 Min Wang
Min Wang Wei Liu
Wei Liu Lianwen Zheng
Lianwen Zheng Shuai Ma
Shuai Ma Lianhai Jin3*
Lianhai Jin3* Donghai Zhao
Donghai Zhao- 1Department of Obstetrics and Gynecology, The Second Hospital of Jilin University, Changchun, China
- 2College of Laboratory Medicine, Jilin Medical University, Jilin, China
- 3Low Pressure and Low Oxygen Environment and Health Intervention Innovation Center, Jilin Medical University, Jilin, China
- 4College of Basic Medicine, Jilin Medical University, Jilin, China
As a heterogeneous disease, endometriosis is associated with diagnostic delay. Delayed diagnosis, physical discomfort, hormone therapy, and inconvenience in daily life and work all contribute to a decreased quality of life for endometriosis patients. Early clinical diagnosis is highly important for the intervention and treatment of endometriosis. Currently, reliable non-invasive diagnostic methods are lacking, and laparoscopic examination combined with pathological diagnosis is considered the “gold standard” for definitively diagnosing endometriosis. An increasing number of studies have confirmed the correlation between endometriosis and microbial ecological changes. Microbial dysbiosis is an important factor in the development and progression of endometriosis. Certain key microbial species and their metabolites can induce functional alterations in endometrial cells through various mechanisms, often preceding the emergence of clinical symptoms. Endometriosis are chronic inflammatory diseases, with an immunoinflammatory response as the pathological foundation. The microbiome may participate in the pathological mechanisms of endometriosis through multiple pathways, including mediating inflammatory responses, regulating immune responses, participating in estrogen regulation, interfering with metabolic activities, and modulating the gut–brain axis. Therefore, the microbiome holds potential as an early non-invasive diagnostic and therapeutic target for endometriosis patients. This study summarizes and analyses the correlations between microorganisms and their metabolites and the onset of endometriosis, aiming to provide novel insights into the etiology, diagnosis, and treatment of endometriosis.
1 Introduction
Endometriosis refers to the presence, growth, infiltration, and recurrent bleeding of endometrial tissue with growth potential, including glands and stroma, in locations outside the uterine cavity (Taylor et al., 2021; Kvaskoff et al., 2021). Endometriosis predominantly affect adolescent and reproductive-aged women, with clinical pathological classifications encompassing ovarian, peritoneal, deep infiltrating, and other site-specific endometriosis (Horne and Missmer, 2022). Despite being a benign condition, endometriosis exhibit characteristics reminiscent of malignancy, such as implantation, invasion, and distant metastasis, significantly impacting the health and quality of life of women of reproductive age (Arion et al., 2020). The etiology and pathogenesis of endometriosis remain elusive, with surgery or hormone therapy being common therapeutic interventions. However, these approaches are associated with high recurrence rates, adverse effects of hormone therapy, and the risk of postoperative recurrence (Lamceva et al., 2023). With the advancement and maturation of genomics and high-throughput sequencing technologies, the correlation between the human microecological environment and female reproductive health has garnered considerable attention, particularly the relationship between microbial dysbiosis and endometriosis, which has emerged as a current research hotspot (Ventolini et al., 2022). Microbial dysbiosis plays a pivotal role in the initiation and progression of endometriosis (Adnane and Chapwanya, 2022). Imbalances in the microbiome lead to elevated proinflammatory factors and compromised immune function, contributing to the onset of endometriosis (Wang et al., 2022). Over time, immune dysregulation and chronic inflammation create an environment conducive to increased adhesion and angiogenesis, driving a vicious cycle of endometriosis pathogenesis and progression (Zhao et al., 2015). Additionally, intestinal microbial dysbiosis, characterized by an increased abundance of harmful bacteria or decreased levels of beneficial bacteria, may damage the integrity of intestinal epithelial cells and impair the mucosal barrier’s defense against pathogens. This, in turn, can alter central nervous system connections via the gut–brain axis, increasing the expression of inflammatory factors and leading to neuroinflammation and immune dysfunction (Amro et al., 2022). There is a definite correlation between endometriosis and the microbiome, suggesting that the microbiome may play a crucial role in both the pathogenesis and diagnosis of endometriosis.
2 Overview of endometriosis
Endometriosis, a chronic inflammatory and estrogen-dependent gynecological disorder, exhibit extensive pathological manifestations that can affect multiple organ systems (Chapron et al., 2019). The precise pathogenesis of endometriosis remains elusive and is intricately linked to various factors, such as epigenetics, epithelial–mesenchymal transition, inflammation, immunity, and angiogenesis (Franca et al., 2022; Fan and Li, 2022). Clinically, endometriosis present with a wide spectrum of symptoms ranging from asymptomatic to chronic pelvic pain, infertility, dysmenorrhea, and pelvic nodules or masses (Terzic et al., 2021). Owing to the non-specific nature of early symptoms and the lack of definitive serological or imaging diagnostic methods, the rate of missed diagnoses for early-stage endometriosis is alarmingly high (Schenken et al., 1984; Opoien et al., 2013). Delayed diagnosis deprives patients of the opportunity for early medication and timely disease control, often leading to surgical intervention when pelvic masses, severe dysmenorrhea, or infertility arise (He et al., 2022). Even after surgical treatment, endometriosis patients often require adjuvant medication to prevent recurrence (Bedaiwy et al., 2002). Endometriosis significantly impact women’s physical and mental health, as well as their overall quality of life (Cohen Ben-Meir et al., 2022). There is an urgent need for deeper insights into the pathogenesis of endometriosis, the development of precise early non-invasive diagnostic methods, and the search for safer and more effective treatment options (Jiang et al., 2021) (Figure 1). Mounting experimental evidence underscores the notable differences in the microbial communities between endometriosis patients, animal models, and healthy controls.
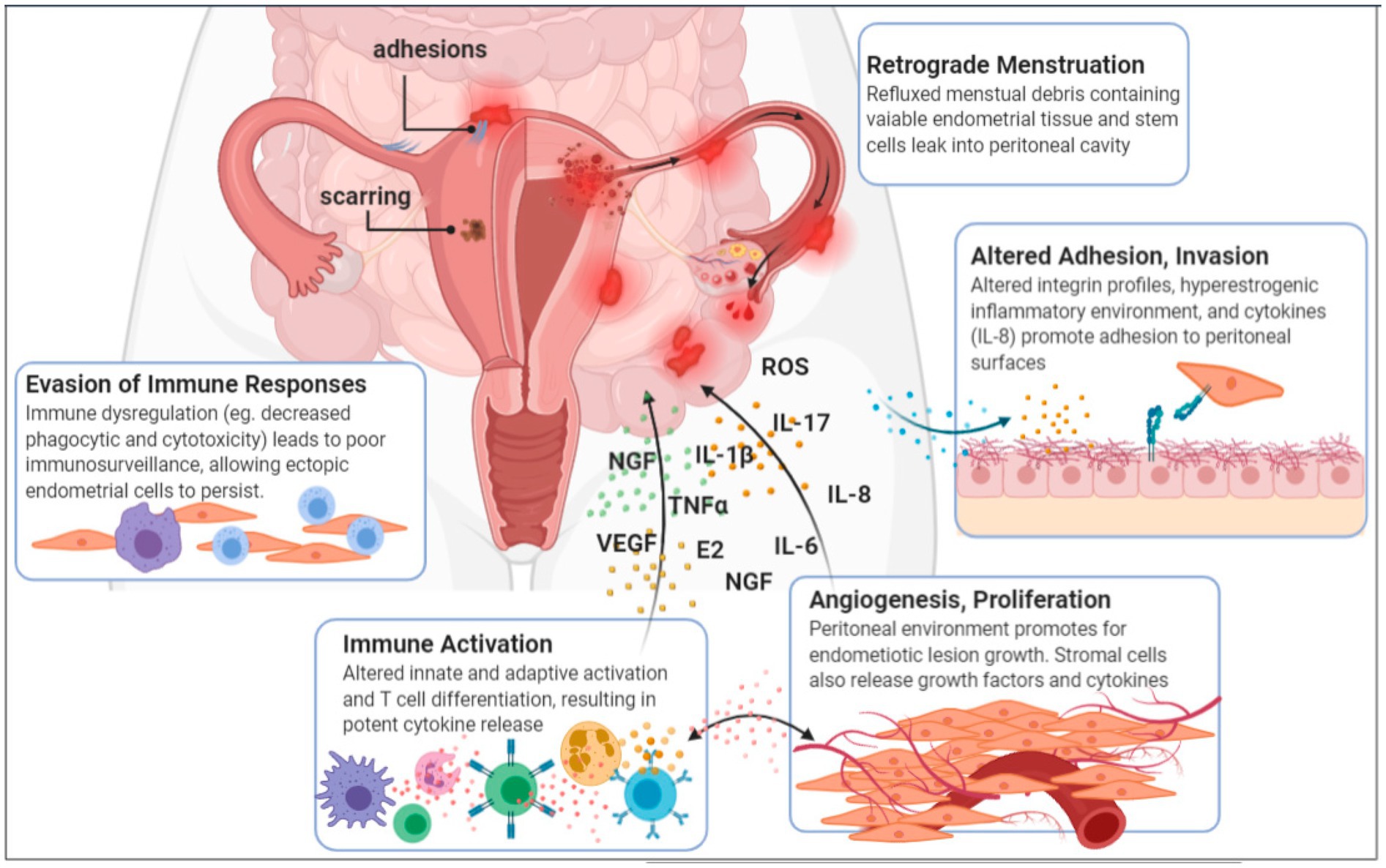
Figure 1. Aetiology and pathogenesis of endometriosis (Jiang et al., 2021).
3 Endometriosis accompanied by microbial dysbiosis
The human microecology encompasses a complex and diverse organic ensemble of symbiotic and pathogenic microorganisms residing both within and on the human body. This intricate ecosystem provides an optimal habitat for these microbes, which, in turn, directly or indirectly influence a range of host vital activities (Molina et al., 2020). The microbial communities engage in an intricate interplay with the host, exchanging energy, materials, and genetic information, fostering a dynamic balance of mutual coordination and restraint (Shabbir et al., 2021). Typically, these microbial populations inhabit the oral cavity, gastrointestinal tract, pelvic and abdominal cavities, vagina, and respiratory tract, encompassing bacteria, fungi, eukaryotes, and viruses. The microbiome, on the other hand, refers to the collective genetic material of these microorganisms (Ser et al., 2023; Berg et al., 2020). Research into the correlations between microbiome and their metabolites and endometriosis has emerged as a pivotal focus in etiological studies (Ravel et al., 2011).
