 Jose Ignacio Marín-Guirao
Jose Ignacio Marín-Guirao Miguel de Cara-García
Miguel de Cara-García- Andalusian Institute of Agricultural and Fisheries Research Training (IFAPA) La Mojonera, Almería, Spain
This study, conducted over two consecutive seasons in an organic-certified Mediterranean tomato greenhouse, aimed to assess the dynamics of soil bacterial composition at pre-planting phase following the incorporation of fresh sheep manure and subsequent solarisation (biosolarisation), as well as at the end of the crop cycle. Gene sequencing using 16S rRNA, and plate culture methods were applied. Additionally, dual culture tests were performed to evaluate the antagonistic activity of 95 soil-isolated bacteria against five soilborne pathogenic fungi and five beneficial fungi sourced from commercial products. Culturable and thermophilic bacterial populations shifted over time, but showed no clear trend. However, alpha diversity was lower at pre-planting phase and increased by the end of the cropping seasons. Significant shifts in beta diversity were also observed across sampling times. Firmicutes showed higher relative abundance at pre-planting phase, while Proteobacteria were consistently more abundant at the end of the cropping season. The genera Bacillus and Thauera were identified as biomarkers, with Bacillus associated with the pre-planting phase and Thauera with the end of the cropping seasons (LDA score > 4.5). Among the 52 ecological pathways detected via FAPROTAX database, nitrogen-related functions showed higher presence at the end of the cropping seasons. Isolates with antagonistic activity were detected at all sampling times. The 37.9% of the isolates showed antagonism, 31.6% against phytopathogenic fungi, 20.0% against beneficial fungi, and 12.6% against both. Colletotrichum gloeosporioides and Botrytis cinerea were the pathogens to which the highest number of isolates showed antagonism, while Trichoderma asperellum and T. atroviride were among the beneficial ones. Among the native antagonistic bacteria, Streptomyces spp. was the dominant genus followed by Bacillus. This information highlights how the functional diversity of native bacteria communities from biosolarised soils, may impact the performance of introduced biocontrol microorganisms, as well as the development of natural soil suppressiveness.
1 Introduction
The current trend on intensification in organic and conventional horticultural production systems, underscores the need for effective crop protection and production methods that ensure environmental sustainability (Lal, 2020). In this regard, disease-suppressive soils (DSS) represent a significant achievement toward sustainability. It refers to soils where disease progression remains minimal, even when soil-borne pathogens and susceptible host plants are present (Baker and Cook, 1974; Schneider, 1982), and is mainly influenced by the intrincate interactions among soil, plant, microbiome and environmental factors (Gómez Expósito et al., 2017; Jayaraman et al., 2021; Sagova-Mareckova et al., 2023). Although the abiotic factors of the soil are believed to play a role, soil biodiversity strongly influences the phenomena through diverse biological mechanisms which are summarized as (Hoitink and Boehm, 1999; Mazzola, 2002; Garbeva et al., 2006; Adesina et al., 2007; Jayaraman et al., 2021): (i) parasitism against pathogens, (ii) production of metabolites, toxins and antibiotics, (iii) competition for nutrients, resources and/or substrates, (iv) activation of disease-resistant genes in the host plant, and (v) improvement of plant nutrition and soil health. In this way, soil microbial community plays a crucial role in the DSS since it is involved in the above mentioned mechanisms (Alabouvette et al., 1993; Mazzola, 2002; Weller et al., 2002; Garbeva et al., 2004; Mazzola and Freilich, 2016; Siegel-Hertz et al., 2018).
Among the native soil microbiota, soil bacteria, including thermophilic bacteria, have been reported as main actors in the suppressiveness of soil-borne diseases in different agricultural soils (Mendes et al., 2011; Gómez Expósito et al., 2017; Schlatter et al., 2017). The composition of the bacterial community is one of the major factors in the mechanism of disease suppression (Hemkemeyer et al., 2024; Kang et al., 2024; Ni et al., 2022), and several bacterial genera including Bacillus, Pseudomonas and Streptomyces, among others, have been proposed as key players in disease suppressiveness of soils (Hoitink et al., 1997; Chng et al., 2015; Xiong et al., 2015; Eberlein et al., 2016). These microorganisms are actively involved in the transformation and decomposition of organic matter (Pankhurst and Lynch, 1995), and the incorporation of organic amendments (OA) can positively impact their communities, improving soil health and enhancing soil disease suppressiveness (Bonanomi et al., 2007, 2010, 2018; Postma and Schilder, 2015; Akanmu et al., 2021), as well as allowing to achieve greenhouse sustainability (Giagnocavo et al., 2022). In addition, OA incorporation impacts on physical and chemical factors that also has an influence on the mechanisms of soil suppressiveness (Amir and Alabouvette, 1993), as well as in greenhouse soil fertility (García-Raya et al., 2019; Marín-Guirao et al., 2019a, 2023a; Castillo-Díaz et al., 2021, 2022). In this regard, the incorporation of fresh organic matter followed by solarisation (i.e., biosolarisation) is an increasing practice in Mediterranean greenhouses, reaching 4.6% of producers in south-eastern Spain during campaign 2019/2020 (García et al., 2024). Biosolarisation is mainly practiced as a soil disinfection technique, but also to promote the decomposition of organic material, what helps to prevent potential phytotoxicity issues that can occur when seedlings are transplanted with fresh, insuficciently decomposed material.
Understanding the impact of soil biosolarisation on the native bacterial community is crucial, as it directly influences community structure and functionality. It is essential to appraises the extent of this impact, and to determine whether bacteria with suppressive properties against phytopathogenic fungi may emerge, following the implementation of such practices. Additionally, evaluating the persistence of these bacteria at the end of consecutive crop cycles is essential, as it is a highly relevant aspect in maintaining crop health (Mendes et al., 2011; Gómez Expósito et al., 2017). Furthermore, given the rising utilization of commercial products containing biological control organisms (BCO) and other beneficial microorganisms for crops, it is highly pertinent to study whether native soil bacteria might also exhibit a suppressive effect on these microorganisms. This currently limited understanding holds substantial significance and is crucial for the efficacy of such products when applied into the soil.
The present study was conducted for two consecutive years in an organic-certified greenhouse with tomato crops. The study aimed to assess the dynamics of soil bacterial composition at pre-planting phase following the incorporation of fresh sheep manure in summer and subsequent solarization (biosolarisation), as well at the finalization of the crop cycle, using 16S rRNA gene sequencing and plate culture methods on soil microbial composition. Additionally, dual culture tests were performed to evaluate the antagonistic activity of 95 soil-isolated bacteria collected at the start and end of the tomato crops against five soilborne pathogenic fungi and five beneficial fungi sourced from commercial products.
2 Materials and methods
2.1 Experimental greenhouse and soil management
The present study was conducted for two consecutive years (the 2019/2020 and 2020/2021 seasons) at the Andalusian Institute for Research and Training in Agriculture and Fisheries (IFAPA) in Almería, Spain (36°48′N, 2°41′W; altitude 142 m), the largest cropping region for Mediterranean greenhouses. The climate in the area is Mediterranean semi-arid, characterized by mild winters and hot summers with no rainfall. The experimental greenhouse, certified for organic production by the Andalusian Organic Farming Committee (C. A. A. E.) since 2006, covered 832 m2 and was representative of the “raspa y amagado” Mediterranean greenhouse. Two subsequent winter cycles of “Valenciano type” tomato plants (Solanum lycopersicum L.), grafted onto Armstrong® rootstock (Syngenta, Basel, Switzerland), were cultivated during 2019/20 and 2020/21 seasons, referred to as season 1 and season 2, respectively. Four-week-old tomato plants were planted on September 17th, 2019, and September 25th, 2020, in seasons 1 and 2, respectively. Prior to planting in both years, fresh sheep manure was uniformly incorporated into the greenhouse soil at a rate of 4 kg m−2 in July. Incorporation involved mixing the manure with the soil using a rotavator. Next, drip lines were deplyed and, with the aim of favoring manure’s decomposition, the soil was covered with transparent polyethylene film (30 μm TIF Desinfección DS®, Sotrafa, Almería, Spain) for a period of 2 months, after a single irrigation application to reach saturation at 15 cm depth. The maximum, minimum, and average temperatures at 20 cm soil depth during biosolarisation in 2020 were 44.8 ± 0.9, 31.8 ± 0.6, and 39.3 ± 0.3, respectively. Planting occurred 2 days after removing the film. Crop management and pest control practices were adhered to Regulation (EU) 2018/848 on organic production. At the start of the study, the soil consisted of 19% clay, 13% silt, and 68% sand. Soil pH was 8.57, electrical conductivity (EC) 6.19 dS m−1, organic matter content 1.20%, total carbonates (HCO3−) 14.33%, active limestone 5.67%, nitric-nitrogen (N) 157 mg L−1, total nitrogen 0.04%, phosphorus Olsen (P Olsen) 19 meq L−1, interchangeable calcium (Ca+2) 1,253 mg L−1, interchangeable sodium (Na+) 37 mg L−1, interchangeable magnesium (Mg+2) 261 mg L−1, interchangeable potassium (K+) 1,841 mg L−1, and C/N ratio 20. The soil had no history of soilborne pathogens.
2.2 Soil collection
Soil samples were taken at four different sampling times. During the 2-year study period, samples were collected 2 days before planting (i.e., Start Season 1, 15th September 2019, and Start Season 2, 23rd September 2020) and at the conclusion of the tomato crop season (i.e., End Season 1, 3rd April 2020, and End Season 2, 23rd April 2021). At Start Season 1, soil sampling was conducted in three designated areas of the greenhouse immediately preceding the planting of tomato seedlings. In subsequent sampling times, systematic soil samples were taken in 12 uniformly distributed plots within the greenhouse. The samples were taken with an auger at a depth of 0–30 cm. Three subsamples were randomly taken from cultivation lines and then mixed and homogenized to ensure representativeness in each sample. For each soil sample, one subsample was air-dried and sieved (<0.2 mm) for microbiological analysis through the plate culture method, and another subsample was frozen at −80°C for DNA extraction used in the assessment of bacterial microbiota through DNA metabarcoding.
2.3 Quantification and isolation of soil culturable bacteria and soil thermophilic bacteria
The preparation of soil samples included a drying, grinding, and sieving process, following the method described by Tello et al. (1991). Drying was conducted at room temperature (20–25°C) for a period of 7–10 days, until soil humidity was constant and homogenous. A porcelain mortar was used for grinding, and a 200 μm mesh-size sieve was used for sieving. Both the mortar and sieve were thoroughly washed and disinfected between samples by covering them with alcohol and lighting it. Then, the soil culturable bacteria and thermophilic bacteria were analyzed by means of the serial dilution method (Tello et al., 1991). The culture medium used for bacteria evaluation was nutrient agar (NA, Oxoid, Basingtoke, U. K.). In the case of thermophilic bacteria, soil samples were previously subjected to a heat treatment in an oven at 70°C for 1 h, using a semi-selective medium for actinobacteria (Crawford et al., 1993). This technique was applied to isolate the live thermophilic culturable fraction and to allow the study of its antagonistic activity (see Section 2.5.). In both cases, five subreplicates (9 cm-diameter Petri dishes) of each soil sample were analised at 10−5 and 10−6 dilutions. The Petri dishes were incubated at 25°C for 7 days. Subsequently, total colony forming units (CFU) of bacteria and thermophilic bacteria were quantified, eventually expressing the results as CFU/g dry soil.
