 Isabela Sguilla Rotta1
Isabela Sguilla Rotta1 Sthefânia Dalva da Cunha Rezende1
Sthefânia Dalva da Cunha Rezende1 Hugo Felix Perini1,2
Hugo Felix Perini1,2 Marcos Vinicius da Silva1
Marcos Vinicius da Silva1 Felipe Alves de Almeida3
Felipe Alves de Almeida3 Uelinton Manoel Pinto4
Uelinton Manoel Pinto4 Alessandra Barbosa Ferreira Machado5
Alessandra Barbosa Ferreira Machado5 Aline Dias Paiva1*
Aline Dias Paiva1*- 1Department of Microbiology, Immunology and Parasitology, Federal University of Triângulo Mineiro, Uberaba, Brazil
- 2Department of Education in Science, Mathematics and Technology, Federal University of Triângulo Mineiro, Uberaba, Brazil
- 3Department of Microbiology, Institute of Biotechnology Applied to Agriculture, Federal University of Viçosa, Viçosa, Brazil
- 4Department of Food and Experimental Nutrition, Food Research Center, São Paulo University, São Paulo, Brazil
- 5Department of Parasitology, Microbiology and Immunology, Federal University of Juiz de Fora, Juiz de Fora, Brazil
Introduction: Weissella paramesenteroides UFTM 2.6.1, isolated from unpasteurized milk, is a potentially probiotic strain exhibiting desirable properties previously demonstrated in vitro, along with a confirmed safe and promising genetic profile based on whole-genome analysis. Due to the limited research on the ability of Weissella species to synthesize compounds with anti-quorum sensing activity, this study aimed to investigate the potential of W. paramesenteroides UFTM 2.6.1 to disrupt quorum sensing (QS) signaling and attenuate the virulence of Listeria monocytogenes, an important foodborne pathogen responsible for the zoonotic disease listeriosis.
Methods: The effects of W. paramesenteroides cell-free supernatant (Wp-CFS) were evaluated on the growth, biofilm formation, motility, and expression of QS- and virulence-related genes in L. monocytogenes.
Results: Wp-CFS exhibited bacteriostatic activity against L. monocytogenes strains isolated from food and food processing environments (n = 21). Additionally, it consistently reduced biofilm formation and swarming motility, two well-known QS-regulated phenotypes in L. monocytogenes. Exposure to Wp-CFS (0.25x MIC; 7.81 mg/mL), at 28°C for 24 h, significantly downregulated the relative expression of the genes luxS, agrA, flaA, motA, motB, and degU, whereas the genes sigB, and prfA were upregulated.
Conclusion: This study represents the first report demonstrating the production of compounds by W. paramesenteroides aimed at disrupting the QS system of L. monocytogenes, offering novel insights into alternative approaches to attenuate pathogen virulence without relying on traditional antimicrobials.
1 Introduction
The genus Weissella belongs to the group of bacteria known as lactic acid bacteria (LAB). The genus was first designated in 1993 after taxonomic studies on atypical Leuconostoc-like microorganisms. Bacteria assigned to the genus Weissella are commonly found in a variety of environments, including the gastrointestinal tract of humans and animals, plant-associated microbiomes, and fermented foods. They are known for their versatility in surviving different environmental conditions, such as varying pH and temperature levels (Collins et al., 1993). Some Weissella strains have been recognized for their probiotic potential, they can also inhibit the growth of bacterial pathogens, modulate gut microbiota, and immune responses (Fhoula et al., 2013). Taken together, these characteristics make Weissella an important genus in the field of food microbiology and human health. Among the species of the genus, Weissella confusa, Weissella cibaria, and Weissella paramesenteroides are the most widely studied (Fusco et al., 2023; Liu et al., 2024; Singh et al., 2024).
W. paramesenteroides UFTM 2.6.1, previously isolated from unpasteurized milk by our research group, shows the ability to survive in the presence of bile salts and acidic pH, and exhibits antagonistic properties against spoilage and pathogenic bacteria (Rotta et al., 2020). Recently, the complete genome sequencing of this strain uncovered genes related to probiotic properties, as well as absence of antimicrobial resistance determinants and other virulence genes, revealing a promising safety profile (Rocha et al., 2024).
Listeria monocytogenes is a well-known foodborne pathogen, notorious for its ability to withstand stress conditions. It can withstand various treatments commonly used in the food processing environment to control microbial growth, including heat, acidification, salt addition, preservatives, sanitizers, and high hydrostatic pressure. In addition, L. monocytogenes is capable of persisting in the environment for extended periods, largely due to its ability to form biofilms (Wiktorczyk-Kapischke et al., 2023; Cheng et al., 2023; Ribeiro et al., 2023; Tuytschaever et al., 2023). Biofilms are defined as aggregates of microbial cells adhered to solid surfaces and embedded in an extracellular polymeric matrix. Within biofilms, microbial cells exhibit increased resistance to sanitizers, detergents, and antimicrobials, and may also express specific virulence genes (Poimenidou et al., 2016; Gemmell et al., 2022).
Biofilm formation is intricately linked to quorum sensing (QS), an inter- and intraspecies communication mechanism that enables bacteria to assess population density through the release and detection of signaling molecules, known as autoinducers (AI). Upon reaching a critical concentration (threshold level), these AIs activate a coordinated response among microbial cells, leading to changes in gene expression. In many pathogens, including L. monocytogenes, QS systems are essential for the regulation of key virulence factors, such as biofilm formation, resilience to environmental stress, motility, and resistance to antimicrobial agents (Rieu et al., 2007; Brackman and Coenye, 2015).
Two main QS systems have been described in L. monocytogenes: the Agr-like system and the LuxS/AI-2 system. The Agr-like system, which is homologous to the Agr system found in Staphylococcus aureus, is used for intraspecies communication, and regulates the expression of genes involved in virulence and stress response (Skandamis and Nychas, 2012; Kocot and Olszewska, 2017). On the other hand, the LuxS/AI-2 system, which is more conserved across different bacterial species, mediates interspecies communication and influences biofilm formation and motility (Belval et al., 2006; Sela et al., 2006; Li et al., 2021; Yu et al., 2022). In this context, disrupting bacterial communication could offer a potential strategy to attenuate L. monocytogenes virulence (Marques et al., 2024).
Unlike bactericidal approaches, QS-targeting molecules, also named quorum-quenching (QQ) molecules, do not kill the target bacteria but instead they interfere with the signaling pathways that coordinate bacterial population behaviors. The QS mechanism can be inhibited by: (I) inhibition of AIs synthesis; (II) inhibition of AI secretion and transport; (III) degradation of AIs using either catalytic antibodies, such as abzymes or enzymes, such as lactonases, acylase, hydrolase, and oxidoreductase; (IV) sequestration of AIs using, for example, antibodies against AIs; (V) antibodies that “cover” and therefore block AI receptors; (VI) antagonists of AIs, such as chemical compounds; (VII) inhibition of targets downstream of the binding of the AI to the receptor; and (VIII) post-transcriptional regulation of QS genes via small regulatory RNAs (sRNAs) (Almeida et al., 2023). Marques et al. (2024) showed that proteins potentially secreted by Lacticaseibacillus rhamnosus GG and Lactobacillus acidophilus NCFM, two probiotic LAB, have the ability to in silico bind to the QS proteins Agr, AgrB and AgrC of L. monocytogenes.
Considering the variety of bioactive compounds synthesized by members of the Weissella genus, we hypothesized that W. paramesenteroides UFTM 2.6.1 could be a source of QQ molecules to disrupt the QS mechanisms in L. monocytogenes, attenuating the virulence of this important pathogen. By elucidating this interaction, we aim to provide insights into a novel strategy to reduce the virulence of L. monocytogenes without relying on bactericidal approaches.
2 Materials and methods
2.1 Microorganisms and growth conditions
W. paramesenteroides UFTM 2.6.1 was grown in Man-Rogosa-Sharpe (MRS) broth (Kasvi, Spain), at 37°C for 18–24 h, under microaerophilic conditions (Rotta et al., 2020). L. monocytogenes ATCC 19112 and 20 strains of L. monocytogenes isolated from different food and food processing sectors were donated by the Listeria Collection (CLIST) of the Bacterial Zoonoses Laboratory at the Oswaldo Cruz Institute (LABZOO/Fiocruz) (Table 1). L. monocytogenes strains were grown in Brain Heart Infusion (BHI) broth (Himedia, India), at 28 or 37°C (depending on the experiment), for 18–24 h, in aerobiosis. Bacteria were kept preserved at −20°C in 20% glycerol-containing BHI broth.
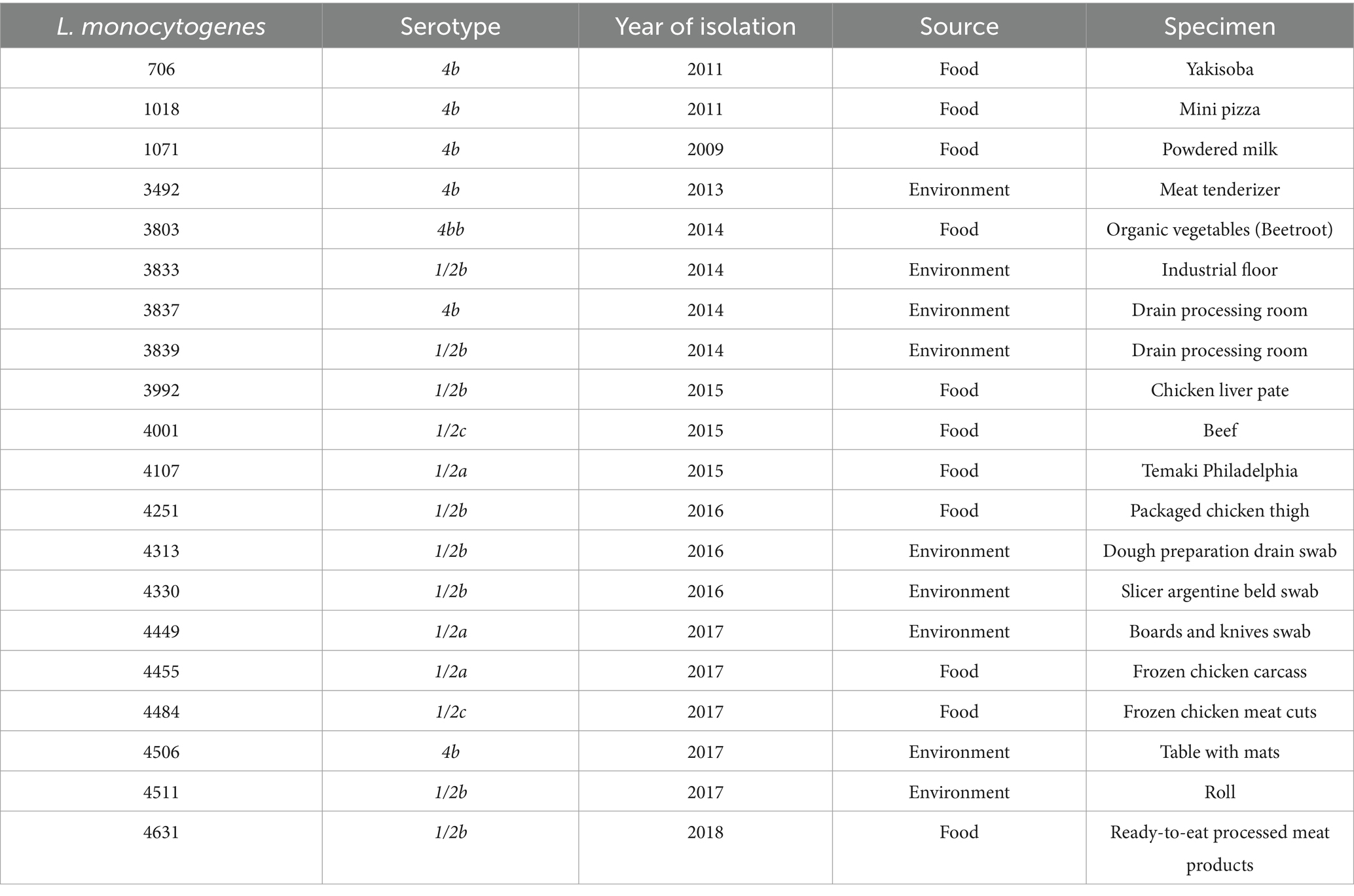
Table 1. Listeria monocytogenes strains: code, serotypes, year of isolation, sources and specimens.
