 Catarina Morais1
Catarina Morais1 Sofia Santos Costa1
Sofia Santos Costa1 Dennis Hanke2,3Ana Santos1
Dennis Hanke2,3Ana Santos1 Henrike Krüger-Haker2,3
Henrike Krüger-Haker2,3 Constança Pomba4
Constança Pomba4 Andrea T. Feßler2,3
Andrea T. Feßler2,3 Stefan Schwarz2,3
Stefan Schwarz2,3 Isabel Couto1*
Isabel Couto1*- 1Global Health and Tropical Medicine, GHTM, LA-REAL, Instituto de Higiene e Medicina Tropical, IHMT, Universidade NOVA de Lisboa, Lisbon, Portugal
- 2Institute of Microbiology and Epizootics, Center for Infection Medicine, School of Veterinary Medicine, Freie Universität Berlin, Berlin, Germany
- 3Veterinary Centre for Resistance Research (TZR), School of Veterinary Medicine, Freie Universität Berlin, Berlin, Germany
- 4Laboratory of Antimicrobial Resistance, CIISA, Faculty of Veterinary Medicine, University of Lisbon, Lisbon, Portugal
The increasing antimicrobial resistance (AMR) in Staphylococcus pseudintermedius causing skin and soft-tissue infections (SSTIs) in companion animals is a public health concern. The aim of this study was to verify if mobile genetic elements (MGEs), in particular plasmids, are related to the carriage of AMR genes among circulating and clinically relevant S. pseudintermedius. In total, 56 S. pseudintermedius, representing predominant and emerging clonal lineages associated with SSTIs in dogs and cats collected in Lisbon (Portugal), were subjected to plasmid DNA extraction and digestion with EcoRI and XbaI. Each unique restriction pattern was assigned to a plasmid profile. A subset of 17 strains was further selected for hybrid whole genome sequencing (WGS) on Oxford Nanopore MinION and Illumina MiSeq platforms. Thirty-one of the 56 S. pseudintermedius strains carried one or more plasmid(s), mostly of small or medium sizes, corresponding to eight plasmid profiles. Two of the identified plasmids carried AMR determinants; plasmid pSP-G3C4, isolated from ST71 strains, carried the tetracycline resistance gene tet(K) and plasmid pSP5912, isolated from a ST2061 strain, harbored the qacG biocide resistance gene. Other AMR determinants were detected as part of MGEs integrated into the bacterial chromosomal DNA, namely Tn552, Tn552-like, Tn553, Tn916, Tn5405-like, Tn5801, Tn5801-like GI6287 and pRE25-like elements. In addition, a new chromosomal cassette, carrying fusC, was identified in a ST1183 strain. The 12 methicillin-resistant S. pseudintermedius studied carried staphylococcal cassette chromosome mec (SCCmec) type III (n = 5), SCCmec type IVg (n = 3), SCCmecNA45 (n = 1), ΨSCCmec57395 (n = 1), the recently described cassettes SCCmec7017–61515 (n = 1), or SCCmec type V(T)SL/154 (n = 1). Most strains carried intact prophages without AMR determinants. Intact restriction-modification systems were detected in 12 out of the 17 strains and CRISPR/Cas in five strains, four of which were methicillin-susceptible. The results of this study suggest that the AMR content in S. pseudintermedius is mainly related to MGEs integrated into the chromosomal DNA rather than located on plasmids. These results provide important insights that may lead to a better understanding of multidrug resistance in S. pseudintermedius towards improved SSTIs treatment in companion animals.
1 Introduction
Staphylococcus pseudintermedius is the most common pathogen associated with skin and soft-tissue infections (SSTIs) in companion animals (Lynch and Helbig, 2021), among which canine pyoderma is the most relevant. The recently updated guidelines for canine pyoderma treatment indicate that the first-line therapy for surface and superficial pyoderma is based on biocides or topical antimicrobials, when necessary (Loeffler et al., 2025). For systemic infections, the treatment includes clindamycin, lincomycin, amoxicillin–clavulanate, or first generation cephalosporins (first-line), fluoroquinolones, tetracyclines or trimethoprim-sulfamethoxazole (second-line) (Loeffler et al., 2025). For cats it was also recommended to apply biocides and the systemic use of amoxicillin–clavulanate, clindamycin or cefovecin (Wildermuth et al., 2006; Miller et al., 2023).
We recently characterized a collection of 155 S. pseudintermedius strains, obtained from SSTIs in companion animals between 2014 and 2018 in Lisbon (Portugal), regarding antimicrobial resistance (AMR) profiles and clonal lineages (Morais et al., 2023). In that earlier study, 45.2% of the strains had a multidrug resistance (MDR) profile, corresponding to resistance to at least one antimicrobial of three different classes (Sweeney et al., 2018), and about a third (31.0%) were methicillin-resistant S. pseudintermedius (MRSP). High rates of resistance were observed to most of the first- and second-line therapeutical antimicrobial agents, following the data from other studies (Feßler et al., 2022; Adiguzel et al., 2022; Afshar et al., 2023; Robb et al., 2024; Calabro et al., 2024). In addition, we detected strains resistant to fusidic acid and rifampicin (Morais et al., 2023). Fusidic acid is a topical antimicrobial agent approved for human and veterinary applications in Europe for the treatment of methicillin-resistant staphylococcal infections (Morris et al., 2017; Loeffler et al., 2025). Rifampicin, an ansamycin, which is part of the first-line treatment of tuberculosis in humans, is indicated for canine pyoderma caused by bacteria resistant to first-line therapy (systemic and topic) (Hillier et al., 2014; Miller et al., 2023; Loeffler et al., 2025) or by MRSP strains with a MDR phenotype (Hicks et al., 2021; Harbour et al., 2022; Loeffler et al., 2025). However, S. pseudintermedius rapidly develops rifampicin resistance (Kadlec et al., 2011; Hicks et al., 2021) and nowadays, this antimicrobial is considered “reserved” (Loeffler et al., 2025). Regarding the S. pseudintermedius clonal lineages circulating in Portugal, our previous study indicated that sequence type (ST) 71 remained the most frequent clonal lineage, associated with methicillin resistance and MDR profiles. Several new clonal lineages (ST258, ST551, ST241 and ST265) were also identified for the first time in Portugal (Morais et al., 2023).
Antimicrobial resistance (AMR) genes can be integrated into the chromosomal DNA or in mobile genetic elements (MGEs) like plasmids, bacteriophages, staphylococcal cassette chromosome (SCC) elements, and transposons. MGEs have been linked to clonal expansion and evolution of different bacteria, including Staphylococcus aureus (Brooks et al., 2020). S. pseudintermedius has an open pangenome with a significant presence of accessory genes, which generally correspond to MGEs (Brooks et al., 2020; Fàbregas et al., 2023; Grist et al., 2025). The presence of AMR genes in this species has been mainly correlated to the carriage of transposons, such as Tn552 (blaZ), Tn917 [erm(B)], Tn5405-like (aadE, sat4, aphA3) and Tn916 [tet(M)] (Kadlec and Schwarz, 2012; Phumthanakorn et al., 2021). However, albeit in lower frequency, some AMR genes were also found on plasmids, like pSTS2 carrying the tet(K) gene or plasmids pSCS1, pSCS11 (Greene and Schwarz, 1992) and pSCS20-23 carrying the cat gene (Schwarz et al., 1995). While phages represent one of the most relevant mechanisms for DNA transfer in S. pseudintermedius (Brooks et al., 2020), AMR or virulence genes are not frequently found in these MGEs. In S. aureus, phages usually also do not carry AMR genes (Haaber et al., 2017), but they allow the mobility of pathogenicity islands and plasmids carrying AMR genes by transduction (Malachowa and DeLeo, 2010).
In the current study, we aimed at further analyzing representative strains from the Lisbon collection (Morais et al., 2023) and, through whole genome sequencing (WGS), obtaining information about the role of the staphylococcal mobilome in the carriage of AMR among circulating and clinically relevant S. pseudintermedius lineages.
2 Material and methods
2.1 Bacterial collection
The study collection comprised 56 S. pseudintermedius strains obtained from SSTIs in companion animals (53 dogs and 3 cats). These 56 strains, described in Supplementary Table 1, were selected from the collection of 155 S. pseudintermedius previously characterized (Morais et al., 2023), according to the following criteria: (i) all strains from ST71 and ST157, which correspond to the two most frequent STs in the collection; (ii) strains from relevant STs in the European context (ST45, ST118, ST241, ST258, ST265 and ST551); (iii) strains with phenotypes of interest, namely resistance to fusidic acid, tetracycline or rifampicin.
2.2 Plasmid DNA profiling
Plasmid DNA (pDNA) was extracted with the NZYMiniprep kit (NZYtech, Portugal) or QIAGEN Plasmid Midi Kit (Qiagen, Germany), adding 35 μg/mL of lysostaphin (Sigma-Aldrich, Missouri, USA) in the cell lysis step, followed by incubation at 37 oC for 90–120 min. Plasmids were classified according to their migration in the gel before and after digestion with XbaI and EcoRI restriction enzymes (NZYtech), as small (≤ 5 kb), medium (> 5 kb and < 23 kb), or large (≥ 23 kb) plasmids, using the weight markers Lambda DNA/HindIII Ladder and GeneRuler 1 kb DNA Ladder (Thermo Fisher Scientific, Waltham, USA). Each unique restriction pattern was assigned to a plasmid profile, later confirmed by WGS data.
2.3 Genomic DNA extraction and whole genome sequencing (WGS)
A subset of 17 strains out of the 56 representative S. pseudintermedius was selected for WGS analysis by a hybrid approach with Oxford Nanopore and Illumina technologies, generating long-reads and short paired-end reads, respectively (Supplementary Table 1). These 17 strains were selected by the following criteria (i) at least one strain from each plasmid profile; (ii) at least one strain per lineage; (iii) all fusidic acid and rifampicin resistant strains, excluding one rifampicin strain (BIOS-V241) sharing the plasmid profile and lineage of BIOS-V240 (sequenced); (iv) strains from the predominant lineages without plasmids.
Genomic DNA was obtained from 1 mL of overnight culture (Tryptic Soy Broth at 37 oC) using the MagAttract HMW DNA Kit (Qiagen) following the manufacturer’s protocol. The Native Barcoding Kit-24 (SQK-NBD112-24, Oxford Nanopore Technologies, Oxford, UK) was used to prepare the sequencing libraries with 400 ng of DNA for MinION. The barcoded libraries were pooled, to carry out multiplexed sequencing, and loaded onto a MinION FLO-MIN106 flow cell v9.4.1 and sequenced in a MinION Mk1C. For Illumina, the libraries were prepared using the Nextera XT DNA Library Preparation Kit (Illumina, Inc., San Diego, USA) according to the manufacturer’s recommendations. The 2 × 300-bp paired-end sequencing in 40-fold multiplexes was performed on the Illumina MiSeq platform with the MiSeq Reagent Kit v3 (Illumina). DNA quantification was carried out using the Qubit™ 4 fluorometer (Invitrogen, NY, USA) with the Qubit™ dsDNA HS assay kit (Invitrogen). Base-calling and demultiplexing of MinION read files were conducted via MinKNOW v23.04.5 and Porechop v0.2.4, respectively. The quality of the long-reads was assessed in LongQC v1.2.0c (Fukasawa et al., 2020) and short fragments with low quality were removed with Filtlong v0.2.1. Short-reads were trimmed with TrimGalore (RRID:SCR_011847) v0.6.10 and their quality assessed through FastQC v0.12.01.
2.4 Genome assembly and annotation
Genomes were de novo assembled with Flye v2.9.3 (Kolmogorov et al., 2019) and polished with NextPolish v1.4.1 (Hu et al., 2020). The results obtained from Flye were compared in Geneious Prime v8.1.9 (Biomatters, Ltd., Auckland, New Zealand) to de novo assemblies performed with Unicycler v0.4.9 (Wick et al., 2017) and MaSuRCA v4.1.0 (Zimin et al., 2017). Annotation was accomplished with Bakta v1.8.2 (Schwengers et al., 2021). The genome completeness was analyzed with the Benchmarking Universal Single-Copy Orthologs (BUSCO) tool (Manni et al., 2021). Extra-chromosomal contigs assembled with Flye were identified as reflecting possible plasmids according to the size, the presence of a rep gene, coverage and circularity in Bandage v0.8.1 (Wick et al., 2015). BLASTn NCBI (Camacho et al., 2009) was used to determine the homology to other plasmids as previously described.
2.5 Antimicrobial resistance (AMR) genes detection
In silico screening for the presence of acquired AMR genes and point mutations was performed through the Comprehensive Antibiotic Resistance Database (CARD) (Jia et al., 2017; Alcock et al., 2023) and ResFinder v4.5.0 (Camacho et al., 2009; Bortolaia et al., 2020). The presence of point mutations in fusA and rpoB genes was confirmed by aligning the sequences with the genes of known susceptible S. pseudintermedius strains: HKU10-03 (accession no.: NC_014925.1) and FDAARGOS_930 (accession no: NZ_CP065635), also deposited at DSMZ repository as S. pseudintermedius strain DSM21284T.
2.6 Mobile genetic elements identification
Mobile genetic elements were identified using bioinformatics tools available online. Plasmids were predicted with PlasmidFinder v2.1 (Camacho et al., 2009; Carattoli et al., 2014), transposons and insertion sequences with MobileElementFinder v1.0.3 (Johansson et al., 2021), both available at the Center for Genomic Epidemiology2. PHIGARO (Starikova et al., 2020) was used to determine the content of prophages in the studied genomes and PHASTEST v3.0 web server (Zhou et al., 2011; Arndt et al., 2016) to identify and classify them as intact, questionable or incomplete prophages. All identified prophages were BLASTn searched against the NCBI Virus database. Staphylococcal cassette chromosome mec (SCCmec) was first screened with SCCmecFinder 1.23, which is available for S. aureus. The results obtained were then compared with the whole genome sequence of the strain, identifying putative integration site sequences (ISSs) for SCCmec as described previously for S. aureus (Ito et al., 2004) and S. pseudintermedius (Perreten et al., 2013). A BLASTn search was performed on the resulting DNA sequence to identify the most similar SCCmec type described for S. pseudintermedius. These putative ISSs were also used to delimit SCCfus.
2.7 Identification of restriction-modification systems and clustered regularly interspaced short palindromic repeats
Restriction-modification (R-M) systems were predicted using the information provided by the rmsFinder tool (Roberts et al., 2015) and REBASE database4. Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR) sequences and Cas proteins were detected with CRISPRCasFinder5 (Couvin et al., 2018), using the software default parameters. The classification provided by the software was compared with the classification suggested by Rossi et al. (2019). Only the complete systems, containing both CRISPR and Cas proteins, were considered for analysis.
3 Results
3.1 Plasmid profiling
A first set of 56 representative S. pseudintermedius strains was selected for analysis of plasmid content. Plasmid DNA extraction revealed the presence of plasmids in 31 out of the 56 strains screened, mostly of small or medium size. These corresponded to eight distinct plasmid profiles (P1–P8) (Table 1). Plasmid profile P1 was the most frequently detected (16/31). Plasmid profile P2 included two different plasmids with indistinguishable restriction profiles, size and main features that were carried by strains from different STs. We also found strains from the same ST carrying different plasmids as well as the same plasmid carried by strains from different lineages (pBIOS236). No plasmids were detected in strains from lineages ST25, ST45, ST157, ST265, ST422, ST497, ST924, ST2055, ST2059, ST2099 and ST2102 (Table 1).
