 Yann Moalic
Yann Moalic Toan Bao Hung Nguyen
Toan Bao Hung Nguyen Jordan Hartunians
Jordan Hartunians Tiphaine Birien
Tiphaine Birien Mohamed Jebbar
Mohamed Jebbar- Univ Brest, Ifremer, CNRS, EMR 6002 BIOMEX, BEEP, Plouzané, France
In Thermococcales, energy metabolism genes are regulated by the sulfur-responsive transcriptional regulator SurR. In the piezophilic archaeon Thermococcus barophilus, these genes are also influenced by hydrostatic pressure. To explore the interaction between pressure, sulfur availability, and gene regulation, we constructed and analyzed several deletion mutants, including a partial surR knockout, under varying conditions. Our results show that hydrostatic pressure modulates the expression of energy metabolism genes and that SurR is essential for activating the hydrogenogenic gene cluster, even in sulfur-rich environments. Under sulfur limitation, the membrane-bound sulfur-reducing complex (MBS) is required for normal growth. These regulatory patterns expand current models derived from non-piezophilic species such as Pyrococcus furiosus and Thermococcus kodakarensis. Overall, our findings indicate that hydrostatic pressure shapes SurR function in T. barophilus, reflecting its adaptive plasticity in extreme environments.
1 Introduction
Thermococcales are ubiquitous extremophilic Archaea found at hydrothermal environments growing optimally at temperature over 80°C (Schut et al., 2014). The chronic energy stress induced by high temperature requires specific metabolic strategies allowing them to thrive in these environments (Valentine, 2007). Additionally, Thermococcales can oxidize different types of carbon-based molecules (proteins, organic acids, carbohydrates…) to produce energy (Nakagawa and Takai, 2006).
A diverse set of hydrogenases, oxidoreductases, and electron transporters enable Thermococcales to generate the electron fluxes for maintaining the ionic gradient across the membrane, which is directly linked to ATP production. Consequently, energy conservation is associated with both H2S production and H2 turnover and recycling (Wu et al., 2018). While some aspects of the functional mechanisms of these catabolic components remain unclear, they are relatively well characterized. The membrane-bound hydrogenase (MBH) and its homologous S0-reducing reductase (MBS) are conserved across all publicly available Thermococcales genomes (Schut et al., 2013). These enzymes oxidize the ferredoxin yielding either H2S or H2, while contributing to energy conservation via Na + pumping. In P. furiosus, one of the most study model of Thermococcales with T. kodakarensis, the addition of elemental sulfur (S0) to growth medium, induces the expression of MBS-encoding genes, while repressing MBH-encoding genes (Chou et al., 2007). This shift results in a two-fold increase in microbial cell yield (Schut et al., 2007), suggesting that MBS is more efficient in energy conservation. MBS facilitates electron transfer for the reduction of polysulfide chains while simultaneous driving Na+ translocation across the membrane. Additionally, it should be noted that in the presence of S0 H2S can be produced abiotically (Wu et al., 2018; Burkhart et al., 2019; Yu et al., 2020).
The master redox-active transcription regulator, SurR, modulates the expression of various enzymes of the energy conservation system based upon whether elemental sulfur is available or not (Lipscomb et al., 2009). When sulfur (S0) is present, SurR becomes oxidized and no longer binds specifically to its target DNA. As a result, the H2-related genes are deactivated, while the H2S-related genes are de-repressed, prompting P. furiosus to produce hydrogen sulfide (Yang et al., 2010). In absence of sulfur, the reduced form of SurR represses genes implicated in H2S production while activating those linked in H2 production, leading P. furiosus to produce hydrogen.
The only available SurR structure was solved for the P. furiosus variant (Yang et al., 2010). SurR is a homodimer with double symmetry, where each monomer consists of three distinct domains. The N-terminal (residues 1–75) and C-terminal (residues 166–219) regions each form a winged helix-turn-helix (wHTH) DNA-binding fold related to that of the ArsR family (Yang et al., 2010).
The SurR recognition motifs (GTTn3ATC or GTTn3AACn5GTT) are located in many promoters of genes involved in the sulfur response of Thermococcales (Lipscomb et al., 2009; Hidese et al., 2017; Lim et al., 2017; Lipscomb et al., 2017; Moalic et al., 2021). A redox-active switch controls DNA binding, through a CxxC motif in the N-terminal wHTH domain, likely by affecting the conformation of the protein’s DNA binding domain. In the reduced free thiol form, in the absence of S0, SurR activates the expression of its own gene as well as genes involved in hydrogen metabolism. SurR also represses the expression of Nsr (NADPH-dependent sulfur reductase), Mbs and other clusters involved in sulfur reduction, such as Pdo (Protein disulfide oxidoreductase) (Lipscomb et al., 2017). However, when S0 is supplied, the CxxC motif forms a disulfide bond, abrogating specific DNA binding by SurR, resulting in deactivation of the hydrogenase genes and de-repression of sulfur reducing genes. Indeed, as described in T. kodakarensis, surR deletion inhibits growth in the absence of sulfur but has no effect in the presence of sulfur (Santangelo et al., 2011). Activation or repression by SurR is linked to its binding site position in the promoter. At target sites located upstream of the TATA box, SurR acts as an activator, whereas it represses transcription when it binds downstream of the TATA box (Yang et al., 2010). In some cases, the presence of both upstream and downstream binding sites is essential for repression (Hidese et al., 2017).
An in vitro study on Thermococcus onnurineus, a member of the Thermococcales, demonstrated that Pdo (protein disulfide reductase) facilitates electron transfer between thioredoxin reductase (TrxR) and the redox-sensitive transcription factor SurR (Lim et al., 2017). These findings indicate that the TrxR-Pdo pair functions as a redox system that reduces SurR. While this reduction has only been observed in vitro, the results are significant, as they suggest that SurR-mediated regulation of the S₀ response could be reversed even in the absence of elemental sulfur (S₀) (Lim et al., 2017). Notably, the genomic organization of surR is highly conserved across all known Thermococcales genomes. Specifically, surR and pdo are positioned adjacent to each other in a divergent arrangement, implying that their functions are tightly coordinated in vivo. Furthermore, pdo has been identified as part of the SurR regulon (Lipscomb et al., 2009). Microarray expression profiling revealed that pdo is upregulated during the initial response to S₀ (Schut et al., 2007) and repressed by SurR, which binds to the pdo promoter in the absence of S₀ (Lipscomb et al., 2009). It has been proposed that once oxidized sulfur species are depleted within the cell, SurR would revert to its reduced, active state. However, the precise mechanism underlying this transition remains undemonstrated (Yang et al., 2010).
Thermococcus barophilus MP, the proposed biological model, is the first true piezophilic hyperthermophile to be isolated from a deep-sea hydrothermal vent (Marteinsson et al., 1999). This archaeon thrives across a wide hydrostatic pressure range of 0.1–80 MPa (Popt 40 MPa) and a temperature range of 48–90°C (Topt 85°C). Elemental sulfur enhances its growth. The complete genome of T. barophilus MP has been sequenced and annotated (Vannier et al., 2011) and a genetic system has been developed to further investigate its adaptations to HHP (Thiel et al., 2014; Birien et al., 2018). Neutron scattering studies have revealed that the proteome of T. barophilus MP exhibits greater flexibility and sensitivity to HHP, a smaller hydration shell, and more confined intracellular water compared to that of a related piezosensitive species T. kodakarensis (Martinez et al., 2016). Interestingly, increased proteome flexibility appears to impair biological function at low pressure, but this effect is mitigated by the accumulation of the osmolyte mannosylglycerate, which stabilizes structures under low pressure conditions (Cario et al., 2016).
Transcriptomic analysis of T. barophilus MP has revealed that hydrostatic pressure influences the expression of numerous genes, particularly those involved in hydrogen and elemental sulfur metabolism (Vannier et al., 2015). Notably, the expression of key hydrogenase-encoding genes remains minimal at 40 MPa in the presence of S0. However, in a surprising deviation from this pattern, when pressure is reduced to 0.1 or increased to 70 MPa, the expression of the hydrogen-related genes (e.g., mbh, mbh-codh, shI, and shII) is upregulated by 2 to 40-fold, even in presence of S0 (Vannier et al., 2015). In contrast, hydrostatic pressure has little to no effect on the expression of genes within the sulfur regulon, including mbs, nsr, surR, pdo, nfn and xfn in T. barophilus MP (Vannier et al., 2015). This suggests that gene regulation in T. barophilus differs significantly from that of related non-piezophilic species such as P. furiosus or T. kodakarensis, for which HHP does not modify the expression of genes under SurR regulation (unpublished results in GSE72783 for P. furiosus and for T. kodakarensis see Vannier et al., 2015) The data further imply that high hydrostatic pressure (HHP) may modulate the DNA-binding affinity of SurR in T. barophilus MP, contributing to its unique regulatory adaptations to extreme pressure conditions.
In this study, we examined the impact of high hydrostatic pressure (HHP) and the redox regulator SurR on physiological growth and the expression of gene clusters involved in energy conservation. Using a combination of genetic approaches and RT-qPCR, we analyzed growth kinetics and gene expression in various mutants.
Our findings confirm that HHP plays a modulatory role in gene regulation and reveal the existence of a sulfur-independent mechanism influencing SurR activity—an adaptation not observed in the other studied non-piezophilic Thermococcales. These results underscore a specialized genetic regulatory strategy that enables T. barophilus to thrive in deep-sea hydrothermal environments.