2.2 Antagonistic activity against Listeria monocytogenes strains
The screening of antagonistic activity against L. monocytogenes strains was performed using the agar overlay method (Booth et al., 1977). Briefly, colonies of W. paramesenteroides UFTM 2.6.1 grown on MRS agar were overlaid with semi-solid BHI agar (0.75% agar) containing 106 CFU/mL of each L. monocytogenes strain separately. The plates were incubated overnight under aerobic conditions at 37°C. Anti-Listeria activity was determined by the presence of zones of inhibition (>6 mm in diameter) of L. monocytogenes growth around the colonies of W. paramesenteroides UFTM 2.6.1.
2.3 Cell-free supernatant obtention and evaluation of anti-Listeria activity
Cell-free supernatant (CFS) was obtained by centrifuging stationary-phase W. paramesenteroides UFTM 2.6.1 cultures (12,500 rpm, 15 min) and filtering the supernatant through a 0.22 μm filter. CFS was lyophilized, suspended in saline solution (0.85%) to a final concentration of 500 mg/mL, referred to as Wp-CFS. The Wp-CFS was stored at −20°C until use.
The inhibitory activity of Wp-CFS was evaluated using the micro-broth dilution technique according to the Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2023). Aliquots of 50 μL of Wp-CFS and its serial dilutions (twofold increments) were added to 96-well plates containing 50 μL of BHI broth (2x concentrated). Suspensions of stationary-phase L. monocytogenes cultures containing 106 CFU/mL (10 μL) were added to the microplates, and incubated at 37°C for 18 h under aerobic conditions. Uninoculated BHI broth was used as negative control, and uninoculated BHI broth was used as negative control, and BHI broth inoculated only with L. monocytogenes was used as positive control. Saline solution was included as an additional control. Minimum inhibitory concentration (MIC) was defined as the lowest concentration at which bacterial growth was completely inhibited visually. Sub-MICs were defined as concentration of the antimicrobial agent below the MIC value, where the agent did not inhibit bacterial growth but could still influence other bacterial processes (CLSI, 2023).
The minimum bactericidal concentration (MBC) for the Wp-CFS was determined after taking aliquots (5 μL) from the wells in the microplates showing no growth after MIC determination and inoculated onto the surface of BHI agar plates (CLSI, 2023). After incubation for 48 h at 37°C, the lowest concentration of Wp-CFS at which no bacterial colonies were formed was considered as MBC (Rodríguez-Melcón et al., 2022).
2.4 Time-kill kinetics study
Time-kill kinetic studies were performed in 1 mL of BHI broth inoculated with logarithmically grown L. monocytogenes to yield a final concentration of 5.0 × 106 CFU/mL, in the presence of different concentrations of Wp-CFS ranging from 0.25 to 4x MIC. All the flasks were incubated at 37°C and samples of 0.1 mL were taken at 0, 3, 6, 9, 12, 24, 48, and 72 h post-inoculation. Serial tenfold dilutions were prepared in sterile saline and used to determine CFU/mL. Time-kill curve was constructed by plotting log10 of CFU/mL versus time. Bactericidal activity was defined as a decrease of 99.99% (≥3 log10) in CFU/mL compared to the initial inoculum. Bacteriostatic activity was defined as maintenance of the original inoculum level or decrease of less than 99.99% (<3 log10) in CFU/mL compared to the initial sample (Petersen et al., 2007).
2.5 Listeria monocytogenes biofilm formation
All 21 L. monocytogenes strains were tested to determine their biofilm-forming ability on polystyrene microplates at 28°C (Guilbaud et al., 2015). Overnight cultures grown in BHI medium were adjusted to 0.5 McFarland turbidity standard, and 10 μL aliquots were transferred to a 96-well polystyrene microplate containing 100 μL of BHI broth. After incubation for 72 h at 28°C, the microplates were washed three times with sterile distilled water to remove planktonic cells. Crystal violet (0.25%, 150 μL) was added to each well of the microplate, and the plate was kept at room temperature for 30 min. The crystal violet solution was removed, washed with running water and allowed to dry; then ethanol (95%, 150 μL) was added to solubilize the stain. Absorbance was determined at 540 nm using a microplate reader. Experiments were performed in triplicate and repeated in three independent experiments. Mean optical density (OD) of the three wells for each sample was compared with the mean absorbance of negative controls, and the strains were classified in weak, moderate, strong or non-biofilm producers (Stepanóvic et al., 2000). Isolates classified as strong biofilm producers were selected for further experiments.
2.6 Anti-biofilm and anti-motility activities of Weissella paramesenteroides cell-free supernatant
The effect of Wp-CFS in L. monocytogenes biofilm formation was evaluated using the crystal violet method (Fan et al., 2018; Liu et al., 2021). The microplates were prepared as described in the biofilm formation assay and different concentrations of Wp-CFS (0.25x, 0.5x, and MIC) were added before the incubation of the microplates. After 72 h at 28°C, biofilm formation inhibition rates were calculated using the following formula:
.
L. monocytogenes biofilms treated with Wp-CFS were also evaluated using fluorescence microscopy and DAPI/PI dual staining. Briefly, the biofilm was prepared as described above was stained with 4′,6-diamidino-2-phenylindole (DAPI) (Sigma, UK; 10 μg/mL, 30 min) and propidium iodide (PI) (Sigma, UK; 10 μg/mL, 10 min). Non-treated biofilms were used as positive control. Images were taken using an inverted fluorescence microscope (Invitrogen EVOS FL) equipped with a 40x phase contrast objective and a Sony™ ICX285AL CCD camera.
Swimming and swarming motilities of L. monocytogenes strains were evaluated in semi-solid agar (Pieta et al., 2017; Jiang et al., 2021). Approximately 3 μL of stationary-phase L. monocytogenes cultures were inoculated into the center of plates containing culture medium for swimming (10 g/L tryptone, 5 g/L NaCl, and 0.3% agar) or swarming (25 g/L Luria-Bertani, 0.5 g/L glucose, and 0.5% agar), supplemented with sub-inhibitory concentrations of the Wp-CFS (0.125x, 0.25x, and 0.5x MIC). After incubation for 48 h at 28°C, the diameter of the motility zones was measured in millimeters (mm). Motility pattern in the absence of CFS was used as a control, and defined as 100% motility.
2.7 Influence of Weissella paramesenteroides-cell-free supernatant on Listeria monocytogenes target gene expression
The effect of Wp-CFS on the expression of genes involved in QS mechanism, motility and biofilm formation in L. monocytogenes was evaluated by quantitative reverse transcription polymerase chain reaction (RT-qPCR). L. monocytogenes was grown in BHI broth in the presence or absence of Wp-CFS (0.25x MIC) at 28°C for 24 h. The cultures were centrifuged (13,500 rpm, 5 min) and washed with sterile phosphate-buffered saline (PBS). Total RNA was extracted using Trizol (Sigma-Aldrich, United States) and converted to cDNA using the High-Capacity cDNA Reverse Transcription Kit (Thermo Fisher, United States). Validated and previously published primers were used (Table 2). The 16S rRNA gene was used as internal control.
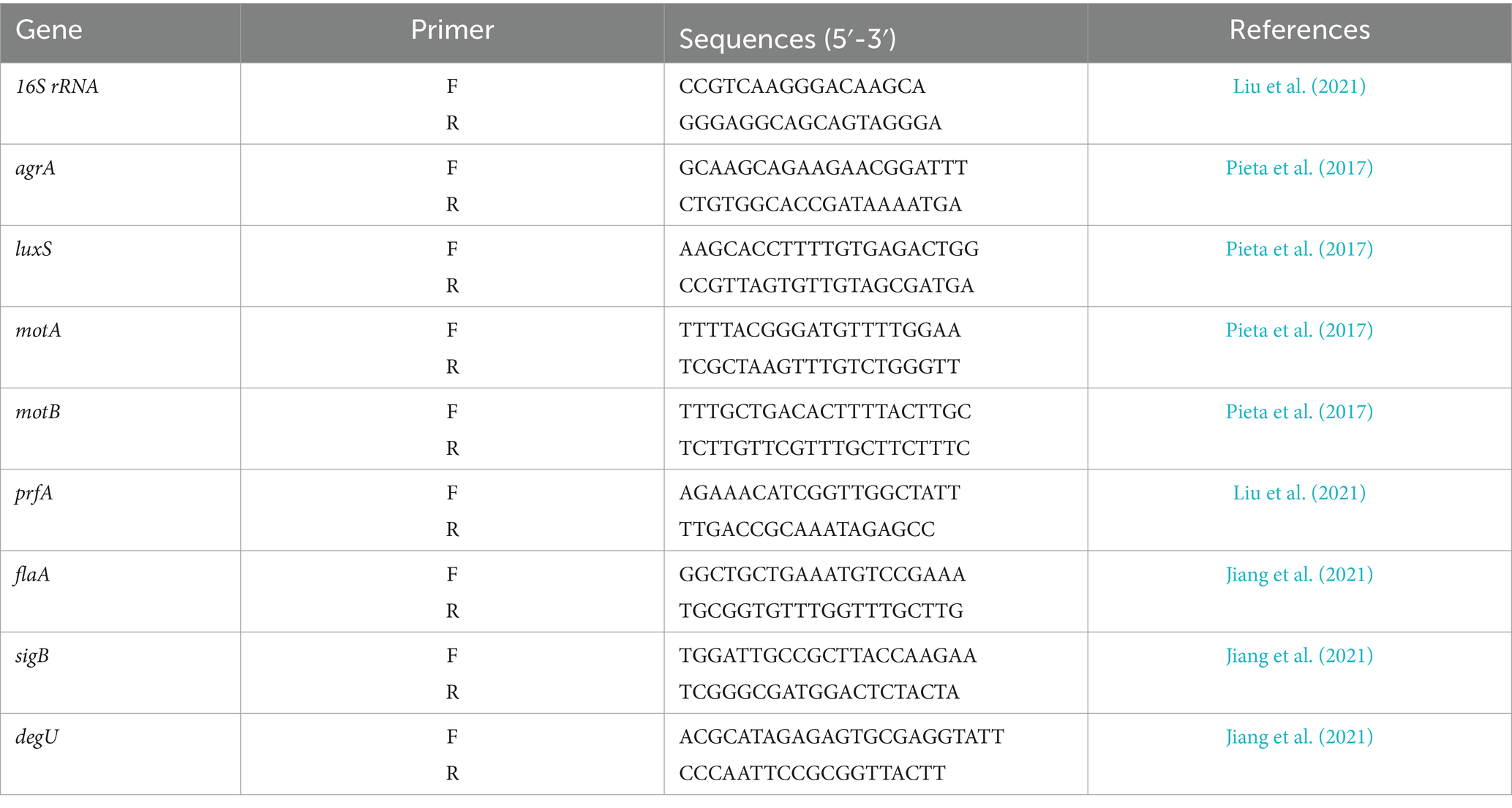
Table 2. Pairs of primers used in the present study for quantitative reverse transcription polymerase chain reaction (RT-qPCR).
