 Wu Xiang
Wu Xiang Han Xiang
Han Xiang Junyu Wang
Junyu Wang Yiqin Jiang1
Yiqin Jiang1- 1Department of Rehabilitation, Beibei Traditional Chinese Medical Hospital, Chongqing, China
- 2Department of Rehabilitation Medicine, Shanghai Fourth People’s Hospital Affiliated to Tongji University School of Medicine, Shanghai, China
- 3Department of Radiology, Daping Hospital, Army Medical University, Chongqing, China
Alzheimer’s disease is a common neurological disorder, which has become one of the major factors affecting human health due to its serious impact on individuals, families and society. It has been confirmed that gut microbiota can affect the occurrence and development of Alzheimer’s disease. Especially, fecal microbiota transplantation plays a positive role in the treatment of Alzheimer’s disease. The mechanisms for improving Alzheimer’s disease might include anti-inflammation and regulation of amyloid β-protein, synaptic plasticity, short-chain fatty acids, and histone acetylation. In this mini-review, the relationship between fecal microbiota transplantation and Alzheimer’s disease was summarized. It is hoped that fecal microbiota transplantation would play a positive role in the prevention and treatment of Alzheimer’s disease in the future.
1. Introduction
Alzheimer’s disease (AD) is a common neurodegenerative disease and the most common cause of dementia. The pathological features of AD mainly include deposition of amyloid β-protein (Aβ) (Duyckaerts et al., 2009; Bloom, 2014; Jouanne et al., 2017), neurofibrillary tangles caused by hyperphosphorylated tau protein (Duyckaerts et al., 2009; Bloom, 2014), and enhancement of neuroinflammation (Felsky et al., 2019). AD is usually characterized by cognitive deficits, as well as impairments in expressive speech, visuospatial processing, and executive function (Knopman et al., 2021). Epidemiology studies show that over 50 million people worldwide have been affected by AD (Hodson, 2018). According to the World Alzheimer’s Disease Report, the number of people with dementia is expected to increase to 139 million by 2050 due to an aging population, among which AD accounting for approximately 60–80% (World Alzheimer Report, 2022).
The etiology of AD may be the result of the interaction between multiple factors, including age, genetic factors, family history, lifestyle actors, and environmental factors. For example, the percentage of AD patients increases dramatically with age. It has been found that 5% of people age 65 to 74, 13.1% of people age 75 to 84 and 33.3% of people age 85 and older suffer from AD (Rajan et al., 2021; Wang et al., 2021). Meanwhile, many genes can increase the risk of AD, among which apolipoprotein E4 is the greatest effect on risk of late-onset AD (Bellenguez et al., 2022). People who have or had a parent or sibling with AD are more likely to develop the disorder (Loy et al., 2014). Genetic and non-genetic factors (e.g., diet and exercise) may play a role when disorders spread within families (Alzheimer’s Association, 2023). In addition, gender (World Alzheimer Report, 2022), smoking (Jeong et al., 2023), education (Manly et al., 2022), traumatic head injury (Schneider et al., 2021), cardiovascular disease (Samieri et al., 2018), gut microbiota (Kim et al., 2020), and other heterogeneous factors (Abolhasani et al., 2023) have been reported to be related to AD. These studies show that AD poses an enormous threat to global health because of high incidence rate, lacking effective efficacious pharmacotherapy and poor prognosis. Therefore, it is necessary to develop new therapeutic approaches to solve this disease.
There are about 100 trillion microorganisms in intestine (Valdes et al., 2018), which play an important role in maintaining the balance of the human body. At the same time, due to their key role in the regulation of the central nervous system, the gut microbiota is named the human “second brain” (Ridaura and Belkaid, 2015). There is a gut-brain axis between gut microbiota and the central nervous system (Morais et al., 2021; Mayer et al., 2022). The gut-brain axis consists of bidirectional communication between the enteric nervous system and the central nervous system, including the immune system, tryptophan metabolism, the vagus nerve and the enteric nervous system (Cryan et al., 2019). Relevant studies have confirmed that gut microbiota can slow down the AD progression by regulating brain function (Dodiya et al., 2019; Elangovan et al., 2019; Fujii et al., 2019; Zhou et al., 2019; Hazan, 2020; Park et al., 2021; Jin et al., 2023).
The gut microbiota of patients with AD is disordered. Research shows that the diversity of α and β of gut microbiota was reduced in patients with AD compared to healthy people (Liu et al., 2019). The gut microbiota disturbance has also been demonstrated in animal models (Harach et al., 2017). Currently, it has become a new strategy in the treatment of AD by targeting regulation of gut microbiota. Prebiotics (Sun et al., 2019a), probiotics (Akbari et al., 2016), and antibiotics (Angelucci et al., 2019) are associated with improvement of prognosis in AD. However, fecal microbiota transplantation (FMT) may be more effective, which involve a broader range of microbiome modifications than prebiotics, probiotics, or antibiotics (Smits et al., 2013). It has been shown that the symptoms of AD can be improved by FMT. Compared with mice from the same background and conventional breeding, FMT can regulate the disturbance of gut microbiota and improve their cognitive function in AD mice (Yang, 2018; Sun et al., 2019b).
In this mini review, the current knowledge about the relationship between gut microbiota and AD has been introduced, including the relationship between AD and gut microbiota, as well as the possible mechanisms of the effects of FMT. In addition, the research progress in FMT for AD has also been summarized (Table 1). Finally, we emphasize that FMT may become a new therapy for the treatment of AD.
