 Angelika Mańkowska1
Angelika Mańkowska1 Paulina Paprocka1
Paulina Paprocka1 Łukasz Suprewicz2Agata Lesiak1Kamila Fortunka1Grzegorz Król1
Łukasz Suprewicz2Agata Lesiak1Kamila Fortunka1Grzegorz Król1 Jakub Spałek3,4
Jakub Spałek3,4 Sławomir Okła3,4Bonita Durnaś1Tamara Daniluk2Ewelina Piktel5
Sławomir Okła3,4Bonita Durnaś1Tamara Daniluk2Ewelina Piktel5 Paul B. Savage6
Paul B. Savage6 Robert Bucki1,2*
Robert Bucki1,2*- 1Department of Microbiology and Immunology, Institute of Medical Science, Collegium Medicum, Jan Kochanowski University in Kielce, Kielce, Poland
- 2Department of Medical Microbiology and Nanobiomedical Engineering, Medical University of Białystok, Białystok, Poland
- 3Institute of Medical Science, Collegium Medicum, Jan Kochanowski University in Kielce, Kielce, Poland
- 4Department of Otolaryngology, Head and Neck Surgery, Holy-Cross Oncology Center of Kielce, Kielce, Poland
- 5Independent Laboratory of Nanomedicine, Medical University of Białystok, Białystok, Poland
- 6Department of Chemistry and Biochemistry, Brigham Young University, Provo, UT, United States
Introduction: The constantly growing resistance of bacteria causing bloodstream infections and the lack of alternative antibiotics generate the need to develop new therapeutic strategies. In this study, the antibacterial properties of synthetic cholic acid derivatives ceragenins CSA-13, CSA-44 and CSA-131, custom-synthesized peptides human cathelicidin LL-37 peptide, synthetic WLBU2 peptide, and antimicrobial VFR12 peptide of human thrombin origin were evaluated to determine their potentials as therapeutic agents for bloodstream infections.
Methods: Minimum inhibitory concentrations/minimum bactericidal concentrations (MIC/MBC) against clinical bacterial strains were measured and compared with activity of clinically used antibiotics colistin and vancomycin. Therapeutic potentials of the tested agents were assessed in the presence of 50% blood plasma, and their hemolytic properties were determined using human red blood cells (RCB). Additionally, the antimicrobial activity of CSA-13 against selected clinical strains was assessed using a killing assay. Plasma cytokine levels were determined, and endothelial cell confluent monolayer permeability was assessed using the FITC-dextran and measurement of transepithelial electrical resistance (TEER).
Results: Under experimental conditions mimicking blood environment, ceragenins display higher antimicrobial activity compared to the cationic peptides regardless of the bacterial species. The presence of blood plasma slightly decreases the effect of ceragenins but does not significantly affect their antibacterial properties or their hemolytic activity, especially in case of ceragenin CSA-13. Furthermore, ceragenins at bactericidal concentrations do not induce hemolysis of red blood cells. CSA-13 dose-dependently regulates the permeability of human umbilical vein endothelial cells (HUVECs) monolayers as well as affects the secretion of cytokines, which may indicate its ability to modulate immune responses.
Conclusion: Results presented herein demonstrate the antibacterial activity of ceragenins against clinical strains of bacteria isolated from blood, their influence on the immune system and the integrity of the endothelial cell monolayer. Further studies are necessary to understand the cell signaling pathway governing these effects.
1 Introduction
Antimicrobial-resistant strains of microorganisms are a major cause of mortality in patients with bloodstream infections. This threat is particularly significant among immunocompromised patients with risk factors, such as damage to mucous membranes and skin, chemotherapy-induced neutropenia, or the use of immunosuppressive drugs in transplant recipients. It is especially concerning in those who have undergone hematopoietic stem cell transplants, as well as in patients with blood cancers or autoimmune disorders (Wang et al., 2024; de Souza et al., 2024; Pezzani et al., 2024). The growing threat of this clinical form of infection requires the search for new therapeutic strategies, especially antimicrobial drugs active against strains of antibiotic-resistant bacteria (Papadimitriou-Olivgeris et al., 2022). A promising experimental direction is the search for new antibiotics that are synthesized using the molecular characteristics of natural antimicrobial peptides that are widely present in nature (Nandi et al., 2022). An example of such molecules is the group of ceragenins (Figure 1A), which are derivatives of cholic acid, characterized by a positive charge and amphipathic nature, which determines their interactions with bacterial membranes and the ability to interfere with membrane structures, resulting in membrane reorganization and leakage (Epand et al., 2008). Ceragenins mimic the membrane-disrupting properties of antimicrobial peptides (Lai et al., 2008). This molecular mechanism of antimicrobial action, based on the physicochemical interaction of molecular charges and membrane insertion (Figure 1B), results in a wide spectrum of antimicrobial activity that covers Gram-positive G (+) and Gram-negative G (−) bacteria, as well as viruses and fungi (Hashemi et al., 2018; Bozkurt-Guzel et al., 2018; Suprewicz et al., 2023). Ceragenins, like natural antimicrobial peptides, have the ability to stimulate the immune system (Suprewicz et al., 2023) and wound healing (Olekson et al., 2017). They also display high activity against bacterial cells surrounded by exopolysaccharides when adopting a biofilm pattern of growth (Wnorowska et al., 2024). The therapeutic potential of ceragenins is also related to the possibility of using metal nanoparticles as carriers for their delivery to sites of infection (Yang et al., 2024; Karasinski et al., 2023; Wnorowska et al., 2020; Hoppens et al., 2014) and control of their toxicity towards host cells using Pluronic F127 (Leszczyńska et al., 2011). They can also serve as molecules that cover the surface of medical devices, preventing colonization and infections associated with their use (Latorre et al., 2021; Mills et al., 2020; Spałek et al., 2021; Zaugg et al., 2023).
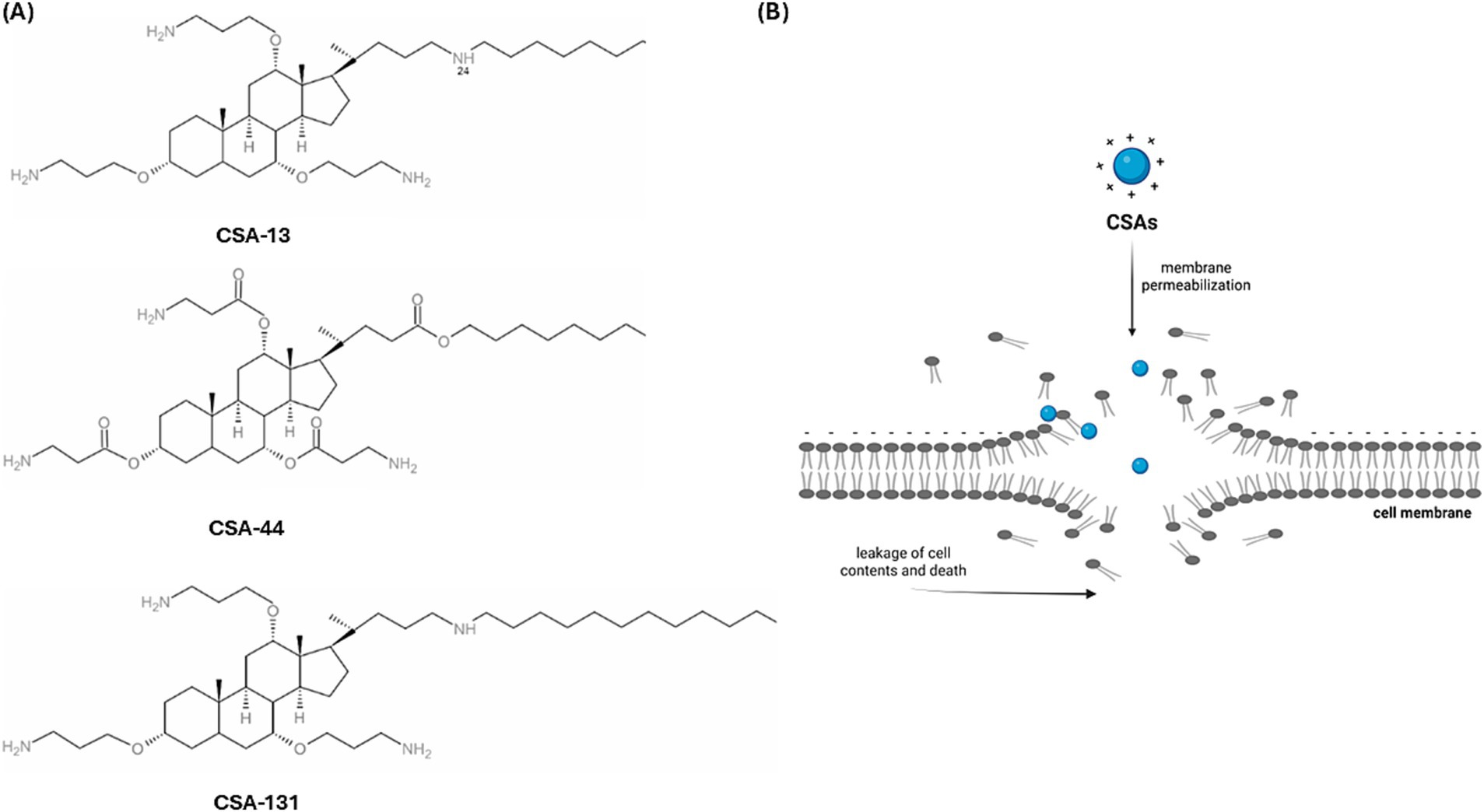
Figure 1. The chemical formula of ceragenins, which are derivatives of cholic acid (A) (Karasiński et al., 2024) Schematic representation of the action of ceragenin on the plasma membrane, which results in damage to its continuity (B) [Biorender.io software].