For the RT-qPCR experiments, a reaction mix was prepared using PowerTrack SYBR Green Master Mix (Thermo Fisher, United States). Each reaction contained the following components: 5 μL of PowerTrack SYBR Green Master Mix, 0.5 μL of forward primer, 0.5 μL of reverse primer, 0.25 μL of yellow dye, 1 μL of cDNA (20 ng), and 3.25 μL of nuclease-free water, resulting in a final reaction volume of 10 μL. Amplification was carried out using the StepOnePlus™ Real-Time PCR System (Thermo Fisher, United States). The reaction conditions were as follows: an initial holding stage at 95°C for 2 min, followed by 40 cycles of denaturation at 95°C for 15 s and 60°C for 60 s. At the end of the amplification protocol, a melting curve analysis was also performed to confirm the specificity of each qPCR reaction (excluding any nonspecific amplification), which included 95°C for 15 s, 60°C for 1 min, and 95°C for 15 s. The 2−ΔΔCt method was used to analyze the relative expression of target genes using Ct values, with melting curve analysis confirming product specificity (Shi et al., 2023; Livak and Schmittgen, 2001). All experiments were performed in biological duplicates and experimental triplicates.
2.8 Statistical analysis
The statistical analyses were performed using GraphPad Prism 8.0 software (GraphPad Software Inc., La Jolla, CA, United States). Comparisons between treated groups and control groups were analyzed by ANOVA followed by Dunnett’s test for multiple comparisons to determine the influence of the Wp-CFS on biofilm formation and motility. Differences were considered statistically significant when p < 0.05. For the relative gene expression, the data were analyzed by ANOVA, and when statistically significant differences were identified (p < 0.05) in relative expression levels calculated by the 2−ΔΔCt method, the genes were considered upregulated (2−ΔΔCt > 1) or downregulated (2−ΔΔCt < 1) in the presence of Wp-CFS.
3 Results
3.1 Antagonistic activity of Weissella paramesenteroides UFTM 2.6.1 against Listeria monocytogenes strains
W. paramesenteroides UFTM 2.6.1 was able to inhibit the growth of L. monocytogenes strains on BHI agar overlaid, with inhibition zones ranging from 18 to 22 mm in diameter, suggesting the secretion of anti-Listeria compound(s) by this bacterium (Figure 1). The Wp-CFS inhibited the growth of all L. monocytogenes strains in liquid medium, and a concentration of 31.25 mg/mL was defined as the MIC. The MBC could not be determined, as L. monocytogenes formed colonies after exposure to Wp-CFS, even at concentrations up to 4x MIC (125.0 mg/mL).
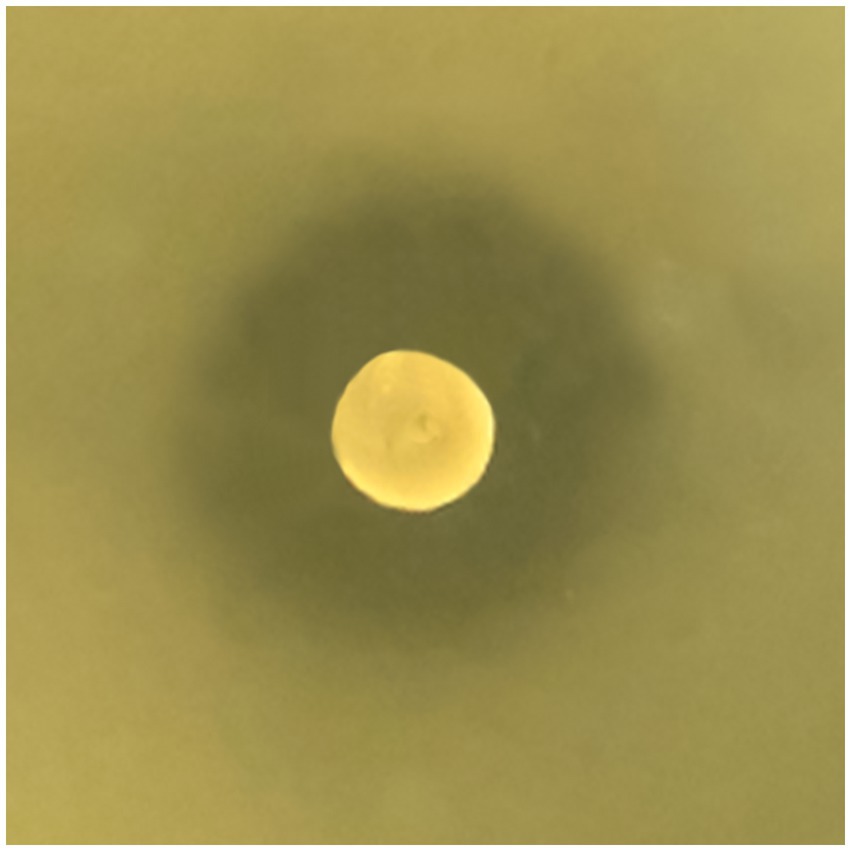
Figure 1. Weissella paramesenteroides UFTM 2.6.1 antagonistic activity against Listeria monocytogenes on solid medium. The representative image shows the growth inhibition of L. monocytogenes strain 4455, with an inhibition zone of 20 mm around the colony of W. paramesenteroides UFTM 2.6.1.
To further assess the bacteriostatic behavior of the compound produced by W. paramesenteroides UFTM 2.6.1, time-kill experiments against L. monocytogenes strains were performed. At a concentration of 4x MIC, the time-kill curve differed significantly from the control group (absence of Wp-CFS) (p < 0.05): the bacterial cell population remained stable at approximately106–107 CFU/mL over the 72 h incubation period, with no evidence of bacterial regrowth (Figure 2). No differences were observed in time kill curves at 0.25x, 0.5x, and MIC value when compared to the control group (p > 0.05). These findings corroborated the MIC/MBC results, and support the conclusion that the compound produced by W. paramesenteroides UFTM 2.6.1 has bacteriostatic activity against L. monocytogenes.
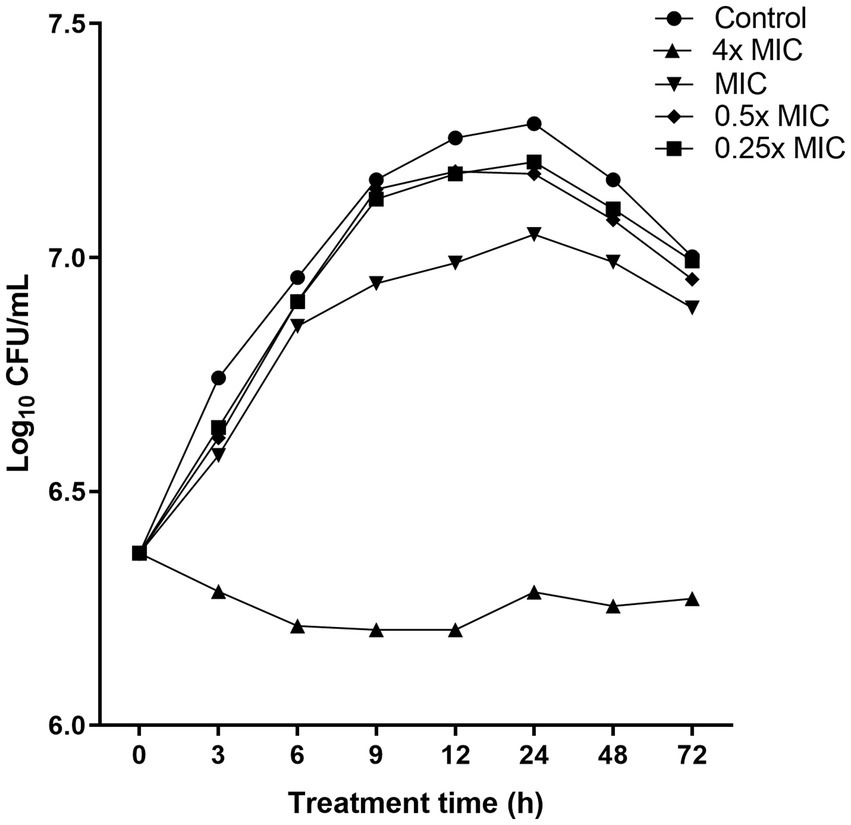
Figure 2. Time-kill curves showing the activity of Weissella paramesenteroides-cell-free supernatant (Wp-CFS; 4x MIC, MIC, 0.5x MIC, and 0.25x MIC) against Listeria monocytogenes strain 4455. Wp-CFS was added at time point 0 and monitored until 72 h. Control represents the curve obtained in absence of Wp-CFS.
3.2 Biofilm formation capacity by Listeria monocytogenes strains
Seventeen of the twenty-one strains of L. monocytogenes formed biofilm after incubation at 28°C for 72 h. The OD540 nm values obtained revealed the variability in biofilm biomass among the L. monocytogenes strains, with 14 strains (3833, 3837, 3839, 4001, 4107, 4251, 4313, 4330, 4449, 4484, 4511, 4455, 4506, and 4631) identified as strong biofilm formers, which were selected for further experiments (Figure 3).

Figure 3. Total biofilm biomass of Listeria monocytogenes strains on polystyrene microplates. Bars represent the mean and standard deviations of three independent experiments.
3.3 Weissella paramesenteroides-cell-free supernatant reduces biofilm formation and motility of Listeria monocytogenes strains
A significant reduction in the development of biofilms by L. monocytogenes strains was observed upon the addition of Wp-CFS (Table 3; Figure 4). Concentrations equal to or above the MIC significantly reduced biofilm formation (up to 100% reduction for several bacterial strains) when compared to the control (non-treated biofilms). At 0.5x MIC (15.6 mg/mL), biofilm formation inhibition rates ranged from 59.8 to 89.5%. A more prominent effect of Wp-CFS was observed against strains 3833, 4330, 4455, and 4631, for which a reduction in biofilm formation was observed even at 0.125x MIC (3.9 mg/mL) (Table 3).