3.1 Disruption of the gut microbiota
The gut microbiota represents the largest microbial population in the human body (Aggarwal et al., 2023). There is a close correlation between alterations in the gut microbiome and the onset of endometriosis (Svensson et al., 2021). Imbalances in the gut microbiota compromise the integrity of the intestinal epithelial barrier, increasing gut epithelial permeability and leading to “leakage” of lipopolysaccharides (LPS), ultimately disrupting the endocrine system, immune system, and lipid metabolism (Dicks, 2022). An increased Firmicutes-to-Bacteroidetes (F/B) ratio is considered a key indicator of gut microbial dysbiosis (Trompette et al., 2014). Disturbances in the gut microbiota can cause pelvic inflammatory adhesions, which alter the pelvic anatomy and hinder the transportation of ova and fertilized eggs through the fallopian tubes, contributing to infertility. To elucidate the causal relationship between gut microbes and endometriosis, researchers have established endometriosis animal models. Yuan et al. (2018) reported significant changes in the gut microbiota of endometriosis mice, with increased abundances of Firmicutes and Actinobacteria and decreased Bacteroidetes, suggesting that endometriosis induce gut microbial dysbiosis. Through a comparative analysis of the vaginal, cervical, and gut microbiota between stage III/IV endometriosis patients and healthy controls, Ata et al. (2019) reported increased abundances of potential pathogens, including Gardnerella, Streptococcus, Escherichia coli, Shigella, and Ureaplasma, in the cervical microbiota of endometriosis patients. The fecal microbiota of endometriosis patients was dominated by Shigella and E. coli. Studies have also revealed that, compared with healthy individuals, patients with ovarian endometriosis and deep infiltrating endometriosis exhibit significantly decreased abundances of Firmicutes and Clostridia and a notable increase in Ruminococcus in their gut microbiota (Shan et al., 2021). These findings point to the potential of the gut microbiota as a novel direction in endometriosis etiology research, and given the ease of sample collection, gut microbes hold promise as non-invasive biomarkers for early endometriosis diagnosis (Leonardi et al., 2020; Liu et al., 2024) (Figure 2).
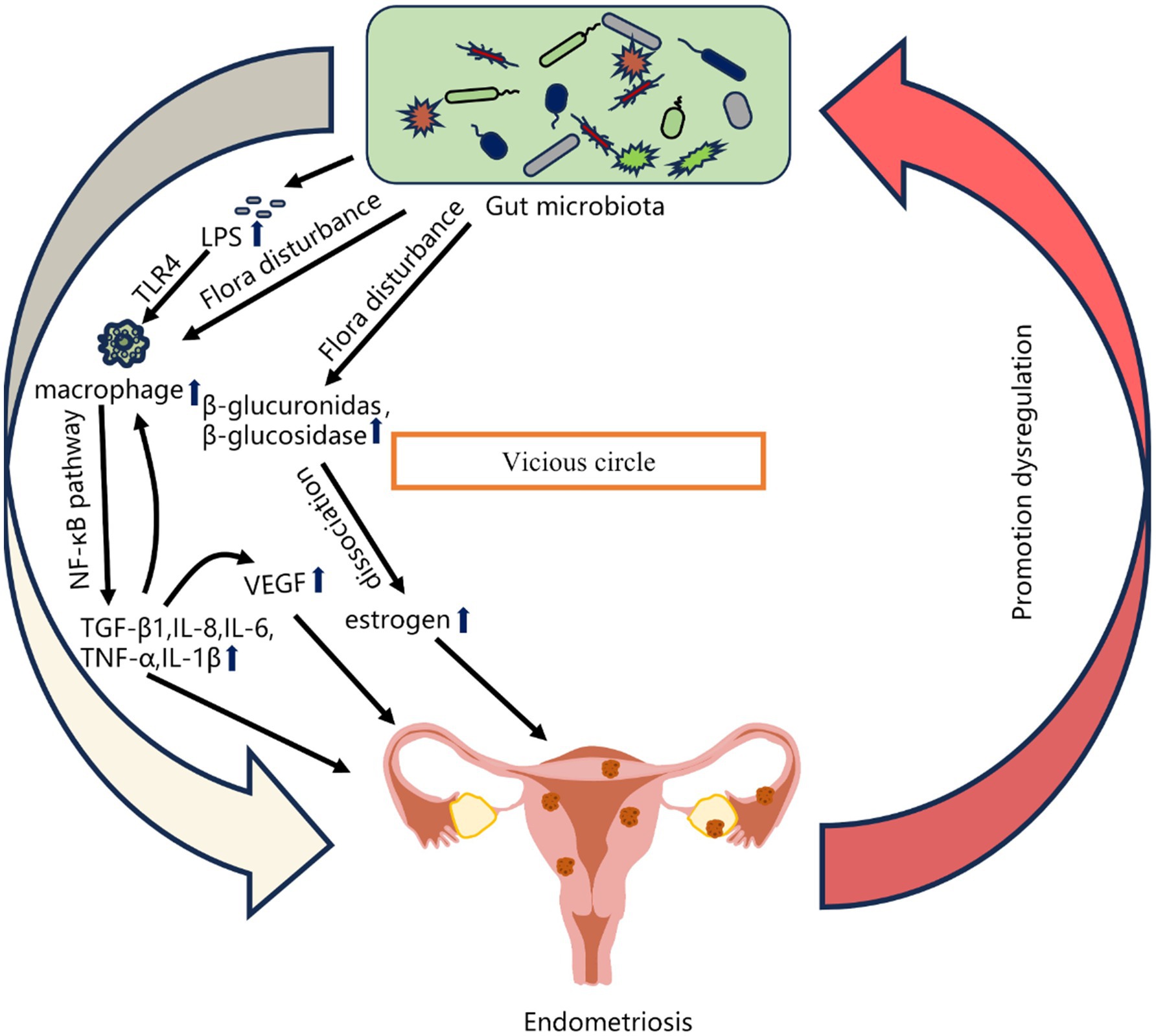
Figure 2. Relationships between gut microbiota imbalance and Endometriosis (Liu et al., 2024).
3.2 Dysbiosis of the reproductive tract microbiota
3.2.1 Vaginal microbiota
The vagina serves as a vast microbial chamber (Gao et al., 2022). In healthy females, the vaginal microbiome has a straightforward structure, with Lactobacillus species occupying a pivotal and dominant position, accompanied by other bacteria such as Gardnerella and Mycoplasma (Onywera et al., 2019). Collectively, they maintain the equilibrium of the vaginal microenvironment, preserving vaginal health. The beneficial bacteria within the vagina suppress the growth of harmful microorganisms to a certain extent, inhibiting inflammatory responses and playing a vital role in sustaining normal physiological functions (Liu et al., 2022). Lactobacillus secretes acidic substances such as lactic acid and H2O2, creating an acidic environment in the vagina with a pH that is consistently maintained below 4.5 (Witkin and Linhares, 2017). Lactic acid blocks protein deacetylases, hinders the attachment of other bacteria to epithelial cells, and enhances gene transcription and DNA repair, thereby killing or inhibiting the growth of other bacteria (Wang et al., 2021). When the colonization of Lactobacillus decreases, the vaginal pH shifts, facilitating the proliferation of anaerobic bacteria. Detrimental metabolic byproducts, which act on the vagina and cervix, disrupt the protective function of the microecological barrier, are produced or excessively accumulate. This increases the risk of vaginal infections and, consequently, female reproductive system diseases (Mahajan et al., 2022). Notably, in patients with endometriosis, the number of Lactobacillus significantly decreases, whereas other harmful bacteria, such as Actinobacteria, particularly Gardnerella and Atopobium, are significantly enriched in the vaginal microbiota. The imbalance of the vaginal microbiota is closely associated with the development of endometriosis, suggesting that delving into the vaginal microbiome for in-depth etiological research on endometriosis could lead to more effective clinical treatments.
3.2.2 Cervical microbiota
The richness and diversity of the cervical microbiome in endometriosis patients are significantly reduced (Kitaya and Yasuo, 2023). Yang et al. (2023) reported that the cervical-to-vaginal microbial ratios of certain genera are correlated with the severity of endometriosis and that these genera are clearly associated with retrograde infection pathways. Akiyama et al. (2019) reported increased levels of Enterobacteriaceae and Streptococcus in the cervical mucus of women undergoing epithelial–mesenchymal transition, where impaired reproductive tract immunity allows pathogens to persist in the cervical mucus, suggesting that bacteria may ascend into the uterine cavity to induce inflammatory responses. Research has revealed not only that the cervical microbiome of endometriosis patients differs from that of healthy women but also that there are some differences in cervical microbial populations between endometriosis patients at stages I/II and III/IV, indicating that the structure of the cervical microbiome may be dynamic during disease progression. These differential microbial populations could serve as biomarkers for endometriosis staging (Chang et al., 2022). The decreased richness and diversity of the cervical microbiota are linked to the severity of endometriosis, including higher CA125 levels and more severe symptoms such as dysmenorrhea and infertility. A systematic review encompassing 28 clinical studies and 6 animal experiments also indicated that an increase in bacterial vaginosis-associated pathogens and a depletion of Lactobacillus in the cervical-vaginal microbiome are associated with endometriosis and infertility (Salliss et al., 2021). Detection of the cervical microbiota may be biased by the influence of the vaginal microbiota during sampling, and there are also subjective differences in research techniques and data analysis.
3.2.3 Uterine microbiota
Compared with the cervical and vaginal microbiota, the endometrial microbiota of healthy women has a lower biomass but greater diversity and a notably distinct microbial composition, dominated by Lactobacillus, Prevotella, Streptomyces, and other species (Medina-Bastidas et al., 2022). Wessels et al. (2021) performed endometrial biopsies on 12 endometriosis patients and 9 non-endometriosis patients and reported a relatively high Shannon diversity index in the microbiota of endometriosis patients. Wei et al. (2020) reported that Pseudomonas and Acinetobacter were the hallmark strains in the uterine cavity of endometriosis patients, with the species richness of bacterial colonies gradually increasing from the vagina to the upper genital tract. Sampling analysis of women undergoing laparoscopic surgery revealed an association between the vaginal microbiota and endometriosis, with a greater abundance of infectious bacteria in the vagina of endometriosis women than in non-endometriosis women. Furthermore, a strong correlation was noted between Lactobacillus in both the vaginal and endometrial microbiomes (Oishi et al., 2022). Khan et al. (2014, 2016) reported that, compared with non-endometriosis patients, endometriosis patients presented a significant decrease in the proportion of Lactobacillaceae in the uterine cavity, accompanied by a marked increase in the ratios of Streptomycetaceae, Staphylococcaceae, and Enterobacteriaceae. Given the inherent difficulty for patients to accept uterine cavity sampling, resulting in limited clinical samples and insufficient research, researchers need to continue collecting data to explore the patterns and characteristics of the endometrial microbiota in endometriosis. Pelzer et al. (2018) reported that anaerobes and staphylococci are the most abundant members of the fallopian tube microbiota and that there are differences in the microbial communities between the left and right fallopian tubes, as well as between the interstitial and ampullary portions, which may be related to the impact of endometriosis lesions on pelvic anatomy and retrograde menstrual flow.
3.3 Dysbiosis of the peritoneal microbiota
The pelvic and abdominal cavities are the primary sites of endometriosis pathogenesis. In females, the pelvic and abdominal cavities communicate with the uterine cavity, allowing the dissemination of the uterine microbiota into these areas during uterine peristalsis and contractions (Yuan et al., 2022). Changes in the gut microbiota composition induced by endometriosis lead to intestinal cell apoptosis, mucus degradation, and disruption of tight junction protein complexes, thereby increasing intestinal wall permeability (Young et al., 2017), which can also result in the translocation of many Gram-negative bacteria outside the intestinal lumen (Mazur-Bialy et al., 2017). Lee et al. (2021) reported that the abundances of Acinetobacter, Pseudomonas, Streptococcus, and Enhydrobacter were significantly elevated in the peritoneal washings of endometriosis patients, whereas the abundances of Propionibacterium and Actinomyces were markedly decreased. Additionally, endometriosis patients presented significantly increased levels of IL-6, IL-10, IL-13, and TNFα in their peritoneal fluid, along with increased epithelial permeability, which allowed the release of gastrointestinal and urogenital microbiota into the peritoneal environment. Consequently, the microbial composition in the peritoneal environment of endometriosis patients also underwent alterations, dominated by Proteobacteria and Firmicutes, followed by Actinobacteria, Bacteroidetes, and Clostridia. Hernandes et al. (2020), in their study of the microbiota in peritoneal fluid from patients with deep endometriosis lesions, reported significantly increased abundances of Alishewanella, Enterococcus, and Pseudomonas, with a scarcity of Lactobacillus.