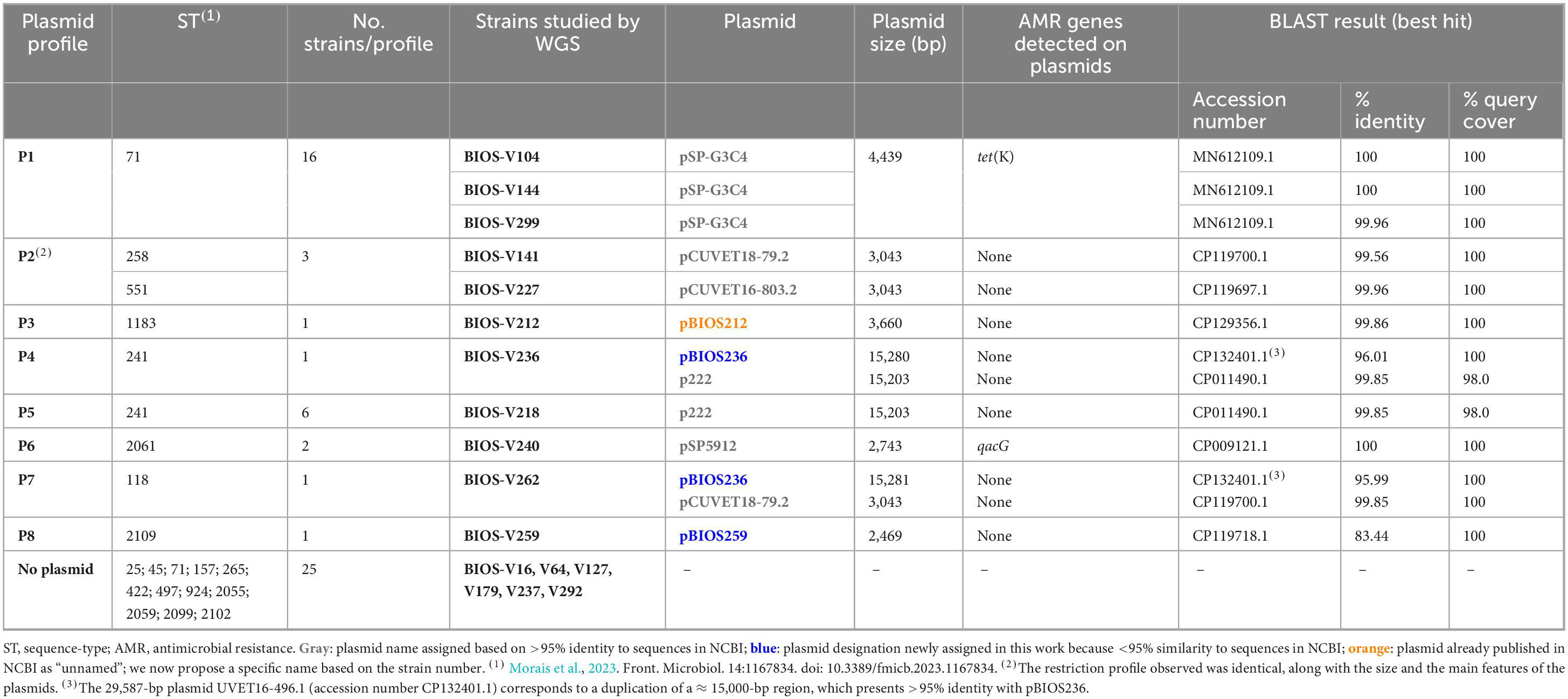
Table 1. Plasmid profiles determined among the 56 representative S. pseudintermedius strains, detailed through whole genome sequencing (WGS) analysis.
3.2 Genomic characterization of S. pseudintermedius by WGS
A subset of 17 strains, selected according to plasmid profile and clonal lineage and including 12 MRSP strains and five methicillin-susceptible (MSSP) strains, was further characterized by hybrid WGS (Figure 1).
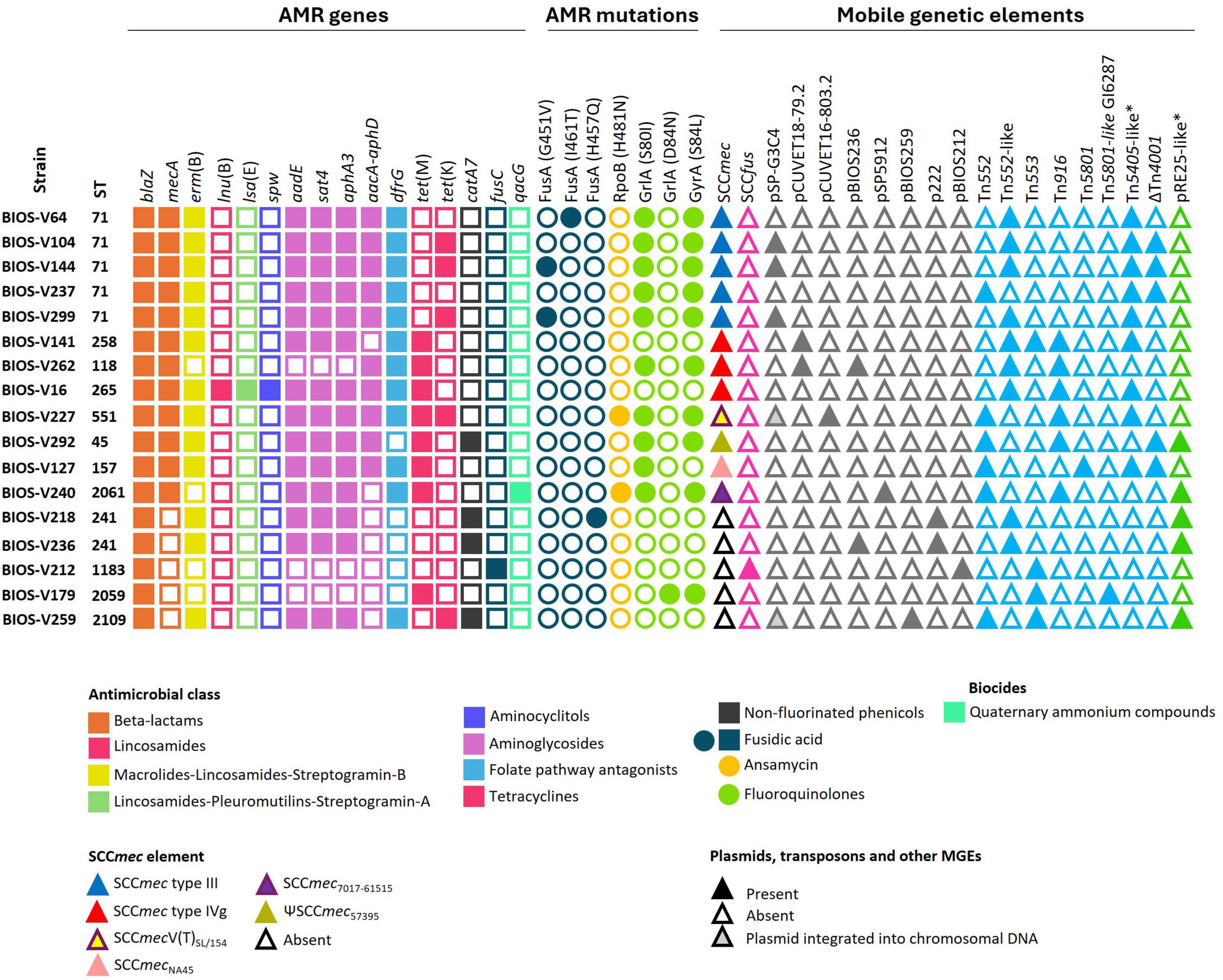
Figure 1. Clonal lineage, antimicrobial resistance (AMR) determinants and mobile genetic elements (MGEs) of the 12 MRSP and 5 MSSP strains selected for whole genome sequencing. Squares and circles correspond to AMR genes and point mutations, respectively. Triangles correspond to MGEs. *Include variants of this element carrying different AMR genes (see Table 3).
The de novo assembly of the 17 genomes resulted in closed circular chromosomes (99.3%–99.6% completeness), with a GC content of about 37.5%. The size of the genomes varied between 2.5 and 2.9 Mbp, with 0–2 plasmids. Interestingly, the GC content of all plasmids identified in this work was lower, ranging between 28.1 and 33.4%. Detailed information on the WGS data is presented in Table 2.
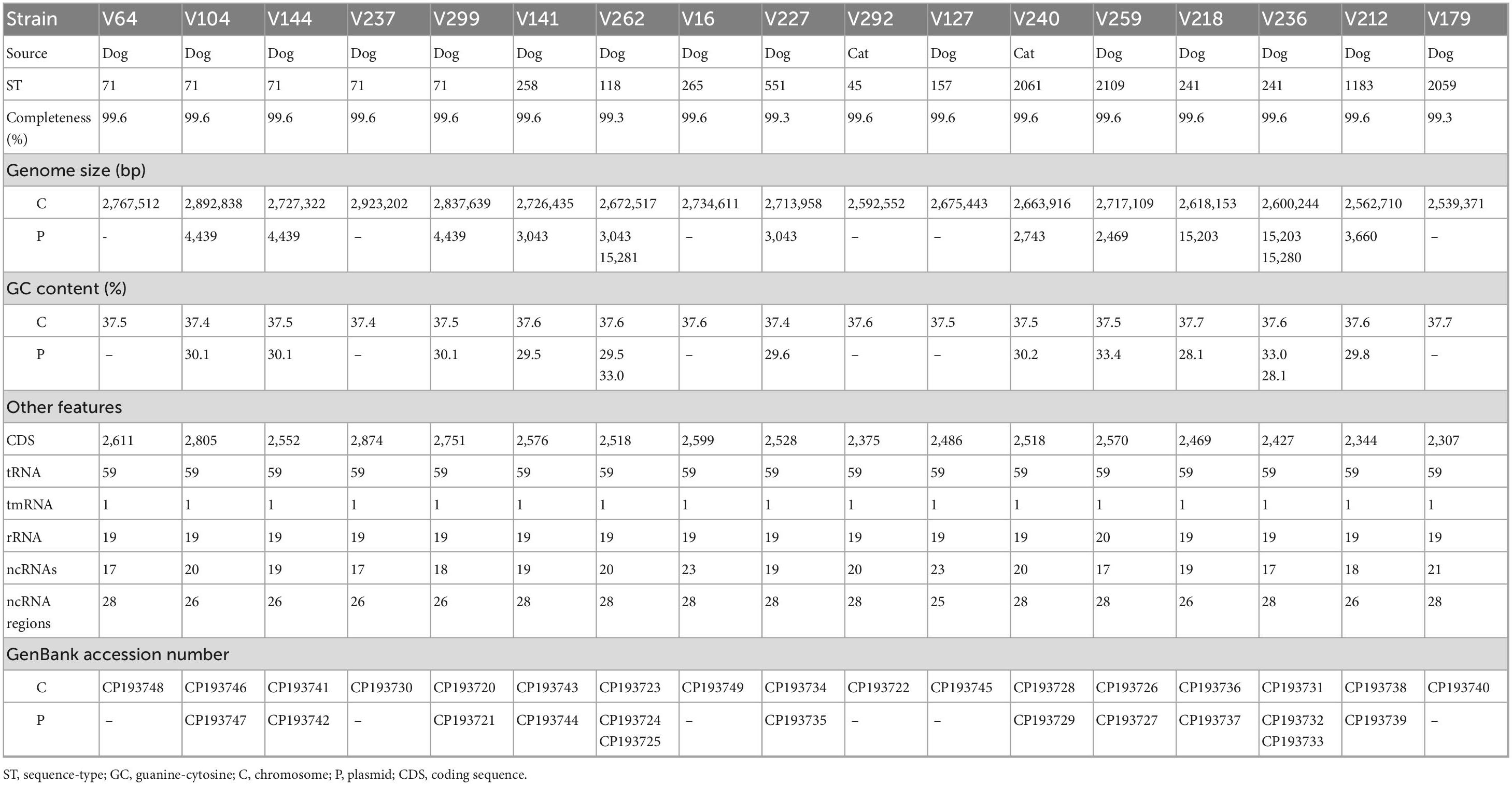
Table 2. Characteristics of the 17 S. pseudintermedius strains and respective genomes studied by whole genome sequencing.
3.2.1 Identification of AMR determinants
In the previous study, the strain collection was characterized regarding antimicrobial susceptibility phenotypes by disk diffusion and PCR screening of several AMR genes (Morais et al., 2023; Supplementary Table 1). WGS analysis allowed the identification of additional resistance genes in the chromosomal DNA of some strains, namely aadE, sat4, lsa(E), and lnu(B) (Figure 1). The fosB gene, related to fosfomycin resistance in S. aureus (Fu et al., 2016) was present in all 17 strains. Mutations in the quinolone resistance determining regions (QRDR) of the target genes grlA and gyrA, and corresponding amino acid exchanges, were previously identified (Morais et al., 2023) and now confirmed through WGS. Four of the five strains resistant to fusidic acid carried mutations in the fusA gene that resulted in the amino acid exchanges G451V, H457Q or I461T in FusA. The remaining strain carried the fusC gene. Rifampicin resistance was associated with a mutation in the rpoB gene that resulted in the amino acid exchange H481N in RpoB of the two resistant strains sequenced (BIOS-V227 and BIOS-V240) (Figure 1). Regarding tetracycline resistance, the determinants previously detected, tet(M) and tet(K) (Morais et al., 2023), were now found located either in the chromosomal DNA [tet(M)] or on free or integrated plasmids [tet(K)], in different combinations, as detailed below.
3.2.2 Mobile genetic elements and AMR genes
Tables 1, 3–5, and Figure 1 detail the distribution of AMR genes and MGEs identified in the genomes of the 17 sequenced strains.