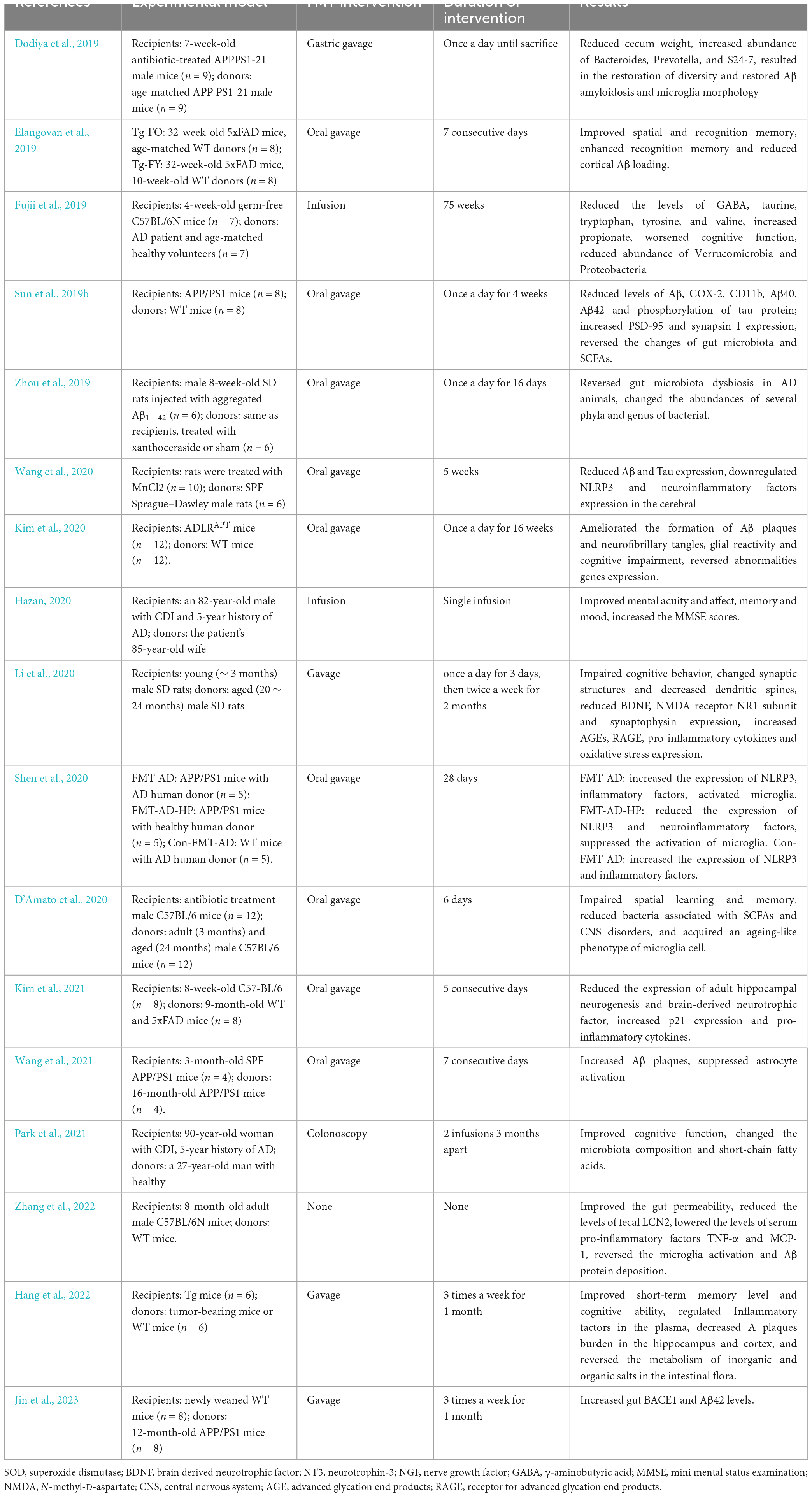
Table 1. Summary of the application of FMT in the intervention of AD.
2. Alzheimer’s disease and gut microbiota
As the largest microbiota in the human body, the human gut microbiota has over 1000 species. “Healthy microbiome” is defined as an ideal set of a healthy “functional core”: a complement of metabolic and other molecular functions that are performed by the microbiome within a particular habitat (Shafquat et al., 2014). Such a core might need to be present as genetic potential, and it must include at least the housekeeping functions necessary for individual microbial life (Lloyd-Price et al., 2016). Meanwhile, a healthy microbiome may be characterized further by the resistance of a microbiome to stress and perturbation and its ability to recover to a healthy functional profile afterward (Bodelier, 2011; Backhed et al., 2012).
Several studies have shown that the gut microbiota of AD patients or model animals is disrupted. For example, compared with age matched wild-type mice, the abundance of Odoribacter and Helicobacter increased in AD mice, while Prevotella decreased (Shen et al., 2017). Another study found that the abundance of Verrucomicrobia and Proteobacteria increased in AD mice, while Ruminococcus and Butyricicoccus decreased (Zhang et al., 2017). In clinical studies, the abundance of Firmicutes and Actinobacteria decreased in AD patients, while Bacteroidetes increased. Meanwhile, the phylum with elevated abundance was negatively correlated with Aβ1-42/Aβ1-40, and positively correlated with P-tau and P-tau/Aβ1-42 (Vogt et al., 2017).
Previous studies have found that transplanting gut microbiota of AD patients or mice into germ-free mice can lead to significant cognitive deficits in object localization and recognition. In contrast, FMT of healthy patients or mice can reverse symptoms and pathologic manifestations in AD model animals. For example, it has been demonstrated that cognitive deficits caused by deposition of Aβ and neurofibrillary tangles can be improved by FMT from healthy mice donors (Kim et al., 2020). Meanwhile, FMT could improve cognitive function by reducing deposition of Aβ, attenuating glial cell hyperactivation and secondary neuroinflammation, and decreasing blood-brain barrier (BBB) permeability (Sun et al., 2019b; Kim et al., 2021). The above evidence suggest that gut microbiota is closely related to the occurrence and development of AD. Based on the existing evidence, it is suggested that improving cognitive function by regulating gut microbiota may provide new ideas for the prevention and treatment of AD.
3. Definition and process of FMT
Fecal microbiota transplantation is a technology that places stool from a healthy donor into another patient’s gastrointestinal tract to change the recipient’s gut microbiota, thereby gaining therapeutic benefits (Gupta and Khanna, 2017). The first records of FMT have been traced back to the fourth century China, where it was used in patients with severe diarrhea (Zhang et al., 2012). Studies have shown that FMT has a positive effect on various diseases, including Clostridium difficile infection (CDI) (Hamilton et al., 2012), inflammatory bowel disease (Paramsothy et al., 2017), metabolic syndrome (Mocanu et al., 2021), autoimmune disorders (Yang et al., 2023), and neurological disorders (Xu et al., 2015). It is worth noting that FMT is also beneficial for AD (Yang, 2018; Sun et al., 2019b).
The process of FMT is rigorous. First of all, strict donor screening tests of FMT are needed (Cammarota et al., 2017), including a donor questionnaire, additional interview, standard donor screening protocols and the time between screening and donation. Secondly, patients undergoing FMT need support and education prior to treatment and antibiotics are avoided 12–48 h before fecal infusion (Blackburn et al., 2015). Finally, the current administration of stool by means include oral capsule, lower gastrointestinal route (via colonoscopy or retention enema) and upper gastrointestinal route (via nasogastric, nasojejunal, esophagogastroduodenoscopy, or nasoduodenal tube) (Wang et al., 2019). For example, a study showed that a non-significant difference in cure rate of recurrent CDI between upper and lower gastrointestinal routes of FMT (Youngster et al., 2014). However, another study found that the lower gastrointestinal route had higher clinical cure rate than the upper gastrointestinal route in CDI patients (Kassam et al., 2013). Furthermore, FMT via oral capsules had comparable results to delivery by colonoscopy in prevention of recurrent CDI (Kao et al., 2017). To sum up, there is no current strong evidence of the optimal FMT measure has been proved in clinical treatment, and it is recommended to select according to individual situation of patients.