2 Materials and methods
2.1 Strains and growth conditions
All the strains used in this study are list in Supplementary Table S1. A modified version of the Thermococcales Rich Medium (TRM) was used for all growth studies, based on the formulation described by Zeng et al. (2009). This modified medium (TRMm) contains 5 g·L−1 yeast extract and 5 g·L−1 tryptone, an increase from the original TRM composition, to support growth under hydrogenogenic conditions with 5 g·L−1 pyruvate as the carbon source. For sulfidogenic growth, TRMm was supplemented with 0.25 g·L−1 colloidal sulfur, except for induction experiments, where higher sulfur concentrations (0.5 g·L−1 or 2 g·L−1) were used. Growth experiments were achieved in biological triplicates at 85°C in anaerobic conditions. At atmospheric pressure condition (0.1 MPa), 50 mL vials were filled with 20 mL of medium while cultivation under high hydrostatic pressure (40 MPa and 70 MPa) requires the use of 15 mL vials entirely filled with medium to avoid the imploding risk due to gas phase. The absence of gas phase has no effect on growth at atmospheric pressure (0.1 MPa) comparatively to cultivation realized with a gas phase (data not shown). Each growth kinetic assay was initiated from overnight pre-cultures (16 h) prepared in the same medium, with an initial cell density of 2 × 106 cells/mL.
For HHP growth experiments, four incubators (Top Industrie) were used (Zeng et al., 2009). Due to compression/decompression constrains, each kinetic point corresponds to a separate incubator containing six 15 mL vials, three replicate of the mutant strain and three replicates of the ∆517 control strain (either ∆517 or ∆517p according to the sulfur condition).
Growth monitoring was realized by cell counting using a Thoma chamber and photonic microscopy at a magnification of X40.
2.2 Mutants constructions
The genetic manipulations were realized following the pop-in/pop-out protocol developed in our laboratory (Birien et al., 2018). This genetic tool was utilized under sulfidogenic conditions, and all the mutant constructions were generated from the T. barophilus strain UBOCC-M3300, a derivative of the T. barophilus MP type strain (Marteinsson et al., 1999), which contains a deletion of the TERMP_00517 gene encoding xanthine-guanine phosphoribosyltransferase, For the purposes of this study strain UBOCC-M3300 is designated as the wild type or reference or ∆517 strain (Supplementary Table S1). The deletion mutations generated in this study included those impacting the Mrp-Mbh and Mrp-Mbs membrane clusters (∆mbh, ∆mbs) and the two cytosolic hydrogenases SHI and SHII (∆shI and ∆shII). In addition, a partial deletion of the SurR coding sequence was produced (∆surR). More precisely, the double copy of Mrp-Mbh complex was deleted (∆mbh strain) and for the ∆surR strain, a partial deletion was built after all attempts to obtain a completed deletion of its locus have failed. Thus, a 137 bp segment at the 5′ end of the surR gene was deleted, corresponding the N-terminal wHTH domain of SurR bearing the CxxC motif located at positions 24 and 27. Thus, clones with this deletion were identified through PCR screening and subsequently verified by Sanger sequencing.
All the strains of this study have followed the same protocol of “pyruvate growth adaptation” before testing the effect of sulfur on the phenotypes. The experiments of adaptation were repeated at least 3 times independently with the ∆517 strain as control. Thus, we consider that if the strain cannot growth, it is more probably due to the absence of the gene/gene clusters in the strain than the toxicity of H2-end product, especially when the H2 production pathway is impaired. The growth curves realized in pyruvate condition were carried out with strains that have been 96 h into TRMm media before pre-culture preparation.
2.3 Gene expression
To assess the effect of SurR regulator on the expression of the cluster of hydrogenases (mbh1, mbh2, shI, shII) and sulfane oxidoreductase (mbs), RT-qPCR were realized. First, RNA of cells in mid-Log phase were extracted with Trizol reagent. 50 mL of culture were centrifuged at 8,000 rpm (6 min at 4°C) and the pellets were suspended in 1 mL of Trizol and transferred in RNase-free 2 mL tubes (Biopur Eppendorf). Then the procedural guidelines of the manufacturer were followed until the RNA is suspended in RNase free water (54 μL). Then a DNase treatment was realized (RQ1 RNase-free DNase kit, Promega) and the RNA were quantified with a nanodrop 8000 (Thermofischer) and conserved at −80°C.
The iScript™ Reverse Transcription Supermix for RT-qPCR (Bio-Rad) was used to get cDNA at the concentration of 2.5 ng.μL−1 About 12.5 ng of cDNA were used as matrix for quantitative real-time PCR reactions (SsoAdvanced™ Universal SYBR® Green supermix - Bio-Rad). The primers were used at the final concentration of 0.5 μM each in a final volume reaction of 20 μL (listed in Supplementary Table S3). The reactions were launched on a CFX96TM (Bio-Rad) with an amplification step of 40 cycles (95°, 15 s followed by 60°, 30 s). The data were saved and analyzed with the Bio-Rad CFX Maestro software.
The ∆∆Ct method was then applied to analyze gene expression variation (Schmittgen and Livak, 2008). Due to the diverse growth culture conditions (Sulfur and Pressure), three references genes were tested: two 30S proteins coding (S19 and S13) and the pcna (see Supplementary Table S3 for their Locus).
2.4 Genome sequencing and analysis
DNA was extracted at mid-log growth phase using protocols previously described (Thiel et al., 2014; Birien et al., 2018). Library preparation and Illumina sequencing were performed at Novogene, UK. Bowtie2 was used for read mapping (Langmead and Salzberg, 2012) and Samtools (Li et al., 2009) was used to manipulate and produce the binary data required for the variants detection. Lofreq (Wilm et al., 2012) is the variant-caller used for inferring single nucleotides variants and indels in the sequencing data comparing to the reference genome of T. barophilus MP (Refseq NC_014804.1). Finally, SnpEff helps to predict the effects of genetic variants (Cingolani et al., 2012).
3 Results and discussion
3.1 Growth of Thermococcus barophilus on pyruvate in batch culture
Since its isolation during the French-American “MAR93” cruise (~30 years ago) (Marteinsson et al., 1999), T. barophilus has been routinely cultivated in yeast extract and peptone-based media (YPS and TRM) with added Sulfur for optimal growth (Marteinsson et al., 1995; Zeng et al., 2009). Growth on minimal media (TAA, Thermococcales amino acid or TBM, Thermococcales basic medium) yields lower but interpretable results with sulfur (Thiel et al., 2014; Cario et al., 2015). In rich media without sulfur, growth was minimal or absent due to H₂ toxicity, which inhibits reduced ferredoxin regeneration (Le Guellec et al., 2021; Malik et al., 1989; Schäfer and Schönheit, 1991). However, continuous culture experiments realized in a gas-lift bioreactor supports high growth rates comparable to sulfur-supplemented batch cultures (Postec et al., 2005). This study required sulfur-free media that did not hinder cell growth for the physiological characterization of energy-conserving gene clusters. After multiple attempts, a modified TRM medium supplemented with pyruvate (5 g/L) successfully supported the growth of T. barophilus at satisfactory yields without sulfur. An adaptation period of at least 72 h was required for cell growth to reach 6.0E+07 cells.mL−1 from an initial density of 1.21E+07 cells.mL−1. However, once adapted, subcultures grew equally well with or without sulfur (Supplementary Figure S1). This pyruvate-adapted strain (∆517p) originated from the laboratory’s genetic strain, Tba ∆517 (Birien et al., 2018). The growth rate of ∆517p on pyruvate (0.36 h−1) was comparable to that of ∆517 with sulfur and increased slightly to 0.43 h−1 when sulfur replaced pyruvate (red curve). In contrast, non-adapted ∆517 exhibited a severe growth impairment (0.004 h−1, blue curve). To assess whether genetic mutations c contributed to metabolic adaptation, a genomic comparison was conducted between Tba ∆517p, Tba ∆517 and the reference genome of Tba MP (NC_014804.1). Notably, Tba ∆517 has lost its plasmid pTBMP (NC_15471.1) for unknown reasons, and it is likewise absent in Tba ∆517p. Both strains also lack the gene encoding a phosphoribosyltransferase (TERMP_00517, 648 bp), resulting in a genome assembly of identical size (2,009,585 pb). To identify genetic variants, LoFreq was used to analyze sequence data (Wilm et al., 2012) and categorized potential effects with SNPeff (Cingolani et al., 2012). After filtering for allele Frequency (AF > 0.1), Tba ∆517 exhibited 47 variants compared to Tba MP while Tba ∆517p had 51. Of these, 45 variants in Tba ∆517 were also present in Tba ∆517p, leaving six unique to Tba ∆517p. SnpEff analysis identified three variants with a predicted HIGH impact due to frameshift mutations (Table 1). Two of these occur at the same locus (TERMP_02062), which encodes an adenylyl-sulfate kinase (F0LLK7), an enzyme involved in active sulfate synthesis. The third is located at TERMP_00520, annotated as an uncharacterized protein (F0LJW0). Additionally, two mutations were classified as MODERATE impact missense variants. One occurred at locus TERMP_00866 coding for an uncharacterized protein (F0LLR6), causing a Glycine to Arginine substitution at position 159. The other affects TERMP_01561, encoding a low affinity inorganic phosphate transporter (F0LIT4), resulting in a Threonine to Methionine substitution at position 95. Finally, a variant was detected in the promoter region of TERMP_02063, which encodes a transposase. Classified as a MODIFIER impact, this variant likely has uncertain or minimal direct effects, as it does not alter protein sequence. These mutations do not provide clear evidence of direct selection for growth on pyruvate. Instead, the adaptation likely results from metabolic reconfiguration via regulatory mechanisms, similar to shifts observed when sulfur is introduced into pyruvate-grown cells or in comparative proteomic studies of sulfur vs. pyruvate grown Thermococcales (Schut et al., 2007; Jager et al., 2014; Moon et al., 2015). Future studies are needed to further elucidate these mechanisms. However, this genomic analysis clarified the effects of a 96-h adaptation period required for growth without sulfur, revealing a metabolic shift rather than a genetic drift. Thus, we established TRMm + pyruvate as a sulfur-free growth condition for this study.
