 Yao Tang
Yao Tang Bo Liu
Bo Liu Aijia Ma
Aijia Ma Bo Wang
Bo Wang Huaiyu Xiong
Huaiyu Xiong Yi Zhou
Yi Zhou Jing Yang
Jing Yang Yan Kang
Yan Kang- 1Department of Critical Care Medicine, West China Hospital of Sichuan University, Chengdu, China
- 2West China Hospital, Institutes for Systems Genetics, Sichuan University, Chengdu, China
The lung microbiota, present in healthy individuals, undergoes alterations in different diseases and is closely linked to changes in both systemic and alveolar immunity. These interactions play a crucial role in the onset and progression of numerous diseases. Acute respiratory distress syndrome (ARDS), one of the most severe conditions encountered in intensive care units (ICU), is characterized by high incidence and mortality rates. The pathophysiology of ARDS involves complex mechanisms, including the activation and dysregulation of overlapping pathways related to injury, inflammation, and coagulation, both locally in the lungs and systemically. Notably, alterations in the microbiota may contribute to the pathogenesis of ARDS. Emerging evidence suggests that changes in the lung microbiota are associated with ARDS development, often marked by increased bacterial burden, reduced microbial diversity, and shifts in microbiota composition. In this review, we focus on the regulatory roles of the lung microbiota in ARDS and their therapeutic potential.
1 Introduction
Advances in culture-independent microbiology, particularly high-throughput sequencing, have revealed that the lungs harbor complex and diverse microbial communities in healthy individuals (Charlson et al., 2011; Dickson et al., 2016a; Dickson et al., 2017), overturned the long-held belief that lungs are sterile (Laurenzi et al., 1961; Pecora, 1963).
Dysbiosis refers to the disruption of the balance between beneficial and harmful microbes, with a relative increase or decrease in certain microbial populations. Lung microbiota dysbiosis can lead to immune imbalances, resulting in excessive inflammation and subsequent tissue damage (Yang et al., 2020). In chronic respiratory diseases, the diversity of the lung microbiome is a predictor of mortality in chronic obstructive pulmonary disease (COPD) (Leitao Filho et al., 2019), and bacterial burden is linked to disease progression and mortality in idiopathic pulmonary fibrosis (IPF) (Molyneaux et al., 2014; O’Dwyer et al., 2019). Additionally, alterations in community composition predict exacerbations in bronchiectasis (Rogers et al., 2014).
In contrast to chronic respiratory diseases, research on the lung microbiome in acute respiratory failure remains in its early stages. ARDS is a common and serious lung problem that often leads to long-term ventilator use and death in critically ill patients. It is characterized by protein-rich pulmonary edema, hypoxemia, and alveolar inflammation. These pathological features may be driven by changes in the pulmonary microbiome. Conversely, the dysbiosis of lung microbiota may result from alveolar nutrient availability following edema onset (Ruff et al., 2020).
Despite advances in optimizing mechanical ventilation settings and antimicrobial therapies, adjunctive treatment options for ARDS remain limited. The lung microbiome represents an underexplored factor contributing to clinical variation in critical illness. A comprehensive understanding of its mechanistic role in ARDS could facilitate the development of targeted therapeutic interventions.
2 Lung microbiota in healthy lung
In most healthy individuals, the lung microbiota harbors a low density but exhibits significant diversity, with various interacting microbial communities. The adult lung microbiota in these individuals is primarily dominated by key genera within the phyla Firmicutes and Bacteroidetes, forming a “core microbiota” of the lungs (Man et al., 2017). This core microbiota, consisting of genera such as Streptococcus, Prevotella, Veillonella, Fusobacterium, and Haemophilus, plays a crucial role in maintaining lung homeostasis (Man et al., 2017; Montassier et al., 2023).
Lung immune cells, especially subpopulations of alveolar macrophages and dendritic cells, exert immunoregulatory functions by promoting the generation of regulatory T cells (Soroosh et al., 2013) and secreting anti-inflammatory molecules such as IL-10, transforming growth factor-beta (TGF-β), and prostaglandin E2 (Hussell and Bell, 2014). Resident microorganisms are integral to maintaining pulmonary immune homeostasis through continuous dialog among lung microbiota, immune cells and airway epithelial cells, which are equipped with pattern recognition receptors. However, the composition of the lung microbiota undergoes significant changes in response to pulmonary pathologies, which can disrupt immune homeostasis and influence the progression of these diseases (Lira-Lucio et al., 2020; Sommariva et al., 2020). Emerging evidence suggests that the lung microbiota plays a critical role in shaping the risk and outcomes of pulmonary diseases by modulating both innate and adaptive immune responses (Khatiwada and Subedi, 2020).
Streptococcus may indirectly promote the excessive production of CXCL8 (interleukin-8) through pulmonary microbiota dysbiosis, with elevated CXCL8 levels in sputum correlating with increased severity of COPD (Wang et al., 2023).
Prevotella, a key part of the airway microbiota, helps activate the innate immune system and protects the respiratory tract. Horn et al. demonstrated that Prevotella facilitated the rapid clearance of Streptococcus pneumoniae from the lungs and improved the outcomes of S. pneumoniae infection by activating neutrophils. Effective neutrophil-mediated clearance requires the recognition of toll-like receptor (TLR) 2, the induction of the pro-inflammatory cytokine tumor necrosis factor-alpha (TNF-α), and the regulation of inflammation by the anti-inflammatory cytokine IL-10 (Horn et al., 2022).
Furthermore, a study showed that specific lung bacteria, including Prevotella spp. and Veillonella spp., were associated with an increased number of lymphocytes in bronchoalveolar lavage fluid (BALF), TH17 cell-mediated lung inflammation, and a reduced TLR4 response by alveolar macrophages (Segal et al., 2016). Although the lung microbiome likely contributes significantly to mucosal immune homeostasis, its precise role in establishing and maintaining respiratory health remains unclear (Man et al., 2017).
Both chronic lung disease and acute lung injury/ARDS are associated with airway dysbiosis and enrichment of Pseudomonas. However, in chronic lung disease, Haemophilus, Streptococcus, and Moraxella are primarily enriched, while in acute lung injury/ARDS, the changes in the lung microbiome are mainly characterized by the enrichment of gut-associated bacteria (i.e., Bacteroides spp.) (Yi et al., 2022).
3 Microbiome features in patients with traditional ARDS
This section provides an overview of the current understanding of the lung microbiome in traditional ARDS and COVID-19-associated ARDS (Table 1), and a few comparisons between bacterial taxa in traditional ARDS and COVID-19 ARDS (Table 2).
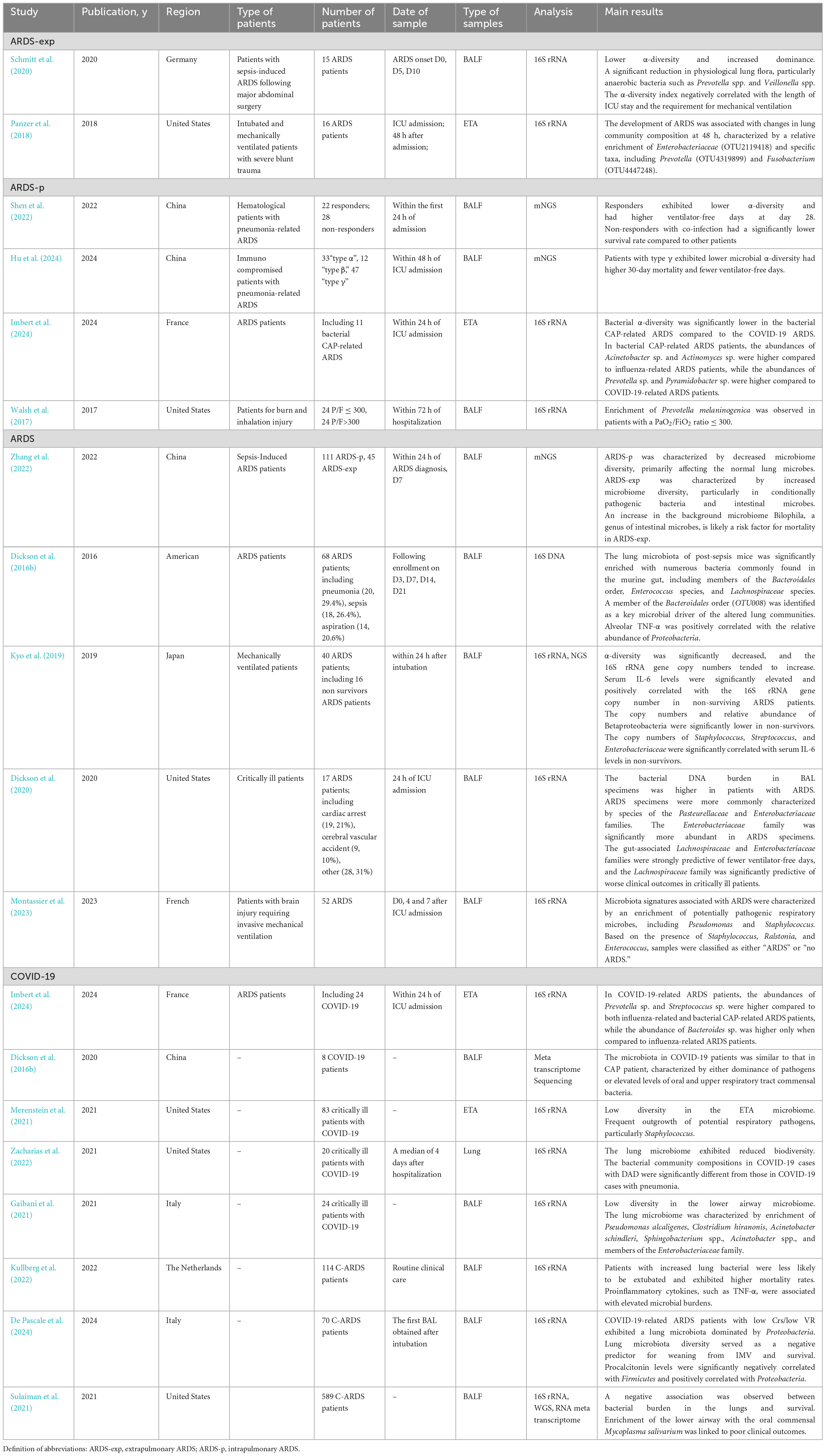
Table 1. Studies concerning lung microbiota of mechanically ventilated patient in traditional and COVID-19 ARDS.
