 Xueqin Tan
Xueqin Tan Beiyi Wu
Beiyi Wu Xue Wen
Xue Wen Yunzhu Li2
Yunzhu Li2 Xuewen Xu
Xuewen Xu- 1Research Laboratory of Plastic and Burns Surgery, West China Hospital of Sichuan University, Chengdu, China
- 2Department of Plastic and Burn Surgery, West China Hospital of Sichuan University, Chengdu, China
Alcohol is inextricably linked with intestinal microbiota as it was absorbed through gut. While mitochondrial aldehyde dehydrogenase 2 (ALDH2), as the major enzyme responsible for metabolizing toxic acetaldehyde to acetate, is important factor influencing alcohol metabolism. However, it is not yet known the relationship between ALDH2 knockout (KO) and gut microbiota profiles in mice under chronic alcohol exposure. Therefore, this study aimed to investigate the effect of 5% v/v alcohol exposure on the gut microbiota of ALDH2 knockout (KO-5%) and wild-type (WT-5%) mice. At the end of 10-week experiment, KO-5% mice exhibited a higher serum acetaldehyde concentration and upregulated expression of pro-inflammatory cytokines in intestine tissue than WT-5% mice. Metagenomic results revealed that the KO-5% mice had a significant decrease in alpha diversities. Moreover, KO-5% mice exhibited gut microbiota dysbiosis with the characteristic of a higher abundance of phylum Proteobacteria, and genera Stenotrophomonas and Ralstonia, whereas the level of genera Lactobacillus, unclassfied Bacilli, and Turicibacter were decreased. Additionally, genera Candidatus Arthromitus and Ralstonia were the most representatives in the KO-5% mice. Further, chronic alcohol exposure resulted in enriched expression of genes associated with bacterial metabolism and cellular processes in gut from WT mice. Taken together, our findings demonstrated a strong interaction between ALDH2 and the gut microbiota to response to alcohol exposure.
1 Introduction
Alcohol is a unique addictive substance consumed worldwide, exhibiting complexing pharmacological qualities in the body (Piano et al., 2025). Harmful alcohol use, encompassing alcohol abuse and chronic exposure has profound adverse effects on the human body and is well documented to be associated with causing social harm and financial burden (Axley et al., 2019). Alcohol is absorbed throughout the entire gastrointestinal tract, with a small proportion of alcohol absorption occurs in the mouse and esophagus, ~20% is absorbed in the stomach, followed by ~70% absorbed from the small intestine (Moreau et al., 1997). The rate of absorption and metabolism was affected by a number of factors, such as host conditions, the concentration of alcohol, the speed of which the drink was consumed, and whether the alcohol are drunk on an empty stomach (Butts et al., 2023). Recently, the possibility has been raised that alcohol intake may also influence the gut microbiota compositions and functions (Engen et al., 2015).
Mammalian gastrointestinal tract harbors trillions of microbes including bacteria, protozoa, viruses, archaea, and fungi. This population of microbes displays a broad range of symbiotic interactions with the host (Mallott and Amato, 2021; Sidhu and van der Poorten, 2017). Moreover, studies have provided compelling evidence for the critical role of gut microbiota in chronic alcohol exposure, one study using 16S rRNA sequencing method found that a lower abundance of Bacteroidetes and a higher abundance of Proteobacteria of mucosa was observed in alcoholics (Mutlu et al., 2012). Another study showed a reduction from Bacteroidetes and an increase from Proteobacteria and Fusobacteria in human subjects with alcohol-related cirrhosis (Chen et al., 2011). In addition, it is now generally accepted that gut microbiota is one of the critical players in alcohol-induced injury in body, and the bacterial products appears to play a key role in the progression of alcohol associated liver disease (Vassallo et al., 2015; Way et al., 2022). Nevertheless, the etiology of alcohol-induced injury is a complex interplay of factors, and these factors determining alcohol toxicity remain unclear, implying that further studies are needed to clarify the interactions between gut microbiome and other factors (Wang et al., 2024).
After absorbed in the gastrointestinal tract, alcohol circulated in the blood and diffused into organs. Then, oxidative alcohol metabolism take place mainly in the liver, where most alcohol was oxidized to toxic acetaldehyde (Zakhari, 2006; Foster and Marriott, 2006). Three separate enzyme systems have been identified to catalyze oxidative alcohol metabolism, among them, mitochondrial aldehyde dehydrogenase 2 (ALDH2), as one of the major rate-limiting enzymes in this reaction, is mainly responsible for detoxifying toxic acetaldehyde to non-toxic acetic acid (Cederbaum, 2012). Excessive accumulation of acetaldehyde has cytotoxicity since acetaldehyde has the capacity to bind to proteins, form carcinogenic with DNA, and forms adducts with neurotransmitter, which can cause DNA even organ damage, resulting in a range of metabolic diseases (Park et al., 2016; Chen et al., 2019). Although the amount of ALDH2 in the liver is abundant, the activity that is closely connected with the ALDH2 genotypes (Chen et al., 2022). Specifically, there is a significant genetic polymorphism (rs671, G>A) of the ALDH2 gene, resulting in allelic variant ALDH2*1 (G) and allelic variant ALDH2*2 (A). Of note, the allelic variant ALDH2*2 has been identified to reduced ALDH2 enzymatic activity in comparison with ALDH2*1 (Edenberg, 2007). People with one or especially two copies of ALDH2*2 allele exhibited higher acetaldehyde levels after alcohol consumption. Moreover, the ALDH2*2 dysfunctional variant causes Asian flush syndrome or other uncomfortable symptoms during alcohol consumption in more than 560 million people worldwide, especially in east Asians (Wall et al., 2000; Chen et al., 2020). Previous works performed on mice revealed that ALDH2 knockout or deficiency led to increased sensitivity to alcohol-related DNA damage and tissue injury (Kwon et al., 2014; Ma et al., 2010; Matsumoto et al., 2014; Rungratanawanich et al., 2023). Additionally, the host gene expression may affect intestinal microbial composition (Goodrich et al., 2017; Davenport, 2016). However, the crosstalk between ALDH2 and the composition of gut microbiota in chronic alcohol exposure and the detailed mechanisms is still unclear. Therefore, exploring the relationship between ALDH2 and gut microbiota in alcohol exposure is meaningful and worthwhile, and we are aimed to investigate the gut microflora composition and function in ALDH2 KO and WT mice after exposure to alcohol in this study.
2 Materials and methods
2.1 Animals
All animal protocols were approved by the Animal Ethics Committee of West China Hospital of Sichuan University (Code: 20230228133). Six-week-old ALDH2 knockout (KO) and wild-type (WT) mice on C57BL/6 background were bought from Cyagen Biosciences lnc. (Suzhou, China). KO individuals were generated by mating heterozygous mice as described previously (Qiu et al., 2023). All mice were housed in a SPF facility (five animals per cage) under a 12-h day and night lighting cycle with free access to food. To confirm the efficiency of our knockouts, the ALDH2 expression at the mRNA and protein level in both liver and ileum tissues was measured by real time PCR and western blot analysis, respectively. The primer sequences of ALDH2 and 18 s were presented in Table 1. GAPDH and ALDH2 protein levels were detected by rabbit anti-GAPDH antibody (Cell Signaling Technology, 1:1000) and rabbit anti-ALDH2 antibody (Proteintech, 1:2000).

Table 1. Information of the primers used for qPCR reactions.
2.2 Chronic alcohol exposure
WT and KO C57BL/6 male mice were used to construct chronic alcohol exposure model through drinking 5% concentration (v/v) of alcohol in their water. The alcohol exposure lasted for 10 weeks. Based on alcohol consumption and ALDH2 genotype, mice were randomly divided into four groups: (1) KO mice were fed 5% alcohol (KO-5%, n = 5), (2) KO mice drinking water (KO-Control, n = 5), (3) WT mice receiving 5% alcohol (WT-5%, n = 5), and (4) WT mice receiving water (WT-Control, n = 5). The water intake (g) and body weight (g) were recorded weekly. At the end of the experiment, all mice were deeply anesthetized with sodium pentobarbital (50 mg/kg), and blood from the heart chambers were collected and stand at room temperature for 1 h, then centrifuged for 15 min at 3,500 × g at 4°C to separate serum. The level of serum ethanol was measured by Ethanol Assay Kit (BioVision; K620-1001, United States) according to manufacturer’s instructions. Concentrations of serum acetaldehyde were determined using aldehyde Quantitation Kit (AAT Bioquest; 10,051, United States). Meanwhile, intestinal contents were collected from approximately the middle region of ileum, and immediately frozen in liquid nitrogen and subsequently transferred to a −80°C freezer until DNA extraction.