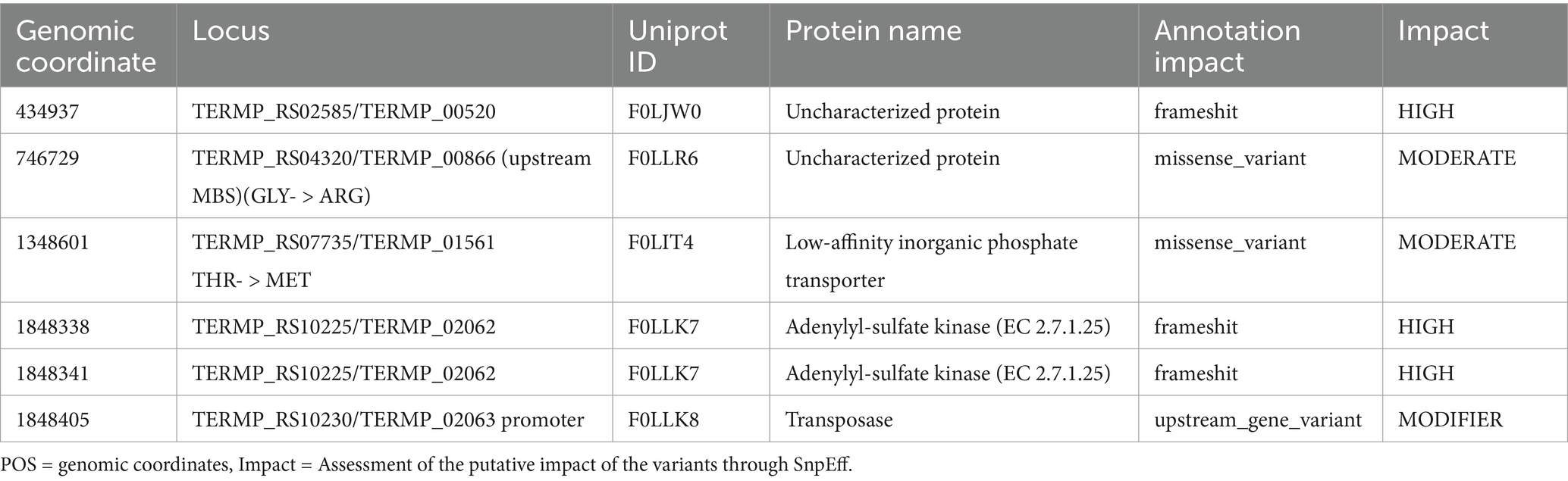
Table 1. Specific variants detected in the ∆517p strain and their potential effects.
3.2 SurR binding motif distribution in energy conservation gene promoters
We extended our genomic analysis by systematically screening the T. barophilus chromosome for SurR recognition motifs (GTTn3ATC or GTTn3AACn5GTT). As observed in other Thermococcales, both long and short motifs were detected upstream of genes clusters involved in electron flow (Table 2). Notably, Thermococcales exhibit diversity in the types and abundance of cytosolic and membrane-bound hydrogenases (Schut et al., 2013). In T. barophilus, the membrane-bound hydrogenase gene cluster exists in two adjacent copies (Mrp-mbh 1 and Mrp-mbh 2, Figure 1), whereas the two cytosolic hydrogenases gene clusters (shI and shII) are separated by ~400 kb in the genome (Table 2; Figure 1). Interestingly, canonical SurR binding motifs were detected only in the promoters of Mrp-mbh2 and shII, while mrp-mbh1 and shI contain alterations to this motif (Table 2). Moreover, these gene clusters (mrp-mbh1 and shI) showed consistently higher expression than their respective duplicates (Vannier et al., 2015; Batista et al., 2024). For the sulfane sulfur reductase complex, mrp-mbs, four SurR binding sites were identified, with half exhibiting mutations. Additionally surR and pdo/glutaredoxin share two SurR binding sites in their operator/promoter region (Table 2; Figure 1). T. barophilus also possesses a unique Mrp-Mbh-Codh complex, enabling growth on carbon monoxide (Kozhevnikova et al., 2016). While its transcription is regulated by pressure variation (Vannier et al., 2015), it has been proposed that in Thermococcus onnurineus, this complex is controlled by a regulator other than SurR (Lee et al., 2016). In T. barophilus, despite the presence of seven short binding sites within its cluster, only one mutated binding site is found in the promoter of the first coding sequence (Table 2). In this study, with the exception of Mrp-Mbh-Codh complex and pdo, all the gene clusters of Table 2 were genetically deleted. Total deletions were made, except for the surR gene, where 137 bp from its 5′ ends were deleted. Their effects on physiological growth were assessed in sulfur-present and sulfur-absent conditions across different hydrostatic pressures (0.1, 40, and 70 MPa).
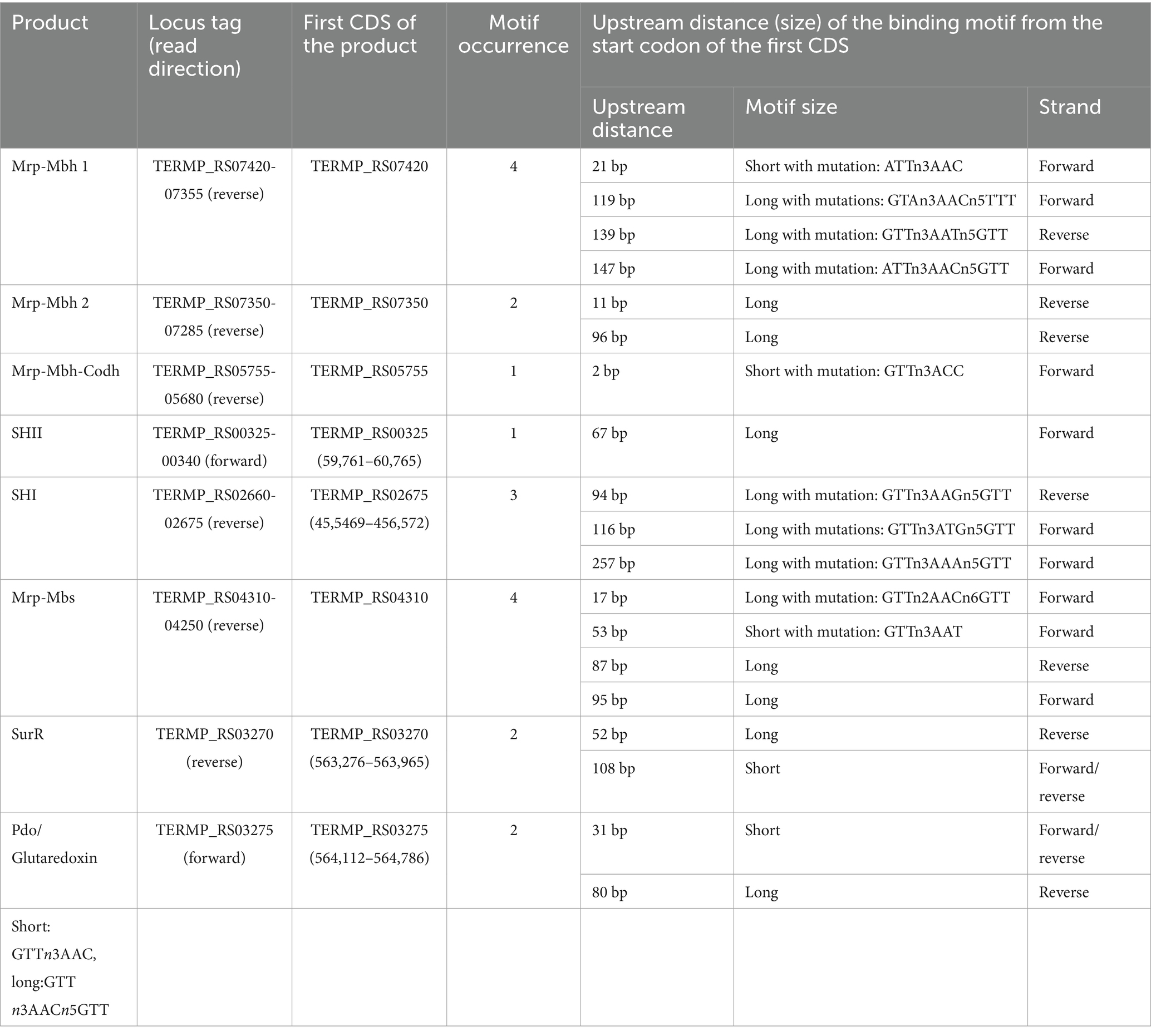
Table 2. SurR binding motifs positions upstream from the start codon of CDS involved in the electron flow.

Figure 1. Genetic organization of gene clusters involved in hydrogenogenic/sulfidogenic metabolisms and their SurR binding motifs. The MBH locus (A) composed of two Mrp-Mbh clusters, the MBS locus (B), the SHII locus (C), the SHI locus (D), and the SurR/Pdo locus (E).