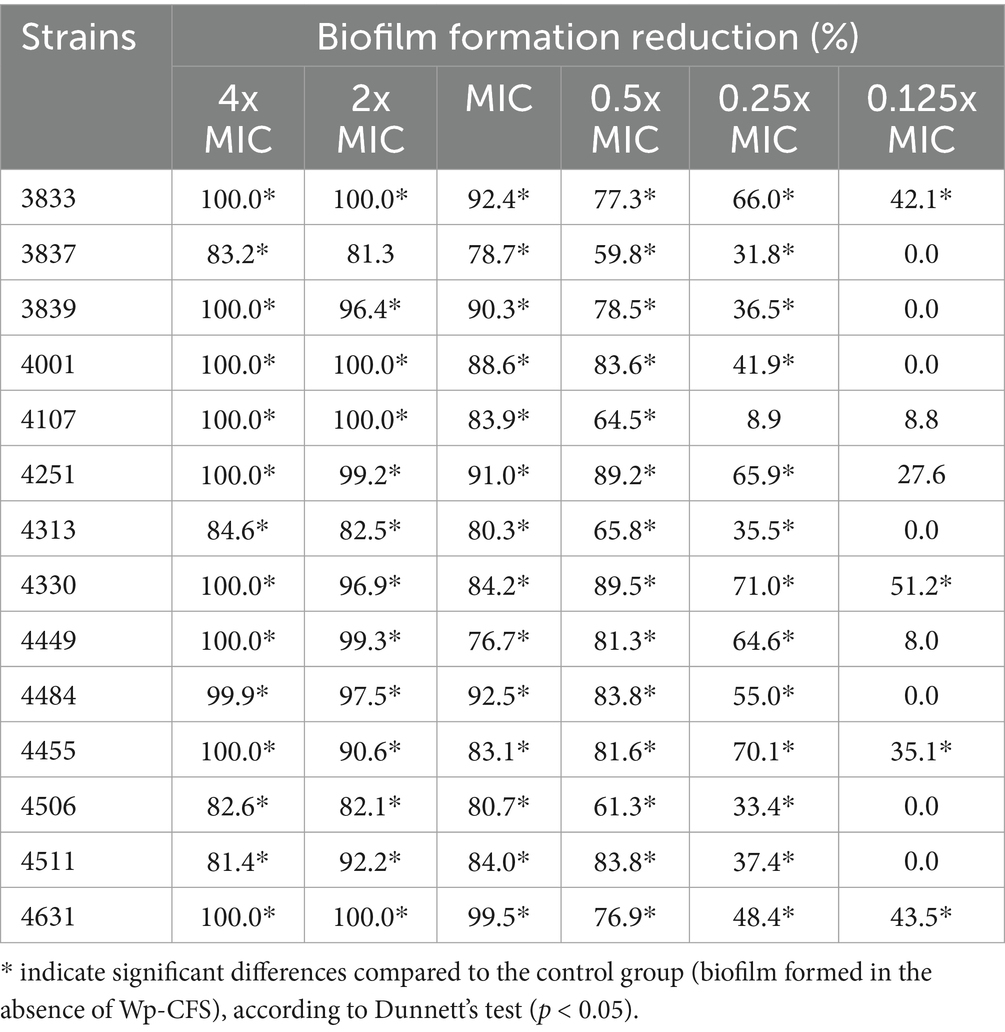
Table 3. Effect of Weissella paramesenteroides-cell-free supernatant (Wp-CFS; 0.125x to 4x MIC) in reducing biofilm formation by Listeria monocytogenes in vitro at 28°C for 72 h.
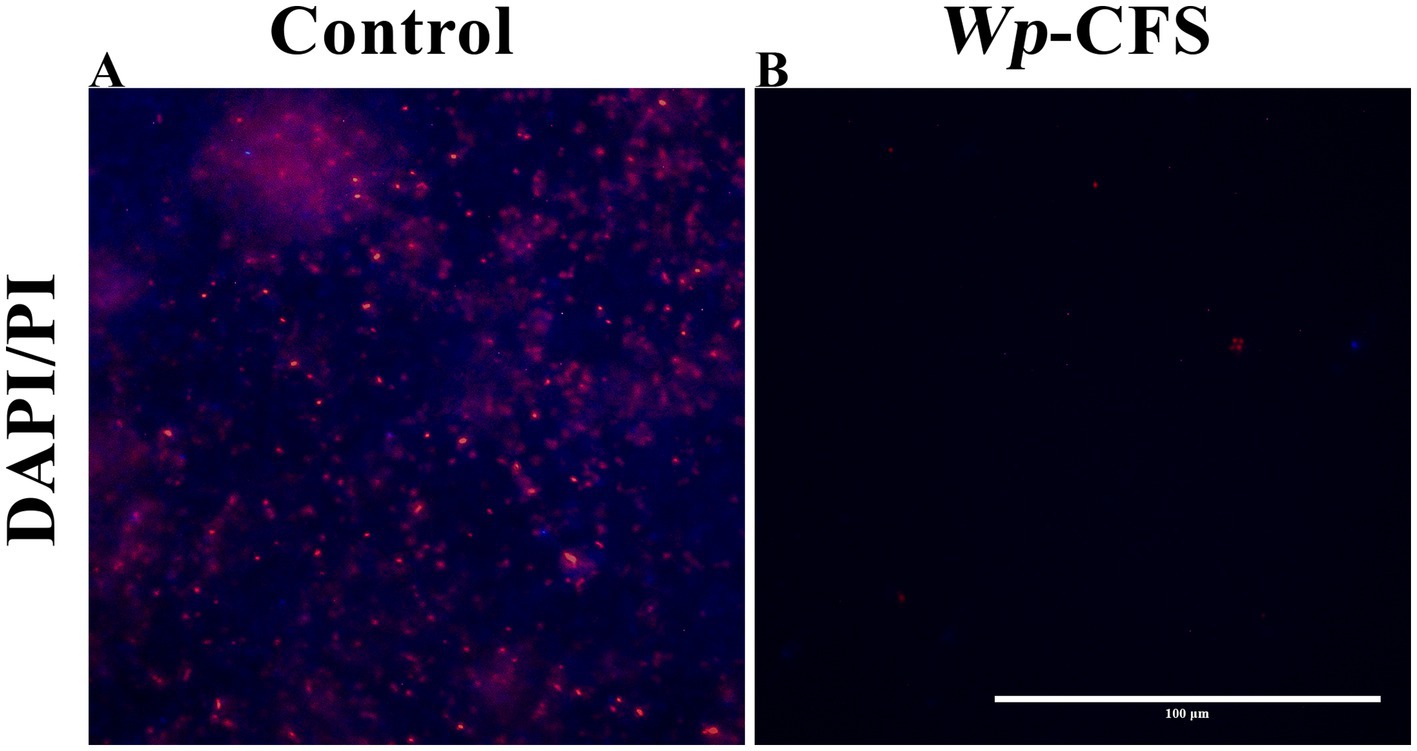
Figure 4. Fluorescence microscopy images of biofilm cells of Listeria monocytogenes strain 4455 on polystyrene microplates, either untreated (control; A) or exposed to Weissella paramesenteroides-cell-free supernatant (Wp-CFS; B). The biofilms were DAPI/PI stained (represented in blue and red on images, respectively). The images are representative of three biological replicates. The bar represents 100 μm.
In the presence of Wp-CFS at concentrations equal to or above the MIC value, a few remaining adhered L. monocytogenes cells were observed under fluorescence microscopy (Figure 4B), unlike the control, which presented a compact and mature biofilm (Figure 4A). This result confirms the anti-biofilm activity of Wp-CFS.
Swimming motility refers to individual movement in liquid powered by rotating flagella, while swarming motility is a multicellular surface movement driven by rotating helical flagella and regulated by quorum sensing. In the presence of 0.25x MIC (7.8 mg/mL) of Wp-CFS, the swarming motility of L. monocytogenes strains 3833, 3837, and 4455 showed a significant reduction compared to non-exposed cells (33.3, 45.0, and 33.3%, respectively) (Figures 5A,C). Swimming motility of L. monocytogenes was not affected by the presence of 0.25x MIC of Wp-CFS, except for strains 4330 and 4511 (30.0 and 55.0%, respectively) (Figures 5B,C). At 0.5x MIC of Wp-CFS, no bacterial growth was observed, while lower concentrations (0.125x MIC) did not affect the motility patterns of the L. monocytogenes strains evaluated (Figure 5C).
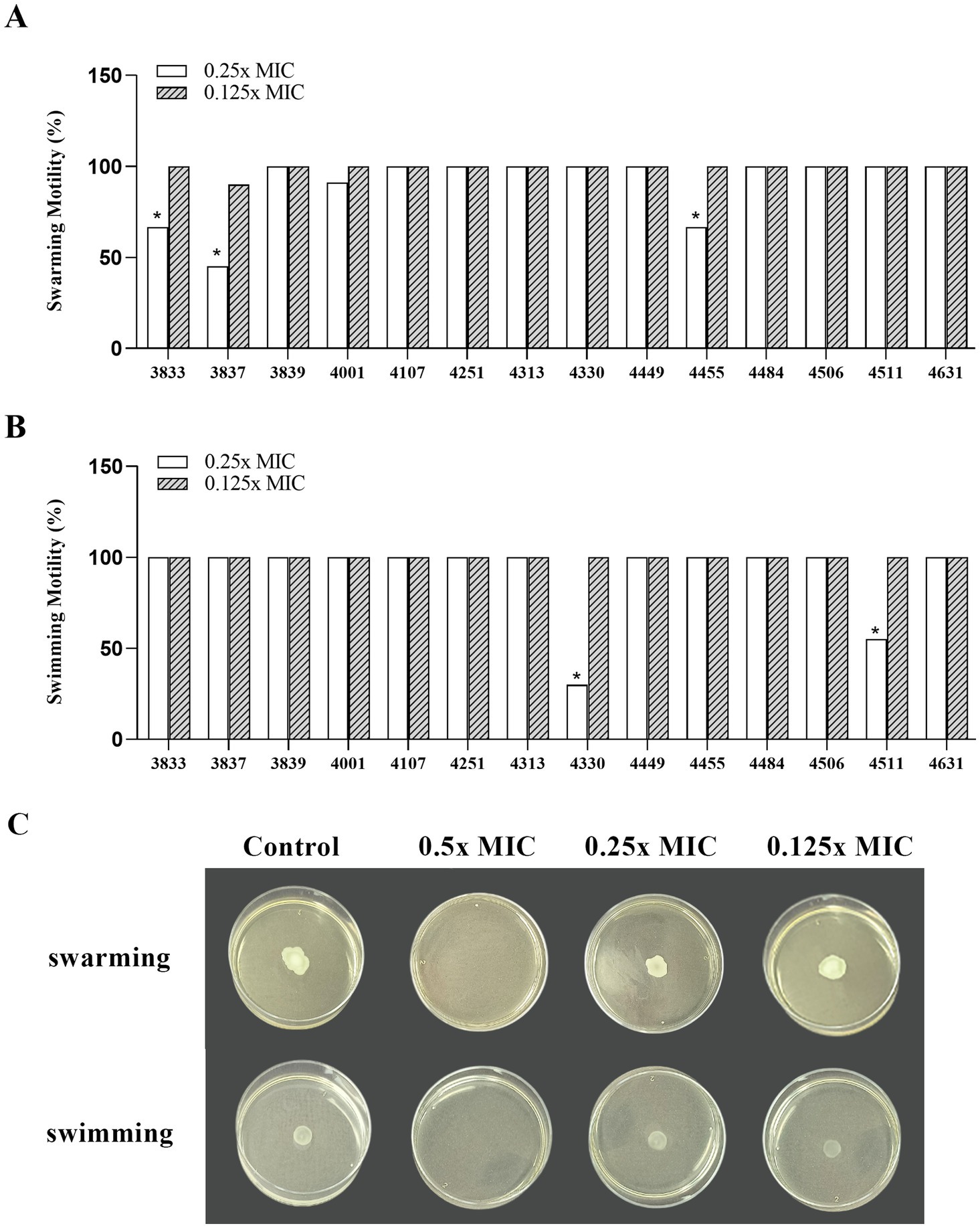
Figure 5. Efficacy of different concentrations of Weissella paramesenteroides-cell-free supernatant (Wp-CFS; 0.25x MIC and 0.125x MIC) in inhibiting motility ability of Listeria monocytogenes, at 28°C. (A) Swarming ability, (B) swimming ability, (C) representative images of the Wp-CFS activity on motility pattern of the Listeria monocytogenes strain 4455. Bars with an asterisk indicate significant differences compared to the control group (non-exposed to Wp-CFS), according to Dunnett’s test (p < 0.05).
3.4 Weissella paramesenteroides-cell-free supernatant modulates the expression of target genes in Listeria monocytogenes 4455
Considering the prominent effect of the Wp-CFS treatment on reducing biofilm formation and swarming motility of the L. monocytogenes strain 4455, this lineage was selected for transcriptional analysis of target genes related to QS, biofilm formation, and motility. Compared to untreated bacterial cells, the treatment with 0.25x MIC of Wp-CFS decreased the expression of the QS genes agrA and luxS. In addition, genes associated with biofilm formation, motility and flagella (degU, flaA, motA, and motB) were all significantly downregulated (p < 0.05) by Wp-CFS (Figure 6). On the other hand, the genes sigB, a global regulator of the stress response, and prfA, the main regulator of virulence factors in L. monocytogenes were upregulated (p < 0.05) in the presence of Wp-CFS (Figure 6).
