 Igor Chebotar
Igor Chebotar Ilya Azizov
Ilya Azizov Mikhail Edelstein
Mikhail Edelstein Roman Kozlov
Roman Kozlov- Institute of Antimicrobial Chemotherapy, Smolensk State Medical University, Smolensk, Russia
Virulence is a property of bacteria that determines the degree of damage inflicted on humans. In modern medicine, there is an underestimation of the importance of testing the virulence of opportunistic pathogens to assess prognosis, optimize therapy, and evaluate the risk of developing probable complications of the infectious process. This review analyzes the basic characteristics of virulence, including multifactoriality, complex regulation, its relationship with fitness and bet-hedging in the context of choosing optimal methods for quantitative virulence testing. The possibilities of various methodological approaches for evaluation of virulence in clinical laboratory settings are considered. Current technology levels allow laboratories to be equipped with test systems for implementing methods to detect the virulence of clinical opportunistic isolates. At the hospital level, determining the virulence of isolates from individual patients will improve the prediction of the course of the infectious process and help to rationalize infection control based on principles similar to those used in combating antibiotic-resistant strains. Knowledge of virulence properties of relevant pathogen clones is required for the successful development of anti-virulence strategies.
Introduction
The global damage caused by bacterial infections results in a staggering loss of life. Each year, approximately 7.7 million people die from bacterial infections worldwide (bacteria-attributable deaths), over 3.7 million people die from respiratory infections and tuberculosis (2017) (Okeke et al., 2024; Naghavi et al., 2024). The dynamic symbiotic relationship between a microbe and its host during an infection is commonly described as an infectious process. The outcome of each infectious process depends on the interaction of three components: (1) the host with its immune defense, (2) the microbe, whose strategy is aimed at multiplying within the host organism while evading and manipulating immune effectors, (3) external medicinal interventions designed to suppress the microbe and protect the host. The microbe’s tactics in the context of infectious diseases often accompanied direct or indirect (immune-mediated) damaging the host tissues. To describe a microbe’s ability to cause harm to the host, the term virulence is employed. The concept of “virulent” began to be actively used in medieval medicine (from about 1380 A.D.) to describe ulcers that emitted poison (Lanfrank, 1894). Later, virulence came to denote toxicity and contagiousness (Simpson, 1665; Wynell, 1670), eventually becoming synonymous with plant toxicity and chemical compounds (Kircheri, 1665). Before Louis Pasteur’s revolutionary discovery, scientists believed that infections were caused by substances such as “morbid poisons,” that “arise from disease, and excite a secretion capable of reproducing the same disease” (Adams, 1807). Even then, to describe the degree of danger posed by these “morbid poisons,” the epithet “virulent” was applied, and attempts were made to destroy their “virulence” (of smallpox, gonorrheal matter) using available chemicals (“chlorine gas,” iodine, etc.), (Browne, 1830). Naturally, the term virulence became associated with newly discovered bacteria causing infections. As early as Pasteur (1881, 1882) widely used the term virulence in relation to pathogenic microbes, understanding it as the degree of danger. Both Pasteur (1881, 1882), Koch et al. (1884) used the term virulence, attributing quantitative characteristics to it. Koch et al. (1884), describing experiments to develop a vaccine against anthrax, confirmed the possibility of reducing the virulence of the “anthrax virus” through cultivation “under certain conditions” (Koch et al., 1884). Today, the meaning of the term virulence essentially corresponds to how it was understood by the pioneers of microbiology - the degree of the injury-producing potential or toxicity of a microorganism. During Koch (1884), Pasteur’s (1881, 1882) time, the criterion for assessing the virulence of a pathogen was its ability to cause disease in animals or humans. This property formed the basis of the classical Henle-Koch postulates, which work well for microbes classified as pathogens. Non-opportunistic pathogens are microorganisms that cause disease in practically healthy persons with high probability (definitions of key terms used in this review are summarized in Supplementary Table 1). For many non-opportunistic pathogens (Bacillus anthracis, Vibrio cholerae, Burkholderia mallei, etc.,) species identification alone is sufficient for accurately predicting their degree of danger. Sometimes, however, additional confirmation of virulence is required, such as evaluating the toxigenicity of a strain, as in the case of Corynebacterium diphtheriae. Overall, assessing virulence of non-opportunistic today does not raise significant questions. Standard protocols exist for such bacteria, allowing precise predictions of their danger and enabling effective control measures.
When dealing with opportunistic pathogens, the situation becomes more complicated. Virulence manifests itself in a dialectical interplay between the host and the microorganism(s). One perspective holds that microbial virulence should be viewed in the context of its interaction with the host (Casadevall and Pirofski, 2015). Indeed, immune effectors can influence virulence levels via up- or down-regulation (Zhou et al., 2023), as well as through mutation-driven processes leading to the emergence of clones with novel virulence profiles (Rossi et al., 2021). This confusion profoundly complicates our understanding of virulence mechanisms and hampers the development of effective evaluation methods. Following the principles of cognition outlined by René Descartes, the complexity of any phenomenon requires its analysis through decomposition into constituent components. In this review, we focus solely on attributes of virulence in opportunistic bacteria, specifically highlighting opportunities and benefits associated with their evaluation.
The material foundation of pathogen virulence lies in virulence factors. A virulence factor is a component of a pathogen that contributes to the progression of the infectious process. Deletion of the virulence factor may compromise virulence but not viability (Casadevall and Pirofski, 1999). Virulence factors can encompass modulins affecting host immune responses (Henderson et al., 1996). The cumulative virulence of a pathogen depends on the integral functioning of its virulence factors. Therefore, assessing virulence factors serves as a vital basis (alongside antibiotic resistance and host condition) for determining pathogen risk. Evaluating virulence in opportunistic pathogens involves numerous uncertainties. Key questions include (1) the specificity of opportunistic pathogens’ virulence manifestations depending on the form of the disease, (2) the feasibility of clinical prediction based on the assessment of the virulence of a specific isolate, (3) the epidemiological threat posed by strains with increased virulence, and (4) the value of information about virulent isolates for optimization of infection control. Answering these questions requires verifying data on the correlation between disease outcomes and virulence profiles of causative agents, as well as accumulating information on the distribution of different pathovars among opportunistic pathogens. To gather such information, species-specific methodologies for virulence evaluation must be defined. While this review does not provide ready-made detailed solutions, it discusses the theoretical foundation for developing virulence assessment methods and analyzes and systematizes the most relevant examples of determining the virulence of current opportunistic pathogens. Modern technological capabilities for detecting virulence using accessible, reproducible, and standardized methods will also be justified.
The main reasons for the complexity of assessing the virulence of opportunistic pathogens can be attributed to (1) difficulties in defining correlations between opportunistic bacteria’s virulence factors and the manifestation of the infectious process, (2) the multifactorial nature of virulence, (3) the complex and unclear regulation of virulence expression, and (4) challenges in choosing appropriate methods for assessing virulence.
Locus-specific virulence factors
Typically, major opportunistic bacteria exhibit diversity in loci and forms of infection, affecting various organs and systems and causing infections of varying severity, acuteness, and duration. This raises the question of whether different virulence factors play equally important roles in realizing pathogenicity across different loci. Such patterns have been observed in some species. For example, in Pseudomonas aeruginosa in chronic ankle wounds, T3SS, responsible for secreting contact toxins like ExoU, ExoS, ExoT, and ExoY, is inhibited (Karash et al., 2021). In cystic fibrosis, exoS(+)/exoU(+) virulotypes often cause chronic forms of cystic fibrosis pneumonia, whereas exoS(+)/exoU(−) isolates are responsible for early clinical lung alterations (Sarges et al., 2020; Savinova et al., 2020). Despite statistically proven correlations, attention should be paid to the absence of absolute rules governing the distribution of virulence genes among isolates obtained from different loci. All results showed numerous exceptions. Traditionally, it has been believed that uropathogenic Escherichia coli strains primarily carry specific adhesins, including type 1 fimbriae, P fimbriae, S fimbriae, F1C fimbriae, and afimbrial adhesin (Sarowska et al., 2019). It is considered that the primary attribute of E. coli strains causing neonatal meningitis is a set of factors, chiefly the K1 capsule, which protects them from immune effectors and is rarely found in uropathogenic strains (Sarowska et al., 2019). Overall, the information on the virulent features of extraintestinal pathogenic E. coli (ExPEC) is enormous and often contradictory. However, some conclusions can be drawn today. Firstly, there are no single molecular/genetic virulence factors that could be unique ExPEC indicators similar to such markers of virulence (toxigenicity) as diphtheria toxin in Corynebacterium diphtheriae or Shiga-toxin in Shigella dysenteriae (Johnson and Russo, 2018). Secondly, differences between ExPEC and commensal E. coli strains lie in complex combinations of true virulence genes and regulatory genes, possibly also in their expression patterns. Emphasizing this point, Russo and Johnson (2000) highlighted the c but rather “various combinations of adhesins (e.g., P and S fimbriae), iron-acquisition systems (e.g., aerobactin), host defense-avoidance mechanisms, and toxins (e.g., hemolysin), which collectively are now regarded as extraintestinal virulence factors.” Commensal strains typically lack specialized virulence traits found in ExPEC. Additionally, ExPEC and commensal E. coli differ in their accessory genomes. Genomic studies have shown the presence of horizontally acquired pathogenicity-associated islands more frequently observed in ExPEC than in commensals (Johnson and Russo, 2018). The more frequent association of ExPEC pathotypes with phylogroup B2 also indicates the specificity of their virulent ensembles (Hazen et al., 2023). Although some specific virulence factors such as adhesins, iron-acquisition systems, toxins, and other “can distinguish ExPEC from typical commensal E. coli, comparative genomics has revealed that this distinction is far from absolute” (Russo and Johnson, 2000).
For other opportunistic pathogens, the correlation between the presence of pathogenicity factors and preferred infection sites remains even less clear. While various intoxications attributed to Staphylococcus aureus are strictly linked to specific toxins, opinions regarding the locus-specificity of virulence factors in S. aureus infections are quite contradictory. In the study by Budzyńska et al. (2021), it was found that sea- and sei-toxin genes were more frequent among blood strains than in wound isolates. Although the difference was statistically significant, it did not support any absolute locus-specificity of toxin genes: the sea-gene was present in 6.0% of blood strains and 19.0% of wound strains, while the sei-gene was found in 46.0% of blood strains and 61.0% of wound strains. The exfoliative toxin eta gene was detected in 14% of wound strains and 13% of blood strains, the toxic shock toxin (tst) gene was found in 13% of wound strains and 9% of blood strains, and the Panton-Valentine leukocidin gene (lukSF-PV) was identified in 3% of wound strains and 1% of blood strains (Budzyńska et al., 2021; Dermota et al., 2024; Gu et al., 2024).
Probably in the future, a detailed deciphering of the mechanisms of pathogenesis and accumulation of information on correlations between the virulence profile of the pathogen and the variant of infection will help obtain a clearer picture of the locus-specificity of various virulence factors.
A practically significant question arises: is it necessary, when detecting virulence in clinical isolates, to pay attention to the phenomenon of locus-specificity and determine only those factors that have predictive significance for a particular type of infection? If we focus solely on the individual patient, it would suffice to identify only the prognostically significant factors for the locus of infection. However, if we think strategically from the perspective of infection control and surveillance tasks, it becomes essential to obtain information about the most critical virulence factors of each isolate. This necessity follows from the existence of cross-pathovars (Lindstedt et al., 2018), which may pose nosocomial epidemic threats. It is likely prudent to follow a strategic course and test all key factors that could yield strategically valuable information about the virulence profiles of pathogens.
Multifactorial nature of virulence in opportunistic pathogens and key virulence factors
Any pathogenic or opportunistic bacterium realizes its virulence during infection by engaging multiple factors. There are no exceptions. Even pathogens causing mono-intoxications require multiple tools to implement the infectious process. For example, typical diphtheria caused by Corynebacterium diphtheriae is determined by the specific diphtheria toxin. However, clinically evident diphtheria would be impossible without the initial stages of infection, including preliminary colonization of mucosal surfaces (or less commonly wound surfaces) by C. diphtheriae and bacterial survival under conditions of localized inflammation. To realize these initial stages, C. diphtheriae expresses a variety of factors, including diverse adhesins, iron uptake systems, cytotoxic proteins, apoptotic and inflammatory inducers (Rogers et al., 2011; Cerdeno-Tarraga et al., 2003; Weerasekera et al., 2019; Ott et al., 2022). According to the Virulence Factor Database (VFDB), the virulome of uropathogenic E. coli includes > 90 genes, Acinetobacter baumannii complex > 110 genes, P. aeruginosa > 260 genes, Klebsiella pneumoniae > 120 genes, S. aureus > 110 genes, and Enterococcus spp. > 40 genes (The Virulence Factor Database, 2025). The virulome is the set of genes that contribute to the virulence of a pathogen, regardless of the significance of each of these genes. Thus, every product of the virulome can contribute directly or indirectly to the overall virulent potential of the pathogen. However, the pathogenetic significance of various virulence factors is not the same. The concept of virulence genes includes three distinct types of genes including (1) true virulence genes, that are directly involved in causing disease, (2) virulence-associated genes that encode regulators/activators of true virulence genes, (3) virulent life-style genes that ensure viability in the host organism (Wassenaar and Gaastra, 2001). The large volume of information about the virulome does not guarantee an understanding of the significance of each virulence gene in the realization of the infectious process. This difficulty arises due to the inability to predict precisely the expression of each gene, which is determined by complex signals from the external environment and enigmatic regulatory mechanisms within the bacterial cell. Redundancy lies in the fact that identifying the entire virulome provides much information already known to the species and represented by constitutively expressed species-specific genes.
This suggests that it is rational only to identify pathogenetic significant factor, which can “enhance” the baseline natural level of virulence of the species. We refer to these as acquired virulence factors. These factors can include (1) chromosomal (non-plasmid-borne) factors that may be overproduced due to “up-regulation” of expression (functional increase either because of mutations in regulator genes), and (2) plasmid-borne factors (Figure 1). A striking example of a species-specific overexpressed factor is the hyperproduction of the alginate by P. aeruginosa isolates due to mutational inactivation of the mucA gene or resulting from up-regulation via activation of the transcriptional regulator σ22 by environmental signals (Boucher et al., 1997; Damron and Goldberg, 2012). Examples of plasmid-borne factors include siderophores aerobactin and salmochelin and the determinant of hypermucoidy (rmpA) in K. pneumoniae (Lam et al., 2018).
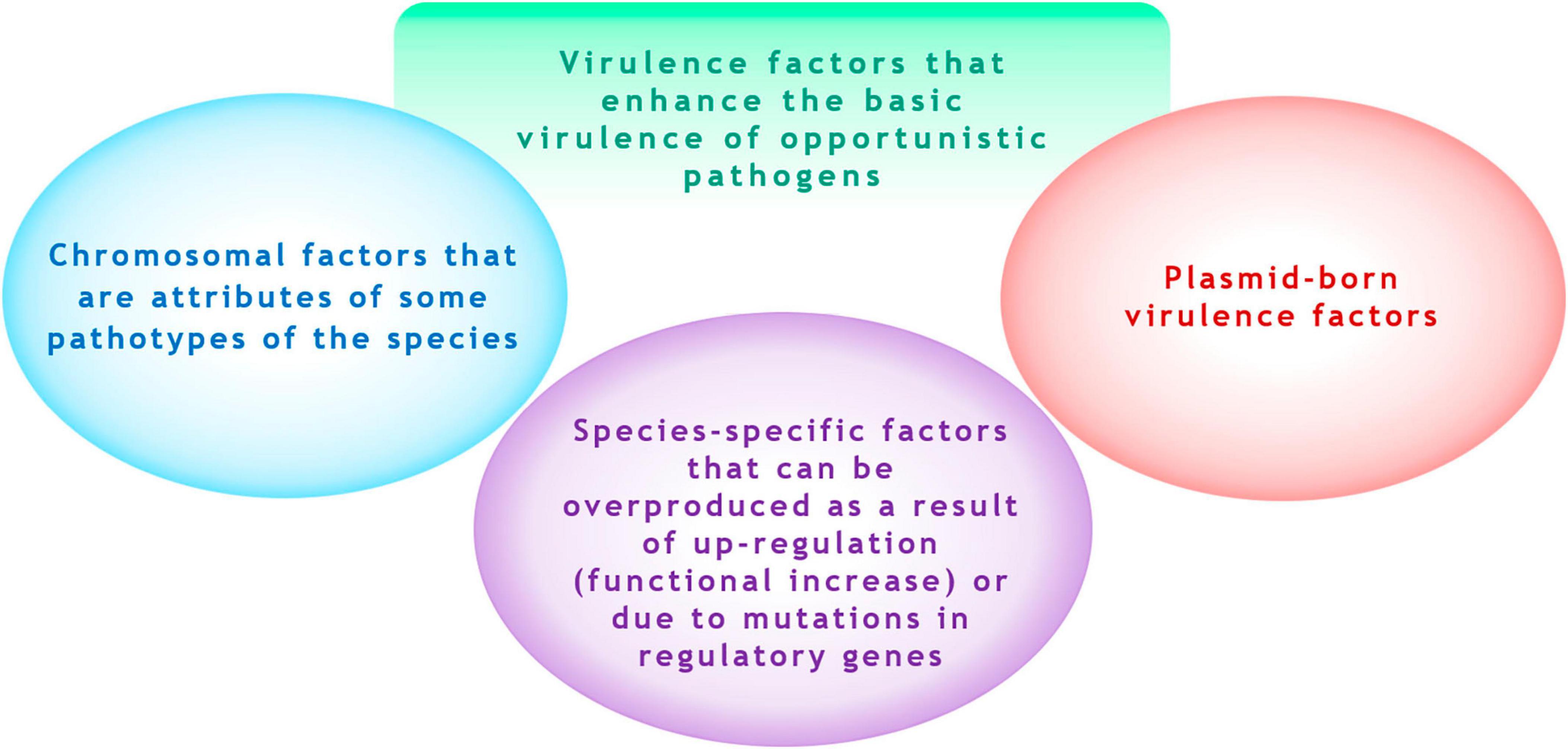
Figure 1. Virulence factors that enhance the basic virulence of opportunistic microorganisms.