3.4 Dysbiosis of the oral microbiota
As a component of the human microbiome, the oral microecology of endometriosis patients reveals Streptococcus, Haemophilus parasuis, and Prevotella as the most prevalent genera in saliva (Hu et al., 2023; Hicks et al., 2025). While no direct correlation between oral microecology and endometriosis has been established thus far, oral microbial imbalance has been linked to certain inflammatory conditions. Following mild brain injury, there is a reduction in species richness and community diversity within the oral microbiota, with Proteobacteria, Bacteroidetes, Firmicutes, and Actinobacteria being the most representative and abundant phyla. Notably, there is a marked increase in Proteobacteria and Bacteroidetes over time following brain injury (Wang et al., 2023). Atherosclerosis fundamentally involves endothelial cell damage, and periodontal infections can either directly or indirectly contribute to immune dysregulation and increased production of systemic inflammatory mediators, thereby exacerbating atherosclerosis (Slocum et al., 2016).
4 Mechanisms by which the microbiota affects endometriosis
4.1 Microbiota-mediated inflammatory reactions contribute to the onset and progression of endometriosis
Inflammation is a pivotal process in the progression of endometriosis. In endometriosis, microecological imbalances within the gut microbiome trigger intestinal inflammation, leading to dysregulation of the immune response and the establishment of an immunosuppressive environment. This environment enables escaped endometrial cells to proliferate and grow outside the uterus, resulting in symptoms such as pain, tissue remodeling, fibrosis, adhesions, and infertility (Salmeri et al., 2024). Bailey and Coe (2002) reported a significant decrease in Lactobacillus counts and an increase in Gram-negative aerobic and facultative anaerobic bacteria in the feces of rhesus monkeys with endometriosis compared with healthy controls. Additionally, the incidence of intestinal inflammation was notably greater in endometriosis-affected monkeys than in control monkeys, suggesting that changes in the gut microbiota may be influenced by endometriosis-related intestinal inflammation (Bailey and Coe, 2002). Periodic bleeding from endometriosis provides a nutrient-rich environment for microorganisms, exacerbating pelvic inflammation and disease progression (Baker et al., 2017). Agostinis et al. (2020) demonstrated reduced T lymphocyte activity and expanded populations of macrophages, eosinophils, and neutrophils in endometriosis patients. Furthermore, abnormal activation of the complement system was detected in the peritoneal cavity of women with endometriosis. The regurgitated menstrual blood and peritoneal fluid of endometriosis patients contain relatively high levels of E. coli, whose endotoxins and LPS, as primary inflammatory mediators, are recognized and bound by Toll-like receptor 4 on macrophages and other immune cells. This interaction triggers the rapid secretion and release of secondary inflammatory mediators, fostering a pelvic inflammatory milieu that promotes inflammatory infiltration, hyperplasia, and angiogenesis in endometriosis (Khan et al., 2010).
4.2 Microbial ecology modulation of immune responses contributes to the onset and progression of endometriosis
Endometriosis is a disease characterized by immune response dysregulation, featuring perturbed polarization of pelvic macrophages. These macrophages aid in evading immune surveillance by ectopic endometrial cells, thereby facilitating the onset and progression of endometriosis (Khan et al., 2019). Campos et al. (2018) revealed that Mycoplasma genitalium can increase interferon-γ and interleukin-1β levels, modulating the local immune response in endometriosis patients. Studies have shown that endometriosis patients exhibit immune deficiencies and alterations in invasion within the endometrial microenvironment. LPS, through Toll-like receptor 4 signaling, is a significant factor contributing to epithelial–mesenchymal transition, a crucial process in endometriosis pathogenesis (Khan et al., 2018). The epithelial–mesenchymal transition plays a pivotal role in the ability of ectopic endometrial cells to successfully invade extrauterine organs and tissues, possibly related to its ability to diminish immune clearance mechanisms (Xu et al., 2020). Microbial dysbiosis impairs the immune clearance function of ectopic endometrial cells, leading to increased LPS levels that continuously activate the programmed death-1 (PD-1) pathway. This activation results in the overexpression of programmed death-1 and programmed death-ligand 1, causing T-cell exhaustion and immune tolerance. Consequently, the body’s immune defenses weaken, promoting infiltration of ectopic lesions, adhesions, and angiogenesis within the endometrial tissue, thus accelerating endometriosis progression (Feng et al., 2023; Santoso et al., 2020).
4.3 The influence of the microbiota on endometriosis through metabolic pathways
4.3.1 Increased estrogen metabolism
The gut microbiota microecology participates in the initiation and progression of endometriosis by modulating circulating estrogen levels (Laschke and Menger, 2016; Lu et al., 2017) (Figure 3). Baker et al. (2017) discovered that the gut microbiota is involved in intestinal estrogen metabolism, which is specifically related to β-glucuronidase, an enzyme that can deconjugate estrogens into their active forms. In cases of gut dysbiosis, when the Firmicutes/Bacteroidetes (F/B) ratio decreases, the secretion of intestinal β-glucuronidase increases, leading to elevated estrogen levels. These active estrogens can subsequently travel via the bloodstream to distant mucosal sites, including the endometrium (Wei et al., 2023). The collective genes of microorganisms capable of metabolizing estrogens within the gut are termed “estrobolomes,” which primarily encompass Bacteroides, Bifidobacteria, Escherichia coli, and Lactobacillus (Flores et al., 2012). In endometriosis research, the estrobolome hypothesis connects the crosstalk mechanism among endometriosis, estrogens, and microorganisms (Pai et al., 2023). Studies on the gut microbiota of endometriosis patients have identified more than 10 bacterial species whose intestinal profiles significantly differ. Additionally, these patients exhibit notably elevated plasma concentrations of estradiol and IL-8, along with an imbalanced ratio of Bacteroidetes to Firmicutes. The enrichment of Erysipelotrichaceae in fecal samples from endometriosis patients and significantly increased levels of estriol, 16-epiestriol, 16α-hydroxyestrone, and 2-methoxyestradiol have also been reported. Research has further indicated a correlation between the composition of the vaginal microbiota and age, with the 40–50 age group being particularly prone to dysbiosis. This age range coincides with the gradual decline in ovarian function and estrogen levels in women, suggesting a link between microorganisms and estrogen metabolism.
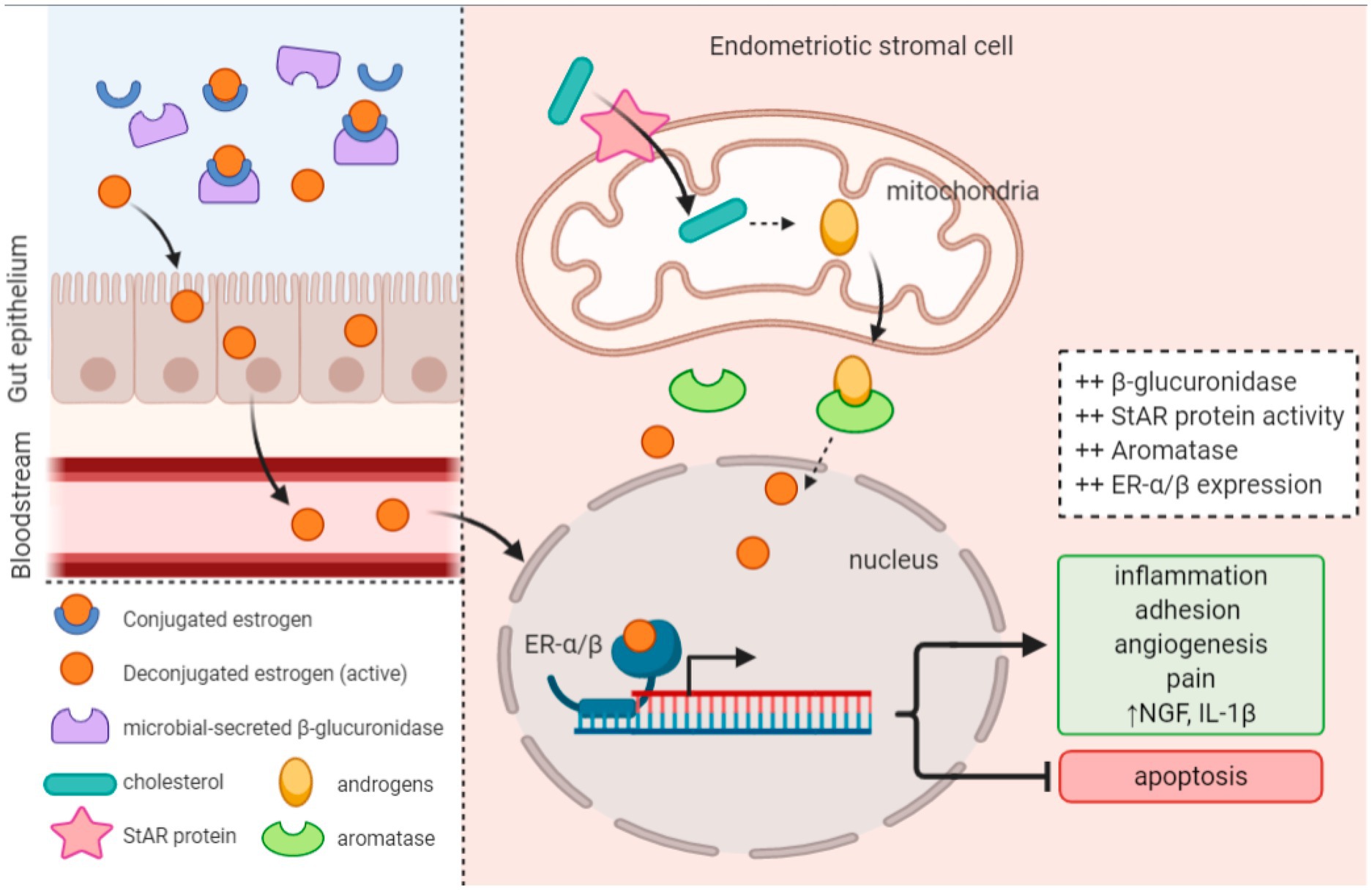
Figure 3. Altered estrobolome activity and upregulated enzyme expression produce a hyperestrogenic environment that promotes endometriosis onset and progression (Jiang et al., 2021).
4.3.2 Abnormal lipid metabolism
Analysis of KEGG pathways associated with the gut microbiota of endometriosis patients suggested enhanced tryptophan metabolism, α-linolenic acid metabolism, and ether lipid metabolism. Similarly, in endometriosis mouse models, abnormally expressed lipids such as phosphatidylcholine, sphingomyelin, phosphatidylethanolamine, and triglycerides were identified, with triglycerides closely linked to peritoneal inflammation. Ni et al. (2020) reported increased levels of chenodeoxycholic acid and ursodeoxycholic acid and a decreased abundance of α-linolenic acid in endometriosis mice. Exogenous supplementation with α-linolenic acid restored the abundance of Firmicutes and Bacteroidetes gut dysbiosis, reduced macrophage aggregation, and improved intestinal barrier function and the inflammatory environment in the abdominal cavity. Matta et al. (2022) reported imbalances in bile acid homeostasis and lipase activity in endometriosis patients, wherein disruptions in bile acid synthesis and metabolism can compromise intestinal barriers, fostering bacterial overgrowth and facilitating LPS entry into the bloodstream, thereby activating aberrant inflammatory responses. A study reported decreased abundances of Firmicutes taxa, such as Lachnospiraceae and Ruminococcaceae, in endometriosis patients, which normally produce short-chain fatty acids to modulate host physiology and immune function (Li et al., 2022). Notably, endometriosis mice presented significantly reduced concentrations of butyrate, isobutyrate, and valerate, with butyrate inhibiting ectopic endometrial growth and inflammatory cell infiltration. Butyrate suppresses the survival and lesion growth of human endometriotic cells through G protein-coupled receptors, histone deacetylases, and the GTPase-activating protein RAP1, thereby inhibiting human endometriosis.