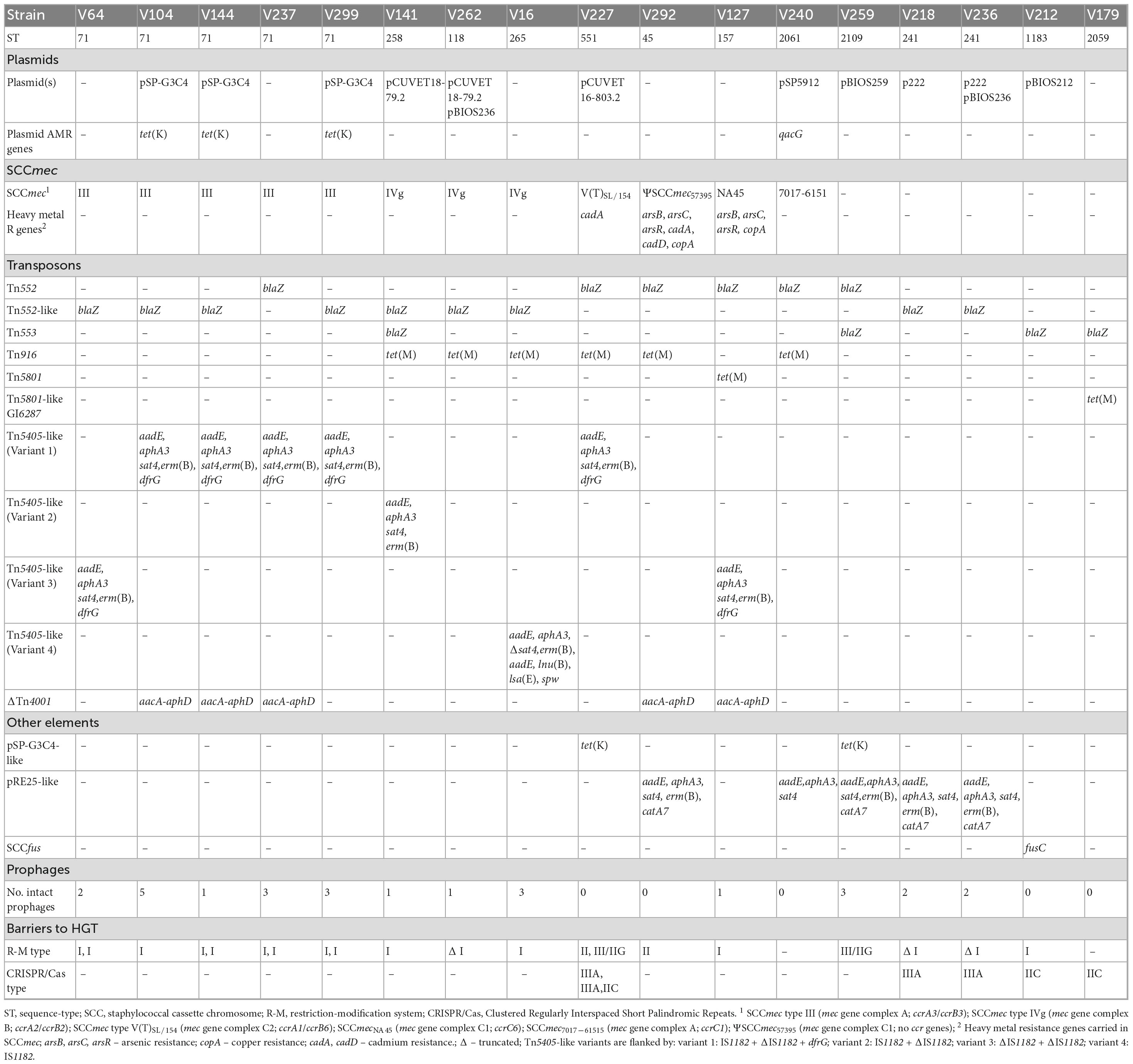
Table 3. Antimicrobial resistance (AMR) genes carried by mobile genetic elements and barriers to horizontal gene transfer (HGT) identified in S. pseudintermedius strains.
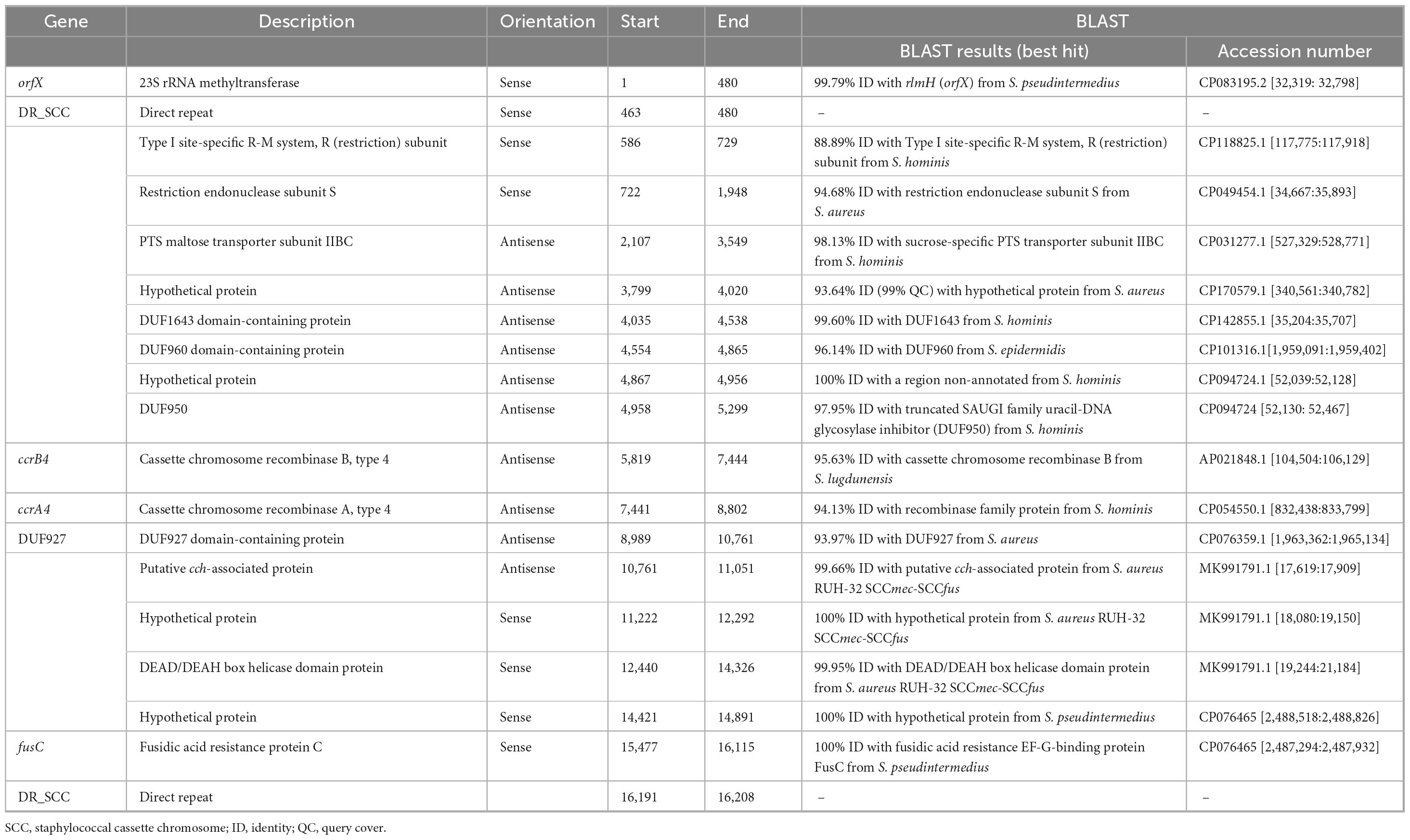
Table 4. Description of the SCCfus element identified in S. pseudintermedius strain BIOS-V212 (lineage ST1183).
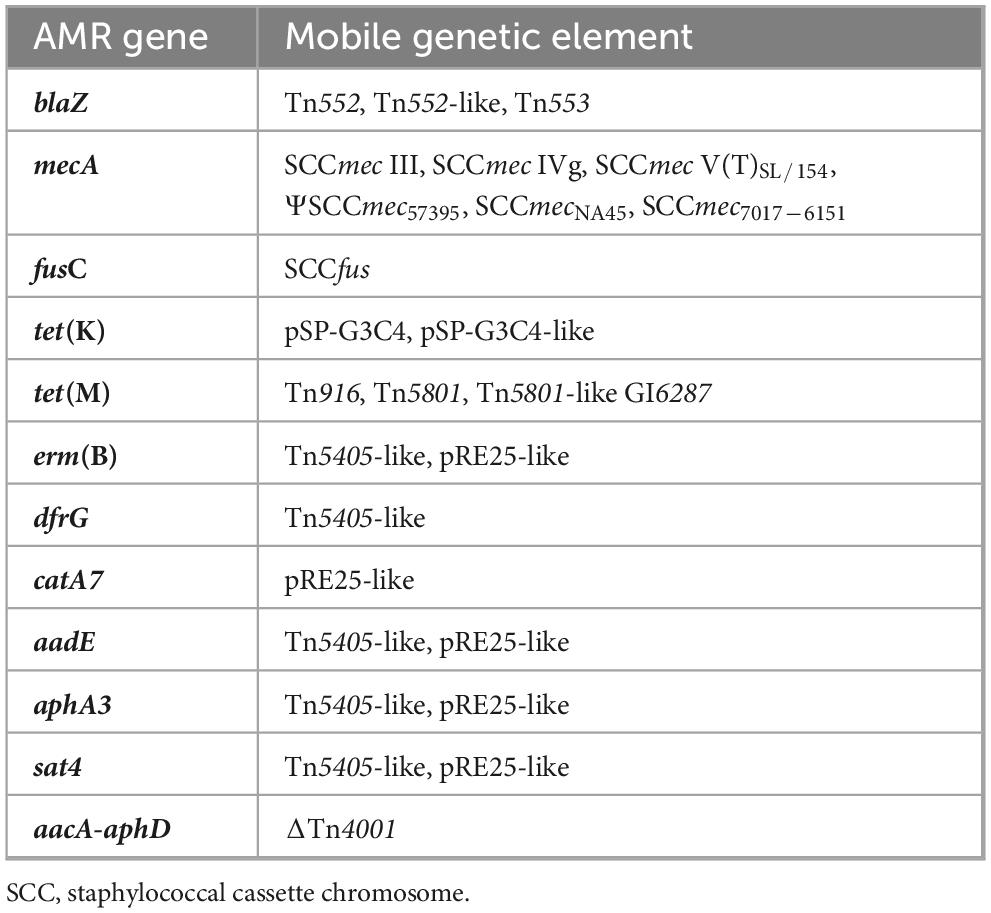
Table 5. Antimicrobial resistance (AMR) genes found in S. pseudintermedius mobile genetic elements (MGEs). All these MGEs were detected integrated into chromosomal DNA, except the plasmid pSP-G3C4.
3.2.2.1 Plasmids
Eleven out of the 17 sequenced strains carried eight different plasmids: pSP-G3C4, pCUVET18-79.2, pCUVET16-803.2, pSP5912, pBIOS212, p222, and the newly described pBIOS236 and pBIOS259. The sizes of these plasmids varied between 2,469 and 15,281 bp. Nine strains carried a single plasmid, and the remaining two (BIOS-V236 and BIOS-V262), had two plasmids (Table 1).
Regarding carriage of AMR genes, the 4,439-bp plasmid pSP-G3C4 shares 100% identity with a tet(K)-harboring plasmid from S. pseudintermedius strain G3C4, responsible for tetracycline resistance through increased efflux activity. The 2,743-bp S. pseudintermedius plasmid pSP5912 carried the qacG gene, associated with the efflux of quaternary ammonium compounds.
The 15,203-bp plasmid p222, carried by strains BIOS-V218 and BIOS-V236, has 99.85% identity with S. pseudintermedius plasmid p222 and includes the gene for the virulence factor bacteriocin BacSp222 (Wladyka et al., 2015).
The plasmid pBIOS236 was detected in BIOS-V236 (15,280 bp) and BIOS-V262 (15,281 bp). This plasmid shares 95.99–96.01% identity with a nearly 15,000-bp region that is duplicated in the 29,587-bp plasmid UVET16-496.1 and harbors tra genes associated with plasmid conjugation.
The 3,043-bp plasmid pCUVET18-79.2, detected in BIOS-V141 and BIOS-V262, is highly similar to the S. pseudintermedius plasmid pCUVET16-803.2 present in BIOS-V227 (Table 1) and to other plasmids deposited in GenBank. These plasmids carry coding sequences (CDS) for hypothetical proteins, a replication protein and CopG, which is involved in plasmid copy number control.
The 3,660-bp plasmid pBIOS212 shows 99.86% identity to S. pseudintermedius strain Dog009_2 plasmid unnamed1, carrying genes for a replication protein and hypothetical proteins.
Plasmid pBIOS259 is described in this study for the first time. This 2,469-bp plasmid shares 83.44% nucleotide sequence identity with plasmid pCUVET18-1255.1 previously isolated in a S. pseudintermedius strain and contains two CDSs for a replication protein and a hypothetical protein.
3.2.2.2 Staphylococcal cassette chromosome mec
The mecA gene is carried in SCCmec elements, classified according to the type of the ccr gene complex and the class of the mec gene complex (International Working Group on The Classification of Staphylococcal Cassette Chromosome Elements [IWG-SCC], 2009). The 17 S. pseudintermedius studied included 12 MRSP strains that carried SCCmec type III (n = 5), SCCmec type IVg (n = 3), SCCmecNA45 (n = 1), ΨSCCmec57395 (n = 1), SCCmec7017–61515 (n = 1), and SCCmec type V(T)SL/154 (n = 1) (Figure 1 and Table 3). SCCmec type III was carried by ST71 strains, while SCCmec type IVg was found among strains of lineages ST265 and ST118 (both from CC227) and ST258 (a double locus variant of the previous ones, assigned to CC258).
3.2.2.3 A novel staphylococcal cassette chromosome harboring fusC in S. pseudintermedius
BIOS-V212 expressed resistance to fusidic acid due to the carriage of the fusC gene, which was located in a novel SCC element (Table 4). This 15,746-bp cassette was delimited using the putative ISS described for the assignment of SCCmec and carried ccrA4 and ccrB4 genes. This new element shared overall 99.72% identity (74% query coverage) with the previous SCCmec-SCCfus element identified in a methicillin-resistant S. aureus (MRSA) (Senok et al., 2019; Figure 2) yet it lacks the orfX-IS1272 region containing the mecA gene and the region downstream of the fusC gene.
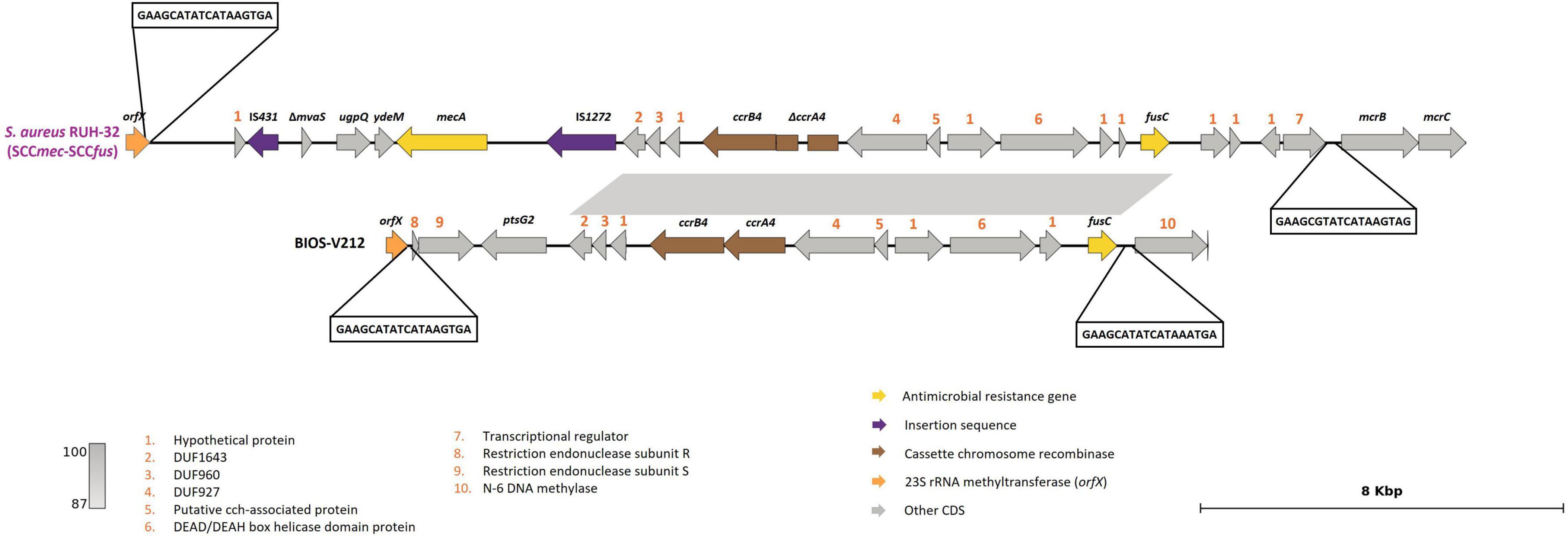
Figure 2. Presentation of the new SCCfus described in S. pseudintermedius carrying the fusC gene. Comparison with the SCCmec-SCCfus described in S. aureus isolate RUH-32 (MK991791). Homology is indicated through a color scale of gray: dark gray (100% homology) to light gray (87% homology). Fusidic acid resistance (fusC) and methicillin-resistance (mecA) genes are represented in yellow; cassette chromosome recombinase genes represented in brown (ccrA4, ccrB4); orfX in orange; insertion sequences in purple. Genes colored in gray represent other genes. The figure was generated using Genofig v1.1.