2.3 RNA extraction and quantitative real-time PCR (qPCR)
Intestinal tissue samples were used to isolate RNA and differential expression of major inflammatory factors was detected through qPCR. Total RNA was extracted using Cell Total RNA Isolation Kit following manufacturer’s protocol (RE-03111, FOREGENE, China), and RNA concentrations and purities were measured by NanoDrop (Thermo Fisher Scientific Inc.). Subsequently, the first-strand cDNA was synthesized with an iScript cDNA Synthesis Kit (Bio-Rad, Hercules, CA, United States) according to the manufacturer’s recommendations. After RNA purification and cDNA synthesis, qPCR was performed with iTaq Universal SYBR Green Supermix (Bio-Rad, Hercules, CA, United States) on the CFX Connect Real-Time System with the cycling conditions: 95°C for 10 min; 40 cycles of 95°C for 10 s and 60°C for 40 s with fluorescent reading. Transcript levels were analyzed by comparative ΔΔCt method and 18 s served as endogenous reference control. Specific primers sequences are listed in Table 1.
2.4 DNA isolation and 16S rDNA sequencing
The microbial DNA was isolated from approximately 0.4 g of intestinal content samples using TIANamp stool DNA isolation Kit (Qiagen, Shanghai, China) according to the manufacturer’s protocol. The concentration and integrity of the DNA was assessed using Nanodrop ND1000 spectrophotometer (Nanodrop Technologies, Mont-chanin, DE, United States) and 2% gel electrophoresis, respectively. Afterwards, 30 ng of DNA was used as template to amplify the V3-V4 region of the 16S rDNA by using the universal primers 338F (5′-ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-GGAC-TACHVGGGTWTCTAAT-3′). Subsequently, the PCR products were checked by visualization with gel electrophoresis, and quantified for sequencing libraries construction. Finally, the qualified libraries were sequenced paired-end on the Illumina HiSeq 2,500 platform.
2.5 Bioinformatics and data analysis
The raw reads were initially processed using the Quantitative Insights into Microbial Ecology2 (QIIME2 v.2023.5) open-source bioinformatics tool (Bolyen et al., 2019). Pair end reads were joined and the Debular program (Amir et al., 2017) was used to demultiplexed sequences. Following this, sequences can be assigned into operational taxonomic units (OTUs), and the OTUs were taxonomically classified based on the Silva database (v.138) (Quast et al., 2013). Then, the alpha diversity metrics including Shannon and Simpson indices were evaluate by using the Kruskal-Wallis test. For beta diversity, calculation of the Bray Curtis distance and Unweighted Unifrac metrics were performed with the QIIME2 platform, and the ggplot2 (v3.5.0) package was used to visualize these metrics. The characterization of bacterial features was performed using the Linear discriminant analysis (LDA) effect size (LEfSe) for biomarker discovery (Segata et al., 2011). Finally, the software PICRUSt2 (v1.7.3) (Douglas et al., 2020) was enrolled to investigate if there were functional similarities between groups, and the resulting pathway abundance data was visualized using the ggpicrust2 package. Statistical analyses were performed using R (v4.3.2) software.1 Differences between pairs of means were considered significant when p < 0.05.
3 Results
3.1 Animal model establishment of chronic alcohol exposure
In order to evaluate the ALDH2 knockout efficiency of KO mice, we examined the ALDH2 expression profiles in both liver and ileum tissues by real time PCR and western blot analysis. The results showed that ALDH2 mRNA (Figure 1A) and protein (Figure 1B) were highly expressed in WT mouse liver and ileum tissues, whereas an undetectably low level in both tissues regarding KO mice. After 10-week ethanol exposure, the average daily water intake in mice of the four groups were similar, without significant difference between the WT and WT-5% groups, or between the KO and KO-5% groups (Figure 1C). Further, the body weight was recorded and we found that both KO-5% and WT-5% mice had a lower body weight gain when compared with KO-Control and WT-Control mice (Figure 1D). Additionally, ethanol clearance in acetaldehyde metabolism was analyzed, which showed that, although sera ethanol concentration did not show significant differences in the KO-5% compared to WT-5% mice (Figure 1E), the serum acetaldehyde level was remarkably elevated in the KO-5% mice than that in the WT-5% mice (p < 0.01) and KO-Control (p < 0.01) (Figure 1F). These findings suggested that the chronic alcohol exposure induced decreases in body weight gain, and the loss of ALDH2 gene can inhibiting the elimination of blood acetaldehyde.
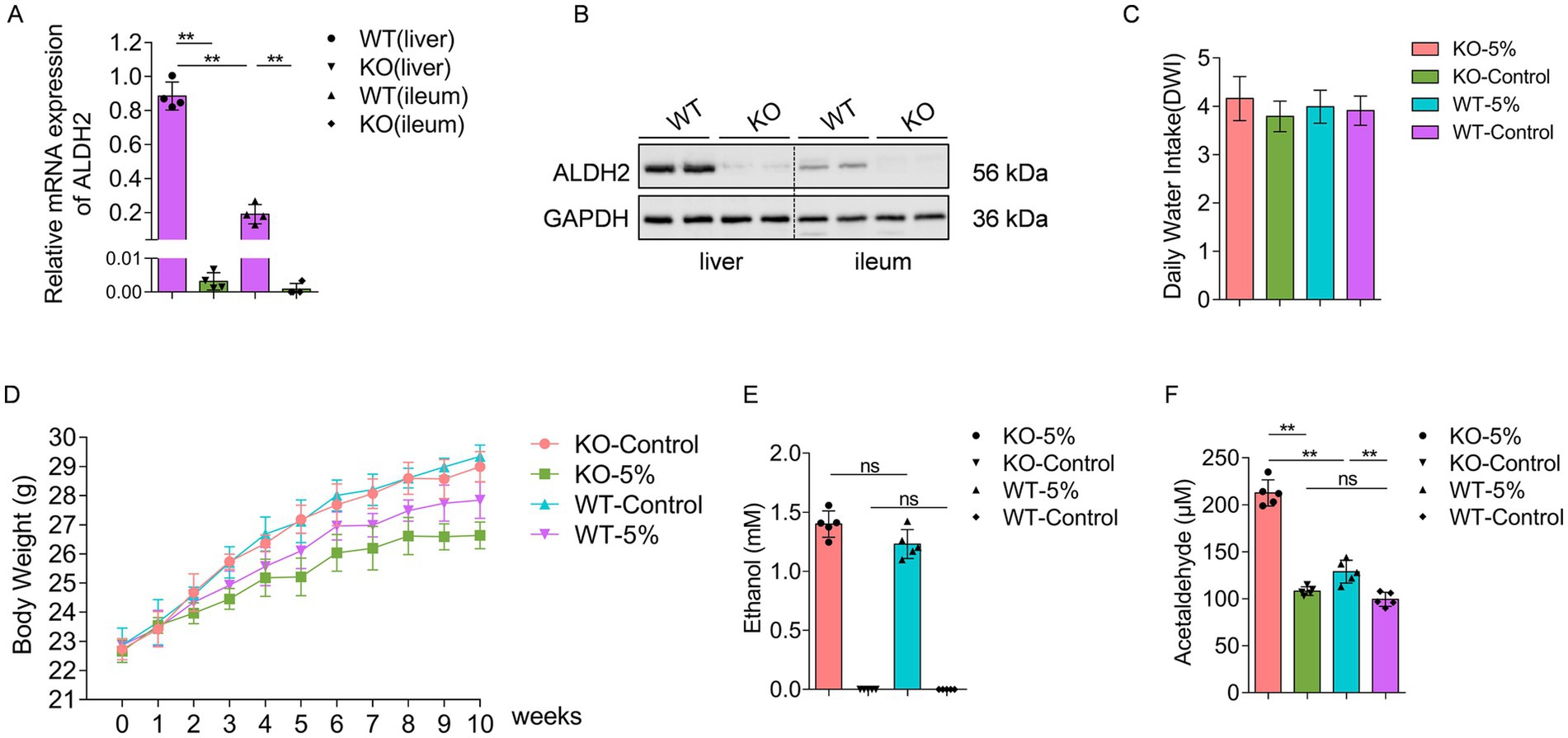
Figure 1. Establish a chronic alcohol exposure animal model. The level of ALDH2 at mRNA (A) and protein (B) was highly expressed in WT mice when compared with KO mice in both liver and ileum tissues. The daily water intake (C) and body weight (D) during the 10-week alcohol exposure had no significant differences across different experimental groups. Ethanol concentrations (E) and acetaldehyde concentrations (F) were measured at the end of experiment among different groups. Data are presented as “mean ± SD,” *p < 0.05, **p < 0.01.