The contribution of each acquired factor to the overall portrait of virulence is incomparable. To reduce the list of tested virulence factors, emphasis should be placed exclusively on key acquired factors. By key virulence factors, we mean virulence factors without which opportunistic pathogens become incapable of causing the disease or result in mild forms of the disease. The concept of the key virulence factor can include both factors encoded by a single gene and factors whose production may depend on a set of different genes. In other words, a key virulence factor must comply with Falkow (2004) rule, which states: “Specific inactivation of the gene(s) associated with the suspected virulence trait should lead to a measurable loss in pathogenicity or virulence… Reversion or allelic replacement of the mutated gene should lead to restoration of pathogenicity….” The most accurate interpretation of the mechanism of action of a virulence factor on host structures should additionally be validated in experiments using animal models knocked out of the virulence factor target gene.
One approach to evaluate virulence involves counting the number of pathogenicity factors in an isolate using phenotypic or genetic methods. One option for such counting is the virulence index, that is defined as the ratio between positive tests for virulence traits and the total amount of virulence traits evaluated and calculated for each isolate (Singh et al., 2017; Cunha et al., 2023). This method has been applied to identify virulence factors without validating their pathogenic significance, such as bacterial hydrophobicity (Cunha et al., 2023). Furthermore, verified examples demonstrating its reliability for predicting infection outcomes in humans are currently unavailable. Nonetheless, we should not overlook the underlying principle of this approach, grounded in quantitative virulence assessment, which could benefit from being complemented by ranking based on expression levels and significance.
Another measure of virulence is the virulence score, which is calculated as the sum of virulence factor genes in each isolate. It should be noted that the virulence score is not a standardized indicator. Different authors calculate it differently, even when analyzing the virulence of the same species. In studies on K. pneumoniae, some authors combine all genes involved in virulence into the virulence score (Maldonado et al., 2024), others count only siderophore genes (Lam et al., 2021), and still others add up genes encoding adhesins, capsules, iron uptake systems, invasins, and toxins (Hsiao et al., 2024). The popular Kleborate software calculates the virulence score for K. pneumoniae, considering virulence loci, including YbST (yersiniabactin), CbST (colibactin), AbST (aerobactin), SmST (salmochelin), and RmpA/RmpA2 (hypermucoidy) genes (Lam et al., 2021). The cited articles do not involve quantitative assessment of virulence factor production, which constitutes their limitation. Other disadvantages of using virulence scoring will be addressed later. Nevertheless, calculating both the virulence score and the virulence index represents reasonable efforts to prioritize the most important virulence factors.
Hypothetically, the existence of key factors can be determined for all nosocomial pathogens, including bacteria with poorly studied virulome. For example, the inactivation of T3SS with the AxoU toxin in Achromobacter spp., predicted merely as an ortholog of the P. aeruginosa ExoU toxin, leads to a reduction in the severity of infection and transforms acute forms into chronic ones (Tessmer et al., 2019; Pickrum et al., 2020). This indicates that the AxoU toxin is a key factor for Achromobacter spp. If key factors cannot be identified in a pathogenic bacterium, this does not imply their absence; rather, it reflects a lack of knowledge about their virulome and how they realize their virulent potential at different stages of infection.
Selecting key factors worth testing is challenging. Given the contradictions in correlations between locus-specificity of the infectious process and the presence of virulence factors in many pathogens (see above), the objects of testing should be key factors whose role in pathogenesis is undeniable. For major opportunists, key virulence factors are well-known. For example, in bloodstream infections, P. aeruginosa isolates actively expressing ExoU are more likely to result in poor outcomes regardless of their antibiotic susceptibility profile (El-Solh et al., 2012; Peña et al., 2015). In cases of septic shock, high mortality rates in P. aeruginosa have been associated with non-motile or pyocyanin hyperproducing isolates (Gupte et al., 2021).
We attempted to present promising acquired key virulence factors or their markers for testing in some opportunistic bacteria in Table 1. The selection of bacteria was based on their relevance, including opportunistic species belonging to the ESKAPE group of pathogens, except Enterobacter spp., as well as extraintestinal pathotypes of E. coli. The criteria for inclusion of virulence factors in the table were (1) confirmed in vivo (experimentally or based on clinical isolate research) pathogenetic significance, including cases of hyperproduction, and (2) variability (qualitative and/or quantitative) in factor production. The exclusion criteria included (1) the presence of the factor in the overwhelming majority of commensal/environmental isolates or clinical isolates from patients with mild, non-invasive, and non-recurrent forms of infection in the absence of hyperproduction in severe forms of infection, and (2) the lack of proper verification of the pathogenetic significance of the virulence factor or its overproduction, verified experimentally in vivo or based on clinical isolate research. The list of virulence factors presented in Table 1 is neither final nor fully verified. Only additional information derived from extensive virulence testing can enhance it.
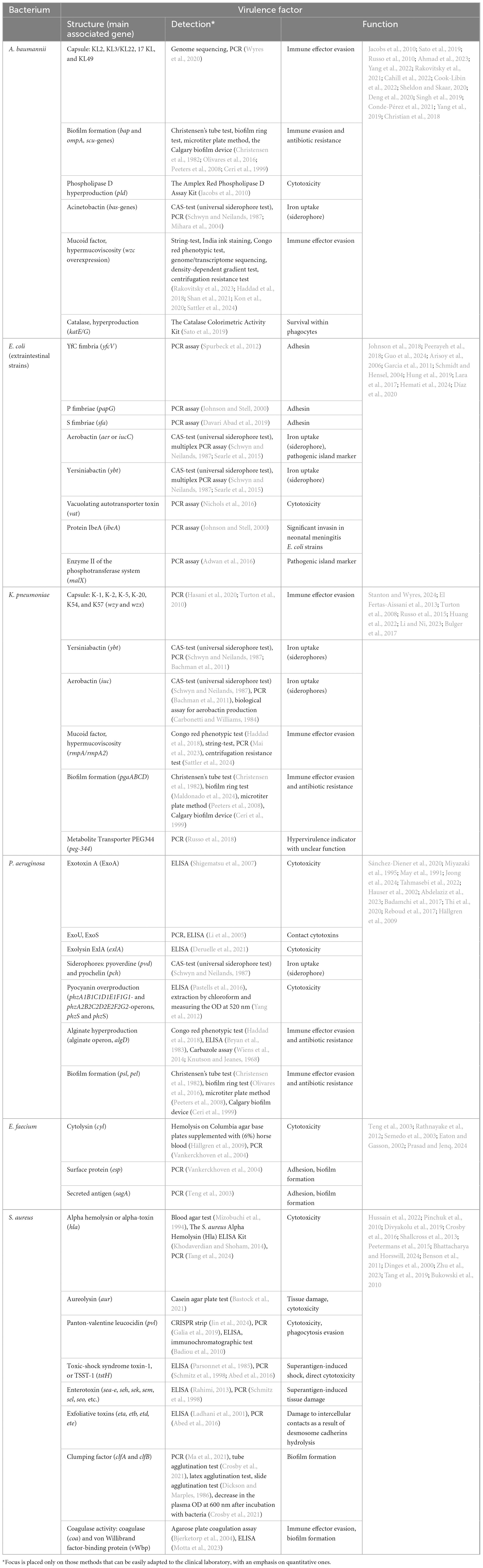
Table 1. Promising virulence factors or their markers for testing important opportunistic pathogens.
Regulatory mechanisms
Virulence factors are encoded by numerous genes, which are either scattered or organized into operons and clusters. The expression of virulence genes, post-translational assembly of their products, and in the case of some exotoxins - final activation inside the target cell - are complex processes controlled by global regulators and influenced by a variety of external conditions, including the microbial environment. Regulatory networks of virulence can vary significantly among different pathogens and have been extensively described in numerous reviews. While staying focused on the purpose of this review, we do not wish to repeat readily available summaries. However, we must emphasize the diverse ways in which external environmental factors influence virulence. For instance, biofilm formation intensity, which can be one manifestation of virulence, is highly sensitive to changes in cultivation conditions. Figure 2 demonstrates examples of how environmental factors affect biofilm formation in various opportunistic bacteria. Figure 2 does not claim to enumerate all possible scenarios but instead illustrates the divergent consequences of the same factors. Optimal Fe3+concentration for K. pneumoniae biofilm formation is approximately 50 μM, lowering Fe3+ concentrations significantly reduce biofilm formation (Chen et al., 2020). Optimal Fe3+ concentration for Burkholderia cenocepacia biofilm formation is roughly 100 μM, decreasing Fe3+ concentration reduces biofilm formation (Berlutti et al., 2005). Optimal Fe3+ concentration for A. baumannii biofilm formation is around 20 μM; increasing Fe3+ concentration drastically reduces biofilm formation (Modarresi et al., 2015). Increase in temperature enhances P. aeruginosa biofilm formation at 20 °C and 42 °C, with reduced formation between 25 °C and 30 °C (Grudniak et al., 2018; Kim et al., 2020). Conversely, E. faecium biofilm formation peaks at 28 °C compared to 37 °C (Maurya et al., 2021). The optimal pH for S. aureus biofilm formation is 7.0, deviations upward or downward reduce biofilm formation (Zmantar et al., 2010). Optimal pH for biofilm formation in Streptococcus agalactiae is 6.5, raising pH to 7.2 or lowering it to 5.5–4.2 diminishes biofilm formation (Miranda et al., 2018). Optimal pH for Streptococcus intermedius biofilm formation is pH 5.7, elevating pH to 6.5 significantly curtails biofilm formation (Ahmed et al., 2008). Addition of 0.2% glucose to growth media suppresses E. coli biofilm formation (Jackson et al., 2002). Meanwhile, Staphylococcus epidermidis biofilm formation increases with elevated glucose concentrations up to 1.5%, concentrations reaching 2.0% do not suppress biofilm formation (Rachid et al., 2000).
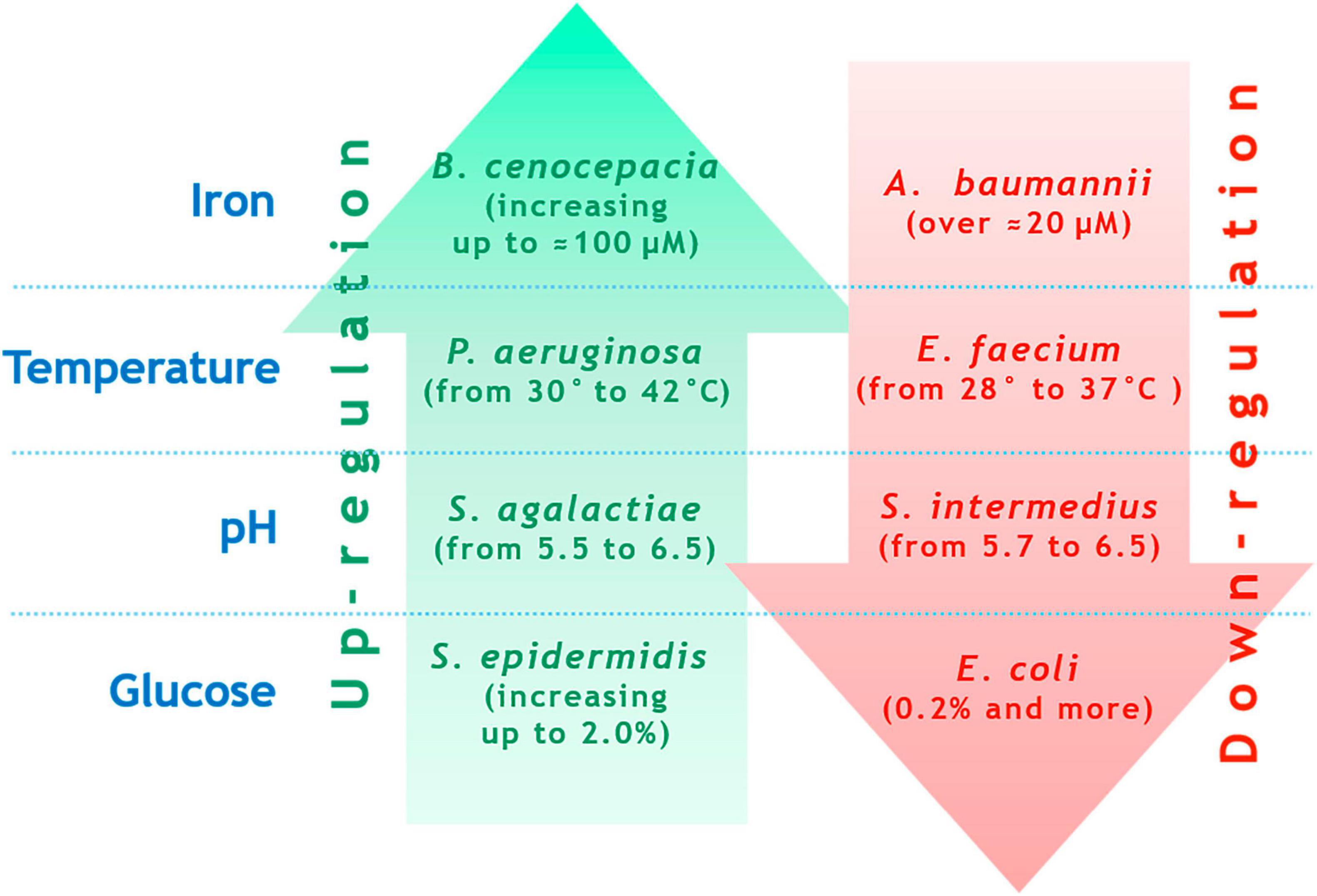
Figure 2. Examples of contrasting biofilm formation regulations for different species under changing external conditions (explanation is in the text).
Biofilm formation is a function of the environment. It can be inferred that this rule applies to other virulence factors as well. For example, decreasing the level of environmental oxygen enhances the production of exotoxin A but reduces pyoverdin synthesis (Gaines et al., 2007). Hemolytic activity in S. aureus is significantly inhibited at 34 °C but realized at 37 °C (Bastock et al., 2021). Conversely, aureolysin production peaks at 34 °C and decreases by approximately 30% at 37 °C (Bastock et al., 2021).
These examples highlight the need for careful selection of standard conditions for phenotypic/biochemical assessments of virulence, ensuring that the production of virulence factors is not inhibited.
Similarities, differences, and interference between the phenomena of virulence and antibiotic resistance
The main characteristics of antimicrobial resistance (AMR) and virulence have a lot in common (Table 2). They negatively impact the course and outcome of disease, can manifest as either constitutive (species-specific) traits or acquired features, and possess analogous principles of genetic regulation and dissemination mechanisms. Numerous instances have been described where enhanced virulence correlates positively with antimicrobial resistance. Genes coding for virulence factors and antibiotic resistance can be associated (Shah et al., 2019), including being co-localized within the same plasmid under unified control (Dong et al., 2022). Nonetheless, despite these commonalities, assessing resistance cannot substitute for evaluating virulence. This distinction arises because combinations of resistant and virulent properties in individual isolates exhibit considerable diversity, which manifests as classic interference in the physical sense of the term. The emergence of antibiotic resistance in a bacterial clone may coincide with decreased virulence, heightened virulence, or have no effect on virulence. The existence of all these variations has been illustrated in experiments involving uniform virulence evaluations of colistin-resistant A. baumannii strains. Colistin-resistant mutants with defects in the lpx gene, leading to complete loss of the polymyxin target lipopolysaccharide (LPS), exhibited diminished virulence in mouse and Caenorhabditis elegans experiments (Beceiro et al., 2014). In another study, colistin-resistant A. baumannii strains harboring mutations in pmrB (insertion of 30 nt or SNP 704C > T) did not diminish virulence toward C. elegans (Wand et al., 2015). An example of increased virulence in colistin-resistant A. baumannii, also confirmed using the C. elegans model, was associated with a mutation in the pmrC homolog eptA, a point mutation in ISAba1 upstream of eptA, and elevated eptA expression (Gerson et al., 2019). Presumably, the outcome of interference between AMR and virulence depends on (1) the chromosomal background of the isolate (its clonal lineage, inherent metabolic profile, baseline arsenal of virulence factors, etc.), (2) specific combinations of resistance and virulence determinants unique to every case, which may be dispersed or clustered as plasmid-borne genes, as well as mutations in chromosomal genes, (3) compensatory mutations that neutralize negative effects on the bacterium (disrupted functions) and partially or fully restore lost capacities arising from acquiring genetic determinants of virulence and resistance (Bæk et al., 2015). An important conclusion from this section is that the global properties of opportunistic pathogens’ virulence closely resemble those of AMR.