3.2.2.4 Transposons
Most of the AMR genes found in the 17 strains sequenced were located on transposons integrated into the chromosomal DNA (Figure 1 and Tables 3, 5).
All strains carried the blaZ gene, located either on a Tn552 (six strains), a Tn552-like element (nine strains) or a Tn553 element (four strains, two of which also carried an additional copy of blaZ in Tn552 or Tn552-like elements) (Table 3). The Tn552 elements correspond to the one identified in S. aureus (accession no: X52734), which is delimited by two inverted repeats (Tn552), or a related element where a reverse transcriptase gene is inserted into one of the inverted repeats (Tn552-like).
The tet(M) gene, associated with tetracycline resistance, was found in eight strains, located either on a Tn916 element (n = 6), Tn5801 (n = 1) or Tn5801-like Genomic Island 6287 (GI6287) (n = 1) (Table 3). The Tn5801-like GI6287 found in our study, with 99.96% identity to GI6287, described previously in S. pseudintermedius, has an additional ISLmo18 encoding an IS256 family transposase (Figure 3).
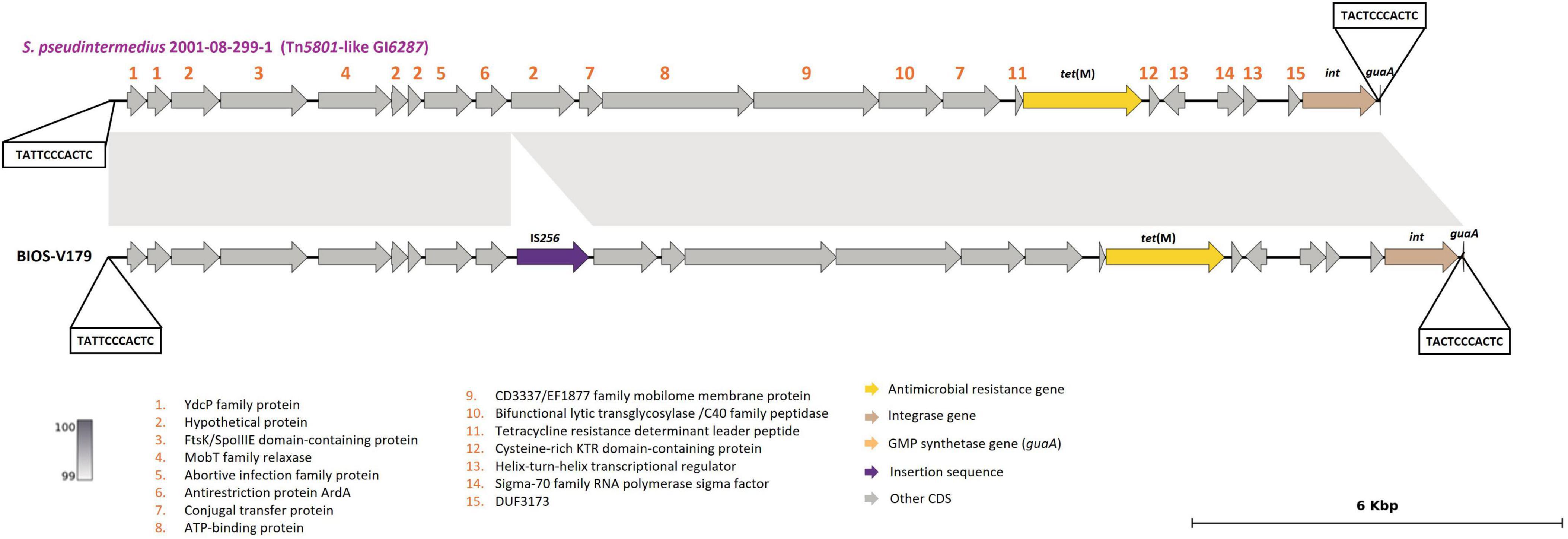
Figure 3. Presentation of Tn5801-like GI6287 (BIOS-V179) carrying the tet(M) gene and comparison with strain S. pseudintermedius 2001-08-299-1 contig_3 (accession number: NZ_JTKO01000003.1, [contig 3, 177,382:198,007]). Minimum homology of 99% was detected between sequences (light gray color). Tetracycline resistance gene [tet(M)] is represented in yellow; integrase gene (int) is represented in brown; GMP synthetase gene (guaA) in orange; insertion sequence in purple. Genes colored in gray represent other genes. The figure was generated using Genofig v1.1.
The genes conferring resistance to aminoglycosides or streptothricin (aphA3, aadE, sat4), macrolide/lincosamide/streptogramin B [erm(B)] and/or trimethoprim (dfrG) were found in nine strains. These genes were carried by four variants of the Tn5405-like element previously identified in S. pseudintermedius ED99, three of them differing in the number and functionality of IS1182 as well as in the presence/absence of dfrG that is linked to an additional insertion sequence in the upstream region of erm(B) (Figure 4A and Table 3). For two strains (BIOS-V237 and BIOS-V299), an integrase downstream of the Tn5405-like element, might have been involved in the insertion of different phages (Figure 4A). The fourth variant carried by BIOS-V16 had a different structure, with a single IS1182, a truncated sat4 gene and four additional genes that confer resistance to aminoglycosides (aadE), aminocyclitols (spw), pleuromutilins/lincosamides/streptogramin A [lsa(E)], and lincosamides [lnu(B)] (Figure 4B).
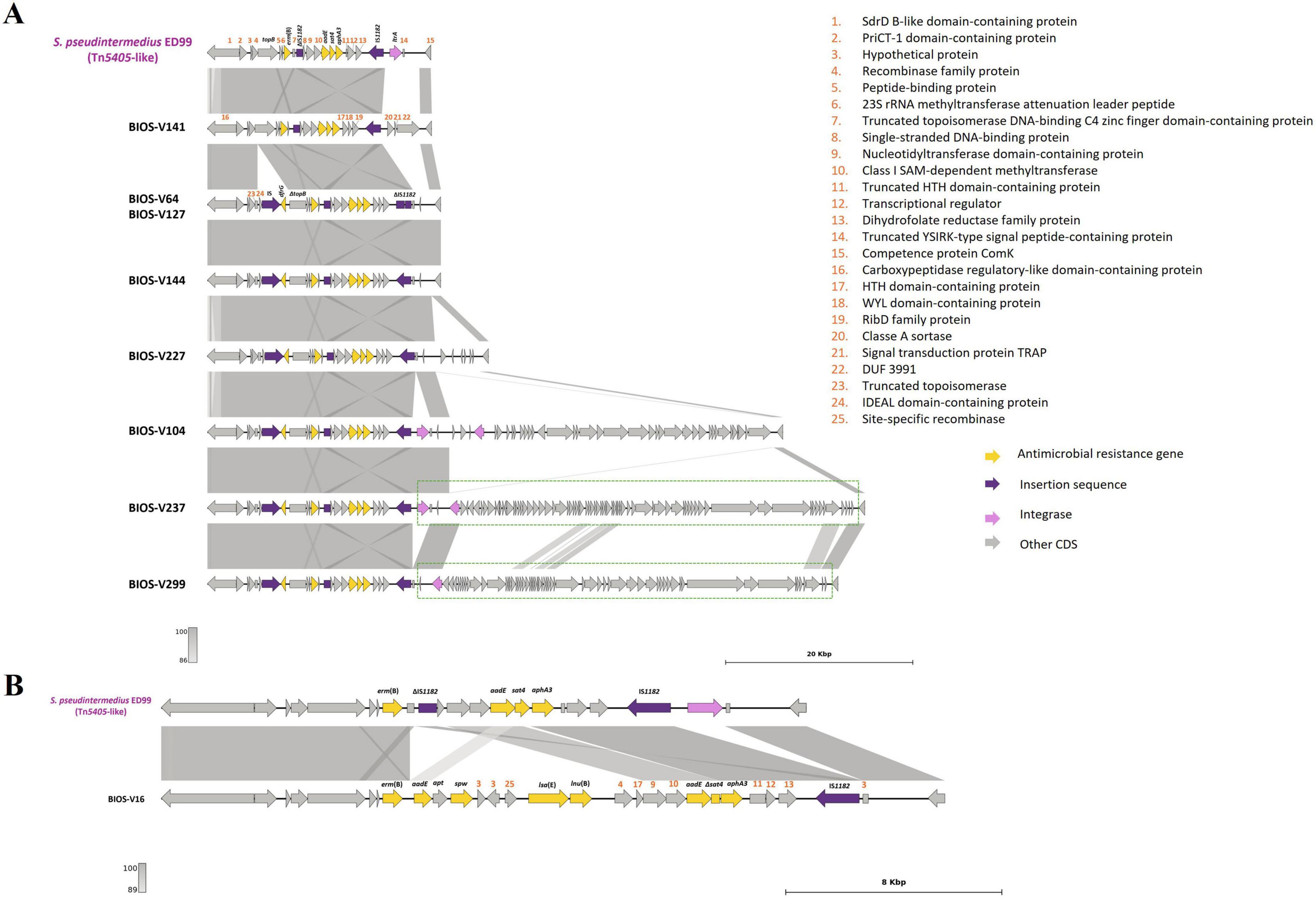
Figure 4. Presentation of Tn5405-like and the region until comK gene and comparison with strain S. pseudintermedius ED99 (NC_017568, [1,833,993:1,857,865]). (A) Homology of Tn5405-like from ED99 with BIOS-V141, a strain without dfrG and seven strains carrying the dfrG gene. Phages detected after the transposon are indicated in a dotted green box. (B) Representation of BIOS-V16 with the additional four antimicrobial resistance genes [aadE, spw, lsa(E) and lnu(B)]. Homology is indicated through a color scale of gray: dark gray (100% homology) to light gray (86% (A) or 89% (B) homology). Antimicrobial resistance genes are represented in yellow; integrase genes are represented in pink; insertion sequences in purple. Genes colored in gray represent other genes. The figure was generated using Genofig v1.1. The inner lines depict additional regions with homology automatically generated by Genofig.
The aacA-aphD gene, conferring resistance to gentamicin, kanamycin and tobramycin, was located in the chromosomal DNA of ten strains, all with phage-related CDS in its vicinity. For five strains (Table 3), this AMR gene is located on a truncated Tn4001, in which the aacA-aphD and orf123 were flanked only by one IS256 and the surrounding phage-related CDS shared 99.97% identity (50% query cover) with the S. epidermidis phage PhiSepi-HH3 (accession no: MT880872.1). In the remaining five strains (BIOS-V16, BIOS-V64, BIOS-V227, BIOS-V262, BIOS-V299) the aacA-aphD gene and the orf123, were also located in the chromosomal DNA but not flanked by IS256 (Figure 5).
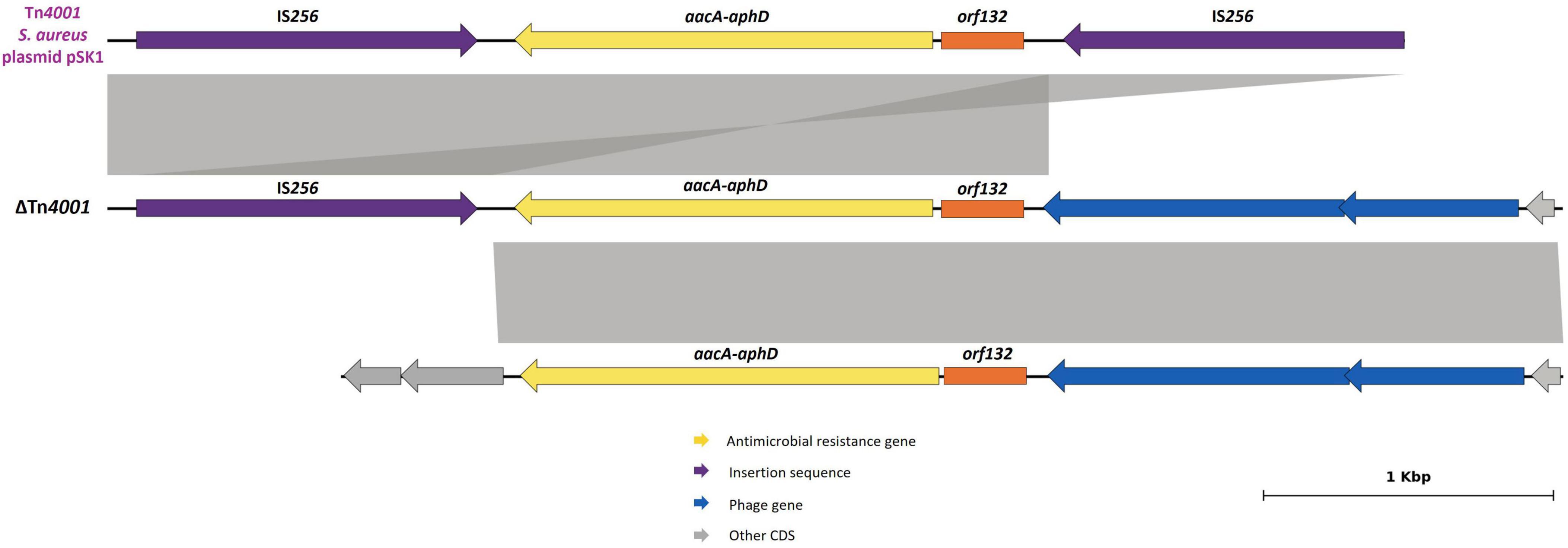
Figure 5. Schematic representations of the truncated Tn4001 and the region in the vicinity of aacA-aphD gene. Comparison with Tn4001 present in S. aureus plasmid pSK1 (accession number: GU565967, [23,577:28,041]). Homology of 100% was detected between sequences. Aminoglycoside resistance gene (aacA-aphD) is represented in yellow; orf132 in orange; insertion sequences in purple; phage-gene related in blue. Genes colored in gray represent other genes. The inner lines depict additional regions with homology automatically generated by Genofig.