4. Biological mechanisms of FMT to improve AD
More and more evidence showed that FMT may have potential for the prevention and treatment of AD. A randomized controlled trial suggested that the abundance of Proteobacteria and Verrucomicrobia decreased, while Bacteroidetes increased, in the FMT-treated mice. Meanwhile, the spatial learning ability and familiarity with novelty performed of FMT-treated mice better than AD model mice (Sun et al., 2019b). A randomized controlled trial suggested that the abundance of Proteobacteria and Verrucomicrobia decreased, while Bacteroidetes increased, in the FMT-treated mice. Meanwhile, the spatial learning ability and familiarity with novelty performed better than AD model mice (Sun et al., 2019b). Another study showed that FMT could reverse the disturbance of gut microbiota in AD mice and enhance the learning and memory ability (Yang, 2018). Numerous studies have shown that FMT can improve AD, which may be related to reducing the abundance of pathogenic bacteria, exerting the anti-inflammatory effects, decreasing the deposition of Aβ, regulating synaptic plasticity, increasing short chain fatty acids, and curbing histone acetylation.
4.1. Anti-inflammatory mechanisms
Inflammation plays an important role in the occurrence and development of many diseases, including metabolic disorders, immune system disorders, cardiovascular diseases and nervous system diseases.
The high expression of inflammatory factors can lead to metabolic disorders of neurotransmitters and disrupt the regulation and signaling mechanisms of behavior and cognition (Johnson et al., 2021). Research have found that the level of interleukin-1β (IL-1β), interleukin-6 (IL-6) and tumor necrosis factor-α (TNF-α) is significantly increased in the cerebrospinal fluid and peripheral blood of AD patients (Cattaneo et al., 2017). The symptoms of AD could be improved by inhibition of pro-inflammatory cytokines and enhancement of anti-inflammatory cytokines. For example, the defect of exercise behavior induced by lipopolysaccharide improved and the number of activated microglia of nigra reduced while the IL-1 receptor antagonists was added to the brain of P70 newborn rats (Pang et al., 2015). Meanwhile, the cognitive and memory function of transgenic male mice with IL-6 knock-out was lower than wild-type control mice (Hryniewicz et al., 2007). A 6-month prospective, single-center, open study showed that the cognitive function of AD patients was improved by TNF-α inhibitor enalapril (Tobinick et al., 2006). In addition, FMT could increase the level of anti-inflammatory cytokines, such as interleukin-10 (IL-10) and interleukin-22 (IL-22) (Kim et al., 2021).
Fecal microbiota transplantation can inhibit the level of pro-inflammatory cytokines and increase anti-inflammatory cytokines. For example, a study showed that manganese could induce the deposition of Aβ and production of tau protein, increasing inflammatory cytokines (e.g., IL-1β) and NLRP3 inflammasomes. FMT from healthy rat donors relieve neurotoxicity by reversing the above changes, but the composition of FMT is unclear (Wang et al., 2020). Another study found that FMT from AD mice donors led to impaired cognitive impairment, activated microglia, and increased NLRP3 and inflammatory factors (including IL-1β, IL-18, and TNF-α). The above changes could be reversed by FMT from healthy people donors, but the composition of FMT is unclear (Shen et al., 2020). In addition, the level of pro-inflammatory cytokines increased, such as TNF-α and monocyte chemoattractant protein-1 (MCP-1), in AD mice. Pearson correlation analysis showed that Ruminiclostridium_5 was positively correlated with inflammatory markers TNF-α and MCP-1. Nevertheless, FMT from wild type mice significantly decreased serum TNF-α and MCP-1 levels, which may be associated with reducing the abundance of Ruminiclostridium_5 (Zhang et al., 2022). Hang et al. (2022) showed that FMT treatment could increase the level of anti-inflammatory factors IL-2 and transforming growth factor-β (TGF-β) and reduce pro-inflammatory factors TNF-α and IL-1β in AD mice. The results may be associated with the increased Firmicutes and Prevotella and the decreased Bacteroidetes, Bacteroides and Sutterella. Yang (2018) found that the level of anti-inflammatory factors increased, such as IL-10, and the pro-inflammatory cytokines decreased, such as TNFα and IL-6, in the FMT-treated mice. The results may be associated with the increased Firmicutes, Bacteroidales, Verrucomicrobia, Clostridia, and Bacteroidia, and the decreased Erysipelotrichia, Clostridia, and Bacteroidia. Xu et al. (2020) showed that yeast β-glucans could increase beneficial bacteria and reduce pathogenic bacteria in the gut microbiota of AD mice, and reverse the increased IL-1β, IL-5, IL-6, and INF- γ and the decreased IL-10. Meanwhile, spearman correlation analyses showed that there were three negatively correlated (Oscillibacter, Butyricicoccus, and Mucispirillum) and two positively correlated (Lactobacillus and Bifidobacterium) with anti-inflammatory. Inversely, there were two negatively correlated (Lactobacillus and Bifidobacterium) and six positively correlated (Alistipes, Oscillibacter, Butyricicoccus, Rikenella, Mucispirillum, and Anaerotruncus) with pro-inflammatory. The above studies suggest that FMT may have a positive effect on cognitive function by regulating the gut microbiota, reducing the expression of pro-inflammatory cytokines, and increasing anti-inflammatory factors.
4.2. Regulating Aβ protein and synaptic plasticity
Previous studies have shown that excessive deposition of Aβ can accelerate the cascade of oxidative stress and neuroinflammation, and induce apoptosis of nerve cells. It has been reported that Aβ42 aggregation is the main cause of Aβ42 toxic protein, and the latter is the major reason of AD. The development of AD can be effectively prevented by inhibiting Aβ42 aggregation (Lansbury and Lashuel, 2006). Meanwhile, high level of Aβ was detected throughout the intestines of both mice and humans. The expression of Aβ42 increased in colons of mice after receiving the gut microbiota of aged APP/PS1 mice donors (Jin et al., 2023). Moreover, low density lipoprotein receptor-related protein 1 (LRP-1) overexpressing expression mice had reduced the level of Aβ in the blood and brain, which was associated with spatial learning, memory consolidation, and spatial recognition memory (Cheng et al., 2023). The above studies suggest a correlation between Aβ42 and AD.
It has been demonstrated that Aβ targets the modulation of synapses (Almeida et al., 2005), and impairment of synaptic plasticity is associated with cognitive decline in AD (D’Amato et al., 2020).
