 Mateusz Szadziul
Mateusz Szadziul Agata Goryluk-Salmonowicz
Agata Goryluk-Salmonowicz Magdalena Popowska
Magdalena Popowska- 1University of Warsaw, Faculty of Biology, Institute of Microbiology, Department of Bacterial Physiology, Warsaw, Poland
- 2Warsaw University of Life Sciences, Faculty of Biology and Biotechnology, Institute of Biology, Department of Biochemistry and Microbiology, Warsaw, Poland
Antimicrobial resistance (AMR) is a critical global health concern. While AMR research has primarily focused on medical and veterinary settings, the spread of antibiotic-resistant bacteria (ARB) and antibiotic resistance genes (ARGs) through natural environments, including soil, remains poorly understood. This study investigates the relationship between soil physico-chemical properties and ARG abundance in environments with varying levels of anthropogenic impact. Soil samples were collected from agricultural fields after harvest (both manured and non-manured, but fertilized with mineral fertilizers) and forests, analyzed for 24 physico-chemical parameters, and subjected to DNA extraction. High-throughput qPCR was used to quantify 27 clinically relevant ARGs and 5 mobile genetic elements (MGEs) in the samples. Results revealed significant differences in soil properties between arable and forest soils, particularly in water content, humus levels, sand and silt proportions, and mercury concentration (p ≤ 0.05). Arable soils exhibited a significantly higher abundance of ARGs and MGEs (p = 0.0247), with certain resistance genes found exclusively in agricultural environments. Correlation analysis identified strong positive associations between MGEs and ARGs, highlighting the role of genetic elements in AMR dissemination. Additionally, soil properties such as aluminum, nitrogen, and magnesium showed positive correlations with ARG and MGE abundance, while sand content and the carbon-to-nitrogen ratio displayed inverse correlations. The results indicate that heavy metal contamination may play a substantial role in AMR spread through co-selection mechanisms. These findings emphasize the influence of environmental factors on AMR dynamics and highlight the need to integrate soil ecology into AMR mitigation strategies within the One Health framework.
1 Introduction
Antimicrobial resistance (AMR) is considered one of the largest health-related issues globally (World Health Organization, 2024). AMR, which exists in both clinical and natural environments, is the subject of numerous studies (Hernando-Amado et al., 2019). However, the issue of AMR spread is described mainly from a medical and veterinary perspective. Reducing antibiotic consumption and implementing other strategies are important measures to lower antimicrobial resistance in a global health context, but the evolution and spread of antibiotic-resistant bacteria (ARB) and antibiotic resistance genes (ARGs) into the environment cannot be reversed (Collignon et al., 2018). Therefore, the dissemination of AMR through the environment leads to various health issues and confirms the present-day consensus that tackling AMR requires action aligned with the ‘One Health’ approach—a holistic framework in which human, veterinary, and environmental settings are interconnected (Aarestrup et al., 2021; Goryluk-Salmonowicz and Popowska, 2022).
Multiple environmental dissemination routes for resistant bacteria have been identified as potentially significant for the proliferation of AMR, particularly in the context of intensive agriculture and increasing urbanization (Thanner et al., 2016). Some critical points have been identified, but the lack of knowledge regarding the propagation and amplification of AMR in the environment complicates AMR prevention and mitigation in human and veterinary medicine (Berendonk et al., 2015). Soil biocenosis is among the most biodiverse systems on Earth, and soils play a crucial role in food production. ARB can disseminate into soils and subsequently be transferred back to animals, through food chain and potentially to humans, while evolving and acquiring new ARGs from environmental bacteria in the process (Forsberg et al., 2012; Delgado-Baquerizo et al., 2022; Liu et al., 2024). The pool of resistance genes prevalent in the environment, known as the ‘resistome,’ poses a threat to human and animal health if transferred to infectious pathogens (Davies and Davies, 2010; Gillings et al., 2017). Therefore, environments influenced by the input of ARB, which also carry genetic mobility determinants (such as plasmids, transposons, or integrons), should not be regarded merely as passive dissemination routes. Additionally, the presence of pesticides, antibiotics, heavy metals, and other toxic substances polluting the environment may promote the acquisition, emergence, and co-selection of resistance mechanisms against these compounds (Pal et al., 2015; Chen et al., 2019; Bazzi et al., 2020). The accumulation of antibiotics in soils depends on their physico-chemical properties. Furthermore, physico-chemical characteristics of the environment have also been identified as potential factors influencing the abundance of ARGs (Song et al., 2016; Zhang et al., 2019).
Currently, the understanding of the fate of ARB and ARGs released into the environment is limited. Both the transfer of ARGs and the balance between retention and out competition of ARB in natural communities remain largely unknown. This is due to the vast array of environmental and ecological factors influencing the spread of ARB and ARGs. In this regard, the biodiversity of a given ecosystem is proposed to be pivotal. Indeed, our previous study revealed that in soil environments, higher biological diversity, evenness, and richness were significantly negatively correlated with the relative abundance of over 85% of the ARGs examined. However, a similar effect was not noticeable in river ecosystems due to their more dynamic nature (Klümper et al., 2024).
To determine whether there is a quantitative and qualitative relationship between ARGs and the physico-chemical properties of soil, research samples were collected after harvest from cultivated fields (both manured and treated with mineral fertilizers) and forest areas. All collected soil samples were analyzed for their physico-chemical properties, including pH value, water content, humus, sand and clay proportions, total nitrogen, carbon-to-nitrogen ratio, and the presence of various chemical elements, including bioavailable elements and heavy metals. In total, 24 different factors were examined. The resistome was characterized by assessing the abundance of 27 clinically relevant ARGs using high-throughput chip-based qPCR. Additionally, the abundance of mobile genetic elements (MGEs) in the samples was evaluated through 5 marker genes associated with AMR. This study reveals that ARG and MGE distribution varies with soil physico-chemical properties and their origin, with higher abundances linked to arable soils and heavy metal concentrations.
2 Materials and methods
2.1 Sample collection
The study was conducted on soil samples collected in autumn 2022 from six areas in Poland with varying levels of anthropogenic activity: four agricultural fields (two manured and two treated with mineral fertilizers) representing high activity, and two forests representing low activity. At each location, five soil subsamples were collected along two 10 m diagonals laid in an X-pattern. The subsamples were combined in a sterile plastic bag, homogenized, and transferred to the laboratory. In total, approximately 1 kg of soil was gathered per sampling location at a single time point. Samples were divided into aliquots of 20 g, sieved (2 mm mesh size) and stored at −20°C. Supplementary Table S1 presents the locations of the sampling sites.
2.2 Physico-chemical properties analysis
Collected soil samples were analyzed for their physico-chemical properties by an external laboratory (AGES, Austria). In total, 24 different factors were examined for each soil sample (Supplementary Table S1).
2.3 DNA isolation
Total DNA was extracted using the DNeasy PowerSoil Pro Kit (Qiagen) according to the manufacturer’s instructions. DNA samples were isolated in four replicates of 0.25 g of soil each and subsequently combined. The quality and quantity of the extracted DNA were assessed using a Qubit 4.0 Fluorometer (dsDNA high-sensitivity assay kit, Invitrogen) and a Colibri spectrophotometer (Titertek Berthold).
