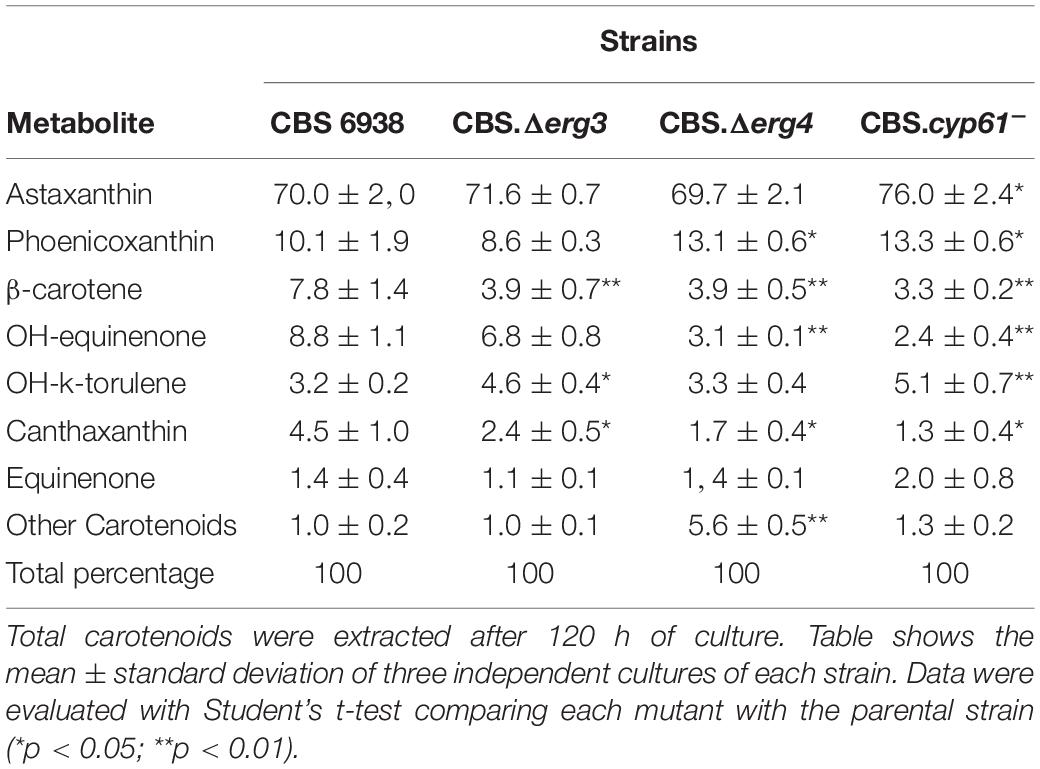
Table 3. Composition (in %) of carotenoids of the mutant and parental strains of X. dendrorhous analyzed by RP-HPLC.
Expression Level of the ERG3, ERG4, and HMGS Genes
In a previous work, we observed that cyp61– mutants derived from different X. dendrorhous wild-type strains had higher transcript levels of the HMGR gene, which is involved in the MVA pathway, in relation to the corresponding parental strains (Loto et al., 2012). Transcriptional activation of several genes involved in the MVA pathway and in the synthesis of ergosterol in different organisms is regulated by sterol levels through proteins SREBPs (named Sre1 in several fungi, including X. dendrorhous). In general, at low sterol levels, SREBP/Sre1 is activated by proteolytic cleavage, allowing the transcription factor domain to travel to the nucleus and bind to specific DNA sequences, known as sterol regulatory elements (SREs), in the promoter region of target genes to regulate their expression. Conversely, when sterol levels increase, SREBP/Sre1 cleavage is inhibited; therefore, the synthesis of sterols is reduced through a negative feedback mechanism (Hughes et al., 2005). Considering that the ERG3 and ERG4 genes encode enzymes involved in ergosterol biosynthesis such that they could be regulated by Sre1, the promoter region of these genes was analyzed to search for potential SREBP/Sre1 binding sites. For comparative purposes, the promoter region of the HMGS gene was also analyzed, as it was demonstrated that Sre1 in X. dendrorhous binds to this DNA (Gutiérrez et al., 2019). The consensus SRE sequence recognized by the human transcription factor SREBF2 was predicted in the promoter region of the three analyzed genes: HMGS has four potential SRE sequences: one located at the [+] strand and three at the [−] strand, ERG3 has two potential SREs at the [+] strand and ERG4 has one potential SRE at the [−] strand (Supplementary Table S3).