4.4 Brain–gut axis
The brain–gut axis encompasses the intricate interplay between the gut microbiota, immune system, neuroendocrine system, and central nervous system (Mayer et al., 2022; Kesika et al., 2021). Disruptions in the gut microbiome can lead to elevated levels of neuroactive metabolites such as glutamate and gamma-aminobutyric acid, which stimulate brain neurons and stimulate estrogen secretion from the ovaries via the H-P-O axis (Liao et al., 2021). In turn, estrogen promotes nerve growth and differentiation, influencing pain perception through various mechanisms. Imbalances in the gut microbiota and disruptions in the brain–gut axis contribute to various types of pain (Yano et al., 2019). Research suggests that the gut microbiota may enhance central sensitization associated with chronic pain in endometriosis by modulating the activities of microglia, astrocytes, and immune cells, thereby intensifying patients’ pain perception (Ding et al., 2022). Moreover, gut microbiota dysbiosis can lead to neuronal death through oxidative stress and neuroinflammation, altering cognitive and immune functions in the body and accelerating the progression of endometriosis (Hu et al., 2023) (Figure 4).
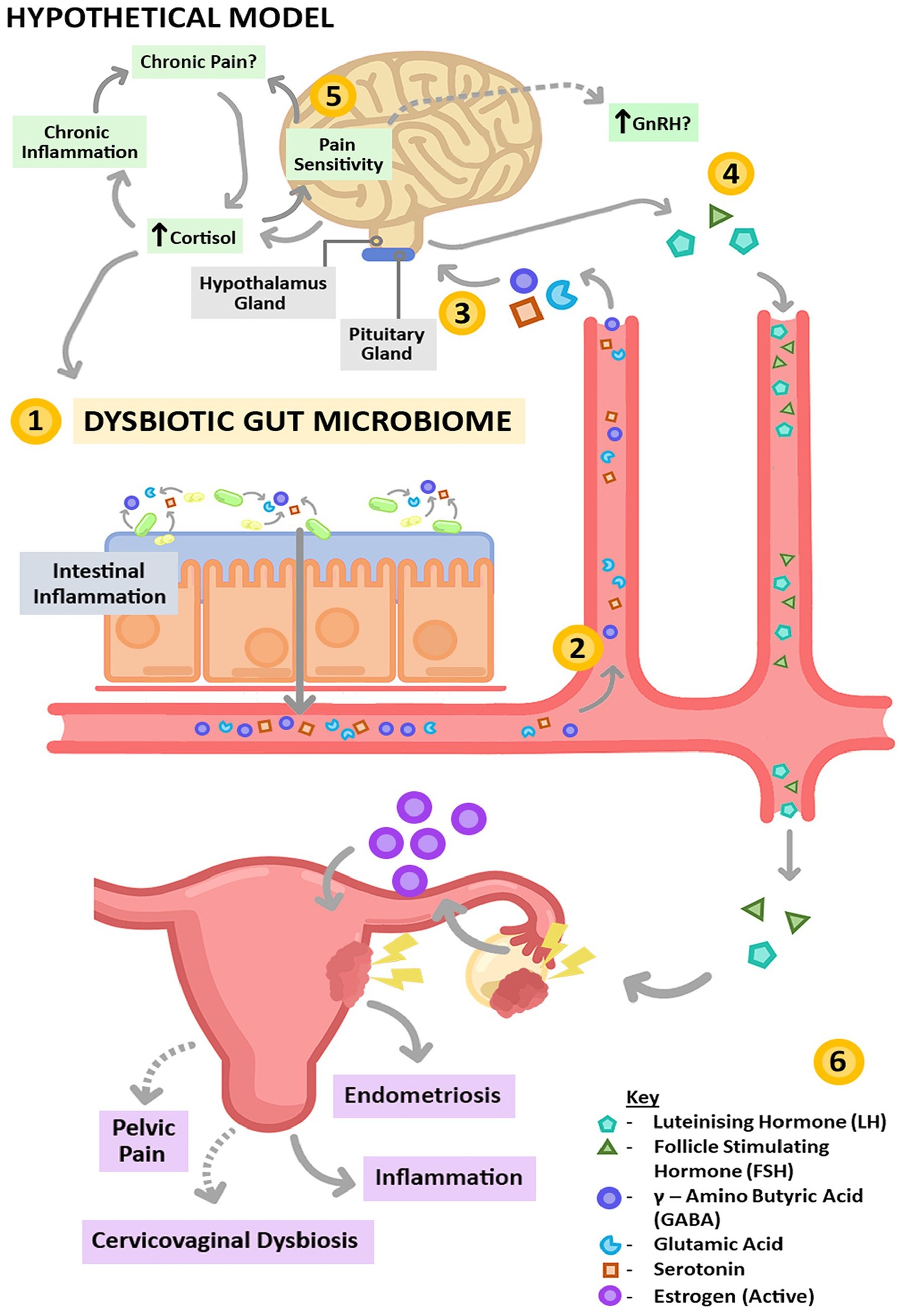
Figure 4. Brain–gut axis. Changes in neuroactive compound/protein production caused by gut dysbiosis can increase the level of GnRH (Salliss et al., 2021).
5 Clinical significance and future directions of the use of the microbiota in endometriosis
5.1 Biomarkers for non-invasive diagnosis
The search for reliable early, non-invasive biomarkers for biological diagnosis has emerged as a pivotal research focus in endometriosis studies (Uzuner et al., 2023). Perrotta et al. (2020) conducted an analysis of vaginal secretions and fecal samples from endometriosis patients and healthy controls during the follicular and menstrual phases and revealed that the Anaerococcus genus within the vaginal microbiome can potentially predict the stage of endometriosis (I/II or III/IV). Chen et al. (2020) developed a model based on the female genital tract microbiome, which can differentiate between infertility caused by endometriosis and other factors, suggesting that the vaginal or cervical microbiome may aid in detecting common upper genital tract diseases. Through screening and analysis of paired samples from feces, cervical mucus, and peritoneal fluid, Ruminococcus in the gut and Pseudomonas in the pelvic cavity were identified as potential biomarkers. A disease prediction model constructed on the basis of the core microbiota indicated that the gut microbiota outperforms the cervical mucus microbiota in the early diagnosis of endometriosis. Furthermore, the combination of the vaginal microbiota with CA125 offers a novel approach for discriminating endometriosis-related pelvic pain. Elevated serum irisin levels and reduced expression of the visfatin and angiogenin genes in endometriosis patients have been proposed as potential biomarkers for endometriosis (Kaya Sezginer et al., 2022; Slate-Romano et al., 2022). These studies underscore the potential of the microbiota as a crucial tool for early, non-invasive diagnosis or screening of endometriosis (Huang et al., 2021).
5.2 Potential approaches for microbiome-based therapies in endometriosis
5.2.1 Diet adjustment
The correlation between diet and the onset of endometriosis has been well established (Parazzini et al., 2013). Dietary interventions can impact the pathogenesis of endometriosis by modulating microbial-mediated metabolic alterations, reducing the release of proinflammatory factors, angiogenesis, and cell proliferation (Tomio et al., 2013). Women with high intakes of fruits, vegetables, dairy products, and fish and nuts rich in polyunsaturated fatty acids (PUFAs) have a decreased risk of developing endometriosis (Fu et al., 2021); conversely, increased consumption of products rich in trans fats, red meat, and alcohol is associated with an elevated risk of endometriosis (Missmer et al., 2010). The anti-inflammatory effects of diets high in ω-3 PUFAs, which are achieved by reducing TNF-α and IL-6 levels, contribute to lowering the risk of endometriosis among women (Yamamoto et al., 2018). This protective effect has also been validated in endometriosis animal models, where diets rich in omega-3 fatty acids and polyunsaturated fatty acids (PUFAs) exhibit anti-inflammatory properties and suppress endometriosis lesions (Attaman et al., 2014). Studies indicate that low-fat diets can increase the α diversity of the gut microbiota, whereas high-fat diets are linked to a reduction in Bacteroides and Coprobacillus species. These diet-induced alterations in the gut microbiota offer novel insights into the development of therapeutic strategies targeting gut bacterial metabolites for endometriosis treatment (Parpex et al., 2024).
5.2.2 Probiotics
Probiotics, as live microorganisms, can modulate the composition of the gut microbiota, thereby influencing β-glucuronidase activity within the body and subsequently regulating estrogen levels. Additionally, they contribute to maintaining immune system homeostasis by promoting anti-inflammatory responses and modulating immune cell activities, which in turn reduces the generation of inflammatory environments associated with endometriosis (You et al., 2022; Markowiak and Slizewska, 2017). Itoh et al. (2011a,b) reported that Lactobacillus gasseri OLL2809 inhibits the development of ectopic endometrial cells in the peritoneal cavity by activating NK cells. Kaczmarczyk et al. (2022) noted significant differences in the levels of IL-1 and IL-6 produced by peripheral blood mononuclear cells between endometriosis patients and healthy controls, whereas Lactobacillus acidophilus was found to induce the production of these cytokines. Furthermore, probiotics enhance neurotransmitter synthesis and signaling within the gut microbiota. By regulating neurotransmitter levels, they may influence pain pathways in endometriosis patients, mitigating pain perception and consequently affecting VAS scores. This modulation is attributed primarily to the regulation of immune responses involving IL-12 and NK cells. Research has shown that exogenous supplementation with the microbial metabolite unsaturated fatty acid alpha-linolenic acid improves the gut microbiota structure, dominant bacterial abundance, and intestinal barrier function in endometriosis mice. This, in turn, regulates intraperitoneal LPS levels and the inflammatory environment, thereby ameliorating endometriosis symptoms (Ni et al., 2021). Other studies have reported that oral Lactobacillus administration is beneficial for reducing the severity of dysmenorrhea, dyspareunia, and chronic pelvic pain among endometriosis patients (Ustianowska et al., 2022).
