 Achraf El Mammery1,2†
Achraf El Mammery1,2† Eva Ramírez de Arellano1,3†
Eva Ramírez de Arellano1,3† Javier E. Cañada-García1,2,3†
Javier E. Cañada-García1,2,3† Emilia Cercenado4,5
Emilia Cercenado4,5 Laura Villar-Gómara6Verónica Casquero-García1Silvia García-Cobos1José Antonio Lepe3,7Enrique Ruiz de Gopegui Bordes3,8
Laura Villar-Gómara6Verónica Casquero-García1Silvia García-Cobos1José Antonio Lepe3,7Enrique Ruiz de Gopegui Bordes3,8 Jorge Calvo-Montes3,9Nieves Larrosa Escartín3,10
Jorge Calvo-Montes3,9Nieves Larrosa Escartín3,10 Rafael Cantón3,11
Rafael Cantón3,11 María Pérez-Vázquez1,3†
María Pérez-Vázquez1,3† Belén Aracil1,3†
Belén Aracil1,3† Jesús Oteo-Iglesias1,3*† and on behalf of the Spanish EARS-Net Group
Jesús Oteo-Iglesias1,3*† and on behalf of the Spanish EARS-Net Group- 1Laboratorio de Referencia e Investigación en Resistencia a Antibióticos e Infecciones Relacionadas con la Asistencia Sanitaria, Centro Nacional de Microbiología, Instituto de Salud Carlos III, Madrid, Spain
- 2Escuela Internacional de Doctorado, Ciencias Biomédicas y Salud Pública - IMIENS (UNED), Madrid, Spain
- 3CIBER de Enfermedades Infecciosas (CIBERINFEC), Instituto Salud Carlos III, Madrid, Spain
- 4Servicio de Microbiología, Hospital General Universitario Gregorio Marañón, Madrid, Spain
- 5CIBER de Enfermedades Respiratorias (CIBERES), Instituto de Salud Carlos III, Madrid, Spain
- 6Agencia Española de Medicamentos y Productos Sanitarios (AEMPS), Madrid, Spain
- 7Servicio de Microbiología, Hospital Universitario Virgen del Rocío, Sevilla, Spain
- 8Servicio de Microbiología, Hospital Universitario Son Espases, Palma de Mallorca, Spain
- 9Servicio de Microbiología, Hospital Universitario Marqués de Valdecilla-IDIVAL, Santander, Spain
- 10Servicio de Microbiología, Hospital Universitario Vall d’Hebron, Barcelona, Spain
- 11Servicio de Microbiología, Hospital Universitario Ramón y Cajal, Madrid, Spain
Objectives: To describe and analyse erythromycin resistance trends in blood isolates of Staphylococcus aureus (EARS-Net Spain, 2004–2020) and the association of these trends with the consumption of macrolide, lincosamide, and streptogramin B (MLSB) antibiotics. To assess molecular changes that could be involved in erythromycin resistance trends by whole genome analysis of representative isolates.
Materials and methods: We collected antibiotic susceptibility data for all first-blood S. aureus isolates in patients from 47 Spanish hospitals according to EARS-Net criteria. MLSB antibiotic consumption was obtained from the Spanish Agency for Medicines and Medical Devices (2008–2020). We sequenced 137 representative isolates for core genome multilocus sequence typing, resistome and virulome analysis.
Results: For the 36,612 invasive S. aureus isolates, methicillin resistance decreased from 26.4% in 2004 to 22.4% in 2020. Erythromycin resistance in methicillin-susceptible S. aureus (MSSA) increased from 13.6% in 2004 to 28.9% in 2020 (p < 0.001); however, it decreased from 68.7 to 61.8% (p < 0.0001) in methicillin-resistant S. aureus (MRSA). Total consumption of MLSB antibiotics increased from 2.72 defined daily doses per 1,000 inhabitants per day (DID) in 2014 to 3.24 DID in 2016. By WGS, the macrolide resistance genes detected were erm (59.8%), msrA (46%), and mphC (45.2%). The erm genes were more prevalent in MSSA (44/57, 77.2%) than in MRSA (38/80, 47.5%). Most of the erm genes identified in MSSA after 2013 differed from the predominant ermC gene (17/22, 77.3%), largely because ermT was significantly associated with MSSA after 2013 (11/29, 37.9%). All 13 ermT isolates in this study, except one, belonged to ST398 and came from 10 hospitals and six Spanish provinces.
Conclusion: The significant increase in erythromycin resistance in blood MSSA correlated with the consumption of the MLSB antibiotics in Spain. These preliminary data seem support the hypothesis that the human ST398 MSSA clade with ermT-mediated resistance to erythromycin may be involved in this trend.
Introduction
The ever-increasing prevalence of antibiotic resistance in bacteria is a serious concern that requires an international approach to management. Hence, the World Health Organization (WHO) and the European Commission both recognize the importance of understanding the emergence and determinants of resistance and the need for control strategies. In Europe, the European Antimicrobial Resistance Surveillance Network (EARS-Net) has collected antimicrobial susceptibility data for isolates from routine blood and cerebrospinal fluid cultures since 1988 (Gagliotti et al., 2011). Funded by the European Commission, EARS-Net is a European network of national surveillance systems coordinated by the European Centre for Disease Prevention and Control (ECDC), whose goal is to collect comparable, reliable data to identify variations in antimicrobial resistance over time and space, providing a basis for infection prevention and control programs (ECDC, 2022).
An EARS-Net indicator organism is Staphylococcus aureus, which is a pathogen of major clinical importance for nosocomial infections. Since the introduction of antibiotics in clinical practice, S. aureus has progressively developed resistance to the most frequently used antibiotics. It is noteworthy that the first clinical isolate of methicillin-resistant S. aureus (MRSA) was reported in 1961, just 1 year after the launch of methicillin (Jevons, 1961). WHO considers MRSA the most important Gram-positive bacterial strain (Priority 2/High) for research and the development of new antibiotics (Tacconelli et al., 2018).
Macrolides, lincosamides, and streptogramin B (MLSB) are alternative antibiotics used to treat severe staphylococcal infections, mainly in penicillin-allergic patients, along with vancomycin and the combination of linezolid and rifampicin (Ji and Xu, 2021). Macrolide resistance emerges quickly and persists, even after a short course of therapy (Malhotra-Kumar et al., 2009; Van Heirstraeten et al., 2012). Total MLSB consumption did not change significantly in Europe from 2007 to 2017 (Adriaenssens et al., 2021); however, the consumption of long-acting macrolides increased, and seasonal variation was high, suggesting that MLSB antibiotics were prescribed inappropriately in many countries (Adriaenssens et al., 2021).
Staphylococcal resistance to macrolides includes target-site modification by methylation, such as erythromycin ribosome methylase (erm) genes; antibiotic efflux pumps, such as ABC-F proteins (msr genes) and major facilitator superfamily transporters (mef genes); and drug inactivation, such as phosphotransferases (mph genes) and esterases (ere genes) (Feßler et al., 2018; Lade et al., 2022). The erm gene products methylate specific targets in the 23S rRNA, preventing the antibiotic from binding to its ribosomal target (di Bonaventura et al., 2019). This is the most widespread mechanism of resistance to macrolides, lincosamides and streptogramin B cross-resistance (MLSB phenotype). The most common erm genes in S. aureus are erm(C) and erm(A), which can be either constitutive or inducible (Fyfe et al., 2016; di Bonaventura et al., 2019).
The goals of this study were (i) to describe and analyse erythromycin resistance trends in the blood isolates of S. aureus collected by EARS-Net Spain (2004–2020), (ii) to compare erythromycin resistance with trends in the consumption of the MLSB antibiotic family (J01F family), and (iii) to characterize antibiotic resistance genes and the prevalence of resistant clones by whole genome sequencing (WGS) in a representative sample of S. aureus isolates.
Materials and methods
Antibiotic resistance
Forty-seven Spanish EARS-Net hospitals collected antibiotic susceptibility data for all S. aureus isolated from the first blood collected from each patient from 2004 to 2020. Using EARS-Net criteria, we selected hospitals that were distributed evenly across the country (TESSy-The European Surveillance System, 2022). These hospitals serve ~13.5 million people, ~32% of the total Spanish population, and are representative of 15 of the 17 Spanish autonomous communities. Each laboratory identified the isolates and tested their susceptibilities according to standard microbiological procedures using commercial microdilution broth assays (Gagliotti et al., 2011, 2021). Results were interpreted according to EUCAST criteria.1 A quality assurance exercise (UK National External Quality Assessment Scheme) was performed annually to ensure comparable results among the hospital laboratories. All the S. aureus isolates included in EARS-Net have information on susceptibility to methicillin (the only mandatory indicator), but the number of isolates tested for other antibiotics may vary (Table 1).