3.3 Growth of wild type and derivative mutants under sulfur and hydrostatic pressure variations
Since genetic modifications were performed at atmospheric pressure (0.1 MPa), we first assessed the physiological impact of mutations under these conditions (Figure 2). Growth was evaluated with (Figure 2A) or without sulfur (Figure 2B). With sulfur, the ∆mbh strain exhibited only a slight growth delay compared to the parental strain ∆517 (see Table 3 for growth rate) while ∆mbs strain showed a more pronounced delay and reduced growth rate, leading to lower biomass after 24 h. Without sulfur, both ∆mbh and ∆mbs strains experienced severe growth impairment, failing to exceed a 1-log increase in cell density within 24 h, with growth rates below 0.1 h−1 (0.023 and 0.032, respectively). This contrasts with previous findings in P. furiosus and T. kodakarensis, where the Mrp-Mbs cluster had no impact on sulfur-free growth (Kanai et al., 2005; Bridger et al., 2011; Kanai et al., 2011; Santangelo et al., 2011). Growth experiments at 40 MPa, the optimal pressure for T. barophilus (Figures 2C,D), showed that in the presence of sulfur, both mutants experienced initial growth delays but reached “parental strain”-like final densities after 24 h. Without sulfur, delays were more pronounced, and final cell concentrations were significantly lower than ∆517 (Figure 2D; Table 3). Notably, despite lower overall growth rates, ∆mbh strain initially compensated for its delay after 15 h, while ∆mbs strain did so after 24 h, suggesting complementarity function or physiological synergy between MBS and MBH in sulfur-independent growth. Additionally, the overexpression of mrp-mbh clusters at 0.1 MPa in sulfur-rich conditions (Vannier et al., 2015) does not appear essential for survival, as their deletion did not dramatically impaired growth.
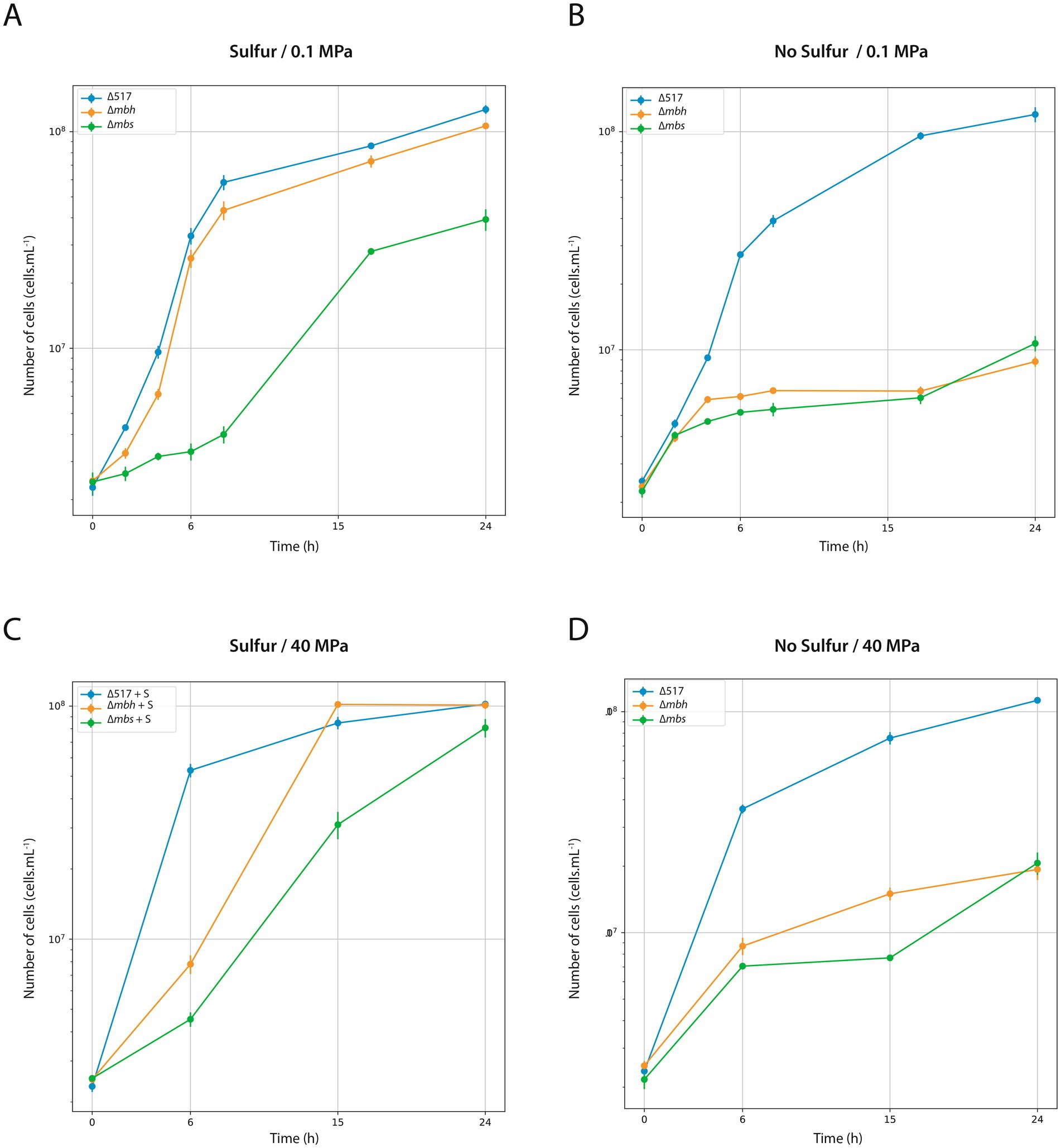
Figure 2. Characterization of ∆517, ∆mbh and ∆mbs mutants at 0.1 MPa (A,B) and 40 MPa (C,D). Growth assays were carried out in TRMm medium at 85°C, with sulfur (A,C) or without sulfur (B,D).
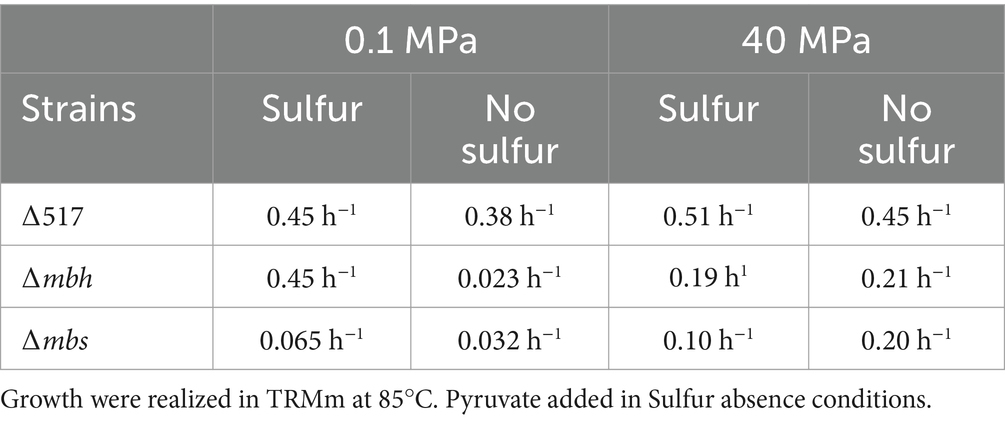
Table 3. Growth rate of ∆517, ∆mbh and ∆mbs mutants according to Sulfur and HHP conditions.
Next, we assessed strains lacking cytosolic hydrogenases under sulfur conditions at 0.1 MPa. With sulfur, ∆shI strain exhibited a more pronounced log-phase delay than ∆shII strain (Supplementary Figure S2A). Without sulfur, ∆shI strain displayed the most significant growth impairment, reaching lower cell concentrations than ∆517 and ∆shII strains after 24 h (Supplementary Figure S2B). These results suggest that some sulfur-grown mutants can metabolically adapt, but those lacking membrane-bound clusters (∆mbh and ∆mbs strains) cannot, likely due to the combined effects of genetic background and sulfur-dependent metabolic constraints.
Finally, we examined SurR’s role in growth across varying pressures and sulfur conditions (Figure 3; Table 4). Under sulfur conditions, ∆surR exhibited a growth delay at all pressures (Figure 3A). At 40 MPa, this delay was nearly compensated within 8 h, whereas at 70 MPa it took 10 h, and at 0.1 MPa over 14 h. Without sulfur, growth was severely impaired at all pressures (Figure 3B), but inhibition lessened as pressure increased, suggesting that pressure promotes cell recovery. Unlike non-piezophilic Thermococcales (e.g., P. furiosus and T. kodakarensis), where surR deletion mainly affects sulfur-free growth (Santangelo et al., 2011; Lipscomb et al., 2017), our results indicate a role for SurR even in sulfur-containing conditions. This supports the hypothesis that SurR regulates hydrogen metabolism genes regardless of sulfur availability, potentially explaining their upregulation under sub-optimal pressure in piezophiles like T. barophilus (Vannier et al., 2015) and T. piezophilus (Moalic et al., 2021). Taken together, these findings highlight how pressure modulates molecular responses for cellular adaptation (Table 5).