5.2.3 Microbial transplantation and antiestrogen therapy
Microbiota transplantation holds promise as a potential tool for future endometriosis treatment (Li et al., 2023; Talwar et al., 2025). Chadchan et al. (2019, 2023) reported that mice with gut microbiota depletion, achieved through broad-spectrum antibiotics or metronidazole treatment, presented significantly reduced endometriosis lesions and alleviated inflammation. When these mice were orally gavaged with feces from endometriosis, their endometriosis lesions and inflammation were restored. In contrast, gavage with feces from healthy mice failed to restore endometriosis lesions (Chadchan et al., 2023; Chadchan et al., 2019). Lu et al. (2022) compared the effects of vaginal microbiota transplantation and gonadotropin-releasing hormone agonist treatment in mice with endometriosis and reported that both approaches had nearly identical inhibitory effects on endometriosis lesion growth, suggesting that the vaginal microbiota can promote endometriosis development. While microbiota transplantation is a promising approach, challenges remain in standardizing the isolation, formulation, dosage, and timing of microbial administration to ensure optimal microbiota transplantation and maintenance for effective clinical responses (Takagi et al., 2019). Studies have shown that antiestrogen therapy increases the α-diversity of gut bacteria in endometriosis patients, and a positive correlation exists between gut bacterial species and urinary estrogen and its metabolite concentrations in endometriosis patients, indicating a link between the microbiota and estrogen metabolism in endometriosis (Le et al., 2021). Currently, one of the mechanisms for treating endometriosis involves inhibiting the H–P–O axis or antagonizing estrogen with progestins to reduce estrogen levels and suppress lesion growth (Rzewuska et al., 2023). However, these drugs are associated with side effects such as hypoestrogenism symptoms, irregular vaginal bleeding, and breast stimulation. If the mechanisms underlying the influence of the microbiota on estrogen metabolism can be elucidated, it may pave the way for the development of novel endometriosis treatment strategies (Quaranta et al., 2019) (Figure 5).
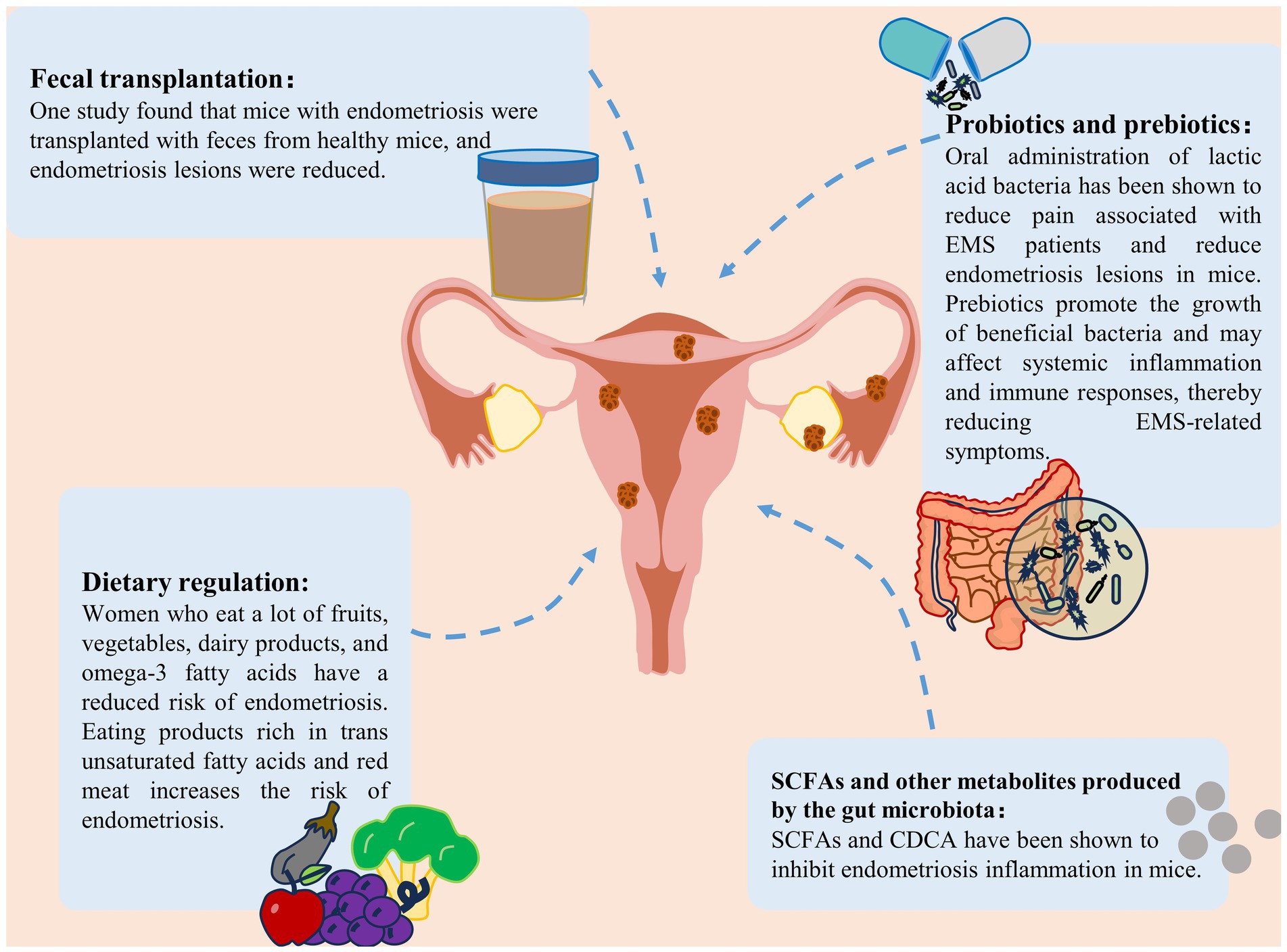
Figure 5. Treatment strategies for endometriosis. Treatment strategies for endometriosis include the use of probiotics and prebiotics, fecal bacterial transplantation, gut microbiota metabolite supplementation and dietary modification (Liu et al., 2024).
6 Conclusion
The human microbiome is intricately linked with endometriosis (Saunders and Horne, 2021; Tang et al., 2024) (Table 1). The microbiota can drive the onset and progression of endometriosis by modulating inflammatory responses, estrogen dynamics, metabolism, immune system function, and oxidative stress and increasing angiogenesis (Guo et al., 2024). Patients with endometriosis exhibit microbial imbalances in various body sites, which are strongly correlated with the pathogenesis of endometriosis. Conversely, the occurrence of endometriosis exacerbates microbial dysbiosis, creating a vicious cycle that accelerates disease progression. Breaking this cycle may yield breakthroughs in the diagnosis and treatment of endometriosis. Modulating microbial homeostasis through diet, medications, probiotics, or microbiota transplantation represents potential therapeutic avenues for endometriosis. Current research on the endometriosis microbiome has focused primarily on the gut microbiota because of its accessibility, diversity, and abundance. However, considering the unique physiological and pathological features of females, the cervical and vaginal microbiota, which are also representative, specific, and easily accessible to diverse microbial populations, merit further investigation. The discovery of microbiome-related biomarkers in the future could facilitate early, non-invasive diagnosis of endometriosis, thereby reducing delayed diagnoses. These findings have significant implications and vast potential for the prevention, diagnosis, and treatment of endometriosis.
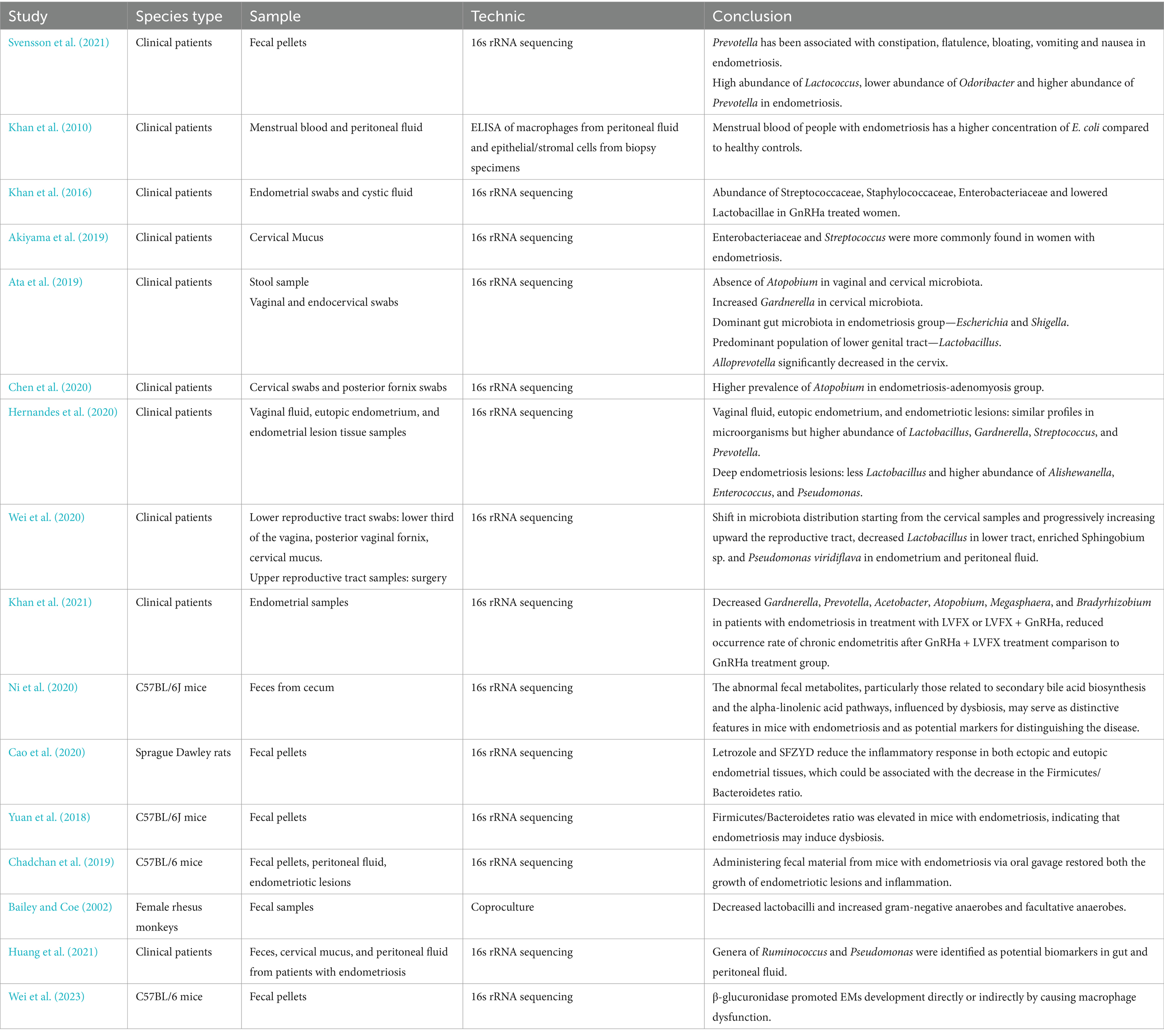
Table 1. A summary of recent literature on the microbiota and endometriosis.
Author contributions
MW: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. WL: Conceptualization, Data curation, Investigation, Methodology, Funding acquisition, Writing – review & editing. LZ: Conceptualization, Data curation, Writing – review & editing. SM: Data curation, Investigation, Writing – review & editing. LJ: Conceptualization, Data curation, Formal analysis, Investigation, Writing – review & editing. DZ: Conceptualization, Data curation, Validation, Visualization, Writing – review & editing. DL: Conceptualization, Data curation, Validation, Project administration, Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by Science and Technology Planning Project of Jilin Province (20230101198JC) and the Jilin Provincial Department of Finance’s Medical and Health Talent Project (2024WSZX-D12).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
LPS, lipopolysaccharides; F/B, Firmicutes-to-Bacteroidetes; PUFAs, polyunsaturated fatty acids.