3.2 Chronic alcohol exposure exacerbated intestinal inflammation in ALDH2 knockout mice
Differential mRNA expression of inflammatory cytokines, including TNF-α, IL-1β, IL-6, and IL-10, was analyzed in the four groups. After 10-week chronic alcohol exposure, the relative mRNA levels of TNF-α, IL-1β, and IL-6 were significantly increased in KO-5% and WT-5% mice compared to KO-Control and WT-Control groups, respectively (Figures 2A–C). Moreover, KO-5% mice exhibited the most pronounced elevation in all pro-inflammatory cytokine levels relative to other groups. This cytokine profile demonstrated that ethanol-induced intestinal injury, with ALDH2 deficiency exacerbating the inflammatory response. Meanwhile, IL-10 exhibited opposite expression patterns across the four groups (Figure 2D), suggesting its potential role in suppressing excessive inflammation responses through negative feedback regulation.
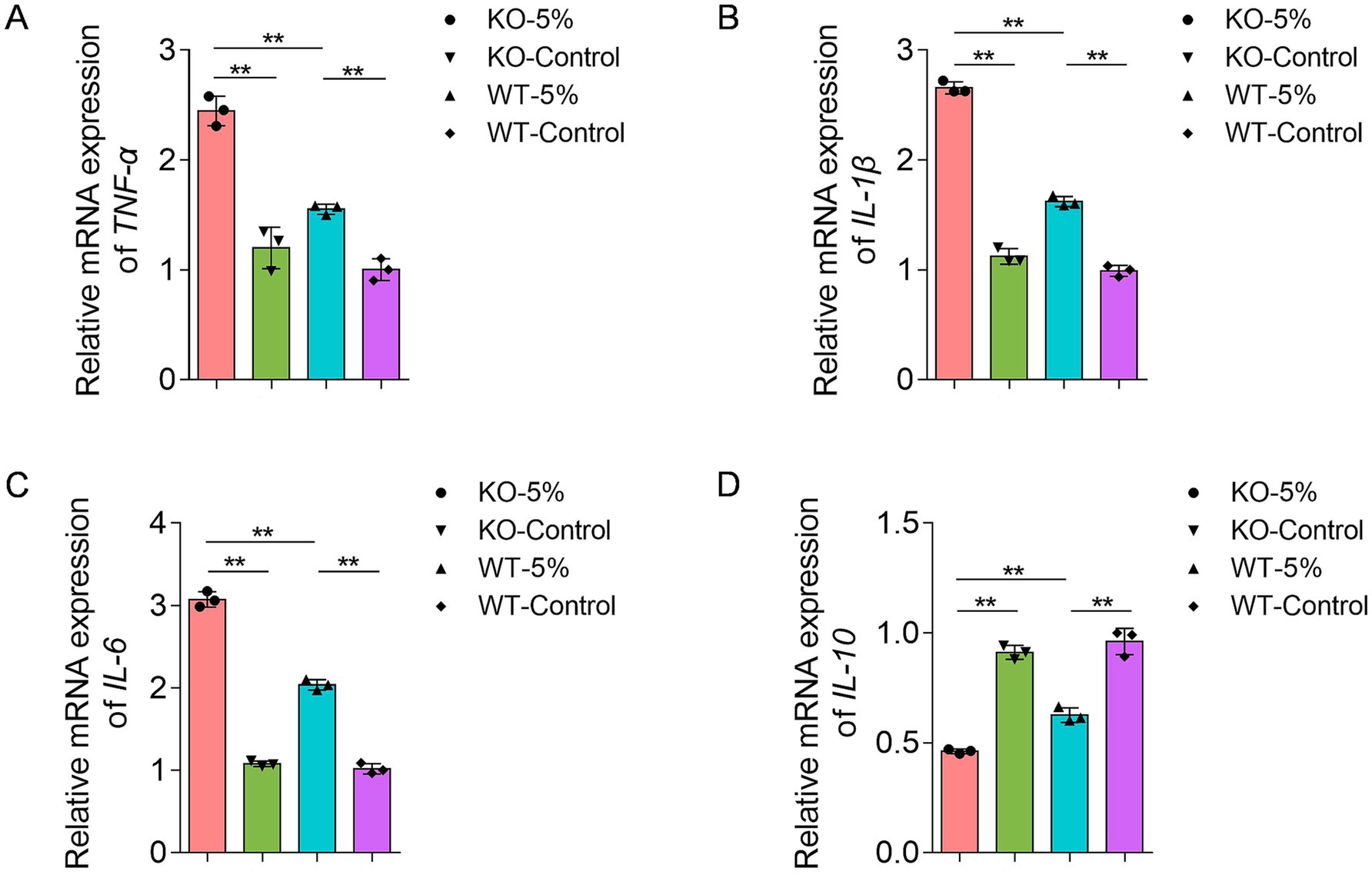
Figure 2. Effects of chronic alcohol exposure on the mRNA expression of inflammatory biomarkers in ALDH2 knockout mice. (A) Expression of TNF-α. (B) Expression of IL-1β. (C) Expression of IL-6. (D) Expression of IL-10. Data are presented as mean ± SD (n = 3). *p < 0.05, **p < 0.01.
3.3 Chronic alcohol exposure reduced microbial community diversity in ALDH2 knockout mice
Changes in gut microbiota diversity between the four groups of mice were analyzed. The Chao1 and Shannon indexes showed that the alpha diversities were significantly reduced in KO-5% when compared to the WT-5% mice (p = 0.028 for Chao1 richness index, Figure 3A; p = 0.011 for Shannon diversity index, Figure 3B). Moreover, the principal-coordinate analysis (PCoA) results based on both Bray Curtis (Figure 3C) and Unweighted Unifrac distances (Figure 3D) were used to measure the beta diversity in each group. The results showed that a notable shift in beta diversities was observed as the bacterial members and structure of the KO-5% mice were different from that of the WT-5% mice. Therefore, it was concluded that the ALDH2 loss might reduce the richness and diversity of intestinal bacterial species when mice were under chronic alcohol exposure.