To obtain further insight, the relative transcript levels of genes ERG3, ERG4 and HMGS were evaluated by RT-qPCR after 120 h of culture in the four X. dendrorhous strains studied in this work (Figure 5). Interestingly, transcript levels of the three genes were higher in strain CBS.cyp61–. Considering this result and that potential SRE sequences were identified in their promoter regions, their expression at the transcriptional level could indeed be regulated by Sre1, which is in its active form in strain CBS.cyp61–. Although mutants CBS.Δerg3 and CBS.Δerg4 do not produce ergosterol, in general, the transcript levels of the three genes did not change with respect to the wild-type strain. The only exception was the HMGS gene in strain CBS.Δerg4, which showed higher transcript levels than the wild-type strain; however, this increment was only approximately 2-fold in contrast to 20-fold observed in strain CBS.cyp61–. In addition, the ERG3 and ERG4 transcripts were not detected in the respective mutant strains, confirming that these mutants do not express these genes.
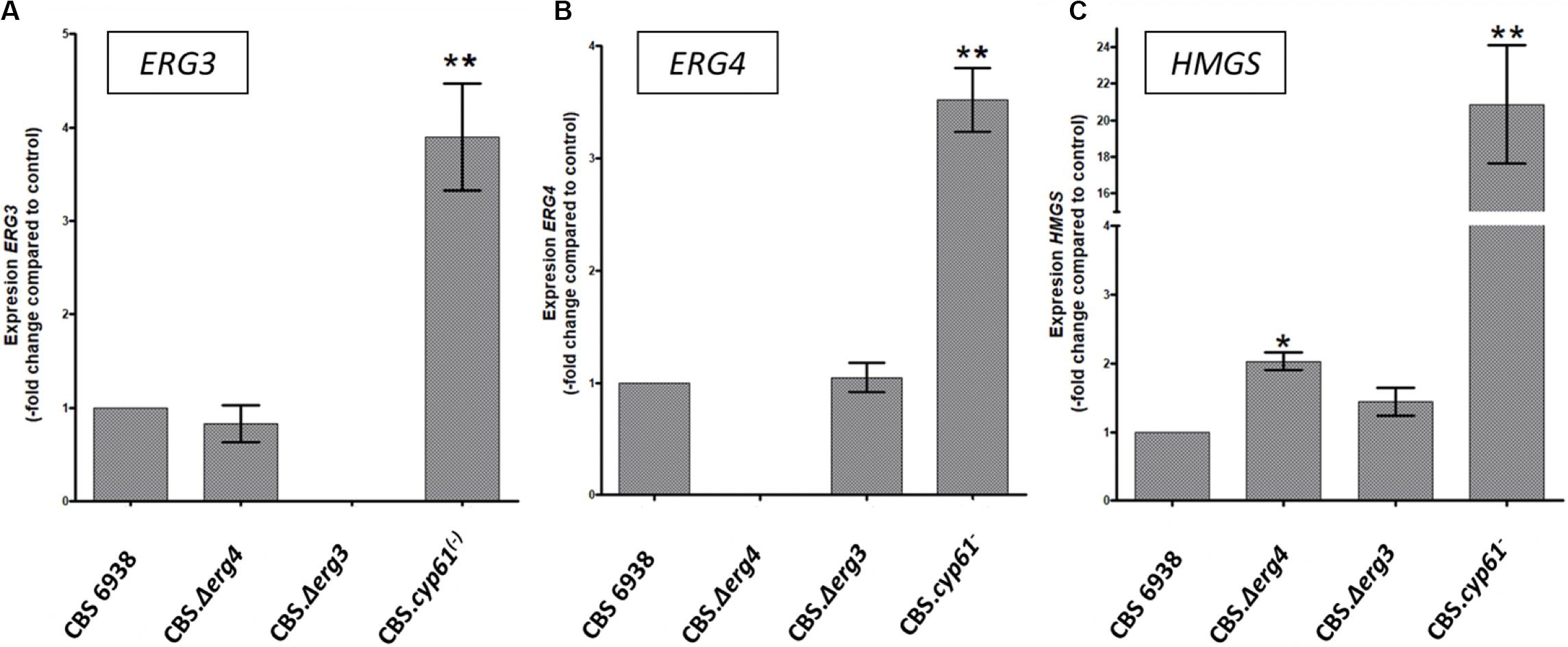
Figure 5. Transcript levels of the HMGS, ERG3 and ERG4 genes from the X. dendrorhous strains studied in this work. Relative transcript levels of genes (A) ERG3, (B) ERG4, and (C) HMGS were determined by RT-qPCR after 120 h of culture and normalized to the housekeeping gene encoding β-actin (GenBank: X89898.1). Values are the average ± standard deviation of three independent cultures (**p < 0.01, *p < 0.05; Student’s t-test).