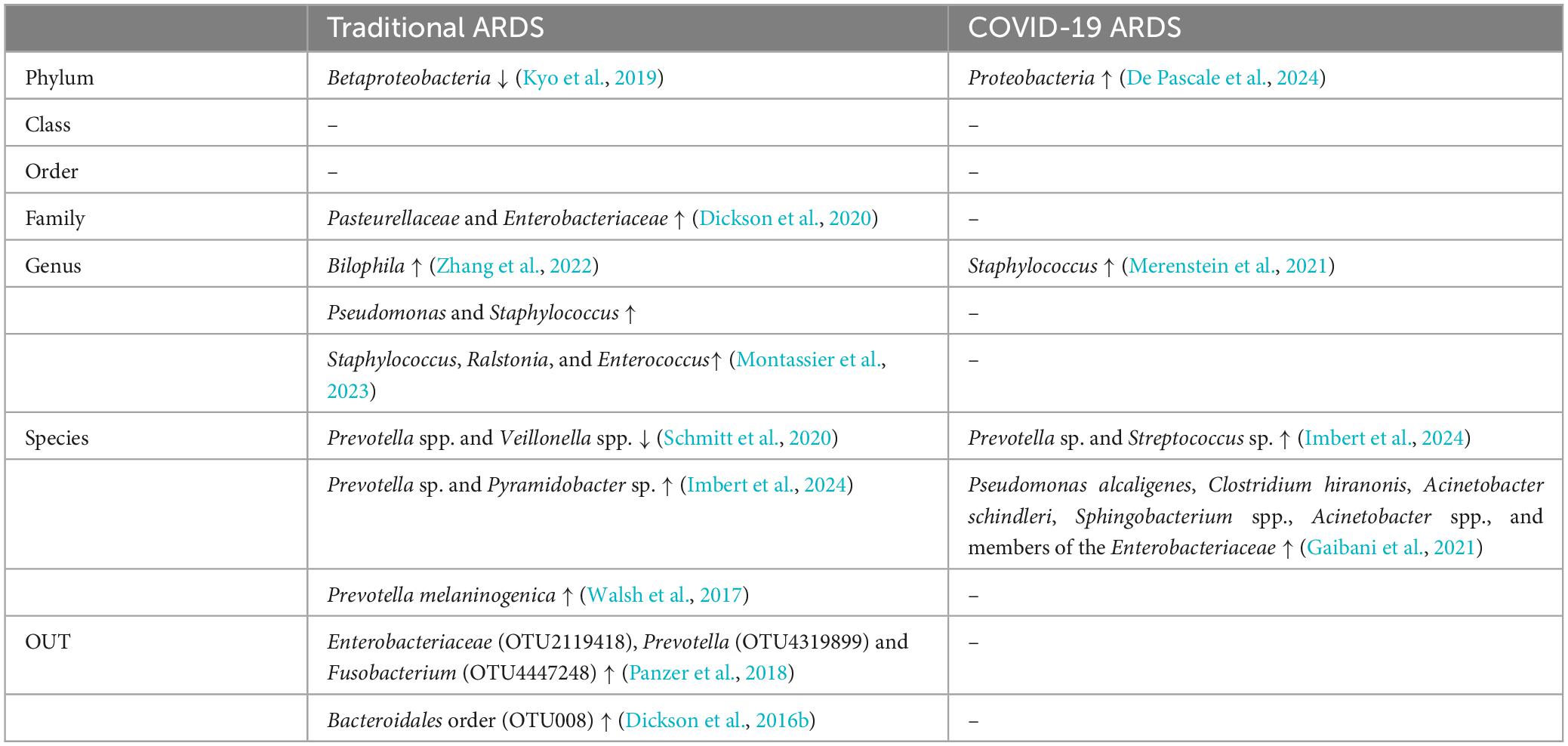
Table 2. Comparisons between bacterial taxa in Traditional ARDS and COVID-19 ARDS.
3.1 Ecological metrics
Several studies have reported that the bacterial DNA burden is higher in patients with ARDS compared to those without ARDS (Dickson et al., 2020; Kyo et al., 2019). Additionally, Kyo et al. found that serum IL-6 levels were significantly elevated in non-survivor ARDS patients and positively correlated with the 16S rRNA copy number (Kyo et al., 2019). Furthermore, Dickson et al. demonstrated that patients with an increased lung bacterial DNA burden had fewer ventilator-free days in critically ill patients (Dickson et al., 2020).
Most studies have shown that alpha diversity is significantly reduced in patients with ARDS (Imbert et al., 2024; Kyo et al., 2019; Schmitt et al., 2020; Zhang et al., 2022). Another study reported that microbial diversity in the lung microbiome of patients with septic ARDS declines progressively over time (Li et al., 2024). However, Zhang et al. found that patients with extrapulmonary infection-induced ARDS exhibited increased microbial diversity (Zhang et al., 2022).
Shen and colleagues observed that responders to early corticosteroid therapy in hematological patients with pneumonia-associated ARDS had lower alpha diversity compared to non-responders. They also found a negative correlation between alpha diversity and levels of inflammatory markers (serum IL-6, IL-8, TNF-α) and CRP in the responders (Shen et al., 2022). In contrast, the microbiota signatures of immunocompromised patients with no obvious inflammatory symptoms and more severe oxygenation failure were characterized by lower alpha diversity and distinct microbial compositions compared to other immunocompromised patients. Hu et al. reported that gut-associated bacteria were more abundant in patients with type α, which is characterized by more active inflammation (Hu et al., 2024).
Interestingly, Walsh et al. found significant differences not in the species dominating the overall community but in the less abundant taxa in patients with burn and inhalation injuries (Walsh et al., 2017). While these taxa did not differ in microbial diversity between patient groups, they exhibited differences in functional diversity, which ultimately had a greater impact on patient outcomes (Dickson et al., 2016a).
Several studies have reported that lower alpha diversity in the lung microbiota is associated with fewer ventilator-free days (De Pascale et al., 2024; Hu et al., 2024; Schmitt et al., 2020). However, Shen and colleagues detected that the responders of corticosteroids had lower alpha diversities and higher ventilator free days in hematological patients with pneumonia-associated ARDS (Shen et al., 2022). One possible explanation is that corticosteroid treatment may alter the lung microbiota composition, though this aspect was not further explored in their study.
3.2 Differential flora
In ARDS patients, the abundance of Prevotella spp. was found to increase in both direct and indirect lung injury-induced ARDS (Imbert et al., 2024; Panzer et al., 2018; Walsh et al., 2017). However, Schmitt et al. reported a significant reduction in physiological lung flora and mostly anaerobic bacteria, such as Prevotella spp. or Veillonella spp., in patients with sepsis-induced ARDS (Schmitt et al., 2020).
In patients with bacterial community-acquired pneumonia (CAP)-related ARDS, the levels of Acinetobacter spp. and Actinomyces spp. were higher compared to those with influenza-related ARDS. When compared to COVID-19-related ARDS patients, only the abundances of Prevotella spp. and Pyramidobacter spp. were found to be elevated (Imbert et al., 2024).
Additionally, several studies have identified gut-associated Bacteroides genus (OTU009) (Dickson et al., 2016b), Pasteurellaceae (Dickson et al., 2020), Enterobacteriaceae (Dickson et al., 2020; Panzer et al., 2018), and Fusobacterium (OTU4319899) (Panzer et al., 2018) were common and abundant in ARDS patients. Furthermore, Dickson et al. found that the relative abundance of Proteobacteria was positively correlated with alveolar TNF-α levels, while the typically abundant Bacteroidetes phylum showed a negative correlation with alveolar TNF-α concentrations. Furthermore, the gut-associated Bacteroides genus (OTU009) was associated with serum TNF-α levels (Dickson et al., 2016b). Four years later, the same team identified the gut-associated Lachnospiraceae and Enterobacteriaceae families as the taxa most strongly predictive of fewer ventilator-free days (Dickson et al., 2020).