2.4 High-throughput qPCR
Determination of the relative abundance of target genes in the samples was performed by Resistomap (Finland) using a SmartChip Real-Time PCR cycler (Takara). A total of 27 ARGs and 5 mobile genetic elements (MGEs) were targeted with customised and independently analysed primer sets designed for assessment of the environmental resistome (Supplementary Table S3) (Zhu et al., 2013; Muziasari et al., 2016; Stedtfeld et al., 2018). The selection of these resistance determinants was based on the recommendations by Berendonk et al., 2015, aiming to include a standardized set of genes that enable global comparability of the results and meet key criteria such as clinical and veterinary relevance, environmental prevalence, association with MGEs and potential for horizontal gene transfer. Quantification included the 16S rRNA gene. The qPCR cycling conditions and initial data processing were carried out as described by Wang et al. (2014). A cycle threshold (CT) of 27 was used as the detection limit (Zhu et al., 2013; Wang et al., 2014). Amplicons with nonspecific melting curves and multiple peaks were discarded from the analysis. The relative abundances of the detected genes, expressed as proportions of the 16S rRNA gene, were calculated using the 2 − ΔCT method (Schmittgen and Livak, 2008).
2.5 Statistical analysis
Relationships between physico-chemical properties of soils and the relative abundances of ARGs/MGEs were determined using Spearman’s correlation coefficient and visualized with GraphPad Prism 9. Correlation networks were created using Cytoscape 3.10.1. Calculations included pairwise correlations between physico-chemical properties and ARGs/MGEs as well as correlations among ARGs/MGEs themselves. Correlation was considered strong for a coefficient value of |Rs| > 0.8 and significant for a p-value ≤ 0.05. Additionally, a t-test (Welch’s t-test for single factors and a paired t-test for overall interaction) was performed using GraphPad Prism 9 to assess whether a significant difference existed between arable and forest soils in terms of physico-chemical properties and ARG/MGE abundances. A difference was considered statistically significant for a p-value ≤ 0.05.
3 Results
3.1 Physico-chemical properties of soils
Physico-chemical properties were determined across all samples of arable and forest soils, including pH value, water content, proportion of humus, sand, and clay, total nitrogen, carbon-to-nitrogen ratio, and the presence of various chemical elements, including bioavailable elements and heavy metals (Supplementary Table S1). A comparison of these properties between arable and forest soils was performed using a t-test with Welch’s correction.
In general, the range of values for the analyzed parameters in forest soils was as follows: pH-value 4.5–5.24; Water content [%] 3–6; Humus [%] 1.8–1.9; Sand [%] 82.3–92.8; Silt [%] 2.7–12.6; Clay [%] 4.6–5.1; Total nitrogen [%] 0.07–0.073; Phosphorous [mg/kg] 79–117; Potassium [mg/kg] 14–33; Magnesium [mg/kg] 12–28; Aluminium [mg/kg] 4.1–6.4; Calcium [mg/kg] 0.51–0.579; Boron [mg/kg] 0–0.2, heavy metals [mg/kg]: Arsenic 1.51–2.2; Chromium 3.29–7.2; Copper 1–1.7; Iron 144–153; Lead 5.86–11.8; Manganese 26–70; Mercury 0.02; Nickel 0.96–4.1; Zinc 1.3–2.9; Vanadium 6.14–24.4. For arable soils, the corresponding ranges were as follows: pH-value 5.7–7.2; Water content [%] 12–26; Humus [%] 2.3–3.8; Sand [%] 34.8–65.9; Silt [%] 27.7–53.4; Clay [%] 6.5–12.6; Total nitrogen [%] 0.12–3; Phosphorous [mg/kg] 61–99; Potassium [mg/kg] 106–733; Magnesium [mg/kg] 70–220; Aluminium [mg/kg] 7.3–11; Calcium [mg/kg] 1.34–4.84, Boron [mg/kg] 0.3–3.1; heavy metals [mg/kg]: Arsenic 2.4–4.1; Chromium 10.6–128.4; Copper 1.6–8.8; Iron 265–1937; Lead 9.2–700.4; Manganese 3.76–308; Mercury 0.03–0.05; Nickel 6.2–19; Zinc 3.8–1433.5; Vanadium 12.3–20.1. Detailed data are provided in Supplementary Table S1.
The analysis revealed a significant difference between arable and forest soils (p ≤ 0.05) in terms of soil water content, humus, sand and silt proportions, and mercury concentration. Several other factors exhibited weaker but noteworthy trends (p ≤ 0.1), including pH value, total nitrogen content, and the concentrations of magnesium, copper, arsenic, nickel, and aluminum. However, no significant differences (p > 0.1) were found for clay content, carbon-to-nitrogen ratio, or the concentrations of phosphorus, potassium, iron, manganese, zinc, boron, lead, chromium, vanadium, and calcium (Supplementary Table S2). Overall, forest and arable soils exhibited significantly different physico-chemical properties (p = 0.034; R2 = 0.1809).
3.2 Diversity and abundance of ARGs and MGEs
The relative abundance of the analyzed antibiotic resistance genes (ARGs) and mobile genetic elements (MGEs) differed between arable and forest soils (p = 0.0247). The average abundance of these genes was lower in forest soils compared to arable soils (total mean difference = −0.05191 copy numbers/16S rRNA copy number; R2 = 0.1524).
Two of the analyzed genes, blaOXA58 (β-lactam resistance) and mcr3 (collistin resistance), were not detected in any of the soil samples. Additionally, several genes were found exclusively in arable soils: intl1_1 (class 1 integron-integrase), Tn5 (transposase), aph3-VIa (aminoglycoside resistance), ermB_1 (MLSB resistance), ermF_1, qnrA (quinolone resistance), qnrS_1, tetM_2 (tetracycline resistance), tetO_1 and dfrA1_1 (trimethoprim resistance). In total, 5 MGEs and 25 ARGs were identified in the analyzed soils (Figure 1). Supplementary Table S1 presents the relative abundances of the identified genes in soil samples.
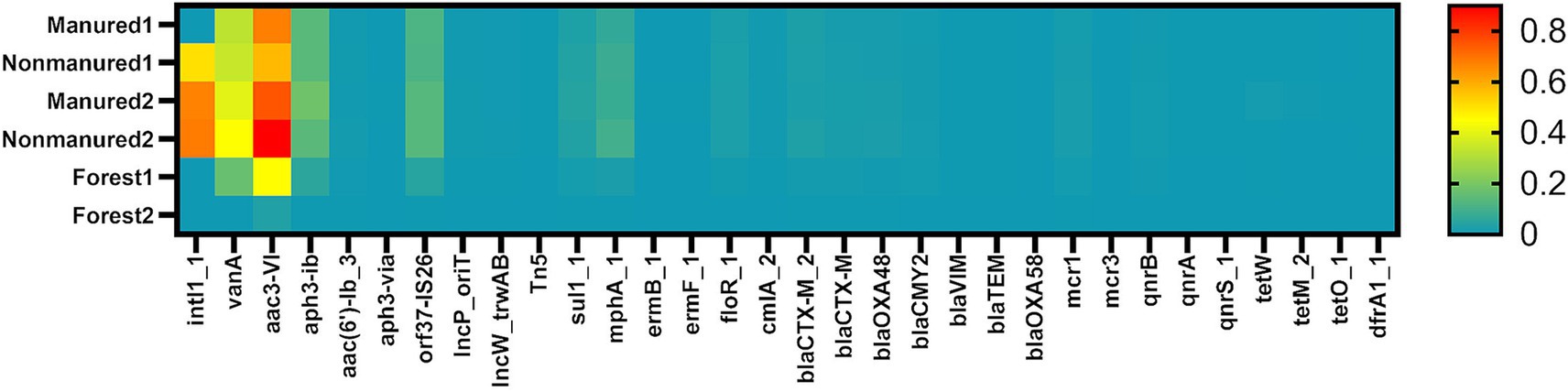
Figure 1. Heatmap of relative ARGs and MGEs abundances in the soils. Values shown are transformed to log₁₀ scale. Samples are color coded from high abundance (red) to below the detection limit (cyan).