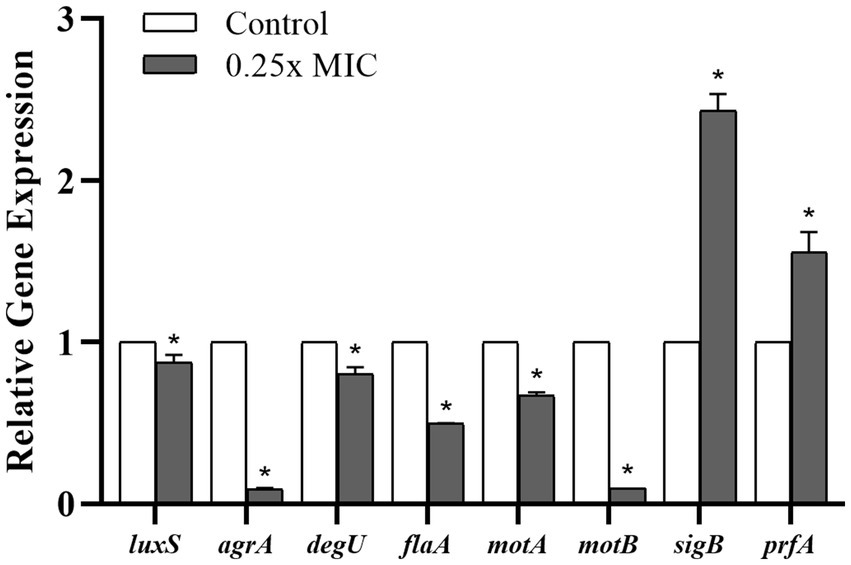
Figure 6. Relative gene expression of Listeria monocytogenes strain 4455 in response to exposure to 0.25x MIC of Weissella paramesenteroides-cell-free supernatant (Wp-CFS), at 28°C for 24 h. *Indicates statistical differences when compared to the control group (non-exposed to Wp-CFS) (p < 0.05). Mean values of three independent experiments are shown.
4 Discussion
Bacteria of the genus Weissella are recognized to produce bioactive compounds with different mechanisms of action. Notable bioactive compounds produced by Weissella species include organic acids, hydrogen peroxide, diacetyl, bacteriocins, exopolysaccharides, antioxidants, vitamins, and short-chain fatty acids (Singh et al., 2024; Deatraksa et al., 2018; Yu et al., 2019; Kavitake et al., 2020; Månberger et al., 2020; Zhao et al., 2021; Fhoula et al., 2022; Hernández-Alcántara et al., 2022; Tuccillo et al., 2022; Lahmar et al., 2024). The diversity of these bioactive metabolites makes Weissella a promising candidate for applications in food preservation, the probiotic industry, and the development of new natural products with antimicrobial and therapeutic properties. Despite the well-recognized antimicrobial properties and beneficial effects in food fermentation, research into the specific quorum-quenching abilities of Weissella species is still underexplored. Most studies on this topic remain focused on other LAB genera, such as Lactobacillus and Enterococcus, which have been screened for the production of QQ molecules that interfere with QS systems of several pathogens (Chatterjee et al., 2017; Boopathi et al., 2017; Ham et al., 2018; Cui et al., 2020; Vadassery and Pillai, 2020; Qiao et al., 2021; Lv et al., 2021; Vasiee et al., 2022; Tomé et al., 2023; Shi et al., 2024).
Some studies have shown that Lactobacillus species produce molecules that reduce L. monocytogenes virulence, motility, biofilm formation, and/or can interfere with QS signaling (Moradi et al., 2019; Singh et al., 2020; Jara et al., 2020; Hossain et al., 2021; Kıran et al., 2021; Lee et al., 2021; Liu et al., 2022; Davares et al., 2022). In the present study, we demonstrate for the first time that W. paramesenteroides UFTM 2.6.1 may also produce QQ compounds, which have the potential to prevent biofilm formation, in addition to reducing motility and downregulating virulence gene expression in L. monocytogenes.
To investigate the effect of Wp-CFS on L. monocytogenes, 21 strains isolated from food and food processing sectors with four different serotypes were selected for this study. After confirming the antagonistic activity of W. paramesenteroides UFTM 2.6.1 on L. monocytogenes strains, both in solid and liquid media, and selection of the best biofilm-producing strains, we demonstrated the bacteriostatic effect of Wp-CFS through time-kill experiments. Those results substantiated the hypothesis that we were addressing a potential QQ compound produced by W. paramesenteroides UFTM 2.6.1, since the premise of a QQ molecule is to attenuate bacterial virulence without directly killing the target bacteria. It is important to highlight that QS assays should be performed in concentrations that do not interfere with bacterial growth, to avoid the interference of cell density differences in QS regulated phenotypes (Defoirdt et al., 2013; Santos et al., 2021).
The ability to form biofilm is an important feature associated with persistence and antimicrobial resistance among bacterial pathogens (Costerton et al., 1999; Donlan and Costerton, 2002; de la Fuente-Núñez et al., 2013a). L. monocytogenes biofilms can serve as a source of food contamination through the release of cells attached to their structure (dispersal phase), enabling the continuous contamination of food products in the production chain (Poimenidou et al., 2009; Miquel et al., 2016). Different compounds extracted from plants or microbial cultures have shown anti-biofilm effects against L. monocytogenes (Riedel et al., 2009; Zamani et al., 2017; Li et al., 2025; Rocha et al., 2019).
Even at lower concentrations, below the MIC value, Wp-CFS consistently reduced biofilm formation and motility, two well-known QS regulated phenotypes in L. monocytogenes. It should be noted that at concentrations higher than the MIC, the reduction in biofilm formation did not significantly differ from the reduction obtained at MIC. Moreover, an inhibition of L. monocytogenes growth was observed in motility assays when Wp-CFS was added at MIC value, which likely affected motility through a QS-independent mechanism.
According to the results obtained, it is likely that the anti-biofilm property of Wp-CFS could be due to reduced L. monocytogenes adhesion capacity in the early stages of biofilm development or preventing the switch to the biofilm phenotype. Our findings can be substantiated with a study conducted by Jamwal et al. (2019), which reported maximum biofilm inhibitory activity (>80%) of probiotic strains at initial stages of biofilm formation, whereas weaker activity was observed after prolonged incubation (48 h). Other studies (Gálvez et al., 2007; Winkelströter et al., 2014; Zhou et al., 2020; de la Fuente-Nunez et al., 2013b; Pimentel-Filho et al., 2014; Segev-Zarko et al., 2015; Ahn et al., 2018) reported that LAB supernatants or compounds produced by LAB, such as bacteriocins and exopolysaccharides, can interfere with the early events of biofilm formation by preventing bacterial cells adhesion to surfaces or to each other cells or by killing cells before they stably become part of the biofilm architecture.
To enlighten possible mechanisms underlying the anti-Listeria activity of WP-CFS, we verified the gene expression of several target genes in L. monocytogenes. Our results showed that the exposure to a sub-inhibitory concentration of Wp-CFS (0.25x MIC) reduced the expression of both luxS and agrA, which, consequently, could impair QS communication in L. monocytogenes. Disruption of a single component within the QS pathway frequently results in the downregulation of QS related genes and subsequent inactivation of the QS mechanism (Galié et al., 2018).
LuxS is the enzyme responsible for the biosynthesis of the signal molecule AI-2. LuxS/AI-2 is an important QS system, present in both Gram-negative and Gram-positive bacteria, and has therefore been proposed as a universal signal enabling interspecies cell–cell communication. In L. monocytogenes, intraspecies communication (Skandamis and Nychas, 2012; Kocot and Olszewska, 2017) occurs via the Agr system, which consists of the four-gene operon agrBDCA. AgrD, the precursor peptide, is processed into an active signaling molecule called autoinducing peptide (AIP) by AgrB, and the AIP is released outside the cells; once the AIP reaches critical concentration, it activates the two-component system AgrC/AgrA (receptor-histidine kinase/response regulator) (Lina et al., 1998).
In some pathogenic bacteria, LuxS was found to be involved in biofilm formation (Sela et al., 2006). However, the contribution of the luxS gene to biofilm formation in L. monocytogenes remains controversial. Belval et al. (2006) found that the luxS mutant gene in L. monocytogenes resulted in an increase in biofilm formation at 25°C. On the other hand, Bonsaglia et al. (2014) noted that L. monocytogenes strains with the luxS gene did not consistently produce biofilms. Zhang et al. (2022) found that the amount of biofilm formation was reduced in the luxS gene deletion strain of L. monocytogenes. In a study conducted by Gao et al. (2024), higher mRNA levels of agrA, agrB, agrC, and luxS in L. monocytogenes strains were not directly correlated with high biofilm capacity in vitro. Those conflicting findings regarding the role of luxS in biofilm formation by L. monocytogenes can be explained, at least in part, by strain-specific responses or experimental differences.
According to Yang et al. (2024), the expression level of the agrA gene in L. monocytogenes strains with strong biofilm-forming capacity is up-regulated after biofilm formation, unlike poor biofilm-forming strains, in which no differential expression is generally observed. This finding demonstrated that agrA gene plays a positive regulatory role in the biofilm formation process. In this sense, the down-regulation of agrA may reduce the development of L. monocytogenes biofilms (Rieu et al., 2007; Riedel et al., 2009), which was clearly demonstrated in the presence of Wp-CFS.
Flagella are recognized as key factors in facilitating the initial contact of the bacterial cell with surfaces in the early stages of biofilm formation (Haiko and Westerlund-Wikström, 2013; Fan et al., 2020; Benyoussef et al., 2022; Coloma-Rivero et al., 2022; Vilas Boas et al., 2024). It has been suggested that flagellum-mediated motility may assist in overcoming repulsive forces at the surface, thereby enabling initial attachment (Benyoussef et al., 2022). Swarming motility is a specialized form of motility on solid surfaces, dependent on extensive flagellation, cell–cell contact, and driven by QS, while swimming motility refers to the movement of individual bacteria, which involves the rotation of flagella to propel the cell through liquid environments.
The presence of sub-inhibitory concentrations of Wp-CFS reduced not only swarming motility but also the expression levels of the motility-associated genes degU, flaA, motA and motB, which additionally provides insights into the mechanism of action of Wp-CFS in disrupting QS-system and reducing in vitro biofilm formation by L. monocytogenes. The degU gene encodes a response regulator in L. monocytogenes that regulates the expression of flagellin and motility genes (Williams et al., 2005; Zhu et al., 2023). The flaA gene encodes flagellin A (FlaA), the structural subunit of the flagellum, while the motA and motB genes encode the flagellar motor proteins MotA and MotB, respectively, which control flagellar movement (Gründling et al., 2004; Casey et al., 2014).
The transcriptional activator PrfA (positive regulatory factor A) has been identified in L. monocytogenes, and it plays a crucial role in the regulation of most genes associated with L. monocytogenes pathogenesis, including the virulence island 1 (LIPI-1) genes (Gaballa et al., 2019; de las Heras et al., 2011). Sigma factor B (sigB) is the global regulator of the stress response, and it has been reported to be involved in biofilm formation and in the adaptation process of L. monocytogenes strains to low-temperature environments (Liu et al., 2021; Hu et al., 2007; Zhou et al., 2020; Vázquez- Armenta et al., 2020). The transcription of both the prfA and sigB genes was upregulated in L. monocytogenes in the presence of 0.25x MIC of Wp-CFS. This result may indicate an attempt by the bacterial cell to activate pathogenesis- and stress-related genes in order to resist and survive the external stimulus caused by the presence of Wp-CFS.