Discussion
The genetic control of ergosterol biosynthesis has been widely studied in S. cerevisiae (Bard et al., 1996; Mo et al., 2002; Shobayashi et al., 2005), whereas in X. dendrorhous, only the CYP61/ERG5 (Loto et al., 2012) and CYP51/ERG11 (Leiva et al., 2015) genes have been functionally described. In this work, two additional genes were described: ERG3 and ERG4, encoding enzymes involved in the late steps of ergosterol biosynthesis. From bioinformatic analyses, it was predicted that the encoded proteins would be located at the endoplasmic reticulum, which is consistent with their function as in S. cerevisiae, the ergosterol biosynthesis ends in this organelle (Nishino, 1981). Additionally, it was deduced that the X. dendrorhous proteins Erg3 (328 aa) and Erg4 (485 aa) have a similar size to that described in S. cerevisiae, 365 and 473 aa, respectively (Smith and Parks, 1993; Zweytick et al., 2000). The X. dendrorhous proteins have the corresponding characteristic domains (Figure 1). Protein Erg4 has two steroid reductase domains that are present in the Erg4 and Erg24 proteins in yeast (Lai et al., 1994; Holmer et al., 1998), as both Erg4 and Erg24 are responsible for reducing double bonds in molecules. In Erg3, the hydroxylase family domain was identified; this family includes the Erg3 and Erg25 proteins of ergosterol biosynthesis. These results support that the gene product of the genes identified in this work could indeed be involved in the ergosterol biosynthesis pathway in X. dendrorhous.
High-performance liquid chromatography (HPLC) is a standard method for separating the major sterols in an extract (Knittelfelder and Kohlwein, 2017). In this work, a reproducible and accurate LC-MS/MS technique was used. Besides ergosterol, the other resulting signals presented a good resolution, indicating the suitability of this methodology to assess sterol profiles. Furthermore, this methodology is faster than the widely used gas liquid chromatography (GLC) analysis and operates under milder column temperatures and non-destructive detection conditions (Madla et al., 2012). The employed method proved to be a useful tool for rapidly determining sterol profile, compared with other chromatographic methodologies and sample preparation protocols. LC-MS/MS analysis was performed with little sample purification, avoiding losses of those molecules in low concentrations. As has been reported in other organisms (Geber et al., 1995; Zweytick et al., 2000), the genes studied in this work were not essential for X. dendrorhous viability. However, growth was slightly reduced in strain CBS.Δerg4, which could be due to the sterols accumulated in this mutant or another kind of interaction of this gene with the metabolism of the yeast. As strain CBS.Δerg4 accumulates a larger amount of ergosta-5,7,22,24(28)-tetraenol than the wild-type (metabolite that was not detected in the other strains), it is possible that this intermediary could be affecting the yeast growth, but further studies are required to support this idea. It was confirmed that the identified ERG3 and ERG4 genes are indeed involved in the biosynthesis of ergosterol, since mutants of these genes were not able to produce ergosterol, but they accumulate other sterols. Strain CBS.Δerg3 mainly accumulates episterol and strain CBS.Δerg4 mainly accumulates ergosta-5,7,22,24(28)-tetraenol, similar to what has been described in homologous S. cerevisiae mutants (Geber et al., 1995; Zweytick et al., 2000). By the approximation used in this work, it was observed that strain CBS.cyp61– mainly accumulates ergosta-5,7,24(28)-trienol.