A critical element in the pathophysiology of sepsis is endothelial dysfunction, which correlates with disease severity and mortality. The endothelium performs several vital functions in the human body. It constitutes a barrier between blood and tissues (Dolmatova et al., 2021). Increased endothelial permeability accompanies many pathological conditions, including: atherosclerosis, diabetes, hypertension, cancer metastases and sepsis (Harris and Nelson, 2010), where it leads to the inability to maintain the volume of the vascular system, leading to hypoxia and impaired functions of internal organs (Lee and Slutsky, 2010). Here we evaluate the bactericidal activity of ceragenins CSA-13, CSA-44, and CSA-131 against bacteria isolated from the blood of oncological patients in an experimental setting that includes the presence of human blood plasma or whole human blood. This assessment was motivated by previous reports indicating limited activity of antibacterial peptides in the presence of blood, especially blood lipoproteins and divalent cations (Deslouches et al., 2005; Wang et al., 2004; Wang et al., 1998; Vaara, 2009). Since previous studies have shown that a natural antimicrobial peptide, cathelicidin LL-37, regulates the permeability of the endothelium (Byfield et al., 2011), we assessed whether the effect of its synthetic analogue, CSA-13, would be conserved. Restoring vascular integrity using synthetic analogues of natural antimicrobial peptides may constitute a new therapeutic direction in patients with sepsis.
2 Materials and methods
2.1 Reference strains and clinical isolates
In accordance with European Committee on Antimicrobial Susceptibility Testing (EUCAST) recommendations (EUCAST, 2023), four reference strains were used for quality control of MIC determinations. This group included: Escherichia coli ATCC 25922, Pseudomonas aeruginosa ATCC 27853, Enterococcus faecalis ATCC 29212, and Staphylococcus aureus ATCC 29213 (all purchased from the American Type Culture Collection, Manassas, United States). The study also included 83 clinical strains isolated from the blood of patients hospitalized at the Świętokrzyskie Oncology Center in Kielce, Poland, with clinical symptoms of bloodstream infection. Clinical isolates were obtained from a bank of strains collected between 2019 and 2023 at the Department of Clinical Microbiology of the Świętokrzyskie Oncology Center in Kielce, Poland. Along with the strains, information including the patient’s gender, age, and diagnosis was recorded. The characterization of this collection of strains is presented in Figure 2. The study was approved by the Bioethics Committee of the Collegium Medicum of the Jan Kochanowski University in Kielce (No. 17/2023) and the guidelines contained in the Declaration of Helsinki were followed. The patient’s consent was not necessary because the material used for the research was the leftover material collected during laboratory tests. Before the tests, the samples were plated on ready-made Columbia Agar with sheep blood (Oxoid, United Kingdom) and incubated for 24 h in aerobic conditions at 37 °C. Then, single colonies were identified on a Vitek 2 Compact and Vitek MS Prime MALDI-TOF mass spectrophotometer (bioMérieux, France). Strains for further studies were stored in the MAST CRYOBANK system (Mast Diagnostic, United Kingdom) at −80 °C.
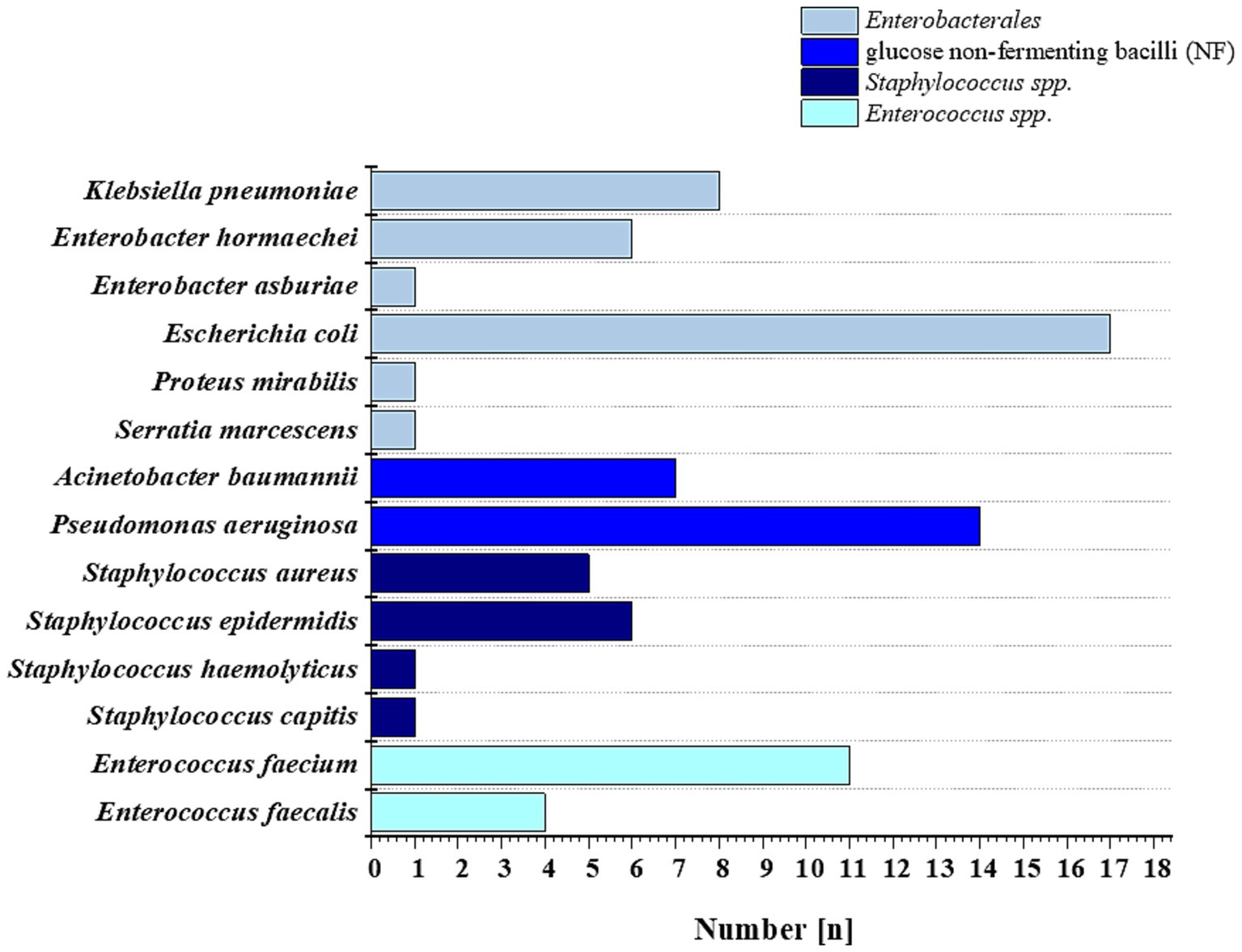
Figure 2. Bacterial strains used in this study were isolated from the blood of patients of the Świętokrzyskie Oncology Center in Kielce, Poland, in 2019–2023.