2.4 Assessment of bacterial microbiota through DNA metabarcoding
2.4.1 Soil DNA extraction, amplicon high-throughput sequencing, and processing of sequencing data
DNA from soil samples was isolated using the DNeasy PowerSoil Pro DNA isolation kit (Qiagen, Hilden, Germany), strictly following the manufacturer’s instructions. An extraction blank for cross-contamination was included. DNA was quantified using the Qubit High Sensitivity dsDNA Assay (Thermo Fisher Scientific, Waltham, MA, United States). For bacterial library preparation, a fragment of the 16S genomic region (of about 460 bp) was amplified using the primers Bakt_341F (5′ CCT ACG GGN GGC WGC AG 3′) and Bakt_805R (5′ GAC TAC HVG GGT ATC TAA TCC 3′) (Herlemann et al., 2011). Illumina sequencing primers were attached to these primers at their 5′ ends. Polymerase chain reactions (PCRs) were carried out in a final volume of 12.5 μL, containing 1.25 μL of template DNA, 0.5 μM of the primers, 3.25 μL of Supreme NZYTaq 2x Green Master Mix (NZYTech, Lisbon, Portugal), and ultrapure water up to 12.5 μL. The reaction mixture was incubated as follows: an initial denaturation step at 95°C for 5 min, followed by 25 cycles of 95°C for 30 s, 50°C for 45 s, 72°C for 45 s, and a final extension step at 72°C for 7 min. A negative control that contained no DNA (BPCR) was included in every PCR round to check for contamination during library preparation. The libraries were run on a 2% agarose gel stained with GreenSafe (NZYTech) and imaged under UV light to verify the library size. Libraries were purified using the Mag-Bind RXNPure Plus magnetic beads (Omega Biotek, Beijing, China), following the instructions provided by the manufacturer. Then, libraries were pooled in equimolar amounts according to the quantification data provided by the Qubit dsDNA HS Assay (Thermo Fisher Scientific). This pool also contained a testimonial amount (1 μL) of the PCR blank and DNA extraction blank (Bex). The pool was sequenced in the MiSeq PE300 run (Illumina). DNA metabarcoding analyses were carried out by AllGenetics & Biology S. L. (La Coruña, Spain; www.allgenetics.eu). Illumina paired-end raw forward (R1) and reverse (R2) FASTQ reads were stored in separate files. The quality of the FASTQ files was checked with the software (v0.11.9) FastQC (Andrews, 2010) and the output was summarized using MultiQC (Ewels et al., 2016). The obtained amplicon reads were processed using QIIME 2 (release 2022.2) (Bolyen et al., 2019). Specifically, the tool DADA2 (Callahan et al., 2016) (implemented in QIIME 2) was used to remove the PCR primers, filter low-quality reads, denoise, merge the forward and reverse reads, remove the chimeric reads, and cluster the resulting sequences into amplicon sequence variants (ASVs). The resulting output of the DADA2 pipeline was a table containing the number of occurrences of every observed ASV in each sample. The taxonomic assignment of ASVs was conducted using a pre-trained classifier of the SILVA reference database (Quast et al., 2013) (updated in August 2020). Specifically, the feature-classifier classify-sklearn approach implemented in QIIME 2 (Bokulich et al., 2018) was employed. The following ASVs were excluded: singletons (i.e., ASVs containing only one member sequence in the whole dataset), ASVs occurring at a frequency below 0.01%, non-bacterial ASVs such as eukaryotic sequences of chloroplast (Moore et al., 2019) and mitochondrial (Emelyanov, 2001) origin, unidentified sequences, and sequences assigned only at the kingdom level. The final filtered ASV table was converted into a Biological Observation Matrix (biom) file that was directly imported into R 3.6.1 (R-Team-Core, 2019) using the package phyloseq 1.24.2 (McMurdie and Holmes, 2013). The final filtered ASV and the biom file were used for further analysis and plotting.
2.4.2 Bacterial community description and data processing
Four indexes of biodiversity were selected to study the alpha diversity of the bacterial community of each sample: Margalef genus richness (d) (Margalef, 1958); the Shannon–Wiener diversity index (H′) log basis (Shannon and Weaver, 1949); Pielou’s evenness (J) (Pielou, 1969) of the distribution of individuals among genera; and Simpson’s dominance (D) (Simpson, 1949). All calculations were carried out with the software PRIMER version 6.0 (Primer-E Ltd., Plymouth, U. K.) for Windows (Clarke and Gorley, 2006). Regarding beta diversity, bacterial community data at the operational taxonomic unit (OTU) level through DNA metabarcoding evaluation method, were compared among sampling times using permutational analysis of variance (PERMANOVA). Also, pairwise PERMANOVA comparisons were performed. These tests calculate Pseudo-F statistics to obtain p-values (Anderson, 2001). All PERMANOVA tests used 9,999 permutations from unrestricted permutations of raw data. Previously, data were square root transformed. The calculation of similarity matrices in the PERMANOVA analysis was based on Bray–Curtis distance. In addition, principal coordinate analysis (PCoA) was used to visualize the potential differences in the soil bacterial community structure among sampling times or within sampling times. PERMANOVA and PCoA analyses were performed using the multivariate statistical software package PRIMER-6 with the PERMANOVA+ extension (Primer-E Ltd., Plymouth, UK) for Windows. The linear discriminant analysis effect size (LEfSe) algorithm (Segata et al., 2011) was generated from Python (version 3.12) and used to identify and compare unique bacterial taxa significantly related to any of the four sampling times (Start Season 1, End Season 1, Start Season 2, End Season 2). In addition, data from Start Season 1 and Start Season 2, as well as those from End Season 1 and End Season 2, were pooled in order to identify and compare unique bacterial taxa significantly related to Start Season and End Season, respectively. The threshold for the logarithmic linear discriminant analysis (LDA) score was established at 4.5 and the Wilcoxon p-value was set at 0.05.
Bacterial functional potential was investigated using Functional Annotation of Prokaryotic Taxa (FAPROTAX) database (Louca et al., 2016). FAPROTAX is a database that categorizes bacterial taxa into recognized metabolic or ecologically significant groups (considered as functional groups), utilizing current literature on cultured strains as a reference. Following the procedure of Vido et al. (2024), previously bacterial ASVs was filtered to reduce noise (i.e., removal of those with <10 counts). In addition, filtering was completed by removing functional groups with reads abundances ≤100. The abundances of each taxa assigned to an functional group were pooled to form a total relative abundance count for each group, and the abundances of unassigned taxa were excluded. Then, the diversity and composition of ecological groups within each sampling time were compared and contrasted using the same method outlined for taxonomy beta diversity. Thus, PERMANOVA, pairwise PERMANOVA and PCoA were conducted on the top 15 ecological groups to focus on the most relevant ecological pathways. In addition, LEfSe was performed to detect differentially abundant ecological pathways across groups of soil samples depending on sampling time (LDA score established at 3).
2.5 In vitro antagonism toward phytopathogenic and beneficial fungi
2.5.1 Strains isolation and target fungi
The isolates used in the antagonism assays were obtained from the thermophilic bacteria analyses described in section 2.3. Following the evaluation specified in that section, colonies from soil samples taken at different sampling times were randomly selected and isolated on Petri dishes with 15 mL of NA medium. Eleven isolates from Start Season 1, 51 from End Season 1, and 33 from Start Season 2 were selected. Thus, the antagonistic activity of 95 soil-isolated bacteria against nine fungi and one oomycete (Phytophthora capsici), referred to as fungi throughout the rest of the document, was studied. Specifically, the evaluations were conducted against five soilborne pathogenic fungi, as well as five beneficial fungi sourced from commercial products (Table 1).
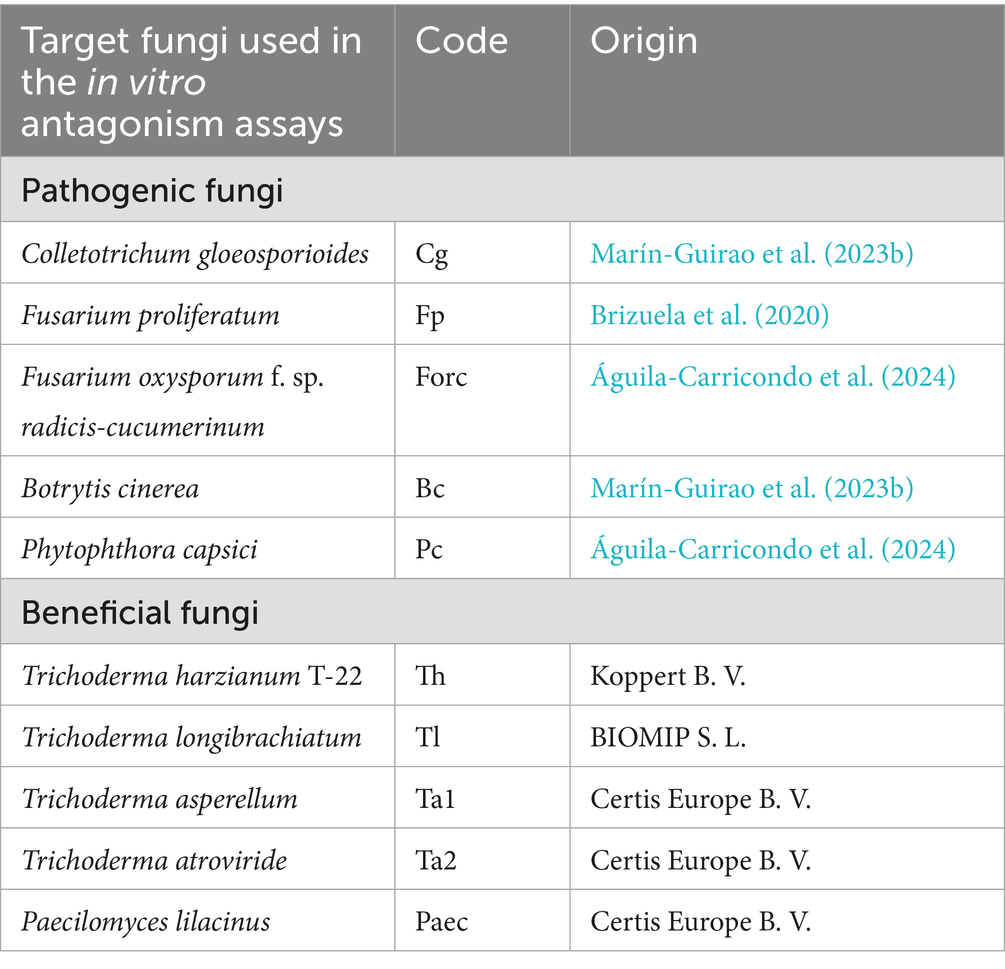
Table 1. Phytopathogenic and beneficial fungi used in dual culture tests and origin of the strains.
2.5.2 Dual culture protocol
Dual culture tests on Potato Glucose Agar (PGA, Oxoid, Basingtoke, U. K.) were performed to evaluate the antagonistic activity of soil-isolated bacteria against the ten target fungi (Table 1). Plates (9 cm diameter), which contained 15 mL of PGA, were inoculated with strain culture by streak and incubated at 25 ± 1°C in the dark for 8 days. Later, a 5-mm-diameter disk of the target fungi was placed at an equidistant distance between the bacteria and the edge of the plate. The disk was obtained from the edge of the colony where the growth is most active. The initial distance between the bacteria and the fungus was 3.25 cm, and the bacteria were streaked up to 2 cm from the edge of the plate. The plates containing the bacteria and the fungus in confrontation were sealed and incubated at 25 ± 1°C in darkness until the fungus reached the edge of the plate opposite to the bacteria. At this point, measurements were performed. The distance between the edge of the bacterial growth and the edge of the fungal colony (inhibition distance) was measured. The approximate incubation times varied depending on the target fungus: 3–4 days for Botrytis cinerea, Trichoderma longibrachiatum, Trichoderma harzianum T-22, and Trichoderma atroviride, 7–8 days for Fusarium oxysporum f. sp. radicis-cucumerinum (FORC), Fusarium proliferatum, Colletotrichum gloeosporioides, and Trichoderma asperellum, and 7–10 days for Phytophthora capsici, while Paecilomyces lilacinus exhibited the slowest growth, taking 20–30 days until measurement. Antagonistic activity was considered when the inhibition distance was equal to or greater than 4 mm to any of the target fungus, and additionally, the growth of the target fungi did not continue after the 5-day observation period. Five replicates were prepared for each combination bacteria-fungi.
2.5.3 Identification of soil-isolated bacteria with suppressive properties
The isolates that showed antagonistic activity were analyzed by means of PCR sequencing of amplicons of the 16S rRNA gene using the primers 27F-1492R (Condori et al., 2019) and subsequent database searches using BLASTN software, based on the consensus sequences created by aligning the forward and reverse sequences of the target isolates. The PCR conditions were 5 min at 94°C, 35 cycles of 1 min at 94°C, 1 min at 55°C, 2 min at 72°C, and a final elongation of 7 min at 72°C. Purified PCR products were sequenced using the Sanger sequencing method (Instituto de Biología Molecular y Celular de Plantas, Valencia, Spain) and edited via base calling on the BioEdit program (Hall, 1999).