Panzer et al. explored the relationship between microbiota composition at 48 h and inflammation, finding that this relationship was driven by the presence or absence of specific taxa. They suggested that the loss or overgrowth of certain bacteria during the first 48 h in the ICU might contribute to the inflammation observed at this time. Their study revealed that lung bacterial community variation at 0 h was significantly linked to endothelial injury (measured by soluble intercellular adhesion molecule-1), epithelial injury [vascular endothelial growth factor (VEGF)], and inflammation (IL-8) in critically ill blunt trauma patients. At 48 h, community variation was also associated with elevated IL-6 and IL-8 levels. Interestingly, the composition of the microbiota at 0 h was significantly correlated with 48-hour levels of VEGF, receptor for advanced glycation end-products (RAGE), angiopoietin-2 (ANG-2), pentraxin 3 (PENT3), and IL-8 (Panzer et al., 2018).
Montassier et al. identified microbiota signatures associated with ARDS, characterized by an enrichment of potentially pathogenic respiratory microbes, including Pseudomonas and Staphylococcus, through data harmonization and the pooling of individual patient data. Furthermore, patients with the presence of Staphylococcus, Ralstonia and Enterococcus, had a lower probability of successful extubation (Montassier et al., 2023). Zhang et al. observed that an increase in Escherichia coli and Staphylococcus aureus in the lungs may serve as risk factors for mortality in ARDS induced by intrapulmonary infections. While no specific pathogens were linked to prognosis, they demonstrated that the increased presence of Bilophila, a genus of intestinal bacteria, could be a potential risk factor for death in ARDS caused by extrapulmonary infections. In contrast, Hydrobacter might play a protective role in ARDS induced by intrapulmonary infections (Zhang et al., 2022). However, it remains unclear whether Hydrobacter is part of the normal lung microbiota or whether it functions independently or synergistically with other microbiomes.
In addition, Kyo et al. discovered that the ratio of the relative abundance of Betaproteobacterial operational taxonomic units (OTUs) to the maximum relative abundance of three other OTUs (Staphylococcus, Streptococcus, and Enterobacteriaceae) was significantly associated with hospital mortality in ARDS patients. They found that the Betaproteobacteria class was strongly negatively correlated with serum IL-6 levels in the non-survivor. Conversely, Staphylococcus and Streptococcus at the genus level and Enterobacteriaceae at the family level were significantly positively correlated with serum IL-6 levels in the non-survivors (Kyo et al., 2019).
4 Microbiome features in severe COVID-19 ARDS patients
4.1 Ecological metrics
Dysbiosis of the lung microbiome has been observed in COVID-19 patients (Merenstein et al., 2021; Shen et al., 2020). Compared to healthy lung samples, lung microbiota from mechanically ventilated COVID-19 patients exhibited lower diversity (Gaibani et al., 2021; Merenstein et al., 2021; Zacharias et al., 2022). Pascale et al. further reported that patients with reduced lung microbiota diversity had a longer duration of invasive mechanical ventilation (IMV) and higher mortality (De Pascale et al., 2024).
Interestingly, Kullberg et al. found a significant association between bacterial burden and beta diversity in COVID-19-related ARDS among mechanically ventilated patients, suggesting that bacterial overgrowth may drive compositional changes in critically ill lungs. They also discovered that increased bacterial DNA burden correlated with elevated alveolar concentrations of proinflammatory cytokines (TNF-α, IL-6, IL-1). Furthermore, they reported that the overall lung microbiota composition, rather than individual bacterial genera, was linked to successful extubation in COVID-19-related ARDS (Kullberg et al., 2022). Several studies have also indicated that higher bacterial burden is negatively associated with successful extubation (Kullberg et al., 2022) and survival (Sulaiman et al., 2021) in mechanically ventilated patients with COVID-19-related ARDS. These findings are consistent with those observed in traditional ARDS.
4.2 Differential flora
Merenstein et al. found that lung microbiome specimens from COVID-19 patients, sampled via endotracheal aspirates (ETA), showed frequent outgrowth of potential respiratory pathogens, particularly Staphylococcus (Merenstein et al., 2021). Additionally, Gaibani et al. reported that the lung microbiota of critically ill COVID-19 patients was enriched with Pseudomonas alcaligenes, Clostridium hiranonis, Acinetobacter schindleri, Sphingobacterium spp., Acinetobacter spp., and Enterobacteriaceae. In contrast, COVID-19-negative patients displayed a higher abundance of lung commensal bacteria, such as Haemophilus influenzae, Veillonella dispar, Granulicatella spp., Porphyromonas spp., and Streptococcus spp. (Gaibani et al., 2021). Compared to influenza-related and bacterial CAP-related ARDS patients, Prevotella and Streptococcus were more abundant in COVID-19-related ARDS patients, while Bacteroides was only more abundant compared to influenza-related ARDS patients (Imbert et al., 2024). However, Shen et al. reported that the microbiota in COVID-19 patients was similar to that in CAP patients, characterized by either the dominance of pathogens or elevated levels of oral and upper respiratory tract commensal bacteria (Shen et al., 2020).
Moreover, Pascale et al. observed that Firmicutes dominated the lung microbiota of patients with high respiratory system compliance/predicted body weight (Crs) phenotype, while in COVID-19-related ARDS patients with low Crs/low ventilatory ratio (VR), Proteobacteria predominated. They also found a positive correlation between serum procalcitonin and Proteobacteria, and a negative correlation with Firmicutes (De Pascale et al., 2024). Additionally, Sulaiman et al. discovered that the oral commensal Mycoplasma salivarum was enriched in patients who died or required mechanical ventilation for more than 28 days. In contrast, Prevotella oris was more abundant in patients with mechanical ventilation duration ≤ 28 days (Sulaiman et al., 2021).
4.3 Microbiome features in acute lung injury animal model
This section reviews current understanding of the lung microbiome in ALI/ARDS animal model (Table 3).
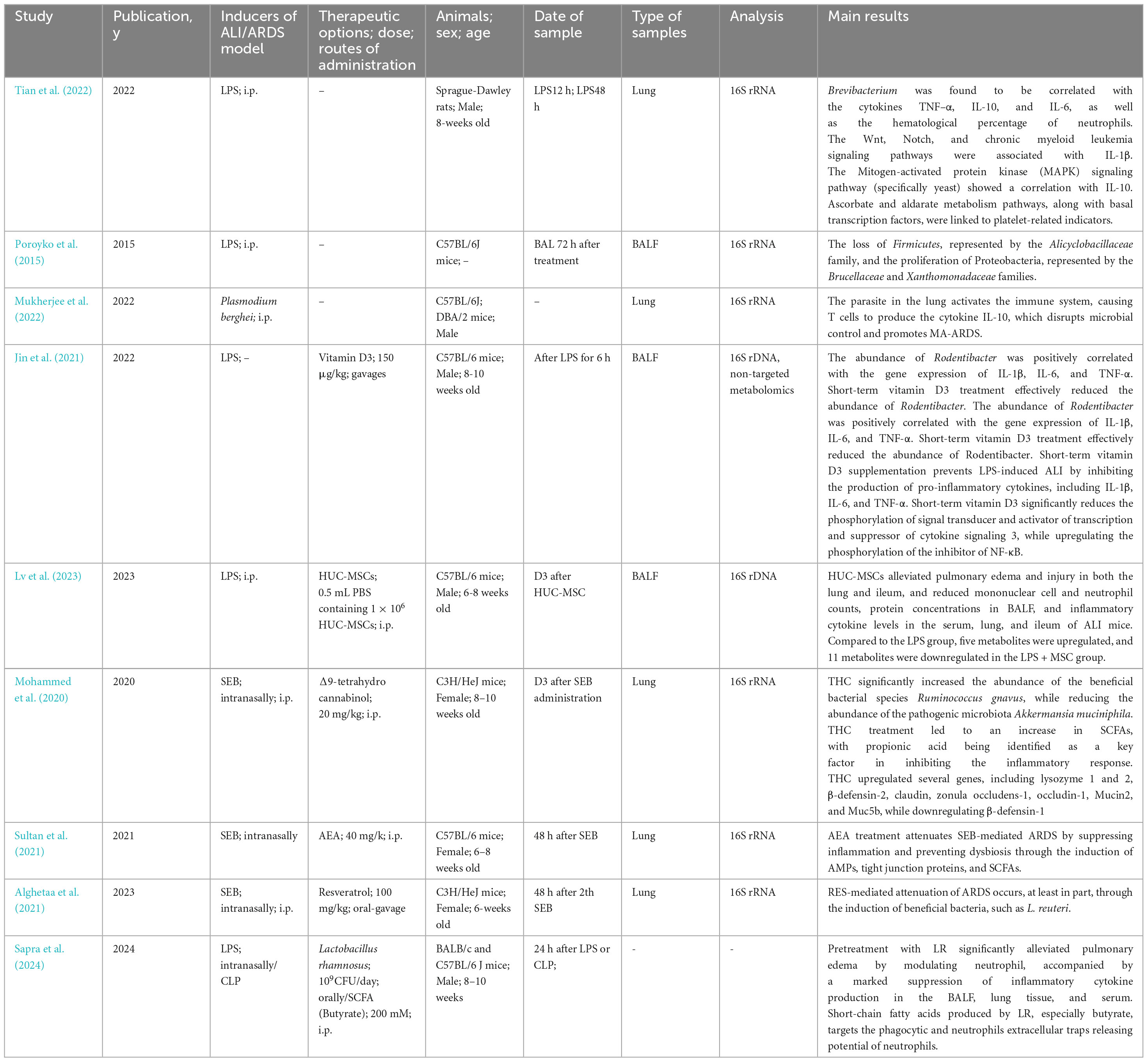
Table 3. Studies concerning lung microbiota of ALI/ARDS in animal model and Therapeutic options.