The production of sterols increased in both mutants constructed in this work compared to the wild-type (Figure 4B), suggesting an ergosterol-mediated regulatory feedback mechanism to regulate sterol biosynthesis in X. dendrorhous, such as the SREBP pathway (Hughes et al., 2005). However, unlike in strain CBS.cyp61–, the production of carotenoids was not affected in the CBS.Δerg3 and CBS.Δerg4 mutants compared to the parental strain, nor was their content, nor was their composition (Figure 4C). These results support that it is not the absence of ergosterol that enhances carotenoid production in X. dendrorhous, but rather, what favors carotenogenesis in this yeast is the different sterol composition that the cyp61– mutants have. This finding suggests that the activation of the SREBP pathway in X. dendrorhous is not mainly due to the absence of ergosterol, but it is due to changes in the composition of sterols in the membrane, as was evidenced in S. pombe (Hughes et al., 2007). It is possible that ergosta-5,7,24(28)-trienol could play a role in the activation of the SREBP pathway in X. dendrorhous, as this was the main sterol accumulated in strain CBS.cyp61–, but further studies are required.
The step controlled by the HMGS gene has been reported as an important regulatory step of the MVA pathway; for example, the overexpression of this gene together with other genetic modifications favored the production of isoprenoid compounds in S. cerevisiae (Bröker et al., 2018). In previous works, we demonstrated that the HMGS gene is a Sre1 gene target in X. dendrorhous (Gutiérrez et al., 2019; Gómez et al., 2020). In this work, the relative transcript levels of the HMGS gene were determined, and no differences were observed between the parental strain and strain CBS.Δerg3. However, a 2-fold increase was observed in strain CBS.Δerg4 in relation to the parental strain. Even though this increment was not as high as the increment observed in strain CBS.cyp61– (Figure 5C), it is interesting to note that strain CBS.Δerg4 follows strain CBS.cyp61– regarding ergosta-5,7,24(28)-trienol content and that the wild-type strain is in third place in this aspect. Then, it is possible to speculate or suggest that the ergosta-5,7,24(28)-trienol content could be somehow related to Sre1 activation in X. dendrorhous: high levels of this sterol (as in strain CBS.cyp61–) would promote Sre1 activation, middle levels of this sterol (as in strain CBS.Δerg4) would promote some level of Sre1 activation and low levels of this sterol (as in the wild-type strain) would allow having a basal Sre1 activation level. This speculation is supported by western blot analysis of proteins extracted from the wild-type strain cultured under standard conditions (Gómez et al., 2020), where a weak band coinciding with the size of the Sre1 active form was observed, indicating that there is indeed a basal level of Sre1 proteolytic activation in the wild-type strain. However, further studies are required to support this possibility.
In summary, in this work, two genes, ERG3 and ERG4, involved in ergosterol biosynthesis in X. dendrorhous were identified and functionally described. Deletion of both genes avoided ergosterol production but exhibited a wild-type color phenotype, unlike the cyp61– ergosterol mutant, which overproduces carotenoids. The results presented in this study support that is not the absence of ergosterol what enhances carotenoid production through the activation of the SREBP pathway in X. dendrorhous, but rather this phenotype depends on the altered sterol composition observed in the cyp61– mutant.
Conclusion
The deletion of the ERG3 and ERG4 genes in X. dendrorhous prevents the synthesis of ergosterol and mutants Δerg3 and Δerg4 accumulate episterol and ergosta-5,7,22,24(28)-tetraenol, respectively. The phenotypic analysis results (production of sterols and carotenoids, and transcript levels of the studied genes), strongly suggest the SREBP pathway is activated in the mutant cyp61–, but not in the mutants constructed in this work (Δerg3 and Δerg4) as sterols and carotenoids levels (and transcript levels) are not affected in the same way. The activation of the SREBP pathway in X. dendrorhous could be somehow related to sterol composition, rather than to the absence of ergosterol in the strains.