A similar result was observed in a study conducted by Huang et al. (2020), in which the photodynamic inactivation treatment upregulated the expression of the prfA while significantly reducing the adhesion ability of L. monocytogenes biofilms. In contrast, Masebe and Thantsha (2022) found that the expression of the prfA gene was significantly reduced in the presence of CFS of LAB strains, with a consequent inhibition and/or dispersion of L. monocytogenes biofilms. In a study conducted by Liu et al. (2021), the natural compounds cinnamaldehyde and eugenol down-regulated the transcription of the prfA and sigB, a result associated with the inhibition of L. monocytogenes biofilm formation.
The downregulation of key regulatory (luxS, agrA, degU) and motility-associated genes (flaA, motA, motB) in the presence of Wp-CFS, alongside its ability to significantly reduce Listeria monocytogenes biofilm formation in vitro, suggests that Wp-CFS interferes with upstream regulatory systems involved in QS and environmental signal processing. A plausible hypothesis for quorum-quenching activity of Wp-CFS is that it disrupts QS-mediated regulatory cascades either by reducing QS signal molecules (e.g., AI-2) or by antagonizing signal perception. The repression of luxS, a gene central to AI-2 biosynthesis, and agrA, a critical component of the Agr QS system, implies interference at both signal production and response levels. This disruption likely affects intermediate regulators such as degU, which ultimately downregulating downstream targets involved in motility and biofilm formation. Since motility is often a prerequisite for initial surface colonization and biofilm development, the coordinated repression of motility and QS genes by Wp-CFS provides a plausible mechanistic explanation for its antibiofilm activity. Together, these results support a model in which the QQ activity of Wp-CFS impairs QS-regulated phenotypes in L. monocytogenes, offering a promising strategy for attenuating virulence and persistence of this pathogen.
5 Conclusion
This study presents a novel contribution to the field by demonstrating the previously unreported potential of Weissella paramesenteroides in disrupting the QS mechanism of the foodborne pathogen Listeria monocytogenes. This discovery broadens the functional scope of W. paramesenteroides UFTM 2.6.1 as a beneficial microorganism, highlighting its promise as a biocontrol agent in food-related and biomedical sector. The identification of compounds with anti-biofilm and anti-virulence properties supports its potential application.
Further investigations are underway to identify the active constituents of Wp-CFS, to characterize the molecular mechanisms underlying the QQ activity. Additionally, future studies should focus on validating the effects of Wp-CFS in complex environments and in animal models. These insights will be pivotal for advancing its practical application in food bioprotection, to ensure food safety.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found at: https://www.ebi.ac.uk/ena/browser/view/SAMEA115664826.
Author contributions
IS: Data curation, Investigation, Writing – original draft, Formal analysis. SR: Writing – original draft, Investigation, Formal analysis. HP: Writing – review & editing, Investigation, Formal analysis, Data curation, Methodology. MS: Methodology, Writing – review & editing, Investigation. FA: Methodology, Writing – review & editing, Formal analysis, Conceptualization, Validation. UP: Writing – review & editing, Validation, Methodology, Conceptualization. AF: Formal analysis, Supervision, Writing – review & editing, Conceptualization. AP: Data curation, Writing – original draft, Funding acquisition, Conceptualization, Supervision, Formal analysis, Project administration, Writing – review & editing, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work has been supported by Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, Brazil -APQ 01597-22). ISR and SDCR were FAPEMIG scholarship folders.
Acknowledgments
The authors thank FAPEMIG for financial support and the Programa de Pós-Graduação em Biociências (UFTM, Brazil).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahn, K. B., Baik, J. E., Yun, C. H., and Han, S. H. (2018). Lipoteichoic acid inhibits Staphylococcus aureus biofilm formation. Front. Microbiol. 9:327. doi: 10.3389/fmicb.2018.00327
Almeida, F. A., Freitas, L. L., Carneiro, D. G., and Vanetti, M. C. D. (2023). “Bacterial cell-to-cell communication and its relevance to food safety” in Present knowledge in food safety: A risk-based approach through the food chain. eds. M. E. Knowles, L. E. Anelich, A. R. Boobis, and B. Popping, vol. 1 (The Netherlands: Elsevier), 829–845.
Belval, S. C., Gal, L., Margiewes, S., Garmyn, D., Piveteau, P., and Guzzo, J. (2006). Assessment of the roles of LuxS, S-ribosyl homocysteine, and autoinducer 2 in cell attachment during biofilm formation by Listeria monocytogenes EGD-e. Appl. Environ. Microbiol. 72, 2644–2650. doi: 10.1128/AEM.72.4.2644-2650.2006
Benyoussef, W., Deforet, M., Monmeyran, A., and Henry, N. (2022). Flagellar motility during E. coli biofilm formation provides a competitive disadvantage which recedes in the presence of co-colonizers. Front. Cell. Infect. Microbiol. 12:896898. doi: 10.3389/fcimb.2022.896898
Bonsaglia, E. C. R., Silva, N. C. C., Fernandes, J., Júniora, J. P. A., Tsunemib, M. H., and Ralla, V. L. M. (2014). Production of biofilm by Listeria monocytogenes in different materials and temperatures. Food Control 35, 386–391. doi: 10.1016/j.foodcont.2013.07.023
Boopathi, S., Selvakumar, N., and Sivakumar, G. (2017). Quorum quenching potentials of probiotic Enterococcus durans Lab38 against methicillin resistant Staphylococcus aureus. Asian J. Pharm. Clin. Res. 10, 445–450. doi: 10.22159/ajpcr.2017.v10i4.17039
Booth, S. J., Johnson, J. L., and Wilkins, T. D. (1977). Bacteriocin production by strains of Bacteroides isolated from human feces and the role of these strains in the bacterial ecology of the colon. Antimicrob. Agents Chemother. 11, 718–724. doi: 10.1128/AAC.11.4.718
Brackman, G., and Coenye, T. (2015). Quorum sensing inhibitors as anti-biofilm agents. Curr. Pharm. Des. 21, 5–11. doi: 10.2174/1381612820666140905114627
Casey, A., Fox, E. M., Schmitz-Esser, S., Coffey, A., McAuliffe, O., and Jordan, K. (2014). Transcriptome analysis of Listeria monocytogenes exposed to biocide stress reveals a multi-system response involving cell wall synthesis, sugar uptake, and motility. Front Microbiol. 68, 1–10. doi: 10.3389/fmicb.2014.00068
Chatterjee, M., D’Morris, S., Paul, V., Warrier, S., Vasudevan, A. K., Vanuopadath, M., et al. (2017). Mechanistic understanding of phenyllactic acid mediated inhibition of quorum sensing and biofilm development in Pseudomonas aeruginosa. Appl. Microbiol. Biotechnol. 101, 8223–8236. doi: 10.1007/s00253-017-8546-4
Cheng, Y., Mousavi, Z. E., Pennone, V., Hurley, D., and Butler, F. (2023). Association between the presence of resistance genes and sanitiser resistance of Listeria monocytogenes isolates recovered from different food-processing facilities. Microorganisms 11:2989. doi: 10.3390/microorganisms11122989
CLSI (2023). Performance standards for antimicrobial susceptibility testing. 33rd Edn. CLSI supplement M100. Wayne, PA: CLSI.
Collins, M. D., Samelis, J., Metaxopoulos, J., and Wallbanks, S. (1993). Taxonomic studies on some Leuconostoc-like organisms from fermented sausages: description of a new genus Weissella for the Leuconostoc paramesenteroides group of species. J. Appl. Bacteriol. 75, 595–603. doi: 10.1111/j.1365-2672.1993.tb01600.x
Coloma-Rivero, R. F., Flores-Concha, M., Molina, R. E., Soto-Shara, R., Cartes, Á., and Oñate, Á. A. (2022). Brucella and its hidden flagellar system. Microorganisms 10:83. doi: 10.3390/microorganisms10010083
Costerton, J. W., Stewart, P. S., and Greenberg, E. P. (1999). Bacterial biofilms: a common cause of persistent infections. Science 284, 1318–1322. doi: 10.1126/science.284.5418.1318
Cui, T., Bai, F., Sun, M., Lv, X., Li, X., Zhang, D., et al. (2020). Lactobacillus crustorum ZHG 2-1 as novel quorum-quenching bacteria reducing virulence factors and biofilms formation of Pseudomonas aeruginosa. LWT 117:108696. doi: 10.1016/j.lwt.2019.108696
Davares, A. K. L., Arsene, M. M. J., Viktorovna, P. I., Vyacheslavovna, Y. N., Vladimirovna, Z. A., Aleksandrovna, V. E., et al. (2022). Quorum-sensing inhibitors from probiotics as a strategy to combat bacterial cell-to-cell communication involved in food spoilage and food safety. Fermentation 8:711. doi: 10.3390/fermentation8120711
de la Fuente-Núñez, C., Reffuveille, F., Fairfull-Smith, K. E., and Hancock, R. E. W. (2013a). Effect of nitroxides on swarming motility and biofilm formation, multicellular behaviors in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 57, 4877–4881. doi: 10.1128/AAC.01381-13