Table 1. Evolution of antibiotic resistance of blood isolates of Staphylococcus aureus in according to EARS-Net (Spain, 2004–2020).
Antibiotic consumption
Community consumption of the MLSB antibiotic family (WHO code J01F) according to the public health prescriptions for the period 2012–2020 was provided by the Spanish National Action Plan on Antimicrobial Resistance (PRAN), coordinated by the Spanish Agency for Medicines and Medical Devices (AEMPS) from the Ministry of Health, and was obtained from the database of retail pharmacy sales from National Health System prescriptions (covering near 100% of the Spanish population). In addition, the community use of MLSB antibiotics from private health prescriptions was obtained from market research companies provided by the PRAN for the period 2012–2020. MLSB hospital dispensing data were also available from public hospital pharmacies and market research companies for private hospital pharmacies, both provided by the PRAN.
The consumption data were tabulated, and the number of units was converted into defined daily doses (DDD) of active drug ingredients according to WHO methodology (WHO Collaborating Centre for Drug Statistics Methodology, 2023). The number of DDD per 1,000 inhabitants per day (DID) was calculated for each active drug ingredient.
WGS of Staphylococcus aureus isolates
To study the population structure and macrolide resistance genes in S. aureus by WGS, we selected a total of 137 isolates sent to the S. aureus Reference Laboratory of the Spanish National Microbiology Centre according to the following characteristics: 46 MRSA isolated before 2013, 34 MRSA isolated after 2013, 28 MSSA isolated before 2013, and 29 MSSA isolates after 2013. These isolates were selected to have a broad geographic representation, and to represent MSSA and MRSA from periods before and after the start of the trend change in erythromycin resistance detected by EARS-Net at MSSA in 2013.
These isolates, which were isolated from blood (41, 29.9%), skin (35, 25.5%), the respiratory tract (34, 24.8%), and other samples (27, 19.7%), came from a total of 71 healthcare centers in 30 Spanish provinces.
Genomic library preparation and sequence analysis
Genomic library preparation and sequence analysis were conducted as described (Pérez-Vázquez et al., 2019). Raw sequence data were submitted to the European Nucleotide Archive (PRJEB61102). The quality of the short reads was assessed using FASTQC, and they were assembled into contigs with Unicycler 0.4.8 (Wick et al., 2017). The quality of the assembly was assessed with QUAST.2 Prokka v1.14-beta (Seemann, 2014) was used for automatic de novo assembly annotation.
Phylogenetic analyses
Sequence types (STs) were calculated according to the multilocus sequence typing (MLST) scheme of the Public databases for molecular typing and microbial genome diversity (PubMLST)3 using Ariba v2.6.2 (Hunt et al., 2017). A simple diversity index (SDI; Gastmeier et al., 2006) was applied to analyze population diversity. Core genome MLST (cgMLST), consisting of 1861 targets for S. aureus provided by SeqSphere+3.5.0 (Ridom, Münster, Germany), was performed with the 137 sequenced isolates. Additionally, ST398 isolates from this study were analyzed using cgMLST together with a collection of 239 S. aureus of this ST downloaded from the NCBI database using “chromosome” and “complete” as filtering criteria for assembly level and the absence of mec genes as the genotypic criterion.
Analysis of antimicrobial resistance and virulence genes
Antibiotic resistance genes were analyzed by Ariba v2-6.2 using the CARD database4 and ResFinder (CGE server5). Virulence genes were analyzed with the previous methodology using the database Virulencefinder_db.6
Statistical analysis
The significance of the trends in macrolide resistance was calculated by the χ2 test for trend. Trends in J01F antibiotic family consumption were examined by simple linear regression analysis. The strength of the association between MLSB antibiotic use and erythromycin resistance was determined by linear regression analysis (Cuevas et al., 2011). The resistance proportion was transformed to the natural logarithm of the odds of resistance. The log of the odds of resistance (as the dependent variable) was expressed as a simple linear function of the independent variable (antibiotic use) (Cuevas et al., 2011). Macrolide resistance for 2013–2020 was correlated with antibiotic use in the prior year. Fisher’s exact test was used to compare the prevalence of resistance or virulence genes by groups. p values < 0.05 were considered statistically significant. Statistical analyses were performed using GraphPad Prism software v.7.02 (GraphPad Software Inc., San Diego, CA, United States).
Results
Sentinel hospitals, patients, and isolates
Most of the participating hospitals (29, 61.7%) were tertiary university hospitals with >500 beds, and 9 (19.1%) had >1,000 beds; however, three primary (6.4%) and 15 secondary (31.9%) hospitals also participated. Data on 36,612 consecutive S. aureus blood infections, corresponding to the same number of patients, were collected between 2004 and 2020; 23,887 patients (65.2%) were males, and 34,146 (93.3%) were adults (>14 years). The blood cultures were obtained from patients in internal medicine (45.9%), the emergency room (28.2%), intensive care units (8.7%), surgery (5.9%), pediatrics (3.1%), and other departments (8.2%).
Antibiotic resistance trends in blood isolates of Staphylococcus aureus
Global resistance rates to methicillin, erythromycin, ciprofloxacin, gentamicin, and clindamycin were 24.4% (36,612 isolates analyzed), 32.7% (35,186 analyzed), 27.6% (27,838 analyzed), 11.8% (31,070 analyzed), and 5.5% (34,328 analyzed), respectively. Methicillin resistance decreased from 26.4% in 2004 to 22.4% in 2022 (Table 1) (p < 0.0001). There was a similar decrease in resistance to other antibiotics, including ciprofloxacin (from 27.7% in 2004 to 26.1% in 2020, p = 0.001) and gentamicin (from 8.5% in 2004 to 5.5% in 2020, p < 0.0001) (Table 1). For MLSB antibiotics monitored in EARS-Net, erythromycin resistance increased from 31.2% in 2004 to 36.1% in 2020 (p < 0.0001; the highest value of the series was 41.6% in 2017), and clindamycin resistance increased from 13.4% in 2004 to 18.4% in 2020 (the highest value) (p < 0.0001) (Table 1).
There was greater resistance to ciprofloxacin, gentamicin, erythromycin, and clindamycin in MRSA isolates (89.5, 13.9, 66, and 27.1%, respectively) than in MSSA isolates (7.2, 3.1, 21.9, and 7.4%, respectively) (p < 0.001). However, between 2004 and 2020, MRSA isolates showed a significant decrease in ciprofloxacin resistance, from 91.6 to 74.2% (χ2 for trend = 76.7, p < 0.0001), in gentamicin resistance, from 22.1 to 15.3% (χ2 for trend = 21.9, p < 0.0001), in erythromycin resistance, from 68.7 to 61.8% (χ2 for trend = 25.7, p < 0.0001), and in clindamycin resistance from 42.8 to 25.6% (χ2 for trend = 26.79, p < 0.0001) (Figure 1). In contrast, MSSA isolates showed an increase in erythromycin resistance from 13.6% in 2004 to 28.9% in 2020 (χ2 for trend = 747.9, p < 0.0001), with the peak in 2019 at 35.5%. This trend was also seen for clindamycin resistance, from 3.7% in 2004 to 14.4% in 2020 (χ2 for trend = 208.9, p < 0.0001), and, although more moderate, for ciprofloxacin resistance, from 4.8% in 2004 to 7.2% in 2020 (χ2 for trend = 14.6; p = 0.0001). No significant variations over time in gentamicin resistance in MSSA isolates were identified (Figure 1).