The definition of synaptic plasticity is the ability of synapses to adjust their function or shift shape in response to changes of internal and external environment. The time-dependent appearance of neurofibrillary tangles after deposition of Aβ can alter synaptic function, affect synaptic plasticity and cause synaptic loss in AD patients. Meanwhile, the memory and synaptic plasticity were damaged by oxidative stress induced by the imbalance between antioxidants and free radicals (Sehar et al., 2022). The change of synaptic plasticity in AD patients is characterized by synaptic dysfunction and impairment of synaptic morphology and structure, which regulated by neurotransmitters, synaptic plasticity related proteins and signaling pathways.
In recent years, the main target of synaptic plasticity mechanism studies is synapse-associated proteins, including growth-associated protein-43, synaptophysin, and postsynaptic density protein-95 (PSD-95). The synaptic function is inhibited by reduction of synapse-associated proteins. The expression of synaptic protein and PSD-95 decreased in AD mice. For example, the expression of PSD-95 in Tg2576 APP mutant neurons was lower than wild-type neurons (Almeida et al., 2005). In contrast, the synaptic plasticity enhanced by increasing the expression of PSD-95, thereby improving the reduction of synapse associated protein. Research showed that FMT from aged donors led to changed expression of proteins involved in synaptic plasticity and neurotransmission in young adult recipients. Meanwhile, four genera (Prevotellaceae, Faecalibaculum, Lachnospiraceae, and Ruminococcaceae) were found to be significantly differentially abundant compared to adult mice. There were three (Faecalibaculum, Lachnospiraceae, and Ruminococcaceae) were significantly correlated with proteins implicated in mitochondrial energy metabolism and neurotransmitter transport (D’Amato et al., 2020). Furthermore, FMT from aged donors led to significant changes in synaptic structure of young rats, which may be associated with reducing the abundance of Bacteroidetes, Prevotella, Bacteroides, and Parabacteroides. However, FMT treatment could reduce the brain deposition of Aβ40 and Aβ42 and increase the expression of PSD-95 and synapsin I in AD model mice, which may be associated with increasing the abundance of Desulfovibrionaceae (Sun et al., 2019b). Therefore, we speculate that FMT may improve AD by reducing deposition of Aβ and inducing synapse recurrent (Li et al., 2020).
4.3. Regulating the short chain fatty acids and histone acetylation
Short chain fatty acids (SCFAs) are the main metabolites of the gut microbiota, including acetate, propionate, butyrate, pentanoate and Caproate. SCFAs can regulate human homeostasis and play critical roles in biological functions. Many studies have shown that SCFAs are closely related to the occurrence and development of AD. A study found that the acetate concentration decreased in the AD model drosophila induced by Aβ deposition (Kong et al., 2021). Meanwhile, the concentrations of propionate, butyrate, and isobutyric acid in AD model mice was lower than wild-type mice (Zheng et al., 2019). In clinical studies, the expression of SCFAs in AD patients was the lowest compared to healthy humans and patients with mild cognitive impairment (Wu et al., 2021). An increase in SCFAs can reduce deposition of Aβ and improve cognitive impairment in AD mice. For example, the reshaped gut microbiome and enhanced butyrate formation are highly associated with behavioral alteration and brain oxidative status, and SCFAs attenuated the behavioral disorders and Aβ accumulation in AD mice (Liu et al., 2021a). Furthermore, there was a strong correlation between the levels of increased SCFAs and reduced cognitive functions in AD mice (Liu et al., 2021b). These studies show that SCFAs play an important role in the occurrence and development of AD.
There is a closely relation between histone deacetylases (HDACs) and AD. In the hippocampus, HDAC1, HDAC2, and HDAC8 are reported to strengthen neuroinflammation associated with cognitive dysfunction, while HDAC3 decreases dendritic spine density and levels of proteins associated with synaptic plasticity (Yang et al., 2022). Meanwhile, it has been reported that the increased activity of HDAC6 related to memory impairment (Li et al., 2021). However, treatment with acetate was shown to reduce mRNA levels of HDAC2, HDAC5, HDAC7, and HDAC8 (Huang et al., 2021), which indicate that SCFAs can regulate the expression of HDACs. According to the report, SCFAs (such as butyrate) can inhibit the activity of histone deacetylase (Tan et al., 2014), reduce deposition of Aβ in neurons. Treatment with butyrate can improve cognitive functioning in AD mice by adding hippocampal acetylation and increasing the expression of genes associated with synaptic plasticity (Govindarajan et al., 2011). At the same time, butyrate can serve as a histone deacetylase inhibitor to enhance the acetylation of histones adjacent to the neurotrophic factor promoter, thereby improving memory (Barichello et al., 2015). Furthermore, acetyl CoA converted by acetate can serve as an acetyl donor for histone acetylation, thus altering the inflammatory signaling in the microglia of brains (Shi and Tu, 2015). The above studies suggest that SCFAs may improve AD by affecting histone acetylation.
Many studies showed that FMT treatment could improve AD by regulating gut microbiota and increasing the expression of SCFAs. For example, a study showed that FMT from aged donors led to impaired spatial learning and memory in young adult recipients, which may be associated with a strong reduction of bacteria associated with SCFAs production (Lachnospiraceae, Faecalibaculum, and Ruminococcaceae) (D’Amato et al., 2020). However, yeast β-glucans could reverse the increased Firmicutes, Oscillibacter, Mucispirillum, and Butyricicoccus, and the decreased Bacteroidetes, Lactobacillus, and Bifidobacterium in AD mice (Xu et al., 2020). Meanwhile, yeast β-glucans treatment was able to trigger the generation of SCFAs, which may be associated with increasing the abundance of Bacteroidetes, Lactobacillus and Bifidobacterium (Xu et al., 2020). Moreover, another study found that FMT could reverse the decreased Bacteroidetes in the Tg mice. Meanwhile, the level of butyrate was significantly increased in the FMT-treated mice, which may be associated with increasing the abundance of Bacteroidetes (Sun et al., 2019b). Therefore, we speculate that FMT may improve AD by increasing the inhibition of histone acetylation by SCFAs. Therefore, we speculate that FMT may improve AD by increasing the inhibition of histone acetylation by SCFAs.
5. Conclusion and future directions
Based on the above evidence, we conclude that FMT is beneficial for AD. AD has a serious impact on the health and quality of life of patients, and it is urgent to find low cost and few side effects intervention measures. However, there is relatively less research on the effects of FMT on AD, especially in clinical studies. So far, only three articles involved in FMT and AD, but no clinical trials have been published, among which one was terminated early due to the COVID-19 pandemic. Therefore, there is an urgent need for large-scale clinical trials to verify whether FMT can be used as a strategy for the treatment of AD. At the same time, seven gut microbiotas were defined as a good FMT, such as, Lachnospiraceae, Faecalibaculum, Ruminococcaceae, Bacteroidetes, Lactobacillus, Bifidobacterium, and Desulfovibrionaceae, which still needs to be further validation.