2.2 Tested compounds
Ceragenins CSA-13, CSA-44, and CSA-131 were synthesized according to previously described procedures (Ding et al., 2002). Since ceragenins are synthetic analogues of antimicrobial peptides, are positively charged, have an amphipathic nature, and are able of inserting into plasma membrane structures, their activity was compare with that of human cathelicidin LL-37 (LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES), which has an α-helical structure containing positively charged amino acids Lys and Arg on the hydrophilic side and several different hydrophobic amino acid residues (Phe, Val, Ile) in the hydrophobic domain (Deslouches et al., 2005; Gudmundsson et al., 1996). The LL-37 peptide is characterized by a broad spectrum of antibacterial activity. Additionally, we decided to evaluate the activity of a synthetic analogue of the LL-37 peptide, namely the WLBU2 peptide (RRWVRRVRRWVRRVVRVVRRWVRR), consisting mainly of Arg (13 residues) and Val (8 residues), with 3 Trp residues on the hydrophobic side, separated by at least 7 amino acids to optimize its activity in blood (Deslouches et al., 2005), and the VFR12 (VFRLKKWIQKVI) peptide, which is derived from thrombin, taking into account the fact that coagulation disorders occur in sepsis (Kasetty et al., 2011) and it can be assumed that its concentration increases with sepsis development. Custom synthesis of the tested peptides was performed at Lipopharm.pl (Zblewo, Poland). Colistin and vancomycin were purchased from Sigma Aldrich (Saint Louis, United States) and Pol-aura (Morąg, Poland) respectfully. Phosphate-buffered saline (PBS) was purchased from Thermo Fisher Scientific, United States.
2.3 Assessment of antimicrobial activity
Minimum inhibitory concentrations (MIC) and minimum bactericidal concentrations (MBC) of CSA-13, CSA-44, CSA-131, LL-37, WLBU2, VFR12, colistin, and vancomycin against tested bacterial strains were determined using the serial broth microdilution method in accordance with EUCAST recommendations (EUCAST, 2024). Mueller-Hinton Broth (Oxoid, United Kingdom) or Mueller-Hinton Broth with 50% plasma was used to prepare a series of two-fold dilutions of the appropriate antimicrobial substance, ranging from 0.25 μg/mL to 64 μg/mL. Plasma was obtained by collecting whole blood from healthy, adult donors who gave informed consent (Study was approved by Bioethics Committee of the Collegium Medicum of the Jan Kochanowski University in Kielce – no. 17/2023). Blood was centrifuged at 2,500 g × 10 min and the plasma was collected. A suspension of each strain was then prepared to reach a final concentration of bacteria equal to 5 × 105 CFU/mL. MIC values were taken as the first value at which no turbidity or sediment was observed at the bottom of the well after 18 ± 2 h of bacterial growth. MIC endpoints were assessed visually. To determine MBC, 10 μL of the suspension was taken from the wells corresponding to 1/2MIC, MIC, MICx2 and MICx4, inoculated on LB Agar (BD, Becton Dickinson, Le Pont de Claix, France) and incubated for 24 h at 37 °C. The lowest value at which no growth was observed on solid media was considered the MBC.
2.4 Hemolysis measurement
Human red blood cells (RBC) were used to assess the biocompatibility of the tested compounds (CSA-13, CSA-44, CSA-131, LL-37, WLBU2, VFR12, colistin and vancomycin) with respect to host cells. The test was performed in 96-well plates containing RBCs (hematocrit ~ 5%) and tested agents at concentrations ranging from 1 μg/mL to 50 μg/mL. RBCs were suspended in PBS and incubated at 37 °C for 1, 6, and 12 h. After incubation, the plates were centrifuged at 2500 g × 10 min, and the supernatant was transferred to new plates. The amount of hemoglobin released was determined by measuring the absorbance at 540 nm using a Tecan Spark plate reader (Tecan, Männedorf, Switzerland). A sample of RBC containing 1% Triton X-100 was considered as 100% of hemolysis. A value up to 10% is considered non-toxic to the erythrocyte membrane, above 10–49% moderately toxic, 50–89% toxic and 90–100% highly toxic (Pedrozo-Peñafiel et al., 2025). In another set of experiments, the hemolytic activity of CSA-13 was assessed in whole blood. In this setting CSA-13 was tested at 20 μg/mL and 50 μg/mL in 96-well plates and incubated with whole blood. We also assessed if CSA-13 addition affects the response of blood cells to the presence of tested bacterial isolates, including Pseudomonas aeruginosa, Escherichia coli and Enterococcus faecium. The prepared bacterial suspension was added to blood sample at final concentration of 105 CFU/mL and measurements were carried out at time intervals of 1, 3, 6 and 8 h at a temperature of 37 °C. The plates were then centrifuged at 2,500 g for 10 min, the collected sediment was transferred to a sterile plate and the absorbance was measured at 540 nm using a Tecan Spark (Tecan, Männedorf, Switzerland). The positive control was a sample with 1% Triton X-100 (100% hemolysis). This study was conducted as previously described (Chmielewska et al., 2020).
2.5 Time-kill kinetics assay in whole blood
A time-kill assay was performed to determine the bactericidal activity of CSA-13. The effect of CSA-13 was also tested in the presence of whole blood against three clinical strains selected from the tested collection: P. aeruginosa, E. coli and E. faecium. Individual bacterial colonies were suspended at a concentration of 108 CFU/mL and diluted to 105 CFU/mL in PBS or whole blood. Blood was collected in heparin tubes from healthy volunteers. In 96-well plates, CSA-13 was prepared in the concentration range of 0.5–50 μg/mL, and then a previously prepared bacterial suspension was added. Incubations were performed in time intervals: 1, 3, 6 and 8 h at 37 °C. The plates were then placed on ice to inhibit the action of the compounds. Dilutions of 1:10, 1:100, and 1:1,000 were made in PBS, and 10 μL of each sample was inoculated onto LB agar and incubated overnight at 37 °C. Colony-forming units (CFU/mL) were then determined for each sample based on the dilution factor.
2.6 Determination of cytokine levels in plasma
Cytokine secretion was assessed using the Human Cytokine Array Kit (Bio-techne, R&D Systems, Minneapolis, MN). Four conditions were prepared on a 96-well plate: whole blood, whole blood + CSA-13 (20 μg/mL), whole blood + CSA-13 (20 μg/mL) + P. aeruginosa and whole blood + P. aeruginosa and incubated for 6 h at 37 °C. P. aeruginosa was suspended in whole blood at 105 CFU/mL. The plates were centrifuged, and the supernatant (plasma) was collected and processed according to the manufacturer’s protocol. Membranes were imaged using a Chemidoc imaging system (Bio-Rad, United States). The image was imported into ImageStudio to quantify the pixel density of each protein on the membrane.
2.7 Cell culture
Human umbilical vein endothelial cells (HUVECs) purchased from Sigma Aldrich (Saint Louis, United States) were used for the experiment. They were cultured in an endothelial cell growth medium (#211-500, Cell Applications, United States), supplemented with antibiotics, and maintained at 37 °C in a humidified incubator with 5% CO2. After reaching 80–90% confluence, the cells were used for further experiments.
2.8 Assessment of endothelial barrier permeability and integrity
To recreate the functions of the endothelial barrier, cells were seeded at a density of 104 cells per transwell insert coated with collagen I (pore size 0.4 μm, diameter 0.33 cm2 #3470, Corning, United States) in 200 μL of complete growth medium. Basolateral chambers were filled with 700 μL of complete growth medium. The medium was changed every 3 days. The formation of the monolayer was confirmed by TEER assessment, reaching maximum, stable values after 5–7 days. After the formation of the monolayer, cells were washed and incubated with CSA-13 at concentrations of 1, 5, and 20 μg/mL, heat-inactivated (autoclaved) bacteria (HI) 108 CFU/mL, and their combinations. Bacterial strains, P. aeruginosa, and E. faecium were isolated from patients diagnosed with sepsis. A schematic representation of the experimental setup is presented in Figure 3. The barrier integrity measurements were performed before (time 0) and after the addition of the compounds at 1, 3, and 6 h. To evaluate the permeability of the endothelial monolayer, a 4-kDa dextran-FITC (#46944, Sigma Aldrich, United States) was used. The tracer was introduced to the apical chamber of the Transwell at a final concentration of 1 mg/mL. The fluorescent intensity in the lower chamber was tracked using a Varioskan Lux microplate reader (Thermo Fisher Scientific, United States). Percentage permeability was calculated as the relative fluorescence of the medium in treated versus untreated conditions.