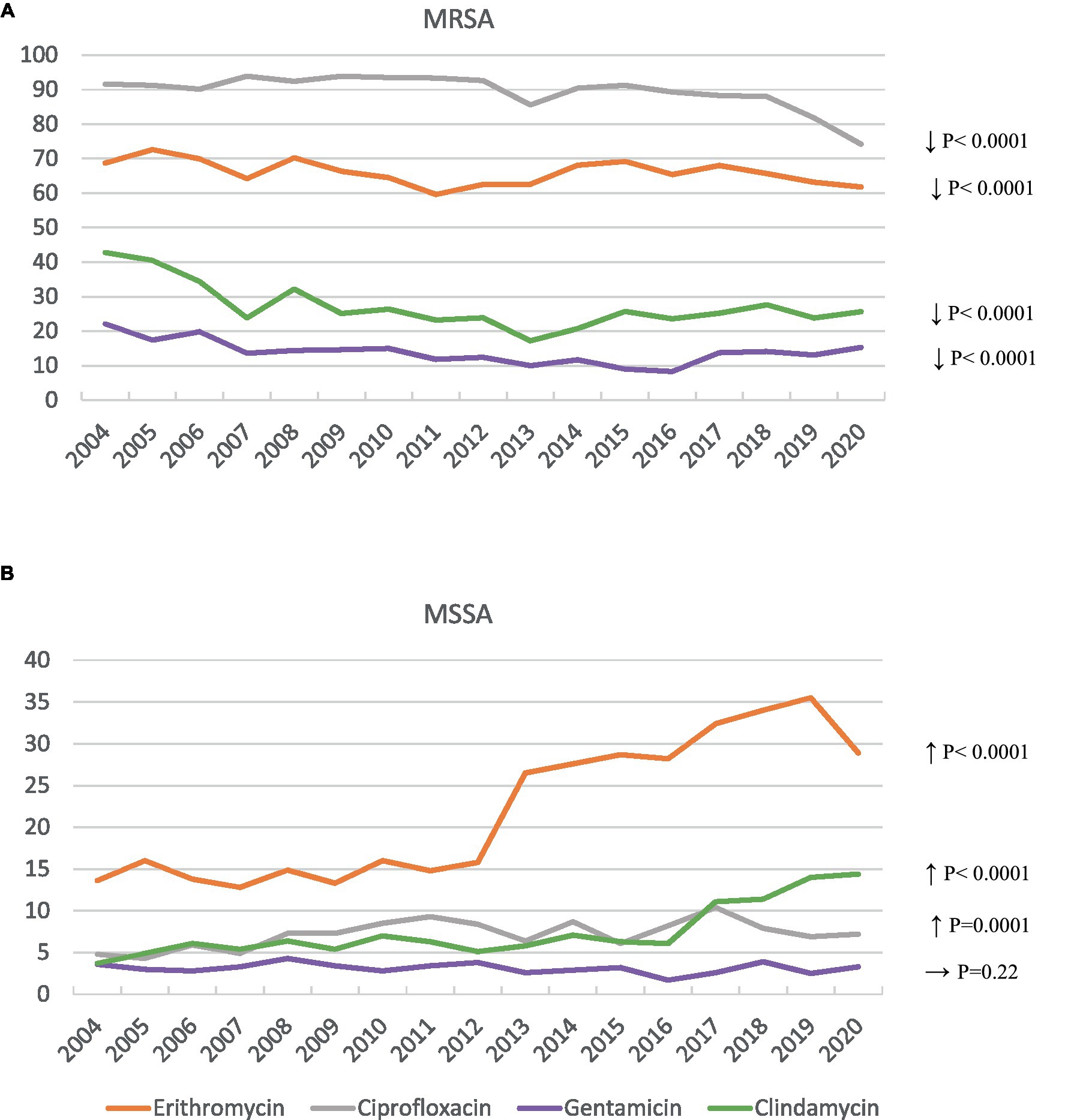
Figure 1. Antibiotic resistance trends in MRSA (A) and MSSA (B) blood isolates. (A) Shows significant decrease in resistance to ciprofloxacin, gentamicin, erythromycin and clindamycin; while (B) displays significant increase in resistance to erythromycin, clindamycin and ciprofloxacin.
Macrolides, lincosamides, and streptogramins consumption
Community use of MLSB antibiotics (WHO code J01F), including both public and private health prescriptions, increased in Spain from 2.61 DIDs in 2012 to 3.22 DIDs in 2016 (22.9%, r2 = 0.91, p = 0.012). It stabilized at around ~3.10 DIDs in 2017–2018 and decreased to 2.80 DIDs in 2019 prior to the COVID-19 pandemic in 2020, the year with the lowest use (2.04 DID) (Table 2). Nosocomial consumption of MLSB between 2012 and 2020 (Table 2) increased from 0.11 DIDs (2012) to 0.14 DIDs (2019) (27.3%, r2 = 0.84, p = 0.006) with a peak (0.17 DIDs) in 2020 likely due the pandemic and the use of macrolides in admitted COVID-19 patients. The total consumption of MLSB antibiotics (community and hospital consumption) increased between 2012 (2.72 DID) and 2016 (3.34 DID) (22.8%, r2 = 0.91, p = 0.01) stabilizing between 2017 and 2018 at ~3.23 DID (Table 2).
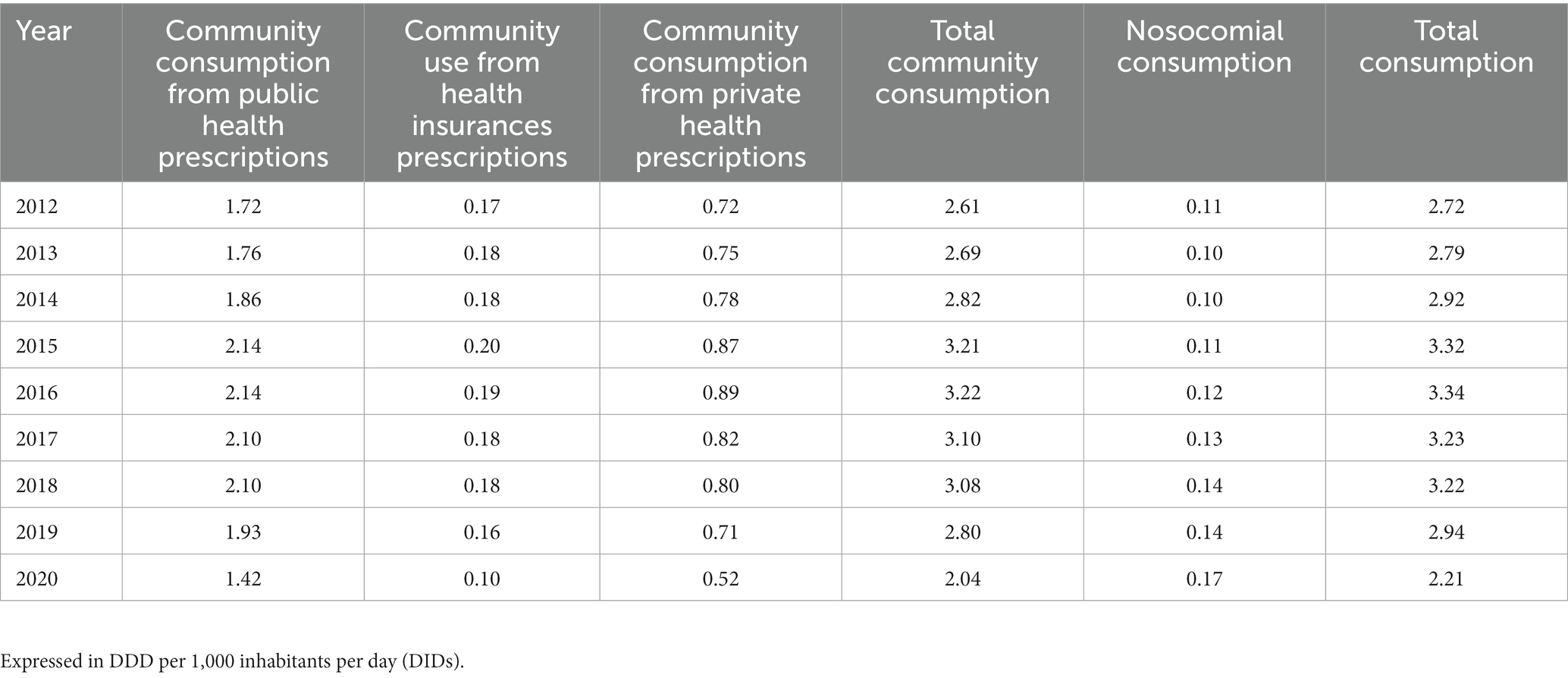
Table 2. Community and nosocomial consumption of macrolide, lincosamine, and streptogramin antibiotics group (WHO code J01F).