4.4 Ecological metrics
Poroyko et al. observed a fivefold increase in bacterial load in a mouse model 72 h after intratracheal administration of sterile bacterial wall lipopolysaccharide (LPS) (Poroyko et al., 2015). In addition, Tian et al. found a significant reduction in the volatility of α-diversity 48 hours after LPS was injected intraperitoneally in rats (Tian et al., 2022).
In contrast, Dickson et al. reported that the lungs of post-sepsis mice exhibited increased bacterial diversity 3 days after exposure in a mouse model of lung injury induced by cecal ligation and puncture (CLP) (Dickson et al., 2016b).
4.5 Differential flora
In addition, Dickson et al. found that the lung microbiota of post-sepsis mice were significantly enriched with numerous bacteria commonly found in the murine gut, including members of the Bacteroidales order, Enterococcus species, and Lachnospiraceae (sp.). Notably, a member of the Bacteroidales order (OTU008) was identified as a key driver of community alterations (Dickson et al., 2016b).
Poroyko et al. observed that the primary microbial response to LPS-induced ALI was characterized by a loss of Firmicutes—represented by the family Alicyclobacillaceae—and a proliferation of Proteobacteria, particularly from the Brucellaceae and Xanthomonadaceae families (Poroyko et al., 2015).
Previous studies have shown that Stenotrophomonas directly contributes to the inflammatory process and impairs respiratory function (Di Bonaventura et al., 2010). Similarly, Ochrobactrum has been implicated in lung inflammation (Dawkins et al., 2006; Naik et al., 2013). Additionally, Poroyko et al. detected bacterial substrates suitable for Ochrobactrum anthropi and Stenotrophomonas maltophilia in BALF, identifying 35 lung metabolites that could serve as substrates. Notably, they found eight metabolites shared by both pathogens, including four carbohydrates, two nucleotide/nucleoside compounds, and two organic acids (citrate and lactate). Furthermore, six metabolites—putrescine, desmosterol, tetracosanoic acid, uric acid, 3-hydroxycholestane, and lactic acid—were significantly elevated in samples from injured lungs (Poroyko et al., 2015).
Additionally, Mukherjee et al. found that the adherence of Plasmodium-infected red blood cells to the vascular endothelium induces persistent immune activation, leading to the production of the anti-inflammatory cytokine IL-10 by T cells in the lung. Elevated IL-10 levels promote bacterial expansion in the lung, thereby contributing to the development of malaria-associated acute respiratory distress syndrome (MA-ARDS) (Mukherjee et al., 2022).
Tian et al. found that Brevibacterium was positively correlated with serum cytokines TNF-α, IL-10, and IL-6 levels. Additionally, they observed that the functionality of the lung microbiota appeared to be more stable and less variable compared to its composition, with only three differential pathways identified in terms of functional abundance. Changes in the abundance of the ABC transporter pathway may reflect the adaptation of lung bacteria to inflammatory conditions.
The upregulation of bacterial chemotaxis functions in the lung microbiota may result from bacterial adaptation to the edema environment. The reduction in the pentose phosphate pathway could indicate a microbiota with lower antioxidant capacities and diminished DNA repair abilities following lung injury. Regarding lung microbiota functionality, the abundances of four signaling pathways—Wnt (ko04310), Notch (ko04330), chronic myeloid leukemia (ko05220), and MAPK-yeast (ko04011)—were strongly negatively correlated with serum IL-1β and IL-10 levels, suggesting that these pathways may play a role in the interaction between the lung microbiota and host immunity (Tian et al., 2022).
4.6 Prospect of therapy
Here are some potential therapeutic targets and promising drugs for attenuating ALI/ARDS by modulating the lung microbiota, as illustrated in Figure 1.
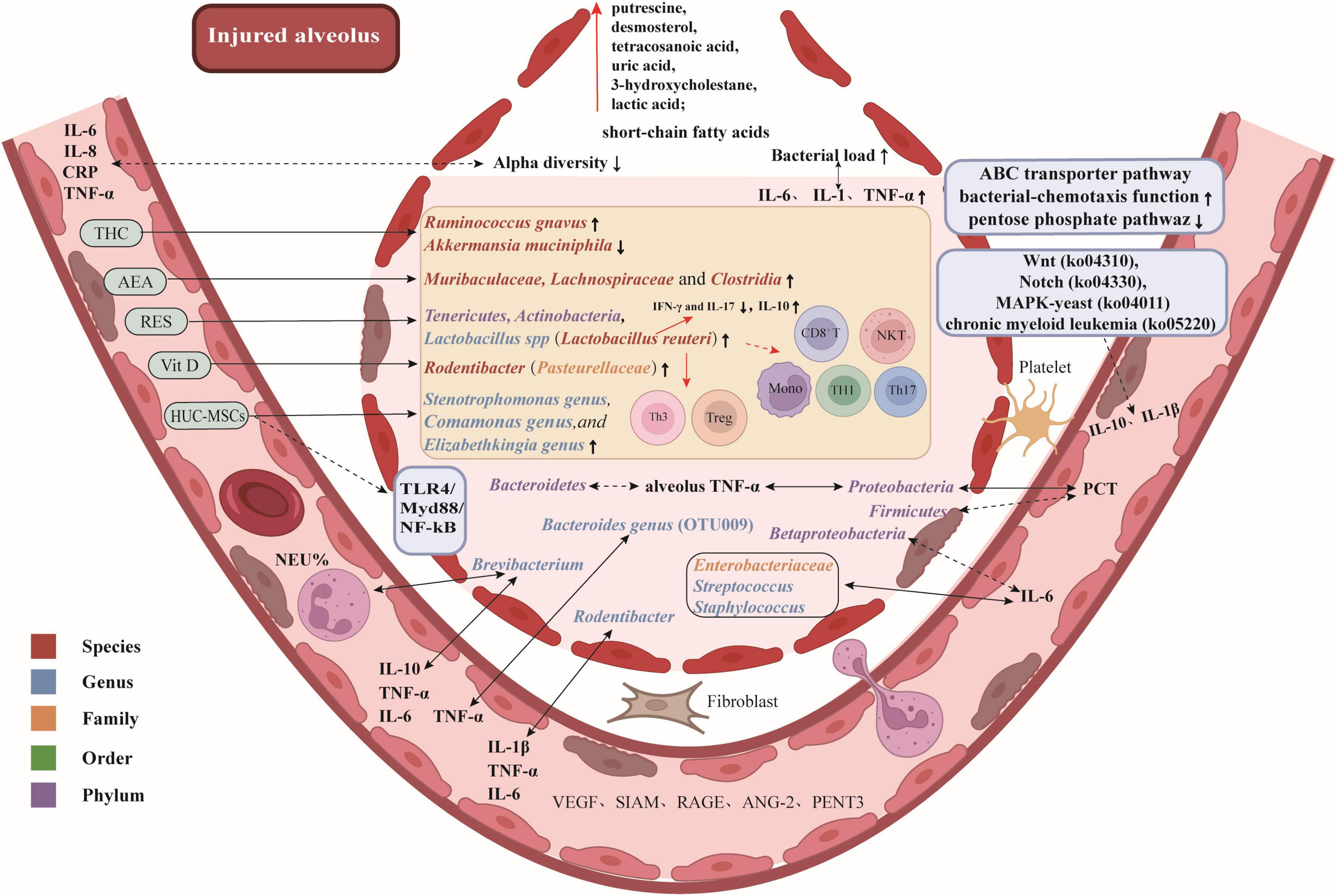
Figure 1. Potential therapeutic targets and promising drugs for attenuating ALI/ARDS by modulating lung microbiota. Gut-associated bacteria, bacteria that are enriched in the gut, such as Bacteroides spp.; Responders to early corticosteroid therapy, patients who showed a significant improvement in their PaO2/FiO2 ratio after receiving corticosteroid treatment. Arterial blood gas measurements were obtained upon ICU admission, prior to the initial administration of corticosteroids, and repeated 24 hours later, following (or prior to) corticosteroid treatment. The difference in PaO2/FiO2 between these two time points, referred to as ΔPaO2/FiO2, was calculated. Patients with a ΔPaO2/FiO2 ≥ 150 mmHg were categorized as responders, while those with a ΔPaO2/FiO2 < 150 mmHg were classified as non-responders.