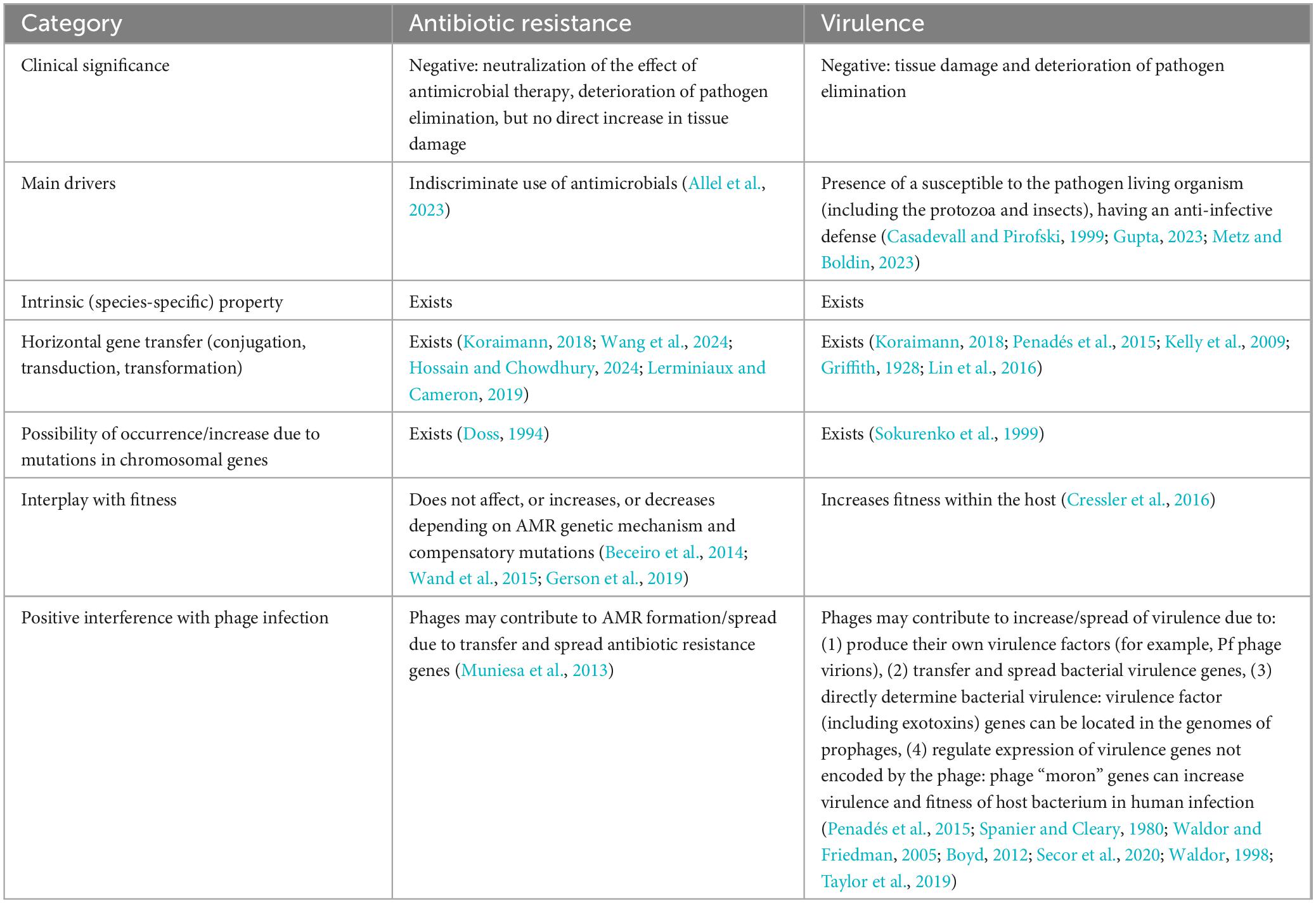
Table 2. Similarities and differences between antibiotic resistance and virulence.
Is the evaluation of virulence and fitness equivalent?
When addressing the issue of bacterial fitness, we seek to understand whether the assessment of bacterial fitness is entirely identical to the evaluation of virulence. The roots of applying the term “fitness” in microbiology can be traced back to the work of Novick and Szilard (1950), wherein the authors modeled spontaneous mutagenesis and used the verb “to fit” (meaning “to correspond, to adapt”) in reference to mutants that evolved by acquiring properties better suited to changes in the nutrient medium than those of their ancestors (Novick and Szilard, 1950). Currently, there are several definitions of the concept of “fitness.” In a broader sense, it refers to the evolutionary success of an organism (Botelho et al., 2019). A more detailed interpretation describes fitness as the ability of a microbe to reproduce in a competitive environment under specific conditions dictated by physicochemical parameters of the microenvironment, nutrient availability, and the presence of antibiotics and other antimicrobial factors. This distinguishes fitness from virulence, which reflects the ability to inflict harm on the host organism. Numerous fitness evaluation methods often use reproduction times or replication speed under defined conditions as a basis (Yasui, 2022). The array of fitness assessment techniques is illustrated in Figure 3. In vitro methods, excluding cytotoxicity in cell cultures, rely on quantitatively determining bacterial survival and multiplication and do not entail evaluating the destructive potential of the pathogen, i.e., virulence. Methods for assessing fitness in animal models and cell culture systems may account for the integrative destructive impact on the object without specifying the activity of individual virulence factors. Additionally, animal models can characterize an essential element of the infectious process dependent on pathogen virulence - the ability to replicate within a living organism. Analyzing fitness using data related to infections is based on indirect integral outcomes. For instance, in the work of Boral et al. (2023), conclusions about fitness levels are drawn from analyzing disease outcomes. Although indicators such as proteolytic and lipolytic enzyme activities, efflux, iron exchange, toxin production, biofilm formation, or gene expression may sometimes correlate with fitness, they should not be equated with it completely, as this contradicts the definition of “fitness” outlined earlier. Instead, they can serve as good indicators of virulence.
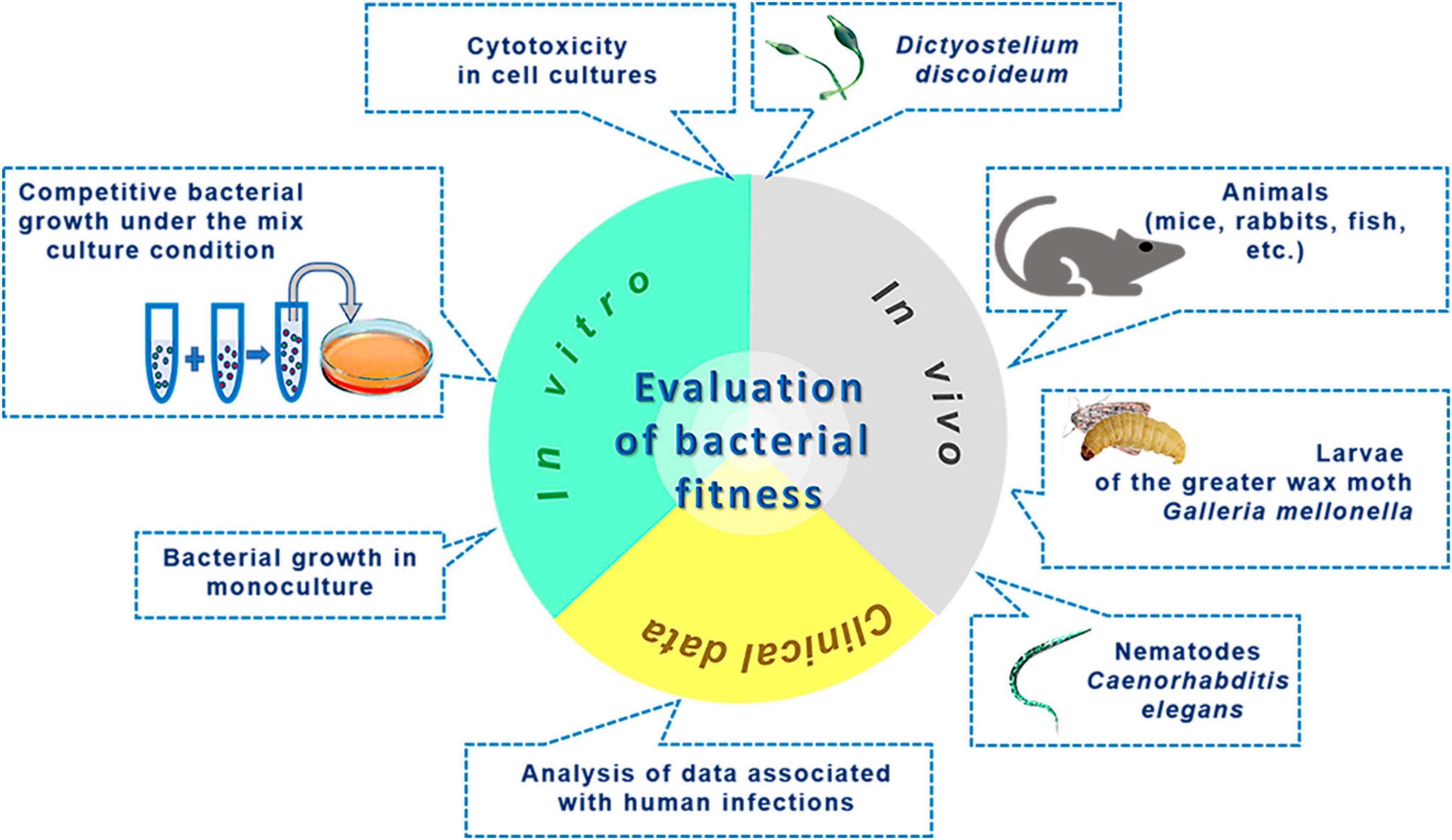
Figure 3. Methods to investigate microbial fitness.
Summarizing the content of this section, we conclude that the evaluation of fitness under in vitro conditions is not equivalent to the evaluation of virulence since it does not aim to determine the harm that the pathogen can inflict on the host. Meanwhile, studying fitness in in vivo models and cell cultures can yield useful insights into virulence, including the pivotal element of pathogenesis - the ability to multiply within live systems.
Virulence in the context of bet-hedging
When discussing methodology choices for assessing virulence, it is essential to consider the bet-hedging strategy. Bet-hedging implies phenotypic and genetic heterogeneity within populations of microbial cells where different variants are adapted to different conditions (Veening et al., 2008). The aim of the bet-hedging is to increase fitness in temporally variable environment. In practical terms, this means that grown on artificial media subpopulations of the pathogen with the highest virulence may be suppressed. They may only be activated upon changes in the surrounding environment, such as invasion into the host organism (Byrne et al., 2025; Spratt and Lane, 2022). It stands to reason that upon invading the host, an opportunistic pathogen experiences stress due to pressure from the immune system. Under these circumstances, the microbe’s lifestyle undergoes a dramatic shift. Only those bacteria whose metabolism can sustain the infection process using virulence factors to achieve this, gain an advantage. Mechanisms of bet-hedging may vary: they occur through (1) shifts in dominant subpopulations within an initially heterogeneous population (Nuss et al., 2016) and/or (2) switching gene expression (Stewart and Cookson, 2012). Regardless of the mechanisms, under infection conditions the more virulent microbial population predominates. Therefore, the bacterial population within a patient may therefore differ substantially in its properties, including virulence, from populations grown on artificial media. This premise was proven in classical pneumococcal experiments dating back to the 19th century. Clinical and attenuated strains grown on artificial media increased their virulence within mice (Kruse and Pansini, 1892; Eyre and Washbourn, 1899; Wadsworth and Kirkbride, 1918). Had these researchers relied solely on the virulence assessment of pneumococci directly from artificial media to forecast their clinical hazard, they would have committed grave errors.
From these observations emerges a vital insight, resonant with the conclusion drawn earlier: when designing systems for cultivating bacteria to evaluate their virulence phenotypically, conditions should not be optimized for the best growth and proliferation of a species. Instead, conditions fostering maximal production of virulence factors should be prioritized.
Methods for virulence evaluation
In vivo evaluation is the gold-standard method for testing virulence, reflecting the integrated interaction between the host and the microbe. The significance of each virulence factor should ideally be confirmed through animal studies and clinical observations. Exceptions may only be made for close orthologs of already tested factors in closely related species or intraspecific variants. Unfortunately, in vivo virulence evaluation is not possible in routine clinical practice, which requires alternative testing methods.
While identifying individual key virulence factors does not equate to the comprehensive in vivo evaluation of virulence, the combination of these key factors (the “virulence puzzle”) can correlate with in vivo assessments. This assertion is supported by various experimental findings (Pimenta et al., 2006; Wikraiphat et al., 2009; Lavender et al., 2004; Williamson et al., 2014).
For virulence evaluation methods to be effective as monitoring tools, they must meet the following requirements:
1. Be technologically and financially accessible to all laboratories performing routine clinical microbiology tests.
2. Provide results within a reasonably short timeframe, ideally not exceeding the duration of phenotypic AMR testing.
3. Yield consistently reproducible results on reference strains.
4. Accurately and linearly measure acceptable quantitative fluctuations in the activity of specific virulence factors that are key factors to the tested species.
5. Have optimal sensitivity and specificity in detecting virulence factor.
6. Generate digitized results suitable for statistical analysis.
7. Employ a comprehensive approach, encompassing the determination of a set of virulence factors for each species, where the cumulative index or score correlates with the level of virulence verified in vivo or based on clinical data.
8. Ensure safety by avoiding the use of live organisms, such as laboratory animals.
At first glance, whole genome sequencing (WGS) appears ideal for virulence assessment (Wolff et al., 2021). However, its application as a monitoring tool faces challenges. First, the ambiguous role of numerous genes and mutations in virulence realization complicates interpretation. Second, the complexity, expense, and prolonged execution render WGS inaccessible to many laboratories. Although sequencing cost has decreased by a factor of 1,000 over the past 20 years, it still remains too high for routine use in clinical laboratory practices. The cost of raw data (“reads”) of whole genome sequencing (WGS) varies depending on local conditions and study volume, ranging from US$40 to US$100 per bacterial genome. The minimum time required from DNA isolation to obtaining raw sequencing data (assuming automatic library preparation and maximum sequencer loading) is at least 3 days. Another reason hindering personalized utilization of WGS in virulence assessment is the imperfection of bioinformatic interpretation of virulome data. We are confident that in the future, accessible sequencing technologies and advanced automated bioinformatic tools will become available, enabling reliable information retrieval on the virulence of each clinical isolate. Currently, practical feasibility dictates focusing on identifying a limited number of virulence factors with proven pathogenic relevance, capable of enhancing the baseline intrinsic virulence level inherent to members of a given species.
Polymerase chain reaction (PCR)-based methods excel in detecting individual resistance genes, as shown in Table 1. However, their inability to quantify virulence factor production limits their utility for virulence detection. Similarly, newer pathogenicity testing methods based on CRISPR strips (Jin et al., 2024) suffer from the same limitations as PCR.
Proteomic techniques, such as mass spectrometry, hold great promise for future applications in virulence assessment (Man et al., 2021; Flores-Trevino et al., 2019). Specifically, encouraging results have been achieved using MALDI-TOF MS to identify hypervirulent variants of Clostridioides (Clostridium) difficile (Flores-Trevino et al., 2019; Godmer et al., 2024). Unfortunately, the current application of MS for virulence monitoring is limited by its inability to quantitatively determine virulence factor production by direct profiling without complex and time-consuming sample preparation. Consequently, mass spectrometry currently fails to meet the aforementioned requirements for virulence monitoring.
Below, we discuss methodological approaches that could serve as a basis for detection of virulence and have demonstrated successful usage.
For clinical microbiology practices, paralleling resistance testing, two options for quantitatively assessing virulence are convenient: (1) direct determination with precise measurement of the concentration of the virulence factor, and (2) indirect determination with quantitative metrics correlated with the level of virulence but lacking exact concentration measurements. Potential methods for detection of virulence are depicted in Figure 4.
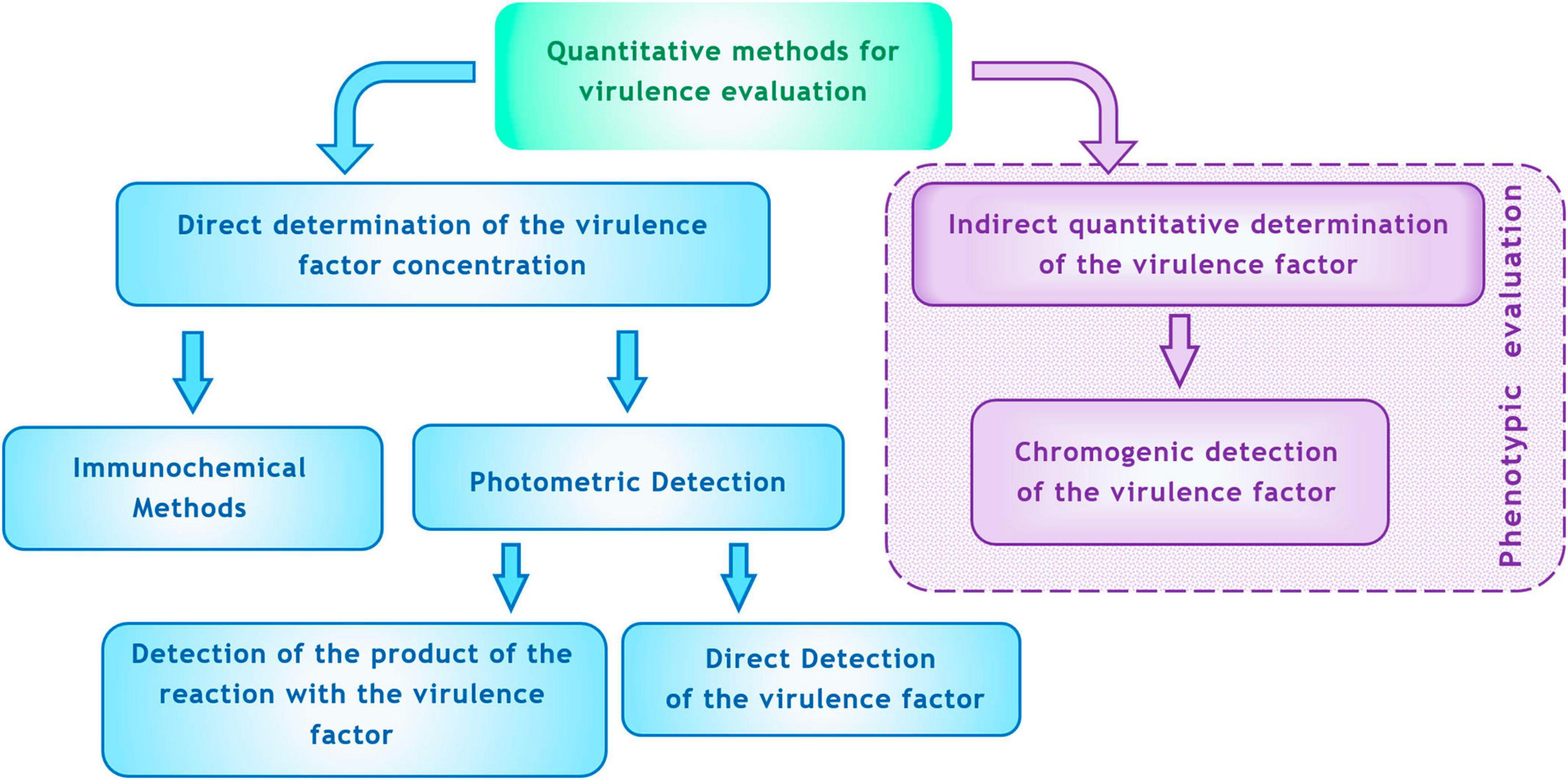
Figure 4. Methods for quantitative determination of virulence factors in vitro (explanations are in the text).