Correlation of erythromycin resistance in MSSA with total use of J01F antibiotics
The rates of erythromycin resistance in MSSA from 2013 to 2020 correlated with the total use of the J01F family of antibiotics (r2 = 0.55, p = 0.04) in the previous year (2012–2019) (Figure 2). However, when the resistance data for 2016 (representing atypical outlier data of unknown origin) were removed from the series, the correlation improved (r2 = 0.85, p = 0.003) (Figure 2).
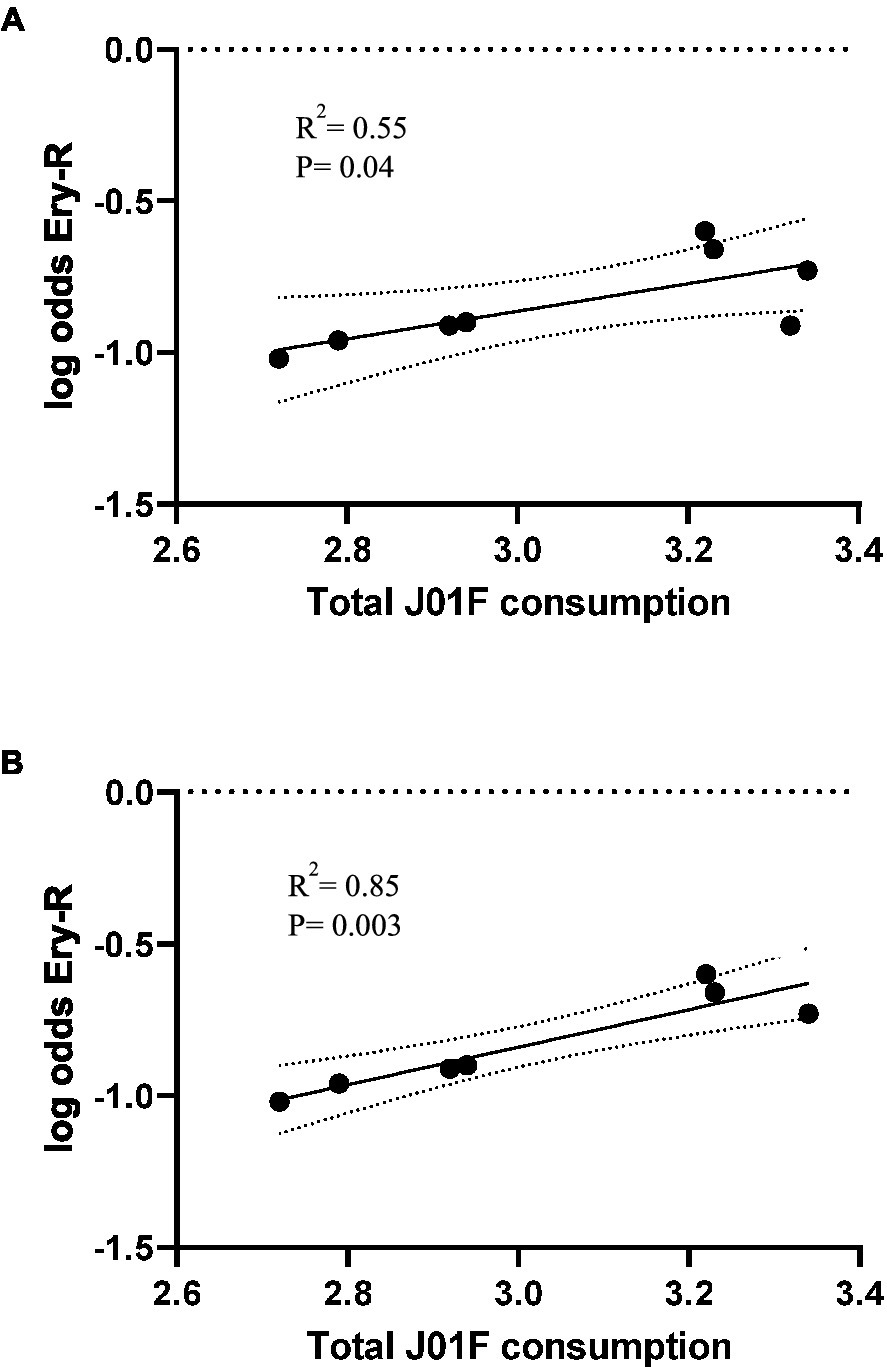
Figure 2. Occurrence of erythromycin resistance in S. aureus causing blood infections (years 2013–2020), plotted against total use of family J01F antibiotics (2012–2019) in Spain with 95% confidence intervals (A). In (B), the outlier erythromycin resistance data of 2016 (consumption of 2015) is excluded. Consumption is expressed in DIDs, DDDs/1000 inhabitants/day. Log odds is the natural logarithm of the OR.
Resistance genes in erythromycin-resistant Staphylococcus aureus isolates by WGS
WGS of the 137 isolates identified erm genes in 83 (60.6%) isolates, a msr(A) gene in 63 (46%), and an mph(C) gene in 62 (45.2%). Five erm genes were identified, including erm(C) (49, 35.8%), erm(A) (21, 15.3%), erm(T) (13, 9.5%), erm(B) (one, 0.7%), and erm(X) (one, 0.7%); two isolates had two erm genes (Supplementary Table S1). A total of 61 isolates (44.5%) had both msr(A) and mph(C) genes, and 9 (6.6%) also carried an erm gene (Table 3). Of the 73 erythromycin-resistant isolates that had only erm genes, 19 (26%) showed a constitutive expression (84.2% had the erm(C) gene), and 54 (74%) were inducible (48.1% had the erm(C) gene). This predominance of constitutive erm(C) genes was statistically significant (p = 0.01). The erm genes were more prevalent in MSSA (45/57, 78.9%) than in MRSA (38/80, 47.5%) (p = 0.0007), whereas msr(A) predominated in MRSA (50/80, 62.5%) vs. MSSA (13/57, 22.8%) (p < 0.0001) (Table 3). There was no significant difference in macrolide resistance genes present in the isolates collected before or after 2013, although the erm genes were more prevalent after 2013 (41/63; 65.1%) than before 2013 (42/74, 56.7%).
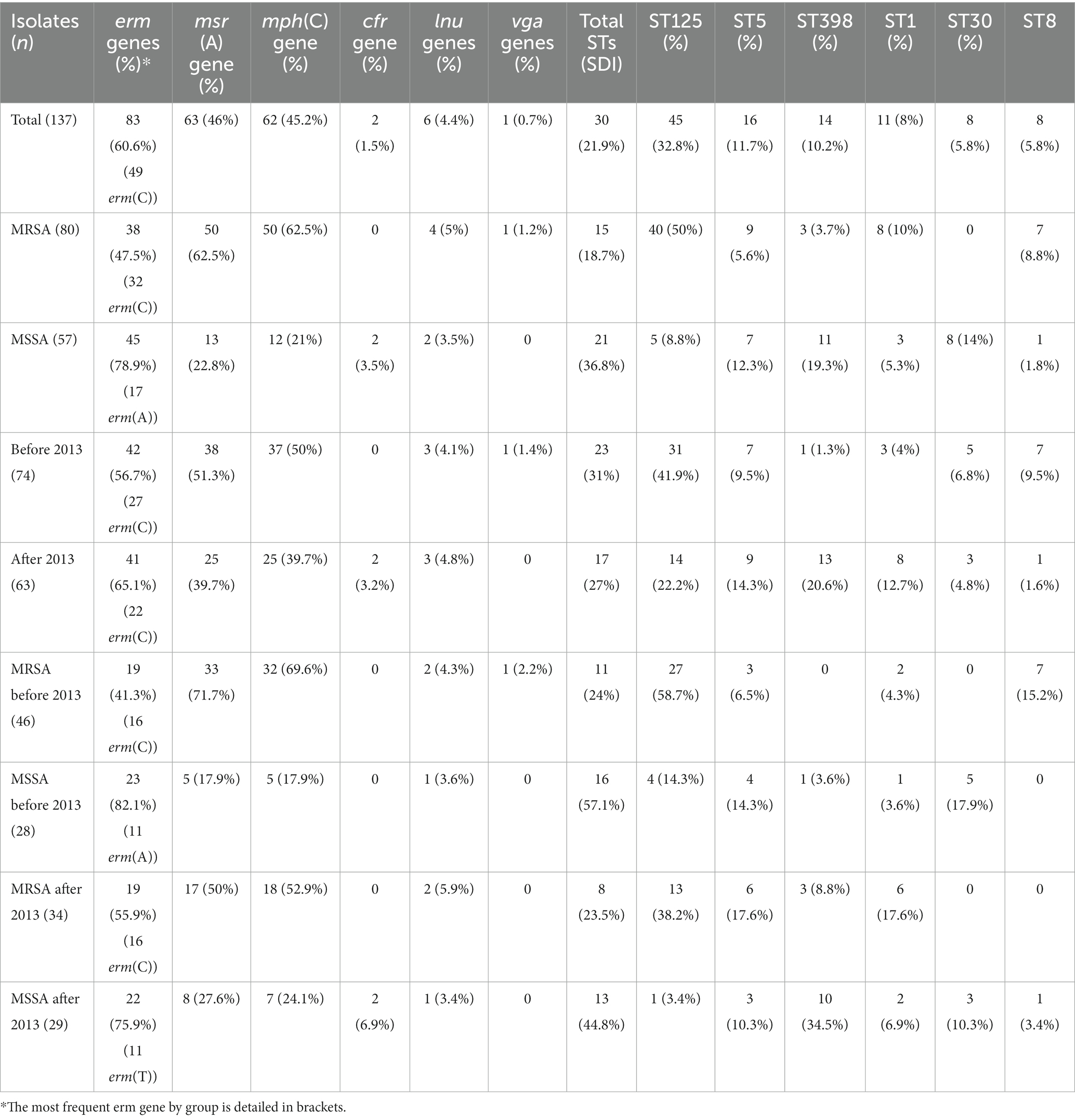
Table 3. Distribution of macrolide and lincosamide resistance genes and main sequence types in S. aureus according to the groups studied.