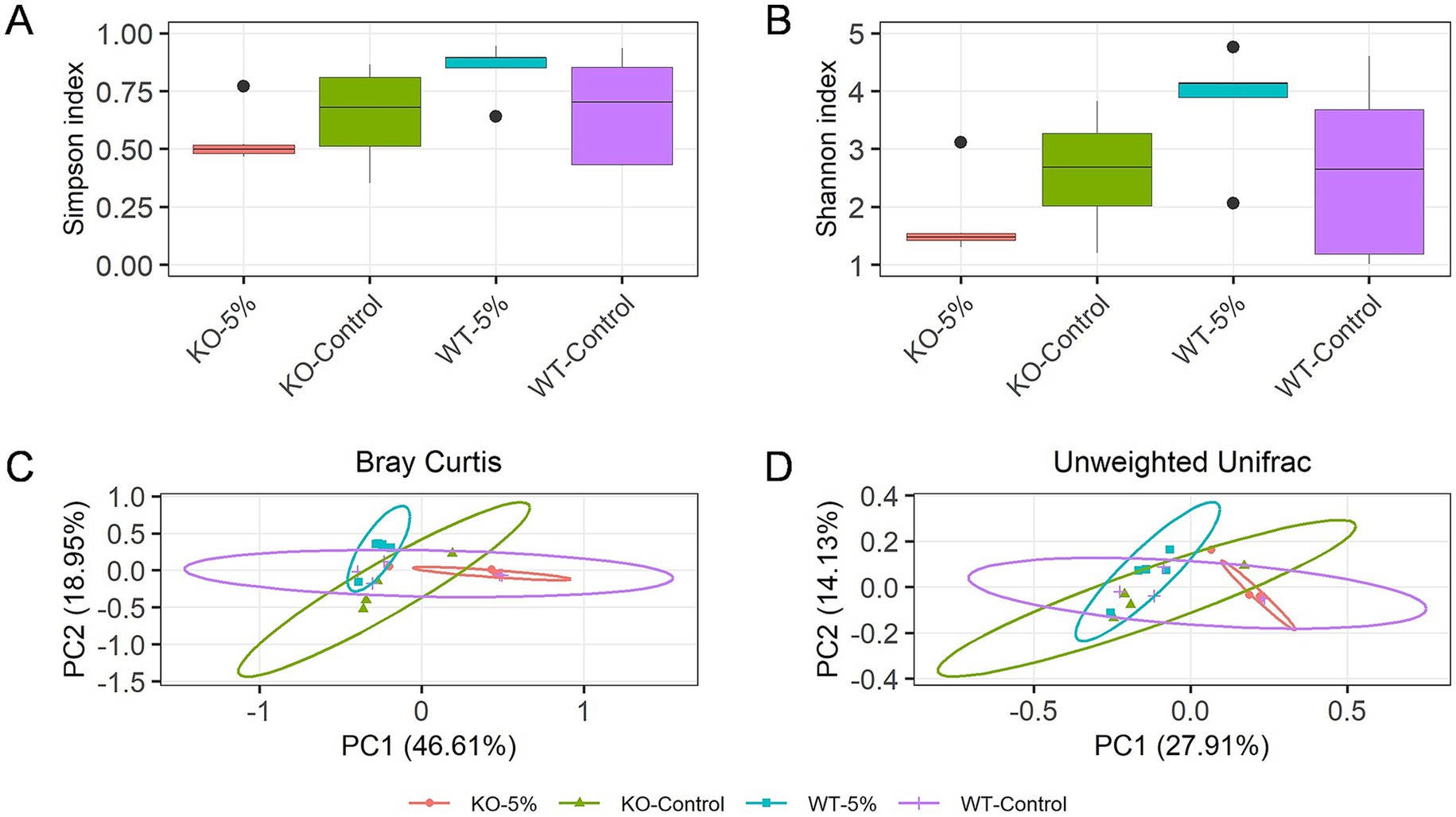
Figure 3. Chronic alcohol exposure resulted in distinct gut microbiota profiles in KO and WT mice. Alpha diversity within samples was measured by Chao1 (A) and Shannon index (B). Scatterplot from Principal Coordinate Analysis (PCoA) of beta diversity was measured by Bray-Curtis index (C) and Unweighted Unifrac distance (D). Results were determined by Kruskal–Wallis test when *p < 0.05, **p < 0.01.
3.4 Chronic alcohol exposure changed gut bacterial composition in ALDH2 knockout mice
We focused on the bacterial taxa relative abundance at the phylum and genus level. Among these phyla, Proteobacteria (95.97% ± 3.31) was the predominant phylum in KO-5% mice, but a significant decreased abundance of Proteobacteria was observed in KO-Control, WT-5%, and WT-Control animals with 16.38, 57.97, and 26.78%, respectively (Figure 4A). Moreover, in KO-Control and WT-5% mice, the predominant phylum was Firmicutes (79.33 and 57.97%, respectively), whereas that was decreased sharply in KO-5% mice. At the genus level, the most abundant taxa were Stenotrophomonas in KO-5% mice, followed by Ralstonia, Delftia, unclassfied Bacilli and Lactobacillus (Figure 4B). Contrarily, the WT-5% inhibited the growth of these taxa. Of note, Lactobacillus, unclassfied Bacilli, Vibrio, and Pseudoalteromonas were increased in WT-5% compared to WT-Control mice but they were less abundant in KO-5% compared to KO-Control mice (Figure 4C).
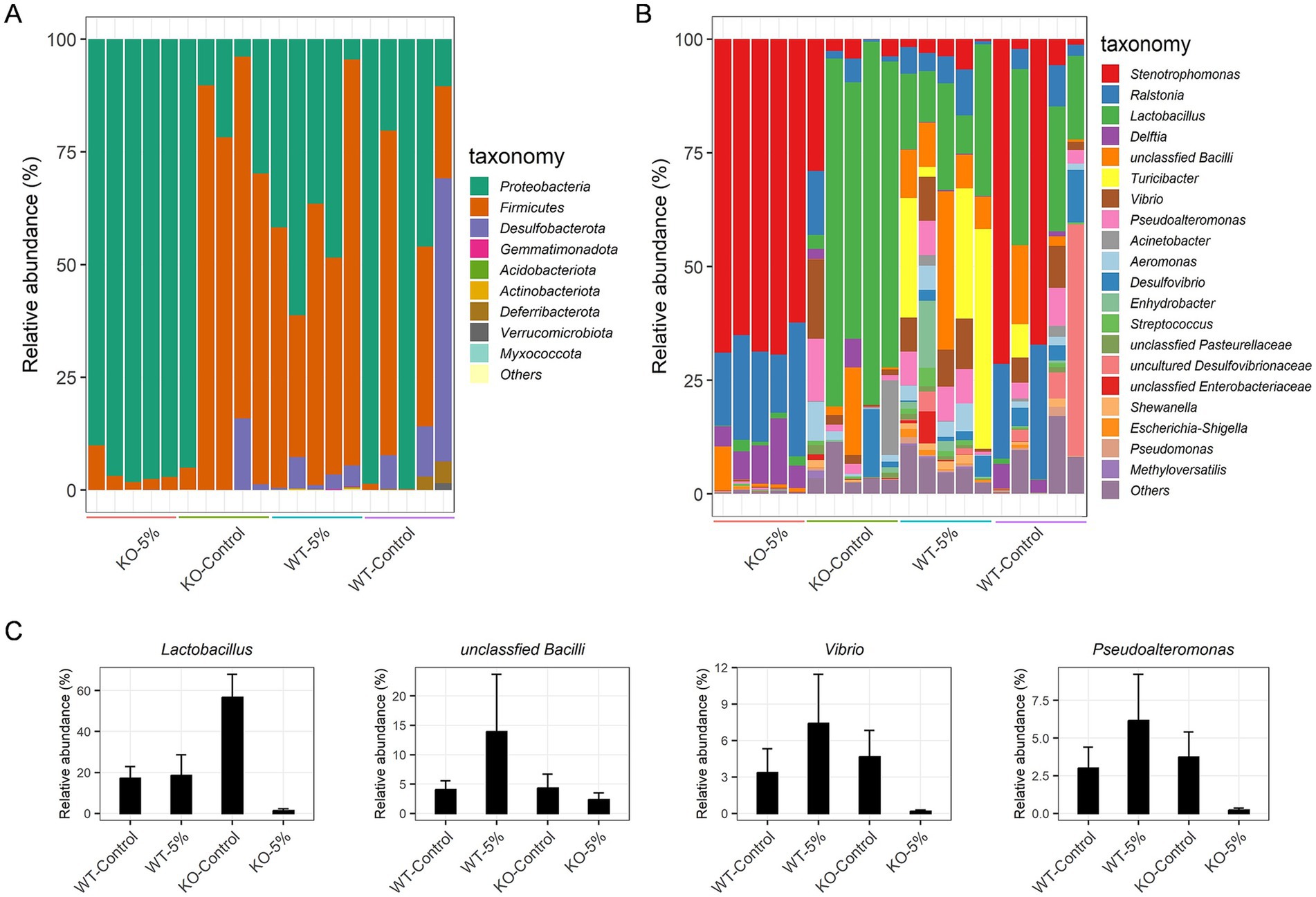
Figure 4. Chronic alcohol exposure resulted in distinct community structure in KO and WT mice. Summary of the relative abundances of bacterial phyla (A) and the top 20 most abundant bacterial genera (B). Each color represents one bacterial species on the stacked bar chart. Relative abundance of special bacteria genera that expressed higher in WT-5% than WT-Control but which has lower abundance in KO-5% than KO-Control mice (C).