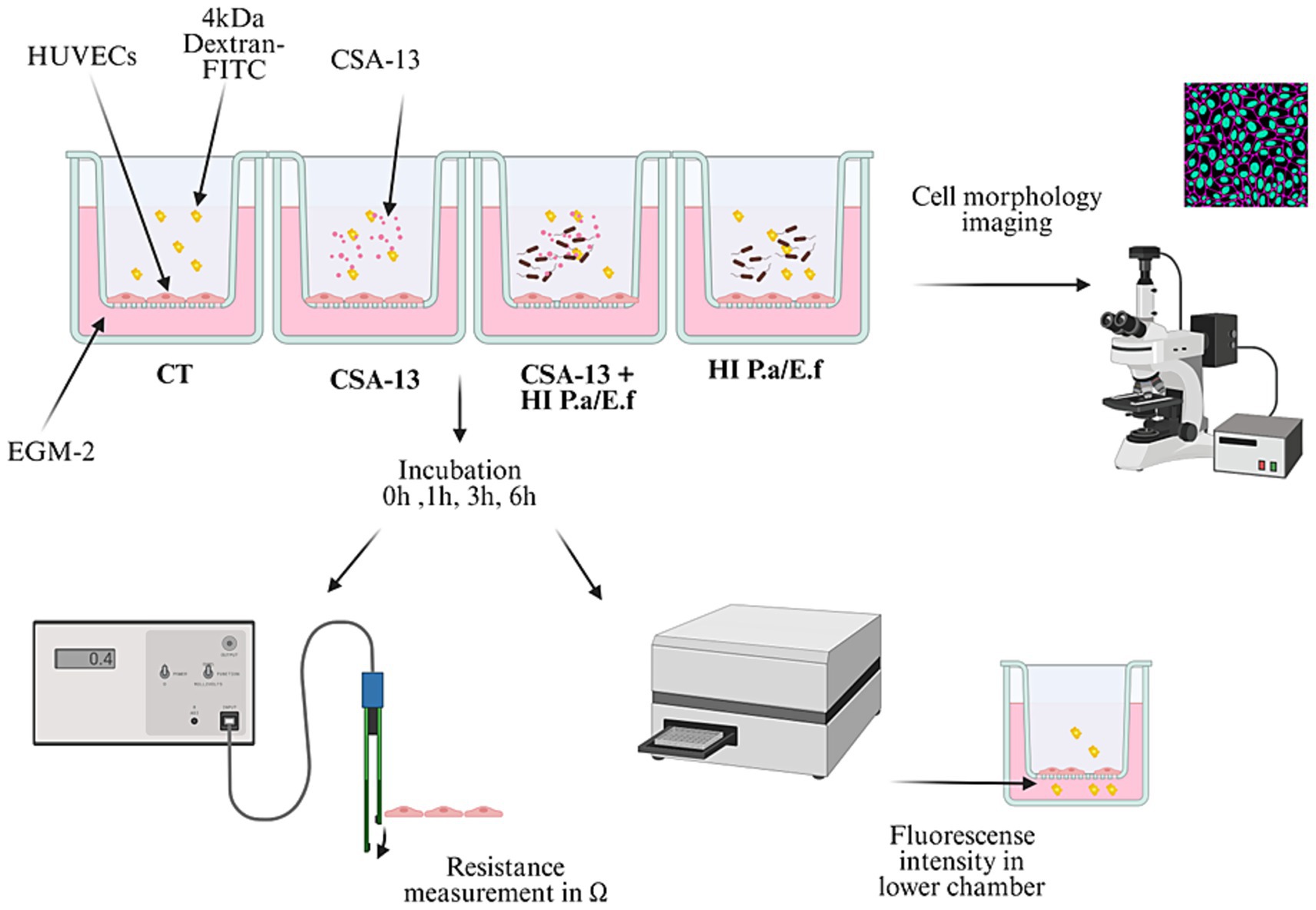
Figure 3. Schematic representation of experimental setting to conduct the permeability/integrity assays using HUVECs monolayers upon addition of ceragenins or their combination with heat-inactivated bacteria [Biorender.io software].
At the same time, transendothelial electrical resistance (TEER) was used to assess the permeability of the endothelial barrier. Each sample was measured separately using an EVOM voltmeter (World Precision Instruments, United States) equipped with an STX-2 stick electrode. Barrier resistance readings (Ω) were obtained for each well and, after subtracting the resistance of the blank, were multiplied by the membrane area (0.33 cm2) to calculate Ω*cm2. The resistance at time 0 for each sample was set to 1.0 to normalize the data. Values of treated samples were normalized to those of an untreated sample (Suprewicz et al., 2022).
2.9 VE-cadherin imaging
After 72 h of incubation, treated and untreated sample inserts with HUVECs were washed with PBS and fixed in 4% paraformaldehyde (Sigma Aldrich, United States) for 20 min at room temperature. Cells were permeabilized by adding 0.1% Triton X-100 for 10 min at room temperature and then blocked in 0.3% BSA for 30 min at 37 °C. Samples were then incubated for 48 h at 4 °C with a 1:500 dilution of mouse VE-cadherin monoclonal antibody (Invitrogen, United States). Samples were washed with PBS and incubated with AlexaFluor 647-conjugated secondary antibody (1:1,000) for 1 h in the dark at room temperature. The nuclei were counterstained with Hoechst for 30 min in the dark. Then, the membranes in the cell monolayer were cut out, placed onto a glass coverslip, and mounted with an antifade solution (Abcam, United Kingdom). Fluorescence images were obtained using a Leica microscope DMi8 (Suprewicz et al., 2022).
2.10 Statistical analysis
Experiments were performed in 3–6 replicates, values presented are mean ± standard deviation (SD)/standard error (SE). Depending on the type of study, experiments were performed as technical or biological replicates, as described in the figure legends. Significance of differences was determined using a two-tailed Student’s t-test with GraphPad software (San Diego, CA) or a one-way ANOVA with Tukey’s post hoc test. p ≤ 0.05*, p < 0.02**, p < 0.001 *** were considered statistically significant.
3 Results
3.1 Antibacterial activity of tested agents
Sensitivity testing of clinical isolates showed that the MIC values of the tested molecules differ depending on the bacterial species. The results are presented in ranges (lower-higher MIC/MBC values) corresponding to species of G (−) and G (+) bacteria (Tables 1, 2, respectively). These data show that in Mueller Hinton Broth (MH) the lowest MIC for E. coli was obtained using CSA-13 (0.5–1 μg/mL), for K. pneumoniae CSA-131 (1–8 μg/mL), E. hormaechei CSA-44 and CSA-131 (1–4 μg/mL), E. asburiae CSA-13 (2 μg/mL), S. marcescens CSA-13 and CSA-131 (8 μg/mL), A. baumannii CSA-131 (1–2 μg/mL) and P. aeruginosa CSA-13 and CSA131 (1–2 μg/mL). Interestingly, MICs for CSAs against P. mirabilis exceeded > 64 μg/mL, suggesting low susceptibility (natural resistance) of this strain to the tested compounds. In the presence of 50% plasma, CSA-13 retained the highest activity except for activity against E. asburiae and P. mirabilis (> 64 μg/mL). Generally, in the presence of human plasma, the antibacterial activities of CSA-44 and CSA-131 were decreased compared to their activity in MH media. As for cationic peptides, the lowest MIC values were recorded in the presence of the WLBU2 peptide for most of the above-mentioned species, except S. marcescens and P. mirabilis, where the MIC was outside the range of tested concentrations > 64 μg/mL. Considering that the MIC values for ceragenins of these two strains were above 64 μg/mL, it is suggested that both strains display lower susceptibility to the tested molecules, whose antibacterial activity requires membrane insertion. This study also confirmed the limited activity of cationic amphipathic peptides in the presence of blood components, particularly for LL-37 and VFR12, where the range of values exceeded 64 μg/mL. Low colistin MIC values were obtained for most of the tested bacterial strains (except for 2 colistin-resistant (R) strains and Serratia marcescens and Proteus mirabilis, which have natural resistance to colistin) (Aghapour et al., 2019).