A comparison of specific ARG and MGE mean abundances between arable and forest soils revealed significant differences (p ≤ 0.05) for sul1_1 (sulfonamide resistance), mphA_1 (macrolide resistance), cmlA_2 (chloramphenicol resistance), blaOXA48 (β-lactam resistance), blaTEM (β-lactam resistance), qnrA, qnrS_1. Weaker evidence (p ≤ 0.1) suggested a similar difference for intl1_1, Tn5, aac(6′)-Ib_3 (aminoglycoside resistance), floR_1 (chloramphenicol resistance) and blaCTX-M_2 (β-lactam resistance). In all cases, including genes with no significant differences, mean abundances were higher in arable soils (Supplementary Table S2). However, no significant differences were observed when comparing overall abundances of MGEs or specific antibiotic classes.
3.3 Correlation between ARGs and MGEs
Correlation analysis between mobile genetic elements (MGEs) and antibiotic resistance genes (ARGs) revealed a network consisting of 25 nodes (20 ARGs and 5 MGEs) and 147 edges, representing strong and significant correlations (|Rs| > 0.8; p ≤ 0.05) (Figure 2A). All correlations were positive. Figure 2B represents a heatmap of correlations between all studied ARGs and MGEs.
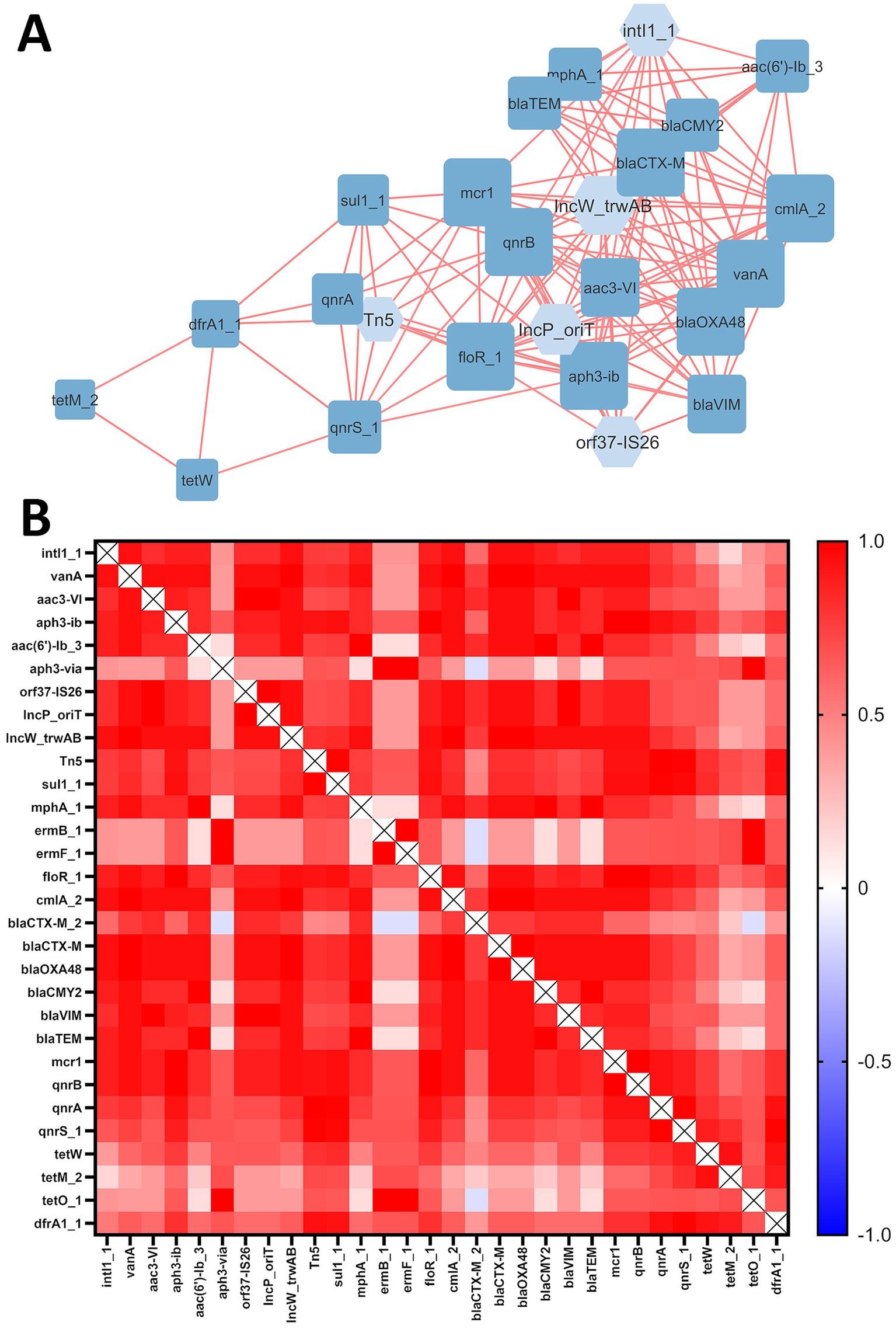
Figure 2. (A) ARG and MGE interaction network in examined soils. The ‘organic layout’ network is presented, where edges indicate strong and significant correlations (|Rs| > 0.8; p-value ≤ 0.05). Positive correlations are shown in red. The size of the nodes represents the degree of interaction, while node colors distinguish MGEs (light blue) from ARGs (dark blue). (B) Heatmap of correlations between ARGs and MGEs, where color represents positive (red) or negative (blue) correlations.
A total of 47 interactions were associated with MGEs, highlighting their role in network formation. All examined MGEs (intl1_1, orf37-IS26, IncP_oriT, IncW_trwAB and Tn5) interacted with ARGs. Furthermore, all of 20 ARGs present in the network were positively correlated with each other. No significant interactions were observed for the following ARGs identified in soil samples: aph3-via, ermB_1, ermF_1, blaCTX-M_2 and tetO_1.
3.4 Correlation between ARGs/MGEs and physico-chemical properties of soils
A correlation analysis was conducted to examine potential interactions between antibiotic resistance genes (ARGs), mobile genetic elements (MGEs), and various physico-chemical soil properties. The resulting network consisted of 44 nodes (20 ARGs, 5 MGEs, and 19 physico-chemical factors) and 275 edges (excluding interactions between ARGs and MGEs), representing strong and significant correlations (|Rs| > 0.8; p ≤ 0.05) (Figure 3A). Of these interactions, 26 were negative, while 249 were positive. Figure 3B represents a heatmap of correlations between all studied physico-chemical properties and ARGs/MGEs.