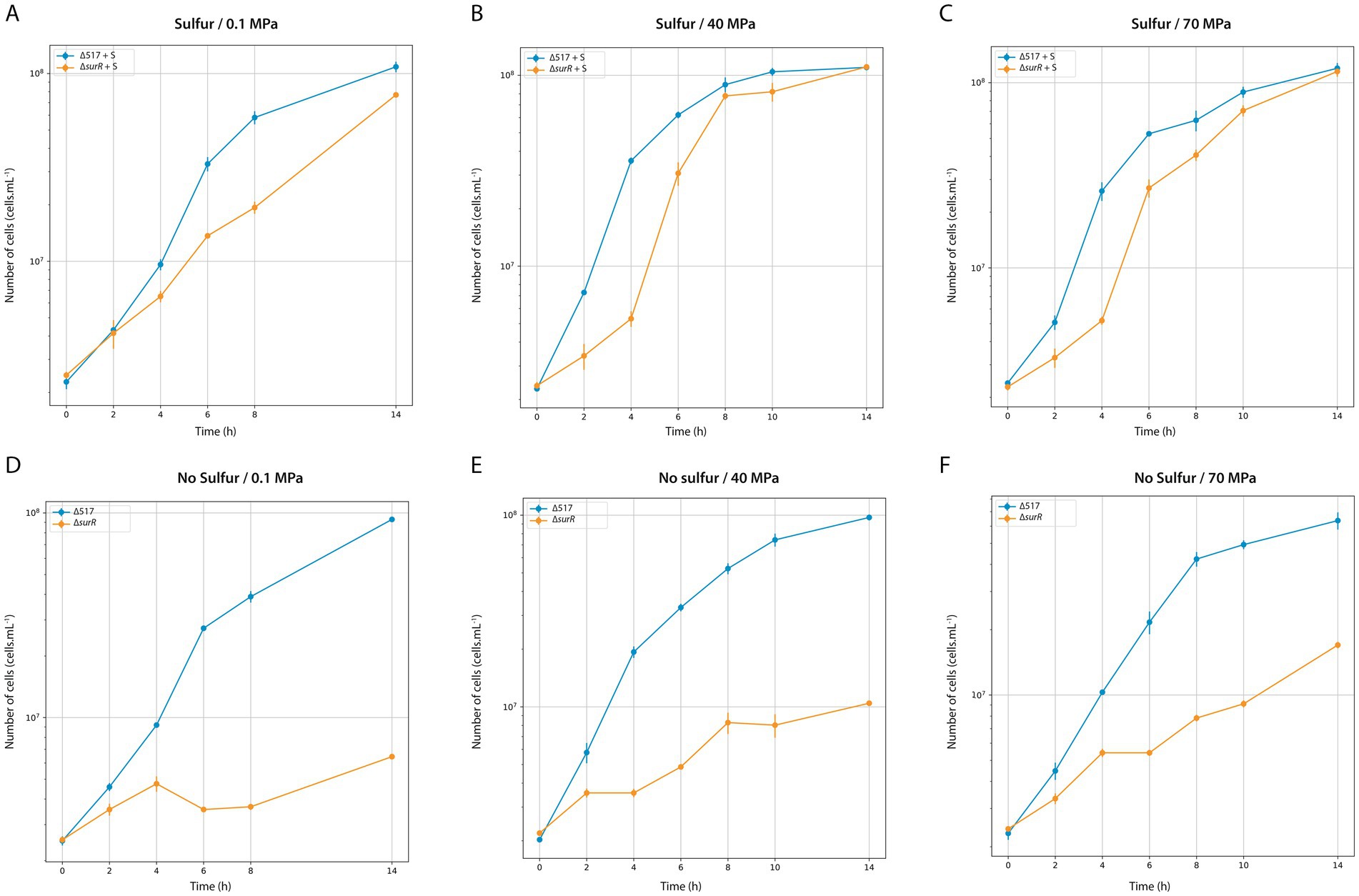
Figure 3. Characterization of ∆517 and ∆surR mutants at 0.1 MPa (A,D), 40 MPa (B,E) and 70 MPa (C,F). Growth assays were carried out in TRMm medium at 85°C, with sulfur (A,C) or without sulfur (B,D).
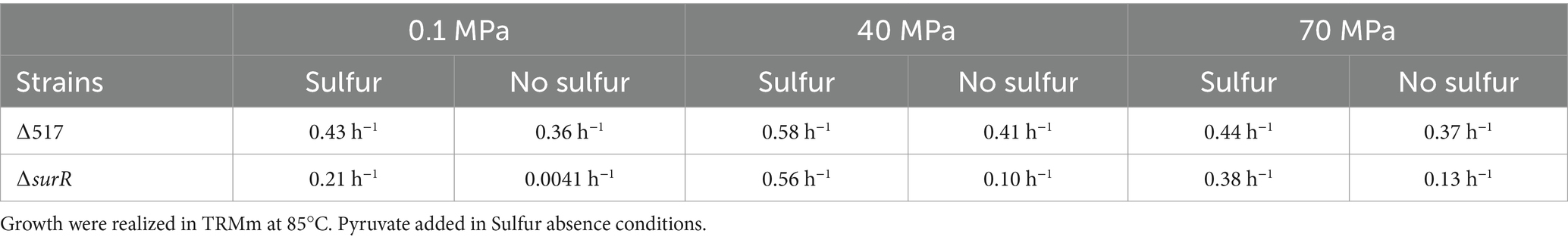
Table 4. Growth rate of ∆517 and ∆surR mutant according to Sulfur and HHP conditions.
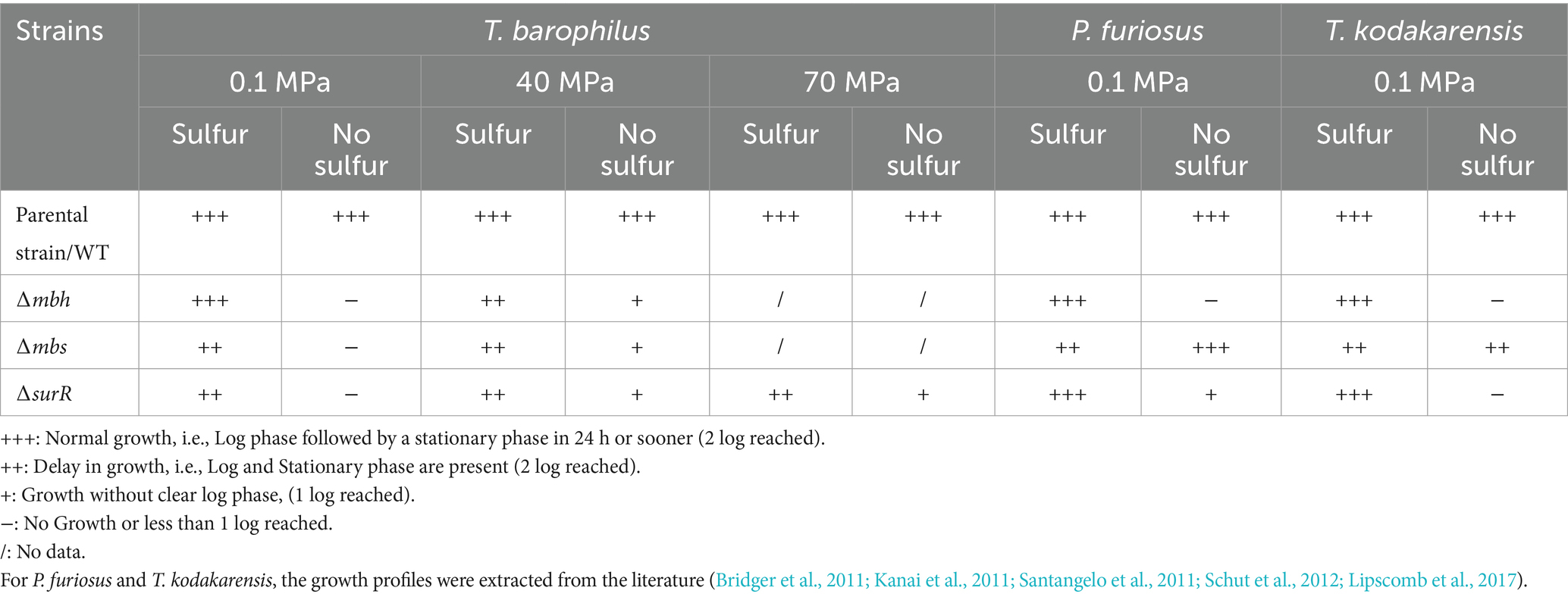
Table 5. Summary of the growth capacity of Thermococcales genetic strain models T. barophilus, P. furiosus, and T. kodakarensis under sulfidogenic or hydrogenogenic conditions.
3.4 Influence of SurR on the expression of targeted energetic metabolism genes
Given the distribution of canonical and variant SurR Binding sites in the promoters of genes and gene clusters involved in energy metabolism (Figure 1; Table 2), we performed real-time quantitative PCR to assess SurR’s regulatory impact under varying sulfur and pressure conditions. Among the three reference genes tested, only pcna showed stable Ct values across experiments, and ∆Ct calculations were based on this reference. Six target genes were selected from clusters encoding SHI, SHII, Mrp-Mbh1, Mrp-Mbh2, MBS, and SurR, based on their expression levels in sulfur conditions (Vannier et al., 2015). Since hydrogenogenic gene cluster expression increases at 0.1 MPa compared to 40 MPa in sulfur conditions, we compared their expression in the parental strain (∆517) and SurR mutant strain (∆surR) under three pressures (0.1, 40, and 70 MPa) with and without sulfur (Figure 4). At 0.1 MPa, in the presence of sulfur, ∆surR exhibited significantly reduced expression for all target genes except mbs and surR, confirming SurR’s role in stimulating gene expression under suboptimal pressure despite sulfur availability (Figure 4A, left panel). Without sulfur, expression remained lower in ∆surR, though standard deviations overlapped, except for mbhI and shI (Figure 4A, right panel). A At 40 MPa, in the presence of sulfur, all genes were downregulated in ∆surR, though standard deviations overlapped for mbs and surR (Figure 4B, left panel), reinforcing SurR’s role in regulating hydrogenogenic genes under sulfur conditions. Without sulfur, all genes were downregulated except mbs and surR, which were overexpressed, suggesting distinct regulatory mechanisms (Figure 4B, right panel). At 70 MPa, a distinct regulatory pattern emerged, with shII upregulated 18-fold and mbhII threefold (Figure 4C, left panel). Without sulfur, expression patterns resembled those at 40 MPa, but with stronger mbs upregulation (Figure 4C, right panel). These findings confirm SurR’s influence on hydrogen metabolism gene regulation, particularly for mbhI and shI, under suboptimal and optimal pressure conditions. At 70 MPa, SHII and Mrp-Mbh2 appear to play a greater role, suggesting a specific physiological synergy between membrane-bound complexes and cytosolic hydrogenases: SHII with Mrp-Mbh2, and SHI with Mrp-Mbh1. This aligns with the similarity of their SurR binding motifs (Figure 1; Table 2).