This mini-review summarized the evidence of FMT in the treatment of AD in recent years, which can bring significant benefits to the prevention and treatment of AD. FMT play a positive effect on AD through the anti-inflammatory effects, regulating deposition of Aβ, synaptic plasticity, SCFAs, and the histone acetylation. Our findings may provide evidence for fecal microbiota-related drug preparations as adjuvant therapy for the prevention and treatment of AD. In the future, the biological mechanism of improvement of AD of FMT still needs to be further clarified.
Author contributions
WX: Writing – original draft, Writing – review and editing. HX: Writing – original draft, Writing – review and editing. JW: Writing – original draft, Writing – review and editing. YJ: Writing – review and editing. CP: Writing – review and editing. BJ: Conceptualization, Writing – review and editing. AZ: Conceptualization, Writing – review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the General Program of National Natural Science Foundation of China (81973927), the Key R&D Program of Sichuan Department of Science and Technology (2021YFS0133), the Special Project for Scientific Research of Traditional Chinese Medicine in Sichuan Province (2020), the Special Project of Traditional Chinese Medicine Scientific Research of Sichuan Provincial Administration of Traditional Chinese Medicine (2020LC0224), the Hospital Project of the General Hospital of the Western Theater (2019), and the Chongqing Beibei District Science and Technology Bureau Project (2022-18).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abolhasani, E., Hachinski, V., Ghazaleh, N., Azarpazhooh, M. R., Mokhber, N., and Martin, J. (2023). Air pollution and incidence of dementia: A systematic review and meta-analysis. Neurology 100, e242–e254. doi: 10.1212/WNL.0000000000201419
Akbari, E., Asemi, Z., Kakhaki, R. D., Bahmani, F., Kouchaki, E., Tamtaji, O., et al. (2016). Effect of probiotic supplementation on cognitive function and metabolic status in Alzheimer’s Disease: A randomized, double-blind and controlled trial. Front. Aging Neurosci 8:256. doi: 10.3389/fnagi.2016.00256
Almeida, C. G., Tampellini, D., Takahashi, R. H., Greengard, P., Lin, M. T., Snyder, E. M., et al. (2005). Beta-amyloid accumulation in APP mutant neurons reduces PSD-95 and GluR1 in synapses. Neurobiol. Dis. 20, 187–198. doi: 10.1016/j.nbd.2005.02.008
Alzheimer’s Association (2023). 2023 Alzheimer’s disease facts and figures. Alzheimers Dement. 19, 1598–1695. doi: 10.1002/alz.13016
Angelucci, F., Cechova, K., Amlerova, J., and Hort, J. (2019). Antibiotics, gut microbiota, and Alzheimer’s disease. J. Neuroinflamm. 16:108. doi: 10.1186/s12974-019-1494-4
Backhed, F., Fraser, C. M., Ringel, Y., Sanders, M. E., Sartor, R. B., Sherman, P. M., et al. (2012). Defining a healthy human gut microbiome: Current concepts, future directions, and clinical applications. Cell. Host Microbe 12, 611–622. doi: 10.1016/j.chom.2012.10.012
Barichello, T., Generoso, J. S., Simoes, L. R., Faller, C. J., Ceretta, R. A., Petronilho, F., et al. (2015). Sodium butyrate prevents memory impairment by re-establishing BDNF and GDNF expression in experimental pneumococcal meningitis. Mol. Neurobiol. 52, 734–740. doi: 10.1007/s12035-014-8914-3
Bellenguez, C., Küçükali, F., Jansen, I. E., Kleineidam, L., Moreno-Grau, S., Amin, N., et al. (2022). New insights into the genetic etiology of Alzheimer’s disease and related dementias. Nat. Genet. 54, 412–436. doi: 10.1038/s41588-022-01024-z
Blackburn, L. M., Bales, A., Caldwell, M., Cordell, L., Hamilton, S., and Kreider, H. (2015). Fecal microbiota transplantation in patients with cancer undergoing treatment. Clin. J. Oncol. Nurs. 19, 111–114. doi: 10.1188/15.CJON.111-114
Bloom, G. S. (2014). Amyloid-β and tau: The trigger and bullet in Alzheimer disease pathogenesis. JAMA Neurol. 71, 505–508. doi: 10.1001/jamaneurol.2013.5847
Bodelier, P. L. (2011). Toward understanding, managing, and protecting microbial ecosystems. Front. Microbiol. 2:80. doi: 10.3389/fmicb.2011.0008
Cammarota, G., Ianiro, G., Tilg, H., Rajilić-Stojanović, M., Kump, P., Satokari, R., et al. (2017). European consensus conference on faecal microbiota transplantation in clinical practice. Gut 66, 569–580. doi: 10.1136/gutjnl-2016-313017
Cattaneo, A., Cattane, N., Galluzzi, S., Provasi, S., Lopizzo, N., Festari, C., et al. (2017). Association of brain amyloidosis with proinflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 49, 60–68. doi: 10.1016/j.neurobiolaging.2016.08.019
Cheng, Y., He, C. Y., Tian, D. Y., Chen, S. H., Ren, J. R., Sun, H. L., et al. (2023). Physiological β-amyloid clearance by the liver and its therapeutic potential for Alzheimer’s disease. Acta Neuropathol. 145, 717–731. doi: 10.1007/s00401-023-02559-z
Cryan, J. F., O’Riordan, K. J., Cowan, C. S. M., Sandhu, K. V., Bastiaanssen, T. F. S., Boehme, M., et al. (2019). The microbiota-gut-brain axis. Physiol. Rev. 99, 1877–2013. doi: 10.1152/physrev.00018.2018
D’Amato, A., Di Cesare Mannelli, L., Lucarini, E., Man, A. L., Le Gall, G., Branca, J. J. V., et al. (2020). Faecal microbiota transplant from aged donor mice affects spatial learning and memory via modulating hippocampal synaptic plasticity- and neurotransmission-related proteins in young recipients. Microbiome 8:140. doi: 10.1186/s40168-020-00914-w
Dodiya, H. B., Kuntz, T., Shaik, S. M., Baufeld, C., Leibowitz, J., Zhang, X., et al. (2019). Sex-specific effects of microbiome perturbations on cerebral Abeta amyloidosis and microglia phenotypes. J. Exp. Med. 216, 1542–1560. doi: 10.1084/jem.20182386
Duyckaerts, C., Delatour, B., and Potier, M. C. (2009). Classification and basic pathology of Alzheimer disease. Acta Neuropathol. 118, 5–36. doi: 10.1007/s00401-009-0532-1
Elangovan, S., Borody, T. J., and Holsinger, R. M. D. (2019). Fecal microbiota transplantation decreases amyloid load and improves cognition in Alzheimer’s. BioRxiv[Preprint]. doi: 10.1101/687376
Felsky, D., Roostaei, T., Nho, K., Risacher, S. L., Bradshaw, E. M., Petyuk, V., et al. (2019). Neuropathological correlates and genetic architecture of microglial activation in elderly human brain. Nat. Commun. 10:409. doi: 10.1038/s41467-018-08279-3
Fujii, Y., Nguyen, T. T. T., Fujimura, Y., Kameya, N., Nakamura, S., Arakawa, K., et al. (2019). Fecal metabolite of a gnotobiotic mouse transplanted with gut microbiota from a patient with Alzheimer’s disease. Biosci. Biotechnol. Biochem 83, 2144–2152.