As a direct method for determining virulence factors, immunochemical techniques can be employed. These methods have a long history. Serological methods for detecting antigenic structures associated with exotoxins (such as diphtheria toxin, botulinum toxin, etc.), LPS (O-antigen), flagella (H-antigen), capsules (e.g., for serotyping S. pneumoniae), and streptococcal M-antigens have become routine. Historically, these methods focused on qualitative determination of toxigenicity or taxonomic classification (species, serogroups, serotypes). More recently, immunoassays have been developed to quantitatively measure virulence factors. For instance, ELISA has successfully been used to assess virulence factors in S. aureus, enabling the quantitative determination of staphyllococcal enterotoxin (SE) (Rahimi, 2013), alpha-hemolysin (Kehoe et al., 1983), toxic shock syndrome toxin-1 (TSST-1) (Parsonnet et al., 1985), protein A (Löfdahl et al., 1983), exfoliative toxin (ET) (Ladhani et al., 2001), and von Willebrand factor-binding protein (vWbp) (Motta et al., 2023). Immunoassays have also proved effective for quantitatively measuring P. aeruginosa virulence factors, including pyocyanin and 1-hydroxyphenazine (Pastells et al., 2016), alkaline proteinase, elastase, and exotoxin A (Shigematsu et al., 2007), and exolysin ExlA (Deruelle et al., 2021) using ELISA.
Moreover, commercial kits based on immunoassays are available not just for research purposes. For example, Cusabio Biotech Co., Ltd. (Hubei, China) manufactures a commercial Human Pseudomonas Exotoxin A PEA ELISA Kit for detecting P. aeruginosa exotoxin A. Oxoid Ltd. (Hampshire, United Kingdom) produces sets for detecting exotoxins, including the ProSpecT™ STEC (Shiga Toxin E. coli) Microplate Assay, ProSpecT™ C. difficile Toxin A/B Microplate Assay, SET-RPLA Kit Toxin Detection Kit for semi-quantitative detection of staphylococcal enterotoxins A, B, C, and D, and TST-RPLA Toxin Detection Kit for semi-quantitative detection of staphylococcal toxic shock syndrome toxin. Abcam Ltd. (Cambridge, United Kingdom) offers Protein A ELISA Kits for the quantitative determination of S. aureus protein A. These developments suggest that modern technologies and manufacturing capabilities can facilitate global monitoring of virulence among opportunistic pathogens using direct quantitative immunoassays.
Despite their advantages, immunoassays face certain limitations. They cannot detect virulence factors composed of simple small molecules (e.g., cyanides produced by some glucose-non-fermenting Gram-negative bacteria) (Enderby et al., 2009) or molecules exhibiting substantial variability within a single species (e.g., P. aeruginosa strains produce around 60 variants of rhamnolipids) (Abdel-Mawgoud et al., 2010).
Another category of methods relies on direct optical measurements to determine the concentration of virulence factors. Table 3 summarizes known methods for quantitatively assessing virulence factors using spectrophotometry. Spectrophotometric techniques have been applied to (1) measure reaction outcomes where a virulence factor exhibits enzymatic properties, (2) directly quantify virulence factors based on their intrinsic optical characteristics (e.g., pyocyanin), and (3) detect virulence factors by staining them with specific dyes (e.g., siderophores, rhamnolipids, cyanide, biofilm matrix, alginate, LPS). Notably, some of these methods have been incorporated into commercially available kits, such as SideroTec™ Total Assay Kits (Accuplex Diagnostics Ltd., Kildare, Ireland) for siderophore determination, Thermo Scientific™ Pierce™ Colorimetric Protease Assay Kit (Thermo Fisher Scientific Inc., MA, United States) for protease detection, Lipase Activity Assay Kit (Abcam Ltd., Cambridge, United Kingdom) for lipase detection, and ToxinSensor™ Chromogenic LAL Endotoxin Assay Kit (GenScript Biotech Corporation, NJ, United States) for endotoxin (LPS) detection.
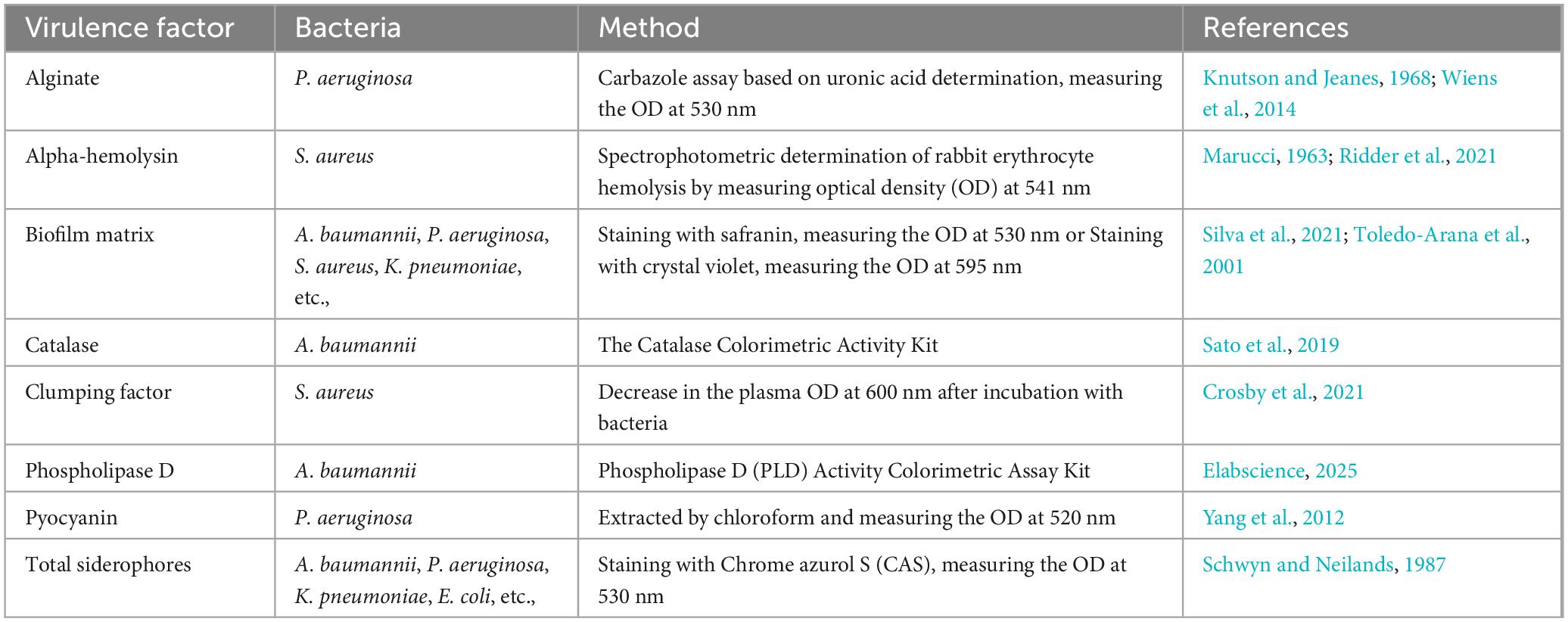
Table 3. Known spectrophotometric methods for quantitative determination of some key virulence factors.
While spectrophotometric methods based on direct in vitro determination of virulence factors offer many benefits, they have yet to be applied to the detection of exotoxins. This limitation is easily explained. Many exotoxins only display their enzymatic properties after undergoing final modifications within the target cell. For instance, P. aeruginosa exotoxin A must undergo disulfide bond reduction facilitated by disulfide isomerase, which occurs in the early endosomes of eukaryotic target cells, before acquiring its full toxic properties as an NAD+-diphtamide-ADP-ribosyltransferase (Michalska and Wolf, 2015; Krueger and Barbieri, 1995). Contact toxins operate via a distinct activation mechanism within eukaryotic cells. For example, P. aeruginosa ExoU must be ubiquitinated to acquire its toxic and enzymatic attributes (Foulkes et al., 2019). Superantigens generally lack enzymatic activity, making their detection through spectrophotometric reactions challenging.
Nevertheless, we believe that detecting exotoxins via photometric methods is feasible. To accomplish this, it is necessary to recreate the conditions for toxin activation in vitro and subsequently measure the enzymatic activity of the toxin in a reaction with a substrate. No theoretical barriers prevent this approach. In fact, the history of ExoU investigation demonstrates that restoring ExoU toxicity in vitro has already been achieved. Sato et al. (2006) treated pro-toxin ExoU with superoxide dismutase, after which it gained toxic properties. While the authors concluded that superoxide dismutase acted as an obligate cofactor for ExoU, subsequent studies revealed that ubiquitination is the true driver behind ExoU activation (Stirling et al., 2006). The observed activation by superoxide dismutase likely resulted from insufficient purification of the enzyme from ubiquitin. Fortunately, this methodological error holds immense positive implications: it proves that exotoxin activity can indeed be restored in vitro, thereby opening doors for novel detection methods. In, Zhang et al. (2017) corroborated this idea by demonstrating that treating ExoU with phosphatidylinositol 4,5-bisphosphate enabled it to acquire phospholipase activity and thus toxicity.
Another intriguing and highly promising approach for detecting virulence factors involves the use of chromogenic substrates. Within the scope of this review, we define “chromogenic substance” as a compound that can be cleaved by specific enzymes at predefined locations, resulting in a visible color change. Chromogenic substrates boast several notable qualities: (1) high substrate specificity when interacting with a particular enzyme, (2) versatility, as there are no theoretical constraints preventing the design of custom chromogens for virtually any enzyme, and (3) cost-effectiveness and ease of interpreting reaction outcomes.
These attributes position chromogenic substrates as cornerstones for building quantitative assessments of virulence factors. Chromogenic virulence testing can be performed on agar-based media. Previous experience with semiquantitative evaluations of virulence factors on agar plates enriched with special additives - whereby the virulence factor generates a colorless or colored halo around the bacterial colony - provides a solid foundation. The diameter of this halo correlates with the quantity of the virulence factor. One of the most well-known examples is the determination of hemolysins on blood agar. Bacterial isolates are plated on sheep blood agar plates, and after incubation, the diameter of the hemolysis zone around the colonies is measured (Stulik et al., 2014; Sankomkai et al., 2020). Using this technique, key virulence factors such as S. aureus α-toxin and E. faecium cytolysin have been assessed (Hällgren et al., 2009; Mizobuchi et al., 1994). S. aureus aureolysin was analyzed using TSB agar supplemented with 15% non-fat milk (Bastock et al., 2021). S. aureus coagulase activity was determined on agarose plates (0.9%) containing bovine fibrinogen and 1% plasma (Bjerketorp et al., 2004). For lipase detection, triglycerides (e.g., tributyrin) are added to agarized agar, and results are evaluated by measuring the clear zone diameters around colonies (Sankomkai et al., 2020). DNAse activity is measured on DNA-agar or DNA-agar-Toluidine Blue-O dye diffusion systems. After bacterial incubation, the agar surface is treated with HCl, and the diameter of the resultant clearing zone around colonies correlates with DNAse activity (Sankomkai et al., 2020). In the TDA system, a pink zone of activity around colonies is measured (Kamman and Tatini, 1977). To assess extracellular protease production, bacterial suspensions are plated onto casein agar plates incorporating bromocresol green dye. The transparent zone of protease activity around colonies reflects protease activity (Vijayaraghavan and Vincent, 2013). For qualitative analysis of siderophore production, the Chrome Azurol Sulfonate (CAS) agar plate assay is employed. Here, the release of CAS dye from the Fe3+-dye complex causes a color change from blue to orange, signifying siderophore production (Kronda et al., 2013). There are numerous variations of these methods, differing in technical details and dyes. The underlying principle remains constant measuring the diameter of the virulence factor diffusion zone in agar. It’s worth noting the existing precedent for qualitatively determining virulence factors based on specific enzymatic properties using chromogenic agar. CHROMagar has introduced CHROMagar™ STEC for detecting Shiga-Toxin-producing E. coli (Gouali et al., 2013). This medium exemplifies how a chromogenic substance can act as an enzyme target, the presence of which correlates with the production of virulence factors (in this case, Shiga toxin). This underscores the potential of chromogenic testing to reflect the production of virulence factors, even those lacking enzymatic activity. This is especially pertinent for detecting superantigens, which are not enzymes.
In our view, combining virulence factor diffusion with chromogenic indication presents a promising pathway for advancing virulence factor determination, analogous to the disk-diffusion method for AMR testing. Implementation of such methods for quantitative virulence testing will require standardizing bacterial quantities. This task is technically achievable using auxiliary devices, such as membrane micro-inserts, which limit bacterial spread during incubation. Another standardization option could involve basing results not on the diameter of color changes in agar around the colony but on the ratio between the area of altered-color agar and the colony diameter.
The merits of employing chromogenic agars for virulence detection include technological accessibility, relative cost-effectiveness, ergonomics, and user-friendly result recording.
A visualization of the comparison of virulence detection methods is presented in Figure 5. Likely, the decisive factor in choosing a detection method will be determined by the balance between result quality (quality criteria described above) and accessibility for routine practice (Supplementary Table 2).
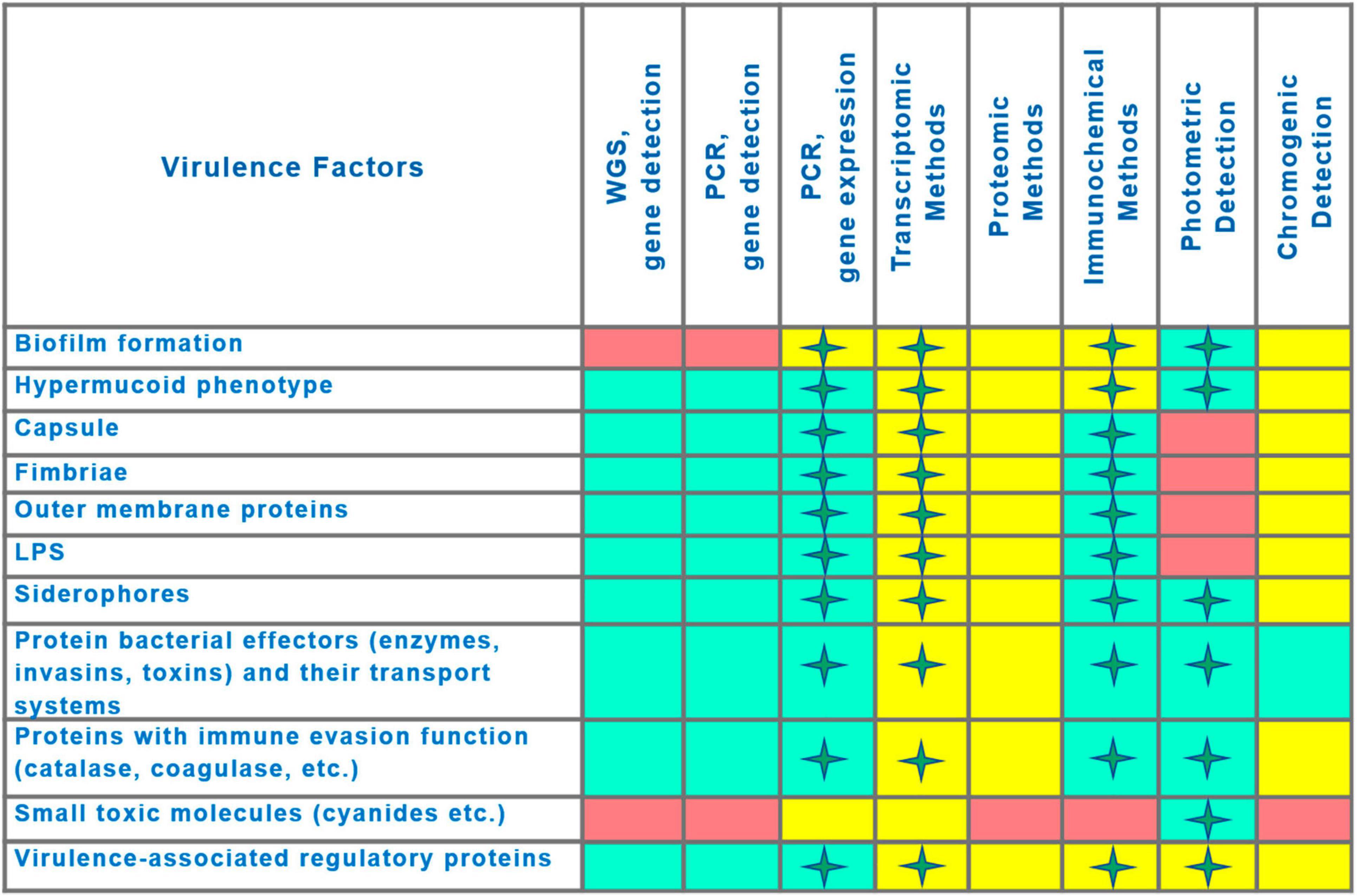
Figure 5. Comparison of capabilities of methods for virulence detection. Asterisks indicate the possibility of quantitative measurement of virulence factor production. Capability of method to detect the virulence factor is indicated by the rectangle fill color as follows: Green, application is feasible; Pink, application is not feasible; Yellow, capabilities have been demonstrated, but practical applicability at present is uncertain (technological or financial limitations).