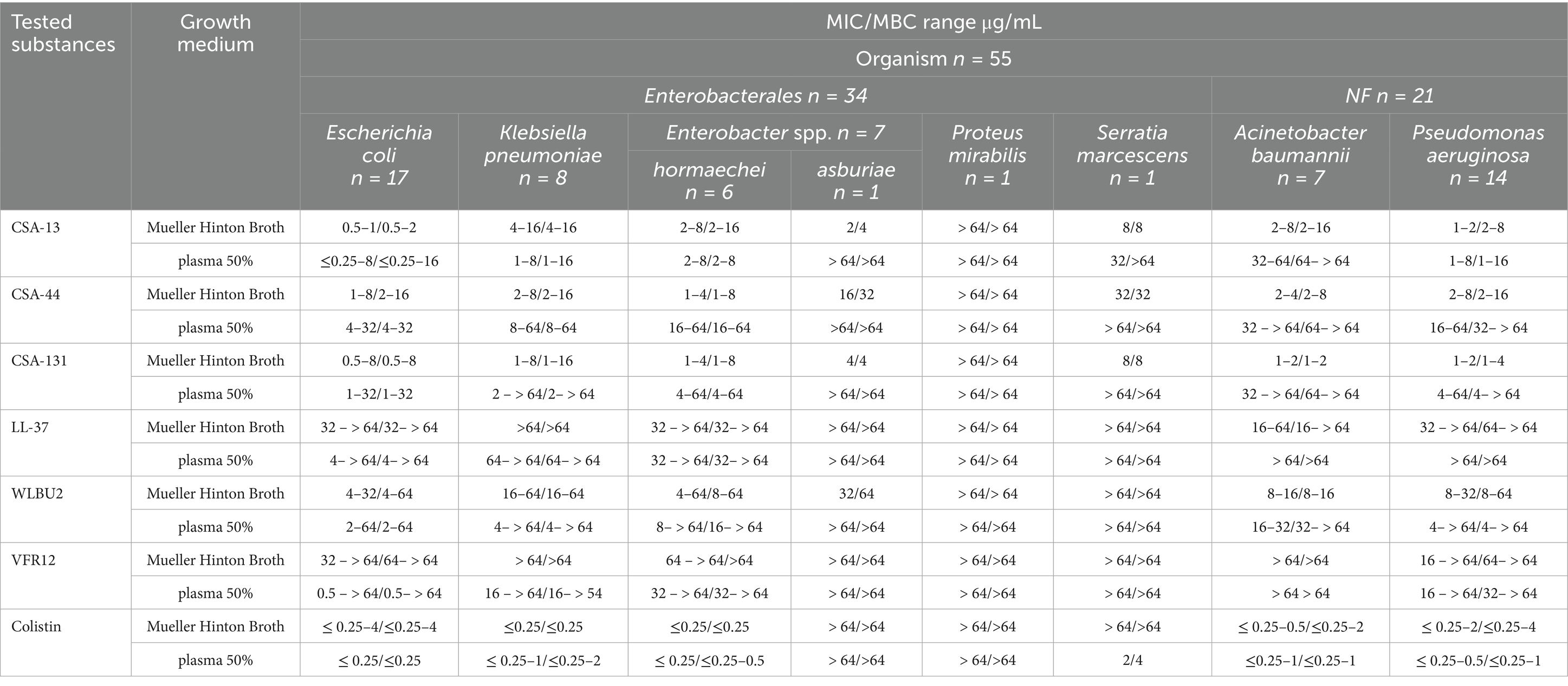
Table 1. MIC/MBC range for CSA-13, CSA-44, CSA-131, LL-37, WLBU2, VFR12 and colistin in the presence of Mueller Hinton Broth and 50% human blood plasma for the clinical G (−) bacteria strain.
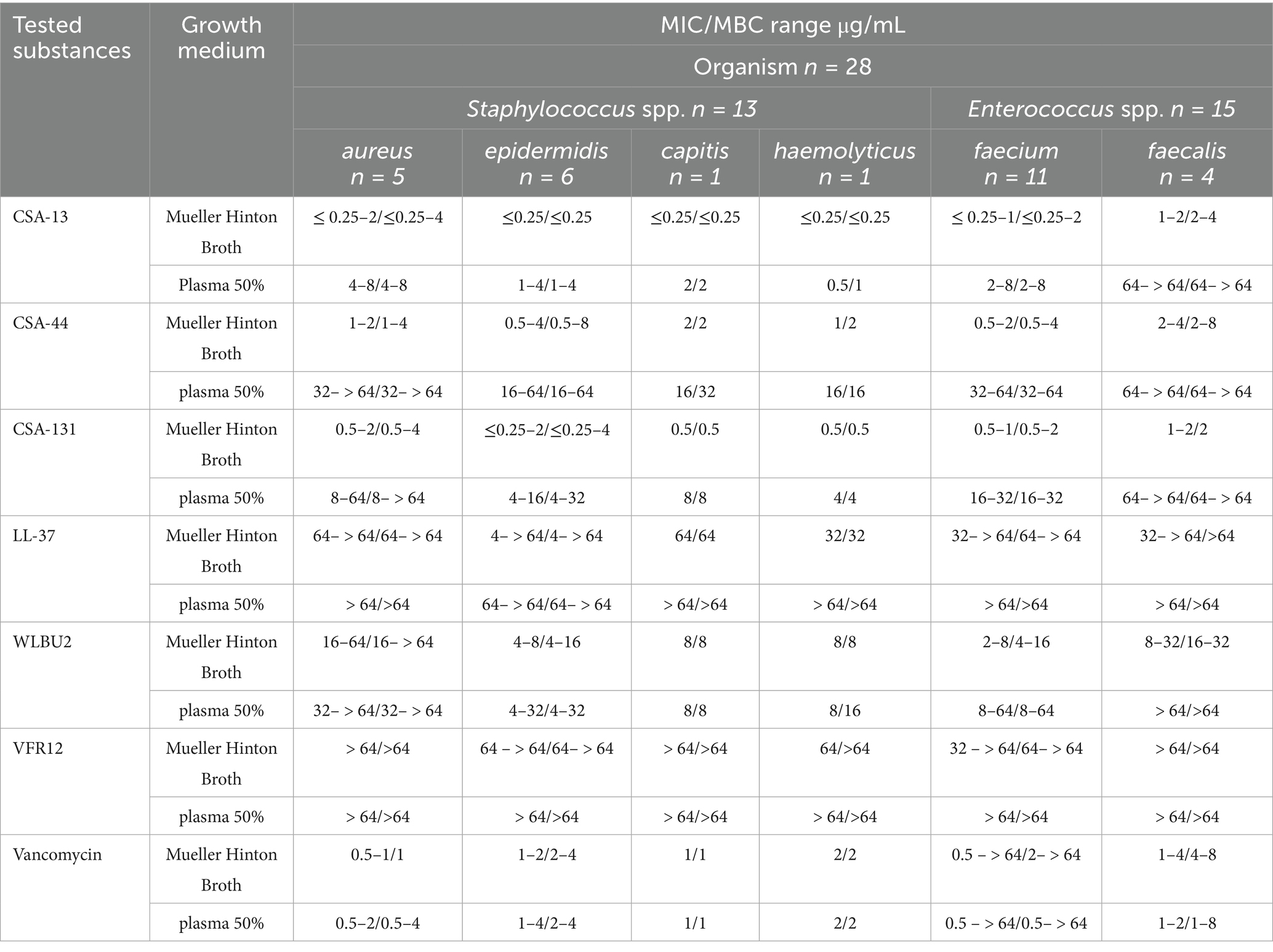
Table 2. MIC/MBC range for CSA-13, CSA-44, CSA-131, LL-37, WLBU2, VFR12 and vancomycin in the presence of Mueller Hinton Broth and 50% human blood plasma for the clinical G (+) bacteria strain.
The highest activity against S. aureus, S. epidermidis, S. capitis, and S. haemolyticus in MH was observed with CSA-13, giving a range of MIC values of ≤0.25–2 μg/mL, ≤0.25 μg/mL, ≤0.25 μg/mL, and ≤0.25 μg/mL, respectively. In the presence of 50% plasma, the effect of ceragenins decreased; however, CSA-13 had the lowest MIC values among the tested ceragenins and retained its high antibacterial activity. All tested strains of Staphylococcus spp. were found to be sensitive to vancomycin, regardless of the type of broth used for MIC determinations. E. faecium and E. faecalis were the species for which the lowest MICs using CSA-13 and CSA-131 were observed. CSA-13 showed the highest antimicrobial activity in the presence of plasma against E. faecium; however, all tested ceragenins partially lost their antimicrobial properties against E. faecalis strains in the presence of blood plasma (MIC ranges exceeded > 64 μg/mL). Among the tested peptides against G (+) bacteria, WLBU2 showed the highest antimicrobial activity, at the same time the MIC range for LL-37 and VFR12 exceeded the highest tested concentration. In the case of G (−) bacteria, a similar pattern of activity was observed.
All tested strains of E. faecalis were sensitive to vancomycin; however, among the tested strains of E. faecium, eight strains were vancomycin-resistant enterococci (VRE). MIC values for reference strains from the ATCC collection were within the range of MIC values for clinical strains. MBCs were determined in the study and the values were close to MIC values (data included in Tables 1, 2).