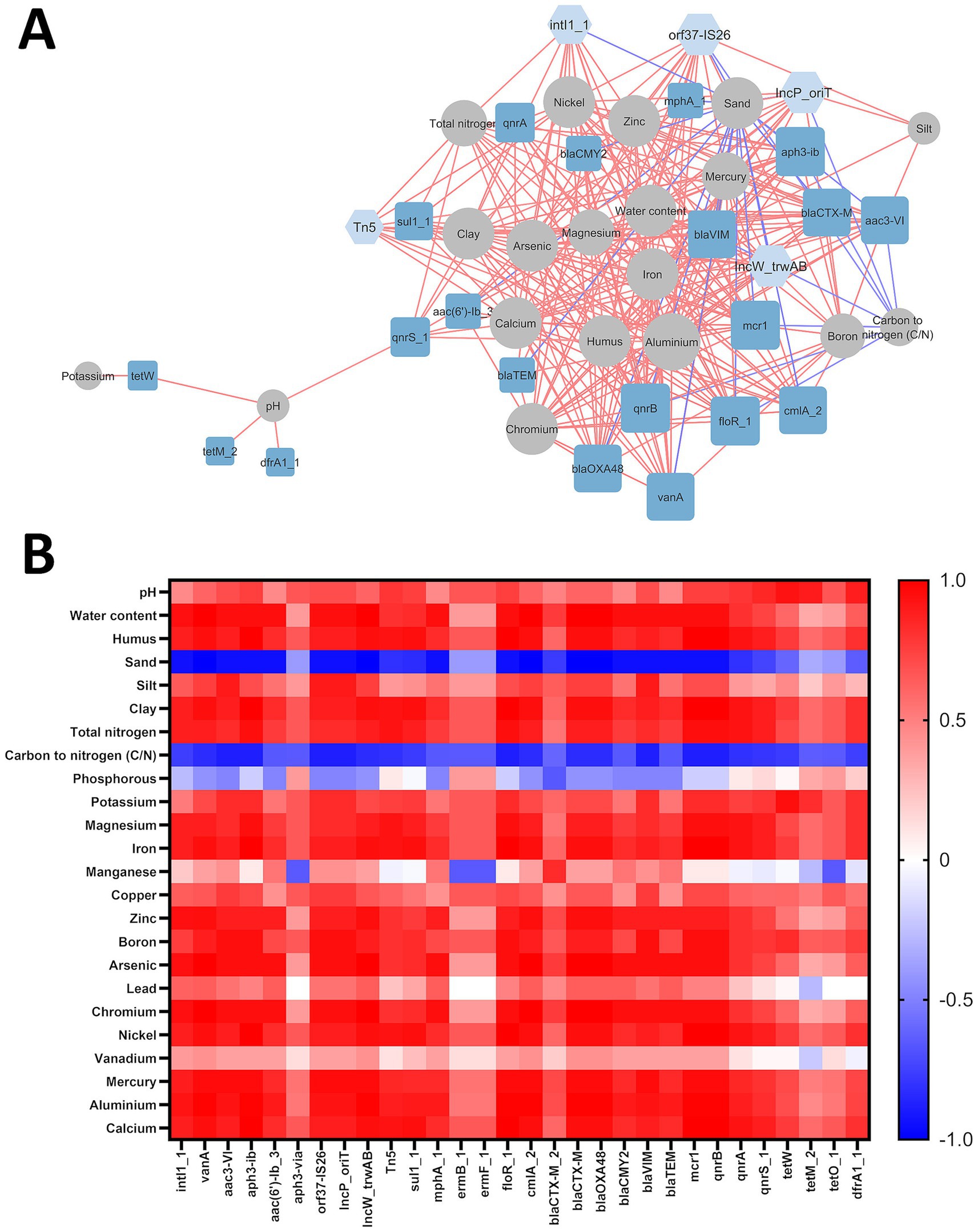
Figure 3. (A) Interaction network of ARGs/MGEs with physico-chemical factors in examined soils. The ‘organic layout’ network is presented, with edges indicating strong and significant correlations (|Rs| > 0.8; p ≤ 0.05). Red edges denote positive correlations. Node sizes represent the degree of interaction, while node colors distinguish MGEs (light blue), ARGs (dark blue), and physico-chemical factors (gray). (B) Heatmap of correlations between ARGs/MGEs and soil physico-chemical properties, where red represents positive correlations and blue represents negative correlations.
Specifically, sand content and the carbon-to-nitrogen ratio were inversely correlated with ARG and MGE abundances, whereas most other physico-chemical factors showed positive correlations. No significant correlations were observed between ARG/MGE abundances and copper, lead, manganese, phosphorus, or vanadium. The degree of interaction (number of correlated genes) among physico-chemical factors varied, ranging from 1 to 22 edges. Potassium, silt content, and soil pH exhibited the fewest correlations (1 for potassium and 4 for silt content and pH), while aluminum abundance had the highest number of connections (22).
All examined MGE-encoding genes were correlated with at least one soil parameter. However, five ARGs (aph3-via, ermB_1, ermF_1, blaCTXM-2 and tetO_1) did not exhibit significant interactions with any tested physico-chemical factors.
Aluminum abundance showed a strong positive correlation with all tested MGE-related genes (intl1_1, orf37-IS26, IncP_oriT, IncW_trwAB, Tn5), and 17 out of 20 ARGs, excluding tetW, tetM_2 and dfrA1_1. Interestingly, tetracycline resistance genes correlated with only two soil parameters: potassium (tetW) and soil pH (tetW, tetM_2).
Physico-chemical properties were categorized into three groups based on their correlation profiles with ARGs:
• Group I: Arsenic (As), chromium (Cr), zinc (Zn), and water content. These factors positively correlated with 14 ARGs, including vanA (vancomycin resistance), aac(3)-VI, aac(6′)-Ib_3, aph3-Ib (aminoglycoside resistance), mphA (macrolide resistance), floR_1, cmlA (phenicol resistance), blaCTX-M, blaOXA-48, blaCMY-2, blaVIM, blaTEM (β-lactam resistance), mcr1 (colistin resistance), and qnrB (quinolone resistance).
• Group II: Calcium (Ca), iron (Fe), nickel (Ni), clay content, and humus content. These factors positively correlated with 13 ARGs, including vanA, aac3-VI, aph3-Ib, floR_1, cmlA, blaCTX-M, blaOXA-48, blaVIM, mcr1, qnrA, qnrB, qnrS, and sul1.
• Group III: Magnesium (Mg) and nitrogen (N). These parameters had nearly the same correlation profile as Group II, except they lacked interactions with aac3-VI and blaVIM.
Boron and mercury were also analyzed separately. Boron showed positive correlations similar to those in Group II, except for sul1_1 and qnrS. Mercury, however, was positively correlated with all genes in the boron group as well as qnrA.
All MGE-related genes were positively correlated with Group II parameters (clay, humus, Ca, Fe, and Ni). Additionally, four out of five MGEs (intl1_1, orf37-IS26, IncP_oriT, IncW_trwAB) correlated with aluminum and Group I parameters (water, As, Cr, Zn). Nitrogen and magnesium correlated positively with intl1_1, IncW_trwAB, and Tn5, while boron and mercury were associated with orf37-IS26, IncP_oriT, and IncW_trwAB.
Analysis of the network topology revealed that most genes and physicochemical parameters form a single, interconnected cluster. However, potassium content and dfrA1_1, tetM_2, tetW genes deviate from this group and pH-value interacts with only one gene (qnrS_1) from the main cluster.