2.6 Statistical analysis
Data on the total culturable bacteria and thermophilic culturable bacteria populations, sequencing depth for the 16S library, the relative abundance of dominant bacterial phyla and genera, and the indices of biodiversity were analyzed via an analysis of variance (one-way ANOVA) to compare differences among sampling times. Previously, normality and homoscedasticity were tested using the Shapiro–Wilk and Levene tests, respectively. The Kruskal–Wallis one-way non-parametric test (p = 0.05) was performed when normality or homoscedasticity of data was not evident (p < 0.05 Shapiro–Wilk or Levene tests, respectively). Arcsine square root transformation was applied to percentages before analyses. Duncan’s post hoc test was then applied to perform a pairwise comparison between sampling times at a 95% confidence level. These statistical analyses were carried out using the statistical software package Statgraphics Centurion XIX (Statgraphics Technologies, Inc., The Plains, VA, United States) for Windows (Microsoft Corporation, WA, United States).
3 Results
3.1 Taxonomic compositions of the bacterial communities
The total culturable bacteria population exhibited values ranging from 2,920,000 to 24,200,000 CFU/g dry soil depending on the sampling time (Figure 1). The highest values were observed at the end of Season 1, which showed significant differences (p < 0.001) compared to the other sampling times. For thermophilic culturable bacteria, the population ranged from 2,170,000 to 11,300,000 CFU/g dry soil. In this case, the population was lower at the beginning of Season 1 (p < 0.05) compared to the other sampling times (Figure 1).
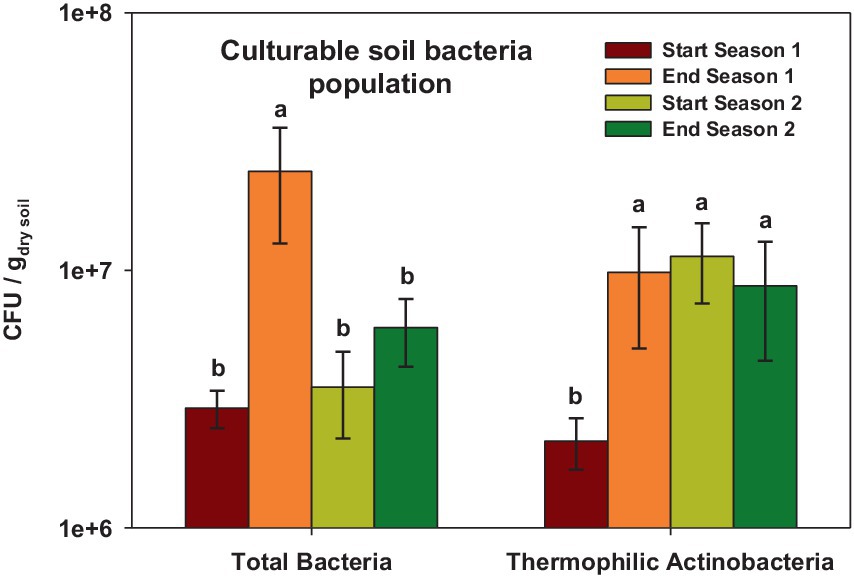
Figure 1. Total Colony Forming Units (CFU) of soil culturable bacteria and thermophilic bacteria at the start and at the end of season 1 (2019/20) and season 2 (2020/2021). In each case, different letters denote significant differences between sampling times at p ≤ 0.05 (Duncan’s test).
Figure 2 illustrates the relative abundance of different microbial phyla (Figures 2A,B) and genera (Figures 2C,D) at the start and at the end of the two seasons. Regarding phyla, Proteobacteria dominated in both start and end seasons, with a significant increase (p < 0.001) from 28.6% at the start to 52.3% at the end in season 1, and from 17.6% at the start to 32.6% at the end in season 2. Firmicutes showed an increase from 7.8% in start season 1 to 30.1% in start season 2, followed by a decrease to 16.3% in end season 2 (p < 0.001). Actinobacteriota also showed fluctuations (p < 0.001), peaking at 13.5% in end season 2. Acidobacteriota, Gemmatimonadota, Methylomirabilota, and Myxococcota showed the highest relative abundance in start season 1 (p < 0.001). Other phyla, such as Bacteroidota, Planctomycetota, Patescibacteria, Chlorofexi displayed varying patterns but maintained presence throughout the two seasons.
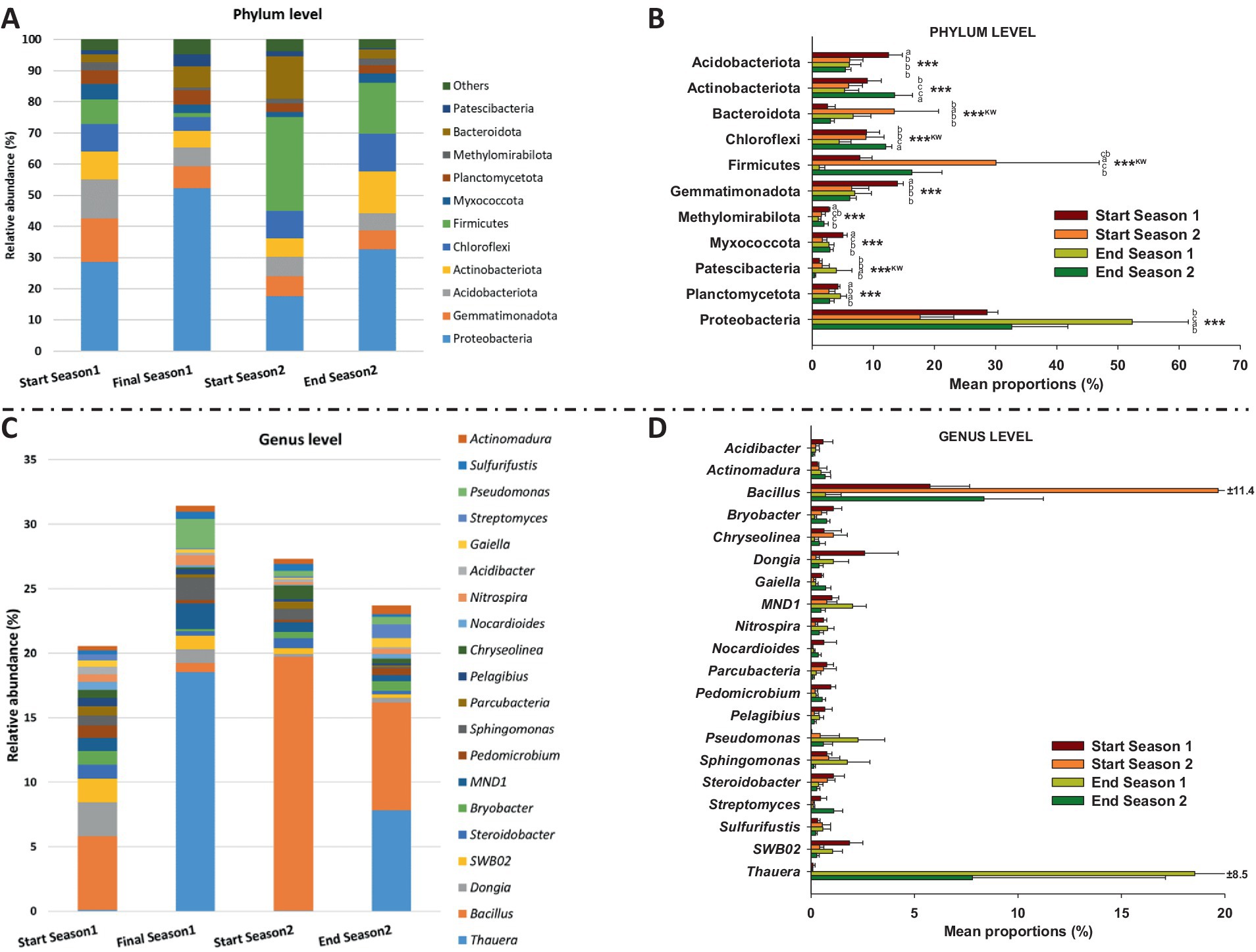
Figure 2. Bacterial community data found in the soil by 16S RNA sequencing. Relative abundance (%) of dominant bacterial phyla (A,B) and genera (C,D) divided into the start and end of the 2 years (seasons). “***” indicates significance at p ≤ 0.001 (Duncan’s test). “KW” p-value through the Kruskal–Wallis non-parametric test. Note: All genera in (D), except Acidibacter sp., showed significance at p ≤ 0.01 through the Kruskal–Wallis non-parametric test.
Similarly, the relative abundance of bacterial genera varied across seasons (Figure 2D). Thauera experienced increase in both seasons, rising from 0.09 to 18.55% in Season 1 and from 0.03 to 7.80% in Season 2. In contrast, Bacillus showed decrease, dropping from 5.74 to 0.69% in Season 1 and from 19.68 to 8.36% in Season 2. Other notable trends include MND1, which increased from 1.00 to 2.00% in Season 1. Some genera, like Pseudomonas, exhibited notable fluctuations, with a peak of 2.26% in End Season 1. Others, such as Streptomyces, Dongia and SWB02 showed sporadic occurrences throughout the seasons, with values ranging from 0.09 to 1.09%. All genera, except Acidibacter, showed significant differences among sampling times (p ≤ 0.01) with no common pattern among them.
3.2 Alpha diversity of soil bacterial community
Generally, the alpha diversity indices showed significant differences among different sampling times (p < 0.01), except for the Dominance-Simpson index (Figures 3A–D). Thus, species richness (Margalef index) was higher at the end of the crop in both years compared to values at the start of the crops (p = 0.0000), with average values ranging from 53.30 ± 7.01 to 37.59 ± 3.99 (Figure 3A). The Evenness-Pielou Index did not offer this trend, showing higher values at the start and end of the study compared to values at the start of the second year (p = 0.0064), although these differences were not detected at the end of the crop in year 1. In the case of the Diversity-Shannon index, values were lower at the start of the crop in year 2 compared to the other time points considered (p = 0.0007), with average values ranging from 4.35 ± 0.41 to 5.07 ± 0.35.
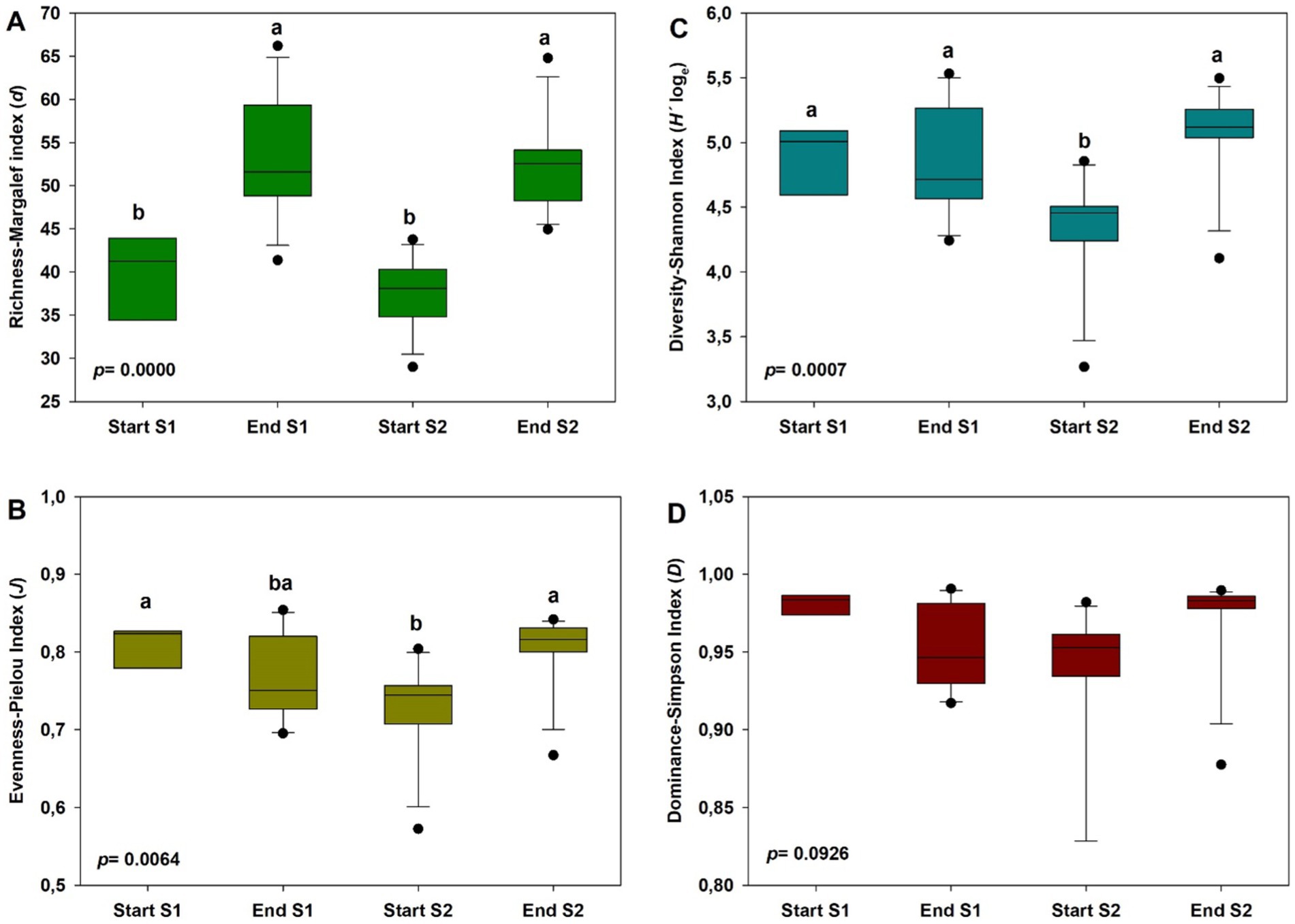
Figure 3. Alpha diversity indices of soil bacterial community at the start and at the end of season 1 (S1; 2019/20) and season 2 (S2; 2020/2021): Margalef richness index (A), Shannon–Wiener diversity index (B), Pielou’s evenness index (C) and Simpson’s dominance index. In each case, different letters denote significant differences between sampling times at p ≤ 0.01 (Duncan’s test).