References
Adnane, M., and Chapwanya, A. (2022). Role of genital tract Bacteria in promoting endometrial health in cattle. Microorganisms 10:2238. doi: 10.3390/microorganisms10112238
Aggarwal, N., Kitano, S., Puah, G. R. Y., Kittelmann, S., Hwang, I. Y., and Chang, M. W. (2023). Microbiome and human health: current understanding, engineering, and enabling technologies. Chem. Rev. 123, 31–72. doi: 10.1021/acs.chemrev.2c00431
Agostinis, C., Balduit, A., Mangogna, A., Zito, G., Romano, F., Ricci, G., et al. (2020). Immunological basis of the endometriosis: the complement system as a potential therapeutic target. Front. Immunol. 11:599117. doi: 10.3389/fimmu.2020.599117
Akiyama, K., Nishioka, K., Khan, K. N., Tanaka, Y., Mori, T., Nakaya, T., et al. (2019). Molecular detection of microbial colonization in cervical mucus of women with and without endometriosis. Am. J. Reprod. Immunol. 82:e13147. doi: 10.1111/aji.13147
Amro, B., Ramirez Aristondo, M. E., Alsuwaidi, S., Almaamari, B., Hakim, Z., Tahlak, M., et al. (2022). New understanding of diagnosis, treatment and prevention of endometriosis. Int. J. Environ. Res. Public Health 19:6725. doi: 10.3390/ijerph19116725
Arion, K., Orr, N. L., Noga, H., Allaire, C., Williams, C., Bedaiwy, M. A., et al. (2020). A quantitative analysis of sleep quality in women with endometriosis. J Womens Health 29, 1209–1215. doi: 10.1089/jwh.2019.8008
Ata, B., Yildiz, S., Turkgeldi, E., Brocal, V. P., Dinleyici, E. C., Moya, A., et al. (2019). The Endobiota study: comparison of vaginal, cervical and gut microbiota between women with stage 3/4 endometriosis and healthy controls. Sci. Rep. 9:2204. doi: 10.1038/s41598-019-39700-6
Attaman, J. A., Stanic, A. K., Kim, M., Lynch, M. P., Rueda, B. R., and Styer, A. K. (2014). The anti-inflammatory impact of omega-3 polyunsaturated fatty acids during the establishment of endometriosis-like lesions. Am. J. Reprod. Immunol. 72, 392–402. doi: 10.1111/aji.12276
Bailey, M. T., and Coe, C. L. (2002). Endometriosis is associated with an altered profile of intestinal microflora in female rhesus monkeys. Hum. Reprod. 17, 1704–1708. doi: 10.1093/humrep/17.7.1704
Baker, J. M., Al-Nakkash, L., and Herbst-Kralovetz, M. M. (2017). Estrogen-gut microbiome axis: physiological and clinical implications. Maturitas 103, 45–53. doi: 10.1016/j.maturitas.2017.06.025
Bedaiwy, M. A., Falcone, T., Sharma, R. K., Goldberg, J. M., Attaran, M., Nelson, D. R., et al. (2002). Prediction of endometriosis with serum and peritoneal fluid markers: a prospective controlled trial. Hum. Reprod. 17, 426–431. doi: 10.1093/humrep/17.2.426
Berg, G., Rybakova, D., Fischer, D., Cernava, T., Verges, M. C., Charles, T., et al. (2020). Microbiome definition re-visited: old concepts and new challenges. Microbiome. 8:103. doi: 10.1186/s40168-020-00875-0
Campos, G. B., Marques, L. M., Rezende, I. S., Barbosa, M. S., Abrao, M. S., and Timenetsky, J. (2018). Mycoplasma genitalium can modulate the local immune response in patients with endometriosis. Fertil. Steril. 109, 549–560.e4. doi: 10.1016/j.fertnstert.2017.11.009
Cao, Y., Jiang, C., Jia, Y., Xu, D., and Yu, Y. (2020). Letrozole and the traditional Chinese medicine, Shaofu Zhuyu decoction, reduce Endometriotic disease progression in rats: a potential role for gut microbiota. Evid. Based Complement. Alternat. Med. 2020:3687498. doi: 10.1155/2020/3687498
Chadchan, S. B., Cheng, M., Parnell, L. A., Yin, Y., Schriefer, A., Mysorekar, I. U., et al. (2019). Antibiotic therapy with metronidazole reduces endometriosis disease progression in mice: a potential role for gut microbiota. Hum. Reprod. 34, 1106–1116. doi: 10.1093/humrep/dez041
Chadchan, S. B., Naik, S. K., Popli, P., Talwar, C., Putluri, S., Ambati, C. R., et al. (2023). Gut microbiota and microbiota-derived metabolites promotes endometriosis. Cell Death Discov. 9:28. doi: 10.1038/s41420-023-01309-0
Chang, C. Y., Chiang, A. J., Lai, M. T., Yan, M. J., Tseng, C. C., Lo, L. C., et al. (2022). A more diverse cervical microbiome associates with better clinical outcomes in patients with endometriosis: a pilot study. Biomedicines 10:174. doi: 10.3390/biomedicines10010174
Chapron, C., Marcellin, L., Borghese, B., and Santulli, P. (2019). Rethinking mechanisms, diagnosis and management of endometriosis. Nat. Rev. Endocrinol. 15, 666–682. doi: 10.1038/s41574-019-0245-z
Chen, S., Gu, Z., Zhang, W., Jia, S., Wu, Y., Zheng, P., et al. (2020). Microbiome of the lower genital tract in Chinese women with endometriosis by 16s-rRNA sequencing technique: a pilot study. Ann Transl Med. 8:1440. doi: 10.21037/atm-20-1309
Cohen Ben-Meir, L., Soriano, D., Zajicek, M., Yulzari, V., Bouaziz, J., Beer-Gabel, M., et al. (2022). The association between gastrointestinal symptoms and transvaginal ultrasound findings in women referred for endometriosis evaluation: a prospective pilot study. Ultraschall Med. 43, e81–e89. doi: 10.1055/a-1300-1887
Dicks, L. M. T. (2022). Gut Bacteria and neurotransmitters. Microorganisms 10:9. doi: 10.3390/microorganisms10091838
Ding, X., Zhou, J., Zhao, L., Chen, M., Wang, S., Zhang, M., et al. (2022). Intestinal Flora composition determines microglia activation and improves epileptic episode Progress. Front. Cell. Infect. Microbiol. 12:835217. doi: 10.3389/fcimb.2022.835217
Fan, P., and Li, T. (2022). Unveil the pain of endometriosis: from the perspective of the nervous system. Expert Rev. Mol. Med. 24:e36. doi: 10.1017/erm.2022.26
Feng, D., Wang, X., Huang, Y., Chen, X., Xu, R., Qu, Z., et al. (2023). Warming menstruation and analgesic soup inhibits PD-1/PD-L1 pathway in rats with endometriosis. Altern. Ther. Health Med. 29, 152–157.
Flores, R., Shi, J., Fuhrman, B., Xu, X., Veenstra, T. D., Gail, M. H., et al. (2012). Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: a cross-sectional study. J. Transl. Med. 10:253. doi: 10.1186/1479-5876-10-253
Franca, P. R. C., Lontra, A. C. P., and Fernandes, P. D. (2022). Endometriosis: a disease with few direct treatment options. Molecules 27:4034. doi: 10.3390/molecules27134034
Fu, Y., Wang, Y., Gao, H., Li, D., Jiang, R., Ge, L., et al. (2021). Associations among dietary Omega-3 polyunsaturated fatty acids, the gut microbiota, and intestinal immunity. Mediat. Inflamm. 2021, 1–11. doi: 10.1155/2021/8879227
Gao, J., Peng, Y., Jiang, N., Shi, Y., and Ying, C. (2022). High-throughput sequencing-based analysis of changes in the vaginal microbiome during the disease course of patients with bacterial vaginosis: a case-control study. Biology 11:1797. doi: 10.3390/biology11121797
Guo, J., Yan, X., and Han, L. (2024). The potential role of gut microbiota in the occurrence and development of endometriosis. Front. Cell. Infect. Microbiol. 14:1454931. doi: 10.3389/fcimb.2024.1454931
He, G., Chen, J., Peng, Z., Feng, K., Luo, C., and Zeng, X. (2022). A study on the correlation between quality of life and unhealthy emotion among patients with endometriosis. Front. Psychol. 13:830698. doi: 10.3389/fpsyg.2022.830698
Hernandes, C., Silveira, P., Rodrigues Sereia, A. F., Christoff, A. P., Mendes, H., Valter de Oliveira, L. F., et al. (2020). Microbiome profile of deep endometriosis patients: comparison of vaginal fluid, endometrium and lesion. Diagnostics 10:163. doi: 10.3390/diagnostics10030163
Hicks, C., Leonardi, M., Chua, X. Y., Mari-Breedt, L., Espada, M., El-Omar, E. M., et al. (2025). Oral, vaginal, and stool microbial signatures in patients with endometriosis as potential diagnostic non-invasive biomarkers: a prospective cohort study. BJOG 132, 326–336. doi: 10.1111/1471-0528.17979
Horne, A. W., and Missmer, S. A. (2022). Pathophysiology, diagnosis, and management of endometriosis. BMJ 379:e070750. doi: 10.1136/bmj-2022-070750
Hu, J., Li, G., Huang, M., Shen, Q., Gu, H., Xue, N., et al. (2023). Analysis of microbiota reveals the underlying mechanism of PHF11 in the development of Enterococcus-regulated endometriotic cysts. iScience. 26:108158. doi: 10.1016/j.isci.2023.108158
Hu, Z., Zhao, P., Liao, A., Pan, L., Zhang, J., Dong, Y., et al. (2023). Fermented wheat germ alleviates depression-like behavior in rats with chronic and unpredictable mild stress. Food Secur. 12:920. doi: 10.3390/foods12050920
Huang, L., Liu, B., Liu, Z., Feng, W., Liu, M., Wang, Y., et al. (2021). Gut microbiota exceeds cervical microbiota for early diagnosis of endometriosis. Front. Cell. Infect. Microbiol. 11:788836. doi: 10.3389/fcimb.2021.788836
Itoh, H., Sashihara, T., Hosono, A., Kaminogawa, S., and Uchida, M. (2011a). Lactobacillus gasseri OLL2809 inhibits development of ectopic endometrial cell in peritoneal cavity via activation of NK cells in a murine endometriosis model. Cytotechnology 63, 205–210. doi: 10.1007/s10616-011-9343-z
Itoh, H., Uchida, M., Sashihara, T., Ji, Z. S., Li, J., Tang, Q., et al. (2011b). Lactobacillus gasseri OLL2809 is effective especially on the menstrual pain and dysmenorrhea in endometriosis patients: randomized, double-blind, placebo-controlled study. Cytotechnology 63, 153–161. doi: 10.1007/s10616-010-9326-5
Jiang, I., Yong, P. J., Allaire, C., and Bedaiwy, M. A. (2021). Intricate connections between the microbiota and endometriosis. Int. J. Mol. Sci. 22:5644. doi: 10.3390/ijms22115644
Kaczmarczyk, M., Szulinska, M., Loniewski, I., Kregielska-Narozna, M., Skonieczna-Zydecka, K., Kosciolek, T., et al. (2022). Treatment with multi-species probiotics changes the functions, not the composition of gut microbiota in postmenopausal women with obesity: a randomized, double-blind, placebo-controlled study. Front Cell Infect Microbiol. 12:815798. doi: 10.3389/fcimb.2022.815798