3.2.2.5 Prophages
Regarding prophage carriage, PHIGARO identified 15 genomes with an average of 136 prophage-like genes per genome (varying from 19 to 345) and two genomes, from ST45 and ST2059 strains, without any prophage-like gene. Forty-six intact, questionable or incomplete prophages were identified, although none harbored AMR genes (Table 3 and Supplementary Table 2). Most prophages were considered intact or incomplete in the different strains. The five ST71 strains analyzed carried 14 out of 27 intact prophages. Four of these five ST71 strains carried S. pseudintermedius phage SpST71A (Brooks et al., 2020; Supplementary Table 2). This phage encodes a putative protein of the class B metallo β-lactamase (MBL) superfamily, although no studies have established its β-lactam hydrolytic activity. The gene encoding this putative MBL was also detected in the chromosomal DNA next to comGA of the remaining 12 S. pseudintermedius strains sequenced, irrespective of beta-lactam resistance phenotype. All ST71 strains harbored one or more phages or phage remnants carrying a gene annotated as virE, encoding a putative virulence-associated protein previously detected in S. aureus SaPI1 (Lindsay et al., 1998) and in ST71 S. pseudintermedius strains (Papić et al., 2021). Several phages found in other lineages also carried the putative virulence gene virE. Strains from ST45, ST551, ST1183, ST2059 and ST2061 did not carry intact prophages.
3.2.2.6 Other mobile genetic elements
BIOS-V227 and BIOS-V259 carried the tet(K) gene on a plasmid integrated into the chromosomal DNA located between two group II intron reverse transcriptase genes, named pSP-G3C4-like, since it shares identity with pSP-G3C4 described in the S. pseudintermedius strain G3C4. This integrated plasmid is closely related to pSP-G3C4 found as free plasmid in the other tet(K)-positive strains (BIOS-V104, BIOS-V144 and BIOS-V299). Out of the 12 tetracycline-resistant strains sequenced, BIOS-V227 was the only one with resistance to tetracycline mediated by both tet(M) and tet(K) (Figure 1).
Variants of the chromosomally integrated 22,000-bp pRE25-like element harboring genes conferring resistance to aminoglycosides and streptothricin (aphA3, aadE, sat4), macrolides/lincosamides/streptogramin B [erm(B)], and chloramphenicol (catA7) were detected in five strains (Supplementary Figure 1). Four of these variants carried the complete element with four copies of IS1216 and one IS1252, yet lacking IS256. One of them also carried an ISL3 family transposase next to IS1252. The fifth variant, present in BIOS-V240, lost an internal 8,103-bp segment, which contained the erm(B) and catA7 genes as well as one IS1216.
3.2.3 R-M systems and CRISPR
R-M genes were identified in 15 out of the 17 S. pseudintermedius genomes analyzed and were mostly related to R-M Type I (Table 3). These strains contained full intact R-M Type I systems with restriction (HsdR), modification (HsdM) and DNA sequence-recognition (HsdS) subunits. R-M Type II systems with Res and Mod subunits were detected in two MRSP strains, one of which had an element integrated in ΨSCCmec57395 (BIOS-V292). Strain BIOS-V227 harbored a SCCmecV(T)SL/154, recently described as carrying a R-M Type III downstream of orfX (Duim et al., 2018), which according to REBASE, can be considered an R-M Type IIG.
CRISPR-Cas systems were detected in five of the 17 strains, namely one MRSP (BIOS-V227) and four MSSP (BIOS-V179, BIOS-V212, BIOS-V218, BIOS-V236). The ST551 BIOS-V227 harbored three CRISPR systems; CAS-Type IIC and CAS-Type IIIA, each with the cas9 and cas10 signature genes, respectively. The CAS-Type IIIA system was present in two copies, one of them integrated into SSCmec V(T)SL/154 (Table 3).
4 Discussion
Our previous work found a high frequency of resistance to first- and second-line therapeutics recommended for SSTIs caused by S. pseudintermedius (Morais et al., 2023) and other staphylococci in companion animals (Costa et al., 2021; Costa et al., 2022; Leal et al., 2023). For S. aureus, the literature demonstrates that most resistance genes are found on plasmids or transposons integrated into plasmids, facilitating the transfer of these genes between strains (Malachowa and DeLeo, 2010). The previous data regarding plasmid carriage for S. pseudintermedius, initially described as S. intermedius (Devriese et al., 2005), indicated a high frequency of plasmid carriage, mostly smaller than 5 kb and associated with resistance to tetracycline and chloramphenicol (Greene and Schwarz, 1992; Schwarz et al., 1995; Schwarz et al., 1998; Werckenthin et al., 2001). The more recent studies, using WGS techniques, confirm the presence of small plasmids in the S. pseudintermedius genome, albeit in a variable frequency (Ferrer et al., 2021; Fàbregas et al., 2023; Zehr et al., 2025), and indicate the carriage of tet(K) (Viñes et al., 2020; Soimala et al., 2020) but also qac genes (Hritcu et al., 2020; Fàbregas et al., 2023) in these plasmids.
Our data suggest that plasmids may not be the most relevant MGE involved in AMR in S. pseudintermedius. Despite a high frequency of plasmid carriage among the initial S. pseudintermedius collection (31/56, 55.4%), the WGS results indicate that most of the AMR determinants found (blaZ, mecA, aacA-aphD, aphA3, erm(B), dfrG, tet(M), fusC) were not related to plasmids (Table 5). Indeed, only tet(K) was located on plasmid pSP-G3C4, carried by ST71 strains, in accordance with literature (Worthing et al., 2018a; Menandro et al., 2019). pSP-G3C4 and structurally very similar tet(K)-carrying plasmids are widespread among various staphylococcal species (Schwarz et al., 1998), including S. aureus (Diep et al., 2006), S. hominis (Belhout et al., 2023) and S. epidermidis (Zhang et al., 2003). Interestingly, a plasmid with high homology to pSP-G3C4 was also found integrated into the chromosomal DNA of ST551 and ST2109 strains. This pSP-G3C4-like plasmid, located between two group II intron reverse transcriptase genes, is a mobilizable plasmid that carries a mobV gene and also the gene tet(M) as part of a transposon and was previously found in the chromosomal DNA of ST551 strains (Viñes et al., 2022; Viñes et al., 2024). Detection of the tet(K) gene is relevant, as it also confers resistance to doxycycline (Weese et al., 2013), recommended as a second-line therapy for canine pyoderma in dogs (Loeffler et al., 2025). We also detected the efflux gene qacG, linked to reduced susceptibility to quaternary ammonium compounds (Costa et al., 2013), located on plasmid pSP5912, previously described in S. pseudintermedius (Worthing et al., 2018b). Reduced susceptibility to biocides has been reported in staphylococci that carry qac genes (Furi et al., 2013; Costa et al., 2016; Hardy et al., 2018; Worthing et al., 2018b), although the corresponding minimal inhibitory or bactericidal concentrations do not reach the in use recommended concentrations (Couto et al., 2013; Worthing et al., 2018b).
Regarding the MGEs in the chromosomal DNA, SCCmec was detected in the 12 MRSP strains. The most frequent type of SCCmec was type III, which is usually found in S. pseudintermedius ST71 strains (Perreten et al., 2013; Krapf et al., 2019; Wegener et al., 2020), as we observed. SCCmec type IVg was found in the ST258 strain and the ΨSCCmec57395, carrying heavy metal resistance genes, was detected in a ST45 strain, in accordance with other studies (Perreten et al., 2013; Worthing et al., 2018c; Wegener et al., 2020; Bruce et al., 2022). BIOS-V240, from ST2061, previously detected for the first time in our collection, carried SCCmec7017–61515, a cassette recently described in an ST1200 S. pseudintermedius isolated from a dog wound, being the first cassette identified with mec gene complex A and a ccrC1 gene in a non-composite element (MacFadyen and Paterson, 2024). Strain BIOS-V227 (ST551) carries the SCCmecV(T)SL/154, described by Duim et al. for an ST121 strain (a triple-locus variant of ST551) and harbors a type III R-M systems, a CRISPR/Cas complex and the cadmium resistance gene cadA (Duim et al., 2018).
In addition to SCCmec, several transposons, integrated into chromosomal DNA, carried AMR determinants. The blaZ gene, conferring penicillin resistance, was located either in Tn552, Tn552-like and/or Tn553 elements. Tn552 is frequently detected on S. aureus plasmids (Schwarz et al., 2014; Partridge et al., 2018) or integrated into S. pseudintermedius chromosomal DNA (McCarthy et al., 2015; Phumthanakorn et al., 2021). Tn553, a member of the Tn554 family, was recently described by Krüger et al. in a porcine MRSA strain and detected in silico in MSSP strains (Krüger et al., 2021). In our study, this element was identified in one MRSP and three MSSP strains.
Resistance to tetracycline mediated by tet(M) was linked to transposons of the conjugative Tn916-like family, namely Tn916, Tn5801 or Tn5801-like GI6287, all previously reported in S. pseudintermedius (McCarthy et al., 2015; de Vries et al., 2016). Genomic islands are relevant for the evolution of bacterial species since they are conserved within strain lineages. The mechanism of mobilization of these elements is not established for S. pseudintermedius, but it is expected that horizontal transfer of GI6287 occurs at a low frequency or under specific conditions, since it lacks the xis-like genes necessary for its excision. In S. aureus, mobilization of GI6287 is assisted by temperate helper phages that have specific tail proteins to target the recipient cells that are maintained in related clonal lineages (Moon et al., 2016).
The carriage of Tn5405-like variants contributes to AMR, conferring a MDR profile in S. pseudintermedius strains (Phumthanakorn et al., 2021). A Tn5405-like element was detected in almost all the MRSP strains (9/12), a result in accordance with other reports that verified the predisposition of staphylococci to acquire this transposon following the acquisition of SCCmec (McCarthy et al., 2015; Fàbregas et al., 2023). Of interest, the Tn5405-like element carried by BIOS-V16 harbored four additional antimicrobial resistance genes (Table 3) in an arrangement previously identified in S. pseudintermedius (Viñes et al., 2022). The region encompassing these four genes shares homology with plasmids isolated from a human MRSA (Lozano et al., 2012; Figure 6A) and a porcine Enterococcus faecium (Li et al., 2014; Figure 6B).
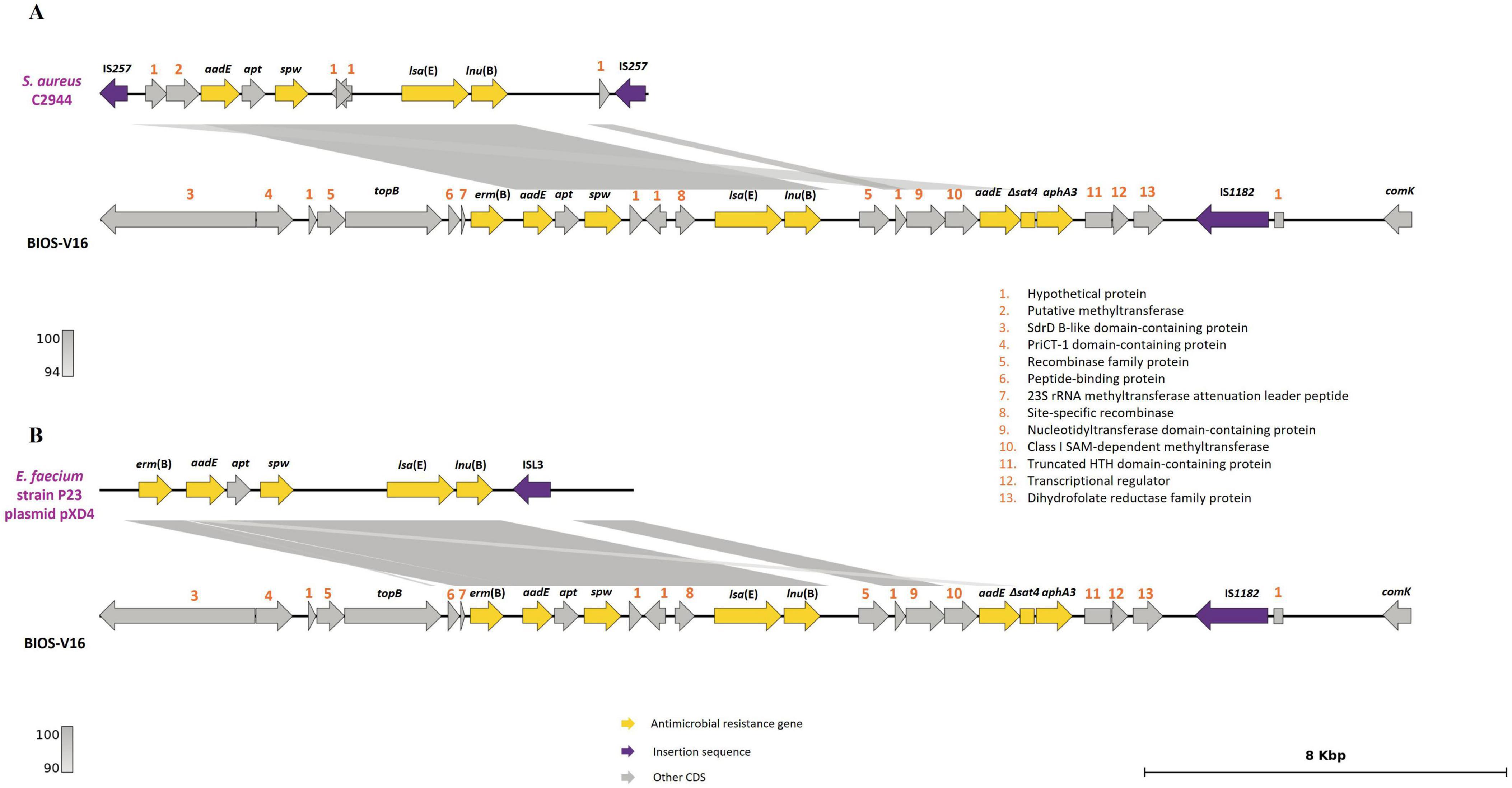
Figure 6. Analysis of the carriage of erm(B), aadE, spw, lsa(E) and lnu(B) genes by BIOS-V16 strain. (A) Comparison with S. aureus strain C2944 (JQ861959.1) and (B) Enterococcus faecium strain P23 plasmid pXD4 (KF421157). Homology is indicated through a color scale of gray: dark gray (100% homology) to light gray (94% (A) or 90% (B) homology). Antimicrobial resistance genes are represented in yellow; insertion sequence in purple. Genes colored in gray represent other genes. The figure was generated using Genofig v1.1. The inner lines depict additional regions with homology automatically generated by Genofig.
The aacA-aphD gene was detected in a ΔTn4001 with a single IS256 integrated in an incomplete phage sequence. Five strains carried a putative Tn4001-like variant, not integrated in a phage and lacking IS256, suggesting the additional presence of a Tn4001-like variant in S. pseudintermedius. Several variants of Tn4001-like have been reported for Staphylococcus spp. (Byrne et al., 1990; Lange et al., 2003; Schwarz et al., 2011; Chanchaithong et al., 2024), mostly caused by the partial deletion of IS256 or the integration of IS257. Zhang et al. identified a Tn4001 variant without IS256 on both termini in Enterococcus faecalis (Zhang et al., 2018).