de la Fuente-Nunez, C., Reffuveille, F., Fernández, L., and Hancock, R. E. W. (2013b). Bacterial biofilm development as a multicellular adaptation: antibiotic resistance and new therapeutic strategies. Curr Opin Microbiol. 16, 580–589. doi: 10.1016/j.mib.2013.06.013
de las Heras, A., Cain, R. J., Bielecka, M. K., and Vázquez-Boland, J. A. (2011). Regulation of Listeria virulence: PrfA master and commander. Curr. Opin. Microbiol. 14, 118–127. doi: 10.1016/j.mib.2011.01.005
Deatraksa, J., Sunthornthummas, S., Rangsiruji, A., Sarawaneeyaruk, S., Suwannasai, N., and Pringsulaka, O. (2018). Isolation of folate-producing Weissella spp. from Thai fermented fish (Plaa Som fug). LWT 89, 388–391. doi: 10.1016/j.lwt.2017.11.016
Defoirdt, T., Brackman, G., and Coenye, T. (2013). Quorum sensing inhibitors: how strong is the evidence? Trends Microbiol. 21, 619–624. doi: 10.1016/j.tim.2013.09.006
Donlan, R. M., and Costerton, J. W. (2002). Biofilms: survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 15, 167–193. doi: 10.1128/CMR.15.2.167-193.2002
Fan, Y., Qiao, J., Lu, Z., Fen, Z., Tao, Y., Lv, F., et al. (2020). Influence of different factors on biofilm formation of Listeria monocytogenes and the regulation of cheY gene. Food Res. Int. 137:109405. doi: 10.1016/j.foodres.2020.109405
Fan, Q., Zhang, Y., Yang, H., Wu, Q., Shi, C., Zhang, C., et al. (2018). Effect of coenzyme Q0 on biofilm formation and attachment-invasion efficiency of Listeria monocytogenes. Food Control 90, 274–281. doi: 10.1016/j.foodcont.2018.02.047
Fhoula, I., Boumaiza, M., Tayh, G., Rehaiem, A., Klibi, N., and Ouzari, I. H. (2022). Antimicrobial activity and safety features assessment of Weissella spp. from environmental sources. Food Sci. Nutr. 10, 2896–2910. doi: 10.1002/fsn3.2885
Fhoula, I., Najjari, A., Turki, Y., Jaballah, S., Boudabous, A., and Ouzari, H.-I. (2013). Diversity and antimicrobial properties of lactic acid bacteria isolated from rhizosphere of olive trees and desert truffles of Tunisia. Microorganisms. 10:405708. doi: 10.1155/2013/405708
Fusco, V., Chieffi, D., Fanelli, F., Montemurro, M., Rizzello, C. G., and Franz, C. M. A. P. (2023). The Weissella and Periweissella genera: up-to-date taxonomy, ecology, safety, biotechnological, and probiotic potential. Front. Microbiol. 14:1289937. doi: 10.3389/fmicb.2023.1289937
Gaballa, A., Guariglia-Oropeza, V., Wiedmann, M., and Boor, K. J. (2019). Cross Talk between SigB and PrfA in Listeria monocytogenes Facilitates Transitions between Extra- and Intracellular Environments. Microbiol Mol Biol Rev. 83:1128. doi: 10.1128/mmbr.00034-19
Galié, S., García-Gutiérrez, C., Miguélez, E. M., Villar, C. J., and Lombó, F. (2018). Biofilms in the Food Industry: Health Aspects and Control Methods. Front Microbiol. 9:898. doi: 10.3389/fmicb.2018.00898
Gálvez, A., Abriouel, H., and Lópes, L. R. (2007). Bacteriocin-based strategies for food biopreservation. Int J Food Microbiol. 120, 51–70. doi: 10.1016/j.ijfoodmicro.2007.06.001
Gao, B. R., Cai, H., Xu, B., Yang, F., Dou, X., Dong, Q., et al. (2024). Growth, biofilm formation, and motility of Listeria monocytogenes strains isolated from food and clinical samples located in Shanghai (China). Food Res. Int. 184:114232. doi: 10.1016/j.foodres.2024.114232
Gemmell, C. T., Parreira, V. R., and Farber, J. M. (2022). Controlling Listeria monocytogenes growth and biofilm formation using flavonoids. J. Food Prot. 85, 639–646. doi: 10.4315/JFP-21-135
Gründling, A., Burrack, L. S., Bouwer, H. G., and Higgins, D. E. (2004). Listeria monocytogenes regulates flagellar motility gene expression through MogR, a transcriptional repressor required for virulence. Proc. Natl. Acad. Sci. USA 101, 12318–12323. doi: 10.1073/pnas.0404924101
Guilbaud, M., Piveteau, P., Desvaux, M., and Brisse, S. (2015). Briandet R Exploring the Diversity of Listeria monocytogenes Biofilm Architecture by High-Throughput Confocal Laser Scanning Microscopy and the Predominance of the Honeycomb-Like Morphotype. Appl Environ Microbiol. 81, 1813–1819. doi: 10.1128/AEM.03173-14
Haiko, J., and Westerlund-Wikström, B. (2013). The role of the bacterial flagellum in adhesion and virulence. Biology 2, 1242–1267. doi: 10.3390/biology2041242
Ham, S. Y., Kim, H. S., Cha, E., Park, J. H., and Park, H. D. (2018). Mitigation of membrane biofouling by a quorum quenching bacterium for membrane bioreactors. Bioresour. Technol. 258, 220–226. doi: 10.1016/j.biortech.2018.03.007
Hernández-Alcántara, A. M., Chiva, R., Mohedano, M. L., Russo, P., Ruiz-Masó, J. A., del Solar, G., et al. (2022). Weissella cibaria riboflavin-overproducing and dextran-producing strains useful for the development of functional bread. Front. Nutr. 9:978831. doi: 10.3389/fnut.2022.978831
Hossain, M. I., Mizan, M. F. R., Roy, P. K., Nahar, S., Toushik, S. H., Ashrafudoulla, M., et al. (2021). Listeria monocytogenes biofilm inhibition on food contact surfaces by application of postbiotics from Lactobacillus curvatus B.67 and Lactobacillus plantarum M.2. Food Res. Int. 148:110595. doi: 10.1016/j.foodres.2021.110595
Hu, Y., Oliver, H. F., Raengpradub, S., Palmer, M. E., Orsi, R. H., Wiedmann, M., et al. (2007). Transcriptomic and phenotypic analyses suggest a network between the transcriptional regulators HrcA and σB in Listeria monocytogenes. Appl. Environ. Microbiol. 73, 7981–7991. doi: 10.1128/aem.01281-07
Huang, J., Chen, B., Li, H., Zeng, Q. H., Wang, J. J., Liu, H., et al. (2020). Enhanced antibacterial and antibiofilm functions of the curcumin-mediated photodynamic inactivation against Listeria monocytogenes. Food Control 108:106886. doi: 10.1016/j.foodcont.2019.106886
Jamwal, A., Sharma, K., Chauhan, R., Bansal, S., and Goel, G. (2019). Evaluation of commercial probiotic lactic cultures against biofilm formation by Cronobacter sakazakii. Intest Res. 17, 192–201. doi: 10.5217/ir.2018.00106
Jara, J., Pérez-Ramos, A., del Solar, G., Rodríguez, J. M., Fernández, L., and Orgaz, B. (2020). Role of Lactobacillus biofilms in Listeria monocytogenes adhesion to glass surfaces. Int. J. Food Microbiol. 334:108804. doi: 10.1016/j.ijfoodmicro.2020.108804
Jiang, X., Ren, S., G, Y., Jiang, C., Liu, G., Wang, H., et al. (2021). Role of the VirSR-VirAB system in biofilm formation of Listeria monocytogenes EGD-e. Food Res. Int. 145:110394. doi: 10.1016/j.foodres.2021.110394
Kavitake, D., Bruntha Devi, P., and Shetty, P. H. (2020). Overview of exopolysaccharides produced by Weissella genus – a review. Int. J. Biol. Macromol. 164, 2964–2973. doi: 10.1016/j.ijbiomac.2020.08.185
Kıran, F., Akoğlu, A., and Çakır, İ. (2021). Control of Listeria monocytogenes biofilm on industrial surfaces by cell-free extracts of Lactobacillus plantarum. J. Food Process. Preserv. 45:e15042. doi: 10.1111/jfpp.15042
Kocot, A. M., and Olszewska, M. A. (2017). Biofilm formation and microscopic analysis of biofilms formed by Listeria monocytogenes in a food processing context. LWT-Food Sci. Techonol. 84:47–57. doi: 10.1016/j.lwt.2017.05.042
Lahmar, M., Besrour-Aouam, N., Hernández-Alcántara, A. M., Diez-Ozaeta, I., Fhoula, I., López, P., et al. (2024). Riboflavin and dextran-producing Weissella confusa FS54 B2: characterization and testing for development of fermented plant-based beverages. Food Secur. 13:4112. doi: 10.3390/foods13244112
Lee, J. E., Lee, N. K., and Paik, H. D. (2021). Antimicrobial and anti-biofilm effects of probiotic Lactobacillus plantarum KU200656 isolated from kimchi. Food Sci. Biotechnol. 30, 97–106. doi: 10.1007/s10068-020-00837-0
Li, P., Chen, X., Aziz, T., Shami, A., Al-Asmari, F., Al-Joufi, F. A., et al. (2025). The anti-biofilm and anti-virulence mechanisms of carvacrol against Listeria monocytogenes and the application in food systems. Food Bioscience 69:106950. doi: 10.1016/j.fbio.2025.106950
Li, J., Fan, Q., Jin, M., Mao, C., Zhang, H., and Zhang, X. (2021). Paeoniflorin reduces luxS/AI-2 system-controlled biofilm formation and virulence in Streptococcus suis. Virulence 12, 3062–3073. doi: 10.1080/21505594.2021.2010398
Lina, G., Jarraud, S., Ji, G., Greenland, T., Pedraza, A., Etienne, J., et al. (1998). Transmembrane topology and histidine protein kinase activity of AgrC, the agr signal receptor in Staphylococcus aureus. Mol. Microbiol. 28, 655–662. doi: 10.1046/j.1365-2958.1998.00830.x
Liu, C., An, C., Zhang, J., Liu, Y., Zhang, Q., Ding, H., et al. (2024). Evaluation of safety and probiotic properties of Weissella spp. in fermented vegetables from Xi’an, Shaanxi, China. Food Sci. Nutr. 13:e4592. doi: 10.1002/fsn3.4592
Liu, Y., Bu, Y., Li, J., Liu, Y., Liu, A., Gong, P., et al. (2022). Inhibition activity of Plantaricin Q7 produced by Lactobacillus plantarum Q7 against Listeria monocytogenes and its biofilm. Fermentation 8:75. doi: 10.3390/fermentation8020075
Liu, Y., Wu, L., Han, J., Dong, P., Luo, X., Zhang, Y., et al. (2021). Inhibition of biofilm formation and related gene expression of Listeria monocytogenes in response to four natural antimicrobial compounds and sodium hypochlorite. Front. Microbiol. 11:617473.