As a complementary and validating method to identify specific differences in taxa, the LEfSe analysis was performed between WT-5% and KO-5% mice with a threshold of LDA values > 4. Firstly, we generated a bar plot of the effect size of taxa and found 7 significantly enriched genera in the WT-5% mice, including Turicibacter, Lactobacillus, Vibrio, Pseudoalteromonas, Aeromonas, Alidiomarina, and Desulfovibrio. Whereas genera Candidatus Arthromitus and Ralstonia were the most representatives in the KO-5% mice (Figure 5A). Of note, all genera except for Alidiomarina, and Candidatus Arthromitus were presented in the top 20 abundant taxa. Secondly, we performed a cladogram to show the relationship between different taxa (Figure 5B). This figure illustrated several taxonomic groups such as Lactobacillus and Turicibacter within Fimicutes phylum, in particular the class Bacilli, were at a significant higher abundance in WT-5% mice. Furthermore, genus Ralstonia was found to be a significantly high abundant taxa in KO-5% mice (Figure 5B).
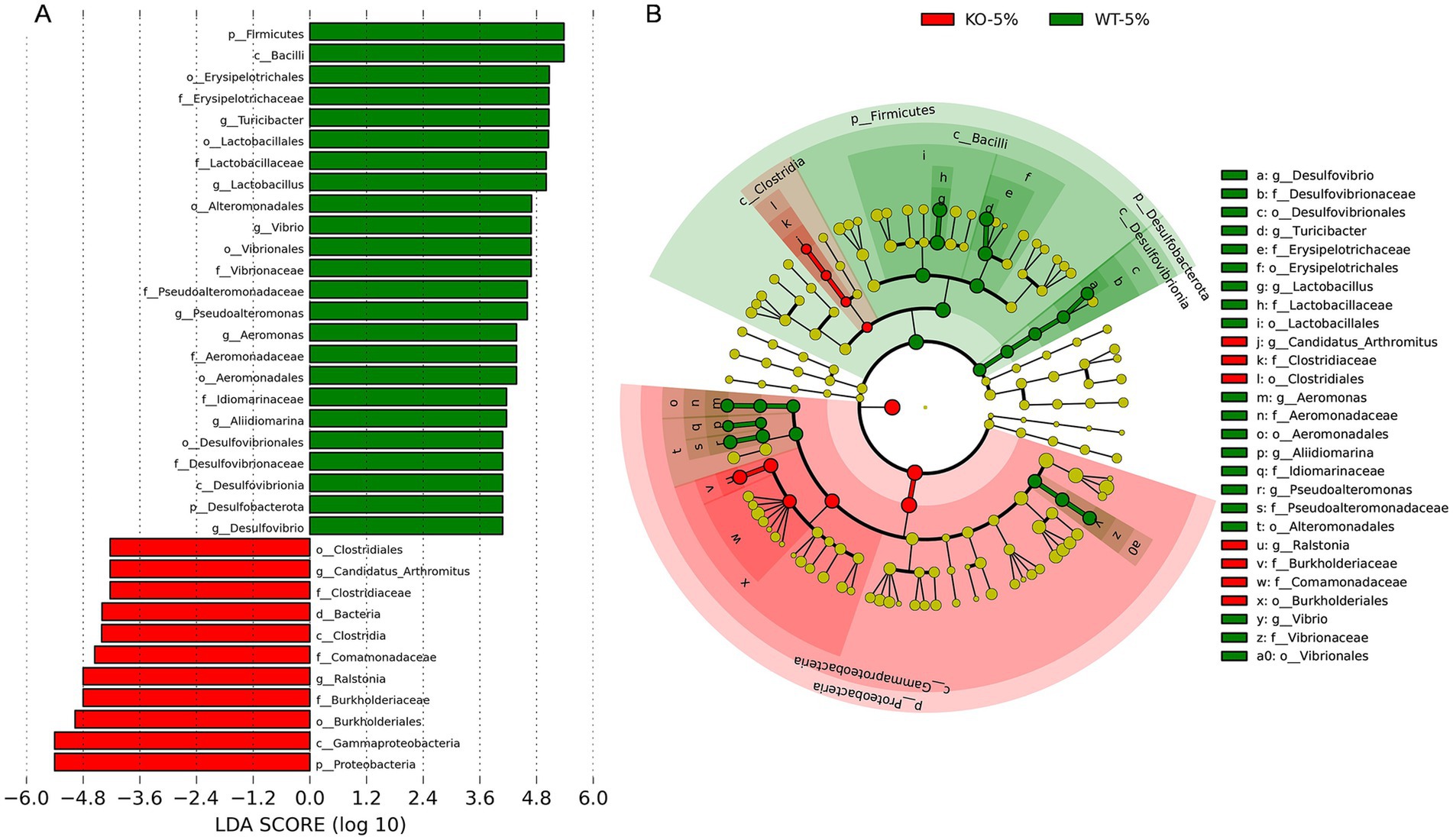
Figure 5. Differential represent taxa between KO-5% and WT-5% mice on the basis of Linear discriminant analysis effect size (LEfSe) analysis. Histogram of Linear Discriminant Analysis (LDA) scores indicating statistical different relative levels across all taxa (A). Cladogram revealing the phylogenetic relationships between taxa with the statistically different (B). Circle sizes in the cladogram plot represent the proportion to bacterial abundance, and the yellow circles represent different taxon with no obvious difference.
3.5 Predicted functional differences of the gut microbiota between KO-5% and WT-5% mice
Using PICRUSt2 software and KEGG database, we compared the gene abundance of the microbiota and inferred the profile of putative microbial functions. As illustrated in Figure 6, these genes were merged into level 3 KEGG orthologs, based on their cellular processes, environmental information processing, genetic information processing, human disease, metabolism, organismal systems. Notably, we found that the gut microbiota of KO-5% mice had relatively less bacterial containing genes for several pathways, such as peroxisome, primary immunodeficiency, fatty acid degradation, alpha-linolenic acid metabolism, pantothenate and CoA biosynthesis, terpenoid backbone biosynthesis, etc. The majority of these pathways were linked to aspects of bacterial metabolism and cellular processes, suggesting that the KO-5% showed a decreased metabolic response on the gut microbiota compared to WT-5% mice.