Other MGEs identified in our collection included the elements pRE25-like, a new SCCfus and prophages.
The pRE25-like element, first described in Enterococcus spp. (Werner et al., 2003), is integrated into the S. pseudintermedius chromosomal DNA and carries cat-erm(B)-aadE-sat4-aphA3 genes (Kang and Hwang, 2020). We identified variants of this mobile element, one of which carrying only aadE-sat4-aphA3, in ST241 and ST45 strains, in line with other studies (Wegener et al., 2022), but also in the newly described lineages ST2061 and ST2109 (Morais et al., 2023).
Resistance to fusidic acid was detected in five strains, one of them carrying fusC in a SCC element described for the first time in S. pseudintermedius in this study. This element carries ccrA4 and ccrB4 genes and shares similarity with the SCCmec-SCCfus described earlier in a MRSA strain (Senok et al., 2019). The other four strains resistant to fusidic acid had point mutations resulting in amino acid exchanges in FusA, two of them (FusA:H457Q and FusA:G451V) described previously in S. aureus strains with low-level resistance (Besier et al., 2003; O’Neill et al., 2004; Castanheira et al., 2010; Chen et al., 2010) and detected now for the first time in S. pseudintermedius. The FusA:I461T alteration was reported previously in S. pseudintermedius together with two other mutations and it was related to growth fitness compensation (Frosini et al., 2019; Lim et al., 2020). We also took into consideration the resistance phenotype to rifampicin. The amino acid exchange RpoB:H481N, here reported also for the first time for S. pseudintermedius, was earlier described in a MRSA strain of porcine origin, associated with low-level resistance to rifampicin (Li et al., 2016; Schwarz et al., 2018).
The five ST71 strains carried SCCmec, Tn552 and Tn552-like as well as Tn5405-like variants. Strains belonging to ST258 and ST551, which are considered emerging lineages in the North of Europe, replacing ST71 (Damborg et al., 2016; Kizerwetter-Świda et al., 2017), harbored the same MGEs integrated into the chromosomal DNA, and additionally Tn553, Tn916 and pSP-G3C4-like. Other studies have found different combinations of AMR gene(s)/MGEs/lineages (McCarthy et al., 2015; Phumthanakorn et al., 2021; Fàbregas et al., 2023). ST241, the most frequent ST among the MSSP strains previously studied (Morais et al., 2023) and recently associated to human S. pseudintermedius infection (Wegener et al., 2021), showed a MDR profile, conferred by the pRE25-like element. The two MSSP-ST241, MSSP-ST2109, and all MRSP strains studied showed resistance to at least one of the first- and second-line systemic treatment options recommended for SSTIs (clindamycin and cephalosporins). This is highly relevant for the therapy of SSTIs in companion animals since it suggests a possible inefficacy of these antimicrobials as a treatment option, not only for MRSP but also for MSSP strains.
Prophages can carry genes that contribute to AMR, virulence, fitness and adaptation to the host (McCarthy et al., 2015). Intact prophages or prophage-like genes were detected in 88% of the sequenced S. pseudintermedius strains and none of them carried AMR determinants, in agreement with previous studies (Wipf et al., 2019; Moodley et al., 2019; Brooks et al., 2020; Phumthanakorn et al., 2021). All ST71 strains except one (BIOS-V299), carried phage SpST71A, described previously in this lineage (Brooks et al., 2020), that disrupts the comG operon, a genetic barrier to horizontal gene transfer (HGT). Few data about prophages in S. pseudintermedius are available in public databases. BLASTn analysis of our sequences revealed high percentages of identity with several phage sequences but with low query coverage using the NCBI Viruses database (Supplementary Table 2). The BLASTn also allowed the detection of similar prophages in different S. pseudintermedius genomes deposited in GenBank, however, these are not identified or classified as phages, hampering prophage identification. Our data suggest that prophages are not related to AMR gene carriage in S. pseudintermedius, yet it is known that these MGEs increase strain plasticity, contributing to the genetic diversity of the bacterial population. In addition, they allow a better adaptation of the bacteria to new environments by increasing their pathogenic potential and the transfer of MGEs harboring factors that confer unique virulence characteristics to the bacteria (Naorem et al., 2021; Gummalla et al., 2023).
Restriction-modification and CRISPR/Cas systems are significant genetic barriers that regulate HGT among bacteria, including staphylococci. Four types of R-M systems were reported in Staphylococcus species (Sadykov, 2016). In S. pseudintermedius, Types I and II are the most frequently described (McCarthy et al., 2015; Brooks et al., 2020; Phumthanakorn et al., 2021). We verified that almost all the strains carried at least one type of R-M system, independently of the number of AMR genes and MGEs carried. R-M type I was the most frequently detected, particularly associated with ST71. A previous study suggested a relation between R-M type and S. pseudintermedius clonal lineage (Brooks et al., 2020). That study identified R-M type I in ST71 and ST258, and R-M type II in ST45, in accordance with our findings. Although more studies are needed to confirm the linkage between ST and R-M type observed in S. pseudintermedius, data from other bacteria suggest that R-M systems facilitate HGT within the same clonal lineage or between lineages with cognate R-M systems (Oliveira et al., 2016).
The CRISPR/Cas regulatory capacity to control HGT was already described for Staphylococcus spp. (Rossi et al., 2017; Mortensen et al., 2021). These systems are not frequently detected in staphylococci of canine origin (Rossi et al., 2019), and only types II and IIIA are known in S. pseudintermedius (Brooks et al., 2020; Phumthanakorn et al., 2021). Following previous studies (Brooks et al., 2020; Wegener et al., 2021), these systems were not detected among the ST71 and ST45 strains of our collection. On the other hand, the single ST551 strain carried several CRISPR/Cas systems, also in line with recent reports (Grist et al., 2025), despite its MDR phenotype and multiple MGEs. Since CRISPR/Cas may function as molecular clock, future studies characterizing the spacers from the CRISPR/Cas system could provide information about the origin and the time of these genetic events.
5 Conclusion
This study highlights a low diversification of the S. pseudintermedius mobilome between clonal lineages. The results obtained in this study indicated that S. pseudintermedius has a high proportion of plasmid carriage (> 50%), although with a low diversity and not frequently related to AMR gene carriage. AMR determinants were found mostly within other MGEs integrated into chromosomal DNA, namely Tn552, Tn552-like, Tn553, Tn5405-like, Tn916, Tn5801, Tn5801-like GI6287 and a pRE25-like element. One MSSP strain harbored SCCfus, a new element carrying the fusC gene, detected, to the best of our knowledge, for the first time in S. pseudintermedius. Most of the strains analyzed also carried prophages in their genomes however without AMR genes, suggesting a role in other biological processes such as adaptation to the host and bacterial fitness. The transfer of these MGEs in S. pseudintermedius can be controlled by R-M systems, which were present in almost all strains, and CRISPR/Cas systems. The results obtained in this study provide important insights that may lead to a better understanding of MDR in S. pseudintermedius towards improved SSTIs treatment in companion animals.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
CM: Data curation, Formal analysis, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. SSC: Conceptualization, Formal analysis, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. DH: Data curation, Formal analysis, Investigation, Validation, Visualization, Writing – original draft, Writing – review & editing. AS: Investigation, Writing – original draft, Writing – review & editing. HK-H: Validation, Writing – original draft, Writing – review & editing. CP: Writing – review & editing. AF: Conceptualization, Validation, Writing – original draft, Writing – review & editing. SS: Conceptualization, Funding acquisition, Project administration, Validation, Writing – original draft, Writing – review & editing. IC: Conceptualization, Funding acquisition, Project administration, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by Project BIOSAFE funded by FEDER and Fundação para a Ciência e a Tecnologia (FCT, Portugal) [LISBOA-01-0145-FEDER-030713, PTDC/CAL-EST/30713/2017]. Further support by FCT to GHTM (UID/04413/2020), LA-REAL (LA/P/0117/2020), grant UI/BD/151061/2021 to CM ( doi: 10.54499/UI/BD/151061/2021) and Bilateral PT-Germany (FCT/DAAD) mobility project 2022.15300.CBM as well as 57664784 FU Berlin. SSC was supported by FCT through CEECINST/00042/2021/CP1773/CT0009, doi: 10.54499/CEECINST/00042/2021/CP1773/CT0009.
Acknowledgments
The authors thank Patrícia Abrantes (IHMT/NOVA) and Sofia Seabra (IHMT/NOVA) for their support in MinION use and data visualization.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1640322/full#supplementary-material
Footnotes
1. ^https://www.bioinformatics.babraham.ac.uk/projects/fastqc
2. ^https://www.genomicepidemiology.org
3. ^https://cge.food.dtu.dk/services/SCCmecFinder
4. ^https://tools.neb.com/genomes/
5. ^https://crisprcas.i2bc.paris-saclay.fr/CrisprCasFinder/Index
References
Adiguzel, M. C., Schaefer, K., Rodriguez, T., Ortiz, J., and Sahin, O. (2022). Prevalence, mechanism, genetic diversity, and cross-resistance patterns of methicillin-resistant Staphylococcus isolated from companion animal clinical samples submitted to a veterinary diagnostic laboratory in the Midwestern United States. Antibiotics 11:609. doi: 10.3390/antibiotics11050609
Afshar, M. F., Zakaria, Z., Cheng, C. H., and Ahmad, N. I. (2023). Prevalence and multidrug-resistant profile of methicillin-resistant Staphylococcus aureus and methicillin-resistant Staphylococcus pseudintermedius in dogs, cats, and pet owners in Malaysia. Vet. World 16, 536–545. doi: 10.14202/vetworld.2023.536-545
Alcock, B. P., Huynh, W., Chalil, R., Smith, K. W., Raphenya, A. R., Wlodarski, M. A., et al. (2023). CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the comprehensive antibiotic resistance database. Nucleic Acids Res. 51, D690–D699. doi: 10.1093/nar/gkac920
Arndt, D., Grant, J. R., Marcu, A., Sajed, T., Pon, A., Liang, Y., et al. (2016). PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 44, W16–W21. doi: 10.1093/nar/gkw387
Belhout, C., Boyen, F., Vereecke, N., Theuns, S., Taibi, N., Stegger, M., et al. (2023). Prevalence and molecular characterization of Methicillin-resistant Staphylococci (MRS) and Mammaliicocci (MRM) in dromedary camels from Algeria: First detection of SCCmec-mecC hybrid in methicillin-resistant Mammaliicoccus lentus. Antibiotics 12:674. doi: 10.3390/antibiotics12040674
Besier, S., Ludwig, A., Brade, V., and Wichelhaus, T. A. (2003). Molecular analysis of fusidic acid resistance in Staphylococcus aureus. Mol. Microbiol. 47, 463–469. doi: 10.1046/j.1365-2958.2003.03307.x
Bortolaia, V., Kaas, R. S., Ruppe, E., Roberts, M. C., Schwarz, S., Cattoir, V., et al. (2020). ResFinder 4.0 for predictions of phenotypes from genotypes. J. Antimicrob. Chemother. 75, 3491–3500. doi: 10.1093/jac/dkaa345
Brooks, M. R., Padilla-Vélez, L., Khan, T. A., Qureshi, A. A., Pieper, J. B., Maddox, C. W., et al. (2020). Prophage-mediated disruption of genetic competence in Staphylococcus pseudintermedius. mSystems 5:e00684–19. doi: 10.1128/mSystems.00684-19
Bruce, S. A., Smith, J. T., Mydosh, J. L., Ball, J., Needle, D. B., Gibson, R., et al. (2022). Accessory genome dynamics of local and global Staphylococcus pseudintermedius populations. Front. Microbiol. 13:798175. doi: 10.3389/fmicb.2022.798175
Byrne, M. E., Gillespie, M. T., and Skurray, R. A. (1990). Molecular analysis of a gentamicin resistance transposonlike element on plasmids isolated from North American Staphylococcus aureus strains. Antimicrob. Agents Chemother. 34, 2106–2113. doi: 10.1128/AAC.34.11.2106
Calabro, C., Sadhu, R., Xu, Y., Aprea, M., Guarino, C., and Cazer, C. L. (2024). Longitudinal antimicrobial susceptibility trends of canine Staphylococcus pseudintermedius. Prev. Vet. Med. 226:6170. doi: 10.1016/j.prevetmed.2024.106170
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: Architecture and applications. BMC Bioinform. 10:421. doi: 10.1186/1471-2105-10-421
Carattoli, A., Zankari, E., Garcia-Fernandez, A., Voldby, L. M., Lund, O., Villa, L., et al. (2014). In silico detection and typing of plasmids using plasmid finder and plasmid multilocus sequence typing. Antimicrob. Agents Chemother. 58, 3895–3903. doi: 10.1128/AAC.02412-14
Castanheira, M., Watters, A. A., Bell, J. M., Turnidge, J. D., and Jones, R. N. (2010). Fusidic acid resistance rates and prevalence of resistance mechanisms among Staphylococcus spp. isolated in North America and Australia, 2007-2008. Antimicrob. Agents Chemother. 54, 3614–3617. doi: 10.1128/AAC.01390-09
Chanchaithong, P., Chueahiran, S., Pinpimai, K., Sroithongkham, P., Leelapsawas, C., Indra, R., et al. (2024). Chromosomal and plasmid localization of ileS2 in high-level mupirocin-resistant Staphylococcus pseudintermedius and Staphylococcus aureus isolated from canine and feline origins. J. Antimicrob. Chemother. 79, 1856–1864. doi: 10.1093/jac/dkae172