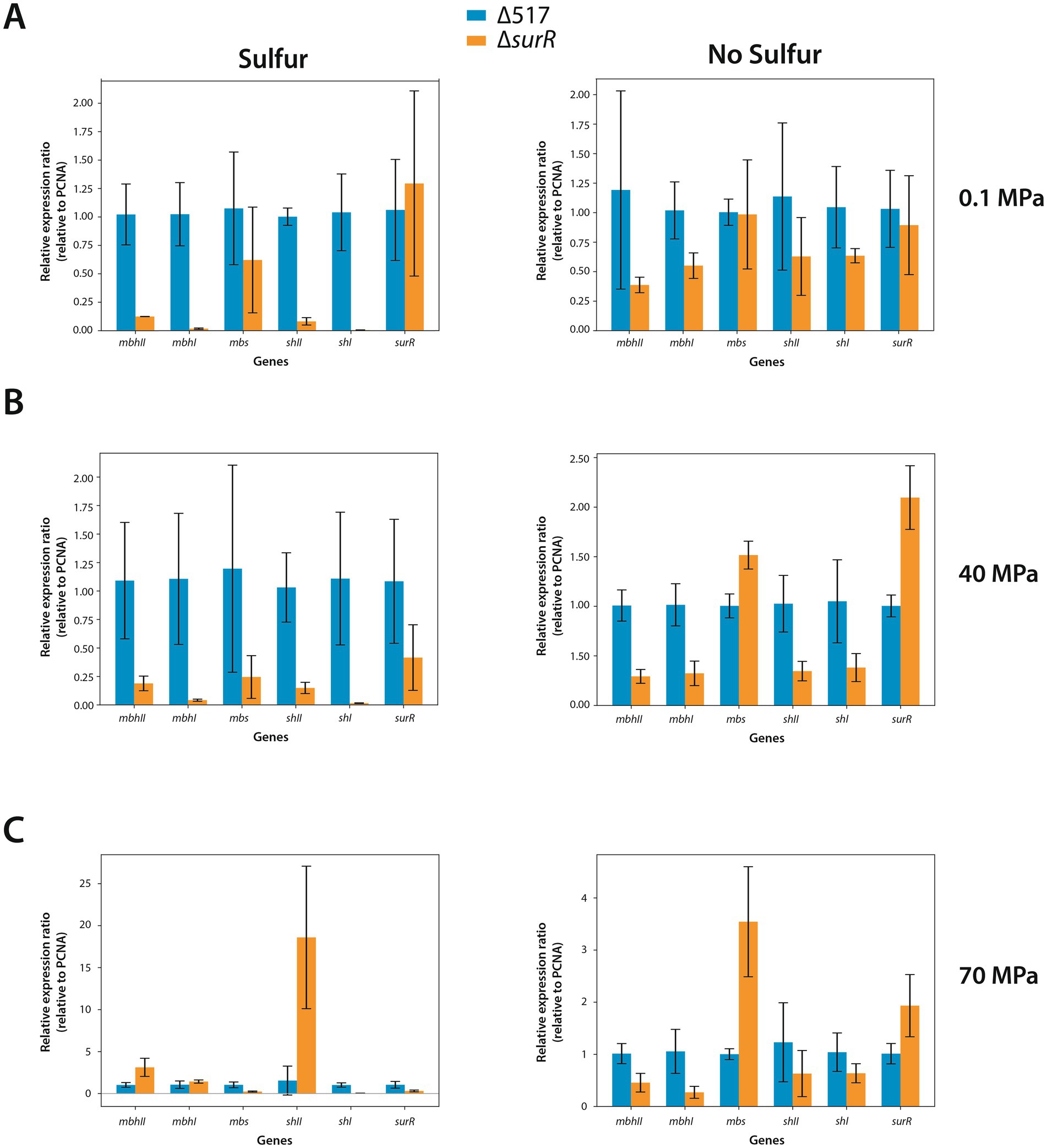
Figure 4. Effect of SurR DNA binding domain deletion on Gene expression variation of genes involved in hydrogenogenic/sulfidogenic metabolisms. The bars represent the expression ratio relative to PCNA gene (proliferating cell nuclear antigen). The blue bars symbolize the expression ratio of the ∆517 strain while the orange bars symbolize the expression ratio of the ∆surR strain. Growth rates are indicated in Table 4. Experiments were realized at 0.1 MPa (A), 40 MPa (B) and 70 MPa (C).
To further investigate sulfur-driven regulation, we measured gene expression 30 min after sulfur addition (0.5 g/L or 2 g/L). As 0.25 g/L supports normal T. barophilus growth, these concentrations were chosen to enhance regulatory signals. In the parental strain (∆517), sulfur addition strongly repressed hydrogenogenic gene expression (Figure 5, upper panels), particularly mbhI and shI, whereas mbs and surR showed inconsistent trends between sulfur concentrations. In ∆surR, at 0.5 g/L, expression responses were more variable, with mbhI and shII showing strong fluctuations, while mbhII and shI decreased clearly (Figure 5, lower left panel). At 2 g/L, all hydrogenogenic genes except shI were downregulated, while mbs and surR displayed high variability (Figure 5, lower right panel). These results, obtained at 0.1 MPa, confirm that sulfur addition triggers a metabolic shift, reducing hydrogenogenic gene expression. They also reinforce SurR’s role in sulfur regulation, as repression was less pronounced in ∆surR, consistent with findings in P. furiosus (Lipscomb et al., 2017).
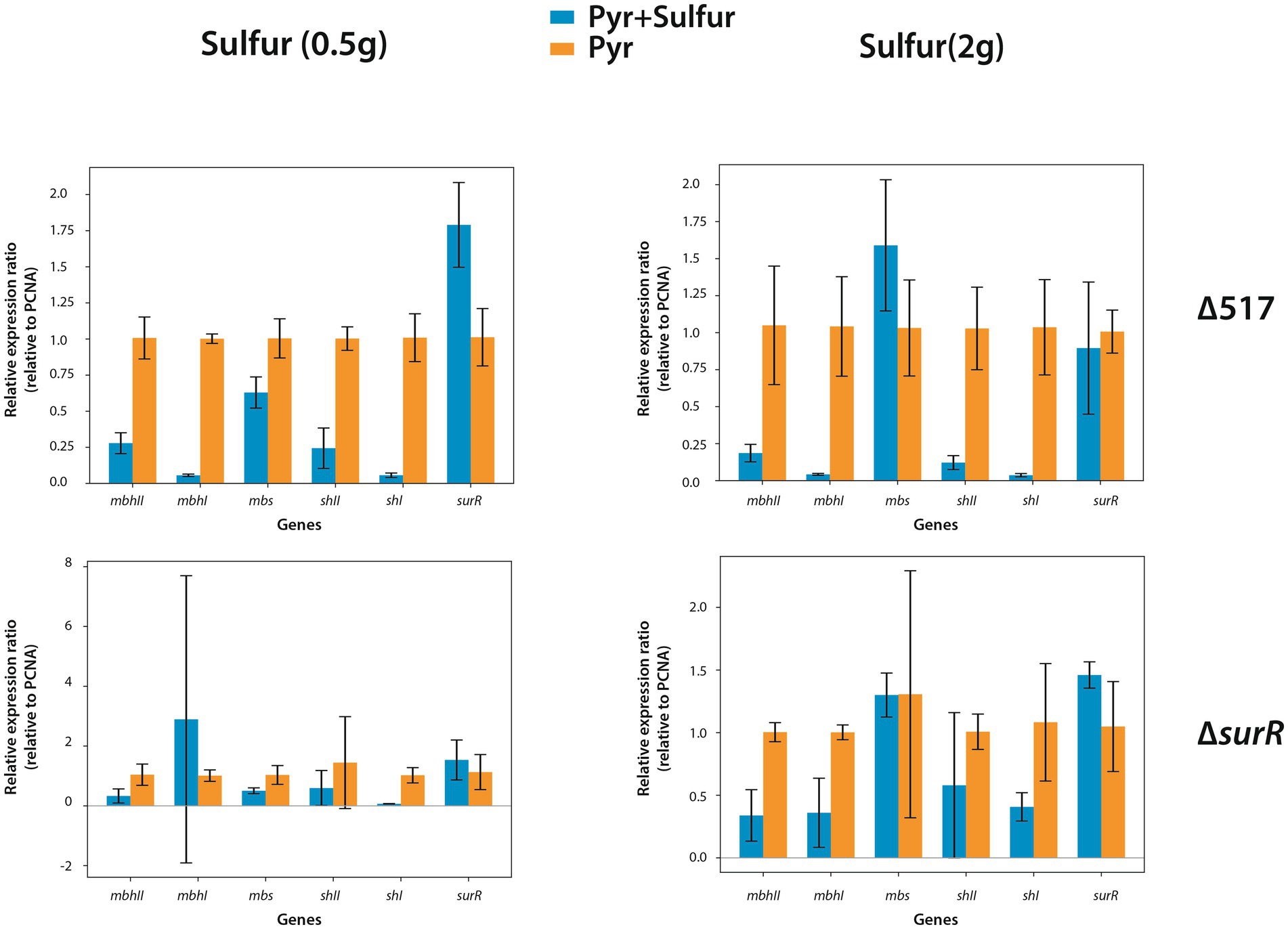
Figure 5. Effect of elemental sulfur (S°) addition on the expression of genes involved in hydrogenogenic and sulfidogenic metabolisms. Gene expression levels are shown as ratios relative to the housekeeping gene PCNA. Orange bars represent gene expression in strains prior to S° addition, while blue bars indicate expression levels 30 minutes after S° supplementation.