4 Discussion
ARGs presence in the soil environment is a complex issue. Various interlinked factors contribute to the distribution of ARGs in soils, including antibiotic residues, anthropogenic pressure, microbial community structure, and soil physico-chemical properties (Čermák et al., 2008; Popowska et al., 2012; Xie et al., 2018; Cycoń et al., 2019). In our study, we analyzed the relationship between ARGs/MGEs presence and soil physico-chemical properties.
The differences in the physico-chemical properties of forest and arable soils were likely influenced by anthropogenic activities in agricultural soil, including fertilization with organic or inorganic fertilizers, which may explain the lower total nitrogen levels in forest soils. Previous studies have reported that fertilization affects soil physico-chemical properties (Li et al., 2018; Zhang X. et al., 2023). Additionally, agricultural soils may be more sensitive to external inputs, leading to significant changes in their physico-chemical properties compared to forest soils (Bang-Andreasen et al., 2020).
In general, highly fertile soils are typically used for agriculture, whereas forest soils exhibit a broader range of physico-chemical properties compared to arable land. Forest soils generally have lower pH values and fewer available nutrients, which aligns with our findings (Plassart et al., 2019). Our study revealed that sand content was higher (82.3–92.8%) and silt content lower (2.7–12.6%) in forest soils compared to arable soils (34.8–65.9% and 27.7–53.4%, respectively). However, the variation in clay content between the examined soils was minimal, and no significant differences were observed. Previous research has reported higher silt and clay contents in arable soils compared to forest soils (Tewari et al., 2016). Polish soils generally have high sand content, and its lower levels in arable soils could be attributed to continuous agricultural use, increased fertility, and fertilization practices (Ballabio et al., 2016). Water content was also significantly lower in forest soils in our study. In addition to anthropogenic influences such as field irrigation, the sandy soil structure of forests, characterized by low compaction resistance and macropore formation, may also be a contributing factor (Gregory et al., 2007).
Our previous research on forest soils across several European countries identified the most abundant ARGs as aac3-VI (aminoglycoside resistance), dfrA1 (trimethoprim resistance), mphA (MLSB) and qnrS (quinolone resistance) (Klümper et al., 2024). In this study, we obtained similar results, but with a few significant differences: vanA (vancomycin resistance) was the most abundant ARG, while dfrA1 was the least abundant ARG. Other studies on ARGs in agricultural soils have shown that genes conferring resistance to ß-lactams, glycopeptides (e.g., vancomycin), and aminoglycosides comprise up to 35% of detected ARG subtypes, which aligns with our findings, although with lower ß-lactamase abundances (Zhang et al., 2021). Our study revealed that ARGs and MGEs are more abundant and diverse in arable soils, suggesting that agricultural areas are important hotspots for ARG pollution (Sun et al., 2020). However, while manure application plays a key role in ARG distribution and increasing abundance in arable fields, we did not observe this relationship in our data (Huang et al., 2021; Zalewska et al., 2021; Zhang Y. et al., 2023). The agricultural soils we examined had been fertilized with either organic or inorganic fertilizers within the last 2 years before this study, therefore the recent application of manure or mineral fertilizers may explain the lack of a detectable relationship. Additionally, we lack information on the long-term fertilization history of the examined fields; it is possible that earlier manure applications could still impact ARG abundance (Zhang Y. et al., 2023). Another potential explanation is that extreme levels of metal concentrations in one set of soils contributed to ARG abundances elevation via co-selection mechanisms (Zhang et al., 2018; Goryluk-Salmonowicz and Popowska, 2022; Zou et al., 2025). Similar evidence exists for the co-selection of MGEs in metal-polluted soils (Zhao et al., 2019). Further studies are needed to elucidate the mechanisms of ARG and MGE co-selection in soils affected by significant metal pollution.
Our analysis revealed a substantial number of correlations between ARGs and MGEs, with all MGEs linked to at least one ARG, suggesting their role in establishing genetic networks. Indeed, in many environments crucial for antibiotic resistance spread, MGEs are key contributors to ARG abundance (Yang et al., 2021; Delgado-Baquerizo et al., 2022; Li et al., 2024). According to our findings, the presence of 20 out of 27 examined ARGs was positively correlated with other ARGs. Research on Chinese soils heavily polluted with industrial waste also identified similar coexistence patterns between ARGs and MGEs, with single genes linked to up to 13 other ARGs (Yang et al., 2021). Multiple correlations between ARGs and MGEs were also observed in Northern Ireland, where several ARGs corresponded to class 1 integrons and transposons, with intl1_1 showing the highest number of correlations, in agreement with our results (Zhao et al., 2019). Among the ARGs examined in this study, vanA exhibited the highest number of interactions (16) with other ARGs/MGEs. This could be attributed to the widespread presence and relatively high abundance of vancomycin resistance genes in the environment (Han et al., 2022; Zheng et al., 2022; Zhang et al., 2025). Previous study has suggested that high ARG abundance is associated with decreased phylogenetic diversity (Klümper et al., 2024). Future research should further investigate the relationship between ARG and MGE abundance and microbiome composition.
The presence of antibiotic resistance genes may be influenced by soil metal concentrations and other elemental factors. In our study, one of the arable soils exhibited extremely elevated concentrations of zinc, lead, and chromium, alongside the highest ARG/MGE abundances among all examined soils. While the direct effects of soil physico-chemical properties and heavy metal presence on ARG variation are relatively small compared to the impact of microbial community structure (3.57 and 1.19% versus 79.76%, respectively), these factors significantly shape the bacterial community and, in turn, contribute substantially to variations in ARG and MGE profiles (Yang et al., 2021; Zhang et al., 2021).
The introduction of heavy metals into agricultural soils through fertilization with both organic and inorganic fertilizers is also of considerable importance. Inorganic fertilizers, also known as chemically manufactured/synthetic fertilizers, are composed of inorganic substances and chemical materials. Long-term fertilization can lead to a gradual buildup of heavy metals in soil (Wang et al., 2020; Gao et al., 2021). Additionally, soil properties play a key role in determining heavy metal availability - clay soils, for example, tend to retain more metals than sandy soils and organic matter content and soil pH also influence the bioavailability of heavy metals in soil environments (Alengebawy et al., 2021). In fact, recent studies indicate the pivotal role of soil pH, which often shows a negative correlation with the availability of many heavy metals (Wang et al., 2024). Soil pH affects the availability and mobility of heavy metals in the environment. Lower pH levels (indicating increased acidity) tend to enhance the solubility and bioavailability of heavy metals, whereas higher pH levels (indicating alkalinity) generally reduce their mobility and uptake. This phenomenon is attributed to the effect of pH on the speciation and solubility of heavy metals within the soil solution, which affects their interaction with soil constituents and subsequent uptake by plant roots. At high pH levels, metals are more likely to form poorly soluble phosphates and carbonates. In contrast, lower pH levels favor the presence of metals in more bioavailable, free ionic forms, increasing their potential uptake (Olaniran et al., 2013). However, this relationship is not reflected in our results, where mean pH was lower in forest soils compared to agricultural soils, but metal concentrations were significantly higher in arable soils, probably due to the buildup of heavy metals that can be attributed to fertilization, as discussed before.