For the MSSA isolates collected after 2013, the group showing the most EARS-Net-based increase in erythromycin resistance, erm gene prevalence was 75.9% of (22/29), similar to MSSA strains isolated before 2013 (23/28, 82.1%) (Table 3). However, after 2013, the erm genes in MSSA were mostly different from the predominant erm(C) and included 11 strains with erm(T) and six with erm(A) (17/22, 77.3%) (Table 3; Supplementary Table S1). In fact, the erm(T) gene was significantly associated with MSSA isolated after 2013 (11/29, 37.9%), whereas it was identified in only two of the other isolates (2/108, 1.8%) (p < 0.0001). All 13 isolates with erm(T), except one, belonged to ST398 and were collected from 10 hospitals in six different Spanish provinces. The sequence of the genetic environment of erm(T) genes identified the rep13 gene, which is involved in plasmid replication.
Nine isolates (6.6%) carried resistance genes for lincosamides (Supplementary Table S1), including lincosamide nucleotidyltransferases (lnu) genes in six isolates [five lnu(A) and one lnu(B)], genes encoding the ABC-F proteins vga and lsa in one isolate each, and the cfr methylase gene in two isolates. One isolate had both lnu(B) and lsa (Table 3).
All MRSA had the mecA gene, and 80% (64/80) of MRSA and 29.8% (17/57) of MSSA had resistance genes to at least one of the three main aminoglycosides (gentamicin, tobramycin, or amikacin) (Supplementary Table S1). The predominant genes encoding resistance to aminoglycosides were the aph (3′)-IIIa gene encoding a phosphotransferase with resistance to amikacin (60/137, 43.8%), the aadD1 gene encoding a nucleotidyl transferase with resistance to tobramycin (45/137, 32.8%), and the aac (6′)-Ie/aph (2″)-Ia encoding a two-domain acetyltransferase/phosphotransferase enzyme with resistance to the three aminoglycosides (12/137, 8.8%). The aac (6′)-Ie/aph (2″)-Ia gene was observed in 10% (8/80) of MRSA and 7% (4/57) of MSSA. No changes over time were observed for aminoglycoside resistance genes.
We identified mutations in the genes encoding topoisomerase II and/or topoisomerase IV in 53.3% (73/137) of the isolates, 78.7% (63/80) were in MRSA, and 17.5% (10/57) were in MSSA. Frequent mutations were S84L in gyrA (71/73, 97.3% of all isolates with mutations in this gene) and S80F in parC (95.9%) (Supplementary Table S1). Fifteen isolates had more than two mutations in topoisomerase II/IV genes.
Phylogenetic analysis of erythromycin-resistant Staphylococcus aureus isolates by WGS
The 137 erythromycin-resistant S. aureus studied by MLST were grouped into 30 STs with an SDI of 21.9 and a mean of 4.6 isolates per ST (range = 1–45). The most prevalent STs (≥5 isolates) were ST125 (45, 32.8%), ST5 (16, 11.7%), ST398 (14, 10.2%), ST1 (11, 8%), ST30 (8, 5.8%), and ST8 (8, 5.8%), accounting for 74.4% of all STs (Supplementary Table S1).
Genome assemblies of 134 S. aureus isolates were analyzed using a gene-by-gene approach, and the allelic distance from cgMLST was visualized in a minimum spanning tree (Figure 3). All isolates differed by an average of 880 alleles (range = 0–1779), and the average allelic difference between ST125 isolates from this study was 60 (range = 0–121). ST5 isolates differed by an average of 188 alleles (range = 38–339); in ST398 isolates, it was 133 alleles (range = 1–265); in ST1 isolates, it was 126 alleles (range = 0–253); in ST30 isolates it was 213 alleles (range = 134–237); and in ST8 isolates, it was 155 alleles (range = 61–250).

Figure 3. Population structure of Staphylococcus aureus isolates from this study: minimum-spanning tree. Distances shown are based on cgMLST of 1861 genes using the parameter “pairwise ignoring missing values.” Fill colors in each circle indicate MSSA and MRSA and the year of isolation, color of the dashed line in circles indicates macrolide resistance mechanism type.
ST125 was more prevalent in MRSA than in MSSA isolates (40/80 vs. 5/57, respectively, p < 0.0001); however, ST30 (0/80 vs. 8/57, respectively; p = 0.0007) and ST398 (3/80 vs. 11/57, respectively; p = 0.004) were more prevalent in MSSA (Figure 3). There were no differences in the distribution of STs in isolates before or after 2013, except for ST8, which was more prevalent in isolates prior to 2013 (7/74 vs. 1/63, p = 0.047), and ST398, which was more prevalent in isolates after 2013 (1/74 vs. 13/63, p = 0.0003).
Regarding to erythromycin-resistant mechanism, ST1, ST30 and ST398 were more frequent in erm isolates (10/82, 12.2%; 8/82, 9.6%; and 13/82, 15.8%; respectively) than in msr(A) isolates (1/63, 1.6%; 0/63; and 1/63, 1.6%; respectively) (p < 0.05). The prevalence of ST398/erm(T) in MSSA strains isolated after 2013 (10/29, 34.5%) was much higher compared to the other isolates (2/108, 1.8%) (p < 0.0001) (Figure 3).
The comparative analysis of ST398/erm(T) isolates included in this study with a collection S. aureus ST398 isolates from NCBI database showed that the most frequent mechanism of macrolide resistance was erm gene production, being erm(C) most frequent one followed by erm(A), erm(B) and erm(T). All MSSA isolates with erm(T) are grouped, MSSA erm(T) isolates from this study are grouped in a cluster together with animal-independent ST398 MSSA isolates reported in New York (Uhlemann et al., 2012) (Figure 4).

Figure 4. Population structure of ST398 Staphylococcus aureus in comparison with other publicly available whole genomes of the same ST. Minimum spanning tree, distance based in a cgMLST scheme of 1861 genes. Fill colors in each circle indicate MSSA and MRSA from this study or from other origins. Blue dashed line around circles remark isolates with erm(T) genes and green dashed line represent isolate NC_017631.1 (Uhlemann et al., 2012). Gray shadows represent groups of strains; a threshold of 5 alleles was applied.
Virulence genes in erythromycin-resistant Staphylococcus aureus isolates identified by WGS
The most common virulence genes (detected in ≥14 isolates) in the 137 isolates were the alpha-toxin gene hla (136, 99.3%), complement inhibitor gene scn (107, 78.1%), staphylokinase gene sak (90, 65.7%), chemotaxis inhibitory protein gene chp (87, 63.5%), beta-toxin gene hlb (32, 23.4%), hyaluronidase gene hysA (20, 14.6%), enterotoxin gene sea (15, 10.9%), and toxic shock syndrome toxin 1 gene tsst-1 (14, 10.2%) (Supplementary Table S1).