4 Conclusions and perspectives
This study provides new insights into the regulatory mechanisms controlling energy metabolism in Thermococcus barophilus under varying pressure and sulfur conditions. We demonstrate that SurR, a redox-sensitive transcriptional regulator, plays a central role in metabolic adaptation, balancing hydrogen and sulfur metabolism in response to pressure. Under suboptimal pressure (0.1 MPa), SurR activates hydrogenogenic genes despite sulfur availability. At optimal (40 MPa) and high pressure (70 MPa), SurR’s regulatory dynamics shift, leading to a distinct reconfiguration of gene expression.
We also identify a regulatory association between membrane-bound hydrogenases (Mrp-Mbh1/Mrp-Mbh2) and cytosolic hydrogenases (SHI/SHII), in the adaptive response to pressure, which could suggest a functional association Given that pressure affects protein folding activity (Chen and Makhatadze, 2017), further investigation into the three-dimensional structure of SurR and its physical interaction with DNA promoters under these pressure conditions is needed. Techniques such as real-time In Vitro Fluoresence anisotropy (Heisler et al., 2021) or Fluorescence Cross-Correlation Spectroscopy (FCCS), which can be applied under pressure (Bourges et al., 2017) provide valuable insights. Finally, integrating global “omic” approaches such as RNA-seq and Chip-seq will offer a comprehensive view of SurR regulation, helping to link its position at promoter regions to its role in gene regulation. These findings contribute to the broader field of extremophile biology and have implications for biotechnological applications and astrobiology, where life must adapt to fluctuating energy landscapes in high-pressure environments.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://www.ncbi.nlm.nih.gov/, accession number: PRJNA1234085.
Author contributions
YM: Data curation, Visualization, Conceptualization, Writing – review & editing, Investigation, Validation, Methodology, Writing – original draft, Supervision, Formal analysis. TN: Formal analysis, Investigation, Writing – review & editing. JH: Investigation, Formal analysis, Validation, Conceptualization, Methodology, Writing – review & editing. TB: Writing – review & editing, Investigation. AT: Conceptualization, Methodology, Writing – review & editing, Investigation. MJ: Formal analysis, Writing – review & editing, Writing – original draft, Validation, Supervision, Funding acquisition, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Agence Nationale de la Recherche (ANR-10-BLAN-1725 01-Living deep; ANR-16-CE12-0016 CASPAR and ANR-22-CE02-0019-01 Hot Dog). AT received a postdoctoral fellowship from the Conseil départemental du Finistère and Ifremer. YM and TB were supported by UBO.
Acknowledgments
We sincerely thank Marion Gardette, Betty Emonot, and Fatima-Zohra Masmoudi for their invaluable assistance in constructing and screening of the ∆mbs and ∆surR strains. We also extend our gratitude to Nadège Quintin for providing the strains used in this study, which are deposited in the UBO culture collection (https://ent.univ-brest.fr/lm2e/home/#/).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. Generative AI was used for English correction.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1593936/full#supplementary-material
SUPPLEMENTARY FIGURE 1 | Characterization of ∆517 strains with or without sulfur at 0.1 MPa. Growth assay were carried out at 85°C in TRM and TRMm media. TRMm containing Pyruvate (5g.L-1) was used for ∆517p strain growths. The ∆517p strain was previously adapted 96h in this medium before being subcultured for the growth kinetics. The growth rates are 0.059 h-1 (∆517, blue curve), 0.35 h-1 (∆517 + S, orange curve), 0.41 h-1 (∆517p, green curve) and 0.48 h-1 (∆517p + S, red curve).
Supplementary FIGURE 2 | Characterization of ∆shI and ∆shII mutants at 0.1 MPa. Growth assays were carried out in TRMm medium at 85°C, with sulfur (A) or without Sulfur (B). With sulfur, their respective growth rates are 0.41 h-1 and 0.28 h-1, while without sulfur, their respective growth rates are 0.33 h-1 and 0.11 h-1.
References
Batista, M., Langendijk-Genevaux, P., Kwapisz, M., Canal, I., and Phung, D. (2024). Evolutionary and functional insights into the Ski2-like helicase family in Archaea: a comparison of Thermococcales ASH-Ski2 and Hel308 activities. NAR Genom. Bioinform. 6:lqae026. doi: 10.1093/nargab/lqae026
Birien, T., Thiel, A., Henneke, G., Flament, D., Moalic, Y., and Jebbar, M. (2018). Development of an effective 6-methylpurine Counterselection marker for genetic manipulation in Thermococcus barophilus. Genes (Basel) 9:77. doi: 10.3390/genes9020077
Bourges, A. C., Torres Montaguth, O. E., Ghosh, A., Tadesse, W. M., Declerck, N., Aertsen, A., et al. (2017). High pressure activation of the Mrr restriction endonuclease in Escherichia coli involves tetramer dissociation. Nucleic Acids Res. 45, 5323–5332. doi: 10.1093/nar/gkx192
Bridger, S. L., Clarkson, S. M., Stirrett, K., Debarry, M. B., Lipscomb, G. L., Schut, G. J., et al. (2011). Deletion strains reveal metabolic roles for key elemental sulfur-responsive proteins in Pyrococcus furiosus. J. Bacteriol. 193, 6498–6504. doi: 10.1128/JB.05445-11
Burkhart, B. W., Febvre, H. P., and Santangelo, T. J. (2019). Distinct physiological roles of the three ferredoxins encoded in the hyperthermophilic archaeon Thermococcus kodakarensis. MBio 10:e02807-18. doi: 10.1128/mBio.02807-18
Cario, A., Jebbar, M., Thiel, A., Kervarec, N., and Oger, P. M. (2016). Molecular chaperone accumulation as a function of stress evidences adaptation to high hydrostatic pressure in the piezophilic archaeon Thermococcus barophilus. Sci. Rep. 6:29483. doi: 10.1038/srep29483
Cario, A., Lormieres, F., Xiang, X., and Oger, P. (2015). High hydrostatic pressure increases amino acid requirements in the piezo-hyperthermophilic archaeon Thermococcus barophilus. Res. Microbiol. 166, 710–716. doi: 10.1016/j.resmic.2015.07.004
Chen, C. R., and Makhatadze, G. I. (2017). Molecular determinant of the effects of hydrostatic pressure on protein folding stability. Nat. Commun. 8:14561. doi: 10.1038/ncomms14561
Chou, C. J., Shockley, K. R., Conners, S. B., Lewis, D. L., Comfort, D. A., Adams, M. W., et al. (2007). Impact of substrate glycoside linkage and elemental sulfur on bioenergetics of and hydrogen production by the hyperthermophilic archaeon Pyrococcus furiosus. Appl. Environ. Microbiol. 73, 6842–6853. doi: 10.1128/AEM.00597-07
Cingolani, P., Platts, A., Wang Le, L., Coon, M., Nguyen, T., Wang, L., et al. (2012). A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly (Austin) 6, 80–92. doi: 10.4161/fly.19695
Heisler, J., Chavan, A., Chang, Y.-G., and Liwang, A. (2021). “Real-time in vitro fluorescence anisotropy of the cyanobacterial circadian clock” in Circadian clocks: Methods and protocols. ed. S. A. Brown (New York, NY: Springer US), 3–18.