4.7 The target of potential treatment: microbial community
4.7.1 Anandamide
Endocannabinoids, which are host-derived lipid hormones, possess potent anti-inflammatory properties (Ahmed et al., 2021; Jackson et al., 2014; Osafo et al., 2021; Pandey et al., 2009; Sido et al., 2016). Some studies have suggested that endocannabinoids can modulate the diversity of the gut microbiota (Finn et al., 2021; Minichino et al., 2021). Sultan et al. demonstrated that staphylococcal enterotoxin B (SEB) induces alterations in both the lung and gut microbiota, particularly leading to an overgrowth of pathogenic bacteria such as Pseudomonas. However, AEA treatment reversed these changes by promoting the induction of antimicrobial peptides (AMPs) and tight junction proteins. Additionally, AEA treatment enhanced the abundance of beneficial bacteria in the lungs of mice, including Muribaculaceae, Lachnospiraceae, and Clostridia, which are known to produce butyrate (Sultan et al., 2021).
4.8 The target of drug intervention: microbial community and immune system
4.8.1 Δ9-tetrahydrocannabinol
Δ9-Tetrahydrocannabinol (THC), a cannabinoid found in Cannabis sativa L., is well known for its anti-inflammatory properties (Nagarkatti et al., 2009). Studies have shown that THC and cannabidiol can influence gut microbiota in experimental models of autoimmune diseases (Al-Ghezi et al., 2019; Cluny et al., 2015). Mohammed et al. found that THC administration protected mice from SEB-induced mortality and attenuated ARDS by suppressing lung inflammation and modulating microbial dysbiosis and their metabolites in the lungs. Specifically, they observed that THC increased the abundance of the beneficial bacterial species Ruminococcus gnavus in Firmicutes phylum, while decreasing the abundance of Akkermansia muciniphila (Mohammed et al., 2020). R. gnavus is a human gut symbiont known to enhance the expression of glycoproteins (Graziani et al., 2016), which are critical for maintaining lung tissue integrity, preventing vascular leaks, and defending against bacterial pneumonia (Johansson et al., 2011; Roy et al., 2014). In contrast, A. muciniphila, commonly found in the intestines of humans and animals, can elevate the expression of pro-inflammatory cytokines (Ganesh et al., 2013) and has a specialized ability to degrade mucin (Derrien et al., 2017; Ganesh et al., 2013).
Additionally, THC was found to significantly increase the levels of short-chain fatty acids (SCFAs) such as propionic acid, butyric acid, and acetic acid in the colon. Notably, propionic acid inhibited the production of pro-inflammatory cytokines while promoting the release of anti-inflammatory cytokines. Furthermore, investigations have shown that R. gnavus produces propionate and propanol as end products of metabolism (Crost et al., 2013). These findings suggest that THC may promote the abundance of beneficial bacterial species like R. gnavus, enhance the production of SCFAs such as propionic acid, and suppress pro-inflammatory responses, thereby attenuating acute lung injury.
4.8.2 Human umbilical cord mesenchymal cells
A previous study has demonstrated that HUC-MSCs promote the expression of PD-L1 on macrophages and attenuate ALI in mice (Tu et al., 2022). Lv et al. observed that HUC-MSCs can improve pulmonary edema, lung injury, and endothelial barrier function in LPS-induced ALI mice, while also reducing the expression of inflammatory cytokines in both serum and lung tissue. Additionally, HUC-MSC treatment enhanced the integrity of the endothelial barrier in the lungs. More importantly, they found that HUC-MSCs ameliorated ALI by reducing the abundance of pathogenic bacteria, including Stenotrophomonas, Comamonas, and Elizabethkingia genera, in the BALF of mice. Furthermore, Haemophilus may play a critical role in the improvement of ALI by HUC-MSCs. Moreover, Lactobacillus, Bacteroides, and an unidentified Rikenellaceae genus were identified as potential biomarkers for evaluating the therapeutic efficacy of HUC-MSCs. The study also revealed that HUC-MSC treatment of ALI mice significantly attenuated the expression of the TLR4/Myd88/NF-κB signaling pathway in lung tissue. Metabolomic analysis further indicated that MSC treatment upregulated 5 metabolites and downregulated 11 metabolites, primarily related to purine metabolism and taste transduction signaling pathways. Notably, metabolites involved in drug metabolism, tyrosine metabolism, autophagy, and endocytosis were significantly associated with the improvement of ALI by HUC-MSC treatment (Lv et al., 2023).
4.8.3 Vitamin D3
Jin et al. observed an increased relative abundance of Rodentibacter in the BALF of LPS-treated mice. Short-term vitamin D3 treatment was found to effectively reduce Rodentibacter abundance and attenuate ALI. Furthermore, they noted a positive correlation between the abundance of Rodentibacter and serum levels of pro-inflammatory cytokines, including IL-1β, IL-6, and TNF-α (Jin et al., 2021).
Rodentibacter, primarily found in rodents, belongs to the Pasteurellaceae family (Adhikary et al., 2017; Benga et al., 2018), and has been reported as an opportunistic pathogen, or even a primary pathogen, under certain conditions (Benga et al., 2015).
4.9 The target of potential treatment: microbial community and immune system
4.9.1 Resveratrol
Resveratrol (RES), a stilbenoid, is well known for its potent anti-inflammatory and antioxidant properties (Chen et al., 2020; Salehi et al., 2018). Studies have shown that RES can significantly alter gut microbiota composition (Alrafas et al., 2019; Li F. et al., 2020), and alleviate SEB-induced ARDS in mice (Alghetaa et al., 2018; Rieder et al., 2012). Alghetaa et al. reported that SEB exposure promotes the growth of pathogenic bacteria, such as those from the Proteobacteria phylum and Propionibacterium acnes species, in lung microbiota, contributing to ARDS pathogenesis. Their findings indicated that RES treatment significantly enhanced lung microbiota diversity in SEB-exposed mice, increasing the abundance of beneficial bacteria, including the Tenericutes and Actinobacteria phyla, as well as the Lactobacillus genus, particularly Lactobacillus reuteri. Colonic microbiota transplantation (CMT) experiments further confirmed that RES-induced enrichment of beneficial bacteria, particularly L. reuteri, plays a crucial role in mitigating ARDS. Specifically, L. reuteri reduced the number of lung-infiltrating mononuclear cells, cytotoxic CD8+ T cells, NKT cells, Th1 cells, and Th17 cells, while increasing the proportions of regulatory T cells (Tregs) and Th3 cells. Furthermore, L. reuteri inhibited the SEB-induced production of pro-inflammatory cytokines such as IFN-γ and IL-17, while promoting the production of the anti-inflammatory cytokine IL-10 (Alghetaa et al., 2021).
L. reuteri is a well-studied probiotic bacterium found in various human organs and tissues. It has been shown to produce antimicrobial molecules, which inhibit the colonization of pathogenic microbes (Mu et al., 2018). Studies also suggest that L. reuteri can suppress the production of pro-inflammatory cytokines while enhancing the development and function of regulatory T cells (Tregs) (Li L. et al., 2020). Additionally, L. reuteri has been linked to increased butyric acid concentrations in the gut, contributing to improved gut health and immune regulation (Li L. et al., 2020; Liu et al., 2017).
4.9.2 Lactobacillus rhamnosus
Pretreatment with LR notably improved lung vascular permeability (edema) by modulating neutrophils, while also significantly decreasing the expression of inflammatory cytokines in the BALF, lungs, and serum in both pulmonary and extrapulmonary mouse models. From a mechanistic perspective, LR, through its short-chain fatty acids (with butyrate being the most potent and effective in improving the pathophysiology of both pulmonary and extrapulmonary ARDS), influences neutrophil phagocytosis and their capacity to release extracellular traps. Moreover, butyrate demonstrates enhanced potential in alleviating the pathophysiology of ARDS by reducing neutrophil infiltration into the lungs (Sapra et al., 2024).
5 Conclusion and prospects
The lung microbiome undergoes significant alterations in ARDS compared to healthy individuals, with these changes closely linked to disease severity. While the literature partially agrees on the specific microbial changes, certain patterns are consistently observed. ARDS is characterized by an increased bacterial burden and reduced microbial diversity. Patients with lower alpha diversity and higher bacterial loads in their lung microbiota tend to have fewer ventilator-free days and higher mortality rates. Physiological lung flora, such as Firmicutes, is notably diminished, while respiratory pathogens, including Staphylococcus, Streptococcus, Enterobacteriaceae, Pseudomonas, and Proteobacteria, proliferate in ARDS patients. Furthermore, the lung microbiome is closely associated with immune and inflammatory responses in ARDS. However, the impact of different initial infection sites on lung microbiome diversity and composition remains unclear, and study heterogeneity may significantly influence findings. We summarized that these studies involved populations with different causes of ARDS, varying in sample sizes, countries and regions, all of which may contribute to the inconsistency of the findings. Given that lower airway microbiota composition can vary depending on the underlying cause of ARDS, future research and experimental models should account for the etiology when investigating the role of the lung microbiome in ARDS pathophysiology.