3.3 Beta diversity of soil bacterial community. Comparison of soil bacterial community structure among sampling times
The global PERMANOVA test indicated a significant effect (Pseudo-F = 9.6531, p < 0.0001) of the sampling time on bacterial communities’ beta diversity. In addition, pairwise tests revealed significant differences between bacterial communities across different seasons (Supplementary Table S1). Notably, comparisons between the start and end of each year exhibit considerable dissimilarities (p < 0.003). However, comparisons between the different sampling times highlight significant variations in bacterial composition in all cases (p < 0.0001).
Principal coordinates analysis (PCoA) further supports the statistical findings of the PERMANOVA analysis. The PCoA results show that the PCoA1 and PCoA2 explained 29.6 and 13.2% of the difference in the compositions of the bacterial communities among samples (Figure 4). The results of PCoA showed that the bacterial communities in the samples of the four different sampling time had a discrete distribution, with relatively large distances among the samples of each moment and a clear clustering of samples from each time.
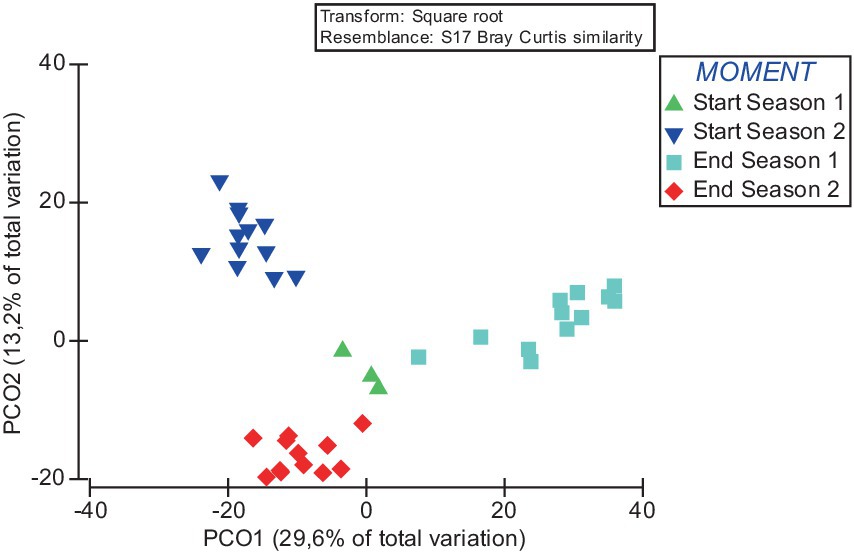
Figure 4. Principal coordinate analysis (PCoA) of the influence of sampling time on the beta diversity of soil bacterial communities. Over the 2 years of the study, soil samples were taken at the start of the crop (Start Season: after incorporating fresh sheep manure in summer followed by solarisation) and at the end (End Season).
3.4 Compositional differences and biomarkers of the bacterial communities
LEfSe analysis identified biomarkers that caused significant differences among the four sampling times (Figure 5). A total of 5, 10, 5, and 2 biomarkers (LDA score > 4.5) were identified in the soil bacterial community analyzed by means of DNA metabarcoding at the Start Season 1, Start Season 2, End Season 1 and End Season 2, respectively.
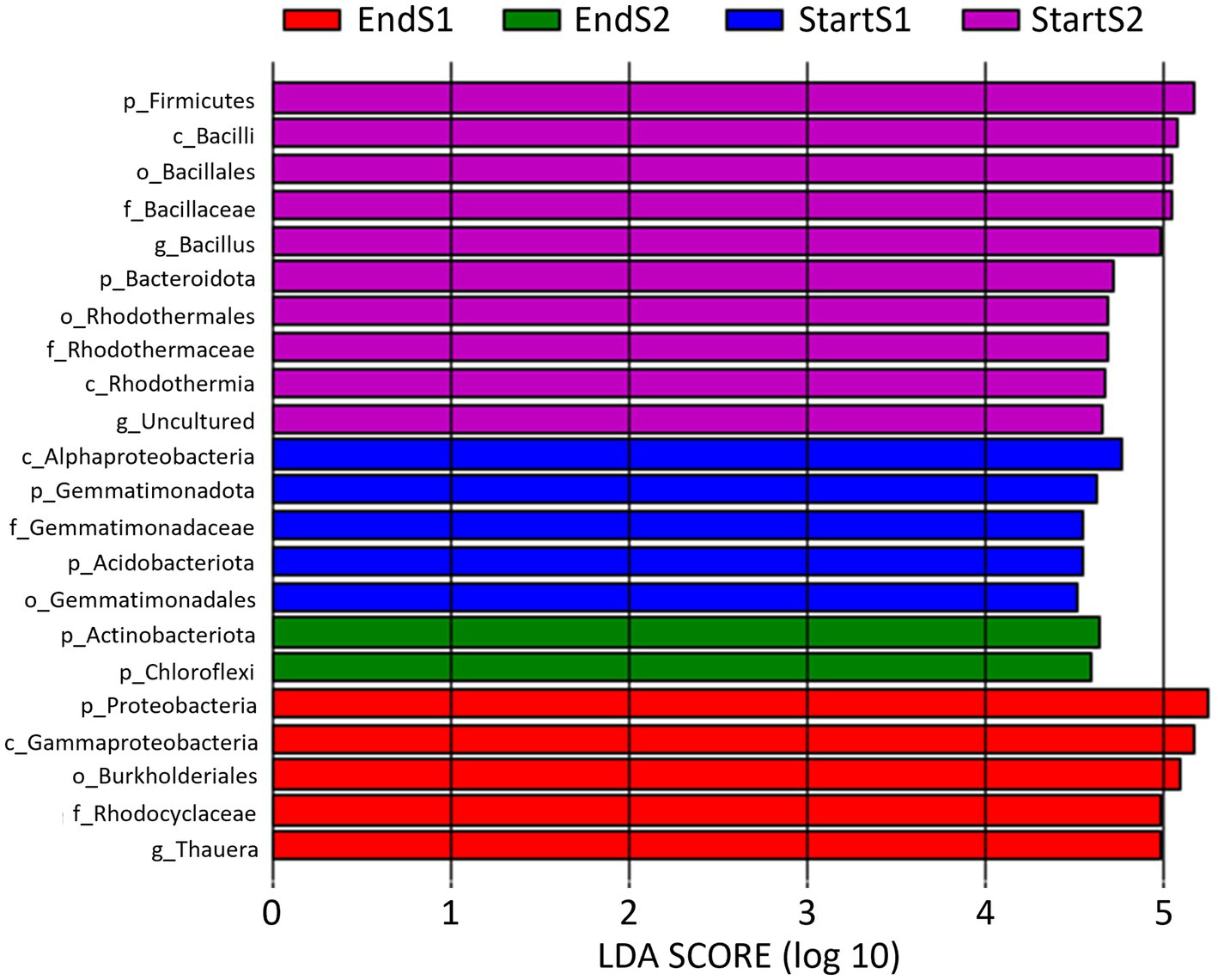
Figure 5. Linear discriminant analysis (LDA) to detect differentially abundant taxa across groups of soil samples depending on sampling time: start and end of season 1 (S1; 2019/20) and season 2 (S2; 2020/2021). Only taxa with an LDA score >4.5 (p < 0.05) are shown.
When data from the 2 years of study are pooled based on whether they come from soil samples taken at the start or at the end of the seasons, a total of 9 and 5 biomarkers were identified at the beginning and end of the crop, respectively (Figure 6). Thus, the phylum Firmicutes, class Bacilli, order Bacillales, family Bacillaceae, and genus Bacillus, along with the class Rhodothermia, order Rhodothermales, family Rhodothermaceae, and an uncultured genus, were the biomarkers identified at the start of the crop. Whereas at the end of the crop, they were the phylum Proteobacteria, class Gammaproteobacteria, order Burkholderiales, family Rhodocyclaceae, and genus Thauera.
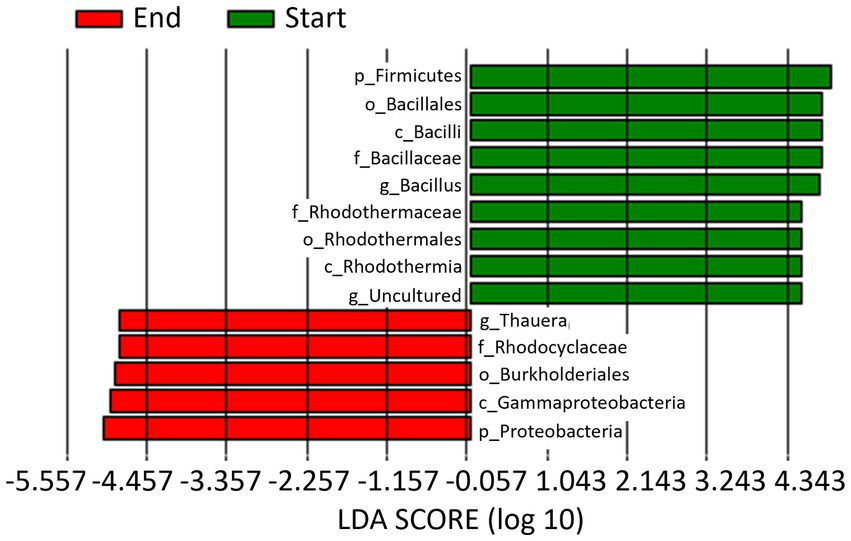
Figure 6. Linear discriminant analysis (LDA) to detect differentially abundant taxa across groups; only taxa with an LDA score >4.5 (p < 0.05) are shown. Data from the 2 years of study are pooled based on whether they come from soil samples taken at the start (Start) or at the end (End) of the seasons.
3.5 Soil bacterial function prediction
The abundances of different bacterial functional pathways were predicted using the FAPROTAX database.
3.5.1 Functional annotation overview based on 16S rRNA data
From a total sequencing reads average per sample of 56,976, an average of 29,763 reads per sample received functional assignment to any of the 52 functional groups detected via FAPROTAX database. This represents 45 ± 23.4% of the total sequencing reads. Considering the percentage of readings assigned to functionality at the different sampling times, the highest value was observed in the End Season 1 (68.9 ± 15.7%), which showed differences compared to the other sampling moments (p ≤ 0.001; Duncan’s test), including End Season 2, which presented an average value of 50.4 ± 14.4%. It is noteworthy that the lowest values were detected at the Start Season in both years of study, with values of 22.0 ± 4.2% and 21.4 ± 5.8% in Season 1 and Season 2, respectively.