Kaya Sezginer, E., Kirlangic OFEskin Tanriverdi, M. D., Topcu, H. O., and Gur, S. (2022). Analysis of changes in serum levels and gene expression profiles of novel Adipocytokines (Omentin, Vaspin, Irisin and Visfatin) and their correlation with serum C-reactive protein levels in women diagnosed with endometriosis. Turk J Pharm Sci. 19, 48–53. doi: 10.4274/tjps.galenos.2021.52284
Kesika, P., Suganthy, N., Sivamaruthi, B. S., and Chaiyasut, C. (2021). Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer's disease. Life Sci. 264:118627. doi: 10.1016/j.lfs.2020.118627
Khan, K. N., Fujishita, A., Hiraki, K., Kitajima, M., Nakashima, M., Fushiki, S., et al. (2018). Bacterial contamination hypothesis: a new concept in endometriosis. Reprod Med Biol. 17, 125–133. doi: 10.1002/rmb2.12083
Khan, K. N., Fujishita, A., Kitajima, M., Hiraki, K., Nakashima, M., and Masuzaki, H. (2014). Intra-uterine microbial colonization and occurrence of endometritis in women with endometriosis dagger. Hum. Reprod. 29, 2446–2456. doi: 10.1093/humrep/deu222
Khan, K. N., Fujishita, A., Koshiba, A., Kuroboshi, H., Mori, T., Ogi, H., et al. (2019). Biological differences between intrinsic and extrinsic adenomyosis with coexisting deep infiltrating endometriosis. Reprod. Biomed. Online 39, 343–353. doi: 10.1016/j.rbmo.2019.03.210
Khan, K. N., Fujishita, A., Masumoto, H., Muto, H., Kitajima, M., Masuzaki, H., et al. (2016). Molecular detection of intrauterine microbial colonization in women with endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 199, 69–75. doi: 10.1016/j.ejogrb.2016.01.040
Khan, K. N., Fujishita, A., Muto, H., Masumoto, H., Ogawa, K., Koshiba, A., et al. (2021). Levofloxacin or gonadotropin releasing hormone agonist treatment decreases intrauterine microbial colonization in human endometriosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 264, 103–116. doi: 10.1016/j.ejogrb.2021.07.014
Khan, K. N., Kitajima, M., Hiraki, K., Yamaguchi, N., Katamine, S., Matsuyama, T., et al. (2010). Escherichia coli contamination of menstrual blood and effect of bacterial endotoxin on endometriosis. Fertil. Steril. 94, 2860–2863.e3. doi: 10.1016/j.fertnstert.2010.04.053
Kitaya, K., and Yasuo, T. (2023). Commonalities and disparities between endometriosis and chronic Endometritis: therapeutic potential of novel antibiotic treatment strategy against ectopic endometrium. Int. J. Mol. Sci. 24:2059. doi: 10.3390/ijms24032059
Kvaskoff, M., Mahamat-Saleh, Y., Farland, L. V., Shigesi, N., Terry, K. L., Harris, H. R., et al. (2021). Endometriosis and cancer: a systematic review and meta-analysis. Hum. Reprod. Update 27, 393–420. doi: 10.1093/humupd/dmaa045
Lamceva, J., Uljanovs, R., and Strumfa, I. (2023). The Main theories on the pathogenesis of endometriosis. Int. J. Mol. Sci. 24:4254. doi: 10.3390/ijms24054254
Laschke, M. W., and Menger, M. D. (2016). The gut microbiota: a puppet master in the pathogenesis of endometriosis? Am. J. Obstet. Gynecol. 215:68.e1. doi: 10.1016/j.ajog.2016.02.036
Le, N., Cregger, M., Brown, V., Loret de Mola, J., Bremer, P., Nguyen, L., et al. (2021). Association of microbial dynamics with urinary estrogens and estrogen metabolites in patients with endometriosis. PLoS One 16:e0261362. doi: 10.1371/journal.pone.0261362
Lee, S. R., Lee, J. C., Kim, S. H., Oh, Y. S., Chae, H. D., Seo, H., et al. (2021). Altered composition of microbiota in women with ovarian Endometrioma: microbiome analyses of extracellular vesicles in the peritoneal fluid. Int. J. Mol. Sci. 22:4608. doi: 10.3390/ijms22094608
Leonardi, M., Hicks, C., El-Assaad, F., El-Omar, E., and Condous, G. (2020). Endometriosis and the microbiome: a systematic review. BJOG 127, 239–249. doi: 10.1111/1471-0528.15916
Li, Y., Wang, K., Ding, J., Sun, S., Ni, Z., and Yu, C. (2022). Influence of the gut microbiota on endometriosis: potential role of chenodeoxycholic acid and its derivatives. Front. Pharmacol. 13:954684. doi: 10.3389/fphar.2022.954684
Li, M., Yang, H., Shao, C., Liu, Y., Wen, S., and Tang, L. (2023). Application of dominant gut microbiota promises to replace fecal microbiota transplantation as a new treatment for Alzheimer's disease. Microorganisms 11:2854. doi: 10.3390/microorganisms11122854
Liao, B., Qiao, J., and Pang, Y. (2021). Central regulation of PCOS: abnormal neuronal-reproductive-metabolic circuits in PCOS pathophysiology. Front Endocrinol 12:667422. doi: 10.3389/fendo.2021.667422
Liu, H., Liang, H., Li, D., Wang, M., and Li, Y. (2022). Association of Cervical Dysbacteriosis, HPV oncogene expression, and cervical lesion progression. Microbiol Spectr. 10:e0015122. doi: 10.1128/spectrum.00151-22
Liu, M., Peng, R., Tian, C., Shi, J., Ma, J., Shi, R., et al. (2024). Effects of the gut microbiota and its metabolite short-chain fatty acids on endometriosis. Front. Cell. Infect. Microbiol. 14:1373004. doi: 10.3389/fcimb.2024.1373004
Lu, N., Li, M., Lei, H., Jiang, X., Tu, W., Lu, Y., et al. (2017). Butyric acid regulates progesterone and estradiol secretion via cAMP signaling pathway in porcine granulosa cells. J. Steroid Biochem. Mol. Biol. 172, 89–97. doi: 10.1016/j.jsbmb.2017.06.004
Lu, F., Wei, J., Zhong, Y., Feng, Y., Ma, B., Xiong, Y., et al. (2022). Antibiotic therapy and vaginal microbiota transplantation reduce endometriosis disease progression in female mice via NF-kappa B signaling pathway. Front Med 9:831115. doi: 10.3389/fmed.2022.831115
Mahajan, G., Doherty, E., To, T., Sutherland, A., Grant, J., Junaid, A., et al. (2022). Vaginal microbiome-host interactions modeled in a human vagina-on-a-chip. Microbiome. 10:201. doi: 10.1186/s40168-022-01400-1
Markowiak, P., and Slizewska, K. (2017). Effects of probiotics, prebiotics, and Synbiotics on human health. Nutrients 9:1021. doi: 10.3390/nu9091021
Matta, K., Lefebvre, T., Vigneau, E., Cariou, V., Marchand, P., Guitton, Y., et al. (2022). Associations between persistent organic pollutants and endometriosis: a multiblock approach integrating metabolic and cytokine profiling. Environ. Int. 158:106926.
Mayer, E. A., Nance, K., and Chen, S. (2022). The gut-brain Axis. Annu. Rev. Med. 73, 439–453. doi: 10.1146/annurev-med-042320-014032
Mazur-Bialy, A. I., Pochec, E., and Zarawski, M. (2017). Anti-inflammatory properties of Irisin, mediator of physical activity, are connected with TLR4/MyD88 signaling pathway activation. Int. J. Mol. Sci. 18:701. doi: 10.3390/ijms18040701
Medina-Bastidas, D., Camacho-Arroyo, I., and Garcia-Gomez, E. (2022). Current findings in endometrial microbiome: impact on uterine diseases. Reproduction 163, R81–R96. doi: 10.1530/REP-21-0120
Missmer, S. A., Chavarro, J. E., Malspeis, S., Bertone-Johnson, E. R., Hornstein, M. D., Spiegelman, D., et al. (2010). A prospective study of dietary fat consumption and endometriosis risk. Hum. Reprod. 25, 1528–1535. doi: 10.1093/humrep/deq044
Molina, N. M., Sola-Leyva, A., Saez-Lara, M. J., Plaza-Diaz, J., Tubic-Pavlovic, A., Romero, B., et al. (2020). New opportunities for endometrial health by modifying uterine microbial composition: present or future? Biomol. Ther. 10:593. doi: 10.3390/biom10040593
Ni, Z., Ding, J., Zhao, Q., Cheng, W., Yu, J., Zhou, L., et al. (2021). Alpha-linolenic acid regulates the gut microbiota and the inflammatory environment in a mouse model of endometriosis. Am. J. Reprod. Immunol. 86:e13471. doi: 10.1111/aji.13471
Ni, Z., Sun, S., Bi, Y., Ding, J., Cheng, W., Yu, J., et al. (2020). Correlation of fecal metabolomics and gut microbiota in mice with endometriosis. Am. J. Reprod. Immunol. 84:e13307. doi: 10.1111/aji.13307
Oishi, S., Mekaru, K., Tanaka, S. E., Arai, W., Ashikawa, K., Sakuraba, Y., et al. (2022). Microbiome analysis in women with endometriosis: does a microbiome exist in peritoneal fluid and ovarian cystic fluid? Reprod Med Biol. 21:e12441. doi: 10.1002/rmb2.12441
Onywera, H., Williamson, A. L., Mbulawa, Z. Z. A., Coetzee, D., and Meiring, T. L. (2019). Factors associated with the composition and diversity of the cervical microbiota of reproductive-age black south African women: a retrospective cross-sectional study. Peer J. 7:e7488. doi: 10.7717/peerj.7488
Opoien, H. K., Fedorcsak, P., Polec, A., Stensen, M. H., Abyholm, T., and Tanbo, T. (2013). Do endometriomas induce an inflammatory reaction in nearby follicles? Hum. Reprod. 28, 1837–1845. doi: 10.1093/humrep/det087
Pai, A. H., Wang, Y. W., Lu, P. C., Wu, H. M., Xu, J. L., and Huang, H. Y. (2023). Gut microbiome-Estrobolome profile in reproductive-age women with endometriosis. Int. J. Mol. Sci. 24:24 (22). doi: 10.3390/ijms242216301
Parazzini, F., Vigano, P., Candiani, M., and Fedele, L. (2013). Diet and endometriosis risk: a literature review. Reprod. Biomed. Online 26, 323–336. doi: 10.1016/j.rbmo.2012.12.011
Parpex, G., Chassaing, B., Bourdon, M., Santulli, P., Doridot, L., Thomas, M., et al. (2024). Western diet promotes endometriotic lesion growth in mice and induces depletion of Akkermansia muciniphila in intestinal microbiota. BMC Med. 22:513. doi: 10.1186/s12916-024-03738-9
Pelzer, E. S., Willner, D., Buttini, M., Hafner, L. M., Theodoropoulos, C., and Huygens, F. (2018). The fallopian tube microbiome: implications for reproductive health. Oncotarget 9, 21541–21551. doi: 10.18632/oncotarget.25059
Perrotta, A. R., Borrelli, G. M., Martins, C. O., Kallas, E. G., Sanabani, S. S., Griffith, L. G., et al. (2020). The vaginal microbiome as a tool to predict rASRM stage of disease in endometriosis: a pilot study. Reprod. Sci. 27, 1064–1073. doi: 10.1007/s43032-019-00113-5
Quaranta, G., Sanguinetti, M., and Masucci, L. (2019). Fecal microbiota transplantation: a potential tool for treatment of human female reproductive tract diseases. Front. Immunol. 10:2653. doi: 10.3389/fimmu.2019.02653