Author contributions
YT: Writing – original draft. BL: Writing – original draft. AM: Writing – original draft. BW: Writing – review & editing. HX: Writing – original draft. YZ: Writing – review & editing. JY: Writing – review & editing. YK: Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by 1.3.5 Project for Disciplines of Excellence, West China Hospital, Sichuan University (ZYGD23012).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Abbreviations
ALI, acute lung injury; AEA, anandamide; AMPs, antimicrobial peptides; ANG-2, angiopoietin-2; ARDS, acute respiratory distress syndrome; ARDS-exp, extrapulmonary ARDS; ARDS-p, intrapulmonary ARDS; BALF, bronchoalveolar lavage fluid; CLP, cecal ligation and puncture; CMT, colonic microbiota transplantation; COPD, chronic obstructive pulmonary disease; COVID-19, corona virus disease 2019; Crs, respiratory system compliance; ETA, endotracheal aspirates; HUC-MSCs, human umbilical cord mesenchymal cells; ICU, intensive care unit; IMV, invasive mechanical ventilation; i.p., intraperitoneally; LPS, lipopolysaccharide; MA-ARDS, malaria-associated acute respiratory distress syndrome; MV, mechanical ventilation; OTUs, operational taxonomic units; PENT3, pentraxin 3; RAGE, receptor for advanced glycation end-products; RES, resveratrol; SCFAs, short-chain fatty acids; SEB, staphylococcal enterotoxin-b; THC, Δ9-tetrahydrocannabinol; TNF-α, tumor necrosis factor-α; TLR, toll-like receptor; VEGF, vascular endothelial growth factor; VR, ventilatory ratio.
References
Adhikary, S., Nicklas, W., Bisgaard, M., Boot, R., Kuhnert, P., Waberschek, T., et al. (2017). Rodentibacter gen. nov. including Rodentibacter pneumotropicus comb. nov., Rodentibacter heylii sp. nov., Rodentibacter myodis sp. nov., Rodentibacter ratti sp. nov., Rodentibacter heidelbergensis sp. nov., Rodentibacter trehalosifermentans sp. nov., Rodentibacter rarus sp. nov., Rodentibacter mrazii and two genomospecies. Int. J. Syst. Evol. Microbiol. 67, 1793–1806. doi: 10.1099/ijsem.0.001866
Ahmed, I., Rehman, S. U., Shahmohamadnejad, S., Zia, M. A., Ahmad, M., Saeed, M. M., et al. (2021). Therapeutic attributes of endocannabinoid system against neuro-inflammatory autoimmune disorders. Molecules 26:3389. doi: 10.3390/molecules26113389
Alghetaa, H., Mohammed, A., Sultan, M., Busbee, P., Murphy, A., Chatterjee, S., et al. (2018). Resveratrol protects mice against SEB-induced acute lung injury and mortality by miR-193a modulation that targets TGF-β signalling. J. Cell Mol. Med. 22, 2644–2655. doi: 10.1111/jcmm.13542
Alghetaa, H., Mohammed, A., Zhou, J., Singh, N., Nagarkatti, M., and Nagarkatti, P. (2021). Resveratrol-mediated attenuation of superantigen-driven acute respiratory distress syndrome is mediated by microbiota in the lungs and gut. Pharmacol. Res. 167:105548. doi: 10.1016/j.phrs.2021.105548
Al-Ghezi, Z. Z., Busbee, P. B., Alghetaa, H., Nagarkatti, P. S., and Nagarkatti, M. (2019). Combination of cannabinoids, delta-9-tetrahydrocannabinol (THC) and cannabidiol (CBD), mitigates experimental autoimmune encephalomyelitis (EAE) by altering the gut microbiome. Brain Behav. Immun. 82, 25–35. doi: 10.1016/j.bbi.2019.07.028
Alrafas, H. R., Busbee, P. B., Nagarkatti, M., and Nagarkatti, P. S. (2019). Resveratrol modulates the gut microbiota to prevent murine colitis development through induction of Tregs and suppression of Th17 cells. J. Leukoc. Biol. 106, 467–480. doi: 10.1002/JLB.3A1218-476RR
Benga, L., Benten, W. P., Engelhardt, E., Gougoula, C., and Sager, M. (2015). Spontaneous bacterial and fungal infections in genetically engineered mice: Is Escherichia coli an emerging pathogen in laboratory mouse? Berl. Munch Tierarztl. Wochenschr. 128, 278–284.
Benga, L., Sager, M., and Christensen, H. (2018). From the [Pasteurella] pneumotropica complex to Rodentibacter spp.: An update on [Pasteurella] pneumotropica. Vet. Microbiol. 217, 121–134. doi: 10.1016/j.vetmic.2018.03.011
Charlson, E. S., Bittinger, K., Haas, A. R., Fitzgerald, A. S., Frank, I., Yadav, A., et al. (2011). Topographical continuity of bacterial populations in the healthy human respiratory tract. Am. J. Respir. Crit. Care Med. 184, 957–963. doi: 10.1164/rccm.201104-0655OC
Chen, M., Fu, Q., Song, X., Muhammad, A., Jia, R., Zou, Y., et al. (2020). Preparation of resveratrol dry suspension and its immunomodulatory and anti-inflammatory activity in mice. Pharm. Biol. 58, 8–15. doi: 10.1080/13880209.2019.1699123
Cluny, N. L., Keenan, C. M., Reimer, R. A., Le Foll, B., and Sharkey, K. A. (2015). Prevention of diet-induced obesity effects on body weight and gut microbiota in mice treated chronically with Δ9-Tetrahydrocannabinol. PLoS One 10:e0144270. doi: 10.1371/journal.pone.0144270
Crost, E. H., Tailford, L. E., Le Gall, G., Fons, M., Henrissat, B., and Juge, N. (2013). Utilisation of mucin glycans by the human gut symbiont Ruminococcus gnavus is strain-dependent. PLoS One 8:e76341. doi: 10.1371/journal.pone.0076341
Dawkins, P., Robertson, A., Robertson, W., Moore, V., Reynolds, J., Langman, G., et al. (2006). An outbreak of extrinsic alveolitis at a car engine plant. Occup. Med. (Lond) 56, 559–565. doi: 10.1093/occmed/kql110
De Pascale, G., Posteraro, B., De Maio, F., Pafundi, P. C., Tanzarella, E. S., Cutuli, S. L., et al. (2024). Lung microbiota composition, respiratory mechanics, and outcomes in COVID-19-related ARDS. Microbiol. Spectr. 12:e0357423. doi: 10.1128/spectrum.03574-23
Derrien, M., Belzer, C., and de Vos, W. M. (2017). Akkermansia muciniphila and its role in regulating host functions. Microb. Pathog. 106, 171–181. doi: 10.1016/j.micpath.2016.02.005
Di Bonaventura, G., Pompilio, A., Zappacosta, R., Petrucci, F., Fiscarelli, E., Rossi, C., et al. (2010). Role of excessive inflammatory response to Stenotrophomonas maltophilia lung infection in DBA/2 mice and implications for cystic fibrosis. Infect. Immun. 78, 2466–2476. doi: 10.1128/IAI.01391-09
Dickson, R. P., Erb-Downward, J. R., Freeman, C. M., McCloskey, L., Falkowski, N. R., Huffnagle, G. B., et al. (2017). Bacterial topography of the healthy human lower respiratory tract. mBio 8:e02287-16. doi: 10.1128/mBio.02287-16
Dickson, R. P., Erb-Downward, J. R., Martinez, F. J., and Huffnagle, G. B. (2016a). The microbiome and the respiratory tract. Annu. Rev. Physiol. 78, 481–504. doi: 10.1146/annurev-physiol-021115-105238
Dickson, R. P., Schultz, M. J., van der Poll, T., Schouten, L. R., Falkowski, N. R., Luth, J. E., et al. (2020). Lung microbiota predict clinical outcomes in critically Ill patients. Am. J. Respir. Crit. Care Med. 201, 555–563. doi: 10.1164/rccm.201907-1487OC
Dickson, R. P., Singer, B. H., Newstead, M. W., Falkowski, N. R., Erb-Downward, J. R., Standiford, T. J., et al. (2016b). Enrichment of the lung microbiome with gut bacteria in sepsis and the acute respiratory distress syndrome. Nat. Microbiol. 1:16113. doi: 10.1038/nmicrobiol.2016.113
Finn, D. P., Haroutounian, S., Hohmann, A. G., Krane, E., Soliman, N., and Rice, A. S. C. (2021). Cannabinoids, the endocannabinoid system, and pain: A review of preclinical studies. Pain 162(Suppl. 1), S5–S25. doi: 10.1097/j.pain.0000000000002268
Gaibani, P., Viciani, E., Bartoletti, M., Lewis, R. E., Tonetti, T., Lombardo, D., et al. (2021). The lower respiratory tract microbiome of critically ill patients with COVID-19. Sci. Rep. 11:10103. doi: 10.1038/s41598-021-89516-6