3.2 Hemolysis
The hemolytic activity of the tested agents was determined using a hemoglobin release assay. Our studies using RBCs showed that CSA-44 and CSA-131 display low hemolysis after 1 h of incubation at a dose of 1–10 μg/mL. After 12 h of incubation, the toxicity of CSA-44 and CSA-131 increased with higher doses ≥10 μg/mL. CSA-13 became slightly toxic above 20 μg/mL. After the application of the WLBU2 peptide, slight toxicity was observed at 5 μg/mL, and this toxicity increased with increasing concentrations of the compound, regardless of the incubation time. LL-37, VFR12, colistin, and vancomycin did not cause significant lysis of human RBCs even at high concentrations (50 μg/mL) after both 1 and 12 h incubation (Figure 4).
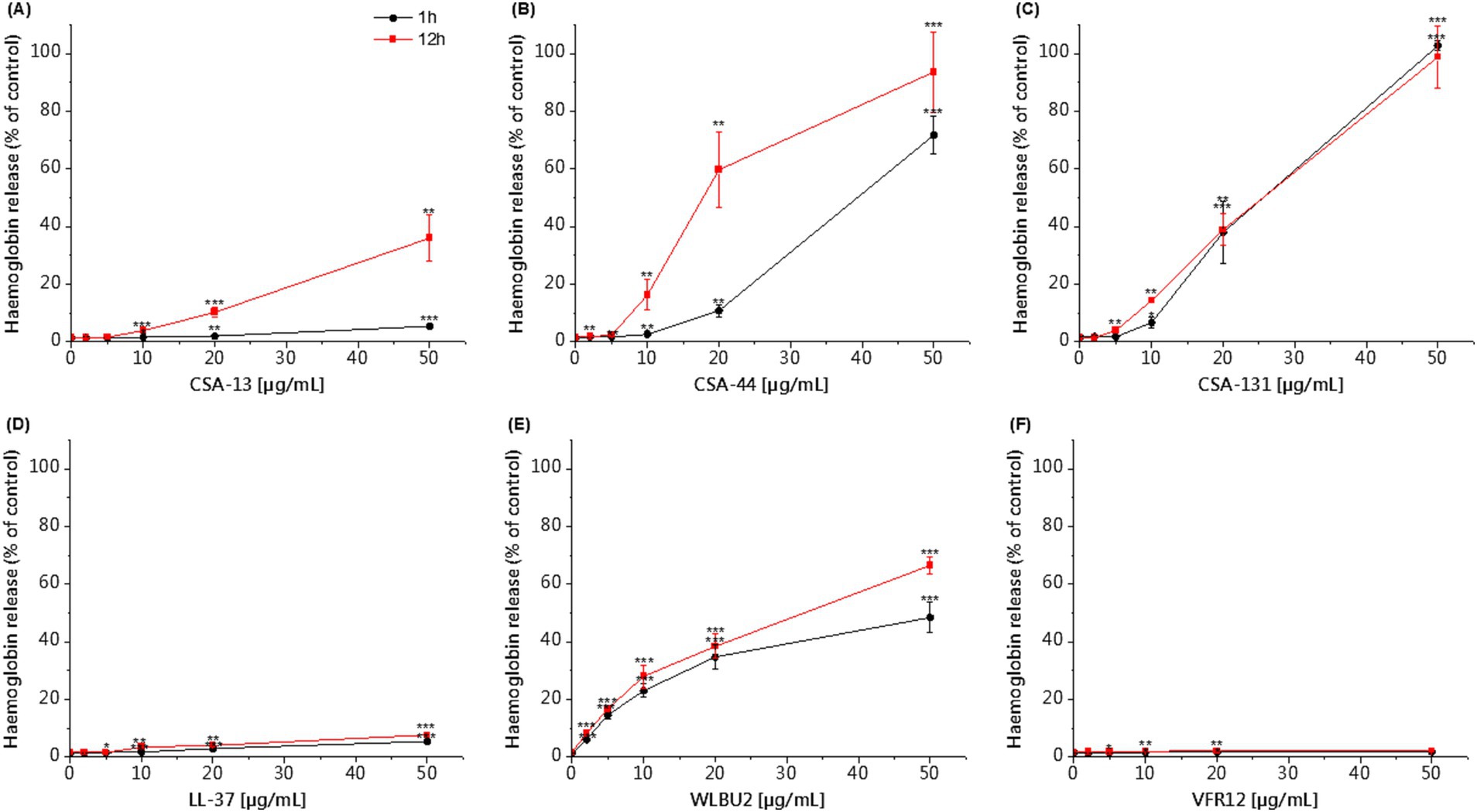
Figure 4. Hemoglobin release from human RBCs after 1 and 12 h of incubation in the presence of CSA-13 (A), CSA-44 (B), and CSA-131 (C), LL-37 (D), WLBU2 (E), VFR12 (F), in the range of 0–50 μg/mL. The results show: mean ± SD, n = 3 (biological replicates); * indicates statistical significance compared to the untreated sample, p ≤ 0.05, ** < 0.02 and *** < 0.001.
CSA-13 showed the strongest antibacterial activity against the tested clinical strains and the highest biocompatibility among the tested compounds. Therefore, it was used to perform additional tests (killing assay, cytokines secretion profile, hemolysis in whole blood and endothelial cell monolayer permeability). From the collection of strains, three clinical isolates were selected for further testing based on their clinical contribution as an important etiological factor of sepsis.
3.3 Antimicrobial and hemolytic activity of CSA-13 in the presence of whole blood
The killing assay in whole blood enabled us to evaluate the activity of CSA-13 against clinical strains under conditions that mimic the pathophysiological environment developed during sepsis. We observed reduced antibacterial activity of CSA-13 against tested clinical isolates in the presence of whole blood compared to experiments performed in PBS. We recorded the highest activity against E. coli. After 3 h, a dose of ≥ 20 μg/mL resulted in growth inhibition. Therefore, we did not perform the determination after 6 and 8 h for this strain. As the dose increased and the incubation time increased, CSA-13 reduced the growth of both P. aeruginosa and E. faecium. The results are presented in Figure 5. The determination of hemolysis in whole blood in the presence of CSA-13 and three clinical isolates (Figure 6), allowed for the assessment of the safety of ceragenin in an experimental setting simulating blood infection. Our data show that the tested ceragenin, under various experimental conditions, induced hemolysis of some red blood cells. However, this effect was observed at doses that were several times higher than the MIC values. At various time points (1, 3, 6 and 8 h), this effect was observed at 50 μg/mL of CSA-13. In the presence of bacteria, no increased levels of hemolysis were observed compared to untreated samples.
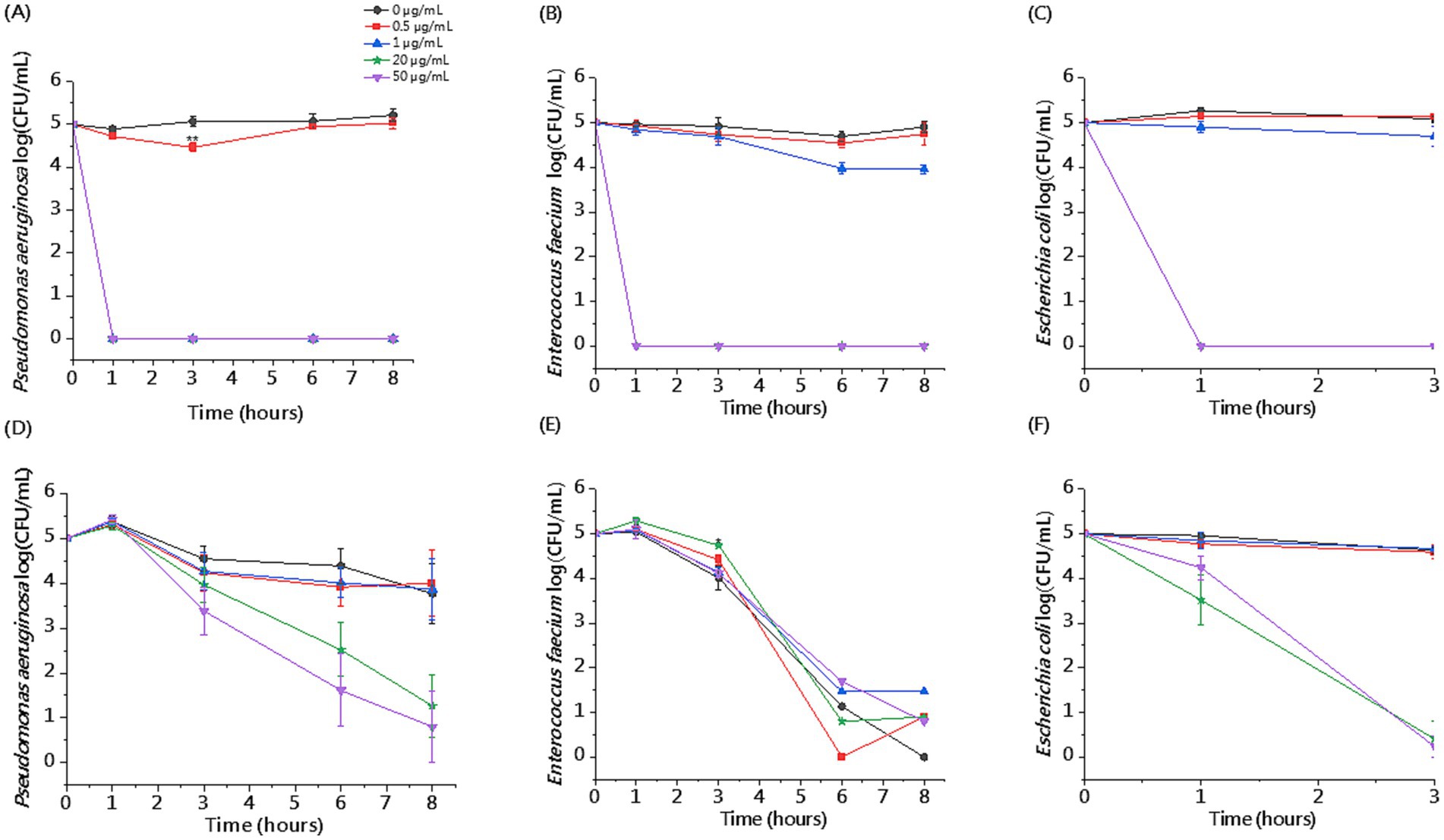
Figure 5. Killing activity of CSA-13 against representative clinical isolates of P. aeruginosa, E. faecium and E. coli at the dose range of 0.5–50 μg/mL in the presence of PBS (A–C) or whole blood (D–F). Results are presented as mean ± SE, n = 3–6 (3 biological replicates/6 technical replicates); * indicates statistical significance compared to an untreated sample, p ≤ 0.05.