Data Availability Statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author Contributions
All authors contributed significantly to the work and agreement with the content of the manuscript. MV, GZ, and AG performed the experiments. SB, DS, MB, VC, and JA provided strategic inputs. MV, GZ, and JA wrote the manuscript.
Funding
This work was supported by the FONDECYT 1160202 and by the graduate scholarship María Ghilardi Venegas Foundation to MV.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01312/full#supplementary-material
Footnotes
References
Adrio, J. L., and Veiga, M. (1995). Transformation of the astaxanthin-producing yeast Phaffia rhodozyma. Biotechnol. Tech. 9, 509–512. doi: 10.1007/bf00159567
An, G.-H., Schuman, D. B., and Johnson, E. A. (1989). Isolation of Phaffia rhodozyma mutants with increased astaxanthin content. Appl. Environ. Microbiol. 55, 116–124. doi: 10.1128/aem.55.1.116-124.1989
Baeza, M., Alcaíno, J., Barahona, S., Sepúlveda, D., and Cifuentes, V. (2015). Codon usage and codon context bias in Xanthophyllomyces dendrorhous. BMC Genomics 16:293. doi: 10.1186/s12864-015-1493-5
Bard, M., Bruner, D. A., Pierson, C. A., Lees, N. D., Biermann, B., Frye, L., et al. (1996). Cloning and characterization of ERG25, the Saccharomyces cerevisiae gene encoding C-4 sterol methyl oxidase. Proc. Natl. Acad. Sci. U.S.A. 93, 186–190. doi: 10.1073/pnas.93.1.186
Bien, C., and Espenshade, P. (2010). Sterol regulatory element binding proteins in fungi: hypoxic transcription factors linked to pathogenesis. Eukaryot Cell. 9, 352–359. doi: 10.1186/1471-2180-11-190
Bröker, J. N., Müller, B., van Deenen, N., Prüfer, D., and Schulze Gronover, C. (2018). Upregulating the mevalonate pathway and repressing sterol synthesis in Saccharomyces cerevisiae enhances the production of triterpenes. Appl. Microbiol. Biotechnol. 102, 6923–6934. doi: 10.1007/s00253-018-9154-7
Córdova, P., González, A.-M., Nelson, D. R., Gutiérrez, M.-S., Baeza, M., Cifuentes, V., et al. (2017). Characterization of the cytochrome P450 monooxygenase genes (P450ome) from the carotenogenic yeast Xanthophyllomyces dendrorhous. BMC Genomics 18:540. doi: 10.1186/s12864-017-3942-9
Degtyarenko, K. N., and Archakov, A. I. (1993). Molecular evolution of P450 superfamily and P450-containing monooxygenase systems. FEBS Lett. 332, 1–8. doi: 10.1016/0014-5793(93)80470-f
Geber, A., Hitchcock, C. A., Swartz, J. E., Pullen, F. S., Marsden, K. E., Kwon-chung, K. J., et al. (1995). Deletion of the Candida glabrata ERG3 and ERG11 genes: effect on cell viability, cell growth, sterol composition, and antifungal susceptibility. Antimicrob. Agents Chemother. 39, 2708–2717. doi: 10.1128/aac.39.12.2708
Golubev, W. I. (1995). Perfect state of Rhodomyces dendrorhous (Phaffia rhodozyma). Yeast 11, 101–110. doi: 10.1002/yea.320110202
Gómez, M., Gutiérrez, M. S., González, A. M., Gárate-Castro, C., Sepúlveda, D., Barahona, S., et al. (2020). Metallopeptidase Stp1 activates the transcription factor Sre1 in the carotenogenic yeast Xanthophyllomyces dendrorhous. J. Lipid Res. 61, 229–243. doi: 10.1194/jlr.RA119000431
Guerin, M., Huntley, M. E., and Olaizola, M. (2003). Haematococcus astaxanthin: applications for human health and nutrition. Trends Biotechnol. 21, 210–216. doi: 10.1016/s0167-7799(03)00078-7
Gutiérrez, M. S., Campusano, S., González, A. M., Gómez, M., Barahona, S., Sepúlveda, D., et al. (2019). Sterol Regulatory Element-Binding Protein (Sre1) promotes the synthesis of carotenoids and sterols in Xanthophyllomyces dendrorhous. Front. Microbiol. 10:586. doi: 10.3389/fmicb.2019.00586
Higuera-Ciapara, I., Felix-Valenzuela, L., and Goycoolea, F. M. (2006). Astaxanthin: a review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 46, 185–196. doi: 10.1080/10408690590957188
Hofmann, K., and Stoffel, W. (1993). TMbase - A database of membrane spanning protein segments. Biol. Chem. Hoppe Seyler 374:166.