3.5.2 Dominant functional groups and their temporal distribution
Based on the FAPROTAX database, the top 15 functional groups which represented an average of 97.1 ± 2.2%, 87.2 ± 15.5%, 95.2 ± 2.7% and 92.7 ± 6.3% of the total sequencing reads assigned to functionality of Start Season 1, Start Season 2, End Season 1 and End Season 2 soil samples, respectively, were obtained. The relative proportion (i.e., percentage) of these top 15 functional groups across all sampling times is showed as a heatmap (Figure 7). In all sampling times, chemoheterotrophy emerges as the primary ecological pathway, representing more than 20% of the sequences in all cases, and surpassing 50% in some of them. This is followed by aerobic chemoheterotrophy, which similarly exhibits values above or close to 20%. It is remarkable that no entry was found for group “plant pathogen.” Figure 7 represents similarities in the relative abundance of the 15 bacterial functional groups within the soil bacterial communities at each sampling time, particularly between Start Season 1 and Start Season 2, as well as between End Season 1 and End Season 2. Nitrogen respiration, nitrate respiration, nitrite respiration, and nitrate reduction are observed with higher relative abundance in the samples from End Season 1 and End Season 2, whereas Fermentation emerges as the ecological pathway that appears to be more relevant in the samples from Start Season 1 and Start Season 2. In this regard, When the global PERMANOVA test was conducted, a significant main effect (Pseudo-F = 31.467, p < 0.0001) of sampling time on the composition of functional groups was detected (Supplementary Table S2). However, the only case where pairwise PERMANOVA tests did not find significant differences was between the samples from Start Season 1 and Start Season 2. Principal Coordinates Analysis supported the statistical findings of the PERMANOVA analysis. In this case, the PCoA results indicated that PCoA1 and PCoA2 explained 80.3 and 7.5% of the differences in compositions and diversity of the bacterial functional groups (Figure 8). The results of PCoA showed that the functional bacterial groups in the samples of the four different sampling time had a discrete distribution, with relatively large distances among the samples of each moment and a clear clustering of samples from each sampling time, except for the samples from Start Season 1 and Start Season 2, which exhibited mixing. Although pairwise PERMANOVA tests found differences between End Season 1 and End Season 2, the PCoA results showed all samples from End Season of both years separated from those of Start Season of both years along PCoA1.
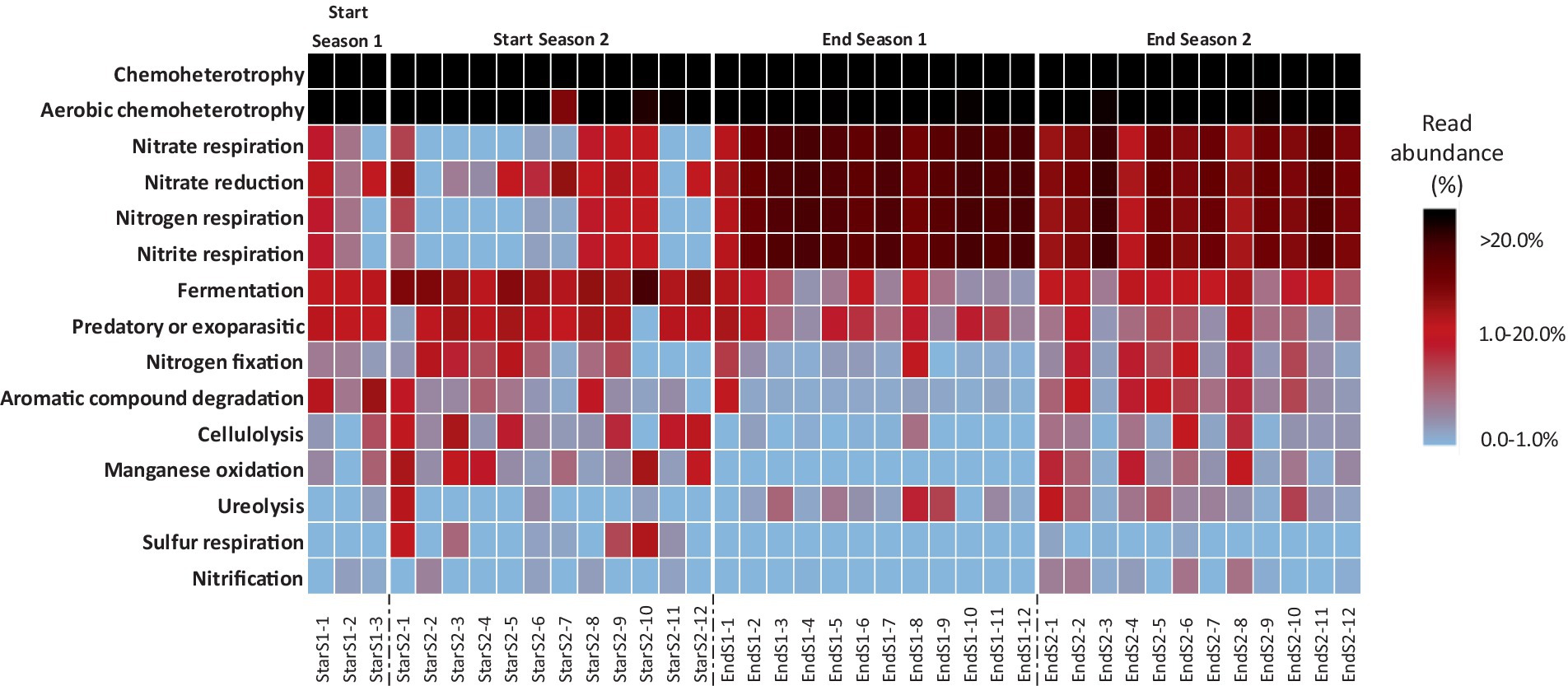
Figure 7. Relative abundance heat map showing the relative abundance of 15 bacterial functional groups within the bacterial communities found in the soil at each sampling time. Functional groups were normalized within each sample via total sum scaling resulting in a color scale denoting the proportion within each sample.
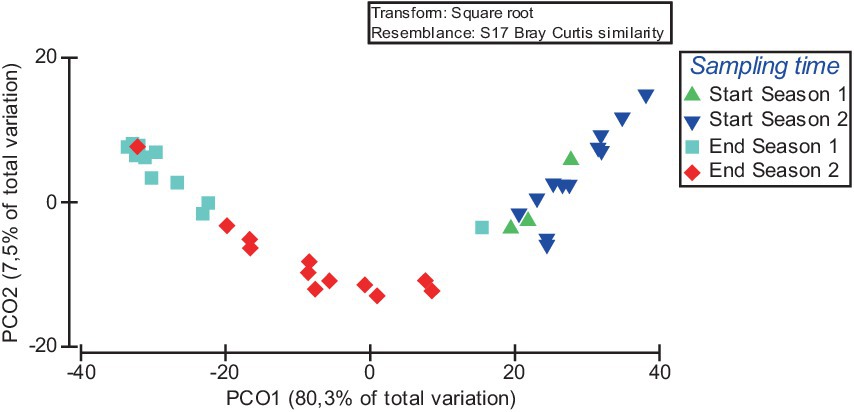
Figure 8. Principal coordinate analysis (PCoA) of the influence of sampling time on the composition and diversity of the top 15 bacterial functional groups. Over the 2 years of the study, soil samples were taken at the start of the crop (Start Season: after incorporating fresh sheep manure in summer followed by solarisation) and at the end (End Season).
3.5.3 Differential functional pathways identified by LEfSe
LEfSe analysis identified a total of 13 functional pathways that caused significant differences (LDA score > 3) among the four sampling times (Figure 9): Aromatic compound degradation, Chemoheterotrophy and aerobic chemoheterotrophy at Start Season 1; fermentation, predatory or exoparasitic, manganese oxidation, cellulolysis y Sulfur respiration at Start Season 2, nitrogen, nitrate and nitrite respiration and nitrate reduction at End Season 1, and ureolysis at End Season 2.
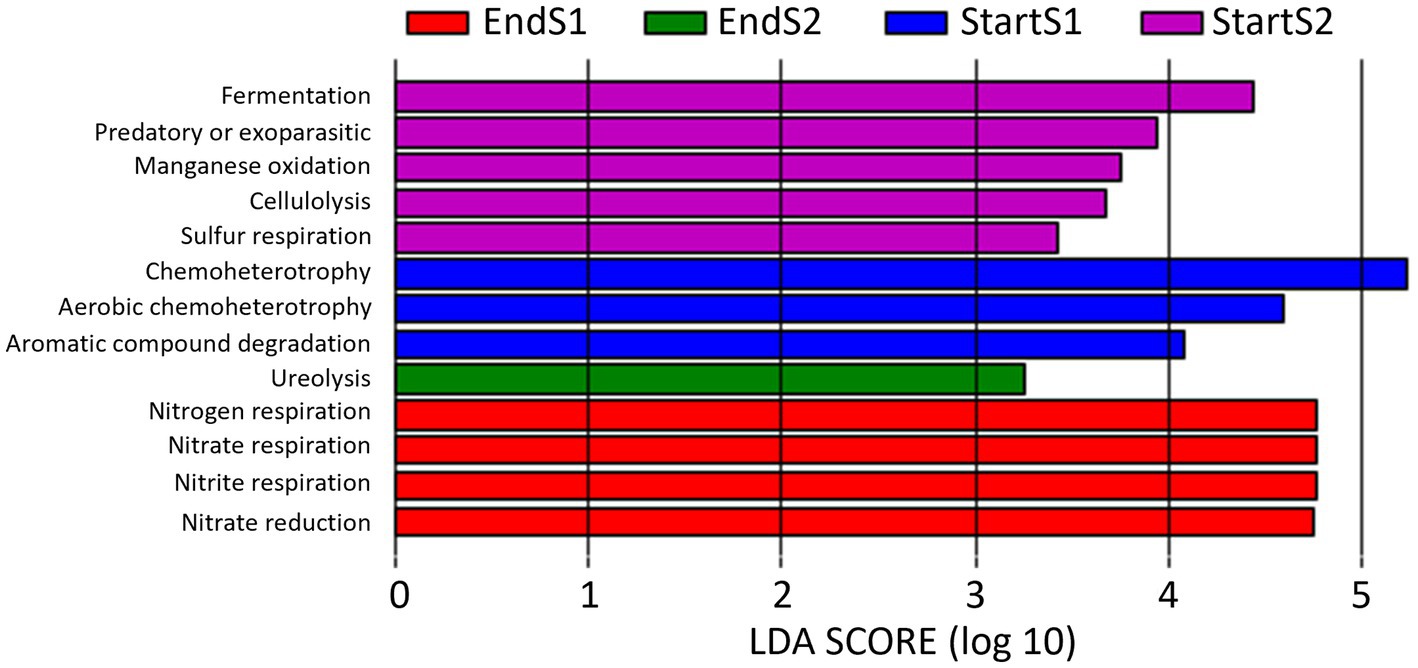
Figure 9. Linear discriminant analysis (LDA) to detect differentially abundant ecological pathways across groups of soil samples depending on sampling time: start and end of season 1 (S1; 2019/20) and season 2 (S2; 2020/2021). Only ecological pathways with an LDA score >3 (p < 0.05) are shown.
When data from the 2 years of study are pooled based on whether they come from soil samples taken at the start or at the end of the seasons, a total of 6 and 5 markers were identified at the start and end of the season, respectively (Figure 10). Thus, Chemoheterotrophy, fermentation, predatory or exoparasitic, manganese oxidation, cellulolysis and sulfur respiration were the biomarkers identified at Start Season, while nitrogen, nitrate and nitrite respiration, nitrate reduction and ureolysis at End Season.
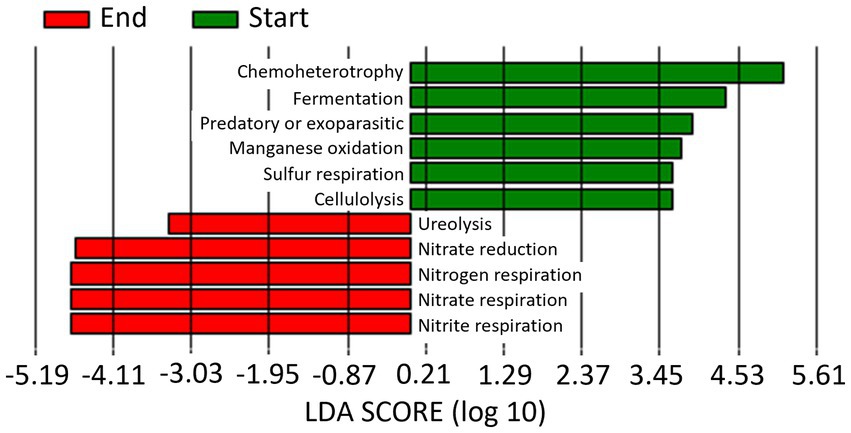
Figure 10. Linear discriminant analysis (LDA) to detect differentially abundant ecological pathways across groups; only ecological pathways with an LDA score >3 (p < 0.05) are shown. Data from the 2 years of study are pooled based on whether they come from soil samples taken at the start (Start) or at the end (End) of the seasons.