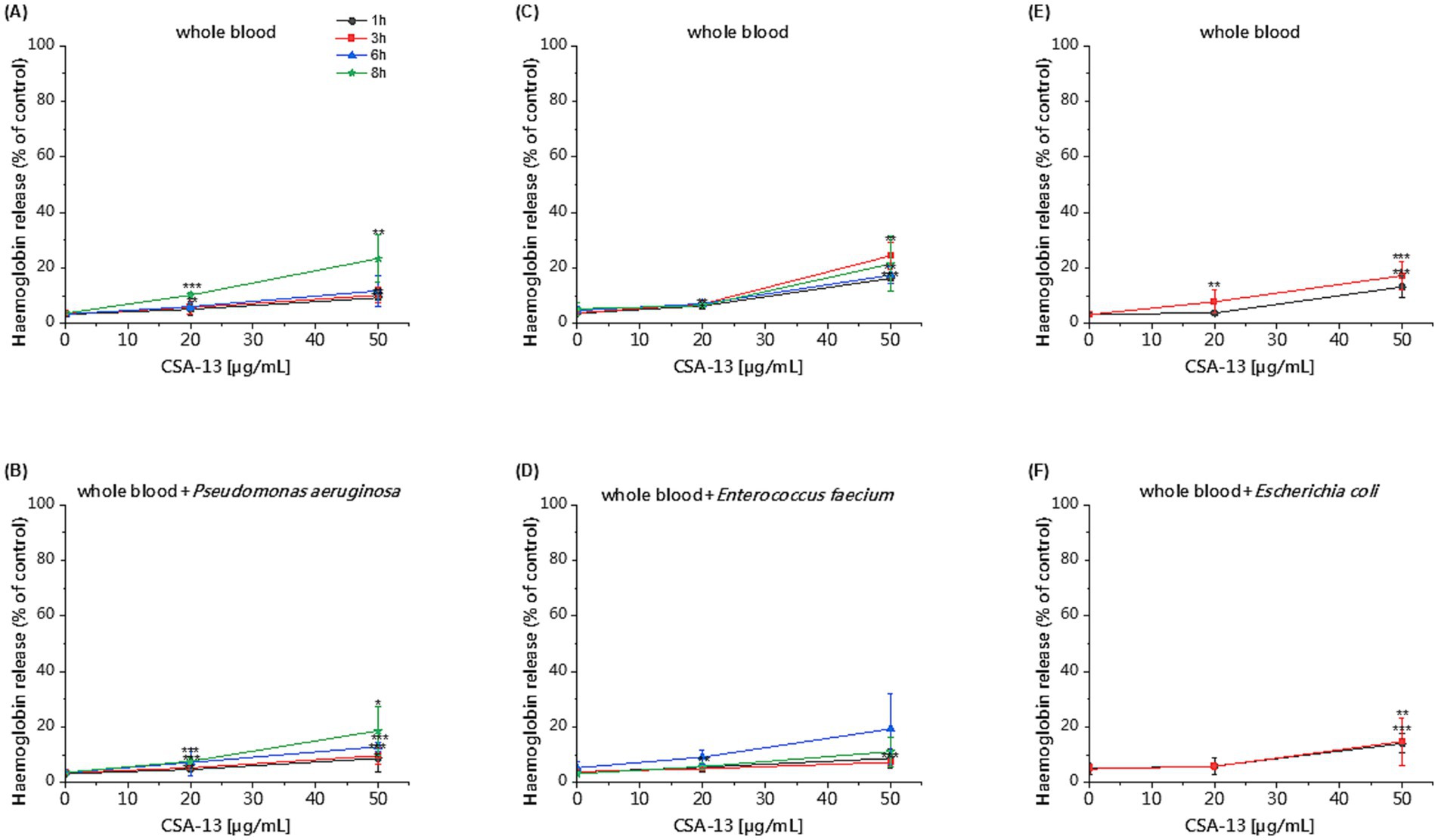
Figure 6. Hemoglobin release after 1, 3, 6 and 8 h of incubation in the presence of CSA-13 at concentrations of 20 and 50 μg/mL in whole blood (A,C,E) and whole blood with P. aeruginosa (B), E. faecium (D) and E. coli (F). The results show: mean ± SD, n = 3–6 (biological replicates); * indicates statistical significance compared to the untreated sample ≤0.05, ** < 0.02 and *** < 0.001.
3.4 Pro-inflammatory response
CSA-13 increases the secretion of proinflammatory cytokines, especially interleukin-8 (IL-8), in the presence of P. aeruginosa after 6 h of incubation (Figure 7).
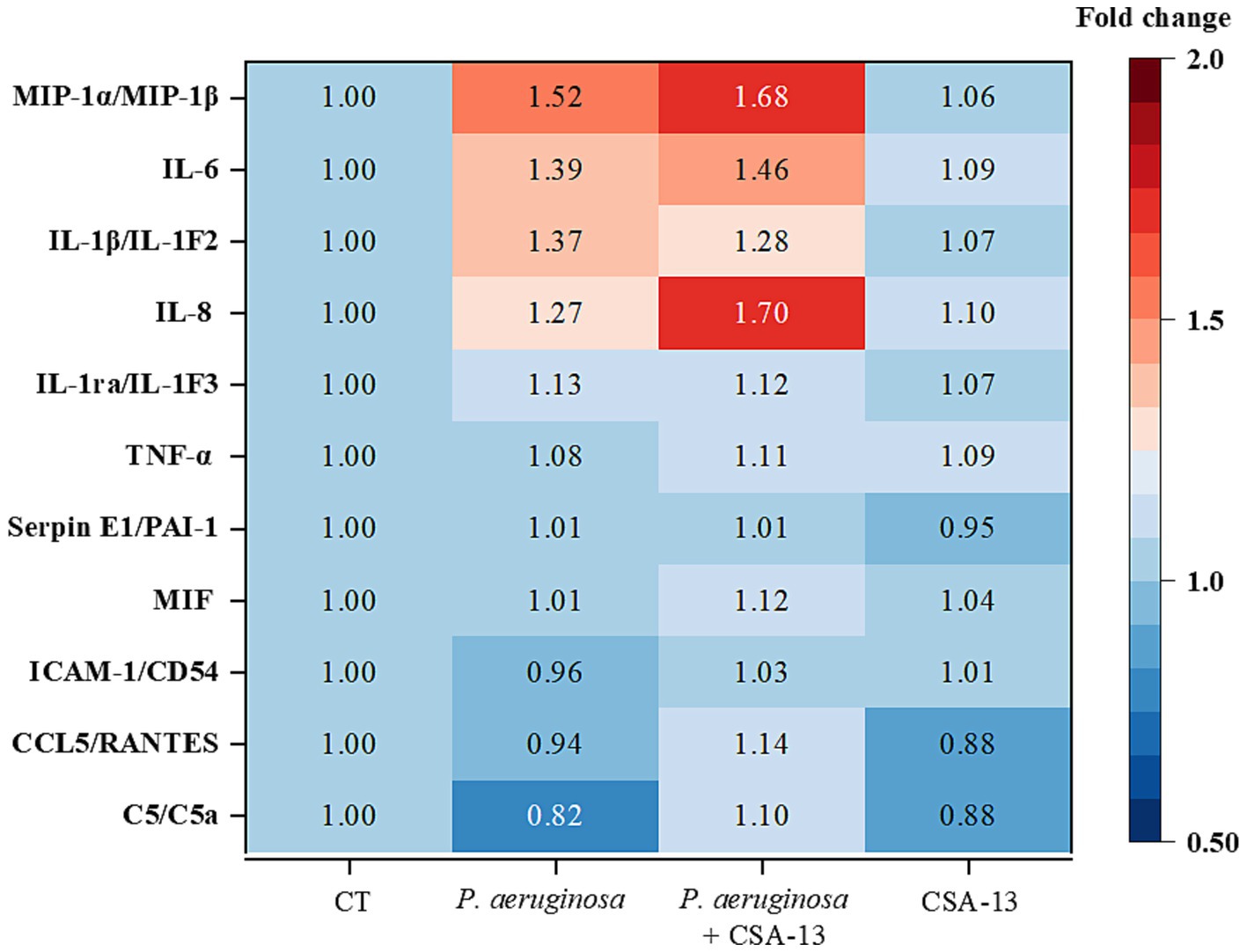
Figure 7. Determination of cytokine levels in plasma after the addition of CSA-13 at a dose of 20 μg/mL (data expressed as fold change compared to the untreated condition, CT). Warmer colors suggest increased cytokine expression, while colder colors suggest decreased expression.