Aluminum is considered a major limiting factor for plant growth and productivity in acidic soils, as it can negatively affect both plants - by inhibiting root system development - and microorganisms, including symbiotic bacteria associated with legumes (Levett et al., 2019; Ofoe et al., 2023). In this study, aluminum content emerged as the most significant physico-chemical factor associated with increased ARG and MGE abundances. Several studies have described the role of aluminum compounds in facilitating antibiotic resistance gene transfer under laboratory conditions (Liu et al., 2019; Aras et al., 2023). A soil study by Ferreira et al. (2024) found that aluminum presence was positively correlated with tetX, sul1 and mphA gene abundance. Similarly, an analysis of residential soil samples in Australia identified aluminum as a major factor influencing ARG abundance, with aluminum levels correlating with the presence of blaOXA, blaTEM, tetM, tetW, sul2 and sul3 (Knapp et al., 2017). Our study, conducted on soils in Poland, supports these findings and confirms the consistency of this relationship.
Knapp et al. (2011) analyzed Scottish soils and identified correlations between 11 ARGs and soil metals. Two ß-lactam resistance genes (blaCTX-M and blaOXA48) were positively correlated with chromium content (53.1 mg/kg), aligning with our findings (mean chromium content: 29.8 mg/kg). However, we observed no correlation between tested ARGs and elements such as copper (1–8.8 mg/kg), lead (9–700.4 mg/kg), manganese (3.76–308 mg/kg), phosphorus (61–117 mg/kg), or vanadium (6.14–24.4 mg/kg). These results are consistent with other studies; Knapp et al. (2011) also observed no correlation of ARGs with lead in concentrations of 10–1,000 mg/kg. Despite existing evidence that copper exposure may act as a selective pressure for antibiotic resistance, our study did not find a significant relationship between copper content and ARG/MGE abundance (Kang et al., 2018). One possible explanation for the absence of such observations is that copper, phosphorus, and magnesium are essential micronutrients required for proper plant growth, with their availability being influenced by factors such as soil pH and their concentrations varying widely depending on soil condition and land use - for instance, copper levels in agricultural soils typically range from 5 to 30 mg/kg (Brun et al., 2003).
In summary, most physico-chemical properties showed a positive correlation with all examined ARGs and MGEs. This study provides a framework for assessing the relationship between ARGs, MGEs, and soil physico-chemical parameters, especially metal content, which could inform the development of targeted fertilization and remediation strategies to mitigate the environmental impact of AMR. Therefore, further research is needed to thoroughly investigate the influence of specific physico-chemical properties on ARG and MGE abundance. Another important variable that should be addressed is the seasonal variation of physico-chemical properties and ARGs/MGEs distribution, as this study was limited to the samples collected in autumn of 2022 (after harvest) and presents only a snapshot of examined interactions. Given that these factors shape the entire soil ecosystem, future studies on ARG pollution should explore the interplay between soil characteristics, microbiome composition, and ARGs diversity and abundance, while possibly including the role of seasonal variations of these characteristics.
5 Conclusion
This study highlights the differences in ARG and MGE distribution based on soil physico-chemical properties, especially the content of heavy metals, and soil origin/utilisation. Forest and arable soils exhibit distinct physico-chemical characteristics, with ARGs and MGEs generally more abundant in arable soils. Strong and significant correlations between MGEs and ARGs were observed across all soils. The results indicate that soil physico-chemical properties influence ARG and MGE abundances, with nearly all examined properties associated with specific ARGs and MGEs. In most cases, these correlations were consistently positive or negative across all genes, with aluminum abundance showing the highest number of associations, followed by calcium, chromium, iron, nickel and zinc, reinforcing the role of metal pollution in promoting AMR spread. A comprehensive assessment of AMR’s impact on soil environments requires further research examining the interaction between all contributing factors, including physico-chemical properties, climatic conditions, seasonal shifts, microbiome composition, and anthropogenic influences such as fertilization and contamination.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MS: Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. AG-S: Formal analysis, Investigation, Writing – original draft. MP: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The research was funded by the National Science Centre of Poland grant (UMO-2019/32/Z/NZ8/00011) as a part of the project: “ANTIVERSA-Biodiversity as an ecological barrier for the spread of clinically relevant antibiotic resistance in the environment” (BiodivERsa2018-A-452).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1584660/full#supplementary-material
References
Aarestrup, F. M., Bonten, M., and Koopmans, M. (2021). Pandemics– one health preparedness for the next. Lancet Reg. Health Eur. 9:100210. doi: 10.1016/j.lanepe.2021.100210
Alengebawy, A., Abdelkhalek, S. T., Qureshi, S. R., and Wang, M.-Q. (2021). Heavy metals and pesticides toxicity in agricultural soil and plants: ecological risks and human health implications. Toxics 9:42. doi: 10.3390/toxics9030042
Aras, A., Rizvanoglu, S. S., Tanriverdi, E. S., Karaca, B., and Eryilmaz, M. (2023). The effects of antiperspirant aluminum Chlorohydrate on the development of antibiotic resistance in Staphylococcus epidermidis. Microorganisms 11:948. doi: 10.3390/microorganisms11040948
Ballabio, C., Panagos, P., and Monatanarella, L. (2016). Mapping topsoil physical properties at European scale using the LUCAS database. Geoderma 261, 110–123. doi: 10.1016/j.geoderma.2015.07.006
Bang-Andreasen, T., Anwar, M. Z., Lanzén, A., Kjøller, R., Rønn, R., Ekelund, F., et al. (2020). Total RNA sequencing reveals multilevel microbial community changes and functional responses to wood ash application in agricultural and forest soil. FEMS Microbiol. Ecol. 96:fiaa016. doi: 10.1093/femsec/fiaa016
Bazzi, W., Abou Fayad, A. G., Nasser, A., Haraoui, L.-P., Dewachi, O., Abou-Sitta, G., et al. (2020). Heavy metal toxicity in armed conflicts potentiates AMR in A. baumannii by selecting for antibiotic and heavy metal co-resistance mechanisms. Front. Microbiol. 11:68. doi: 10.3389/fmicb.2020.00068
Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., et al. (2015). Tackling antibiotic resistance: the environmental framework. Nat. Rev. Microbiol. 13, 310–317. doi: 10.1038/nrmicro3439
Brun, L. A., Le Corff, J., and Maillet, J. (2003). Effects of elevated soil copper on phenology, growth and reproduction of five ruderal plant species. Environ. Pollut. 122, 361–368. doi: 10.1016/S0269-7491(02)00312-3
Čermák, L., Kopecký, J., Novotná, J., Omelka, M., Parkhomenko, N., Plháčková, K., et al. (2008). Bacterial communities of two contrasting soils reacted differently to lincomycin treatment. Appl. Soil Ecol. 40, 348–358. doi: 10.1016/j.apsoil.2008.06.001
Chen, J., Li, J., Zhang, H., Shi, W., and Liu, Y. (2019). Bacterial heavy-metal and antibiotic resistance genes in a copper tailing dam area in northern China. Front. Microbiol. 10:1916. doi: 10.3389/fmicb.2019.01916
Collignon, P., Beggs, J. J., Walsh, T. R., Gandra, S., and Laxminarayan, R. (2018). Anthropological and socioeconomic factors contributing to global antimicrobial resistance: a univariate and multivariable analysis. Lancet Planet. Health 2, e398–e405. doi: 10.1016/S2542-5196(18)30186-4