Hidese, R., Yamashita, K., Kawazuma, K., Kanai, T., Atomi, H., Imanaka, T., et al. (2017). Gene regulation of two ferredoxin:NADP(+) oxidoreductases by the redox-responsive regulator SurR in Thermococcus kodakarensis. Extremophiles 21, 903–917. doi: 10.1007/s00792-017-0952-0
Jager, D., Forstner, K. U., Sharma, C. M., Santangelo, T. J., and Reeve, J. N. (2014). Primary transcriptome map of the hyperthermophilic archaeon Thermococcus kodakarensis. BMC Genomics 15:684. doi: 10.1186/1471-2164-15-684
Kanai, T., Imanaka, H., Nakajima, A., Uwamori, K., Omori, Y., Fukui, T., et al. (2005). Continuous hydrogen production by the hyperthermophilic archaeon, Thermococcus kodakaraensis KOD1. J. Biotechnol. 116, 271–282. doi: 10.1016/j.jbiotec.2004.11.002
Kanai, T., Matsuoka, R., Beppu, H., Nakajima, A., Okada, Y., Atomi, H., et al. (2011). Distinct physiological roles of the three [NiFe]-hydrogenase orthologs in the hyperthermophilic archaeon Thermococcus kodakarensis. J. Bacteriol. 193, 3109–3116. doi: 10.1128/JB.01072-10
Kozhevnikova, D. A., Taranov, E. A., Lebedinsky, A. V., Bonch-Osmolovskaya, E. A., and Sokolova, T. G. (2016). Hydrogenogenic and sulfidogenic growth of Thermococcus archaea on carbon monoxide and formate. Microbiology 85, 400–410. doi: 10.1134/S0026261716040135
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Le Guellec, S., Leroy, E., Courtine, D., Godfroy, A., and Roussel, E. G. (2021). H2-dependent formate production by hyperthermophilic Thermococcales: an alternative to sulfur reduction for reducing-equivalents disposal. ISME J. 15, 3423–3436. doi: 10.1038/s41396-021-01020-x
Lee, S. H., Kim, M.-S., Lee, J.-H., Kim, T. W., Bae, S. S., Lee, S.-M., et al. (2016). Adaptive engineering of a hyperthermophilic archaeon on CO and discovering the underlying mechanism by multi-omics analysis. Sci. Rep. 6:22896. doi: 10.1038/srep22896
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The sequence alignment/map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Lim, J. K., Jung, H.-C., Kang, S. G., and Lee, H. S. (2017). Redox regulation of SurR by protein disulfide oxidoreductase in Thermococcus onnurineus NA1. Extremophiles 21, 491–498. doi: 10.1007/s00792-017-0919-1
Lipscomb, G. L., Keese, A. M., Cowart, D. M., Schut, G. J., Thomm, M., Adams, M. W., et al. (2009). SurR: a transcriptional activator and repressor controlling hydrogen and elemental Sulphur metabolism in Pyrococcus furiosus. Mol. Microbiol. 71, 332–349. doi: 10.1111/j.1365-2958.2008.06525.x
Lipscomb, G. L., Schut, G. J., Scott, R. A., and Adams, M. W. W. (2017). SurR is a master regulator of the primary electron flow pathways in the order Thermococcales. Mol. Microbiol. 104, 869–881. doi: 10.1111/mmi.13668
Malik, B., Su, W. W., Wald, H. L., Blumentals, I., and Kelly, R. M. (1989). Growth and gas production for hyperthermophilic archaebacterium, Pyrococcus furiosus. Biotechnol. Bioeng. 34, 1050–1057. doi: 10.1002/bit.260340805
Marteinsson, V. T., Birrien, J. L., Reysenbach, A. L., Vernet, M., Marie, D., Gambacorta, A., et al. (1999). Thermococcus barophilus sp. nov., a new barophilic and hyperthermophilic archaeon isolated under high hydrostatic pressure from a deep-sea hydrothermal vent. Int. J. Syst. Bacteriol. 49, 351–359. doi: 10.1099/00207713-49-2-351
Marteinsson, V. T., Watrin, L., Prieur, D., Caprais, J. C., Raguénès, G., and Erauso, G. (1995). Phenotypic characterization, DNA similarities and protein profiles of twenty sulfur-metabolizing hyperthermophilic anaerobic archaea isolated from hydrothermal vents in the southwestern Pacific Ocean. Int. J. Syst. Bacteriol. 45, 623–632. doi: 10.1099/00207713-45-4-623
Martinez, N., Michoud, G., Cario, A., Ollivier, J., Franzetti, B., Jebbar, M., et al. (2016). High protein flexibility and reduced hydration water dynamics are key pressure adaptive strategies in prokaryotes. Sci. Rep. 6:32816. doi: 10.1038/srep32816
Moalic, Y., Hartunians, J., Dalmasso, C., Courtine, D., Georges, M., Oger, P., et al. (2021). The piezo-Hyperthermophilic archaeon Thermococcus piezophilus regulates its energy efficiency system to cope with large hydrostatic pressure variations. Front. Microbiol. 12:730231. doi: 10.3389/fmicb.2021.730231
Moon, Y.-J., Kwon, J., Yun, S.-H., Lim, H. L., Kim, J., Kim, S. J., et al. (2015). Proteomic insights into sulfur metabolism in the hydrogen-producing Hyperthermophilic archaeon Thermococcus onnurineus NA1. Int. J. Mol. Sci. 16, 9167–9195. doi: 10.3390/ijms16059167
Nakagawa, S., and Takai, K. (2006). “The isolation of thermophiles from deep-sea hydrothermal environments,” in Extremophiles, methods in microbiology. eds. F. A. Rainey and A. Oren. (London, UK: Elsevier), 35, 57–91.
Postec, A., Le Breton, C., Fardeau, M.-L., Lesongeur, F., Pignet, P., Querellou, J., et al. (2005). Marinitoga hydrogenitolerans sp. nov., a novel member of the order Thermotogales isolated from a black smoker chimney on the mid-Atlantic ridge. Int. J. Syst. Evol. Microbiol. 55, 1217–1221. doi: 10.1099/ijs.0.63550-0
Santangelo, T. J., Cubonova, L., and Reeve, J. N. (2011). Deletion of alternative pathways for reductant recycling in Thermococcus kodakarensis increases hydrogen production. Mol. Microbiol. 81, 897–911. doi: 10.1111/j.1365-2958.2011.07734.x
Schäfer, T., and Schönheit, P. (1991). Pyruvate metabolism of the hyperthermophilic archaebacterium Pyrococcus furiosus. Arch. Microbiol. 155, 366–377. doi: 10.1007/BF00243457
Schmittgen, T. D., and Livak, K. J. (2008). Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 3, 1101–1108. doi: 10.1038/nprot.2008.73
Schut, G. J., Boyd, E. S., Peters, J. W., and Adams, M. W. W. (2013). The modular respiratory complexes involved in hydrogen and sulfur metabolism by heterotrophic hyperthermophilic archaea and their evolutionary implications. FEMS Microbiol. Rev. 37, 182–203. doi: 10.1111/j.1574-6976.2012.00346.x
Schut, G. J., Bridger, S. L., and Adams, M. W. W. (2007). Insights into the metabolism of elemental sulfur by the Hyperthermophilic archaeon Pyrococcus furiosus: characterization of a coenzyme A- dependent NAD(P)H sulfur oxidoreductase. J. Bacteriol. 189, 4431–4441. doi: 10.1128/JB.00031-07
Schut, G. J., Lipscomb, G. L., Han, Y., Notey, J. S., Kelly, R. M., and Adams, M. M. W. (2014). “The order Thermococcales and the family Thermococcaceae” in The prokaryotes: Other major lineages of Bacteria and the Archaea. eds. E. Rosenberg, E. F. Delong, S. Lory, E. Stackebrandt, and F. Thompson (Berlin, Heidelberg: Springer Berlin Heidelberg), 363–383.
Schut, G. J., Nixon, W. J., Lipscomb, G. L., Scott, R. A., and Adams, M. (2012). Mutational analyses of the enzymes involved in the metabolism of hydrogen by the hyperthermophilic archaeon Pyrococcus furiosus. Front. Microbiol. 3.
Thiel, A., Michoud, G., Moalic, Y., Flament, D., and Jebbar, M. (2014). Genetic manipulations of the Hyperthermophilic Piezophilic archaeon Thermococcus barophilus. Appl. Environ. Microbiol. 80, 2299–2306. doi: 10.1128/AEM.00084-14
Valentine, D. L. (2007). Adaptations to energy stress dictate the ecology and evolution of the Archaea. Nat. Rev. Microbiol. 5, 316–323. doi: 10.1038/nrmicro1619
Vannier, P., Marteinsson, V. T., Fridjonsson, O. H., Oger, P., and Jebbar, M. (2011). Complete genome sequence of the hyperthermophilic, piezophilic, heterotrophic, and carboxydotrophic archaeon Thermococcus barophilus MP. J. Bacteriol. 193, 1481–1482. doi: 10.1128/JB.01490-10
Vannier, P., Michoud, G., Oger, P., Marteinsson, V., and Jebbar, M. (2015). Genome expression of Thermococcus barophilus and Thermococcus kodakarensis in response to different hydrostatic pressure conditions. Res. Microbiol. 166, 717–725. doi: 10.1016/j.resmic.2015.07.006
Wilm, A., Aw, P. P., Bertrand, D., Yeo, G. H., Ong, S. H., Wong, C. H., et al. (2012). LoFreq: a sequence-quality aware, ultra-sensitive variant caller for uncovering cell-population heterogeneity from high-throughput sequencing datasets. Nucleic Acids Res. 40, 11189–11201. doi: 10.1093/nar/gks918
Wu, C. H., Schut, G. J., Poole, F. L. 2nd, Haja, D. K., and Adams, M. W. W. (2018). Characterization of membrane-bound sulfane reductase: a missing link in the evolution of modern day respiratory complexes. J. Biol. Chem. 293, 16687–16696. doi: 10.1074/jbc.RA118.005092
Yang, H., Lipscomb, G. L., Keese, A. M., Schut, G. J., Thomm, M., Adams, M. W. W., et al. (2010). SurR regulates hydrogen production in Pyrococcus furiosus by a sulfur-dependent redox switch. Mol. Microbiol. 77, 1111–1122. doi: 10.1111/j.1365-2958.2010.07275.x
Yu, H., Haja, D. K., Schut, G. J., Wu, C.-H., Meng, X., Zhao, G., et al. (2020). Structure of the respiratory MBS complex reveals iron-sulfur cluster catalyzed sulfane sulfur reduction in ancient life. Nat. Commun. 11:5953. doi: 10.1038/s41467-020-19697-7
Keywords: piezophilic archaea, high hydrostatic pressure, energy metabolism, transcriptional regulation, sulfur metabolism, gene expression, adaptation to extreme environments
Citation: Moalic Y, Nguyen TBH, Hartunians J, Birien T, Thiel A and Jebbar M (2025) Regulation of gene expression under high hydrostatic pressure: the versatile role of the master regulator SurR in energy metabolism. Front. Microbiol. 16:1593936. doi: 10.3389/fmicb.2025.1593936
Edited by:
Mark Alexander Lever, The University of Texas at Austin, United StatesReviewed by:
Doug Bartlett, University of California, San Diego, United StatesSatya P. Singh, Saurashtra University, India
Copyright © 2025 Moalic, Nguyen, Hartunians, Birien, Thiel and Jebbar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yann Moalic, eWFubi5tb2FsaWNAaXNlbi1vdWVzdC55bmNyZWEuZnI=; Mohamed Jebbar, bW9oYW1lZC5qZWJiYXJAdW5pdi1icmVzdC5mcg==
†Present address: Yann Moalic, L@BISEN - Laboratoire ISEN, LABISEN, Brest, France