Other important virulence genes in S. aureus, such as Panton-Valentine leucocidin (PVL) genes lukS and lukF, and exfoliative toxin A gene eta were detected in eight (5.8%; 75% of them belonging to ST8) and six (4.4, 66.7% belonging to ST15) isolates, respectively (Supplementary Table S1). We found no pattern of virulence genes associated with the different groups, except for the presence of the lukF/luKS genes in the MRSA isolates. Of the genes mentioned above, the ST398/erm(T) isolates only presented homogeneously the hla (100%), chp (91.7%) and scn (91.7%) genes.
Discussion
In addition to methicillin resistance, the cross-resistance to MLSB antibiotics among S. aureus strains, as well as the rapid transmission of resistance genes, is a major concern for the future efficacy of antibiotic therapy. Specifically, the extensive use of MLSB antibiotics against Gram-positive bacteria is of concern because macrolide-resistant MRSA strains are believed to be a major cause of clinical infections (Miklasińska-Majdanik, 2021), and they are associated with increased mortality rates (Bishr et al., 2021). In our study, erythromycin resistance in MRSA was approximately three times more common than in MSSA. However, our major concern that led to this research was the significant increase in erythromycin-resistant MSSA blood isolates that were not observed in MRSA.
The main strengths of this study were the analysis of bacteraemia caused by S. aureus, representing a broad national caseload over 17 years, in conjunction with antibiotic consumption data. Our molecular analyses represent a pilot study of a representative sample of S. aureus isolates to develop hypotheses about the EARS-Net findings, which will then require further studies. EARS-Net is coordinated by ECDC to collect, analyse, and report data on antimicrobial resistance through a network of national surveillance systems across Europe and to take actions to address antimicrobial resistance. During its more than 20 years of operation, EARS-Net has been effective in detecting changes in trends in antibiotic resistance in S. aureus, both at a national and a European level (Gagliotti et al., 2021).
The increase in erythromycin resistance in MSSA correlated temporally with an increase in the consumption of MLSB in Spain; however, in 2016, outlier erythromycin resistance data reduced the statistical significance of the association. The cause of this single year of atypical data is unknown. The overall rise in MLSB antibiotics consumption in this study was probably mostly due to the increased prescription of these antibiotics for community-acquired respiratory infections, and specifically the prescriptions of 3-day azithromycin courses. The association between macrolide consumption and increased resistance to MLSB antibiotics has been described previously, especially in the context of specific pathologies such as cystic fibrosis (Tramper-Stranders et al., 2007) or trachoma (Bojang et al., 2017). Although the use of antibiotics (even appropriate usage) entails inevitable selection of resistance, this can be mitigated by the implementation of stewardship programs, including the diverse and combined use of antibiotics.
Although erm(C) is the most common macrolide resistance gene in S. aureus worldwide, it varies by geographical region and by the phenotype of susceptibility or resistance to methicillin (Miklasińska-Majdanik, 2021). Although we confirmed the general dominance of erm(C), we found a high prevalence of msr(A), which was most common in MRSA. A study in Spain (2006–2007) showed 23.1% prevalence for erm genes and 15.8% for msr genes in invasive S. aureus (Pérez-Vázquez et al., 2009).
The present study showed a strong association between msr(A) and mph(C) genes; only 2/63 isolates with msr(A) lacked mph(C). Matsuoka et al. (2003) found the mph(C) phosphotransferase gene on plasmid pMS97, 342 bp downstream of the msr(A) gene. These authors suggested, based on their findings, that a region of the msr(A) gene is required for the full expression of mph(C). However, that plasmid also carried erm(Y) (15), a gene that we did not find in our study.
One of the main findings of this study was the prevalence of erm (T)-bearing ST398 isolates in MSSA collected during the period in which EARS-Net found a significant increase in erythromycin resistance in MSSA isolates that produced bacteraemia. Although this pilot study included a limited number of representative isolates, our finding suggests that the increased resistance to macrolides in these MSSA identified by EARS-Net could be due, at least in part, to the spread of ST398 in this group of isolates. ST398 is relevant to public health because methicillin-resistant strains are associated with livestock capable of infecting humans (Silva et al., 2023). However, previous studies suggested two subpopulations in clonal complex 398 S. aureus—a human-adapted clade mostly with the erm(T) gene, and an animal-associated clade with mecA, tet(M), and erm(C) genes (Price et al., 2012; Argudín et al., 2018). A recent Belgian study identified different ST398 subpopulations, including typical human and animal clades, as well as new emerging mixed subpopulations that underlie the ability of this lineage to acquire resistance and virulence genes (Argudín et al., 2018). The proximity of erm(T) and rep13 genes in MSSA ST398 isolates suggests that erm(T) is on a plasmid, as has been described recently (Salgueiro et al., 2023). The MSSA ST398 human clade was reported mainly in China and France (Bouiller et al., 2020), and was frequently implicated in severe infections, whereas the MRSA ST398 animal clade was reported mainly in the skin and soft tissue (Bouiller et al., 2020).
Most well-known staphylococcal virulence genes, such as enterotoxins, toxic shock syndrome toxin, or PVL, were absent in ST398 isolates from this study, as previously communicated (Bouiller et al., 2020). However, previous studies have suggested that MSSA ST398, which is prevalent in bacteraemia, may be a more virulent subtype (Bouiller et al., 2020) with higher mortality (Bouiller et al., 2016). Among the virulence factors previously associated with ST398 MSSA, only scn and chp genes were identified in erm(T) ST398 isolates of our study, but the immune evasion cluster (IEC) sak and sea genes were not found.
A limitation of this research could be the reduced number of sequenced isolates by group, which, although they were strictly chosen to be representative of the issue to study, not all belonged to the same collection in which the increase in resistance was primarily detected.
Conclusion
The highly significant increase in resistance to macrolides in MSSA causing bacteraemia, in contrast to the absence of this trend in MRSA, correlated with the consumption of MLSB antibiotics in Spain. Our data seem to support the hypothesis that the human ST398 MSSA clade with erm(T)-mediated resistance to erythromycin may be involved in this trend. An epidemiological surveillance system for MRSA and MSSA is important to monitor the emergence of dangerous new S. aureus subpopulations. Further WGS research is needed to identify the emergence of these subpopulations in the clinical setting, as well as their correlation with changes in the patterns of antibiotic susceptibility.
Data availability statement
The data presented in the study are deposited in the European Nucleotide Archive (ENA) repository (https://www.ebi.ac.uk/ena), accession number PRJEB61102.