Cycoń, M., Mrozik, A., and Piotrowska-Seget, Z. (2019). Antibiotics in the soil environment—degradation and their impact on microbial activity and diversity. Front. Microbiol. 10:338. doi: 10.3389/fmicb.2019.00338
Davies, J., and Davies, D. (2010). Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 74, 417–433. doi: 10.1128/MMBR.00016-10
Delgado-Baquerizo, M., Hu, H.-W., Maestre, F. T., Guerra, C. A., Eisenhauer, N., Eldridge, D. J., et al. (2022). The global distribution and environmental drivers of the soil antibiotic resistome. Microbiome 10:219. doi: 10.1186/s40168-022-01405-w
Ferreira, P. F. A., Rocha, F. I., Howe, A., Barbosa, D. R., da Conceição Jesus, E., Do Amaral Sobrinho, N. M. B., et al. (2024). Chemical attributes, bacterial community, and antibiotic resistance genes are affected by intensive use of soil in agro-ecosystems of the Atlantic Forest, southeastern Brazil. Environ. Geochem. Health 46:123. doi: 10.1007/s10653-024-01894-8
Forsberg, K. J., Reyes, A., Wang, B., Selleck, E. M., Sommer, M. O. A., and Dantas, G. (2012). The shared antibiotic resistome of soil bacteria and human pathogens. Science 337, 1107–1111. doi: 10.1126/science.1220761
Gao, P., Huang, J., Wang, Y., Li, L., Sun, Y., Zhang, T., et al. (2021). Effects of nearly four decades of long-term fertilization on the availability, fraction and environmental risk of cadmium and arsenic in red soils. J. Environ. Manag. 295:113097. doi: 10.1016/j.jenvman.2021.113097
Gillings, M. R., Paulsen, I. T., and Tetu, S. G. (2017). Genomics and the evolution of antibiotic resistance. Ann. N. Y. Acad. Sci. 1388, 92–107. doi: 10.1111/nyas.13268
Goryluk-Salmonowicz, A., and Popowska, M. (2022). Factors promoting and limiting antimicrobial resistance in the environment – existing knowledge gaps. Front. Microbiol. 13:992268. doi: 10.3389/fmicb.2022.992268
Gregory, A. S., Watts, C. W., Whalley, W. R., Kuan, H. L., Griffiths, B. S., Hallett, P. D., et al. (2007). Physical resilience of soil to field compaction and the interactions with plant growth and microbial community structure. Eur. J. Soil Sci. 58, 1221–1232. doi: 10.1111/j.1365-2389.2007.00956.x
Han, B., Ma, L., Yu, Q., Yang, J., Su, W., Hilal, M. G., et al. (2022). The source, fate and prospect of antibiotic resistance genes in soil: a review. Front. Microbiol. 13:976657. doi: 10.3389/fmicb.2022.976657
Hernando-Amado, S., Coque, T. M., Baquero, F., and Martínez, J. L. (2019). Defining and combating antibiotic resistance from one health and Global Health perspectives. Nat. Microbiol. 4, 1432–1442. doi: 10.1038/s41564-019-0503-9
Huang, J., Mi, J., Yan, Q., Wen, X., Zhou, S., Wang, Y., et al. (2021). Animal manures application increases the abundances of antibiotic resistance genes in soil-lettuce system associated with shared bacterial distributions. Sci. Total Environ. 787:147667. doi: 10.1016/j.scitotenv.2021.147667
Kang, W., Zhang, Y.-J., Shi, X., He, J.-Z., and Hu, H.-W. (2018). Short-term copper exposure as a selection pressure for antibiotic resistance and metal resistance in an agricultural soil. Environ. Sci. Pollut. Res. Int. 25, 29314–29324. doi: 10.1007/s11356-018-2978-y
Klümper, U., Gionchetta, G., Catão, E., Bellanger, X., Dielacher, I., Elena, A. X., et al. (2024). Environmental microbiome diversity and stability is a barrier to antimicrobial resistance gene accumulation. Commun. Biol. 7, 706–713. doi: 10.1038/s42003-024-06338-8
Knapp, C. W., Callan, A. C., Aitken, B., Shearn, R., Koenders, A., and Hinwood, A. (2017). Relationship between antibiotic resistance genes and metals in residential soil samples from Western Australia. Environ. Sci. Pollut. Res. 24, 2484–2494. doi: 10.1007/s11356-016-7997-y
Knapp, C. W., McCluskey, S. M., Singh, B. K., Campbell, C. D., Hudson, G., and Graham, D. W. (2011). Antibiotic resistance gene abundances correlate with metal and geochemical conditions in archived Scottish soils. PLoS One 6:e27300. doi: 10.1371/journal.pone.0027300
Levett, A., Gagen, E. J., Diao, H., Guagliardo, P., Rintoul, L., Paz, A., et al. (2019). The role of aluminium in the preservation of microbial biosignatures. Geosci. Front. 10, 1125–1138. doi: 10.1016/j.gsf.2018.06.006
Li, Y., Li, R., Hou, J., Sun, X., Wang, Y., Li, L., et al. (2024). Mobile genetic elements affect the dissemination of antibiotic resistance genes (ARGs) of clinical importance in the environment. Environ. Res. 243:117801. doi: 10.1016/j.envres.2023.117801
Li, D., Luo, P., Han, X., and Yang, J. (2018). Influence of long-term fertilization on soil physicochemical properties in a brown soil. IOP Conf. Ser. Earth Environ. Sci. 108:42027. doi: 10.1088/1755-1315/108/4/042027
Liu, X., Tang, J., Song, B., Zhen, M., Wang, L., and Giesy, J. P. (2019). Exposure to Al2O3 nanoparticles facilitates conjugative transfer of antibiotic resistance genes from Escherichia coli to Streptomyces. Nanotoxicology 13, 1422–1436. doi: 10.1080/17435390.2019.1669731
Liu, Z.-L., Wang, Y.-F., Zhu, D., Quintela-baluja, M., Graham, D. W., Zhu, Y.-G., et al. (2024). Increased transmission of antibiotic resistance occurs in a soil food chain under pesticide stress. Environ. Sci. Technol. 58, 21989–22001. doi: 10.1021/acs.est.4c07822
Muziasari, W. I., Pärnänen, K., Johnson, T. A., Lyra, C., Karkman, A., Stedtfeld, R. D., et al. (2016). Aquaculture changes the profile of antibiotic resistance and mobile genetic element associated genes in Baltic Sea sediments. FEMS Microbiol. Ecol. 92:fiw052. doi: 10.1093/femsec/fiw052
Ofoe, R., Thomas, R. H., Asiedu, S. K., Wang-Pruski, G., Fofana, B., and Abbey, L. (2023). Aluminum in plant: benefits, toxicity and tolerance mechanisms. Front. Plant Sci. 13:1085998. doi: 10.3389/fpls.2022.1085998
Olaniran, A. O., Balgobind, A., and Pillay, B. (2013). Bioavailability of heavy metals in soil: impact on microbial biodegradation of organic compounds and possible improvement strategies. Int. J. Mol. Sci. 14, 10197–10228. doi: 10.3390/ijms140510197
Pal, C., Bengtsson-Palme, J., Kristiansson, E., and Larsson, D. G. J. (2015). Co-occurrence of resistance genes to antibiotics, biocides and metals reveals novel insights into their co-selection potential. BMC Genomics 16:964. doi: 10.1186/s12864-015-2153-5
Plassart, P., Prévost-Bouré, N. C., Uroz, S., Dequiedt, S., Stone, D., Creamer, R., et al. (2019). Soil parameters, land use, and geographical distance drive soil bacterial communities along a European transect. Sci. Rep. 9:605. doi: 10.1038/s41598-018-36867-2
Popowska, M., Rzeczycka, M., Miernik, A., Krawczyk-Balska, A., Walsh, F., and Duffy, B. (2012). Influence of soil use on prevalence of tetracycline, streptomycin, and erythromycin resistance and associated resistance genes. Antimicrob. Agents Chemother. 56, 1434–1443. doi: 10.1128/aac.05766-11
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Song, L., Li, L., Yang, S., Lan, J., He, H., McElmurry, S. P., et al. (2016). Sulfamethoxazole, tetracycline and oxytetracycline and related antibiotic resistance genes in a large-scale landfill, China. Sci. Total Environ. 551-552, 9–15. doi: 10.1016/j.scitotenv.2016.02.007
Stedtfeld, R. D., Guo, X., Stedtfeld, T. M., Sheng, H., Williams, M. R., Hauschild, K., et al. (2018). Primer set 2.0 for highly parallel qPCR array targeting antibiotic resistance genes and mobile genetic elements. FEMS Microbiol. Ecol. 94:fiy130. doi: 10.1093/femsec/fiy130
Sun, J., Jin, L., He, T., Wei, Z., Liu, X., Zhu, L., et al. (2020). Antibiotic resistance genes (ARGs) in agricultural soils from the Yangtze River Delta, China. Sci. Total Environ. 740:140001. doi: 10.1016/j.scitotenv.2020.140001
Tewari, G., Khati, D., Rana, L., Yadav, P., Pande, C., Bhatt, S., et al. (2016). Assessment of physicochemical properties of soils from different land use Systems in Uttarakhand, India. J. Chem. Eng. Chem. Res. 3, 1114–1118.