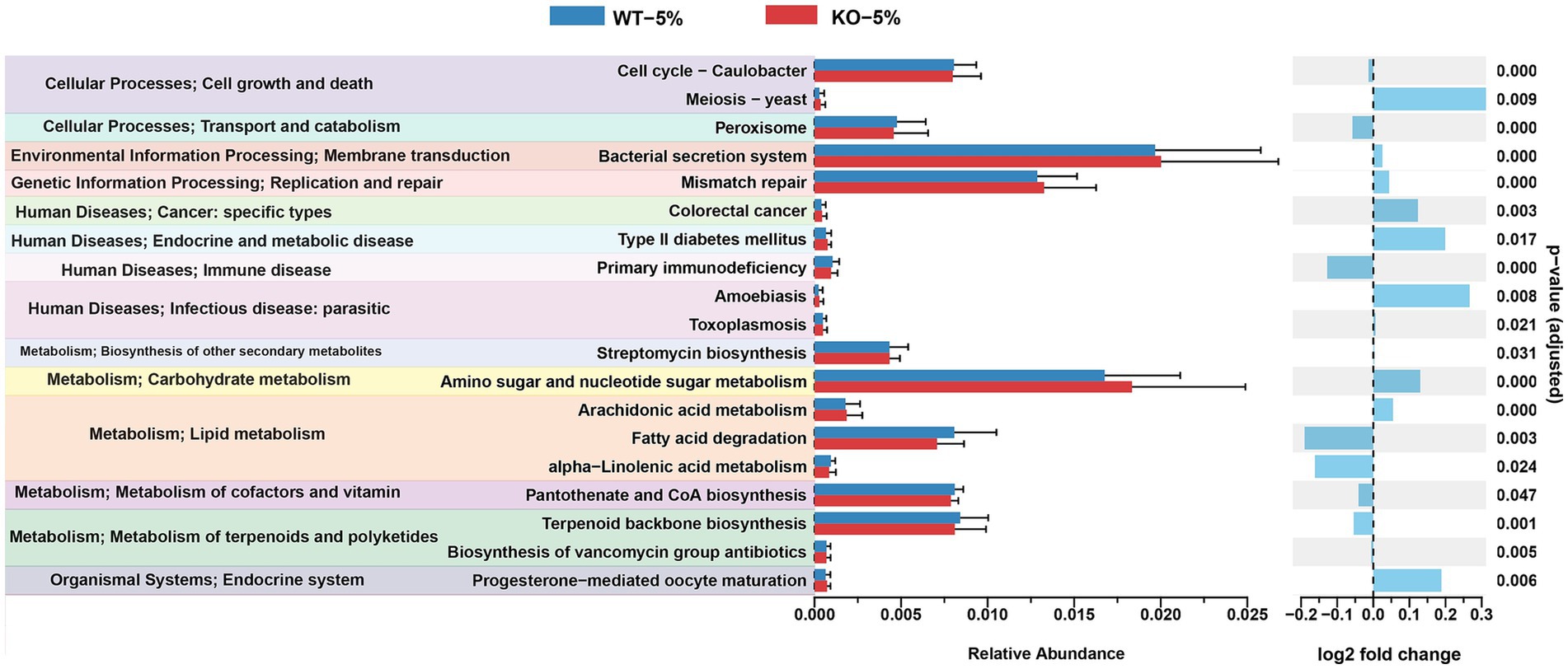
Figure 6. Differential abundances of bacterial functional pathways between KO-5% and WT-5% mice. All KEGG orthologs shown have an adjusted q value cutoff of 0.05.
4 Discussion
The harms associated with alcohol are well documented, and heavy and long-term alcohol consumption were associated with serious physical and psychiatric illnesses such as liver disease, pancreatitis, depression, and dementia (Shimizu et al., 2016). Although various alcohol consumption patterns have been defined and the exact definitions are blurred, to date, acute and chronic drinking are the most relevant two alcohol drinking patterns for studying alcohol influence (Alcohol Research: Current Reviews Editorial Staff, 2018). In fact, the effects of acute and chronic alcohol consumption have been widely studied, most works have focused on alcohol-related liver disease because liver is the main site of alcohol metabolism (Massey and Arteel, 2012; Abbas et al., 2023). Additionally, recent works unveiled that alcohol intake impairs the function and homeostasis of gut microbiota (Wang et al., 2024; Leclercq et al., 2020), and further extended the interactions across host gene and gut microbiota in alcohol exposure murine model (Rungratanawanich et al., 2023; Liu et al., 2022).
It is well established that ALDH2 gene mutation as well as ALDH2 knockout induced an increased sensitivity to tissue injury under alcohol exposure (Chen et al., 2020; Ma et al., 2010; Matsumoto et al., 2014; Chen et al., 2014; Isse et al., 2005). In this study, we found that the ALDH2 knockout contributes to higher sera acetaldehyde accumulation after exposure to 5% alcohol (Figure 1B), although they showed the same sera ethanol level, similar results were observed in mice that the loss of ALDH2 leads to higher concentration of acetaldehyde in blood and multiple organs (Isse et al., 2005). These data indicated that ALDH2 knockout causes a significant decrease in acetaldehyde scavenging capacity, resulting in evaluation of acetaldehyde level after drinking of alcohol (Chen et al., 2009). Although most alcohol is metabolized and converts into acetaldehyde in hepatocytes, the intestinal mucosa also expressed enzymes and contributed to oxidative of metabolism of alcohol (Chen et al., 2015). Besides, evidence has showed that alcohol is not directly metabolized by gut microbiota (Martino et al., 2022), but alcohol exposure can alter gut environment and microbiota structure and function. The significantly reduced Shannon indexes in KO-5% when compared to the WT-5% mice found in the current study are consistent with previous study performed on binge-on-chronic alcohol consumption mice (Shen et al., 2024), and heavy drinkers (Gurwara et al., 2020). In addition, the microbial community composition of KO-5% mice had the most significant dissimilarity from other groups. Therefore, this current study revealed that the alcohol exposure altered gut microbiota biodiversity in ALDH2 knockout mice.
The KO-5% mice had the highest relative abundance of phylum Proteobacteria, while Firmicutes decreased significantly. Prior studies have found that alcohol drinking altered gut microbiota structure, with most of the data focusing on Firmicutes or Bacteroidetes, although the results were inconsistent (Lee et al., 2020; Peterson et al., 2017). Proteobacteria seem to have important role in inflammatory active and they were found with higher abundance in alcohol overconsumers. Moreover, many members of phylum Proteobacteria, including Escherichia have been associated with inflammation (Janket et al., 2020). In this study, the abundance of Ralastonia and Stenotrophomonas were obviously higher in KO-5% mice, suggesting that the inflammatory microbiota of ALDH2 knockout mice may be increased. The genus Lactobacillus belongs to phylum Firmicutes, is a well-known probiotic with multiple health-promoting effects. Jung and colleagues (Jung et al., 2021) revealed that the Lactobacilli species as the indigenous flora had a potential to downregulate the alcohol and acetaldehyde levels, and whose effects depend on host ALDH2 genotype. Increased abundance of Lactobacilli can alleviate the ill effects of alcohol consumption (Kim et al., 2022). Notably, a significantly reduced Lactobacilli was observed in ALDH2 knockout mice, which perturbed intestinal bile acids metabolism and linked to sera proinflammatory cytokines such as IL-1β, IL-6, TNF-α, and C-reactive protein (Li et al., 2023). In line with these findings, we found the significantly decreased abundance of Lactobacilli as well as higher mRNA levels of TNF-α, IL-1β, and IL-6 in KO-5% mice when compared to WT-5% individuals, which suggested that Lactobacilli played critical role in the ALDH2 deficiency mice through the crosstalk with proinflammatory mediators. Based on prediction of functional pathways for gut microbiota, we found that several pathways, such as peroxisome, primary immunodeficiency, fatty acid degradation, alpha-linolenic acid metabolism, pantothenate and CoA biosynthesis, were more enriched in WT-5% mice. Of which, peroxisomes were closely related to lipid metabolism and reactive oxygen species turnover, and which were also required to innate immunity pathways (Cara et al., 2017). Furthermore, alpha-linolenic acid metabolism has been proven to alleviate inflammatory by restraining the NF-κB signaling pathway (Yang et al., 2023). These results suggested that the ALDH2 played critical roles on the metabolic homeostasis.
Limitations of the current study are manifold. In our model setting we tested the sera ethanol concentration, serum acetaldehyde level, inflammatory cytokines, as well as intestinal flora at the end of the chronic alcohol exposure. This failed to present the systemic physiological changes over time, especially for the temporal dynamics of microbial communities. Furthermore, all mice enrolled in the study were selected based on their gender, age, body size and weight before our model setting, without prior characterization of the gut microbiota, which may be the causality that WT-Control mice had a relative diverse microbiome composition. Additionally, the intestinal barrier proteins such as ZO-1, Occludin, and Claudin are proven to be corrected with the dysbiosis of the intestinal microbiome (McMahan et al., 2023). It is critical to collected the corresponding tissues to determine whether gut microbiome dysbiosis corresponds to morphological damage.
5 Conclusion
This study demonstrated the effect of ALDH2 on alcohol mediated gut microbiota dysbiosis upon chronic exposure to 5% alcohol. Host ALDH2 knockout resulted in intestinal inflammation as well as gut dysbiosis, especially the increase of Proteobacteria, Stenotrophomonas, and Ralstonia proportion, while decrease of Firmicutes, Lactobacillus, unclassfied Bacilli, Vibrio, and Pseudoalteromonas proportion. Based on these findings, intestinal microbiota is closely connected to host ALDH2 gene upon chronic alcohol exposure.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found at: https://www.ncbi.nlm.nih.gov/, PRJNA1245522.