Ethics statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Spanish EARS-Net group
Members of the Spanish EARS-Net group are as follows: Amparo Coira Nieto and Daniel Navarro de la Cruz (EOXI de Santiago de Compostela, A Coruña); Elena Escribano, Mª Dolores Crespo and Juan José Palomar (H. Universitario de Albacete, Albacete); Mónica Parra (H. General Universitario de Alicante, Alicante); Nieves Gonzalo and Gloria Royo (H. General de Elche, Elche, Alicante); María Navarro and Nieves Gonzalo (H. de Orihuela, Orihuela, Alicante); Teresa Cabezas (H. de Poniente, El Ejido, Almería); Javier Fernández Domínguez and Ana Mª Fleites (H. Universitario Central de Asturias, Oviedo, Asturias); Eugenio Garduño (H. Infanta Cristina, Badajoz); Belén Viñado and Oscar del Valle (H. Universitari Vall d’Hebrón, Barcelona); Isabel Sanfeliú and Dionisia Fontanals (Corporació Sanitaria Parc Taulí, Sabadell, Barcelona); José Lite and Javier Garau (H. Mutua Terrassa, Terrassa, Barcelona); Pilar Berdonces (H. de Galdakao, Bizkaia); Mª Angeles Mantecón and Eva Ojeda (Complejo Asistencial de Burgos, Burgos); Rosario Sánchez Benito, Jesús Viñuelas and Pilar Teno (H. San Pedro de Alcántara, Cáceres); Carmen Martínez Rubio, Carolina Freyre and Manuel Antonio Rodríguez Iglesias (H. Universitario de Puerto Real, Cádiz); Natalia Montiel and Manuel Antonio Rodríguez Iglesias (H. Universitario Puerta del Mar, Cádiz); José Luis de Francisco Ramírez, Juan Carlos Alados and Mª Dolores López Prieto (H. Universitario de Jerez de la Frontera, Cádiz); Susana Sabater and Rosario Moreno (H. General de Castellón, Castellón); Alberto Yagüe, Óscar Pérez Olaso and María Gil Fortuño (H. La Plana, Villareal, Castellón); Ana Bordes Benítez and Raúl Guilarranz Luengo (H. Universitario Dr. Negrín, Las Palmas de Gran Canaria); Fernando Cobo Martínez, Mª Dolores Pérez Ramírez and Dolores Rojo (H. Virgen de las Nieves, Granada); Alejandro González Praetorius (H. Universitario de Guadalajara, Guadalajara); José Antonio Lepe (H. Río tinto, Huelva); Inocente Cuesta and Concepción Carazo (H. Universitario Ciudad de Jaén, Jáen); Carmen Amores (H. San Agustín, Linares, Jaén); M. José Gastañares (H. San Millán, Logroño, La Rioja); Isabel Fernández- Natal (Complejo Asistencial Universitario de León, León); María Rodríguez Velasco, Carmen Raya and Carlos Fúster (H. del Bierzo, Ponferrada, León); Ana Mª Sánchez Díaz, Elena Loza and Ana Verónica Halperin (H. Universitario Ramón y Cajal, Madrid); Mª José González and Mercedes Menéndez Rivas (H. Niño Jesús, Madrid); José Francisco Valverde Cánovas and Alberto Delgado Iribarren (Fundación H. de Alcorcón, Madrid); Jesús García Martínez (H. Universitario de Fuenlabrada, Madrid); Sara Quevedo, Isabel Wilhelmi and Pilar Reyes (H. Severo Ochoa, Leganés, Madrid); Isabel Sánchez Romero (H. General Universitario Puerta de Hierro, Majadahonda, Madrid); Mª Victoria García-López and Alfonso Pinedo (H. Virgen de la Victoria, Málaga); Fernando Fernández Sánchez and Natalia Montiel (H. Costa del Sol, Marbella, Málaga); Xabier Berastain (H. Universitario de Navarra, Navarra); Isabel Paz Vidal, Begoña Fernández, Gloria Esteban and Almudena Tinajas (Complejo Hospìtalario de Orense, Orense); Almudena Tinajas (Complejo Hospitalario Asistencial de Palencia); Antonio Oliver and José Luis Pérez (H universitario son Espases, Palma de Mallorca); Mª Ángeles Pallarés, Marta García Campello, Victoria Pullian and Mª Ángeles Pascual (Complejo Hospitalario de Pontevedra, Pontevedra); Francisco Vasallo (H. Universitario de Vigo, Pontevedra); Ana Isabel Aller, Estrella Martín and Samuel Bernal (H. Universitario Valme, Sevilla); José Antonio Lepe (H. Universitario Virgen del Rocío, Sevilla); Carmen Aldea Mansilla, Ángel Campos and Teresa Nebreda (Complejo Asistencial de Soria, Soria); Mar Olga Pérez Moreno, Ignacio Buj and Mª José Centelles (H. Virgen de la Cinta, Tortosa, Tarragona); Teresa Delgado Melián, Mª Antonia de Miguel and Antonio Sierra (H. Universitario de Canarias, Tenerife); Juan Frasquet (H. Universitari i Politècnic La Fe, Valencia); Mª Fe Brezmes and Juana Rodríguez-Hernández (H. Virgen de la Concha, Zamora); Ana López Caleja, Mª Cruz Villuendas, Javier Pereira, Luisa Marco and Mª José Revillo (H. Universitario Miguel Servet, Zaragoza); Carmen Aspiroz (H. Royo Villanova, Zaragoza).
Author contributions
JO-I, BA, and MP-V conceived, designed, and coordinated the study. AE, ERa, JC-G, MP-V, VC-G, JL, ERu, JC-M, NL, RC, EC, SG-C, and Spanish EARS-Net Group collected data and isolates and performed the experiments. AE, BA, and JO-I analyzed resistance trends. LV-G and JO-I collected and analyzed antibiotic consumption trends. JO-I, AE, ERa, JC-G, and MP-V wrote the manuscript. All authors have read, edited and approved the final manuscript.
Funding
This research was supported by CIBER—Consorcio Centro de Investigación Biomédica en Red (CB21/13/00095, CB21/13/00006, CB21/13/00054, CB21/13/00068, CB21/13/00084, CB21/13/00099 groups of CIBERINFEC; CB06/06/0058 group of CIBERES), Instituto de Salud Carlos III, Ministerio de Ciencia e Innovación and Unión Europea-NextGenerationEU. This research was also supported by Personalized and precision medicine grant from the Instituto de Salud Carlos III (MePRAM Project, PMP22/00092), and by the Antibiotic Resistance and Staphylococcus aureus Surveillance Programs of the National Center for Microbiology, Instituto de Salud Carlos III.
Acknowledgments
We thank all participating hospitals and the Genomics Unit of the Centro Nacional de Microbiología for support with DNA sequencing. We also thank the European publicly funded system for antimicrobial resistance of the ECDC EARS-Net, and the Spanish National Plan against Antibiotic Resistance (PRAN).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2023.1220286/full#supplementary-material
Footnotes
1. ^https://www.eucast.org/clinical_breakpoints
2. ^http://quast.bioinf.spbau.ru/, accessed on 3 March 2023.
3. ^https://pubmlst.org/organisms/staphylococcus-aureus
4. ^https://card.mcmaster.ca, accessed on 3 December 2022.
5. ^https://cge.cbs.dtu.dk, accessed on 3 December 2022.
6. ^https://bitbucket.org/genomicepidemiology/virulencefinder_db/src/master/, version 2022-12-02.
References
Adriaenssens, N., Bruyndonckx, R., Versporten, A., Hens, N., Monnet, D. L., Molenberghs, G., et al. (2021). Consumption of macrolides, lincosamides and streptogramins in the community, European Union/European economic area, 1997-2017. J. Antimicrob. Chemother. 76, ii30–ii36. doi: 10.1093/jac/dkab175
Argudín, M. A., Deplano, A., Vandendriessche, S., Dodémont, M., Nonhoff, C., Denis, O., et al. (2018). CC398 Staphylococcus aureus subpopulations in Belgian patients. Eur. J. Clin. Microbiol. Infect. Dis. 37, 911–916. doi: 10.1007/s10096-018-3205-y
Bishr, A. S., Abdelaziz, S. M., Yahia, I. S., Yassien, M. A., Hassouna, N. A., and Aboshanab, K. M. (2021). Association of macrolide resistance genotypes and synergistic antibiotic combinations for combating macrolide-resistant MRSA recovered from hospitalized patients. Biology 10:624. doi: 10.3390/biology10070624
Bojang, E., Jafali, J., Perreten, V., Hart, J., Harding-Esch, E. M., Sillah, A., et al. (2017). Short-term increase in prevalence of nasopharyngeal carriage of macrolide-resistant Staphylococcus aureus following mass drug administration with azithromycin for trachoma control. BMC Microbiol. 17:75. doi: 10.1186/s12866-017-0982-x
Bouiller, K., Bertrand, X., Hocquet, D., and Chirouze, C. (2020). Human infection of methicillin-susceptible Staphylococcus aureus CC398: a review. Microorganisms 8, 1–19. doi: 10.3390/microorganisms8111737
Bouiller, K., Gbaguidi-Haore, H., Hocquet, D., Cholley, P., Bertrand, X., and Chirouze, C. (2016). Clonal complex 398 methicillin-susceptible Staphylococcus aureus bloodstream infections are associated with high mortality. Clin. Microbiol. Infect. 22, 451–455. doi: 10.1016/j.cmi.2016.01.018
Cuevas, O., Oteo, J., Lazaro, E., Aracil, B., de Abajo, F., Garcia-Cobos, S., et al. (2011). Significant ecological impact on the progression of fluoroquinolone resistance in Escherichia coli with increased community use of moxifloxacin, levofloxacin and amoxicillin/clavulanic acid. J. Antimicrob. Chemother. 66, 664–669. doi: 10.1093/jac/dkq471
di Bonaventura, G., Pompilio, A., Monaco, M., Pimentel de Araujo, F., Baldassarri, L., Pantosti, A., et al. (2019). Adhesion and biofilm formation by Staphylococcus aureus clinical isolates under conditions relevant to the host: relationship with macrolide resistance and clonal lineages. J. Med. Microbiol. 68, 148–160. doi: 10.1099/jmm.0.000893
ECDC (2022). European antimicrobial resistance surveillance network (EARS-net). Eur. Cent. Dis. Prev. Control. Available at: https://www.ecdc.europa.eu/en/about-us/partnerships-and-networks/disease-and-laboratory-networks/ears-net (Accessed March 28, 2023).