Thanner, S., Drissner, D., and Walsh, F. (2016). Antimicrobial Resistance in Agriculture. MBio 7, e02227–e02215. doi: 10.1128/mBio.02227-15
Wang, X., Liu, W., Li, Z., Teng, Y., Christie, P., and Luo, Y. (2020). Effects of long-term fertilizer applications on peanut yield and quality and plant and soil heavy metal accumulation. Pedosphere 30, 555–562. doi: 10.1016/S1002-0160(17)60457-0
Wang, F.-H., Qiao, M., Su, J.-Q., Chen, Z., Zhou, X., and Zhu, Y.-G. (2014). High throughput profiling of antibiotic resistance genes in urban park soils with reclaimed water irrigation. Environ. Sci. Technol. 48, 9079–9085. doi: 10.1021/es502615e
Wang, Y., Zhang, Z., Li, Y., Liang, C., Huang, H., and Wang, S. (2024). Available heavy metals concentrations in agricultural soils: relationship with soil properties and total heavy metals concentrations in different industries. J. Hazard. Mater. 471:134410. doi: 10.1016/j.jhazmat.2024.134410
World Health Organization (2024). Global research agenda for antimicrobial resistance in human health. World Health Organization. Available online at: https://www.who.int/publications/i/item/9789240102309 (accessed February 6, 2025)
Xie, W.-Y., Yuan, S.-T., Xu, M.-G., Yang, X.-P., Shen, Q.-R., Zhang, W.-W., et al. (2018). Long-term effects of manure and chemical fertilizers on soil antibiotic resistome. Soil Biol. Biochem. 122, 111–119. doi: 10.1016/j.soilbio.2018.04.009
Yang, F., Zhang, F., Li, H., Wu, H., Zhao, H., Cheng, X., et al. (2021). Contribution of environmental factors on the distribution of antibiotic resistance genes in agricultural soil. Eur. J. Soil Biol. 102:103269. doi: 10.1016/j.ejsobi.2020.103269
Zalewska, M., Błażejewska, A., Czapko, A., and Popowska, M. (2021). Antibiotics and antibiotic resistance genes in animal manure – consequences of its application in agriculture. Front. Microbiol. 12:610656. doi: 10.3389/fmicb.2021.610656
Zhang, Y., Cheng, D., Zhang, Y., Xie, J., Xiong, H., Wan, Y., et al. (2021). Soil type shapes the antibiotic resistome profiles of long-term manured soil. Sci. Total Environ. 786:147361. doi: 10.1016/j.scitotenv.2021.147361
Zhang, Y., Hao, X., Thomas, B. W., McAllister, T. A., Workentine, M., Jin, L., et al. (2023). Soil antibiotic resistance genes accumulate at different rates over four decades of manure application. J. Hazard. Mater. 443:130136. doi: 10.1016/j.jhazmat.2022.130136
Zhang, Y.-J., Hu, H.-W., Yan, H., Wang, J.-T., Lam, S. K., Chen, Q.-L., et al. (2019). Salinity as a predominant factor modulating the distribution patterns of antibiotic resistance genes in ocean and river beach soils. Sci. Total Environ. 668, 193–203. doi: 10.1016/j.scitotenv.2019.02.454
Zhang, X., Li, J., Shao, L., Qin, F., Yang, J., Gu, H., et al. (2023). Effects of organic fertilizers on yield, soil physico-chemical property, soil microbial community diversity and structure of Brassica rapa var. Chinensis. Front. Microbiol. 14:1132853. doi: 10.3389/fmicb.2023.1132853
Zhang, D., Li, J., Zhang, G., and Xu, Y. (2025). Decoding the trajectory of antibiotic resistance genes in saline and alkaline soils: insights from different fertilization regimes. Environ. Int. 195:109247. doi: 10.1016/j.envint.2024.109247
Zhang, F., Zhao, X., Li, Q., Liu, J., Ding, J., Wu, H., et al. (2018). Bacterial community structure and abundances of antibiotic resistance genes in heavy metals contaminated agricultural soil. Environ. Sci. Pollut. Res. 25, 9547–9555. doi: 10.1007/s11356-018-1251-8
Zhao, Y., Cocerva, T., Cox, S., Tardif, S., Su, J.-Q., Zhu, Y.-G., et al. (2019). Evidence for co-selection of antibiotic resistance genes and mobile genetic elements in metal polluted urban soils. Sci. Total Environ. 656, 512–520. doi: 10.1016/j.scitotenv.2018.11.372
Zheng, D., Yin, G., Liu, M., Hou, L., Yang, Y., Van Boeckel, T. P., et al. (2022). Global biogeography and projection of soil antibiotic resistance genes. Sci. Adv. 8:eabq8015. doi: 10.1126/sciadv.abq8015
Zhu, Y.-G., Johnson, T. A., Su, J.-Q., Qiao, M., Guo, G.-X., Stedtfeld, R. D., et al. (2013). Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. 110, 3435–3440. doi: 10.1073/pnas.1222743110
Keywords: antimicrobial resistance, mobile genetic elements, metals, co-selection, soil, physico-chemical properties
Citation: Szadziul M, Goryluk-Salmonowicz A and Popowska M (2025) The link between antibiotic resistance level and soil physico-chemical properties. Front. Microbiol. 16:1584660. doi: 10.3389/fmicb.2025.1584660
Edited by:
Razi Ahmad, Indian Institute of Technology Delhi, IndiaReviewed by:
Anuradha Goswami, University of Alabama at Birmingham, United StatesDuc Phan, University of Texas at San Antonio, United States
Copyright © 2025 Szadziul, Goryluk-Salmonowicz and Popowska. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Magdalena Popowska, bWEucG9wb3dza2FAdXcuZWR1LnBs