Govindarajan, N., Agis-Balboa, R. C., Walter, J., and Sananbenesi, F. (2011). Fischer A Sodium butyrate improves memory function in an Alzheimer’s disease mouse model when administered at an advanced stage of disease progression. J. Alzheimers Dis. 26:187197. doi: 10.3233/JAD-2011-110080
Gupta, A., and Khanna, S. (2017). Fecal microbiota transplantation. JAMA 318:102. doi: 10.1001/jama.2017.6466
Hamilton, M. J., Weingarden, A. R., Sadowsky, M. J., and Khoruts, A. (2012). Standardized frozen preparation for transplantation of fecal microbiota for recurrent Clostridium difficile infection. Am. J. Gastroenterol. 107, 761–767. doi: 10.1038/ajg.2011.482
Hang, Z., Cai, S., Lei, T., Zhang, X., Xiao, Z., Wang, D., et al. (2022). Transfer of tumor-bearing mice intestinal flora can ameliorate cognition in Alzheimer’s Disease Mice. J. Alzheimers Dis. 86, 1287–1300. doi: 10.3233/JAD-215495
Harach, T., Marungruang, N., Duthilleul, N., Cheatham, V., Mc Coy, K. D., Frisoni, G., et al. (2017). Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 7:41802. doi: 10.1038/srep41802
Hazan, S. (2020). Rapid improvement in Alzheimer’s disease symptoms following fecal microbiota transplantation: A case report. J. Int. Med. Res. 48:0300060520925930. doi: 10.1177/030006052092593
Hryniewicz, A., Bialuk, I., Kamiński, K. A., and Winnicka, M. M. (2007). Impairment of recognition memory in interleukin-6 knock-out mice. Eur. J. Pharmacol. 577, 219–220.
Huang, W., Hu, W., Cai, L., Zeng, G., Fang, W., Dai, X., et al. (2021). Acetate supplementation produces antidepressant-like effect via enhanced histone acetylation. J. Affect. Disord. 281, 51–60. doi: 10.1016/j.jad.2020.11.121
Jeong, S. M., Park, J., Han, K., Yoo, J., Yoo, J. E., Lee, C. M., et al. (2023). Association of changes in smoking intensity with risk of dementia in Korea. JAMA Netw. Open 6:e2251506. doi: 10.1001/jamanetworkopen.2022.51506
Jin, J., Xu, Z., Zhang, L., Zhang, C., Zhao, X., Mao, Y., et al. (2023). Gut-derived β-amyloid: Likely a centerpiece of the gut-brain axis contributing to Alzheimer’s pathogenesis. Gut Microbes 15:2167172. doi: 10.1080/19490976.2023.2167172
Johnson, D., Thurairajasingam, S., Letchumanan, V., Chan, K. G., and Lee, L. H. (2021). Exploring the role and potential of probiotics in the field of mental health: Major depressive disorder. Nutrients 13:1728. doi: 10.3390/nu13051728
Jouanne, M., Rault, S., and Voisin-Chiret, A. S. (2017). Tau protein aggregation in Alzheimer’s disease: An attractive target for the development of novel therapeutic agents. Eur. J. Med. Chem. 139, 153–167. doi: 10.1016/j.ejmech.2017.07.070
Kao, D., Roach, B., Silva, M., Beck, P., Rioux, K., Kaplan, G. G., et al. (2017). Effect of oral capsule- vs colonoscopy-delivered fecal microbiota transplantation on recurrent Clostridium difficile infection: A randomized clinical trial. JAMA 318, 1985–1993. doi: 10.1001/jama.2017.17077
Kassam, Z., Lee, C. H., Yuan, Y., and Hunt, R. H. (2013). Fecal microbiota transplantation for Clostridium difficile infection: Systematic review and meta-analysis. Am. J. Gastroenterol. 108, 500–508. doi: 10.1038/ajg.2013.59
Kim, M. S., Kim, Y., Choi, H., Kim, W., Park, S., Lee, D., et al. (2020). Transfer of a healthy microbiota reduces amyloid and tau pathology in an Alzheimer’s disease animal model. Gut 69, 283–294. doi: 10.1136/gutjnl-2018-317431
Kim, N., Jeon, S. H., Ju, I. G., Gee, M. S., Do, J., Oh, M. S., et al. (2021). Transplantation of gut microbiota derived from Alzheimer’s disease mouse model impairs memory function and neurogenesis in C57BL/6 mice. Brain Behav. Immun. 98, 357–365. doi: 10.1016/j.bbi.2021.09.002
Knopman, D. S., Amieva, H., Petersen, R. C., Chételat, G., Holtzman, D. M., Hyman, B. T., et al. (2021). Alzheimer disease. Nat. Rev. Dis. Primers 7:33. doi: 10.1038/s41572-021-00269-y
Kong, Y., Wang, L., and Jiang, B. (2021). The role of gut microbiota in aging and aging related neurodegenerative disorders: Insights from drosophila model. Life 11:855. doi: 10.3390/life11080855
Lansbury, P. T., and Lashuel, H. A. (2006). A century-old debate on protein aggregation and neurodegeneration enters the clinic. Nature 443, 774–779. doi: 10.1038/nature05290
Li, Y., Ning, L., Yin, Y., Wang, R., Zhang, Z., Hao, L., et al. (2020). Age-related shifts in gut microbiota contribute to cognitive decline in aged rats. Aging 12, 7801–7817. doi: 10.18632/aging.103093
Li, Y., Sang, S., Ren, W., Pei, Y., Bian, Y., Chen, Y., et al. (2021). Inhibition of Histone Deacetylase 6 (HDAC6) as a therapeutic strategy for Alzheimer’s disease: A review (2010-2020). Eur. J. Med. Chem. 226:113874. doi: 10.1016/j.ejmech.2021.113874
Liu, P., Wu, L., Peng, G., Han, Y., Tang, R., Ge, J., et al. (2019). Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain Behav. Immun. 80, 633–643. doi: 10.1016/j.bbi.2019.05.008
Liu, Q., Xi, Y., Wang, Q., Liu, J., Li, P., Meng, X., et al. (2021a). Mannan oligosaccharide attenuates cognitive and behavioral disorders in the 5xFAD Alzheimer’s disease mouse model via regulating the gut microbiota-brain axis. Brain Behav. Immun. 95, 330–343. doi: 10.1016/j.bbi.2021.04.005
Liu, Q., Xie, T., Xi, Y., Li, L., Mo, F., Liu, X., et al. (2021b). Sesamol attenuates amyloid peptide accumulation and cognitive deficits in APP/PS1 Mice: The mediating role of the gut-brain axis. J. Agric. Food. Chem. 69, 12717–12729. doi: 10.1021/acs.jafc.1c04687