Holmer, L., Pezhman, A., and Worman, H. J. (1998). The human lamin B receptor/sterol reductase multigene family. Genomics 54, 469–476. doi: 10.1006/geno.1998.5615
Hughes, A. L., Powell, D. W., Bard, M., Eckstein, J., Barbuch, R., Link, A. J., et al. (2007). Dap1/PGRMC1 binds and regulates cytochrome P450 enzymes. Cell Metab. 5, 143–149. doi: 10.1016/j.cmet.2006.12.009
Hughes, A. L., Todd, B. L., and Espenshade, P. J. (2005). SREBP pathway responds to sterols and functions as an oxygen sensor in fission yeast. Cell 120, 831–842. doi: 10.1016/j.cell.2005.01.012
Johnson, E. A. (2003). Phaffia rhodozyma: colorful odyssey. Int. Microbiol. 6, 169–174. doi: 10.1007/s10123-003-0130-3
Jones, P., Binns, D., Chang, H. Y., Fraser, M., Li, W., McAnulla, C., et al. (2014). InterProScan 5: genome-scale protein function classification. Bioinformatics 30, 1236–1240. doi: 10.1093/bioinformatics/btu031
Knittelfelder, O. L., and Kohlwein, S. D. (2017). Lipid extraction from yeast cells. Cold Spring Harb. Protoc. 2017, 408–411. doi: 10.1101/pdb.prot085449
Lai, M. H., Bard, M., Pierson, C. A., Alexander, J. F., Goebl, M., Carter, G. T., et al. (1994). The identification of a gene family in the Saccharomyces cerevisiae ergosterol biosynthesis pathway. Gene 140, 41–49. doi: 10.1016/0378-1119(94)90728-5
Leiva, K., Werner, N., Sepulveda, D., Barahona, S., Baeza, M., Cifuentes, V., et al. (2015). Identification and functional characterization of the CYP51 gene from the yeast Xanthophyllomyces dendrorhous that is involved in ergosterol biosynthesis. BMC Microbiol. 15:89. doi: 10.1186/s12866-015-0428-2
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Loto, I., Gutiérrez, M. S., Barahona, S., Sepúlveda, D., Martinez-Moya, P., Baeza, M., et al. (2012). Enhancement of carotenoid production by disrupting the C22-sterol desaturase gene (CYP61) in Xanthophyllomyces dendrorhous. BMC Microbiol. 12:235. doi: 10.1186/1471-2180-12-235
Madla, S., Miura, D., and Wariishi, H. (2012). Optimization of extraction method for GC-MS based metabolomics for filamentous fungi. J. Microb. Biochem. Technol. 4, 5–9. doi: 10.4172/1948-5948.1000064
Mathelier, A., Fornes, O., Arenillas, D. J., Chen, C. Y., Denay, G., Lee, J., et al. (2016). JASPAR 2016: a major expansion and update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 44, D110–D115. doi: 10.1093/nar/gkv1176
Miao, L., Chi, S., Tang, Y., Su, Z., Yin, T., Guan, G., et al. (2011). Astaxanthin biosynthesis is enhanced by high carotenogenic gene expression and decrease of fatty acids and ergosterol in a Phaffia rhodozyma mutant strain. FEMS Yeast Res. 11, 192–201. doi: 10.1111/j.1567-1364.2010.00705.x
Miao, L., Wang, Y., Chi, S., Yan, J., Guan, G., Hui, B., et al. (2010). Reduction of fatty acid flux results in enhancement of astaxanthin synthesis in a mutant strain of Phaffia rhodozyma. J. Ind. Microbiol. Biotechnol. 37, 595–602. doi: 10.1007/s10295-010-0706-9
Mo, C., Valachovic, M., Randall, S. K., Nickels, J. T., and Bard, M. (2002). Protein-protein interactions among C-4 demethylation enzymes involved in yeast sterol biosynthesis. Proc. Natl. Acad. Sci. U.S.A. 99, 9739–9744. doi: 10.1073/pnas.112202799
Nakai, K., and Horton, P. (1999). PSORT: a program for detecting sorting signals in proteins and predicting their subcellular localization. Trends Biochem. Sci. 24, 34–36.