Ganesh, B. P., Klopfleisch, R., Loh, G., and Blaut, M. (2013). Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PLoS One 8:e74963. doi: 10.1371/journal.pone.0074963
Graziani, F., Pujol, A., Nicoletti, C., Dou, S., Maresca, M., Giardina, T., et al. (2016). Ruminococcus gnavus E1 modulates mucin expression and intestinal glycosylation. J. Appl. Microbiol. 120, 1403–1417. doi: 10.1111/jam.13095
Horn, K. J., Schopper, M. A., Drigot, Z. G., and Clark, S. E. (2022). Airway Prevotella promote TLR2-dependent neutrophil activation and rapid clearance of Streptococcus pneumoniae from the lung. Nat. Commun. 13:3321. doi: 10.1038/s41467-022-31074-0
Hu, Y., Shen, J., An, Y., Jiang, Y., and Zhao, H. (2024). Phenotypes and lung microbiota signatures of immunocompromised patients with pneumonia-related acute respiratory distress syndrome. J. Inflamm. Res. 17, 1429–1441. doi: 10.2147/JIR.S453123
Hussell, T., and Bell, T. J. (2014). Alveolar macrophages: Plasticity in a tissue-specific context. Nat. Rev. Immunol. 14, 81–93. doi: 10.1038/nri3600
Imbert, S., Revers, M., Enaud, R., Orieux, A., Camino, A., Massri, A., et al. (2024). Lower airway microbiota compositions differ between influenza, COVID-19 and bacteria-related acute respiratory distress syndromes. Crit. Care 28:133. doi: 10.1186/s13054-024-04922-2
Jackson, A. R., Hegde, V. L., Nagarkatti, P. S., and Nagarkatti, M. (2014). Characterization of endocannabinoid-mediated induction of myeloid-derived suppressor cells involving mast cells and MCP-1. J. Leukoc. Biol. 95, 609–619. doi: 10.1189/jlb.0613350
Jin, A., Zhao, Y., Yuan, Y., Ma, S., Chen, J., Yang, X., et al. (2021). Single treatment of vitamin D3 ameliorates LPS-induced acute lung injury through changing lung Rodentibacter abundance. Mol. Nutr. Food Res. 66:e2100952. doi: 10.1002/mnfr.202100952
Johansson, M. E., Ambort, D., Pelaseyed, T., Schütte, A., Gustafsson, J. K., Ermund, A., et al. (2011). Composition and functional role of the mucus layers in the intestine. Cell Mol. Life Sci. 68, 3635–3641. doi: 10.1007/s00018-011-0822-3
Khatiwada, S., and Subedi, A. (2020). Lung microbiome and coronavirus disease 2019 (COVID-19): Possible link and implications. Hum. Microb. J. 17:100073. doi: 10.1016/j.humic.2020.100073
Kullberg, R. F. J., de Brabander, J., Boers, L. S., Biemond, J. J., Nossent, E. J., Heunks, L. M. A., et al. (2022). Lung microbiota of critically Ill patients with COVID-19 are associated with nonresolving acute respiratory distress syndrome. Am. J. Respir. Crit. Care Med. 206, 846–856. doi: 10.1164/rccm.202202-0274OC
Kyo, M., Nishioka, K., Nakaya, T., Kida, Y., Tanabe, Y., Ohshimo, S., et al. (2019). Unique patterns of lower respiratory tract microbiota are associated with inflammation and hospital mortality in acute respiratory distress syndrome. Respir. Res. 20:246. doi: 10.1186/s12931-019-1203-y
Laurenzi, G. A., Potter, R. T., and Kass, E. H. (1961). Bacteriologic flora of the lower respiratory tract. N. Engl. J. Med. 265, 1273–1278. doi: 10.1056/NEJM196112282652601
Leitao Filho, F. S., Alotaibi, N. M., Ngan, D., Tam, S., Yang, J., Hollander, Z., et al. (2019). Sputum microbiome is associated with 1-year mortality after chronic obstructive pulmonary disease hospitalizations. Am. J. Respir. Crit. Care Med. 199, 1205–1213. doi: 10.1164/rccm.201806-1135OC
Li, F., Han, Y., Cai, X., Gu, M., Sun, J., Qi, C., et al. (2020). Dietary resveratrol attenuated colitis and modulated gut microbiota in dextran sulfate sodium-treated mice. Food Funct. 11, 1063–1073. doi: 10.1039/c9fo01519a
Li, L., Fang, Z., Liu, X., Hu, W., Lu, W., Lee, Y. K., et al. (2020). Lactobacillus reuteri attenuated allergic inflammation induced by HDM in the mouse and modulated gut microbes. PLoS One 15:e0231865. doi: 10.1371/journal.pone.0231865
Li, N. N., Kang, K., Zhou, Y., Liu, Y. Q., Zhang, Q. Q., Luo, P. Y., et al. (2024). Throat microbiota drives alterations in pulmonary alveolar microbiota in patients with septic ARDS. Virulence 15:2350775. doi: 10.1080/21505594.2024.2350775
Lira-Lucio, J. A., Falfán-Valencia, R., Ramírez-Venegas, A., Buendía-Roldán, I., Rojas-Serrano, J., Mejía, M., et al. (2020). Lung microbiome participation in local immune response regulation in respiratory diseases. Microorganisms 8:1059. doi: 10.3390/microorganisms8071059
Liu, H., Hou, C., Wang, G., Jia, H., Yu, H., and Zeng, X. (2017). Lactobacillus reuteri I5007 modulates intestinal host defense peptide expression in the model of IPEC-J2 cells and neonatal piglets. Nutrients 9:559. doi: 10.3390/nu9060559
Lv, L., Cui, E. H., Wang, B., Li, L. Q., Hua, F., Lu, H. D., et al. (2023). Multiomics reveal human umbilical cord mesenchymal stem cells improving acute lung injury via the lung-gut axis. World J. Stem Cells 15, 908–930. doi: 10.4252/wjsc.v15.i9.908
Man, W. H., de Steenhuijsen Piters, W. A., and Bogaert, D. (2017). The microbiota of the respiratory tract: Gatekeeper to respiratory health. Nat. Rev. Microbiol. 15, 259–270. doi: 10.1038/nrmicro.2017.14
Merenstein, C., Liang, G., Whiteside, S. A., Cobián-Güemes, A. G., Merlino, M. S., Taylor, L. J., et al. (2021). Signatures of COVID-19 severity and immune response in the respiratory tract microbiome. mBio 12:e0177721. doi: 10.1128/mBio.01777-21
Minichino, A., Jackson, M. A., Francesconi, M., Steves, C. J., Menni, C., Burnet, P. W. J., et al. (2021). Endocannabinoid system mediates the association between gut-microbial diversity and anhedonia/amotivation in a general population cohort. Mol. Psychiatry 26, 6269–6276. doi: 10.1038/s41380-021-01147-5
Mohammed, A., Alghetaa, H. K., Zhou, J., Chatterjee, S., Nagarkatti, P., and Nagarkatti, M. (2020). Protective effects of Δ9 -tetrahydrocannabinol against enterotoxin-induced acute respiratory distress syndrome are mediated by modulation of microbiota. Br. J. Pharmacol. 177, 5078–5095. doi: 10.1111/bph.15226
Molyneaux, P. L., Cox, M. J., Willis-Owen, S. A., Mallia, P., Russell, K. E., Russell, A. M., et al. (2014). The role of bacteria in the pathogenesis and progression of idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 190, 906–913. doi: 10.1164/rccm.201403-0541OC
Montassier, E., Kitsios, G. D., Radder, J. E., Le Bastard, Q., Kelly, B. J., Panzer, A., et al. (2023). Robust airway microbiome signatures in acute respiratory failure and hospital-acquired pneumonia. Nat. Med. 29, 2793–2804. doi: 10.1038/s41591-023-02617-9
Mu, Q., Tavella, V. J., and Luo, X. M. (2018). Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 9:757. doi: 10.3389/fmicb.2018.00757
Mukherjee, D., Chora, ÂF., Lone, J. C., Ramiro, R. S., Blankenhaus, B., Serre, K., et al. (2022). Host lung microbiota promotes malaria-associated acute respiratory distress syndrome. Nat. Commun. 13:3747. doi: 10.1038/s41467-022-31301-8
Nagarkatti, P., Pandey, R., Rieder, S. A., Hegde, V. L., and Nagarkatti, M. (2009). Cannabinoids as novel anti-inflammatory drugs. Future Med. Chem. 1, 1333–1349. doi: 10.4155/fmc.09.93
Naik, C., Kulkarni, H., Darabi, A., and Bhanot, N. (2013). Ochrobactrum anthropi: A rare cause of pneumonia. J. Infect. Chemother. 19, 162–165. doi: 10.1007/s10156-012-0436-1
O’Dwyer, D. N., Ashley, S. L., Gurczynski, S. J., Xia, M., Wilke, C., Falkowski, N. R., et al. (2019). Lung microbiota contribute to pulmonary inflammation and disease progression in pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 199, 1127–1138. doi: 10.1164/rccm.201809-1650OC