Future directions
Key prerequisites for implementing virulence detection in clinical practice include method standardization/validation and executing a pilot project. Initial implementation steps could comprise sequential actions including (1) selecting, adapting, and standardizing optimal clinically-relevant methods for detecting virulence factors in several relevant opportunistic pathogens (e.g., ESKAPE group) based on literature data, (2) attracting funding from non-profit foundations and interested businesses to support pilot studies in a limited number of clinical laboratories, (3) conducting pilot studies in select clinical labs, (4) collecting, analyzing, and publishing/reporting collected data on the pathogenic role of virulence factors to enhance awareness and understanding of their clinical significance. If positive results are achieved, it would be possible to initiate broader cooperation with interested public organizations (ESCMID research groups, etc.), and manufacturers of diagnostic kits. Our review aligns with the first point of this action plan, and we expect it to serve as a stimulus increasing interest among professional organizations and their leaders in addressing the issue of virulence.
Limitations
Our review has several limitations. Firstly, we analyzed the virulence of opportunistic pathogens, representing just one side of the infection process, without considering the host immune system’s contribution, the impact of microbiota, or comorbidities. Secondly, the review focused primarily on a limited number of opportunistic bacterial species, for which fewer controversies exist in the literature regarding their pathogenic factors. The existing imbalance in the amount of controversy may stem from unequal research intensity directed toward different species and the disparity in the number of published works dedicated to various species. Additionally, the incomplete state of knowledge regarding genetic determinants of virulence in opportunistic pathogens restricts the perfect interpretation of virulence, although it does not preclude the detection of known virulence factors for informational purposes. Lastly, the review did not incorporate an analysis of correlations between virulence factors and the transmissibility of the infectious agent.
Conclusion
Current technological advancements now enable the integration of virulence detection for main opportunistic pathogens into clinical laboratory practice. To briefly justify the necessity of introducing virulence testing into clinical routines, it is imperative to address two central questions. What benefits will virulence detection bring to individual clinical cases? And what advantages will global-level monitoring of virulence offer? Answers to these queries exist:
1. Determining the virulence of isolates from individual patients improves the prediction of the progression of infectious processes. The activity of virulence factors can mirror the actual threat posed by a pathogen, shedding light on the likelihood of infection chronicity or exacerbation.
2. Evaluating the virulence profiles of circulating nosocomial pathogens aids in rationalizing infection control at the hospital level, drawing on principles similar to those guiding the fight against antibiotic-resistant strains.
3. As virulence testing becomes more widespread, data on the global prevalence of highly virulent strains will accumulate, facilitating optimization of anti-epidemic measures and enabling healthcare resources to be reallocated strategically toward the greatest threats.
4. Finally, monitoring virulence may prove invaluable for informing strategies to develop antivirulence therapy.
Author contributions
IC: Conceptualization, Visualization, Writing – original draft. IA: Conceptualization, Writing – review & editing. ME: Formal analysis, Writing – review & editing. RK: Supervision, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
We thank Julia Bocharova, The University of Szeged, Hungary, for sharing her valuable advice on writing the review.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1638925/full#supplementary-material
References
Abdelaziz, A. A., Kamer, A. M. A., Al-Monofy, K. B., and Al-Madboly, L. A. (2023). Pseudomonas aeruginosa’s greenish-blue pigment pyocyanin: Its production and biological activities. Microb. Cell Fact. 22:110. doi: 10.1186/s12934-023-02122-1
Abdel-Mawgoud, A. M., Lépine, F., and Déziel, E. (2010). Rhamnolipids: Diversity of structures, microbial origins and roles. Appl. Microbiol. Biotechnol. 86, 1323–1336. doi: 10.1007/s00253-010-2498-2
Abed, S. M., Raoof, W. M., Assie, A. H. A., Al-Jaafari, E. N. I., and Hadi, Y. A. (2016). Multiplex PCR for detection of exfoliative toxins, and toxic shock syndrome toxin 1 genes in MRSA strains of clinical Staphylococcus aureus isolates. Tikrit J. Pure Sci. 21, 39–44. doi: 10.25130/tjps.v21i3.993
Adams, J. (1807). Observations on morbid poisons, chronic and acute: The first compehending syphilis, yaws, sivvens, elephantiasis, and the anomala confounded with them; the second the acute contagions, particularly the variolous & vaccine. London: Printed for J. Callow, Crown Court, Princes Street, Soho by J. & W. Smith.
Adwan, G., Adwan, K., and Hiba Bourinee, H. (2016). Molecular characterization of some new E. coli strains theoretically responsible for both intestinal and extraintestinal infections. Int. J. Med. Res. Health Sci. 5, 158–163.
Ahmad, I., Nadeem, A., Mushtaq, F., Zlatkov, N., Shahzad, M., Zavialov, A. V., et al. (2023). Csu pili dependent biofilm formation and virulence of Acinetobacter baumannii. NPJ Biofilms Microb. 9:101. doi: 10.1038/s41522-023-00465-6
Ahmed, N. A. A. M., Petersen, F. C., and Scheie, A. A. (2008). Biofilm formation and autoinducer-2 signaling in Streptococcus intermedius: Role of thermal and pH factors. Oral Microbiol. Immunol. 23, 492–497. doi: 10.1111/j.1399-302x.2008.00460.x
Allel, K., Day, L., Hamilton, A., Lin, L., Furuya-Kanamori, L., Moore, C. E., et al. (2023). Global antimicrobial-resistance drivers: An ecological country-level study at the human–animal interface. Lancet Planet Health 7, e291–e303. doi: 10.1016/S2542-5196(23)00026-8
Arisoy, M., Aysev, D., Ekim, M., Özel, D., Köse, S. K., Özsoy, E. D., et al. (2006). Detection of virulence factors of Escherichia coli from children by multiplex polymerase chain reaction. Int. J. Clin. Pract. 60, 170–173. doi: 10.1111/j.1742-1241.2005.00668.x
Bachman, M. A., Oyler, J. E., Burns, S. H., Caza, M., Lépine, F., Dozois, C. M., et al. (2011). Klebsiella pneumoniae yersiniabactin promotes respiratory tract infection through evasion of lipocalin 2. Infect. Immun. 79, 3309–3316. doi: 10.1128/IAI.05114-11
Badamchi, A., Masoumi, H., Javadinia, S., Asgarian, R., and Tabatabaee, A. (2017). Molecular detection of six virulence genes in Pseudomonas aeruginosa isolates detected in children with urinary tract infection. Microb. Pathog. 107, 44–47. doi: 10.1016/j.micpath.2017.03.009
Badiou, C., Dumitrescu, O., George, N., Forbes, A. R., Drougka, E., Chan, K. S., et al. (2010). Rapid detection of Staphylococcus aureus panton-valentine leukocidin in clinical specimens by enzyme-linked immunosorbent assay and immunochromatographic tests. J. Clin. Microbiol. 48, 1384–1390. doi: 10.1128/JCM.02274-09
Bæk, K. T., Thøgersen, L., Mogenssen, R. G., Mellergaard, M., Thomsen, L. E., Petersen, A., et al. (2015). Stepwise decrease in daptomycin susceptibility in clinical Staphylococcus aureus isolates associated with an initial mutation in rpoB and a compensatory inactivation of the clpX gene. Antimicrob. Agents Chemother. 59, 6983–6991. doi: 10.1128/AAC.01303-15
Bastock, R. A., Marino, E. C., Wiemels, R. E., Holzschu, D. L., Keogh, R. A., Zapf, R. L., et al. (2021). Staphylococcus aureus responds to physiologically relevant temperature changes by altering its global transcript and protein profile. mSphere 6:e01303–20. doi: 10.1128/msphere.01303-20
Beceiro, A., Moreno, A., Fernández, N., Vallejo, J. A., Aranda, J., Adler, B., et al. (2014). Biological cost of different mechanisms of colistin resistance and their impact on virulence in Acinetobacter baumannii. Antimicrob. Agents Chemother. 58, 518–526. doi: 10.1128/aac.01597-13
Benson, M. A., Lilo, S., Wasserman, G. A., Thoendel, M., Smith, A., Horswill, A. R., et al. (2011). Staphylococcus aureus regulates the expression and production of the staphylococcal superantigen-like secreted proteins in a Rot-dependent manner. Mol. Microbiol. 81, 659–675. doi: 10.1111/j.1365-2958.2011.07720.x
Berlutti, F., Morea, C., Battistoni, A., Sarli, S., Cipriani, P., Superti, F., et al. (2005). Iron availability influences aggregation, biofilm, adhesion and invasion of Pseudomonas aeruginosa and Burkholderia cenocepacia. Int. J. Immunopathol. Pharmacol. 18, 661–670. doi: 10.1177/039463200501800407
Bhattacharya, M., and Horswill, A. R. (2024). The role of human extracellular matrix proteins in defining Staphylococcus aureus biofilm infections. FEMS Microbiol. Rev. 48:fuae002. doi: 10.1093/femsre/fuae002
Bjerketorp, J., Jacobsson, K., and Frykberg, L. (2004). The von Willebrand factor-binding protein (vWbp) of Staphylococcus aureus is a coagulase. FEMS Microbiol. Lett. 234, 309–314. doi: 10.1111/j.1574-6968.2004.tb09549.x
Boral, J., Pinarlik, F., Ekinci, G., Can, F., and Ergönül, Ö (2023). Does emerging carbapenem resistance in Acinetobacter baumannii increase the case fatality rate? Systematic review and meta-analysis. Infect. Dis. Rep. 15, 564–575. doi: 10.3390/idr15050055
Botelho, J., Grosso, F., and Peixe, L. (2019). Antibiotic resistance in Pseudomonas aeruginosa - mechanisms, epidemiology and evolution. Drug Resist. Rev. Comment Antimicrob. Anticancer Chemother. 44:100640. doi: 10.1016/j.drup.2019.07.002
Boucher, J. C., Yu, H., Mudd, M. H., and Deretic, V. (1997). Mucoid Pseudomonas aeruginosa in cystic fibrosis: Characterization of muc mutations in clinical isolates and analysis of clearance in a mouse model of respiratory infection. Infect. Immun. 65, 3838–3846. doi: 10.1128/iai.65.9.3838-3846.1997
Boyd, E. F. (2012). Bacteriophage-encoded bacterial virulence factors and phage-pathogenicity island interactions. Adv. Virus Res. 82, 91–118. doi: 10.1016/b978-0-12-394621-8.00014-5
Browne, E. (1830). An investigation of the powers of the simple supporters of combustion to destroy the virulence of morbid poisons, and of the poisonous gases, with a view to ascertain the possibility of controuling the extension of contagious or epidemic diseases. London: Royal Society.
Bryan, L. E., Kureishi, A., and Rabin, H. R. (1983). Detection of antibodies to Pseudomonas aeruginosa alginate extracellular polysaccharide in animals and cystic fibrosis patients by enzyme-linked immunosorbent assay. J. Clin. Microbiol. 18, 276–282. doi: 10.1128/jcm.18.2.276-282.1983
Budzyńska, A., Skowron, K., Kaczmarek, A., Wietlicka-Piszcz, M., and Gospodarek-Komkowska, E. (2021). Virulence factor genes and antimicrobial susceptibility of Staphylococcus aureus strains isolated from blood and chronic wounds. Toxins 13:491. doi: 10.3390/toxins13070491
Bukowski, M., Wladyka, B., and Dubin, G. (2010). Exfoliative Toxins of Staphylococcus aureus. Toxins 2, 1148–1165. doi: 10.3390/toxins2051148
Bulger, J., MacDonald, U., Olson, R., Beanan, J., and Russo, T. A. (2017). Metabolite transporter peg344 is required for full virulence of hypervirulent Klebsiella pneumoniae strain hvKP1 after pulmonary but not subcutaneous challenge. Infect. Immun. 85:e0093–17. doi: 10.1128/iai.00093-17
Byrne, A. S., Bissonnette, N., and Tahlan, K. (2025). Mechanisms and implications of phenotypic switching in bacterial pathogens. Can. J. Microbiol. 71, 1–19. doi: 10.1139/cjm-2024-0116
Cahill, S. M., Hall, R. M., and Kenyon, J. J. (2022). An update to the database for Acinetobacter baumannii capsular polysaccharide locus typing extends the extensive and diverse repertoire of genes found at and outside the K locus. Microb. Genom. 8:mgen000878. doi: 10.1099/mgen.0.000878
Carbonetti, N. H., and Williams, P. H. (1984). A cluster of five genes specifying the aerobactin iron uptake system of plasmid ColV-K30. Infect. Immun. 46, 7–12. doi: 10.1128/iai.46.1.7-12.1984
Casadevall, A., and Pirofski, L. A. (1999). Host-pathogen interactions: Redefining the basic concepts of virulence and pathogenicity. Infect. Immun. 67, 3703–3713. doi: 10.1128/iai.67.8.3703-3713.1999
Casadevall, A., and Pirofski, L. A. (2015). What is a host? Incorporating the microbiota into the damage-response framework. Infect. Immun. 83, 2–7. doi: 10.1128/iai.02627-14
Cerdeno-Tarraga, A. M., Efstratiou, A., Dover, L. G., and Holden, M. T. G. (2003). The complete genome sequence and analysis of Corynebacterium diphtheriae NCTC13129. Nucleic Acids Res. 31, 6516–6523. doi: 10.1093/nar/gkg874
Ceri, H., Olson, M. E., Stremick, C., Read, R. R., Morck, D., and Buret, A. (1999). The calgary biofilm device: New technology for rapid determination of antibiotic susceptibilities of bacterial biofilms. J. Clin. Microbiol. 37, 1771–1776. doi: 10.1128/jcm.37.6.1771-1776.1999
Chen, T., Dong, G., Zhang, S., Zhang, X., Zhao, Y., Cao, J., et al. (2020). Effects of iron on the growth, biofilm formation and virulence of Klebsiella pneumoniae causing liver abscess. BMC Microbiol. 20:36. doi: 10.1186/s12866-020-01727-5
Christensen, G. D., Simpson, W. A., Bisno, A. L., and Beachey, E. H. (1982). Adherence of slime-producing strains of Staphylococcus epidermidis to smooth surfaces. Infect. Immun. 37, 318–326. doi: 10.1128/iai.37.1.318-326.1982
Christian, M., Harding, C. M., Hennon, H. W., and Feldman, M. F. (2018). Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 16, 91–102. doi: 10.1038/nrmicro.2017.148
Conde-Pérez, K., Vázquez-Ucha, J. C., Álvarez-Fraga, L., Ageitos, L., Rumbo-Feal, S., Martínez-Guitián, M., et al. (2021). In-depth analysis of the role of the acinetobactin cluster in the virulence of Acinetobacter baumannii. Front. Microbiol. 12:752070. doi: 10.3389/fmicb.2021.752070
Cook-Libin, S., Sykes, E. M., Kornelsen, V., and Kumar, A. (2022). Iron acquisition mechanisms and their role in the virulence of Acinetobacter baumannii. Infect. Immun. 90:e0022322. doi: 10.1128/iai.00223-22
Cressler, C. E., McLeod, D. V., Rozins, C., Van Den Hoogen, J., and Day, T. (2016). The adaptive evolution of virulence: A review of theoretical predictions and empirical tests. Parasitology 143, 915–930. doi: 10.1017/S003118201500092X
Crosby, H. A., Kwiecinski, J. M., and Horswill, A. R. (2021). In vitro assay for quantifying clumping of Staphylococcus aureus. Methods Mol. Biol. 2341, 31–36. doi: 10.1007/978-1-0716-1550-8_5
Crosby, H. A., Kwiecinski, J., and Horswill, A. R. (2016). Staphylococcus aureus aggregation and coagulation mechanisms, and their function in host–pathogen interactions. Adv. Appl. Microbiol. 96, 1–41. doi: 10.1016/bs.aambs.2016.07.018
Cunha, E., Ferreira, A. F., Valente, S., Matos, A., Carreira, L. M., Videira, M., et al. (2023). In vivo effect of a nisin-biogel on the antimicrobial and virulence signatures of canine oral enterococci. Antibiotics 12:468. doi: 10.3390/antibiotics12030468
Damron, F. H., and Goldberg, J. B. (2012). Proteolytic regulation of alginate overproduction in Pseudomonas aeruginosa. Mol. Microbiol. 84, 595–607. doi: 10.1111/j.1365-2958.2012.08049.x
Davari Abad, E., Khameneh, A., and Vahedi, L. (2019). Identification phenotypic and genotypic characterization of biofilm formation in Escherichia coli isolated from urinary tract infections and their antibiotics resistance. BMC Res Notes 12:796. doi: 10.1186/s13104-019-4825-8
Deng, Q., Zhang, J., Zhang, M., Liu, Z., Zhong, Y., Liu, S., et al. (2020). Rapid identification of KL49 Acinetobacter baumannii associated with clinical mortality. Infect. Drug Resist. 13, 4125–4132. doi: 10.2147/IDR.S278891
Dermota, U., Šturm, A. C., Triglav, T., Smrdel, K. S., and Velimirovic, I. (2024). Whole genome sequencing and molecular epidemiology of methicillin-resistant Staphylococcus aureus isolated from patients with bacteraemia in Slovenia. Eur. J. Clin. Microbiol. Infect. Dis. 43, 969–977. doi: 10.1007/s10096-024-04802-1
Deruelle, V., Berry, A., Bouillot, S., Job, V., Maillard, A. P., Elsen, S., et al. (2021). ExlA pore-forming toxin: Localization at the bacterial membrane, regulation of secretion by cyclic-di-GMP, and detection in vivo. Toxins 13:645. doi: 10.3390/toxins13090645
Díaz, J. M., Dozois, C. M., Avelar-González, F. J., Hernández-Cuellar, E., Pokharel, P., de Santiago, A. S., et al. (2020). The vacuolating autotransporter toxin (Vat) of Escherichia coli causes cell cytoskeleton changes and produces non-lysosomal vacuole formation in bladder epithelial cells. Front. Cell. Infect. Microbiol. 10:299. doi: 10.3389/fcimb.2020.00299
Dickson, J. I., and Marples, R. R. (1986). Coagulase production by strains of Staphylococcus aureus of differing resistance characters: A comparison of two traditional methods with a latex agglutination system detecting both clumping factor and protein A. J. Clin. Pathol. 39, 371–375. doi: 10.1136/jcp.39.4.371
Dinges, M. M., Orwin, P. M., and Schlievert, P. M. (2000). Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 13, 16–34. doi: 10.1128/cmr.13.1.16-34.2000
Divyakolu, S., Chikkala, R., Ratnakar, K. S., and Sritharan, V. (2019). Hemolysins of Staphylococcus aureus—an update on their biology, role in pathogenesis and as targets for anti-virulence therapy. Adv. Infect. Dis. 9, 80–104. doi: 10.4236/aid.2019.92007
Dong, N., Yang, X., Chan, E. W. C., Zhang, R., and Chen, S. (2022). Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. eBioMedicine 79:103998. doi: 10.1016/j.ebiom.2022.103998
Doss, S. A. (1994). Chromosomally-mediated antibiotic resistance and virulence. J. Med. Microbiol. 40, 305–306. doi: 10.1099/00222615-40-5-305
Eaton, T. J., and Gasson, M. J. (2002). A variant enterococcal surface protein Esp(fm) in Enterococcus faecium; distribution among food, commensal, medical, and environmental isolates. FEMS Microbiol. Lett. 216, 269–275. doi: 10.1111/j.1574-6968.2002.tb11446.x
El Fertas-Aissani, R., Messai, Y., Alouache, S., and Bakour, R. (2013). Virulence profiles and antibiotic susceptibility patterns of Klebsiella pneumoniae strains isolated from different clinical specimens. Pathol. Biol. 61, 209–216. doi: 10.1016/j.patbio.2012.10.004
Elabscience (2025). Phospholipase D (PLD) activity colorimetric assay Kit (E-BC-K783-M). Available online at: https://www.elabscience.com/p/phospholipase-d-pld-activity-colorimetric-assay-kit–e-bc-k783-m (accessed April 02, 2025).