Niklitschek, M., Alcaíno, J., Barahona, S., Sepulveda, D., Lozano, C., Carmona, M., et al. (2008). Genomic organization of the structural genes controlling the astaxanthin biosynthesis pathway of Xanthophyllomyces dendrorhous. Biol. Res. 41, 93–108.
Nishino (1981). Subcellular localization of the enzymes involved in the late stage. J. Biochem. 89, 1391–1396. doi: 10.1093/oxfordjournals.jbchem.a133330
Parker, J. E., Warrilow, A. G., Price, C. L., Mullins, J. G., Kelly, D. E., and Kelly, S. L. (2014). Resistance to antifungals that target CYP51. J. Chem. Biol. 7, 143–161. doi: 10.1007/s12154-014-0121-1
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: A Laboratory Manual, 3rd Edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Schaller, H. (2003). The role of sterols in plant growth and development. Prog. Lipid Res. 42, 163–175. doi: 10.1016/s0163-7827(02)00047-4
Shang, F., Wen, S., Wang, X., and Tan, T. (2006). Effect of nitrogen limitation on the ergosterol production by fed-batch culture of Saccharomyces cerevisiae. J. Biotechnol. 122, 285–292. doi: 10.1016/j.jbiotec.2005.11.020
Shobayashi, M., Mitsueda, S.-I., Ago, M., Fujii, T., Iwashita, K., and Iefuji, H. (2005). Effects of culture conditions on ergosterol biosynthesis by Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 69, 2381–2388. doi: 10.1271/bbb.69.2381
Skaggs, B. A., Alexander, J. F., Pierson, C. A., Schweitzer, K. S., Chun, K. T., Koegel, C., et al. (1996). Cloning and characterization of the Saccharomyces cerevisiae C-22 sterol desaturase gene, encoding a second cytochrome P-450 involved in ergosterol biosynthesis. Gene 169, 105–109. doi: 10.1016/0378-1119(95)00770-9
Smith, S. J., and Parks, L. W. (1993). The ERG3 gene in Saccharomyces cerevisiae is required for the utilization of respiratory substrates and in heme-deficient cells. Yeast 9, 1177–1187. doi: 10.1002/yea.320091104
Urban, A., Neukirchen, S., and Jaeger, K. E. (1997). A rapid and efficient method for site-directed mutagenesis using one-step overlap extension PCR. Nucleic Acids Res. 25, 2227–2228. doi: 10.1093/nar/25.11.2227
Zhang, Y., and Rao, R. (2010). Beyond ergosterol: linking pH to antifungal mechanisms. Virulence 1, 551–554. doi: 10.4161/viru.1.6.13802
Keywords: ergosterol, ERG3, ERG4, Xanthophyllomyces dendrorhous, carotenoids, sterol regulatory element-binding protein
Citation: Venegas M, Barahona S, González AM, Sepúlveda D, Zúñiga GE, Baeza M, Cifuentes V and Alcaíno J (2020) Phenotypic Analysis of Mutants of Ergosterol Biosynthesis Genes (ERG3 and ERG4) in the Red Yeast Xanthophyllomyces dendrorhous. Front. Microbiol. 11:1312. doi: 10.3389/fmicb.2020.01312
Received: 02 April 2020; Accepted: 25 May 2020;
Published: 16 June 2020.
Edited by:
Christian Sohlenkamp, National Autonomous University of Mexico, MexicoReviewed by:
Gábor Nagy, University of Szeged, HungaryGuoliang Yan, China Agricultural University, China
Copyright © 2020 Venegas, Barahona, González, Sepúlveda, Zúñiga, Baeza, Cifuentes and Alcaíno. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jennifer Alcaíno, amFsY2Fpbm9nQHVjaGlsZS5jbA==