Lloyd-Price, J., Abu-Ali, G., and Huttenhower, C. (2016). The healthy human microbiome. Genome. Med. 8:51. doi: 10.1186/s13073-016-0307-y
Loy, C. T., Schofield, P. R., Turner, A. M., and Kwok, J. B. (2014). Genetics of dementia. Lancet 383, 828–840. doi: 10.1016/S0140-6736(13)60630-3
Manly, J. J., Jones, R. N., Langa, K. M., Ryan, L. H., Levine, D. A., McCammon, R., et al. (2022). Estimating the prevalence of dementia and mild cognitive impairment in the US: The 2016 health and retirement study harmonized cognitive assessment protocol project. JAMA Neurol. 79, 1242–1249. doi: 10.1001/jamaneurol.2022.3543
Mayer, E. A., Nance, K., and Chen, S. (2022). The gut-brain axis. Annu. Rev. Med. 73, 439–453. doi: 10.1146/annurev-med-042320-014032
Mocanu, V., Zhang, Z., Deehan, E. C., Kao, D. H., Hotte, N., Karmali, S., et al. (2021). Fecal microbial transplantation and fiber supplementation in patients with severe obesity and metabolic syndrome: A randomized double-blind, placebo-controlled phase 2 trial. Nat. Med. 27, 1272–1279. doi: 10.1038/s41591-021-01399-2
Morais, L. H., Schreiber, H. L. IV, and Mazmanian, S. K. (2021). The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 19, 241–255. doi: 10.1038/s41579-020-00460-0
Pang, Y., Tien, L. T., Zhu, H., Shen, J., Wright, C. F., Jones, T. K., et al. (2015). Interleukin-1 receptor antagonist reduces neonatal lipopolysaccharide-induced long-lasting neurobehavioral deficits and dopaminergic neuronal injury in adult rats. Int. J. Mol. Sci. 16, 8635–8654.
Paramsothy, S., Kamm, M. A., Kaakoush, N. O., Walsh, A. J., van den Bogaerde, J., Samuel, D., et al. (2017). Multidonor intensive faecal microbiota transplantation for active ulcerative colitis: A randomised placebo-controlled trial. Lancet 389, 1218–1228. doi: 10.1016/S0140-6736(17)30182-4
Park, S. H., Lee, J. H., Shin, J., Kim, J. S., Cha, B., Lee, S., et al. (2021). Cognitive function improvement after fecal microbiota transplantation in Alzheimer’s dementia patient: A case report. Curr. Med. Res. Opin. 37, 1739–1744. doi: 10.1080/03007995.2021.1957807
Rajan, K. B., Weuve, J., Barnes, L. L., McAninch, E. A., Wilson, R. S., and Evans, D. A. (2021). Population estimate of people with clinical Alzheimer’s disease and mild cognitive impairment in the United States (2020-2060). Alzheimers Dement. 17, 1966–1975. doi: 10.1002/alz.12362
Ridaura, V., and Belkaid, Y. (2015). Gut microbiota: The link to your second brain. Cell 161, 193–194. doi: 10.1016/j.cell.2015.03.033
Samieri, C., Perier, M. C., Gaye, B., Proust-Lima, C., Helmer, C., Dartigues, J. F., et al. (2018). Association of cardiovascular health level in older age with cognitive decline and incident dementia. JAMA 320, 657–664. doi: 10.1001/jama.2018.11499
Schneider, A. L. C., Selvin, E., Latour, L., Turtzo, L. C., Coresh, J., Mosley, T., et al. (2021). Head injury and 25-year risk of dementia. Alzheimers Dement. 17, 1432–1441. doi: 10.1002/alz.12315
Sehar, U., Rawat, P., Reddy, A. P., Kopel, J., and Reddy, P. H. (2022). Amyloid beta in aging and Alzheimer’s Disease. Int. J. Mol. Sci. 23:12924. doi: 10.3390/ijms232112924
Shafquat, A., Joice, R., Simmons, S. L., and Huttenhower, C. (2014). Functional and phylogenetic assembly of microbial communities in the human microbiome. Trends Microbiol. 22, 261–266. doi: 10.1016/j.tim.2014.01.011
Shen, H., Guan, Q., Zhang, X., Yuan, C., Tan, Z., Zhai, L., et al. (2020). New mechanism of neuroinflammation in Alzheimer’s disease: The activation of NLRP3 inflammasome mediated by gut microbiota. Prog. Neuropsychopharmacol. Biol. Psychiatry 100:109884. doi: 10.1016/j.pnpbp.2020.109884
Shen, L., Liu, L., and Ji, H. F. (2017). Alzheimer’s disease histological and behavioral manifestations in transgenic mice correlate with specific gut microbiome state. J. Alzheimers Dis. 56, 385–390. doi: 10.3233/JAD-160884
Shi, L., and Tu, B. P. (2015). Acetyl-CoA and the regulation of metabolism: Mechanisms and consequences. Curr. Opin. Cell Biol. 33, 125–131. doi: 10.1016/j.ceb.2015.02.003
Smits, L. P., Bouter, K. E., de Vos, W. M., Borody, T. J., and Nieuwdorp, M. (2013). Therapeutic potential of fecal microbiota transplantation. Gastroenterology 145, 946–953. doi: 10.1053/j.gastro.2013.08.058
Sun, J., Liu, S., Ling, Z., Wang, F., Ling, Y., Gong, T., et al. (2019a). Fructooligosaccharides ameliorating cognitive deficits and neurodegeneration in APP/PS1 transgenic mice through modulating gut microbiota. J. Agric. Food. Chem. 67, 3006–3017. doi: 10.1021/acs.jafc.8b07313
Sun, J., Xu, J., Ling, Y., Wang, F., Gong, T., Yang, C., et al. (2019b). Fecal microbiota transplantation alleviated Alzheimer’s disease-like pathogenesis in APP/PS1 transgenic mice. Transl. Psychiatry. 9:189. doi: 10.1038/s41398-019-0525-3
Tan, J., McKenzie, C., Potamitis, M., Thorburn, A. N., Mackay, C. R., and Macia, L. (2014). The role of short-chain fatty acids in health and disease. Adv. Immunol. 121, 91–119. doi: 10.1016/B978-0-12-800100-4.00003-9
Tobinick, E., Gross, H., Weinberger, A., and Cohen, H. (2006). TNF-alpha modulation for treatment of Alzheimer’s disease: A 6-month pilot study. Med. Gen. Med. 8:25.