El-Solh, A. A., Hattemer, A., Hauser, A. R., Alhajhusain, A., and Vora, H. (2012). Clinical outcomes of type III Pseudomonas aeruginosa bacteremia. Crit. Care Med. 40, 1157–1163. doi: 10.1097/CCM.0b013e3182377906
Enderby, B., Smith, D., Carroll, W., and Lenney, W. (2009). Hydrogen cyanide as a biomarker for Pseudomonas aeruginosa in the breath of children with cystic fibrosis. Pediatr. Pulmonol. 44, 142–147. doi: 10.1002/ppul.20963
Eyre, J. W., and Washbourn, J. W. (1899). Varieties and virulence of the pneumococcus. Lancet 153, 19–22. doi: 10.1016/S0140-6736(01)78950-7
Falkow, S. (2004). Molecular Koch’s postulates applied to bacterial pathogenicity – a personal recollection 15 years later. Nat. Rev. Microbiol. 2, 67–72. doi: 10.1038/nrmicro799
Flores-Trevino, S., Garza-González, E., Mendoza-Olazarán, S., Morfín-Otero, R., Camacho-Ortiz, A., Rodríguez-Noriega, E., et al. (2019). Screening of biomarkers of drug resistance or virulence in ESCAPE pathogens by MALDI-TOF mass spectrometry. Sci. Rep. 9:18945. doi: 10.1038/s41598-019-55430-1
Foulkes, D. M., McLean, K., Haneef, A. S., Fernig, D. G., Winstanley, C., Berry, N., et al. (2019). Pseudomonas aeruginosa toxin ExoU as a therapeutic target in the treatment of bacterial infections. Microorganisms 7:707. doi: 10.3390/microorganisms7120707
Gaines, J. M., Carty, N. L., Tiburzi, F., Davinic, M., Visca, P., Colmer-Hamood, J. A., et al. (2007). Regulation of the Pseudomonas aeruginosa toxA, regA and ptxR genes by the iron-starvation sigma factor PvdS under reduced levels of oxygen. Microbiology 153, 4219–4233. doi: 10.1099/mic.0.2007/011338-0
Galia, L., Ligozzi, M., Bertoncelli, A., and Mazzariol, A. (2019). Real-time PCR assay for detection of Staphylococcus aureus, panton-valentine leucocidin and methicillin resistance directly from clinical samples. AIMS Microbiol. 5, 138–146. doi: 10.3934/microbiol.2019.2.138
Garcia, E. C., Brumbaugh, A. R., and Mobley, H. L. (2011). Redundancy and specificity of Escherichia coli iron acquisition systems during urinary tract infection. Infect. Immun. 79, 1225–1235. doi: 10.1128/IAI.01222-10
Gerson, S., Betts, J. W., Lucaßen, K., Nodari, C. S., Wille, J., Josten, M., et al. (2019). Investigation of novel pmrB and eptA mutations in isogenic Acinetobacter baumannii isolates associated with colistin resistance and increased virulence in vivo. Antimicrob. Agents Chemother. 63:e01586–18. doi: 10.1128/AAC.01586-18
Godmer, A., Giai Gianetto, Q., Le Neindre, K., Latapy, V., Bastide, M., Ehmig, M., et al. (2024). Contribution of MALDI-TOF mass spectrometry and machine learning including deep learning techniques for the detection of virulence factors of Clostridioides difficile strains. Microb. Biotechnol. 17:e14478. doi: 10.1111/1751-7915.14478
Gouali, M., Ruckly, C., Carle, I., Lejay-Collin, M., and Weill, F. X. (2013). Evaluation of CHROMagar STEC and STEC O104 chromogenic agar media for detection of shiga toxin-producing Escherichia coli in stool specimens. J. Clin. Microbiol. 51, 894–900. doi: 10.1128/JCM.03121-12
Griffith, F. (1928). The significance of pneumococcal types. J. Hyg. 27, 113–159. doi: 10.1017/s0022172400031879
Grudniak, A. M., Klecha, B., and Wolska, K. I. (2018). Effects of null mutation of the heat-shock gene htpg on the production of virulence factors by Pseudomonas aeruginosa. Future Microbiol. 13, 69–80. doi: 10.2217/fmb-2017-0111
Gu, F., He, W., Zhu, D., Zeng, Q., Li, X., Xiao, S., et al. (2024). Genome-wide comparative analysis of CC1 Staphylococcus aureus between colonization and infection. Eur. J. Med. Res. 29:474. doi: 10.1186/s40001-024-02076-z
Guo, Y., Xiao, R., Feng, J., Wang, X., Lai, J., Kang, W., et al. (2024). Distribution of virulence genes and antimicrobial resistance of Escherichia coli isolated from hospitalized neonates: A multi-center study across China. Heliyon 10:e35991. doi: 10.1016/j.heliyon.2024.e35991
Gupta, S. (2023). Evolution of pathogen virulence: Studying the complex interplay of pathogen interactions, virulence and transmission helps us understand how they evolve and spread. EMBO Rep. 24:e57611. doi: 10.15252/embr.202357611
Gupte, A., Jyot, J., Ravi, M., and Ramphal, R. (2021). High pyocyanin production and non-motility of Pseudomonas aeruginosa isolates are correlated with septic shock or death in bacteremic patients. PLoS One 16:e0253259. doi: 10.1371/journal.pone.0253259
Haddad, O., Merghni, A., Elargoubi, A., Rhim, H., Kadri, Y., and Mastouri, M. (2018). Comparative study of virulence factors among methicillin resistant Staphylococcus aureus clinical isolates. BMC Infect. Dis. 18:560. doi: 10.1186/s12879-018-3457-2
Hällgren, A., Claesson, C., Saeedi, B., Monstein, H. J., Hanberger, H., and Nilsson, L. E. (2009). Molecular detection of aggregation substance, enterococcal surface protein, and cytolysin genes and in vitro adhesion to urinary catheters of Enterococcus faecalis and E. faecium of clinical origin. Int. J. Med. Microbiol. 299, 323–332. doi: 10.1016/j.ijmm.2008.10.001
Hasani, A., Soltani, E., Ahangarzadeh Rezaee, M., Pirzadeh, T., Ahangar Oskouee, M., Hasani, A., et al. (2020). Serotyping of Klebsiella pneumoniae and its relation with capsule-associated virulence genes, antimicrobial resistance pattern, and clinical infections: A descriptive study in medical practice. Infect. Drug Resist. 13, 1971–1980. doi: 10.2147/IDR.S243984
Hauser, A. R., Cobb, E., Bodí, M., Mariscal, D., Vallés, J., Engel, J. N., et al. (2002). Type III protein secretion is associated with poor clinical outcomes in patients with ventilator-associated pneumonia caused by Pseudomonas aeruginosa. Crit. Care Med. 30, 521–528. doi: 10.1097/00003246-200203000-00005
Hazen, T. H., Michalski, J. M., Tennant, S. M., and Rasko, D. A. (2023). Genomic diversity of non-diarrheagenic fecal Escherichia coli from children in sub-Saharan Africa and south Asia and their relatedness to diarrheagenic E. coli. Nat. Commun. 14:1400. doi: 10.1038/s41467-023-36337-y
Hemati, S., Halimi, S., Jabalameli, F., Emaneini, M., and Beigverdi, R. (2024). Phylogenetic group, antibiotic resistance, virulence gene, and genetic diversity of Escherichia coli causing bloodstream infections in Iran. Front. Microbiol. 15:1426510. doi: 10.3389/fmicb.2024.1426510
Henderson, B., Poole, S., and Wilson, M. (1996). Bacterial modulins: A novel class of virulence factors which cause host tissue pathology by inducing cytokine synthesis. Microbiol. Rev. 60, 316–341. doi: 10.1128/mr.60.2.316-341.1996
Hossain, A. Z., and Chowdhury, A. M. A. (2024). Understanding the evolution and transmission dynamics of antibiotic resistance genes: A comprehensive review. J. Basic Microbiol. 64:e2400259. doi: 10.1002/jobm.202400259
Hsiao, C. Y., Lee, Y. C., Shyu, D. J., Su, C. P., Lin, M. Y., Guo, N. Y., et al. (2024). Molecular characterization of high and low virulent Escherichia coli clinical strains isolated from patients with urinary tract infections with or without bacteremia in southern Taiwan. Infect. Drug Resist. 17, 2389–2399. doi: 10.2147/IDR.S458925
Huang, X., Li, X., An, H., Wang, J., Ding, M., Wang, L., et al. (2022). Capsule type defines the capability of Klebsiella pneumoniae in evading Kupffer cell capture in the liver. PLoS Pathog. 18:e1010693. doi: 10.1371/journal.ppat.1010693
Hung, W. T., Cheng, M. F., Tseng, F. C., Chen, Y. S., Lee, S. S. J., Chang, T. H., et al. (2019). Bloodstream infection with extended-spectrum beta-lactamase–producing Escherichia coli: The role of virulence genes. J Microbiol. Immunol. Infect. 52, 947–955. doi: 10.1016/j.jmii.2019.03.005
Hussain, K., Bandyopadhyay, A., Roberts, N., Mughal, N., Moore, L. S., and Fuller, L. C. (2022). Panton–Valentine leucocidin-producing Staphylococcus aureus: A clinical review. Clin. Exp. Dermatol. 47, 2150–2158. doi: 10.1111/ced.15392
Jackson, D. W., Simecka, J. W., and Romeo, T. (2002). Catabolite repression of Escherichia coli biofilm formation. J. Bacteriol. 184, 3406–3410. doi: 10.1128/JB.184.12.3406-3410.2002
Jacobs, A. C., Hood, I., Boyd, K. L., Olson, P. D., Morrison, J. M., Carson, S., et al. (2010). Inactivation of phospholipase D Diminishes Acinetobacter baumannii pathogenesis. Infect. Immun. 78, 1952–1962. doi: 10.1128/IAI.00889-09
Jeong, G. J., Khan, F., Tabassum, N., Jo, D. M., Jung, W. K., and Kim, Y. M. (2024). Roles of Pseudomonas aeruginosa siderophores in interaction with prokaryotic and eukaryotic organisms. Res. Microbiol. 175:104211. doi: 10.1016/j.resmic.2024.104211
Jin, L., Hu, X., Tian, Y., Fang, M., Dong, X., Jiang, Y., et al. (2024). Detection of Staphylococcus aureus virulence gene pvl based on CRISPR strip. Front. Immunol. 15:1345532. doi: 10.3389/fimmu.2024.1345532
Johnson, J. R., and Russo, T. A. (2018). Molecular epidemiology of extraintestinal pathogenic Escherichia coli. EcoSal Plus 8:10. doi: 10.1128/ecosalplus.esp-0004-2017
Johnson, J. R., and Stell, A. L. (2000). Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 181, 261–272. doi: 10.1086/315217
Johnson, J. R., Magistro, G., Clabots, C., Porter, S., Manges, A., Thuras, P., et al. (2018). Contribution of yersiniabactin to the virulence of an Escherichia coli sequence type 69 (“clonal group A”) cystitis isolate in murine models of urinary tract infection and sepsis. Microb. Pathog. 120, 128–131. doi: 10.1016/j.micpath.2018.04.048
Kamman, J. F., and Tatini, S. R. (1977). Optimal conditions for assay of staphylococcal nuclease. J. Food Sci. 42, 421–424. doi: 10.1111/j.1365-2621.1977.tb01513.x
Karash, S., Nordell, R., Ozer, E. A., and Yahr, T. L. (2021). Genome sequences of two Pseudomonas aeruginosa isolates with defects in type III secretion system gene expression from a chronic ankle wound infection. Microbiol. Spectr. 9:e0034021. doi: 10.1128/Spectrum.00340-21
Kehoe, M., Duncan, J., Foster, T., Fairweather, N., and Dougan, G. (1983). Cloning, expression, and mapping of the Staphylococcus aureus alpha-hemolysin determinant in Escherichia coli K-12. Infect. Immun. 41, 1105–1111. doi: 10.1128/iai.41.3.1105-1111.1983
Kelly, B. G., Vespermann, A., and Bolton, D. J. (2009). Horizontal gene transfer of virulence determinants in selected bacterial foodborne pathogens. Food Chem. Toxicol. 47, 969–977. doi: 10.1016/j.fct.2008.02.007
Khodaverdian, V. Y., and Shoham, M. (2014). ELISA for alpha-hemolysin (Hla) in methicilin-resistant Staphylococcus aureus (MRSA). BioProtocol 4:e1118. doi: 10.21769/BioProtoc.1118
Kim, S., Li, X. H., Hwang, H. J., and Lee, J. H. (2020). Thermoregulation of Pseudomonas aeruginosa biofilm formation. Appl. Environ. Microbiol. 86:e01584–20. doi: 10.1128/AEM.01584-20
Kircheri, A. (1665). Mundus Subterraneus, in XII libros digestus; quo divinum subterrestris mundi opificium, mira ergasteriorum naturæ in eo distributio, verbo pantámorphou Protei regnum, universæ denique naturæ majestas. Amsterodams: Joannem Janssonium & Elizeum Weyerstraten.
Knutson, C. A., and Jeanes, A. (1968). A new modification of the carbazole analysis: Application to heteropolysaccharides. Anal. Biochem. 24, 470–481. doi: 10.1016/0003-2697(68)90154-1
Koch, R., Gaffky, G., and Loeffler, F. (1884). The virulence of cultivated anthrax virus: Experimental studies on the artificial attenuation of the infectious properties of the bacillus of anthrax by means of cultivation. Science IV, 276–277. doi: 10.1126/science.ns-4.85.276
Kon, H., Schwartz, D., Temkin, E., Carmeli, Y., and Lellouche, J. (2020). Rapid identification of capsulated Acinetobacter baumannii using a density-dependent gradient test. BMC Microbiol. 20:285. doi: 10.1186/s12866-020-01971-9
Koraimann, G. (2018). Spread and persistence of virulence and antibiotic resistance genes: A ride on the f plasmid conjugation module. EcoSal Plus 8:10. doi: 10.1128/ecosalplus.esp-0003-2018
Kronda, J. M., Cooper, R. A., and Maddocks, S. E. (2013). Manuka honey inhibits siderophore production in Pseudomonas aeruginosa. J. Appl. Microbiol. 115, 86–90. doi: 10.1111/jam.12222
Krueger, K. M., and Barbieri, J. T. (1995). The family of bacterial ADP-ribosylating exotoxins. Clin. Microbiol. Rev. 8, 34–47. doi: 10.1128/cmr.8.1.34
Kruse, W., and Pansini, S. (1892). Untersuchungen über den Diplococcus pneumoniae und verwandte Streptokokken. Z. Hyg. Infektionskrankheiten 11, 279–380. doi: 10.1007/BF02284301
Ladhani, S., Robbie, S., Garratt, R. C., Chapple, D. S., Joannou, C. L., and Evans, R. W. (2001). Development and evaluation of detection systems for staphylococcal exfoliative toxin A responsible for scalded-skin syndrome. J. Clin. Microbiol. 39, 2050–2054. doi: 10.1128/JCM.39.6.2050-2054.2001
Lam, M. M., Wick, R. R., Watts, S. C., Cerdeira, L. T., Wyres, K. L., and Holt, K. E. (2021). A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 12:4188. doi: 10.1038/s41467-021-24448-3
Lam, M. M., Wyres, K. L., Judd, L. M., Wick, R. R., Jenney, A., Brisse, S., et al. (2018). Tracking key virulence loci encoding aerobactin and salmochelin siderophore synthesis in Klebsiella pneumoniae. Genome Med. 10:77. doi: 10.1186/s13073-018-0587-5
Lanfrank. (1894). Science of cirurgie” by Lanfranco, of Milan, 13th cent. London: Pub. for the Early English Text Society, K. Paul, Trench, Trübner & Co.