Ethics statement
The animal study was approved by Animal Ethics Committee of West China Hospital of Sichuan University. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
XT: Formal analysis, Project administration, Methodology, Writing – original draft, Investigation, Software. BW: Writing – original draft, Methodology, Data curation, Investigation. XW: Validation, Software, Writing – review & editing. XX: Conceptualization, Supervision, Funding acquisition, Formal analysis, Resources, Writing – review & editing. YL: Validation, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Natural Science Foundation of China (grant no. 82472580), and Young Scientists Fund of the Natural Science Foundation of Sichuan (grant no. 2024NSFSC1505).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Abbas, D., Ciricillo, J. A., Elom, H. A., and Moon, A. M. (2023). Extrahepatic health effects of alcohol use and alcohol-associated liver disease. Clin. Ther. 45, 1201–1211. doi: 10.1016/j.clinthera.2023.08.018
Alcohol Research: Current Reviews Editorial Staff (2018). Drinking patterns and their definitions. Alcohol Res. 39, 17–18
Amir, A., McDonald, D., Navas-Molina, J. A., Kopylova, E., Morton, J. T., Zech Xu, Z., et al. (2017). Deblur rapidly resolves single-nucleotide community sequence patterns. mSystems 2, e00191–e00116. doi: 10.1128/mSystems.00191-16
Axley, P. D., Richardson, C. T., and Singal, A. K. (2019). Epidemiology of alcohol consumption and societal burden of alcoholism and alcoholic liver disease. Clin. Liver Dis. 23, 39–50. doi: 10.1016/j.cld.2018.09.011
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Butts, M., Sundaram, V. L., Murughiyan, U., Borthakur, A., and Singh, S. (2023). The influence of alcohol consumption on intestinal nutrient absorption: a comprehensive review. Nutrients 15:1571. doi: 10.3390/nu15071571
Cara, D., Francesca, A. S., Braverman, N. E., Rachubinski, R. A., and Simmonds, A. J. (2017). Peroxisome-mediated metabolism is required for immune response to microbial infection. Immunity 47, 93–106.e7. doi: 10.1016/j.immuni.2017.06.016
Cederbaum, A. I. (2012). Alcohol metabolism. Clin. Liver Dis. 16, 667–685. doi: 10.1016/j.cld.2012.08.002
Chen, C. H., Ferreira, J. C., Gross, E. R., and Mochly-Rosen, D. (2014). Targeting aldehyde dehydrogenase 2: new therapeutic opportunities. Physiol. Rev. 94, 1–34. doi: 10.1152/physrev.00017.2013
Chen, C. H., Ferreira, J. C. B., Joshi, A. U., Stevens, M. C., Li, S. J., Hsu, J. H. M., et al. (2020). Novel and prevalent non-east Asian ALDH2 variants; implications for global susceptibility to aldehydes' toxicity. EBioMedicine 55:102753. doi: 10.1016/j.ebiom.2020.102753
Chen, C. H., Ferreira, J. C. B., and Mochly-Rosen, D. (2019). ALDH2 and cardiovascular disease. Adv. Exp. Med. Biol. 1193, 53–67. doi: 10.1007/978-981-13-6260-6_3
Chen, Y., Liu, H., Yu, Z., Yang, Y., Huang, Q., Deng, C., et al. (2022). ALDH2 polymorphism rs671 *1/*2 genotype is a risk factor for the development of alcoholic liver cirrhosis in Hakka alcoholics. Int. J. Gene. Med. 15, 4067–4077. doi: 10.2147/ijgm.S356761
Chen, P., Miyamoto, Y., Mazagova, M., Lee, K. C., Eckmann, L., and Schnabl, B. (2015). Microbiota protects mice against acute alcohol-induced liver injury. Alcohol. Clin. Exp. Res. 39, 2313–2323. doi: 10.1111/acer.12900
Chen, Y. C., Peng, G. S., Tsao, T. P., Wang, M. F., Lu, R. B., and Yin, S. J. (2009). Pharmacokinetic and pharmacodynamic basis for overcoming acetaldehyde-induced adverse reaction in Asian alcoholics, heterozygous for the variant ALDH2*2 gene allele. Pharmacogenet. Genomics 19, 588–599. doi: 10.1097/FPC.0b013e32832ecf2e
Chen, Y., Yang, F., Lu, H., Wang, B., Lei, D., Wang, Y., et al. (2011). Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 54, 562–572. doi: 10.1002/hep.24423
Davenport, E. R. (2016). Elucidating the role of the host genome in shaping microbiome composition. Gut Microbes 7, 178–184. doi: 10.1080/19490976.2016.1155022
Douglas, G. M., Maffei, V. J., Zaneveld, J. R., Yurgel, S. N., Brown, J. R., Taylor, C. M., et al. (2020). PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 38, 685–688. doi: 10.1038/s41587-020-0548-6
Edenberg, H. J. (2007). The genetics of alcohol metabolism: role of alcohol dehydrogenase and aldehyde dehydrogenase variants. Alcohol Res. Health 30, 5–13.
Engen, P. A., Green, S. J., Voigt, R. M., Forsyth, C. B., and Keshavarzian, A. (2015). The gastrointestinal microbiome: alcohol effects on the composition of intestinal microbiota. Alcohol Res. 37, 223–236. doi: 10.1016/0306-4565(86)90035-5
Foster, R. K., and Marriott, H. E. (2006). Alcohol consumption in the new millennium–weighing up the risks and benefits for our health. Nutr. Bull. 31, 286–331. doi: 10.1111/j.1467-3010.2006.00588.x
Goodrich, J. K., Davenport, E. R., Clark, A. G., and Ley, R. E. (2017). The relationship between the human genome and microbiome comes into view. Annu. Rev. Genet. 51, 413–433. doi: 10.1146/annurev-genet-110711-155532
Gurwara, S., Dai, A., Ajami, N. J., Graham, D. Y., White, D. L., Chen, L., et al. (2020). Alcohol use alters the colonic mucosa-associated gut microbiota in humans. Nutr. Res. 83, 119–128. doi: 10.1016/j.nutres.2020.09.004
Isse, T., Matsuno, K., Oyama, T., Kitagawa, K., and Kawamoto, T. (2005). Aldehyde dehydrogenase 2 gene targeting mouse lacking enzyme activity shows high acetaldehyde level in blood, brain, and liver after ethanol gavages. Alcohol. Clin. Exp. Res. 29, 1959–1964. doi: 10.1097/01.alc.0000187161.07820.21
Janket, S. J., Ackerson, L. K., and Diamandis, E. P. (2020). Drug-resistant bacteremia after fecal microbiota transplant. N. Engl. J. Med. 382, 1960–1962. doi: 10.1056/NEJMc2002496
Jung, S. J., Hwang, J. H., Park, E. O., Lee, S. O., Chung, Y. J., Chung, M. J., et al. (2021). Regulation of alcohol and acetaldehyde metabolism by a mixture of Lactobacillus and Bifidobacterium species in human. Nutrients 13:1875. doi: 10.3390/nu13061875
Kim, J., Ahn, S. W., Kim, J. Y., Whon, T. W., Lim, S. K., Ryu, B. H., et al. (2022). Probiotic lactobacilli ameliorate alcohol-induced hepatic damage via gut microbial alteration. Front. Microbiol. 13:869250. doi: 10.3389/fmicb.2022.869250
Kwon, H. J., Won, Y. S., Park, O., Chang, B., Duryee, M. J., Thiele, G. E., et al. (2014). Aldehyde dehydrogenase 2 deficiency ameliorates alcoholic fatty liver but worsens liver inflammation and fibrosis in mice. Hepatology 60, 146–157. doi: 10.1002/hep.27036
Leclercq, S., Le Roy, T., Furgiuele, S., Coste, V., Bindels, L. B., Leyrolle, Q., et al. (2020). Gut microbiota-induced changes in β-Hydroxybutyrate metabolism are linked to altered sociability and depression in alcohol use disorder. Cell Rep. 33:108238. doi: 10.1016/j.celrep.2020.108238