3.6 Antagonistic bacteria against the in vitro growth of phytopathogenic and beneficial fungi
In total, out of the 95 isolates tested, 36 of them (37.9%) showed antagonism, 30 (31.6%) against phytopathogenic fungi, 19 (20.0%) against beneficial fungi, and 12 (12.6%) against both (Table 2; Figure 11). Colletotrichum gloeosporioides and Botrytis cinerea were the pathogenic fungi against which the highest number of isolates showed antagonism, with 58.3 and 41.7% of the isolates showing antagonism, respectively. Meanwhile, Trichoderma asperellum and T. atroviride were prominent among the beneficial fungi, with 33.3 and 25.0% of the isolates, respectively. Only 8.3% exhibited antagonism against F. proliferatum and Forc, while T. longibrachiatum and Paecilomyces lilacinus were the fungi with the least antagonistic response, with only 1 and 2 isolates (2.8 and 5.6%), respectively. Depending on the sampling time, 36.4% of the isolates from Start Season 1, 54.9% of the isolates from End Season 1, and 22.2% of the isolates from Start Season 2 exhibited antagonism against target fungi. All isolates were identified through PCR (16S rRNA): Streptomyces spp. (26), Bacillus spp. (4), Brevundimonas spp. (2), Pseudochrobactrum spp. (2), Stenotrophomonas sp. (1), Alcaligenes sp. (1).
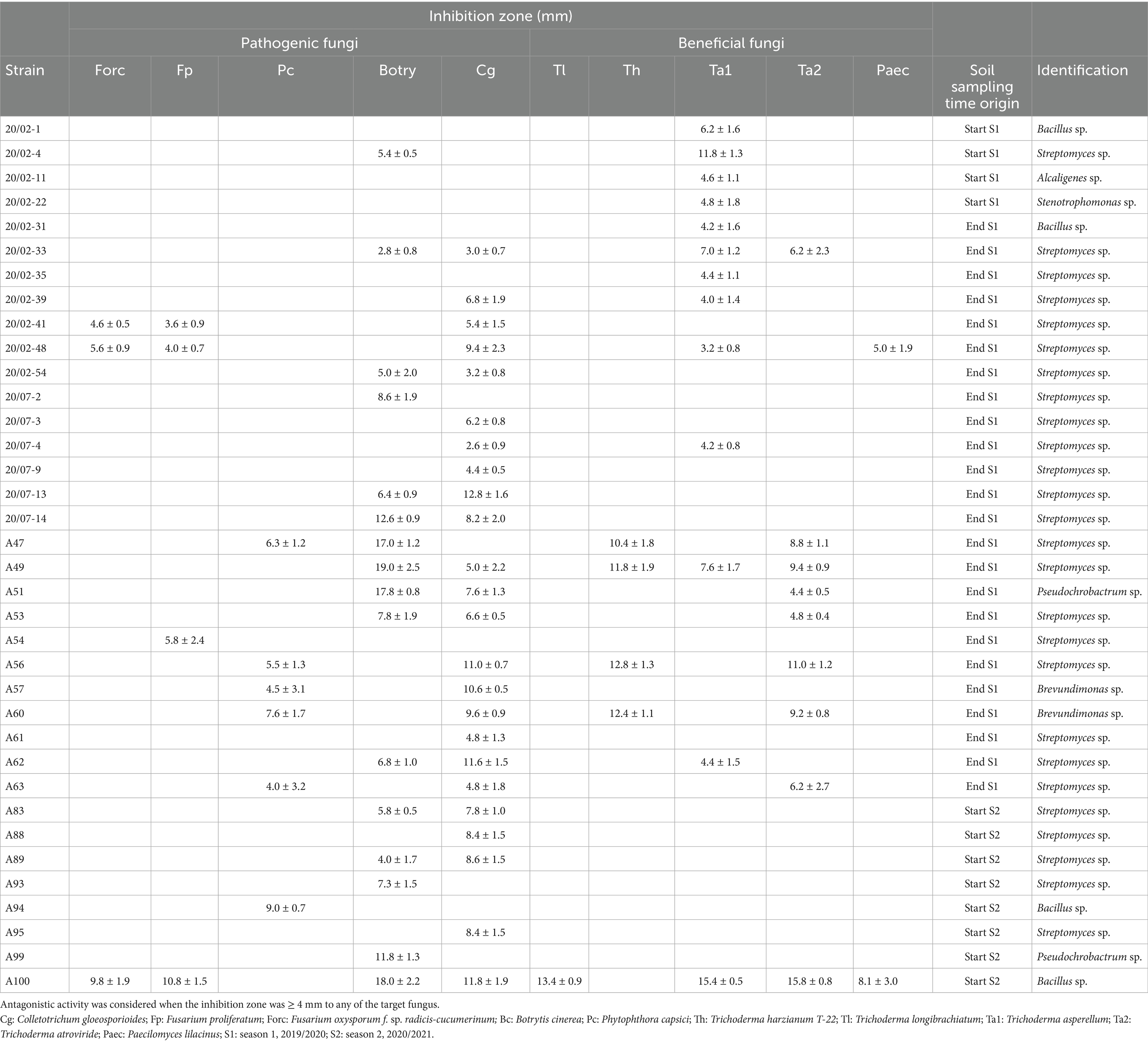
Table 2. Antagonistic activity of soil-isolated bacteria at different sampling times against phytopathogenic and beneficial fungi, and their identification through polymerase chain reaction (PCR) sequencing of amplicons of the 16S rRNA gene.
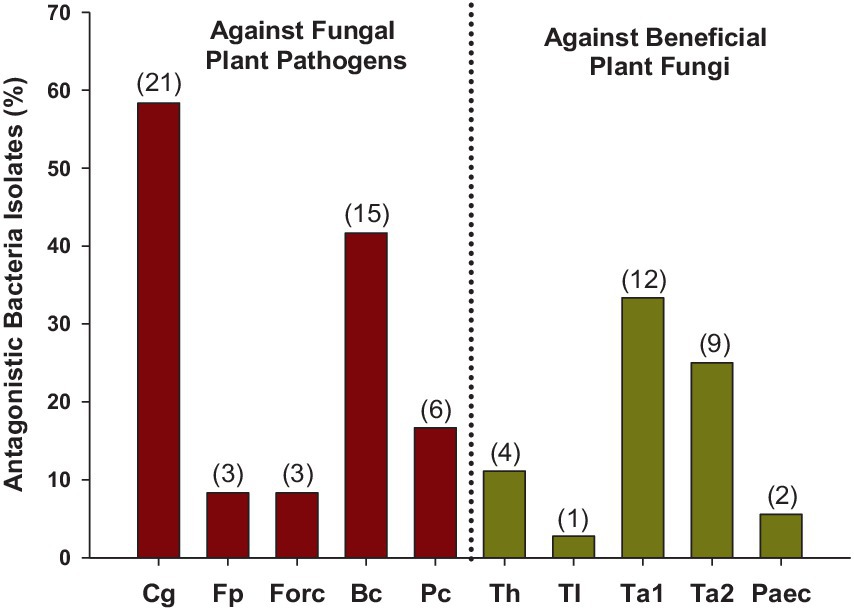
Figure 11. Soil-isolated bacteria with antagonism against the tested fungi. In total, 95 isolates were evaluated. Percentages expressed over the total bacteria that exhibited antagonism, which were 36. In brackets, the number of isolates with antagonism against each fungus. Only those with the inhibition distance equal to or greater than 4 mm to the target fungus were considered. Cg: Colletotrichum gloeosporioides; Fp: Fusarium proliferatum; Forc: Fusarium oxysporum f. sp. radicis-cucumerinum; Bc: Botrytis cinerea; Pc: Phytophthora capsici; Th: Trichoderma harzianum T-22; Tl: Trichoderma longibrachiatum; Ta1: Trichoderma asperellum; Ta2: Trichoderma atroviride; Paec: Paecilomyces lilacinus.
4 Discussion
In this study, the total culturable bacterial populations and thermophilic culturable bacteria experienced shifts depending on the sampling time, but no clear trend was observed and the impact on these bacterial fractions population cannot be conclusively attributed to the treatments. In this regard, it’s important to consider that changes in soil quality parameters and sampling time are factors that highly influence microbial variations, particularly in cultivated and amended soils (Li et al., 2018; Vido et al., 2024).
A noticeable outcome of the study is the dynamics of the bacterial communitys alpha diversity throughout the crop season. Primarily, species richness (Margalef index), but also Shannon diversity and Pielou’s evenness indices, appeared lower at pre-planting phase (Start Season), meanwhile at the end of the cropping seasons, all alpha diversity indices tended to increase and consistently reach higher values. In addition, significant changes in bacterial community beta diversity were recorded across the different sampling times (PERMANOVA p < 0.01). In this sense, changes in the soil microbial community have been previously reported after the application of manure in crop fields (Hartmann et al., 2015; Sun et al., 2015; Tang et al., 2023), as well as after solarisation treatments when manure and other organic amendments had been previously incorporated (Achmon et al., 2020; Marín-Guirao et al., 2019b, 2023a; Shea et al., 2022). According to Vido et al. (2024) our results also highlight the dynamic nature of bacterial community composition over time at phylum and genus level when soils are amended with organic materials and organic crop management. In any case, Proteobacteria and Firmicutes were the most prevalent phyla in all sampling times. These two phyla, along with Actinobacteriota, are consistently associated with soil-borne disease suppression (Raaijmakers et al., 2009; Mendes et al., 2011).
In our study, the relative abundance of Firmicutes was higher at pre-planting phase (Start Season) compared to End Season, contrary to Proteobacteria, which consistently showed higher levels at the End Season in both years. Thus, Bacillus (phylum Firmicutes) was identified as a biomarker in Start Season, while Thauera (phylum Proteobacteria) was the biomarker at the genus level in End Season. These are biomarkers indicating a superior ability to discriminate between the studied conditions and a higher statistical significance and relevance for the study (LDA score >4.5). Bacillus is a widely studied genus for its suppressive properties against soil pathogens (Miljaković et al., 2020; Zhang et al., 2023). However, bacteria of the genus Thauera have been biotechnologically interesting for their ability to degrade aromatic compounds (Elder and Kelly, 1994; Philipp and Schink, 2000), and more recently for their role in the production of a promising alternative to non-degradable plastics (Koller et al., 2017). To the best of our knowledge, Thauera has not been reported to have capabilities for controlling soil pathogens. The observed changes in the abundances of these two genera may have been influenced by the high temperatures reached during the solarization process that followed the incorporation of the organic amendment (Pullman et al., 1981). While, Bacillus spp. are recognized thermophilic bacteria (Edwards et al., 1965; Bond and Favero, 1975), most Thauera species are recognized as mesophilic bacteria (Fida et al., 2016), although a thermophilic Thauera species has been also reported (Yang et al., 2018). In fact, the treatments did not completely eliminate Thauera bacteria but maintained a viable inoculum capable of recolonizing the soil during the crop season. In this regard, the incorporation of an organic amendment rich in nitrogen, such as fresh sheep manure, could favor the presence of these bacteria in the studied soil. Both bacteria genera are active microorganisms involved in the nitrogen cycle and have been recognized as dominant denitrifiers in various environments, including soils (Fuka et al., 2007). In this regard, we identified ecological (functional) pathways related to the nitrogen cycle (nitrate and nitrite respiration, nitrate reduction, and ureolysis) that present a higher percentage of functional related sequences at the End of the Season compared to the Start of the Season, immediately after the treatments. As previously reported, alkeline hydrolyzable organic nitrogen content and soil organic carbon content are key factors that drive shifts in soil microbial composition and could also determine the observed changes in microbial functional communities formed at different successional stages (Dasgupta et al., 2024; Ding et al., 2024).