Lara, F. B., Nery, D. R., De Oliveira, P. M., Araujo, M. L., Carvalho, F. R., Messias-Silva, L. C., et al. (2017). Virulence markers and phylogenetic analysis of Escherichia coli strains with hybrid EAEC/UPEC genotypes recovered from sporadic cases of extraintestinal infections. Front. Microbiol. 8:146. doi: 10.3389/fmicb.2017.00146
Lavender, H. F., Jagnow, J. R., and Clegg, S. (2004). Biofilm formation in vitro and virulence in vivo of mutants of Klebsiella pneumoniae. Infect. Immun. 72, 4888–4890. doi: 10.1128/iai.72.8.4888-4890.2004
Lerminiaux, N. A., and Cameron, A. D. (2019). Horizontal transfer of antibiotic resistance genes in clinical environments. Can. J. Microbiol. 65, 34–44. doi: 10.1139/cjm-2018-0275
Li, L., Ledizet, M., Kar, K., Koski, R. A., and Kazmierczak, B. I. (2005). An indirect enzyme-linked immunosorbent assay for rapid and quantitative assessment of Type III virulence phenotypes of Pseudomonas aeruginosa isolates. Ann. Clin. Microbiol. Antimicrob. 4:22. doi: 10.1186/1476-0711-4-22
Li, Y., and Ni, M. (2023). Regulation of biofilm formation in Klebsiella pneumoniae. Front. Microbiol. 14:1238482. doi: 10.3389/fmicb.2023.1238482
Lin, J., Zhu, L., and Lau, G. W. (2016). Disentangling competence for genetic transformation and virulence in Streptococcus pneumoniae. Curr. Genet. 62, 97–103. doi: 10.1007/s00294-015-0520-z
Lindstedt, B. A., Finton, M. D., Porcellato, D., and Brandal, L. T. (2018). High frequency of hybrid Escherichia coli strains with combined intestinal pathogenic Escherichia coli (IPEC) and extraintestinal pathogenic Escherichia coli (ExPEC) virulence factors isolated from human faecal samples. BMC Infect. Dis. 18:544. doi: 10.1186/s12879-018-3449-2
Löfdahl, S., Guss, B., Uhlén, M., Philipson, L., and Lindberg, M. (1983). Gene for staphylococcal protein A. Proc. Natl. Acad. Sci. U.S.A. 80, 697–701. doi: 10.1073/pnas.80.3.697
Ma, M., Chu, M., Tao, L., Li, J., Li, X., Huang, H., et al. (2021). First report of oxacillin susceptible mecA-positive Staphylococcus aureus in a children’s hospital in Kunming, China. Infect. Drug Resist. 14, 2597–2606. doi: 10.2147/IDR.S317670
Mai, D., Wu, A., Li, R., Cai, D., Tong, H., Wang, N., et al. (2023). Identification of hypervirulent Klebsiella pneumoniae based on biomarkers and Galleria mellonella infection model. BMC Microbiol. 23:369. doi: 10.1186/s12866-023-03124-0
Maldonado, N., López-Hernández, I., García-Montaner, A., López-Cortés, L. E., Pérez-Crespo, P. M. M., Retamar-Gentil, P., et al. (2024). Whole-genome characterisation of Escherichia coli isolates from patients with bacteraemia presenting with sepsis or septic shock in Spain: A multicentre cross-sectional study. Lancet Microbe 5, e390–e399. doi: 10.1016/S2666-5247(23)00369-5
Man, L., Klare, W. P., Dale, A. L., Cain, J. A., and Cordwell, S. J. (2021). Integrated mass spectrometry-based multi-omics for elucidating mechanisms of bacterial virulence. Biochem. Soc. Trans. 49, 1905–1926. doi: 10.1042/bst20191088
Marucci, A. A. (1963). Mechanism of action of staphylococcal alpha-hemolysin. J. Bacteriol. 86, 1182–1188. doi: 10.1128/jb.86.6.1182-1188.1963
Maurya, A., Kumar, R., Singh, A., and Raj, A. (2021). Investigation on biofilm formation activity of Enterococcus faecium under various physiological conditions and possible application in bioremediation of tannery effluent. Bioresour. Technol. 339:125586. doi: 10.1016/j.biortech.2021.125586
May, T. B., Shinabarger, D., Maharaj, R. O., Kato, J., Chu, L., DeVault, J. D., et al. (1991). Alginate synthesis by Pseudomonas aeruginosa: A key pathogenic factor in chronic pulmonary infections of cystic fibrosis patients. Clin. Microbiol. Rev. 4, 191–206. doi: 10.1128/cmr.4.2.191
Metz, J. A., and Boldin, B. (2023). Immunity-driven evolution of virulence and diversity in respiratory diseases. Evolution 77, 2392–2408. doi: 10.1093/evolut/qpad145
Michalska, M., and Wolf, P. (2015). Pseudomonas exotoxin A: Optimized by evolution for effective killing. Front. Microbiol. 6:963. doi: 10.3389/fmicb.2015.00963
Mihara, K., Tanabe, T., Yamakawa, Y., Funahashi, T., Nakao, H., Narimatsu, S., et al. (2004). Identification and transcriptional organization of a gene cluster involved in biosynthesis and transport of acinetobactin, a siderophore produced by Acinetobacter baumannii ATCC 19606T. Microbiology 150, 2587–2597. doi: 10.1099/mic.0.27141-0
Miranda, P. S. D., Lannes-Costa, P. S., Pimentel, B. A. S., Silva, L. G., Ferreira-Carvalho, B. T., Menezes, G. C., et al. (2018). Biofilm formation on different pH conditions by Streptococcus agalactiae isolated from bovine mastitic milk. Lett. Appl. Microbiol. 67, 235–243. doi: 10.1111/lam.13015
Miyazaki, S., Matsumoto, T., Tateda, K., Ohno, A., and Yamaguchi, K. (1995). Role of exotoxin A in inducing severe Pseudomonas aeruginosa infections in mice. J. Med. Microbiol. 43, 169–175. doi: 10.1099/00222615-43-3-169
Mizobuchi, S., Minami, J., Jin, F., Matsushita, O., and Okabe, A. (1994). Comparison of the virulence of methicillin-resistant and methicillin-sensitive Staphylococcus aureus. Microbiol. Immunol. 38, 599–605. doi: 10.1111/j.1348-0421.1994.tb01829.x
Modarresi, F., Azizi, O., Shakibaie, M. R., Motamedifar, M., Mosadegh, E., and Mansouri, S. (2015). Iron limitation enhances acyl homoserine lactone (AHL) production and biofilm formation in clinical isolates of Acinetobacter baumannii. Virulence 6, 152–161. doi: 10.1080/21505594.2014.1003001
Motta, C., Pellegrini, A., Camaione, S., Geoghegan, J., Speziale, P., Barbieri, G., et al. (2023). von Willebrand factor-binding protein (vWbp)-activated factor XIII and transglutaminase 2 (TG2) promote cross-linking between FnBPA from Staphylococcus aureus and fibrinogen. Sci. Rep. 13:11683. doi: 10.1038/s41598-023-38972-3
Muniesa, M., Colomer-Lluch, M., and Jofre, J. (2013). Potential impact of environmental bacteriophages in spreading antibiotic resistance genes. Future Microbiol. 8, 739–751. doi: 10.2217/fmb.13.32
Naghavi, M., Vollset, S. E., Ikuta, K. S., Swetschinski, L. R., Gray, A. P., Wool, E. E., et al. (2024). Global burden of bacterial antimicrobial resistance 1990-2021: A systematic analysis with forecasts to 2050. Lancet 404, 1199–1226. doi: 10.1016/s0140-6736(24)01867-1
Nichols, K. B., Totsika, M., Moriel, D. G., Lo, A. W., Yang, J., Wurpel, D. J., et al. (2016). Molecular characterization of the vacuolating autotransporter toxin in uropathogenic Escherichia coli. J. Bacteriol. 198, 1487–1498. doi: 10.1128/jb.00791-15
Novick, A., and Szilard, L. (1950). Experiments with the chemostat on spontaneous mutations of bacteria. Proc. Natl. Acad. Sci. U.S.A. 36, 708–719. doi: 10.1073/pnas.36.12.708
Nuss, A. M., Schuster, F., Roselius, L., Klein, J., Bücker, R., Herbst, K., et al. (2016). A precise temperature-responsive bistable switch controlling Yersinia virulence. PLoS Pathog. 12:e1006091. doi: 10.1371/journal.ppat.1006091
Okeke, I. N., de Kraker, M. E., Van Boeckel, T. P., Kumar, C. K., Schmitt, H., Gales, A. C., et al. (2024). The scope of the antimicrobial resistance challenge. Lancet 403, 2426–2438. doi: 10.1016/S0140-6736(24)00876-6
Olivares, E., Badel-Berchoux, S., Provot, C., Jaulhac, B., Prévost, G., Bernardi, T., et al. (2016). The biofilm ring test: A rapid method for routine analysis of Pseudomonas aeruginosa biofilm formation kinetics. J. Clin. Microbiol. 54, 657–661. doi: 10.1128/jcm.02938-15
Ott, L., Möller, J., and Burkovski, A. (2022). Interactions between the re-emerging pathogen Corynebacterium diphtheriae and host cells. Int. J. Mol. Sci. 23:3298. doi: 10.3390/ijms23063298
Parsonnet, J., Hickman, R. K., Eardley, D. D., and Pier, G. B. (1985). Induction of human interleukin-l by toxic-shock-syndrome toxin-1. J. Infect. Dis. 151, 514–522. doi: 10.1093/infdis/151.3.514
Pastells, C., Pascual, N., Sanchez-Baeza, F., and Marco, M. P. (2016). Immunochemical determination of pyocyanin and 1-hydroxyphenazine as potential biomarkers of Pseudomonas aeruginosa infections. Anal. Chem. 88, 1631–1638. doi: 10.1021/acs.analchem.5b03490
Pasteur, L. (1882). Remarks on anthracic vaccination as a prophylactic of splenic fever. Br. Med. J. 1:489. doi: 10.1136/bmj.1.1110.489
Peerayeh, S. N., Navidinia, M., Fallah, F., Bakhshi, B., and Jamali, J. (2018). Pathogenicity determinants and epidemiology of uropathogenic E. coli (UPEC) strains isolated from children with urinary tract infection (UTI) to define distinct pathotypes. Biomed. Res. 29, 2035–2043. doi: 10.4066/biomedicalresearch.29-17-1591
Peetermans, M., Verhamme, P., and Vanassche, T. (2015). Coagulase activity by Staphylococcus aureus: A potential target for therapy? Semin. Thromb. Hemost. 41, 433–444. doi: 10.1055/s-0035-1549849
Peeters, E., Nelis, H. J., and Coenye, T. (2008). Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 72, 157–165. doi: 10.1016/j.mimet.2007.11.010
Peña, C., Cabot, G., Gómez-Zorrilla, S., Zamorano, L., Ocampo-Sosa, A., Murillas, J., et al. (2015). Influence of virulence genotype and resistance profile in the mortality of Pseudomonas aeruginosa bloodstream infections. Clin. Infect. Dis. 60, 539–548. doi: 10.1093/cid/ciu866
Penadés, J. R., Chen, J., Quiles-Puchalt, N., Carpena, N., and Novick, R. P. (2015). Bacteriophage-mediated spread of bacterial virulence genes. Curr. Opin. Microbiol. 23, 171–178. doi: 10.1016/j.mib.2014.11.019
Pickrum, A. M., DeLeon, O., Dirck, A., Tessmer, M. H., Riegert, M. O., Biller, J. A., et al. (2020). Achromobacter xylosoxidans cellular pathology is correlated with activation of a type III secretion system. Infect. Immun. 88:e00136–20. doi: 10.1128/IAI.00136-20
Pimenta, A. L., Di Martino, P., and Blight, M. A. (2006). Positive correlation between in vivo and in vitro assays for the evaluation of Pseudomonas virulence. Res. Microbiol. 157, 885–890. doi: 10.1016/j.resmic.2006.06.005
Pinchuk, I. V., Beswick, E. J., and Reyes, V. E. (2010). Staphylococcal enterotoxins. Toxins 2, 2177–2197. doi: 10.3390/toxins2082177
Prasad, R., and Jenq, R. R. (2024). The SagA of E. faecium. eLife 13:e97277. doi: 10.7554/eLife.97277
Rachid, S., Ohlsen, K., Witte, W., Hacker, J., and Ziebuhr, W. (2000). Effect of subinhibitory antibiotic concentrations on polysaccharide intercellular adhesin expression in biofilm-forming Staphylococcus epidermidis. Antimicrob. Agents Chemother. 44, 3357–3363. doi: 10.1128/aac.44.12.3357-3363.2000
Rahimi, E. (2013). Enterotoxigenicity of Staphylococcus aureus isolated from traditional and commercial dairy products marketed in Iran. Braz. J. Microbiol. 44, 393–399. doi: 10.1590/S1517-83822013000200008
Rakovitsky, N., Lellouche, J., Ben David, D., Frenk, S., Elmalih, P., Weber, G., et al. (2021). Increased capsule thickness and hypermotility are traits of carbapenem-resistant Acinetobacter baumannii ST3 strains causing fulminant infection. Open Forum Infect. Dis. 8:ofab386. doi: 10.1093/ofid/ofab386
Rakovitsky, N., Lurie-Weinberger, M. N., Hameir, A., Wulffhart, L., Keren Paz, A., Schwartz, D., et al. (2023). Phenotypic and genomic characterization of nine string-positive carbapenem-resistant Acinetobacter baumannii isolates from Israel. Microbiol. Spectr. 11:e0300222. doi: 10.1128/spectrum.03002-22
Rathnayake, I. U., Hargreaves, M., and Huygens, F. (2012). Antibiotic resistance and virulence traits in clinical and environmental Enterococcus faecalis and Enterococcus faecium isolates. Syst. Appl. Microbiol. 35, 326–333. doi: 10.1016/j.syapm.2012.05.004
Reboud, E., Basso, P., Maillard, A. P., Huber, P., and Attrée, I. (2017). Exolysin shapes the virulence of Pseudomonas aeruginosa clonal outliers. Toxins 9:364. doi: 10.3390/toxins9110364
Ridder, M. J., Daly, S. M., Hall, P. R., and Bose, J. L. (2021). Quantitative hemolysis assays. Methods Mol. Biol. 2341, 25–30. doi: 10.1007/978-1-0716-1550-8_4
Rogers, E. A., Das, A., and Ton-That, H. (2011). “Adhesion by pathogenic corynebacteria,” in Bacterial adhesion. Advances in experimental medicine and biology, Vol. 715, eds D. Linke and A. Goldman (Dordrecht: Springer), 91–103. doi: 10.1007/978-94-007-0940-9_6
Rossi, E., La Rosa, R., and Bartell, J. A. (2021). Pseudomonas aeruginosa adaptation and evolution in patients with cystic fibrosis. Nat. Rev. Microbiol. 19, 331–342. doi: 10.1038/s41579-020-00477-5
Russo, T. A., and Johnson, R. (2000). Proposal for a new inclusive designation for extraintestinal pathogenic isolates of Escherichia coli: ExPEC. J. Infect. Dis. 181, 1753–1754. doi: 10.1086/315418
Russo, T. A., Luke, N. R., Beanan, J. M., Olson, R., Sauberan, S. L., MacDonald, U., et al. (2010). The K1 capsular polysaccharide of Acinetobacter baumannii strain 307-0294 is a major virulence factor. Infect. Immun. 78, 3993–4000. doi: 10.1128/IAI.00366-10
Russo, T. A., Olson, R., Fang, C. T., Stoesser, N., Miller, M., MacDonald, U., et al. (2018). Identification of biomarkers for differentiation of hypervirulent Klebsiella pneumoniae from classical K. pneumoniae. J. Clin. Microbiol. 56:e00776–18. doi: 10.1128/jcm.00776-18
Russo, T. A., Olson, R., MacDonald, U., Beanan, J., and Davidson, B. A. (2015). Aerobactin, but not yersiniabactin, salmochelin, or enterobactin, enables the growth/survival of hypervirulent (hypermucoviscous) Klebsiella pneumoniae ex vivo and in vivo. Infect. Immun. 83, 3325–3333. doi: 10.1128/iai.00430-15
Sánchez-Diener, I., Zamorano, L., Peña, C., Ocampo-Sosa, A., Cabot, G., Gómez-Zorrilla, S., et al. (2020). Weighting the impact of virulence on the outcome of Pseudomonas aeruginosa bloodstream infections. Clin. Microbiol. Infect. 26, 351–357. doi: 10.1016/j.cmi.2019.06.034
Sankomkai, W., Boonyanugomol, W., Kraisriwattana, K., Nutchanon, J., Boonsam, K., Kaewbutra, S., et al. (2020). Characterisation of classical enterotoxins, virulence activity, and antibiotic susceptibility of Staphylococcus aureus isolated from Thai fermented pork sausages, clinical samples, and healthy carriers in northeastern Thailand. J. Vet. Res. 64, 289–297. doi: 10.2478/jvetres-2020-0036
Sarges, E. D. S. N. F., Rodrigues, Y. C., Furlaneto, I. P., de Melo, M. V. H., Brabo, G. L. D. C., Lopes, K. C. M., et al. (2020). Pseudomonas aeruginosa type iii secretion system virulotypes and their association with clinical features of cystic fibrosis patients. Infect. Drug Resist. 13, 3771–3781. doi: 10.2147/IDR.S273759
Sarowska, J., Futoma-Koloch, B., Jama-Kmiecik, A., Frej-Madrzak, M., Ksiazczyk, M., Bugla-Ploskonska, G., et al. (2019). Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 11:10. doi: 10.1186/s13099-019-0290-0
Sato, H., Feix, J. B., and Frank, D. W. (2006). Identification of superoxide dismutase as a cofactor for the Pseudomonas type III toxin, ExoU. Biochemistry 45, 10368–10375. doi: 10.1021/bi060788j
Sato, Y., Unno, Y., Miyazaki, C., Ubagai, T., and Ono, Y. (2019). Multidrug-resistant Acinetobacter baumannii resists reactive oxygen species and survives in macrophages. Sci. Rep. 9:17462. doi: 10.1038/s41598-019-53846-3
Sattler, J., Ernst, C. M., Zweigner, J., and Hamprecht, A. (2024). High frequency of acquired virulence factors in carbapenemase-producing Klebsiella pneumoniae isolates from a large German university hospital, 2013–2021. Antimicrob. Agents Chemother. 68:e0060224. doi: 10.1128/aac.00602-24
Savinova, T., Bocharova, Y., Mayanskiy, N., and Chebotar, I. (2020). The phenomenon of T3SS inactivation for Pseudomonas aeruginosa strains from a chronic infection locus: Do mutations in T3SS-regulators matter? Microbiol. Spectr. 10:e0049422. doi: 10.1128/spectrum.00494-22
Schmidt, H., and Hensel, M. (2004). Pathogenicity islands in bacterial pathogenesis. Clin. Microbiol. Rev. 17, 14–56. doi: 10.1128/cmr.17.1.14-56.2004
Schmitz, F. J., Steiert, M., Hofmann, B., Verhoef, J., Hadding, U., Heinz, H. P., et al. (1998). Development of a multiplex-PCR for direct detection of the genes for enterotoxin B and C, and toxic shock syndrome toxin-1 in Staphylococcus aureus isolates. J. Med. Microbiol. 47, 335–340. doi: 10.1099/00222615-47-4-335
Schwyn, B., and Neilands, J. (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160, 47–56. doi: 10.1016/0003-2697(87)90612-9
Searle, L. J., Méric, G., Porcelli, I., Sheppard, S. K., and Lucchini, S. (2015). Variation in siderophore biosynthetic gene distribution and production across environmental and faecal populations of Escherichia coli. PLoS One 10:e0117906. doi: 10.1371/journal.pone.0117906
Secor, P. R., Burgener, E. B., Kinnersley, M., Jennings, L. K., Roman-Cruz, V., Popescu, M., et al. (2020). Pf bacteriophage and their impact on Pseudomonas virulence, mammalian immunity, and chronic infections. Front. Immunol. 11:244. doi: 10.3389/fimmu.2020.00244
Semedo, T., Santos, M. A., Lopes, M. F., Marques, J. J. F., Crespo, M. T., and Tenreiro, R. (2003). Virulence factors in food, clinical and reference enterococci: A common trait in the genus? Syst. Appl. Microbiol. 26, 13–22. doi: 10.1078/072320203322337263
Shah, C., Baral, R., Bartaula, B., and Shrestha, L. B. (2019). Virulence factors of uropathogenic Escherichia coli (UPEC) and correlation with antimicrobial resistance. BMC Microbiol. 19:204. doi: 10.1186/s12866-019-1587-3
Shallcross, L. J., Fragaszy, E., Johnson, A. M., and Hayward, A. C. (2013). The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect Dis. 13, 43–54. doi: 10.1016/s1473-3099(12)70238-4
Shan, W., Zhang, H., Kan, J., Yin, M., Zhang, J., Wan, L., et al. (2021). Acquired mucoid phenotype of Acinetobacter baumannii: Impact for the molecular characteristics and virulence. Microbiol. Res. 246:126702. doi: 10.1016/j.micres.2021.126702
Sheldon, J. R., and Skaar, E. P. (2020). Acinetobacter baumannii can use multiple siderophores for iron acquisition, but only acinetobactin is required for virulence. PLoS Pathog. 16:e1008995. doi: 10.1371/journal.ppat.1008995
Shigematsu, T., Suda, N., Okuda, K., and Fukushima, J. (2007). Reliable enzyme-linked immunosorbent assay systems for pathogenic factors of Pseudomonas aeruginosa alkaline proteinase, elastase, and exotoxin A: A comparison of methods for labeling detection antibodies with horseradish peroxidase. Microbiol. Immunol. 51, 1149–1459. doi: 10.1111/j.1348-0421.2007.tb04010.x
Silva, N. B. S., Alves, P. G. V., de Andrade Marques, L., Silva, S. F., de Oliveira Faria, G., de Araújo, L. B., et al. (2021). Quantification of biofilm produced by clinical, environment and hands’ isolates Klebsiella species using colorimetric and classical methods. J. Microbiol. Methods 185:106231. doi: 10.1016/j.mimet.2021.106231
Simpson, W. (1665). Zenexton ante-pestilentiale, or, A short discourse of the plague: Its antidotes and cure, according to the placets of the best of physicians, Hippocrates, Paracelsus, and Helmont. London: Printed for George Sawbridge by the Sign of the Bible.