Feßler, A. T., Wang, Y., Wu, C., and Schwarz, S. (2018). Mobile macrolide resistance genes in staphylococci. Plasmid 99, 2–10. doi: 10.1016/j.plasmid.2018.05.001
Fyfe, C., Grossman, T. H., Kerstein, K., and Sutcliffe, J. (2016). Resistance to macrolide antibiotics in public health pathogens. Cold Spring Harb. Perspect. Med. 6, 1–37. doi: 10.1101/cshperspect.a025395
Gagliotti, C., Balode, A., Baquero, F., Degener, J., Grundmann, H., Gür, D., et al. (2011). Escherichia coli and Staphylococcus aureus: bad news and good news from the European antimicrobial resistance surveillance network (EARS-net, formerly EARSS), 2002 to 2009. Eur. Secur. 16, 1–5. doi: 10.2807/ese.16.11.19819-en
Gagliotti, C., Högberg, L. D., Billström, H., Eckmanns, T., Giske, C. G., Heuer, O. E., et al. (2021). Staphylococcus aureus bloodstream infections: diverging trends of meticillin-resistant and meticillin-susceptible isolates, EU/EEA, 2005 to 2018. Eur. Secur. 26:2002094. doi: 10.2807/1560-7917.ES.2021.26.46.2002094
Gastmeier, P., Schwab, F., Bärwolff, S., Rüden, H., and Grundmann, H. (2006). Correlation between the genetic diversity of nosocomial pathogens and their survival time in intensive care units. J. Hosp. Infect. 62, 181–186. doi: 10.1016/j.jhin.2005.08.010
Hunt, M., Mather, A. E., Sánchez-Busó, L., Page, A. J., Parkhill, J., Keane, J. A., et al. (2017). ARIBA: rapid antimicrobial resistance genotyping directly from sequencing reads. Microb. Genomics 3:e000131. doi: 10.1099/mgen.0.000131
Jevons, M. P. (1961). Celbenin-resistant staphylococci. Br. Med. J. 1, 124–125. doi: 10.1136/bmj.1.5219.124-a
Ji, C., and Xu, X. (2021). Recent advancements in macrolide hybrids against Staphylococcus aureus. Curr. Top. Med. Chem. 21, 2455–2473. doi: 10.2174/1568026620999201203213733
Lade, H., Joo, H. S., and Kim, J. S. (2022). Molecular basis of non-β-lactam antibiotics resistance in Staphylococcus aureus. Antibiotics 11, 1–28. doi: 10.3390/antibiotics11101378
Malhotra-Kumar, S., Mazzariol, A., van Heirstraeten, L., Lammens, C., de Rijk, P., Cornaglia, G., et al. (2009). Unusual resistance patterns in macrolide-resistant Streptococcus pyogenes harbouring erm (a). J. Antimicrob. Chemother. 63, 42–46. doi: 10.1093/jac/dkn432
Matsuoka, M., Inoue, M., Endo, Y., and Nakajima, Y. (2003). Characteristic expression of three genes, msr (a), mph (C) and erm (Y), that confer resistance to macrolide antibiotics on Staphylococcus aureus. FEMS Microbiol. Lett. 220, 287–293. doi: 10.1016/S0378-1097(03)00134-4
Miklasińska-Majdanik, M. (2021). Mechanisms of resistance to macrolide antibiotics among Staphylococcus aureus. Antibiotics 10:1406. doi: 10.3390/antibiotics10111406
Pérez-Vázquez, M., Sola Campoy, P. J., Ortega, A., Bautista, V., Monzón, S., Ruiz-Carrascoso, G., et al. (2019). Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob. Chemother. 74, 3489–3496. doi: 10.1093/jac/dkz366
Pérez-Vázquez, M., Vindel, A., Marcos, C., Oteo, J., Cuevas, O., Trincado, P., et al. (2009). Spread of invasive Spanish Staphylococcus aureus spa-type t067 associated with a high prevalence of the aminoglycoside-modifying enzyme gene ant (4′)-Ia and the efflux pump genes msr a/msr B. J. Antimicrob. Chemother. 63, 21–31. doi: 10.1093/jac/dkn430
Price, L. B., Stegger, M., Hasman, H., Aziz, M., Larsen, J., Andersen, P. S., et al. (2012). Staphylococcus aureus CC398: host adaptation and emergence of methicillin resistance in livestock. MBio 3, 1–6. doi: 10.1128/mBio.00305-11
Salgueiro, V., Manageiro, V., Bandarra, N. M., Ferreira, E., Clemente, L., and Caniça, M. (2023). First comparative genomic characterization of the MSSA ST398 lineage detected in aquaculture and other reservoirs. Front. Microbiol. 14, 1–9. doi: 10.3389/fmicb.2023.1035547
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Silva, V., Araújo, S., Monteiro, A., Eira, J., Pereira, J. E., Maltez, L., et al. (2023). Staphylococcus aureus and MRSA in livestock: antimicrobial resistance and genetic lineages. Microorganisms 11:124. doi: 10.3390/microorganisms11010124
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
TESSy-The European Surveillance System (2022). Antimicrobial resistance (AMR) reporting protocol 2022 European antimicrobial resistance surveillance network (EARS-net) surveillance data for 2021. Available at: https://www.ecdc.europa.eu/sites/default/files/documents/EARS-Net-reporting-protocol-2022_0.pdf.
Tramper-Stranders, G. A., Wolfs, T. F. W., Fleer, A., Kimpen, J. L. L., and Van Der Ent, C. K. (2007). Maintenance azithromycin treatment in pediatric patients with cystic fibrosis: long-term outcomes related to macrolide resistance and pulmonary function. Pediatr. Infect. Dis. J. 26, 8–12. doi: 10.1097/01.inf.0000247109.44249.ac
Uhlemann, A. C., Porcella, S. F., Trivedi, S., Sullivan, S. B., Hafer, C., Kennedy, A. D., et al. (2012). Identification of a highly transmissible animal-independent Staphylococcus aureus ST398 clone with distinct genomic and cell adhesion properties. MBio 3, 1–9. doi: 10.1128/mBio.00027-12
Van Heirstraeten, L., Coenen, S., Lammens, C., Hens, N., Goossens, H., and Malhotra-Kumar, S. (2012). Antimicrobial drug use and macrolide-resistant Streptococcus pyogenes, Belgium. Emerg. Infect. Dis. 18, 1515–1518. doi: 10.3201/eid1809.120049
WHO Collaborating Centre for Drug Statistics Methodology (2023). Guidelines for ATC classification and DDD assignment|2023. Oslo: WHO Available at: https://www.whocc.no/filearchive/publications/2023_guidelines_web.pdf.
Keywords: Staphylococcus aureus, macrolides, ST398, ermT, EARS-Net, antibiotic resistance, antibiotic consumption
Citation: El Mammery A, Ramírez de Arellano E, Cañada-García JE, Cercenado E, Villar-Gómara L, Casquero-García V, García-Cobos S, Lepe JA, Ruiz de Gopegui Bordes E, Calvo-Montes J, Larrosa Escartín N, Cantón R, Pérez-Vázquez M, Aracil B and Oteo-Iglesias J (2023) An increase in erythromycin resistance in methicillin-susceptible Staphylococcus aureus from blood correlates with the use of macrolide/lincosamide/streptogramin antibiotics. EARS-Net Spain (2004–2020). Front. Microbiol. 14:1220286. doi: 10.3389/fmicb.2023.1220286
Edited by:
Kristina Kadlec, Independent Researcher, Wunstorf, GermanyReviewed by:
Juan-ignacio Alós, Getafe University Hospital, SpainMeiji Soe Aung, Sapporo Medical University, Japan
Etinosa Igbinosa, University of Benin, Nigeria
Copyright © 2023 El Mammery, Ramírez de Arellano, Cañada-García, Cercenado, Villar-Gómara, Casquero-García, García-Cobos, Lepe, Ruiz de Gopegui Bordes, Calvo-Montes, Larrosa Escartín, Cantón, Pérez-Vázquez, Aracil and Oteo-Iglesias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jesús Oteo-Iglesias, amVzdXMub3Rlb0Bpc2NpaWkuZXM=
†These authors have contributed equally to this work