A relevant result of the present study is the presence of native bacteria with antagonistic properties against prevalent pathogenic fungi, but also against common beneficial fungi sourced from commercial products. Even a large number of these isolates exhibited antagonism against both pathogenic and beneficial fungi, thus highlighting the complexity of the edaphic system, an aspect often overlooked in agricultural soil management plans. In addition, isolates with antagonistic activity were detected at all sampling times throughout the study, both following the treatments performed and at the end of the crop season, which suggests that they are also present during the development of the crops. Among the native antagonistic bacteria (16S rRNA), Streptomyces was the dominant genus followed by Bacillus, while bacteria from the genera Brevundimonas, Pseudochrobactrum, Stenotrophomonas, and Alcaligenes also showed antagonism. Among them, Streptomyces spp. and Bacillus spp. are among the most studied biological control agents. Their role in the suppression of several plant pathogens has been well-documented and attributed to their production of a wide variety of bioactive secondary metabolites and enzymes, including antibiotics, bacteriocins, and volatile compounds (Alabouvette and Steinberg, 2006; Kunova et al., 2016; Zhang et al., 2023).
The evaluation of the specific soil management practices needed for developing and maintaining soil microbial functionality which favor DSS in cultivable lands is a highly complex issue that is difficult to decipher (Alabouvette and Steinberg, 2006). It must be considered that the complexity of the microbial interactions as well as the underlying mechanisms and microbial traits remain elusive for most disease suppressive soils (Gómez Expósito et al., 2017). In any case, in our study, isolates with antagonistic activity were detected at all sampling times, even after incorporating fresh sheep manure in summer followed by solarisation for 2 months. All the isolates used in the antagonism assays, were obtained from the thermophilic bacteria analyses performed in this study, and thus their presence is high in the evaluated soil, with populations ranging from millions to tens of millions of CFU/g dry soil. Thus, the soil management described in this study was compatible with, and possibly responsible for the high presence of these microorganisms in the soil of the greenhouse. This is a highly relevant information for the development of a soil suppressive to soil pathogens, but also for the use of certain commercial products containing BCO and other beneficial microorganisms for crops, as the presence of native bacteria with these properties can influence their effectiveness.
In conclusion, the results of this study show changes in the soil bacterial community and its functionality over the crop cycle following the application of sheep manure combined with summer solarisation in a Mediterranean greenhouse. The study reflects the presence of native bacteria with antagonistic properties inhabiting the mentioned biosolarised soils. Moreover, this fact also demonstrates the differential antagonistic ability of each isolate concerning the phytopathogenic or beneficial profile of the tested fungal species. These results are highly relevant for the application of agronomic soil management practices aimed at improving the natural suppressiveness of soils, to diseases caused by fungi and actinomycetes, as well as for the proper use of formulations based on beneficial fungi. However, further studies are necessary to elucidate whether the suppressive properties against soil-borne pathogens of the studied native bacteria are clearly manifested when considering the intricate interactions among soil, plant, microbiome, and environment. Additionally, these results should guide the development of commercial formulations with microorganisms that ensure farmers the establishment of these microorganisms in their soil and the derived benefits.
Data availability statement
The datasets and sequences from this study are available in the Zenodo open-access repository under the OPTIMSOIL project (PID2021-125545OR-C21): https://zenodo.org/records/15545241.
Author contributions
JM-G: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Software, Validation, Writing – original draft, Writing – review & editing. MC-G: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Grant PID2021-125545OR-C21 funded by MICIU/AEI/10.13039/501100011033 and by “ERDF/EU.”
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1484219/full#supplementary-material
References
Achmon, Y., Claypool, J. T., Fernández-Bayo, J. D., Hernandez, K., McCurry, D. G., Harrold, D. R., et al. (2020). Structural changes in bacterial and fungal soil microbiome components during biosolarization as related to volatile fatty acid accumulation. Appl. Soil Ecol. 153:103602. doi: 10.1016/j.apsoil.2020.103602
Adesina, M. F., Lembke, A., Costa, R., Speksnijder, A., and Smalla, K. (2007). Screening of bacterial isolates from various European soils for in vitro antagonistic activity towards Rhizoctonia solani and fusarium oxysporum: site- dependent composition and diversity revealed. Soil Biol. Biochem. 39, 2818–2828. doi: 10.1016/j.soilbio.2007.06.004
Águila-Carricondo, P., Román, R., Marín-Guirao, J. I., Cantón, Y., and de Cara, M. (2024). Native biocrust-cyanobacteria strains showing antagonism against three soilborne pathogenic fungi. Pathogens 13:579. doi: 10.3390/pathogens13070579
Akanmu, A. O., Babalola, O. O., Venturi, V., Ayilara, M. S., Adeleke, B. S., Amoo, A. E., et al. (2021). Plant disease management: leveraging on the plant-microbe-soil interface in the biorational use of organic amendments. Front. Plant Sci. 12:700507. doi: 10.3389/fpls.2021.700507
Alabouvette, C., Lemanceau, P., and Steinberg, C. (1993). Recent advances in the biological control of fusarium wilts. Pestic. Sci. 37, 365–373. doi: 10.1002/ps.2780370409
Alabouvette, C., and Steinberg, C. (2006). “The soil as a reservoir for antagonists to plant diseases” in An ecological and societal approach to biological control. eds. J. Eilenberg and H. M. T. Hokkanen, vol. 2 (Dordrecht the Netherlands: Springer), 123–144. doi: 10.1007/978-1-4020-4401-4_8
Amir, H., and Alabouvette, C. (1993). Involvement of soil abiotic factors in the mechanisms of soil suppressiveness to fusarium wilts. Soil Biol. Biochem. 25, 157–164. doi: 10.1016/0038-0717(93)90022-4
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070
Andrews, S. (2010). FastQC: a quality control tool for high throughput sequence data ; Cambridge: Babraham Bioinformatics, Babraham Institute. Available online at: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (Accessed April 8, 2024).
Baker, K., and Cook, R. J. (1974). Biological control of plant pathogens. New York: WH Freeman and Company.
Bokulich, N. A., Kaehler, B. D., Rideout, J. R., Dillon, M., Bolyen, E., Knight, R., et al. (2018). Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 6:90. doi: 10.1186/s40168-018-0470-z
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Bonanomi, G., Antignani, V., Capodilupo, M., and Scala, F. (2010). Identifying the characteristics of organic soil amendments that suppress soilborne plant diseases. Soil Biol. Biochem. 42, 136–144. doi: 10.1016/j.soilbio.2009.10.012
Bonanomi, G., Antignani, C., Pane, C., and Scala, F. (2007). Suppression of soil borne fungal diseases with organic amendments. J. Plant Pathol. 89, 311–324.
Bonanomi, G., Lorito, M., Vinale, F., and Woo, S. L. (2018). Organic amendments, beneficial microbes, and soil microbiota: toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 56, 1–20. doi: 10.1146/annurev-phyto-080615-100046
Bond, W. W., and Favero, M. S. (1975). Thermal profile of a Bacillus species (ATCC 27380) extremely resistant to dry heat. Appl. Microbiol. 29, 859–860. doi: 10.1128/am.29.6.859-860.1975
Brizuela, A. M., De La Lastra, E., Marín-Guirao, J. I., Gálvez, L., De Cara-García, M., Capote, N., et al. (2020). Fusarium consortium populations associated with Asparagus crop in Spain and their role on field decline syndrome. J. Fungi 6:336. doi: 10.3390/jof6040336
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Castillo-Díaz, F. J., Belmonte-Ureña, L. J., Camacho-Ferre, F., and Tello Marquina, J. C. (2022). Biodisinfection as a profitable fertilization method for horticultural crops in the framework of the circular economy. Agronomy 12:521. doi: 10.3390/agronomy12020521
Castillo-Díaz, F. J., Marín-Guirao, J. I., Belmonte-Ureña, L. J., and Tello-Marquina, J. C. (2021). Effect of repeated plant debris reutilization as organic amendment on greenhouse soil fertility. Int. J. Environ. Res. Public Health 18:11544. doi: 10.3390/ijerph182111544
Chng, S., Cromey, M., Dodd, S., Stewart, A., Butler, R. C., and Jaspers, M. V. (2015). Take-all decline in New Zealand wheat soils and the microorganisms associated with the potential mechanisms of disease suppression. Plant Soil 397, 239–259. doi: 10.1007/s11104-015-2620-4
Clarke, K. R., and Gorley, R. N. (2006). Primer v6: User manual or tutorial. Plymouth, UK: Plymouth Marine Laboratory, 190.
Condori, S., Fernández, P., and Valderrama, M. R. (2019). Aislamiento y caracterización de Streptomyces spp. rizosféricos promotores del crecimiento vegetal. Idesia 37, 109–116. doi: 10.4067/S0718-34292019000200109
Crawford, D., Lynch, J., Whipps, J., and Ousley, M. (1993). Isolation and characterization of actinomycete antagonists of a fungal root pathogen. Appl. Environ. Microbiol. 59, 3899–3905. doi: 10.1128/aem.59.11.3899-3905.1993
Dasgupta, D., Richardson, A. E., Camuy-Vélez, L. A., Kirkby, C., Kirkegaard, J. A., and Banerjee, S. (2024). Microbial dynamics during in-situ organic matter decomposition reveals the importance of keystone taxa in the core microbiome. Appl. Soil Ecol. 199:105396. doi: 10.1016/j.apsoil.2024.105396
Ding, M., Dai, H., He, Y., Liang, T., Zhai, Z., Zhang, S., et al. (2024). Continuous cropping system altered soil microbial communities and nutrient cycles. Front. Microbiol. 15:1374550. doi: 10.3389/fmicb.2024.1374550
Eberlein, C., Heuer, H., Vidal, S., and Westphal, A. (2016). Microbial communities in Globodera pallida females raised in potato monoculture soil. Phytopathology 106, 581–590. doi: 10.1094/PHYTO-07-15-0180-R
Edwards, J. L. Jr., Busta, F. F., and Speck, M. L. (1965). Thermal inactivation characteristics of Bacillus subtilis spores at ultrahigh temperatures. Appl. Microbiol. 13, 851–857. doi: 10.1128/am.13.6.851-857.1965
Elder, D. J., and Kelly, D. J. (1994). The bacterial degradation of benzoic acid and benzenoid compounds under anaerobic conditions: unifying trends and new perspectives. FEMS Microbiol. Rev. 13, 441–468. doi: 10.1111/j.1574-6976.1994.tb00061.x
Emelyanov, V. V. (2001). Evolutionary relationship of Rickettsiae and mitochondria. FEBS Lett. 501, 11–18. doi: 10.1016/S0014-5793(01)02618-7
Ewels, P., Magnusson, M., Lundin, S., and Käller, M. (2016). MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics 32, 3047–3048. doi: 10.1093/bioinformatics/btw354
Fida, T. T., Chen, C., Okpala, G., and Voordouw, G. (2016). Implications of limited thermophilicity of nitrite reduction for control of sulfide production in oil reservoirs. Appl. Environ. Microbiol. 82, 4190–4199. doi: 10.1128/AEM.00599-16
Fuka, M. M., Braker, S. H. G., and Philippot, L. (2007). “Molecular tools to assess the diversity and density of denitrifying Bacteria in their habitats” in Biology of the nitrogen cycle. eds. H. Bothe, S. J. Ferguson, and W. E. Newton (Amsterdam, Netherlands: Elsevier), 313–330.
Garbeva, P., Postma, J., van Veen, J. A., and van Elsas, J. D. (2006). Effect of above-ground plant species on soil microbial community structure and its impact on suppression of Rhizoctonia solani AG3. Environ. Microbiol. 8, 233–246. doi: 10.1111/j.1462-2920.2005.00888.x
Garbeva, P., van Veen, J. A., and van Elsas, J. D. (2004). Microbial diversity in soil: selection microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 42, 243–270. doi: 10.1146/annurev.phyto.42.012604.135455
García, C., Céspedes, A. J., Páez, F. C., Cano, M., Martín, E., and Parra, S. (2024). Trends in soil disinfection in greenhouses in southeastern Spain. Acta Hortic. 1410, 51–56. doi: 10.17660/ActaHortic.2024.1410.8
García-Raya, P., Ruiz-Olmos, C., Marín-Guirao, J. I., Asensio-Grima, C., Tello-Marquina, J. C., and de Cara-García, M. (2019). Greenhouse soil Biosolarization with tomato plant debris as a unique fertilizer for tomato crops. Int. J. Environ. Res. Public Health 16:279. doi: 10.3390/ijerph16020279
Giagnocavo, C., de Cara-García, M., González, M., Juan, M., Marín-Guirao, J. I., Mehrabi, S., et al. (2022). Reconnecting farmers with nature through agroecological transitions: interacting niches and experimentation and the role of agricultural knowledge and innovation systems. Agriculture 12:137. doi: 10.3390/agriculture12020137
Gómez Expósito, R., de Bruijn, I., Postma, J., and Raaijmakers, J. M. (2017). Current insights into the role of rhizosphere Bacteria in disease suppressive soils. Front. Microbiol. 8:2529. doi: 10.3389/fmicb.2017.02529
Hall, T. A. (1999). Bioedit: a user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98.