Chen, H., Hung, W., Tseng, S., Tsai, J., Hsueh, P., and Teng, L. (2010). Fusidic acid resistance determinants in Staphylococcus aureus clinical isolates. Antimicrob. Agents Chemother. 54, 4985–4991. doi: 10.1128/AAC.00523-10
Costa, S. S., Oliveira, V., Serrano, M., Pomba, C., and Couto, I. (2021). Phenotypic and molecular traits of Staphylococcus coagulans associated with canine skin infections in Portugal. Antibiotics 10:518. doi: 10.3390/antibiotics10050518
Costa, S. S., Palma, C., Kadlec, K., Feßler, A. T., Viveiros, M., Melo-Cristino, J., et al. (2016). Plasmid-borne antimicrobial resistance of Staphylococcus aureus isolated in a hospital in Lisbon, Portugal. Microb. Drug Resist. 22, 617–626. doi: 10.1089/mdr.2015.0352
Costa, S. S., Ribeiro, R., Serrano, M., Oliveira, K., Ferreira, C., Leal, M., et al. (2022). Staphylococcus aureus causing skin and soft tissue infections in companion animals: Antimicrobial resistance profiles and clonal lineages. Antibiotics 11:599. doi: 10.3390/antibiotics11050599
Costa, S. S., Viveiros, M., Amaral, L., and Couto, I. (2013). Multidrug efflux pumps in Staphylococcus aureus: An update. Open Microbiol. J. 7, 59–71. doi: 10.2174/1874285801307010059
Couto, N., Belas, A., Tilley, P., Couto, I., Gama, L. T., Kadlec, K., et al. (2013). Biocide and antimicrobial susceptibility of methicillin-resistant staphylococcal isolates from horses. Vet. Microbiol. 166, 299–303. doi: 10.1016/j.vetmic.2013.05.011
Couvin, D., Bernheim, A., Toffano-Nioche, C., Touchon, M., Michalik, J., Néron, B., et al. (2018). CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 46, W246–W251. doi: 10.1093/nar/gky425
Damborg, P., Moodley, A., Aalbæk, B., Ventrella, G., Pires, dos Santos, T., et al. (2016). High genotypic diversity among methicillin-resistant Staphylococcus pseudintermedius isolated from canine infections in Denmark. BMC Vet. Res. 12:131. doi: 10.1186/s12917-016-0756-y
de Vries, L. E., Hasman, H., Rabadán, S. J., and Agersø, Y. (2016). Sequence-based characterization of Tn5801-like genomic islands in tetracycline-resistant Staphylococcus pseudintermedius and other Gram-positive bacteria from humans and animals. Front. Microbiol. 7:576. doi: 10.3389/fmicb.2016.00576
Devriese, L. A., Vancanneyt, M., Baele, M., Vaneechoutte, M., De Graef, E., Snauwaert, C., et al. (2005). Staphylococcus pseudintermedius sp. nov., a coagulase-positive species from animals. Int. J. Syst. Evol. Microbiol. 55, 1569–1573. doi: 10.1099/ijs.0.63413-0
Diep, B. A., Gill, S. R., Chang, R. F., Phan, T. H., Chen, J. H., Davidson, M. G., et al. (2006). Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 367, 731–739. doi: 10.1016/SO140-6736(06)68231-7
Duim, B., Verstappen, K. M. H. W., Kalupahana, R. S., Ranathunga, L., Fluit, A. C., and Wagenaar, J. A. (2018). Methicillin-resistant Staphylococcus pseudintermedius among dogs in the description of novel SCCmec variants. Vet. Microbiol. 213, 136–141. doi: 10.1016/j.vetmic.2017.11.022
Fàbregas, N., Pérez, D., Viñes, J., Cuscó, A., Migura-García, L., Ferrer, L., et al. (2023). Diverse populations of Staphylococcus pseudintermedius colonize the skin of healthy dogs. Microbiol. Spectr. 11:e0339322. doi: 10.1128/spectrum.03393-22
Ferrer, L., García-Fonticoba, R., Pérez, D., Viñes, J., Fàbregas, N., Madroñero, S., et al. (2021). Whole genome sequencing and de novo assembly of Staphylococcus pseudintermedius: A pangenome approach to unravelling pathogenesis of canine pyoderma. Vet. Dermatol. 32, 654–663. doi: 10.1111/vde.13040
Feßler, A. T., Scholtzek, A. D., Schug, A. R., Kohn, B., Weingart, C., Schink, A. K., et al. (2022). Antimicrobial and biocide resistance among feline and canine Staphylococcus aureus and Staphylococcus pseudintermedius isolates from diagnostic submissions. Antibiotics 11:127. doi: 10.3390/antibiotics11020127
Frosini, S., Bond, R., Rantala, M., Grönthal, T., Rankin, S. C., O’Shea, K., et al. (2019). Genetic resistance determinants to fusidic acid and chlorhexidine in variably susceptible staphylococci from dogs. BMC Microbiol. 19:81. doi: 10.1186/s12866-019-1449-z
Fu, Z., Liu, Y., Chen, C., Guo, Y., Ma, Y., Yang, Y., et al. (2016). Characterization of fosfomycin resistance gene, fosB, in methicillin-resistant Staphylococcus aureus isolates. PLoS One 11:e0154829. doi: 10.1371/journal.pone.0154829
Fukasawa, Y., Ermini, L., Wang, H., Carty, K., and Cheung, M. (2020). LongQC: A quality control tool for third generation sequencing long read data. G3 10, 1193–1196. doi: 10.1534/g3.119.400864
Furi, L., Ciusa, M. L., Knight, D., Di Lorenzo, V., Tocci, N., Cirasola, D., et al. (2013). Evaluation of reduced susceptibility to quaternary ammonium compounds and bisbiguanides in clinical isolates and laboratory-generated mutants of Staphylococcus aureus. Antimicrob. Agents Chemother. 57, 3488–3497. doi: 10.1128/AAC.00498-13
Greene, R. T., and Schwarz, S. (1992). Small antibiotic resistance plasmids in Staphylococcus intermedius. Zentralblatt Bakteriol. 276, 380–389. doi: 10.1016/S0934-8840(11)80545-1
Grist, L. F., Brown, A., Fitzpatrick, N., Mariano, G., La Ragione, R. M., Van Vliet, A. H. M., et al. (2025). Global phylogenomic analysis of Staphylococcus pseudintermedius reveals genomic and prophage diversity in multidrug-resistant lineages. Microb. Genom. 11:001369. doi: 10.1099/mgen.0.001369
Gummalla, V. S., Zhang, Y., Liao, Y., and Wu, V. C. H. (2023). The role of temperate phages in bacterial pathogenicity. Microorganisms 11:541. doi: 10.3390/microorganisms11030541
Haaber, J., Penadés, J. R., and Ingmer, H. (2017). Transfer of antibiotic resistance in Staphylococcus aureus. Trends Microbiol. 25, 893–905. doi: 10.1016/j.tim.2017.05.011
Harbour, L., Schick, A., Mount, R., and White, A. (2022). Rifampicin treatment of canine multidrug-resistant meticillin-resistant staphylococcal pyoderma: A retrospective study of 51 cases. Vet. Dermatol. 33, 384–391. doi: 10.1111/vde.13105
Hardy, K., Sunnucks, K., Gil, H., Shabir, S., Trampari, E., Hawkey, P., et al. (2018). Increased usage of antiseptics is associated with reduced susceptibility in clinical isolates of Staphylococcus aureus. mBio 9:e00894–18. doi: 10.1128/mBio.00894-18
Hicks, K., Tan, Y., Cao, W., Hathock, T., Boothe, D., Kennis, R., et al. (2021). Genomic and in vitro pharmacodynamic analysis of rifampicin resistance in multidrug-resistant canine Staphylococcus pseudintermedius isolates. Vet. Dermatol. 32, 219–e57. doi: 10.1111/vde.12959
Hillier, A., Lloyd, D. H., Weese, J. S., Blondeau, J. M., Boothe, D., Breitschwerdt, E., et al. (2014). Guidelines for the diagnosis and antimicrobial therapy of canine superficial bacterial folliculitis (antimicrobial guidelines working group of the international society for companion animal infectious diseases). Vet. Dermatol. 25, 163–e43. doi: 10.1111/vde.12118
Hritcu, O. M., Schmidt, V. M., Salem, S. E., Maciuca, I. E., Moraru, R. F., Lipovan, I., et al. (2020). Geographical variations in virulence factors and antimicrobial resistance amongst Staphylococci isolated from dogs from the United Kingdom and Romania. Front. Vet. Sci. 7:414. doi: 10.3389/fvets.2020.00414
Hu, J., Fan, J., Sun, Z., and Liu, S. (2020). NextPolish: A fast and efficient genome polishing tool for long-read assembly. Bioinformatics 36, 2253–2255. doi: 10.1093/bioinformatics/btz891
International Working Group on The Classification of Staphylococcal Cassette Chromosome Elements [IWG-SCC] (2009). Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 56, 4961–4967. doi: 10.1128/AAC.00579-09
Ito, T., Ma, X. X., Takeuchi, F., Okuma, K., Yuzawa, H., and Hiramatsu, K. (2004). Novel type V Staphylococcal cassette chromosome mec driven by a novel cassette chromosome recombinase, ccrC. Antimicrob. Agents Chemother. 48, 2637–2651. doi: 10.1128/AAC.48.7.2637-2651.2004
Jia, B., Raphenya, A. R., Alcock, B., Waglechner, N., Guo, P., Tsang, K. K., et al. (2017). CARD 2017: Expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res. 45, D566–D573. doi: 10.1093/nar/gkw1004
Johansson, M. H. K., Bortolaia, V., Tansirichaiya, S., Aarestrup, F. M., Roberts, A. P., and Petersen, T. N. (2021). Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J. Antimicrob. Chemother. 76, 101–109. doi: 10.1093/jac/dkaa390
Kadlec, K., and Schwarz, S. (2012). Antimicrobial resistance of Staphylococcus pseudintermedius. Vet. Dermatol. 23, 276–82, e55. doi: 10.1111/j.1365-3164.2012.01056.x
Kadlec, K., van Duijkeren, E., Wagenaar, J. A., and Schwarz, S. (2011). Molecular basis of rifampicin resistance in methicillin-resistant Staphylococcus pseudintermedius isolates from dogs. J. Antimicrob. Chemother. 66, 1236–1242. doi: 10.1093/jac/dkr118
Kang, J., and Hwang, C. (2020). First detection of multiresistance pRE25-like elements from Enterococcus spp. in Staphylococcus pseudintermedius isolated from canine pyoderma. J. Glob. Antimicrob. Resist. 20, 304–308. doi: 10.1016/j.jgar.2019.08.022
Kizerwetter-Świda, M., Chrobak-Chmiel, D., Rzewuska, M., and Binek, M. (2017). Changes in the population structure of canine methicillin-resistant Staphylococcus pseudintermedius in Poland. Vet. Microbiol. 208, 106–109. doi: 10.1016/j.vetmic.2017.07.025
Kolmogorov, M., Yuan, J., Lin, Y., and Pevzner, P. (2019). Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 37, 540–546. doi: 10.1038/s41587-019-0072-8
Krapf, M., Müller, E., Reissig, A., Slickers, P., Braun, S. D., Müller, E., et al. (2019). Molecular characterisation of methicillin-resistant Staphylococcus pseudintermedius from dogs and the description of their SCCmec elements. Vet. Microbiol. 233, 196–203. doi: 10.1016/j.vetmic.2019.04.002
Krüger, H., Ji, X., Wang, Y., Feßler, A. T., Wang, Y., Wu, C., et al. (2021). Identification of Tn553, a novel Tn554-related transposon that carries a complete blaZ-blaR1-blaI β-lactamase operon in Staphylococcus aureus. J. Antimicrob. Chemother. 76, 2733–2735. doi: 10.1093/jac/dkab210
Lange, C. C., Werckenthin, C., and Schwarz, S. (2003). Molecular analysis of the plasmid-borne aacA/aphD resistance gene region of coagulase-negative staphylococci from chickens. J. Antimicrob. Chemother. 51, 1397–1401. doi: 10.1093/jac/dkg257
Leal, M., Morais, C., Ramos, B., Pomba, C., Abrantes, P., Costa, S. S., et al. (2023). Exploring efflux as a mechanism of reduced susceptibility towards biocides and fluoroquinolones in Staphylococcus pseudintermedius. Animals 13:1270. doi: 10.3390/ani13071270
Li, J., Feßler, A. T., Jiang, N., Fan, R., Wang, Y., Wu, C., et al. (2016). Molecular basis of rifampicin resistance in multiresistant porcine livestock-associated MRSA. J. Antimicrob. Chemother. 71, 3313–3315. doi: 10.1093/jac/dkw294
Li, X., Dong, W., Wang, X., Hu, G., Wang, Y., Cai, B., et al. (2014). Presence and genetic environment of pleuromutilin–lincosamide–streptogramin A resistance gene lsa(E) in enterococci of human and swine origin. J. Antimicrob. Chemother. 69, 1424–1439. doi: 10.1093/jac/dkt502
Lim, Y., Hyun, J., and Hwang, C. (2020). Identification of fusidic acid resistance in clinical isolates of Staphylococcus pseudintermedius from dogs in Korea. Vet. Dermatol. 31, 267–e62. doi: 10.1111/vde.12844
Lindsay, J. A., Ruzin, A., Rosse, H. F., Kurepina, N., and Novick, R. P. (1998). The gene for toxic shock toxin is carried by a family of mobile pathogenicity islands in Staphylococcus aureus. Mol. Microbiol. 29, 527–543. doi: 10.1046/j.1365-2958.1998.00947.x
Loeffler, A., Cain, C. L., Ferrer, L., Nishifuji, K., Varjonen, K., Papich, M. G., et al. (2025). Antimicrobial use guidelines for canine pyoderma by the international society for companion animal infectious diseases (ISCAID). Vet. Dermatol. 36, 234–282. doi: 10.1111/vde.13342
Lozano, C., Aspiroz, C., Sáenz, Y., Ruiz-García, M., Royo-García, G., Gómez-Sanz, E., et al. (2012). Genetic environment and location of the lnu(A) and lnu(B) genes in methicillin-resistant Staphylococcus aureus and other staphylococci of animal and human origin. J. Antimicrob. Chemother. 67, 2804–2808. doi: 10.1093/jac/dks320
Lynch, S. A., and Helbig, K. J. (2021). The complex diseases of Staphylococcus pseudintermedius in canines: Where to next? Vet. Sci. 8:11. doi: 10.3390/vetsci8010011
MacFadyen, A. C., and Paterson, G. K. (2024). Methicillin resistance in Staphylococcus pseudintermedius encoded within novel staphylococcal cassette chromosome mec (SCCmec) variants. J. Antimicrob. Chemother. 79, 1303–1308. doi: 10.1093/jac/dkae096
Malachowa, N., and DeLeo, F. R. (2010). Mobile genetic elements of Staphylococcus aureus. Cell. Mol. Life Sci. 67, 3057–3071. doi: 10.1007/s00018-010-0389-4
Manni, M., Berkeley, M. R., Seppey, M., Simão, F. A., and Zdobnov, E. M. (2021). BUSCO Update: Novel and streamlined workflows along with broader and deeper phylogenetic coverage for scoring of eukaryotic, prokaryotic, and viral genomes. Mol. Biol. Evol. 38, 4647–4654. doi: 10.1093/molbev/msab199
McCarthy, A. J., Harrison, E. W., Stanczak-Mrozek, K., Leggett, B., Waller, A., Holmes, M. A., et al. (2015). Genomic insights into the rapid emergence and evolution of MDR in Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 70, 997–1007. doi: 10.1093/jac/dku496
Menandro, M. L., Dotto, G., Mondin, A., Martini, M., Ceglie, L., and Pasotto, D. (2019). Prevalence and characterization of methicillin-resistant Staphylococcus pseudintermedius from symptomatic companion animals in Northern Italy: Clonal diversity and novel sequence types. Comp. Immunol. Microbiol. Infect. Dis. 66:101331. doi: 10.1016/j.cimid.2019.101331