Osafo, N., Yeboah, O. K., and Antwi, A. O. (2021). Endocannabinoid system and its modulation of brain, gut, joint and skin inflammation. Mol. Biol. Rep. 48, 3665–3680. doi: 10.1007/s11033-021-06366-1
Pandey, R., Mousawy, K., Nagarkatti, M., and Nagarkatti, P. (2009). Endocannabinoids and immune regulation. Pharmacol. Res. 60, 85–92. doi: 10.1016/j.phrs.2009.03.019
Panzer, A. R., Lynch, S. V., Langelier, C., Christie, J. D., McCauley, K., Nelson, M., et al. (2018). Lung microbiota is related to smoking status and to development of acute respiratory distress syndrome in critically Ill trauma patients. Am. J. Respir. Crit. Care Med. 197, 621–631. doi: 10.1164/rccm.201702-0441OC
Pecora, D. V. (1963). A comparison of transtracheal aspiration with other methods of determining the bacterial flora of the lower respiratory tract. N. Engl. J. Med. 269, 664–666. doi: 10.1056/NEJM196309262691304
Poroyko, V., Meng, F., Meliton, A., Afonyushkin, T., Ulanov, A., Semenyuk, E., et al. (2015). Alterations of lung microbiota in a mouse model of LPS-induced lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 309, L76–L83. doi: 10.1152/ajplung.00061.2014
Rieder, S. A., Nagarkatti, P., and Nagarkatti, M. (2012). Multiple anti-inflammatory pathways triggered by resveratrol lead to amelioration of staphylococcal enterotoxin B-induced lung injury. Br. J. Pharmacol. 167, 1244–1258. doi: 10.1111/j.1476-5381.2012.02063.x
Rogers, G. B., Zain, N. M., Bruce, K. D., Burr, L. D., Chen, A. C., Rivett, D. W., et al. (2014). A novel microbiota stratification system predicts future exacerbations in bronchiectasis. Ann. Am. Thorac. Soc. 11, 496–503. doi: 10.1513/AnnalsATS.201310-335OC
Roy, M. G., Livraghi-Butrico, A., Fletcher, A. A., McElwee, M. M., Evans, S. E., Boerner, R. M., et al. (2014). Muc5b is required for airway defence. Nature 505, 412–416. doi: 10.1038/nature12807
Ruff, W. E., Greiling, T. M., and Kriegel, M. A. (2020). Host-microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 18, 521–538. doi: 10.1038/s41579-020-0367-2
Salehi, B., Mishra, A. P., Nigam, M., Sener, B., Kilic, M., Sharifi-Rad, M., et al. (2018). Resveratrol: A double-edged sword in health benefits. Biomedicines 6:91. doi: 10.3390/biomedicines6030091
Sapra, L., Saini, C., Das, S., Mishra, P. K., Singh, A., Mridha, A. R., et al. (2024). Lactobacillus rhamnosus (LR) ameliorates pulmonary and extrapulmonary acute respiratory distress syndrome (ARDS) via targeting neutrophils. Clin. Immunol. 258:109872. doi: 10.1016/j.clim.2023.109872
Schmitt, F. C. F., Lipinski, A., Hofer, S., Uhle, F., Nusshag, C., Hackert, T., et al. (2020). Pulmonary microbiome patterns correlate with the course of the disease in patients with sepsis-induced ARDS following major abdominal surgery. J. Hosp. Infect. doi: 10.1016/j.jhin.2020.04.028 [Epub ahead of print].
Segal, L. N., Clemente, J. C., Tsay, J. C., Koralov, S. B., Keller, B. C., Wu, B. G., et al. (2016). Enrichment of the lung microbiome with oral taxa is associated with lung inflammation of a Th17 phenotype. Nat. Microbiol. 1:16031. doi: 10.1038/nmicrobiol.2016.31
Shen, J., Hu, Y., Lv, J., Zhao, H., Wang, B., Yang, S., et al. (2022). Lung Microbiota Signature and Corticosteroid Responses in Pneumonia-Associated Acute Respiratory Distress Syndrome in Hematological Patients. J. Inflamm. Res. 15, 1317–1329. doi: 10.2147/JIR.S353662
Shen, Z., Xiao, Y., Kang, L., Ma, W., Shi, L., Zhang, L., et al. (2020). Genomic diversity of severe acute respiratory Syndrome-Coronavirus 2 in patients with Coronavirus Disease 2019. Clin. Infect. Dis. 71, 713–720. doi: 10.1093/cid/ciaa203
Sido, J. M., Nagarkatti, P. S., and Nagarkatti, M. (2016). Production of endocannabinoids by activated T cells and B cells modulates inflammation associated with delayed-type hypersensitivity. Eur. J. Immunol. 46, 1472–1479. doi: 10.1002/eji.201546181
Sommariva, M., Le Noci, V., Bianchi, F., Camelliti, S., Balsari, A., Tagliabue, E., et al. (2020). The lung microbiota: Role in maintaining pulmonary immune homeostasis and its implications in cancer development and therapy. Cell Mol. Life Sci. 77, 2739–2749. doi: 10.1007/s00018-020-03452-8
Soroosh, P., Doherty, T. A., Duan, W., Mehta, A. K., Choi, H., Adams, Y. F., et al. (2013). Lung-resident tissue macrophages generate Foxp3+ regulatory T cells and promote airway tolerance. J. Exp. Med. 210, 775–788. doi: 10.1084/jem.20121849
Sulaiman, I., Chung, M., Angel, L., Tsay, J. J., Wu, B. G., Yeung, S. T., et al. (2021). Microbial signatures in the lower airways of mechanically ventilated COVID-19 patients associated with poor clinical outcome. Nat. Microbiol. 6, 1245–1258. doi: 10.1038/s41564-021-00961-5
Sultan, M., Wilson, K., Abdulla, O. A., Busbee, P. B., Hall, A., Carter, T., et al. (2021). Endocannabinoid anandamide attenuates acute respiratory distress syndrome through modulation of microbiome in the gut-lung axis. Cells 10:3305. doi: 10.3390/cells10123305
Tian, Z., Wu, E., You, J., Ma, G., Jiang, S., Liu, Y., et al. (2022). Dynamic alterations in the lung microbiota in a rat model of lipopolysaccharide-induced acute lung injury. Sci. Rep. 12:4791. doi: 10.1038/s41598-022-08831-8
Tu, C., Wang, Z., Xiang, E., Zhang, Q., Zhang, Y., Wu, P., et al. (2022). Human umbilical cord mesenchymal stem cells promote macrophage PD-L1 expression and attenuate acute lung injury in mice. Curr. Stem Cell Res. Ther. 17, 564–575. doi: 10.2174/1574888X17666220127110332
Walsh, D. M., McCullough, S. D., Yourstone, S., Jones, S. W., Cairns, B. A., Jones, C. D., et al. (2017). Alterations in airway microbiota in patients with PaO2/FiO2 ratio = 300 after burn and inhalation injury. PLoS One 12:e0173848. doi: 10.1371/journal.pone.0173848
Wang, R., Huang, C., Yang, W., Wang, C., Wang, P., Guo, L., et al. (2023). Respiratory microbiota and radiomics features in the stable COPD patients. Respir. Res. 24:131. doi: 10.1186/s12931-023-02434-1
Yang, D., Xing, Y., Song, X., and Qian, Y. (2020). The impact of lung microbiota dysbiosis on inflammation. Immunology 159, 156–166. doi: 10.1111/imm.13139
Yi, X., Gao, J., and Wang, Z. (2022). The human lung microbiome-A hidden link between microbes and human health and diseases. Imeta 1:e33. doi: 10.1002/imt2.33
Zacharias, M., Kashofer, K., Wurm, P., Regitnig, P., Schütte, M., Neger, M., et al. (2022). Host and microbiome features of secondary infections in lethal covid-19. iScience 25:104926. doi: 10.1016/j.isci.2022.104926
Keywords: lung microbiota, ARDS, corona virus disease 2019 (COVID-19), prognosis, metabolic products
Citation: Tang Y, Liu B, Ma A, Wang B, Xiong H, Zhou Y, Yang J and Kang Y (2025) Lung microbiota: a new hope for treating acute respiratory distress syndrome? Front. Microbiol. 16:1586949. doi: 10.3389/fmicb.2025.1586949
Received: 04 March 2025; Accepted: 08 May 2025;
Published: 30 May 2025.
Edited by:
George Grant, Independent Researcher, Aberdeen, United KingdomReviewed by:
Sandhya Bansal, St. Joseph’s Hospital and Medical Center, United StatesQiyue Jia, Capital Medical University, China
Copyright © 2025 Tang, Liu, Ma, Wang, Xiong, Zhou, Yang and Kang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jing Yang, eWppbmdzY3VAMTYzLmNvbQ==; Yan Kang, S2FuZ3lhbkBzY3UuZWR1LmNu
†These authors have contributed equally to this work and share first authorship