Livak, K. J., and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Lv, X., Cui, T., Du, H., Sun, M., Bai, F., Li, J., et al. (2021). Lactobacillus plantarum CY 1-1: a novel quorum quenching bacteria and anti-biofilm agent against Aeromonas sobria. LWT 137:110439. doi: 10.1016/j.lwt.2020.110439
Månberger, A., Verbrugghe, P., Guðmundsdóttir, E. E., Santesson, S., Nilsson, A., Hreggviðsson, G. Ó., et al. (2020). Taxogenomic assessment and genomic characterisation of Weissella cibaria strain 92 able to metabolise oligosaccharides derived from dietary fibres. Sci. Rep. 10:5853. doi: 10.1038/s41598-020-62610-x
Marques, P. H., Jaiswal, A. K., de Almeida, F. A., Pinto, U. M., Ferreira-Machado, A. B., Tiwari, S., et al. (2024). Lactic acid bacteria secreted proteins as potential Listeria monocytogenes quorum sensing inhibitors. Mol. Divers. 28, 2897–2912. doi: 10.1007/s11030-023-10722-7
Masebe, R. D., and Thantsha, M. S. (2022). Anti-biofilm activity of cell-free supernatants of selected lactic acid bacteria against Listeria monocytogenes isolated from avocado and cucumber fruits, and from an avocado processing plant. Food Secur. 11:2872. doi: 10.3390/foods11182872
Miquel, S., Martin, R., Rossi, O., Bermudez-Humaran, L. G., Chatel, J. M., Sokol, H., et al. (2016). Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 16, 255–261. doi: 10.1016/j.mib.2013.06.003
Moradi, M., Mardani, K., and Tajik, H. (2019). Characterization and application of postbiotics of Lactobacillus spp. on Listeria monocytogenes in vitro and in food models. LWT 111, 457–464. doi: 10.1016/j.lwt.2019.05.072
Petersen, P. J., Jones, C. H., and Bradford, P. A. (2007). In vitro antibacterial activities of tigecycline and comparative agents by time-kill kinetic studies in fresh Mueller-Hinton broth. Diagn. Microbiol. 59, 347–349. doi: 10.1016/j.diagmicrobio.2007.05.013
Pieta, L., Escudero, F. L. G., Jacobus, A. P., Cheiran, K. P., Gross, J., Moya, M. L. E., et al. (2017). Comparative transcriptomic analysis of Listeria monocytogenes reveals upregulation of stress genes and downregulation of virulence genes in response to essential oil extracted from Baccharis psiadioides. Ann. Microbiol. 67, 479–490. doi: 10.1007/s13213-017-1277-z
Pimentel-Filho, N. J., Montovani, H. C., Carvalho, A. F., Dias, R. S., and Vanetti, M. C. D. (2014). Efficacy of bovicin HC5 and nisin combination against Listeria monocytogenes and Staphylococcus aureus in fresh cheese. Int J Food Sci Technol. 49, 416–422. doi: 10.1111/ijfs.12316
Poimenidou, S. V., Chatzithoma, D. N., Nychas, G. J., and Skandamis, P. N. (2016). Variability in biofilm formation by Listeria monocytogenes strains on stainless steel and polystyrene surfaces and resistance to sanitizers. Int. J. Food Microbiol. 223, 23–30. doi: 10.1016/j.ijfoodmicro.2016.08.029
Poimenidou, S. V., Skarveli, A., Saxami, G., Mitsou, E. K., Kotsou, M., and Kyriacou, A. (2009). Inhibition of Listeria monocytogenes growth, adherence and invasion in Caco-2 cells by potential probiotic lactic acid bacteria isolated from fecal samples of healthy neonates. Microorganisms 11:363. doi: 10.3390/microorganisms11020363
Qiao, Z., Chen, J., Zhou, Q., Wang, X., Shan, Y., Yi, Y., et al. (2021). Purification, characterization, and mode of action of a novel bacteriocin BM173 from Lactobacillus crustorum MN047 and its effect on biofilm formation of Escherichia coli and Staphylococcus aureus. J. Dairy Sci. 104, 1474–1483. doi: 10.3168/jds.2020-18959
Ribeiro, A. C., Almeida, F. A., Medeiros, M. M., Miranda, B. R., Pinto, U. M., and Alves, V. F. (2023). Listeria monocytogenes: an inconvenient hurdle for the dairy industry. Dairy 4, 316–344. doi: 10.3390/dairy4020022
Riedel, C. U., Monk, I. R., Casey, P. G., Waidmann, M. S., Gahan, C. G. M., and Hill, C. (2009). AgrD-dependent quorum sensing affects biofilm formation, invasion, virulence and global gene expression profiles in Listeria monocytogenes. Mol. Microbiol. 71, 1177–1189. doi: 10.1111/j.1365-2958.2008.06589.x
Rieu, A., Weidmann, S., Garmyn, D., Piveteau, P., and Guzzo, J. (2007). Agr system of Listeria monocytogenes EGD-e: role in adherence and differential expression pattern. Appl. Environ. Microbiol. 73, 6125–6133. doi: 10.1128/AEM.00608-07
Rocha, K. R., Perini, H. F., de Souza, C. M., Schueler, J., Tosoni, N. F., Furlaneto, M. C., et al. (2019). Inhibitory effect of bacteriocins from enterococci on developing and preformed biofilms of Listeria monocytogenes, Listeria ivanovii and Listeria innocua. World J. Microbiol. Biotechnol. 35:96. doi: 10.1007/s11274-019-2675-0
Rocha, B. M. O., Sabino, Y. N. V., Almeida, T. C., Palacio, F. B., Rotta, I. S., Dias, V. C., et al. (2024). Unlocking probiotic potential: genomic insights into Weissella paramesenteroides UFTM 2.6.1. Probiot. Antimicrob. Proteins 1:1. doi: 10.1007/s12602-024-10409-x
Rodríguez-Melcón, C., Alonso-Calleja, C., García-Fernández, C., Carballo, J., and Capita, R. (2022). Minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) for twelve antimicrobials (biocides and antibiotics) in eight strains of Listeria monocytogenes. Biology 11:46. doi: 10.3390/biology11010046
Rotta, I. S., Matta, M. F., Santos, C. T. B., Paiva, A. D., and Ferreira-Machado, A. B. (2020). Bactérias do ácido lático potencialmente probióticas isoladas de leite não pasteurizado. Rev. Inst. Latic. Cândido Tostes 75, 178–189. doi: 10.14295/2238-6416.v75i3.820
Santos, C. A., EMF, L., Franco, B. D. G. M., and Pinto, U. M. (2021). Exploring phenolic compounds as quorum sensing inhibitors in Listeria monocytogenes. Front. Microbiol. 12:735931. doi: 10.3389/fmicb.2021.735931
Segev-Zarko, L., Saar-Dover, R., Brumfeld, V., Mangoni, M. L., and Shai, Y. (2015). Mechanisms of biofilm inhibition and degradation by antimicrobial peptides Available to Purchase Biochem J. 468, 259–270. doi: 10.1042/BJ20141251
Sela, S., Frank, S., Belausov, E., and Pinto, R. (2006). A mutation in the luxS gene influences Listeria monocytogenes biofilm formation. Appl. Environ. Microbiol. 72, 5653–5658. doi: 10.1128/AEM.00048-06
Shi, C., Chen, Y., Li, C., Al-Asmari, F., Cui, H., and Lin, L. (2024). Potential application of Lactiplantibacillus plantarum in food bio-preservation – a comprehensive review with a focus on the antibacterial and anti-virulence effects on foodborne pathogens. Food Rev. Int. 40, 2993–3019. doi: 10.1080/87559129.2024.2317283
Shi, W., Zhang, Q., Li, H., Du, D., Ma, X., Wang, J., et al. (2023). Biofilm formation, motility, and virulence of Listeria monocytogenes are reduced by deletion of the gene lmo0159, a novel Listerial LPXTG surface protein. Microorganisms 12:1354. doi: 10.3390/microorganisms12071354
Singh, J. K., Devi, P. B., Reddy, G. B., Jaiswal, A. K., Kavitaki, D., and Shetty, P. H. (2024). Biosynthesis, classification, properties, and applications of Weissella bacteriocins. Front. Microbiol. 15:1406904. doi: 10.3389/fmicb.2024.1406904
Singh, N., Kaur, R., Singh, B. P., Rokana, N., Goel, G., Puniya, A. K., et al. (2020). Impairment of Cronobacter sakazakii and Listeria monocytogenes biofilms by cell-free preparations of lactobacilli of goat milk origin. Folia Microbiol. 65, 185–196. doi: 10.1007/s12223-019-00721-3
Skandamis, P. N., and Nychas, G. J. (2012). Quorum sensing in the context of food microbiology. Appl. Environ. Microbiol. 78, 5473–5482. doi: 10.1128/AEM.00468-12
Stepanóvic, S., Vukovic, D., Dakic, I., Savic, B., and Svabic-Vlahovic, M. (2000). A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 40, 175–179. doi: 10.1016/S0167-7012(00)00122-6
Tomé, A. R., Carvalho, F. M., Teixeira-Santos, R., Burmølle, M., Mergulhão, F. J. M., and Gomes, L. C. (2023). Use of probiotics to control biofilm formation in food industries. Antibiotics 12:754. doi: 10.3390/antibiotics12040754
Tuccillo, F., Wang, Y., Edelmann, M., Lampi, A. M., Coda, R., and Katina, K. (2022). Fermentation conditions affect the synthesis of volatile compounds, dextran, and organic acids by Weissella confusa A16 in faba bean protein concentrate. Food Secur. 11:3579. doi: 10.3390/foods11223579
Tuytschaever, T., Raes, K., and Sampers, I. (2023). Listeria monocytogenes in food businesses: from persistence strategies to intervention/prevention strategies - a review. Compr. Rev. Food Sci. Food Saf. 22, 3910–3950. doi: 10.1111/1541-4337.13219
Vadassery, D. H., and Pillai, D. (2020). Quorum quenching potential of Enterococcus faecium QQ12 isolated from gastrointestinal tract of Oreochromis niloticus and its application as a probiotic for the control of Aeromonas hydrophila infection in goldfish Carassius auratus. Braz. J. Microbiol. 51, 1333–1343. doi: 10.1007/s42770-020-00230-3
Vasiee, A., Falah, F., and Mortazavi, S. A. (2022). Evaluation of probiotic potential of autochthonous lactobacilli strains isolated from Zabuli yellow kashk, an Iranian dairy product. J. Appl. Microbiol. 133, 3201–3214. doi: 10.1111/jam.15772
Vázquez- Armenta, F. J., López-Malo, A., and Palou, E. (2020). Inhibition of biofilm formation and related gene expression of Listeria monocytogenes by natural antimicrobial compounds. Front. Microbiol. 11:617473. doi: 10.3389/fmicb.2020.617473
Vilas Boas, D., Castro, J., Araújo, D., Nóbrega, F. L., Keevil, C. W., Azevedo, N. F., et al. (2024). The role of flagellum and flagellum-based motility on salmonella Enteritidis and Escherichia coli biofilm formation. Microorganisms 12:232. doi: 10.3390/microorganisms12020232
Wiktorczyk-Kapischke, N., Walecka-Zacharska, E., Korkus, J., Grudlewska-Buda, K., Budzynska, A., Wnuk, K., et al. (2023). The influence of stress factors on selected phenotypic and genotypic features of Listeria monocytogenes - a pilot study. BMC Microbiology. 23:259. doi: 10.1186/s12866-023-03006-5
Williams, T., Joseph, B., Beier, D., Goebel, W., and Kuhn, M. (2005). Response regulator DegU of Listeria monocytogenes regulates the expression of flagella specific genes. FEMS Microbiol. Lett. 252, 287–298. doi: 10.1016/j.femsle.2005.09.011
Winkelströter, L. K., Teixeira, F. B. R., Silva, E. P., Alves, V. F., and Martins, E. C. P. (2014). Unraveling Microbial Biofilms of Importance for Food Microbiology. Microb. Ecol. 68, 35–46. doi: 10.1007/s00248-013-0347-4
Yang, Y., Kong, X., Niu, B., Yang, J., and Chen, Q. (2024). Differences in biofilm formation of Listeria monocytogenes and their effects on virulence and drug resistance of different strains. Food Secur. 13:1076. doi: 10.3390/foods13071076
Yu, T., Jiang, X., Xu, X., Jiang, C., Kang, R., and Jiang, X. (2022). Andrographolide inhibits biofilm and virulence in Listeria monocytogenes as a quorum-sensing inhibitor. Molecules 27:3234. doi: 10.3390/molecules27103234
Yu, H. S., Lee, N. K., Choi, A. J., Choe, J. S., Bae, C. H., and Paik, H. D. (2019). Antagonistic and antioxidant effect of probiotic Weissellacibaria JW15. Food Sci. Biotechnol. 28, 851–855. doi: 10.1007/s10068-018-0519-6
Zamani, S., Sadeghi, A., and Ghanbarzadeh, B. (2017). Quorum sensing in Listeria monocytogenes: a review on its role in biofilm formation and virulence. J. Food Sci. 82, 315–322.
Zhang, X., Lu, Z., Zheng, L., Lü, Z., Zhou, L., Meng, F., et al. (2022). Effect of quorum sensing systems on biofilm formation by Listeria monocytogenes. Food Sci. 43:105. doi: 10.7506/spkx1002-6630-20211002-006
Zhao, D., Jiang, J., Liu, L., Wang, S., Ping, W., and Ge, J. (2021). Characterization of exopolysaccharides produced by Weissella confusa XG-3 and their potential biotechnological applications. Int. J. Biol. Macromol. 178, 306–315. doi: 10.1016/j.ijbiomac.2021.02.182
Zhou, L., Zhang, Y., Ge, Y., Zhu, X., and Pan, J. (2020). Regulatory Mechanisms and Promising Applications of Quorum Sensing-Inhibiting Agents in Control of Bacterial Biofilm Formation. Front Microbiol. 11:589640. doi: 10.3389/fmicb.2020.589640
Keywords: biofilm, foodborne pathogen, lactic acid bacteria, probiotic, quorum quenching
Citation: Rotta IS, Rezende SDdC, Perini HF, da Silva MV, de Almeida FA, Pinto UM, Ferreira Machado AB and Paiva AD (2025) Exploring the potential of Weissella paramesenteroides UFTM 2.6.1 in disrupting quorum sensing and attenuating virulence in Listeria monocytogenes. Front. Microbiol. 16:1601203. doi: 10.3389/fmicb.2025.1601203
Edited by:
Vincenzina Fusco, National Research Council (CNR), ItalyReviewed by:
Carlo Giuseppe Rizzello, Sapienza University of Rome, ItalyDigambar Kavitake, National Institute of Nutrition (ICMR), India
Copyright © 2025 Sguilla Rotta, Rezende, Perini, da Silva, de Almeida, Pinto, Ferreira Machado and Paiva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Aline Dias Paiva, YWxpbmUucGFpdmFAdWZ0bS5lZHUuYnI=