Miller, J., Simpson, A., Bloom, P., Diesel, A., Friedeck, A., Paterson, T., et al. (2023). 2023 AAHA Management of allergic skin diseases in dogs and cats guidelines. J. Am. Anm. Hosp. Assoc. 59, 255–284. doi: 10.5326/JAAHA-MS-7396
Moodley, A., Kot, W., Nälgård, S., Jakociune, D., Neve, H., Hansen, L. H., et al. (2019). Isolation and characterization of bacteriophages active against methicillin-resistant Staphylococcus pseudintermedius. Res. Vet. Sci. 122, 81–85. doi: 10.1016/j.rvsc.2018.11.008
Moon, B. Y., Park, J. Y., Robinson, D. A., Thomas, J. C., Park, Y. H., Thornton, J. A., et al. (2016). Mobilization of genomic islands of Staphylococcus aureus by temperate bacteriophage. PLoS One 11:e0151409. doi: 10.1371/journal.pone.0151409
Morais, C., Costa, S. S., Leal, M., Ramos, B., Andrade, M., Ferreira, C., et al. (2023). Genetic diversity and antimicrobial resistance profiles of Staphylococcus pseudintermedius associated with skin and soft-tissue infections in companion animals in Lisbon, Portugal. Front. Microbiol. 14:1167834. doi: 10.3389/fmicb.2023.1167834
Morris, D. O., Loeffler, A., Favis, M. F., Guardabassi, L., and Weese, J. S. (2017). Recommendations for approaches to meticillin-resistant staphylococcal infections of small animals: Diagnosis, therapeutic considerations and preventative measures. Vet. Dermatol. 28, 304–e69. doi: 10.1111/vde.12444
Mortensen, K., Lam, T. J., and Ye, Y. (2021). Comparison of CRISPR–Cas immune systems in healthcare-related pathogens. Front. Microbiol. 12:758782. doi: 10.3389/fmicb.2021.758782
Naorem, R. S., Goswami, G., Gyorgy, S., and Fekete, C. (2021). Comparative analysis of prophages carried by human and animal-associated Staphylococcus aureus strains spreading across the European regions. Sci. Rep. 11:18994. doi: 10.1038/s41598-021-98432-8
O’Neill, A. J., Larsen, A. R., Henriksen, A. S., and Chopra, I. (2004). A fusidic acid-resistant epidemic strain of Staphylococcus aureus carries the fusB determinant, whereas fusA mutations are prevalent in other resistant isolates. Antimicrob. Agents Chemother. 48, 3594–3597. doi: 10.1128/AAC.48.9.3594-3597.2004
Oliveira, P. H., Touchon, M., and Rocha, E. P. C. (2016). Regulation of genetic flux between bacteria by restriction–modification systems. Proc. Natl. Acad. Sci. U.S.A. 113, 5658–5663. doi: 10.1073/pnas.1603257113
Papić, B., Golob, M., Zdovc, I., Kusar, D., and Avbersek, J. (2021). Genomic insights into the emergence and spread of methicillin-resistant Staphylococcus pseudintermedius in veterinary clinics. Vet. Microbiol. 258:109119. doi: 10.1016/j.vetmic.2021.109119
Partridge, S. R., Kwong, S. M., Firth, N., and Jensen, S. O. (2018). Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 31, e88–e17. doi: 10.1128/CMR.00088-17
Perreten, V., Chanchaithong, P., Prapasarakul, N., Rossano, A., Blum, S. E., Elad, D., et al. (2013). Novel pseudo-staphylococcal cassette chromosome mec element (ψSCCmec57395) in methicillin-resistant Staphylococcus pseudintermedius CC45. Antimicrob. Agents Chemother. 57, 5509–5515. doi: 10.1128/AAC.00738-13
Phumthanakorn, N., Schwendener, S., Donà, V., Chanchaithong, P., Perreten, V., and Prapasarakul, N. (2021). Genomic insights into methicillin-resistant Staphylococcus pseudintermedius isolates from dogs and humans of the same sequence types reveals diversity in prophages and pathogenicity islands. PLoS One 16:e0254382. doi: 10.1371/journal.pone.0254382
Robb, A. R., Ure, R., Chaput, D. L., and Foster, G. (2024). Emergence of novel methicillin resistant Staphylococcus pseudintermedius lineages revealed by whole genome sequencing of isolates from companion animals and humans in Scotland. PLoS One 19:e0305211. doi: 10.1371/journal.pone.0305211
Roberts, R. J., Vincze, T., Posfai, J., and Macelis, D. (2015). REBASE-a database for DNA restriction and modification: Enzymes, genes and genomes. Nucleic Acids Res. 43, D298–D299. doi: 10.1093/nar/gku1046
Rossi, C. C., Andrade-Oliveira, A. L., and Giambiagi-deMarval, M. (2019). CRISPR tracking reveals global spreading of antimicrobial resistance genes by Staphylococcus of canine origin. Vet. Microbiol. 232, 65–69. doi: 10.1016/j.vetmic.2019.04.009
Rossi, C. C., Souza-Silva, T., Araújo-Alves, A. V., and Giambiagi-deMarval, M. (2017). CRISPR-Cas Systems Features and the rene-reservoir role of Coagulase-Negative Staphylococci. Front. Microbiol. 8:1545. doi: 10.3389/fmicb.2017.01545
Sadykov, M. R. (2016). Restriction–modification systems as a barrier for genetic manipulation of Staphylococcus aureus. Methods Mol. Biol. 1373, 9–23. doi: 10.1007/7651_2014_180
Schwarz, S., Feßler, A. T., Hauschild, T., Kehrenberg, C., and Kadlec, K. (2011). Plasmid-mediated resistance to protein biosynthesis inhibitors in staphylococci. Ann. N. Y. Acad. Sci. 1241, 82–103. doi: 10.1111/j.1749-6632.2011.06275.x
Schwarz, S., Feßler, A. T., Loncaric, I., Wu, C., Kadlec, K., Wang, Y., et al. (2018). Antimicrobial resistance among staphylococci of animal origin. Microbiol. Spectr. 6:10. doi: 10.1128/microbiolspec.ARBA-0010-2017
Schwarz, S., Roberts, M. C., Werckenthin, C., Pang, Y., and Lange, C. (1998). Tetracycline resistance in Staphylococcus spp. from domestic animals. Vet. Microbiol. 63, 217–227. doi: 10.1016/s0378-1135(98)00234-x
Schwarz, S., Shen, J., Wendlandt, S., Feßler, A. T., Loncaric, I., Kadlec, K., et al. (2014). Plasmid-mediated antimicrobial resistance in Staphylococci and other Firmicutes. Microbiol. Spectr. 2:20. doi: 10.1128/microbiolspec.PLAS-0020-2014
Schwarz, S., Werckenthin, C., Pinter, L., Kent, L. E., and Noble, W. C. (1995). Chloramphenicol resistance in Staphylococcus intermedius from a single veterinary centre: Evidence for plasmid and chromosomal location of the resistance genes. Vet. Microbiol. 43, 151–159. doi: 10.1016/0378-1135(94)00092-b
Schwengers, O., Jelonek, L., Dieckmann, M. A., Beyvers, S., Blom, J., and Goesmann, A. (2021). Bakta: Rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microb. Genom. 7:685. doi: 10.1099/mgen.0.000685
Senok, A., Slickers, P., Hotzel, H., Boswihi, S., Braun, S. D., Gawlik, D., et al. (2019). Characterisation of a novel SCCmec VI element harbouring fusC in an emerging Staphylococcus aureus strain from the Arabian Gulf region. PLoS One 14:e0223985. doi: 10.1371/journal.pone.0223985
Soimala, T., Lübke-Becker, A., Hanke, D., Eichhorn, I., Feßler, A. T., Schwarz, S., et al. (2020). Molecular and phenotypic characterization of methicillin-resistant Staphylococcus pseudintermedius from ocular surfaces of dogs and cats suffering from ophthalmological diseases. Vet. Microbiol. 244:108687. doi: 10.1016/j.vetmic.2020.108687
Starikova, E. V., Tikhonova, P. O., Prianichnikov, N. A., Rands, C. M., Zdobnov, E. M., Ilina, E. N., et al. (2020). Phigaro: High-throughput prophage sequence annotation. Bioinformatics 36, 3882–3884. doi: 10.1093/bioinformatics/btaa250
Sweeney, M. T., Lubbers, B. V., Schwarz, S., and Watts, J. L. (2018). Applying definitions for multidrug resistance, extensive drug resistance and pandrug resistance to clinically significant livestock and companion animal bacterial pathogens. J. Antimicro. Chemother. 73, 1460–1463. doi: 10.1093/jac/dky043
Viñes, J., Cuscó, A., and Francino, O. (2020). Hybrid assembly from a pathogenic methicillin- and multidrug-resistant Staphylococcus pseudintermedius strain isolated from a case of canine otitis in Spain. Microbiol. Resour. Announc. 9:e01121–19. doi: 10.1128/MRA.01121-19
Viñes, J., Fàbregas, N., Pérez, D., Cuscó, A., Fonticoba, R., Francino, O., et al. (2022). Concordance between antimicrobial resistance phenotype and genotype of Staphylococcus pseudintermedius from Healthy Dogs. Antibiotics 11:1625. doi: 10.3390/antibiotics11111625
Viñes, J., Verdejo, M., Horvath, L., Vergara, A., Vila, J., Francino, O., et al. (2024). Isolation of Staphylococcus pseudintermedius in immunocompromised patients from a single center in Spain: A zoonotic pathogen from companion animals. Microorganisms 12:1695. doi: 10.3390/microorganisms12081695
Weese, J. S., Sweetman, K., Edson, H., and Rousseau, J. (2013). Evaluation of minocycline susceptibility of methicillin-resistant Staphylococcus pseudintermedius. Vet. Microbiol. 162, 968–971. doi: 10.1016/j.vetmic.2012.10.002
Wegener, A., Broens, E. M., Bloois, L., Zomer, A. L., Visser, C. E., van Zeijl, J., et al. (2021). Absence of Host-Specific Genes in canine and human Staphylococcus pseudintermedius as inferred from comparative genomics. Antibiotics 10:854. doi: 10.3390/antibiotics10070854
Wegener, A., Damborg, P., Guardabassi, L., Moodley, A., Mughini-Gras, L., Duim, B., et al. (2020). Specific staphylococcal cassette chromosome mec (SCCmec) types and clonal complexes are associated with low-level amoxicillin/clavulanic acid and cefalotin resistance in methicillin-resistant Staphylococcus pseudintermedius. J. Antimicrob. Chemother. 75, 508–511. doi: 10.1093/jac/dkz509
Wegener, A., Duim, B., van der Graaf-van, L. B., Zomer, A. L., Visser, C. E., Spaninks, M., et al. (2022). Within-household transmission and bacterial diversity of Staphylococcus pseudintermedius. Pathogens 11:850. doi: 10.3390/pathogens11080850
Werckenthin, C., Cardoso, M., Martel, J. L., and Schwarz, S. (2001). Antimicrobial resistance in staphylococci from animals with particular reference to bovine Staphylococcus aureus, porcine Staphylococcus hyicus, and canine Staphylococcus intermedius. Vet. Res. 32, 341–362. doi: 10.1051/vetres:2001129
Werner, G., Hildebrandt, B., and Witte, W. (2003). Linkage of erm(B) and aadE-sat4-aphA-3 in multiple-resistant Enterococcus faecium isolates of different ecological origins. Microb. Drug Resist. 9, S9–S16. doi: 10.1089/107662903322541847
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13:e1005595. doi: 10.1371/journal.pcbi.1005595
Wick, R. R., Schultz, M. B., Zobel, J., and Holt, K. E. (2015). Bandage: Interactive visualization of de novo genome assemblies. Bioinformatics 31, 3350–3352. doi: 10.1093/bioinformatics/btv383
Wildermuth, B. E., Griffin, C. E., and Rosenkrantz, W. S. (2006). Feline pyoderma therapy. Clin. Tech. Small Anim. Pract. 21, 150–156. doi: 10.1053/j.ctsap.2006.05.008
Wipf, J. R. K., Deutsch, D. R., Westblade, L. F., and Fischetti, V. A. (2019). Genome sequences of six prophages isolated from Staphylococcus pseudintermedius strains recovered from human and animal clinical specimens. Microbiol. Resour. Announc. 8:e00387–19. doi: 10.1128/MRA.00387-19
Wladyka, B., Piejko, M., Bzowska, M., Pieta, P., Krzysik, L. M., Guevara-Lora, I., et al. (2015). A peptide factor secreted by Staphylococcus pseudintermedius exhibits properties of both bacteriocins and virulence factors. Sci. Rep. 5:14569. doi: 10.1038/srep14569
Worthing, K. A., Abraham, S., Coombs, G. W., Pang, S., Saputra, S., Jordan, A., et al. (2018a). Clonal diversity and geographic distribution of methicillin-resistant Staphylococcus pseudintermedius from Australian animals: Discovery of novel sequence types. Vet. Microbiol. 213, 58–65. doi: 10.1016/j.vetmic.2017.11.018
Worthing, K. A., Marcus, A., Abraham, S., Trott, D. J., and Norris, J. M. (2018b). Qac genes and biocide tolerance in clinical veterinary methicillin-resistant and methicillin-susceptible Staphylococcus aureus and Staphylococcus pseudintermedius. Vet. Microbiol. 216, 153–158. doi: 10.1016/j.vetmic.2018.02.004
Worthing, K. A., Schwendener, S., Perreten, V., Saputra, S., Coombs, G. W., Pang, S., et al. (2018c). Characterization of staphylococcal cassette chromosome mec elements from methicillin-resistant Staphylococcus pseudintermedius infections in Australian animals. mSphere 3:e00491–18. doi: 10.1128/mSphere.00491-18
Zehr, J. D., Sun, Q., Ceres, K., Merrill, A., Tyson, G. H., Ceric, O., et al. (2025). Population and pan-genomic analyses of Staphylococcus pseudintermedius identify geographic distinctions in accessory gene content and novel loci associated with AMR. Appl. Environ. Microbiol. 91:e0001025. doi: 10.1128/aem.00010-25
Zhang, J. M., Wang, Q., Han, T. Y., Liu, J. H., Hu, X. X., Qiao, F., et al. (2018). Structure analysis of transposons carrying the aac(6′)-aph(2″) gene in Enterococcus faecalis isolated in Beijing, China, and comparison of their transfer efficiency. Int. J. Antimicrob. Agents 52, 799–804. doi: 10.1016/j.ijantimicag.2018.08.025
Zhang, Y., Ren, S., Li, H., Wang, Y., Fu, G., Yang, J., et al. (2003). Genome-based analysis of virulence genes in a non-biofilm-forming Staphylococcus epidermidis strain (ATCC 12228). Mol. Microbiol. 49, 1577–1593. doi: 10.1046/j.1365-2958.2003.03671.x
Zhou, Y., Liang, Y., Lynck, K. H., Dennis, J. J., and Wishart, D. S. (2011). PHAST: A fast phage search tool. Nucleic Acids Res. 39, W347–W352. doi: 10.1093/nar/gkr485
Keywords: Staphylococcus pseudintermedius, mobilome, antimicrobial resistance, WGS, plasmids, transposons, SCCfus, mobile genetic elements
Citation: Morais C, Costa SS, Hanke D, Santos A, Krüger-Haker H, Pomba C, Feßler AT, Schwarz S and Couto I (2025) Genomic analysis of the Staphylococcus pseudintermedius mobilome associated with antimicrobial resistance. Front. Microbiol. 16:1640322. doi: 10.3389/fmicb.2025.1640322
Received: 03 June 2025; Accepted: 01 September 2025;
Published: 08 October 2025.
Edited by:
Hazem Ramadan, Mansoura University, EgyptReviewed by:
Mehmet Cemal Adiguzel, Atatürk University, TürkiyeRachana Banerjee, JIS Institute of Advanced Studies and Research, India
Manuela Iurescia, Institute of Experimental Zooprophylactic of the Lazio and Tuscany Regions (IZSLT), Italy
Copyright © 2025 Morais, Costa, Hanke, Santos, Krüger-Haker, Pomba, Feßler, Schwarz and Couto. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Isabel Couto, aWNvdXRvQGlobXQudW5sLnB0