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S., McCulle, S. L., et al. (2011). Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 108, 4680–4687. doi: 10.1073/pnas.1002611107
Rzewuska, A. M., Zybowska, M., Sajkiewicz, I., Spiechowicz, I., Zak, K., Abramiuk, M., et al. (2023). Gonadotropin-releasing hormone antagonists-a new Hope in endometriosis treatment? J. Clin. Med. 12:1008. doi: 10.3390/jcm12031008
Salliss, M. E., Farland, L. V., Mahnert, N. D., and Herbst-Kralovetz, M. M. (2021). The role of gut and genital microbiota and the estrobolome in endometriosis, infertility and chronic pelvic pain. Hum. Reprod. Update 28, 92–131. doi: 10.1093/humupd/dmab035
Salmeri, N., Vigano, P., Cavoretto, P., Marci, R., and Candiani, M. (2024). The kisspeptin system in and beyond reproduction: exploring intricate pathways and potential links between endometriosis and polycystic ovary syndrome. Rev. Endocr. Metab. Disord. 25, 239–257. doi: 10.1007/s11154-023-09826-0
Santoso, B., Sa'adi, A., Dwiningsih, S. R., Tunjungseto, A., Widyanugraha, M. Y. A., Mufid, A. F., et al. (2020). Soluble immune checkpoints CTLA-4, HLA-G, PD-1, and PD-L1 are associated with endometriosis-related infertility. Am. J. Reprod. Immunol. 84:e13296. doi: 10.1111/aji.13296
Saunders, P. T. K., and Horne, A. W. (2021). Endometriosis: etiology, pathobiology, and therapeutic prospects. Cell 184, 2807–2824. doi: 10.1016/j.cell.2021.04.041
Schenken, R. S., Asch, R. H., Williams, R. F., and Hodgen, G. D. (1984). Etiology of infertility in monkeys with endometriosis: luteinized unruptured follicles, luteal phase defects, pelvic adhesions, and spontaneous abortions. Fertil. Steril. 41, 122–130. doi: 10.1016/S0015-0282(16)47552-7
Ser, H. L., Au Yong, S. J., Shafiee, M. N., Mokhtar, N. M., and Ali, R. A. R. (2023). Current updates on the role of microbiome in endometriosis: a narrative review. Microorganisms. 11:360. doi: 10.3390/microorganisms11020360
Shabbir, U., Arshad, M. S., Sameen, A., and Oh, D. H. (2021). Crosstalk between gut and brain in Alzheimer's disease: the role of gut microbiota modulation strategies. Nutrients 13:690. doi: 10.3390/nu13020690
Shan, J., Ni, Z., Cheng, W., Zhou, L., Zhai, D., Sun, S., et al. (2021). Gut microbiota imbalance and its correlations with hormone and inflammatory factors in patients with stage 3/4 endometriosis. Arch. Gynecol. Obstet. 304, 1363–1373. doi: 10.1007/s00404-021-06057-z
Slate-Romano, J. J., Yano, N., and Zhao, T. C. (2022). Irisin reduces inflammatory signaling pathways in inflammation-mediated metabolic syndrome. Mol. Cell. Endocrinol. 552:111676. doi: 10.1016/j.mce.2022.111676
Slocum, C., Kramer, C., and Genco, C. A. (2016). Immune dysregulation mediated by the oral microbiome: potential link to chronic inflammation and atherosclerosis. J. Intern. Med. 280, 114–128. doi: 10.1111/joim.12476
Svensson, A., Brunkwall, L., Roth, B., Orho-Melander, M., and Ohlsson, B. (2021). Associations between endometriosis and gut microbiota. Reprod. Sci. 28, 2367–2377. doi: 10.1007/s43032-021-00506-5
Takagi, T., Naito, Y., Inoue, R., Kashiwagi, S., Uchiyama, K., Mizushima, K., et al. (2019). Differences in gut microbiota associated with age, sex, and stool consistency in healthy Japanese subjects. J. Gastroenterol. 54, 53–63. doi: 10.1007/s00535-018-1488-5
Talwar, C., Davuluri, G. V. N., Kamal, A. H. M., Coarfa, C., Han, S. J., Veeraragavan, S., et al. (2025). Identification of distinct stool metabolites in women with endometriosis for non-invasive diagnosis and potential for microbiota-based therapies. Med 6:100517. doi: 10.1016/j.medj.2024.09.006
Tang, Y., Yang, J., Hang, F., Huang, H., and Jiang, L. (2024). Unraveling the relationship between gut microbiota and site-specific endometriosis: a Mendelian randomization analysis. Front. Microbiol. 15:1363080. doi: 10.3389/fmicb.2024.1363080
Taylor, H. S., Kotlyar, A. M., and Flores, V. A. (2021). Endometriosis is a chronic systemic disease: clinical challenges and novel innovations. Lancet 397, 839–852. doi: 10.1016/S0140-6736(21)00389-5
Terzic, M., Aimagambetova, G., Kunz, J., Bapayeva, G., Aitbayeva, B., Terzic, S., et al. (2021). Molecular basis of endometriosis and endometrial Cancer: current knowledge and future perspectives. Int. J. Mol. Sci. 22:9274. doi: 10.3390/ijms22179274
Tomio, K., Kawana, K., Taguchi, A., Isobe, Y., Iwamoto, R., Yamashita, A., et al. (2013). Omega-3 polyunsaturated fatty acids suppress the cystic lesion formation of peritoneal endometriosis in transgenic mouse models. PLoS One 8:e73085. doi: 10.1371/journal.pone.0073085
Trompette, A., Gollwitzer, E. S., Yadava, K., Sichelstiel, A. K., Sprenger, N., Ngom-Bru, C., et al. (2014). Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 20, 159–166. doi: 10.1038/nm.3444
Ustianowska, K., Ustianowski, L., Machaj, F., Goracy, A., Rosik, J., Szostak, B., et al. (2022). The role of the human microbiome in the pathogenesis of pain. Int. J. Mol. Sci. 23:3267. doi: 10.3390/ijms232113267
Uzuner, C., Mak, J., El-Assaad, F., and Condous, G. (2023). The bidirectional relationship between endometriosis and microbiome. Front Endocrinol 14:1110824. doi: 10.3389/fendo.2023.1110824
Ventolini, G., Vieira-Baptista, P., De Seta, F., Verstraelen, H., Lonnee-Hoffmann, R., and Lev-Sagie, A. (2022). The vaginal microbiome: IV. The role of vaginal microbiome in reproduction and in gynecologic cancers. J. Low. Genit. Tract Dis. 26, 93–98. doi: 10.1097/LGT.0000000000000646
Wang, J., Li, Z., Ma, X., Du, L., Jia, Z., Cui, X., et al. (2021). Translocation of vaginal microbiota is involved in impairment and protection of uterine health. Nat. Commun. 12:4191. doi: 10.1038/s41467-021-24516-8
Wang, S., Shang, Y., Pi, Z., Zhou, Z., Zhang, X., Ren, L., et al. (2023). Temporal changes of the Oral and fecal microbiota after mild traumatic brain injury in rats by 16S rRNA sequencing. Microorganisms 11:1462. doi: 10.3390/microorganisms11061452
Wang, L., Yang, J., Su, H., Shi, L., Chen, B., and Zhang, S. (2022). Endometrial microbiota from endometrial cancer and paired pericancer tissues in postmenopausal women: differences and clinical relevance. Menopause 29, 1168–1175. doi: 10.1097/GME.0000000000002053
Wei, Y., Tan, H., Yang, R., Yang, F., Liu, D., Huang, B., et al. (2023). Gut dysbiosis-derived beta-glucuronidase promotes the development of endometriosis. Fertil. Steril. 120, 682–694. doi: 10.1016/j.fertnstert.2023.03.032
Wei, W., Zhang, X., Tang, H., Zeng, L., and Wu, R. (2020). Microbiota composition and distribution along the female reproductive tract of women with endometriosis. Ann. Clin. Microbiol. Antimicrob. 19:15. doi: 10.1186/s12941-020-00356-0
Wessels, J. M., Dominguez, M. A., Leyland, N. A., Agarwal, S. K., and Foster, W. G. (2021). Endometrial microbiota is more diverse in people with endometriosis than symptomatic controls. Sci. Rep. 11:18877. doi: 10.1038/s41598-021-98380-3
Witkin, S. S., and Linhares, I. M. (2017). Why do lactobacilli dominate the human vaginal microbiota? BJOG 124, 606–611. doi: 10.1111/1471-0528.14390
Xu, H., Zhao, J., Lu, J., and Sun, X. (2020). Ovarian endometrioma infiltrating neutrophils orchestrate immunosuppressive microenvironment. J. Ovarian Res. 13:44. doi: 10.1186/s13048-020-00642-7
Yamamoto, A., Harris, H. R., Vitonis, A. F., Chavarro, J. E., and Missmer, S. A. (2018). A prospective cohort study of meat and fish consumption and endometriosis risk. Am. J. Obstet. Gynecol. 219:178.e1. doi: 10.1016/j.ajog.2018.05.034
Yang, Q., Wang, Y., Cai, H., Zhou, Q., Zeng, L., Li, S., et al. (2023). Translocation of vaginal and cervical low-abundance non-Lactobacillus bacteria notably associate with endometriosis: a pilot study. Microb. Pathog. 183:106309. doi: 10.1016/j.micpath.2023.106309
Yano, M., Matsuda, A., Natsume, T., Ogawa, S., Awaga, Y., Hayashi, I., et al. (2019). Pain-related behavior and brain activation in cynomolgus macaques with naturally occurring endometriosis. Hum. Reprod. 34, 469–478. doi: 10.1093/humrep/dey383
You, S., Ma, Y., Yan, B., Pei, W., Wu, Q., Ding, C., et al. (2022). The promotion mechanism of prebiotics for probiotics: a review. Front. Nutr. 9:1000517. doi: 10.3389/fnut.2022.1000517
Young, V. J., Ahmad, S. F., Duncan, W. C., and Horne, A. W. (2017). The role of TGF-beta in the pathophysiology of peritoneal endometriosis. Hum. Reprod. Update 23, 548–559. doi: 10.1093/humupd/dmx016
Yuan, M., Li, D., Zhang, Z., Sun, H., An, M., and Wang, G. (2018). Endometriosis induces gut microbiota alterations in mice. Hum. Reprod. 33, 607–616. doi: 10.1093/humrep/dex372
Yuan, W., Wu, Y., Chai, X., and Wu, X. (2022). The colonized microbiota composition in the peritoneal fluid in women with endometriosis. Arch. Gynecol. Obstet. 305, 1573–1580. doi: 10.1007/s00404-021-06338-7
Keywords: endometriosis, microbiota, metabolomics, infertility, etiology, biomarkers, therapeutics
Citation: Wang M, Liu W, Zheng L, Ma S, Jin L, Zhao D and Li D (2025) Broadening horizons: microbiota as a novel biomarker and potential treatment for endometriosis. Front. Microbiol. 16:1521216. doi: 10.3389/fmicb.2025.1521216
Edited by:
Philippa T. Saunders, University of Edinburgh, United KingdomReviewed by:
Ökkeş Zortuk, Ministry of Health, TürkiyeSayali Kandari, Cellsure Biotech And Research Centre, India
Copyright © 2025 Wang, Liu, Zheng, Ma, Jin, Zhao and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dandan Li, MjY5NjYxODQ3QHFxLmNvbQ==; Donghai Zhao, emRoNzUxMDI3QDEyNi5jb20=; Lianhai Jin, bGlhbmhhaWVkdUAxNjMuY29t
†These authors have contributed equally to this work and share first authorship