Singh, J. K., Adams, F. G., and Brown, M. H. (2019). Diversity and function of capsular polysaccharide in Acinetobacter baumannii. Front. Microbiol. 9:3301. doi: 10.3389/fmicb.2018.03301
Singh, S. K., Ekka, R., Mishra, M., and Mohapatra, H. (2017). Association study of multiple antibiotic resistance and virulence: A strategy to assess the extent of risk posed by bacterial population in aquatic environment. Environ Monit. Assess. 189:320. doi: 10.1007/s10661-017-6005-4
Sokurenko, E. V., Hasty, D. L., and Dykhuizen, D. E. (1999). Pathoadaptive mutations: Gene loss and variation in bacterial pathogens. Trends Microbiol. 7, 191–195. doi: 10.1016/S0966-842X(99)01493-6
Spanier, J. G., and Cleary, P. P. (1980). Bacteriophage control of antiphagocytic determinants in group A streptococci. J. Exp. Med. 152, 1393–1406. doi: 10.1084/jem.152.5.1393
Spratt, M. R., and Lane, K. (2022). Navigating environmental transitions: The role of phenotypic variation in bacterial responses. mBio 13:e0221222. doi: 10.1128/mbio.02212-22
Spurbeck, R. R., Dinh, P. C. Jr., Walk, S. T., Stapleton, A. E., Hooton, T. M., Nolan, L. K., et al. (2012). Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect. Immun. 80, 4115–4122. doi: 10.1128/IAI.00752-12
Stanton, T. D., and Wyres, K. L. (2024). What defines hypervirulent Klebsiella pneumoniae? EBioMedicine 108:105331. doi: 10.1016/j.ebiom.2024.105331
Stewart, M. K., and Cookson, B. T. (2012). Non-genetic diversity shapes infectious capacity and host resistance. Trends Microbiol. 20, 461–466. doi: 10.1016/j.tim.2012.07.003
Stirling, F. R., Cuzick, A., Kelly, S. M., Oxley, D., and Evans, T. J. (2006). Eukaryotic localization, activation and ubiquitinylation of a bacterial type III secreted toxin. Cell Microbiol. 8, 1294–1309. doi: 10.1111/j.1462-5822.2006.00710.x
Stulik, L., Malafa, S., Hudcova, J., Rouha, H., Henics, B. Z., Craven, D. E., et al. (2014). α-Hemolysin activity of methicillin-susceptible Staphylococcus aureus predicts ventilator-associated pneumonia. Am. J. Respir. Crit. Care Med. 190, 1139–1148. doi: 10.1164/rccm.201406-1012oc
Tahmasebi, H., Dehbashi, S., Nasaj, M., and Arabestani, M. R. (2022). Molecular epidemiology and collaboration of siderophore-based iron acquisition with surface adhesion in hypervirulent Pseudomonas aeruginosa isolates from wound infections. Sci. Rep. 12:7791. doi: 10.1038/s41598-022-11984-1
Tang, A., Caballero, A. R., Bierdeman, M. A., Marquart, M. E., Foster, T. J., Monk, I. R., et al. (2019). Staphylococcus aureus superantigen-like protein SSL1: A toxic protease. Pathogens 8:2. doi: 10.3390/pathogens8010002
Tang, W., Liu, Y., Li, X., Leng, G., Gao, J., Wang, Y., et al. (2024). Microbiological characteristics of clinically isolated Staphylococcus aureus with different hemolytic phenotypes in China. Infect. Drug Resist. 17, 3273–3287. doi: 10.2147/IDR.S466416
Taylor, V. L., Fitzpatrick, A. D., Islam, Z., and Maxwell, K. L. (2019). The diverse impacts of phage morons on bacterial fitness and virulence. Adv. Virus Res. 103, 1–31. doi: 10.1016/bs.aivir.2018.08.001
Teng, F., Kawalec, M., Weinstock, G. M., Hryniewicz, W., and Murray, B. E. (2003). An Enterococcus faecium secreted antigen, SagA, exhibits broad-spectrum binding to extracellular matrix proteins and appears essential for E. faecium growth. Infect. Immun. 71, 5033–5041. doi: 10.1128/IAI.71.9.5033-5041.2003
Tessmer, M. H., Anderson, D. M., Pickrum, A. M., Riegert, M. O., and Frank, D. W. (2019). Identification and verification of ubiquitin-activated bacterial phospholipases. J. Bacteriol. 201:e00623–18. doi: 10.1128/JB.00623-18
The Virulence Factor Database (2025). Virulence factors of pathogenic bacteria. Available online at: https://www.mgc.ac.cn/VFs/search_VFs.htm (accessed April 2, 2025).
Thi, M. T. T., Wibowo, D., and Rehm, B. H. (2020). Pseudomonas aeruginosa biofilms. Int. J. Mol. Sci. 21:8671. doi: 10.3390/ijms21228671
Toledo-Arana, A., Valle, J., Solano, C., Arrizubieta, M. J., Cucarella, C., Lamata, M., et al. (2001). The enterococcal surface protein, Esp, is involved in Enterococcus faecalis biofilm formation. Appl. Environ. Microbiol. 67, 4538–4545. doi: 10.1128/AEM.67.10.4538-4545.2001
Turton, J. F., Baklan, H., Siu, L. K., Kaufmann, M. E., and Pitt, T. L. (2008). Evaluation of a multiplex PCR for detection of serotypes K1, K2 and K5 in Klebsiella sp. and comparison of isolates within these serotypes. FEMS Microbiol. Lett. 284, 247–252. doi: 10.1111/j.1574-6968.2008.01208.x
Turton, J. F., Perry, C., Elgohari, S., and Hampton, C. V. (2010). PCR characterization and typing of Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and virulence gene targets. J. Med. Microbiol. 59, 541–547. doi: 10.1099/jmm.0.015198-0
Vankerckhoven, V., Van Autgaerden, T., Vael, C., Lammens, C., Chapelle, S., Rossi, R., et al. (2004). Development of a multiplex pcr for the detection of asa1, gelE, cylA, esp, and hyl genes in enterococci and survey for virulence determinants among European hospital isolates of Enterococcus faecium. J. Clin. Microbiol. 42, 4473–4479. doi: 10.1128/jcm.42.10.4473-4479.2004
Veening, J. W., Smits, W. K., and Kuipers, O. P. (2008). Bistability, epigenetics, and bet-hedging in bacteria. Annu. Rev. Microbiol. 62, 193–210. doi: 10.1146/annurev.micro.62.081307.163002
Vijayaraghavan, P., and Vincent, S. G. (2013). A simple method for the detection of protease activity on agar plates using bromocresolgreen dye. J. Biochem Technol. 4, 628–630.
Wadsworth, A. B., and Kirkbride, M. B. (1918). A study of the changes in virulence of the pneumococcus at different periods of growth and under different conditions of cultivation in media. J. Exp. Med. 28, 791–821. doi: 10.1084/jem.28.6.791
Waldor, M. K. (1998). Bacteriophage biology and bacterial virulence. Trends Microbiol. 6, 295–297. doi: 10.1016/s0966-842x(98)01320-1
Waldor, M. K., and Friedman, D. I. (2005). Phage regulatory circuits and virulence gene expression. Curr. Opin. Microbiol. 8, 459–465. doi: 10.1016/j.mib.2005.06.001
Wand, M. E., Bock, L. J., Bonney, L. C., and Sutton, J. M. (2015). Retention of virulence following adaptation to colistin in Acinetobacter baumannii reflects the mechanism of resistance. J. Antimicrob. Chemother. 70, 2209–2216. doi: 10.1093/jac/dkv097
Wang, B., Farhan, M. H. R., Yuan, L., Sui, Y., Chu, J., Yang, X., et al. (2024). Transfer dynamics of antimicrobial resistance among gram-negative bacteria. Sci. Total Environ. 954:176347. doi: 10.1016/j.scitotenv.2024.176347
Wassenaar, T. M., and Gaastra, W. (2001). Bacterial virulence: Can we draw the line? FEMS Microbiol. Lett. 201, 1–7. doi: 10.1111/j.1574-6968.2001.tb10724.x
Weerasekera, D., Möller, J., Kraner, M. E., Azevedo Antunes, C., Mattos-Guaraldi, A. L., and Burkovski, A. (2019). Beyond diphtheria toxin: Cytotoxic proteins of Corynebacterium ulcerans and Corynebacterium diphtheriae. Microbiology 165, 876–890. doi: 10.1099/mic.0.000820
Wiens, J. R., Vasil, A. I., Schurr, M. J., and Vasil, M. L. (2014). Iron-regulated expression of alginate production, mucoid phenotype, and biofilm formation by Pseudomonas aeruginosa. mBio 5:e01010–13. doi: 10.1128/mBio.01010-13
Wikraiphat, C., Charoensap, J., Utaisincharoen, P., Wongratanacheewin, S., Taweechaisupapong, S., Woods, D. E., et al. (2009). Comparative in vivo and in vitro analyses of putative virulence factors of Burkholderia pseudomallei using lipopolysaccharide, capsule and flagellin mutants. FEMS Immunol. Med. Microbiol. 56, 253–259. doi: 10.1111/j.1574-695X.2009.00574.x
Williamson, D. A., Mills, G., Johnson, J. R., Porter, S., and Wiles, S. (2014). In vivo correlates of molecularly inferred virulence among extraintestinal pathogenic Escherichia coli (ExPEC) in the wax moth Galleria mellonella model system. Virulence 5, 388–393. doi: 10.4161/viru.27912
Wolff, N., Hendling, M., Schroeder, F., Schönthaler, S., Geiss, A. F., Bedenic, B., et al. (2021). Full pathogen characterisation: Species identification including the detection of virulence factors and antibiotic resistance genes via multiplex DNA-assays. Sci. Rep. 11:6001. doi: 10.1038/s41598-021-85438-5
Wyres, K. L., Cahill, S. M., Holt, K. E., Hall, R. M., and Kenyon, J. J. (2020). Identification of Acinetobacter baumannii loci for capsular polysaccharide (KL) and lipooligosaccharide outer core (OCL) synthesis in genome assemblies using curated reference databases compatible with Kaptive. Microb. Genom. 6:e000339. doi: 10.1099/mgen.0.000339
Yang, C. H., Su, P. W., Moi, S. H., and Chuang, L. Y. (2019). Biofilm formation in Acinetobacter baumannii: Genotype-phenotype correlation. Molecules 24:1849. doi: 10.3390/molecules24101849
Yang, J. L., Yang, C. J., Chuang, Y. C., Sheng, W. H., Chen, Y. C., and Chang, S. C. (2022). Association of capsular polysaccharide locus 2 with prognosis of Acinetobacter baumannii bacteraemia. Emerg. Microbes Infect. 11, 83–90. doi: 10.1080/22221751.2021.2011624
Yang, Y. X., Xu, Z. H., Zhang, Y. Q., Tian, J., Weng, L. X., and Wang, L. H. (2012). A new quorum-sensing inhibitor attenuates virulence and decreases antibiotic resistance in Pseudomonas aeruginosa. J. Microbiol. 50, 987–993. doi: 10.1007/s12275-012-2149-7
Yasui, Y. (2022). Evolutionary bet-hedging reconsidered: What is the mean–variance trade-off of fitness? Ecol. Res. 37, 406–420. doi: 10.1111/1440-1703.12303
Zhang, A., Veesenmeyer, J. L., and Hauser, A. R. (2017). Phosphatidylinositol 4,5-bisphosphate-dependent oligomerization of the Pseudomonas aeruginosa cytotoxin ExoU. Infect. Immun. 86, e402–e417. doi: 10.1128/IAI.00402-17
Zhou, J., Ma, H., and Zhang, L. (2023). Mechanisms of virulence reprogramming in bacterial pathogens. Annu. Rev. Microbiol. 77, 561–581. doi: 10.1146/annurev-micro-032521-025954
Zhu, Z., Hu, Z., Li, S., Fang, R., Ono, H. K., and Hu, D. L. (2023). Molecular characteristics and pathogenicity of Staphylococcus aureus exotoxins. Int. J. Mol. Sci. 25:395. doi: 10.3390/ijms25010395
Keywords: bacterial infection, human opportunistic pathogen, virulence mechanisms, detection of virulence factors, bacterial fitness
Citation: Chebotar I, Azizov I, Edelstein M and Kozlov R (2025) Detection of virulence factors in opportunistic bacteria: advances, challenges, and practical implementation. Front. Microbiol. 16:1638925. doi: 10.3389/fmicb.2025.1638925
Received: 31 May 2025; Accepted: 03 September 2025;
Published: 17 September 2025.
Edited by:
Axel Cloeckaert, Institut national de recherche pour l’agriculture, l’alimentation et l’environnement (INRAE), FranceReviewed by:
Roberto Mauricio Vidal, University of Chile, ChileLia-Mara Ditu, University of Bucharest, Romania
Gene D. Godbold, Signature Science, LLC, United States
Copyright © 2025 Chebotar, Azizov, Edelstein and Kozlov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Igor Chebotar, bGFiMTIzNDU2MTdAZ21haWwuY29t