Valdes, A. M., Walter, J., Segal, E., and Spector, T. D. (2018). Role of the gut microbiota in nutrition and health. BMJ 361:k2179. doi: 10.1136/bmj.k2179
Vogt, N. M., Kerby, R. L., Dill-McFarland, K. A., Harding, S. J., Merluzzi, A. P., Johnson, S. C., et al. (2017). Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 7:13537. doi: 10.1038/s41598-017-13601-y
Wang, H., Yang, F., Xin, R., Cui, D., He, J., Zhang, S., et al. (2020). The gut microbiota attenuate neuroinflammation in manganese exposure by inhibiting cerebral NLRP3 inflammasome. Biomed. Pharmacother. 129:110449.
Wang, J. W., Kuo, C. H., Kuo, F. C., Wang, Y. K., Hsu, W. H., Yu, F. J., et al. (2019). Fecal microbiota transplantation: Review and update. J. Formos. Med. Assoc. 118(Suppl. 1), S23–S31. doi: 10.1016/j.jfma.2018.08.011
Wang, M., Cao, J., Gong, C., Amakye, W. K., Yao, M., and Ren, J. (2021). Exploring the microbiota-Alzheimer’s disease linkage using short-term antibiotic treatment followed by fecal microbiota transplantation. Brain Behav. Immun. 96, 227–238. doi: 10.1016/j.bbi.2021.06.00
World Alzheimer Report (2022). Life after Diagnosis: Navigating Treatment, Care and Support. London: Alzheimer’s Disease International.
Wu, L., Han, Y., Zheng, Z., Peng, G., Liu, P., Yue, S., et al. (2021). Altered gut microbial metabolites in amnestic mild cognitive impairment and Alzheimer’s disease: signals in host-microbe interplay. Nutrients 13:228. doi: 10.3390/nu13010228
Xu, M., Mo, X., Huang, H., Chen, X., Liu, H., Peng, Z., et al. (2020). Yeast β-glucan alleviates cognitive deficit by regulating gut microbiota and metabolites in Aβ1-42-induced AD-like Mice. Int. J. Biol. Macromol. 161, 258–270.
Xu, M. Q., Cao, H. L., Wang, W. Q., Wang, S., Cao, X. C., Yan, F., et al. (2015). Fecal microbiota transplantation broadening its application beyond intestinal disorders. World J. Gastroenterol. 21, 102–111. doi: 10.3748/wjg.v21.i1.102
Yang, L. (2018). Effects of Fecal Microbiota Transplantation on Alzheimer’S Disease Mice and its Molecular Mechanisms. Zhengzhou: Zhengzhou University.
Yang, L., Hao, J. R., Gao, Y., Yang, X., Shen, X. R., Wang, H. Y., et al. (2022). HDAC3 of dorsal hippocampus induces postoperative cognitive dysfunction in aged mice. Behav. Brain Res. 433:114002. doi: 10.1016/j.bbr.2022.114002
Yang, R., Chen, Z., and Cai, J. (2023). Fecal microbiota transplantation: Emerging applications in autoimmune diseases. J. Autoimmun. [Online ahead of print]. doi: 10.1016/j.jaut.2023.103038
Youngster, I., Sauk, J., Pindar, C., Wilson, R. G., Kaplan, J. L., Smith, M. B., et al. (2014). Fecal microbiota transplant for relapsing Clostridium difficile infection using a frozen inoculum from unrelated donors: A randomized, open-label, controlled pilot study. Clin. Infect. Dis. 58, 1515–1522. doi: 10.1093/cid/ciu135
Zhang, B., Chen, T., Cao, M., Yuan, C., Reiter, R. J., Zhao, Z., et al. (2022). Gut microbiota dysbiosis induced by decreasing endogenous melatonin mediates the pathogenesis of Alzheimer’s Disease and obesity. Front. Immunol. 13:900132. doi: 10.3389/fimmu.2022.900132
Zhang, F., Luo, W., Shi, Y., Fan, Z., and Ji, G. (2012). Should we standardize the 1,700-year-old fecal microbiota transplantation? Am. J. Gastroenterol. 107:1755. doi: 10.1038/ajg.2012.251
Zhang, L., Wang, Y., Xiayu, X., Shi, C., Chen, W., Song, N., et al. (2017). Altered gut microbiota in a mouse model of Alzheimer’s Disease. J. Alzheimers Dis. 60, 1241–1257. doi: 10.3233/JAD-170020
Zheng, J., Zheng, S. J., Cai, W. J., Yu, L., Yuan, B. F., and Feng, Y. Q. (2019). Stable isotope labeling combined with liquid chromatography-tandem mass spectrometry for comprehensive analysis of short-chain fatty acids. Anal. Chim. Acta 1070, 51–59. doi: 10.1016/j.aca.2019.04.021
Keywords: fecal microbiota transplantation, gut microbiota, Alzheimer’s disease, review, mechanism
Citation: Xiang W, Xiang H, Wang J, Jiang Y, Pan C, Ji B and Zhang A (2023) Fecal microbiota transplantation: a novel strategy for treating Alzheimer’s disease. Front. Microbiol. 14:1281233. doi: 10.3389/fmicb.2023.1281233
Received: 22 August 2023; Accepted: 31 October 2023;
Published: 16 November 2023.
Edited by:
Nazarii Kobyliak, Bogomolets National Medical University, UkraineReviewed by:
Ahmed Gamal, Case Western Reserve University, United StatesYingli Jing, Capital Medical University, China
Copyright © 2023 Xiang, Xiang, Wang, Jiang, Pan, Ji and Zhang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bingjin Ji, NTcwNjc2MDc4QHFxLmNvbQ==; Anren Zhang, YW4wMTI0QDE2My5jb20=
†These authors have contributed equally to this work