Lee, J. E., Ha, J. S., Park, H. Y., and Lee, E. (2020). Alteration of gut microbiota composition by short-term low-dose alcohol intake is restored by fermented rice liquor in mice. Food Res. Int. 128:108800. doi: 10.1016/j.foodres.2019.108800
Li, Z.-M., Kong, C.-Y., Mao, Y.-Q., Chen, H.-L., Zhang, S.-L., Huang, J.-T., et al. (2023). Host ALDH2 deficiency aggravates nonalcoholic steatohepatitis through gut-liver axis. Pharmacol. Res. 196:106902. doi: 10.1016/j.phrs.2023.106902
Liu, Y., Wu, Z., Zhang, Y., Chen, B., Yu, S., Li, W., et al. (2022). Alcohol-dependent downregulation of apolipoprotein H exacerbates fatty liver and gut microbiota dysbiosis in mice. Lipids Health Dis. 21:89. doi: 10.1186/s12944-022-01699-7
Ma, H., Yu, L., Byra, E. A., Hu, N., Kitagawa, K., Nakayama, K. I., et al. (2010). Aldehyde dehydrogenase 2 knockout accentuates ethanol-induced cardiac depression: role of protein phosphatases. J. Mol. Cell. Cardiol. 49, 322–329. doi: 10.1016/j.yjmcc.2010.03.017
Mallott, E. K., and Amato, K. R. (2021). Host specificity of the gut microbiome. Nat. Rev. Microbiol. 19, 639–653. doi: 10.1038/s41579-021-00562-3
Martino, C., Zaramela, L. S., Gao, B., Embree, M., Tarasova, J., Parker, S. J., et al. (2022). Acetate reprograms gut microbiota during alcohol consumption. Nat. Commun. 13:4630. doi: 10.1038/s41467-022-31973-2
Massey, V. L., and Arteel, G. E. (2012). Acute alcohol-induced liver injury. Front. Physiol. 3:193. doi: 10.3389/fphys.2012.00193
Matsumoto, A., Vasiliou, V., Kawamoto, T., Tanaka, K., and Ichiba, M. (2014). Ethanol reduces lifespan, body weight, and serum alanine aminotransferase level of aldehyde dehydrogenase 2 knockout mouse. Alcohol. Clin. Exp. Res. 38, 1883–1893. doi: 10.1111/acer.12462
McMahan, R. H., Hulsebus, H. J., Najarro, K. M., Giesy, L. E., Frank, D. N., and Kovacs, E. J. (2023). Changes in gut microbiome correlate with intestinal barrier dysfunction and inflammation following a 3-day ethanol exposure in aged mice. Alcohol 107, 136–143. doi: 10.1016/j.alcohol.2022.08.011
Moreau, R., Cailmail, S., Valla, D., and Lebrec, D. (1997). Use of measurements of ethanol absorption from stomach and intestine to assess human ethanol metabolism. Am. J. Physiol. 11, 993–997. doi: 10.1046/j.1365-2036.1997.00238.x
Mutlu, E. A., Gillevet, P. M., Rangwala, H., Sikaroodi, M., Naqvi, A., Engen, P. A., et al. (2012). Colonic microbiome is altered in alcoholism. Am. J. Physiol. Gastrointest. Liver Physiol. 302, G966–G978. doi: 10.1152/ajpgi.00380.2011
Park, B., Lee, H. R., and Lee, Y. J. (2016). Alcoholic liver disease: focus on prodromal gut health. J. Dig. Dis. 17, 493–500. doi: 10.1111/1751-2980.12375
Peterson, V. L., Jury, N. J., Cabrera-Rubio, R., Draper, L. A., Crispie, F., Cotter, P. D., et al. (2017). Drunk bugs: chronic vapour alcohol exposure induces marked changes in the gut microbiome in mice. Behav. Brain Res. 323, 172–176. doi: 10.1016/j.bbr.2017.01.049
Piano, M. R., Marcus, G. M., Aycock, D. M., Buckman, J., Hwang, C.-L., Larsson, S. C., et al. (2025). Alcohol use and cardiovascular disease: a scientific statement from the American Heart Association. Circulation 152:e00, e7–e21. doi: 10.1161/CIR.0000000000001341
Qiu, Y., Xu, S., Chen, X., Wu, X., Zhou, Z., Zhang, J., et al. (2023). NAD+ exhaustion by CD38 upregulation contributes to blood pressure elevation and vascular damage in hypertension. Signal Transduct. Target. Ther. 8:353. doi: 10.1038/s41392-023-01577-3
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Rungratanawanich, W., Lin, Y., Wang, X., Kawamoto, T., Chidambaram, S. B., and Song, B. J. (2023). ALDH2 deficiency increases susceptibility to binge alcohol-induced gut leakiness, endotoxemia, and acute liver injury in mice through the gut-liver axis. Redox Biol. 59:102577. doi: 10.1016/j.redox.2022.102577
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L., Garrett, W. S., et al. (2011). Metagenomic biomarker discovery and explanation. Genome Biol. 12:R60. doi: 10.1186/gb-2011-12-6-r60
Shen, M., Zhao, H., Han, M., Su, L., Cui, X., Li, D., et al. (2024). Alcohol-induced gut microbiome dysbiosis enhances the colonization of Klebsiella pneumoniae on the mouse intestinal tract. mSystems 9:e0005224. doi: 10.1128/msystems.00052-24
Shimizu, C., Oki, Y., Mitani, Y., Tsuchiya, Y., and Nabeshima, T. (2016). Moderate-dose regular lifelong alcohol intake changes the intestinal flora, protects against aging, and keeps spatial memory in the senescence-accelerated mouse prone 8 (SAMP8) model. J. Pharm. Pharm. Sci. 19, 430–447. doi: 10.18433/j3990v
Sidhu, M., and van der Poorten, D. (2017). The gut microbiome. Aust. Fam. Physician 46, 206–211. doi: 10.3316/informit.741870194405168
Vassallo, G., Mirijello, A., Ferrulli, A., Antonelli, M., Landolfi, R., Gasbarrini, A., et al. (2015). Review article: alcohol and gut microbiota - the possible role of gut microbiota modulation in the treatment of alcoholic liver disease. Aliment. Pharmacol. Ther. 41, 917–927. doi: 10.1111/apt.13164
Wall, T. L., Horn, S. M., Johnson, M. L., Smith, T. L., and Carr, L. G. (2000). Hangover symptoms in Asian Americans with variations in the aldehyde dehydrogenase (ALDH2) gene. J. Stud. Alcohol 61, 13–17. doi: 10.15288/jsa.2000.61.13
Wang, X., Pan, L., Gu, J., Gu, L., Lou, M., and Liu, Y. (2024). Associations between gut microbiota and alcohol abuse: a Mendelian randomisation and bioinformatics study. J. Mol. Neurosci. 74:80. doi: 10.1007/s12031-024-02259-7
Way, G. W., Jackson, K. G., Muscu, S. R., and Zhou, H. (2022). Key signaling in alcohol-associated liver disease: the role of bile acids. Cells 11:1374. doi: 10.3390/cells11081374
Yang, J., Liu, S., Zhao, Q., Li, X., and Jiang, K. (2023). Gut microbiota-related metabolite alpha-linolenic acid mitigates intestinal inflammation induced by oral infection with toxoplasma gondii. Microbiome 11:273. doi: 10.1186/s40168-023-01681-0
Keywords: chronic alcohol exposure, mitochondrial acetaldehyde dehydrogenase 2 (ALDH2), acetaldehyde, ileum microbiota, 16S rRNA gene sequencing
Citation: Tan X, Wu B, Wen X, Li Y and Xu X (2025) Host ALDH2 deficiency aggravates acetaldehyde metabolism disturbance and gut microbiota dysbiosis in chronic alcohol exposure mice. Front. Microbiol. 16:1617673. doi: 10.3389/fmicb.2025.1617673
Edited by:
Huan Li, Lanzhou University, ChinaReviewed by:
Won Fen Wong, University of Malaya, MalaysiaYannick Rossez, UMR8576 Unité de Glycobiologie Structurale et Fonctionnelle (UGSF), France
Copyright © 2025 Tan, Wu, Wen, Li and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xuewen Xu, eHh3XzA4MjZAMTYzLmNvbQ==