Hartmann, M., Frey, B., Mayer, J., Mader, P., and Widmer, F. (2015). Distinct soil microbial diversity under long-term organic and conventional farming. ISME J. 9, 1177–1194. doi: 10.1038/ismej.2014.210
Hemkemeyer, M., Schwalb, S. A., Berendonk, C., Geisen, S., Heinze, S., Joergensen, R. G., et al. (2024). Potato yield and quality are linked to cover crop and soil microbiome, respectively. Biol. Fertil. Soils 60, 525–545. doi: 10.1007/s00374-024-01813-0
Herlemann, D. P., Labrenz, M., Jürgens, K., Bertilsson, S., Waniek, J. J., and Andersson, A. F. (2011). Transitions in bacterial communities along the 2000 km salinity gradient of the Baltic Sea. ISME J. 5, 1571–1579. doi: 10.1038/ismej.2011.41
Hoitink, H. A. J., and Boehm, M. J. (1999). Biocontrol within the context of soil microbial communities: a substrate-dependent phenomenon. Annu. Rev. Phytopathol. 37, 427–446. doi: 10.1146/annurev.phyto.37.1.427
Hoitink, H. A. J., Stone, A. G., and Han, D. Y. (1997). Suppression of plant disease by composts. HortScience 32, 184–187.
Jayaraman, S., Naorem, A., Lal, R., Dalal, R. C., Sinha, R. K., Patra, A. K., et al. (2021). Disease-suppressive soils—beyond food production: a critical review. J. Soil Sci. Plant Nutr. 21, 1437–1465. doi: 10.1007/s42729-021-00451-x
Kang, H., Chai, A., Lin, Z., Shi, Y., Xie, X., Li, L., et al. (2024). Deciphering differences in microbial community diversity between Clubroot-diseased and healthy soils. Microorganisms 12:251. doi: 10.3390/microorganisms12020251
Koller, M., Maršálek, L., de Sousa Dias, M. M., and Braunegg, G. (2017). Producing microbial polyhydroxyalkanoate (PHA) biopolyesters in a sustainable manner. New Biotechnol. 37, 24–38. doi: 10.1016/j.nbt.2016.05.001
Kunova, A., Bonaldi, M., Saracchi, M., Pizzatti, C., Chen, X., and Cortesi, P. (2016). Selection of Streptomyces against soil borne fungal pathogens by a standardized dual culture assay and evaluation of their effects on seed germination and plant growth. BMC Microbiol. 16:272. doi: 10.1186/s12866-016-0886-1
Lal, R. (2020). Managing soil quality for humanity and the planet. Front. Agricult. Sci. Eng. 7, 251–253. doi: 10.15302/J-FASE-2020329
Li, L., Xu, M., Eyakub Ali, M., Zhang, W., Duan, Y., and Li, D. (2018). Factors affecting soil microbial biomass and functional diversity with the application of organic amendments in three contrasting cropland soils during a field experiment. PLoS One 13:e0203812. doi: 10.1371/journal.pone.0203812
Louca, S., Parfrey, L. W., and Doebeli, M. (2016). Decoupling function and taxonomy in the global ocean microbiome. Science 353, 1272–1277. doi: 10.1126/science.aaf4507
Marín-Guirao, J. I., De Cara-García, M., Crisol-Martínez, E., Gómez-Tenorio, M. Á., García-Raya, P., and Tello-Marquina, J. C. (2019a). Association of plant development to organic matter and fungal presence in soils of horticultural crops. Ann. Appl. Biol. 174, 339–348. doi: 10.1111/aab.12501
Marín-Guirao, J. I., de Cara-García, M., and Tello-Marquina, J. C. (2019b). Effect of soil biodisinfection on soil fungal communities associated to horticultural crops. Ecosistemas 28, 63–72. doi: 10.7818/ECOS.1708
Marín-Guirao, J. I., García-García, M. C., Martín-Expósito, E., and de Cara-García, M. (2023a). Continued organic fertigation after basal manure application does not impact soil fungal communities, tomato yield or soil fertility. Microorganisms 11:1715. doi: 10.3390/microorganisms11071715
Marín-Guirao, J. I., Páez-Cano, F. C., García-García, M. C., Katsoulas, N., and de Cara-García, M. (2023b). Evaluation of copper-free alternatives to control grey mould in organic Mediterranean greenhouse tomato production. Agronomy 13:137. doi: 10.3390/agronomy13010137
Mazzola, M. (2002). Mechanisms of natural soil suppressiveness to soilborne diseases. Antonie Van Leeuwenhoek 81, 557–564. doi: 10.1023/A:1020557523557
Mazzola, M., and Freilich, S. (2016). Prospects for biological soil-borne disease control: application of indigenous versus synthetic microbiomes. Phytopathology 107, 256–263. doi: 10.1094/PHYTO-09-16-0330-RVW
McMurdie, P. J., and Holmes, S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Mendes, R., Kruijt, M., de Bruijn, I., Dekkers, E., van der Voort, M., and Schneider, J. H. M. (2011). Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 332, 1097–1100. doi: 10.1126/science.1203980
Miljaković, D., Marinković, J., and Balešević-Tubić, S. (2020). The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 8:1037. doi: 10.3390/microorganisms8071037
Moore, K. R., Magnabosco, C., Momper, L. M., Gold, D. A., Bosak, T., and Fournier, G. P. (2019). An expanded ribosomal phylogeny of Cyanobacteria supports a deep placement of plastids. Front. Microbiol. 10:1612. doi: 10.3389/fmicb.2019.01612
Ni, H., Zong, R., Sun, J., Wu, Y., Yu, L., Liu, Y., et al. (2022). Response of bacterial community to the occurrence of Clubroot disease in Chinese cabbage. Front. Microbiol. 13:922660. doi: 10.3389/fmicb.2022.922660
Pankhurst, C. E., and Lynch, J. M. (1995). The role of soil microbiology in sustainable intensive agriculture. Adv. Plant Pathol. 11, 229–247.
Philipp, B., and Schink, B. (2000). Two distinct pathways for anaerobic degradation of aromatic compounds in the denitrifying bacterium Thauera aromatica strain AR-1. Arch. Microbiol. 173, 91–96. doi: 10.1007/s002039900112
Postma, J., and Schilder, M. T. (2015). Enhancement of soil suppressiveness against Rhizoctonia solani in sugar beet by organic amendments. Appl. Soil Ecol. 94, 72–79. doi: 10.1016/j.apsoil.2015.05.002
Pullman, G. S., De Vay, J. E., and Garber, R. H. (1981). Soil solarization and thermal death: algorithmic relationship between time and temperature for four soilborne plant pathogens. Phytopathology 71, 959–964.
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Raaijmakers, J. M., Paulitz, T. C., Steinberg, C., Alabouvette, C., and Moenne-Loccoz, Y. (2009). The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321, 341–361. doi: 10.1007/s11104-008-9568-6
R-Team-Core (2019). R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. Available online at: https://www.R-project.org/.
Sagova-Mareckova, M., Omelka, M., and Kopecky, J. (2023). The Golden goal of soil management: disease-suppressive soils. Phytopathology 113, 741–752. doi: 10.1094/PHYTO-09-22-0324-KD
Schlatter, D., Kinkel, L., Thomashow, L., Weller, D., and Paulitz, T. (2017). Disease suppressive soils: new insights from the soil microbiome. Phytopathology 107, 1284–1297. doi: 10.1094/PHYTO-03-17-0111-RVW
Schneider, R. W. (1982). Suppressive soils and plant disease. St Paul, MN: American Phytopathological Society.
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12:R60. doi: 10.1186/gb-2011-12-6-r60
Shannon, C. E., and Weaver, W. (1949). The mathematical theory of communication. Urbana, IL: University Illinois Press.
Shea, E. A., Fernández-Bayo, J. D., Hodson, A. K., Parr, A. E., Lopez, E., Achmon, Y., et al. (2022). Biosolarization restructures soil bacterial communities and decreases parasitic nematode populations. Appl. Soil Ecol. 172:104343. doi: 10.1016/j.apsoil.2021.104343
Siegel-Hertz, K., Edel-Hermann, V., Chapelle, E., Terrat, S., Raaijmakers, J. M., and Steinberg, C. (2018). Comparative microbiome analysis of a fusarium wilt suppressive soil and a fusarium wilt conducive soil from the Chateaurenard region. Front. Microbiol. 9:568. doi: 10.3389/fmicb.2018.00568
Sun, R., Zhang, X., Guo, X., Wang, D., and Chu, H. (2015). Bacterial diversity in soils subjected to long-term chemical fertilization can be more stably maintained with the addition of livestock manure than wheat straw. Soil Biol. Biochem. 88, 9–18. doi: 10.1016/j.soilbio.2015.05.007
Tang, T., Sun, X., Liu, Q., Dong, Y., and Zha, M. (2023). Treatment with organic manure inoculated with a biocontrol agent induces soil bacterial communities to inhibit tomato fusarium wilt disease. Front. Microbiol. 13:1006878. doi: 10.3389/fmicb.2022.1006878
Tello, J. C., Vares, F., and Lacasa, A. (1991). “Análisis de muestras” in Manual de Laboratorio. Diagnóstico de Hongos. Bacterias y Nematodos Fitopatógenos (Madrid: MAPA), 9–48.
Vido, J. J., Wang, X., Sale, P. W. G., Celestina, C., Shindler, A. E., and Hayden, H. L. (2024). Bacterial community shifts occur primarily through rhizosphere expansion in response to subsoil amendments. Environ. Microbiol. 26:e16587. doi: 10.1111/1462-2920.16587
Weller, D. M., Raaijmakers, J. M., Gardener, B. B., and Thomashow, L. S. (2002). Microbial populations responsible for specific soil suppressiveness. Annu. Rev. Phytopathol. 40, 309–348. doi: 10.1146/annurev.phyto.40.030402.110010
Xiong, W., Zhao, Q., Zhao, J., Xun, W., Li, R., Zhang, R., et al. (2015). Different continuous cropping spans significantly affect microbial community membership and structure in a vanilla-grown soil as revealed by deep pyrosequencing. Microb. Ecol. 70, 209–218. doi: 10.1007/s00248-014-0516-0
Yang, L., Muhadesi, J. B., Wang, M. M., Wang, B. J., Liu, S. J., and Jiang, C. Y. (2018). Thauera hydrothermalis sp. nov., a thermophilic bacterium isolated from hot spring. Int. J. Syst. Evol. Microbiol. 68, 3163–3168. doi: 10.1099/ijsem.0.002960
Keywords: agroecology, antagonism, biosolarisation, FAPROTAX, DNA metabarcoding, organic amendment, sustainability, Trichoderma
Citation: Marín-Guirao JI and de Cara-García M (2025) Native bacteria from a Mediterranean greenhouse associated to soil health and suppressiveness. Front. Microbiol. 16:1484219. doi: 10.3389/fmicb.2025.1484219
Edited by:
Anil K. H. Raghavendra, Elizabeth Macarthur Agricultural Institute, AustraliaReviewed by:
Wensheng Fang, Chinese Academy of Agricultural Sciences, ChinaRafael Hernández Maqueda, University of Almeria, Spain
Ana M. Pastrana, University of California, Agriculture and Natural Resources, United States
Copyright © 2025 Marín-Guirao and de Cara-García. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jose Ignacio Marín-Guirao, am9zZWkubWFyaW5AanVudGFkZWFuZGFsdWNpYS5lcw==; Miguel de Cara-García, ZnJhbmNpc2NvbS5jYXJhQGp1bnRhZGVhbmRhbHVjaWEuZXM=