3.5 CSA-13 reduces bacteria-induced HUVEC monolayer permeability
To evaluate the effect of CSA-13 on endothelial cell permeability under various experimental conditions, we performed a dextran permeability assay (Figures 8A,B) and a TEER measurement (Figures 8C,D). Regardless of the time point, heat-inactivated bacteria increased the permeability of HUVEC cells by up to 172% (Enterococcus faecium after 3 h). CSA-13 (1 μg/mL and 5 μg/mL) did not induce permeability and a decrease of up to 10% was observed compared to the untreated sample. A dose of 20 μg/mL after 1 h increased the permeability to 156%, but with increasing incubation time, the permeability decreased to 129%. In the presence of heat-inactivated E. faecium after 1 h, the permeability reached 163%. After adding CSA-13 at doses of 1, 5, and 20 μg/mL simultaneously with bacteria, the permeability decreased to 101, 121, and 115%, respectively. After 3 h, heat-inactivated E. faecium caused an increase to 172%, while 1 μg/mL CSA-13 reduced this effect to 101%, representing a 71% decrease in permeability. After 6 h, heat-inactivated E. faecium induced a 152% increase in permeability, while 1 μg/mL CSA-13 induced a 101% increase, representing a 51% decrease in permeability. After 6 h, heat-inactivated P. aeruginosa increased permeability to over 160% in 1–6 h. Adding CSA-13 at concentrations of 1 and 5 μg/mL after 1 h reduced permeability to 70 and 82%. A 6 h incubation with CSA-13, 1 μg/mL reduced permeability to 89% and a dose of 5 μg/mL to 110%. CSA-13 regulates the dose-dependent increase in HUVECs permeability induced by bacterial LPS or LTA, with a clear beneficial effect observed at a dose of 1 μg/mL. Measurement of TEER confirms the protective role of CSA-13, which increases the strength of the endothelial cell barrier in a dose-dependent manner. Higher resistance indicates reduced permeability, while lower resistance is associated with impaired permeability. Inactivated bacteria significantly reduced resistance at all time points, while the addition of CSA-13 alleviated the reduction in TEER measurements.
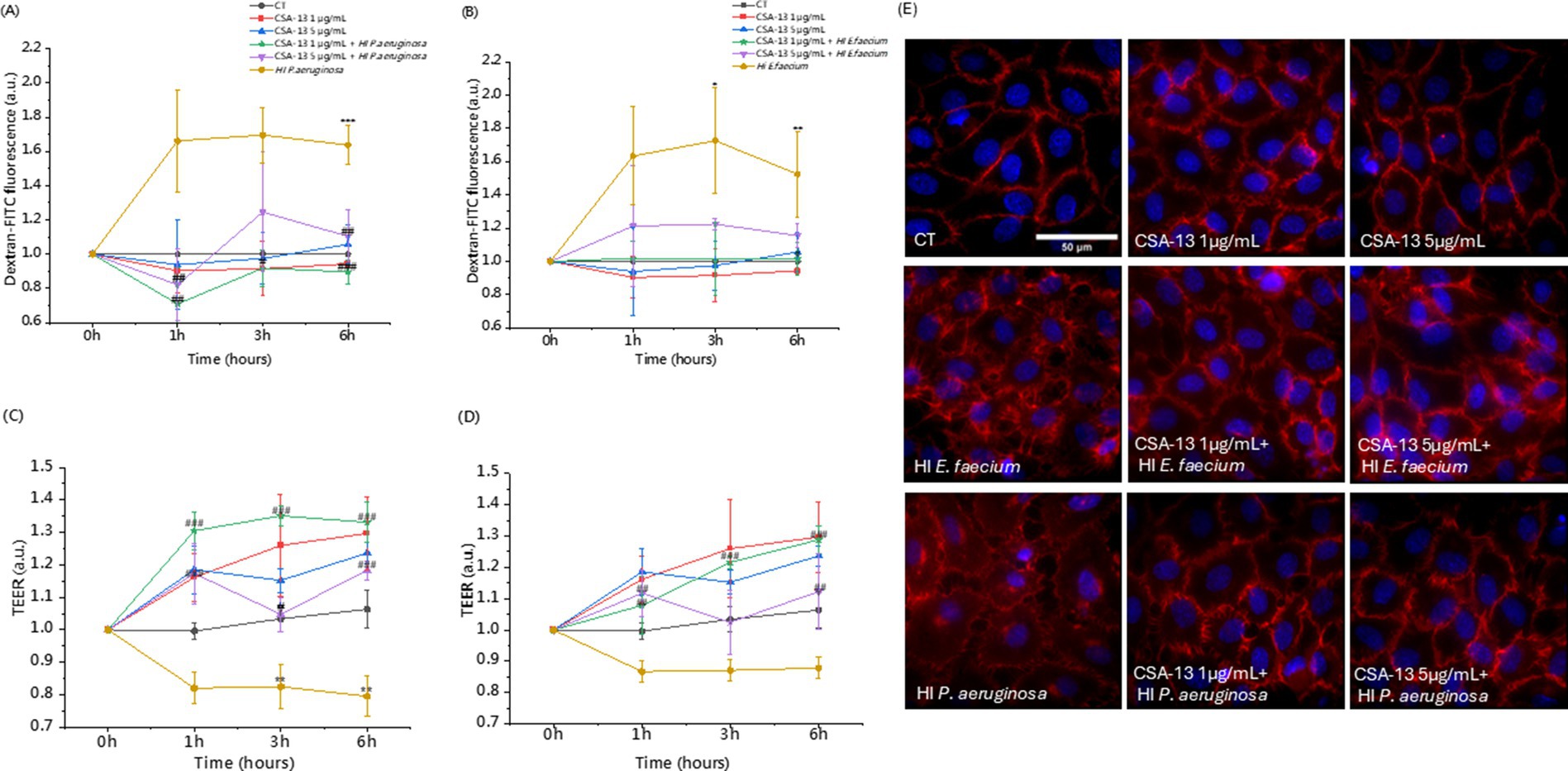
Figure 8. Assessment of the permeability of HUVEC monolayers without and with heat-inactivated bacteria in the presence of CSA-13. Measurements were performed using a fluorometric method based on dextran-FITC permeability (A,B) and transendothelial electrical resistance (TEER) (C,D). The results represent the mean ± SD of three measurements (biological replicates), * and # indicate statistical significance at p ≤ 0.05, ** /## < 0.02 and *** /### < 0.001 compared to CT and heat-inactivated P. aeruginosa (A,C)/heat-inactivated E. faecium (B,D). VE-cadherin (red) rearrangement in response to CSA-13 with the addition of heat-inactivated E. faecium or P. aeruginosa (E), nuclei are in blue.
We also evaluated the morphological changes occurring in the monolayer shown in Figure 8E. CSA-13 at doses of 1 and 5 μg/mL in the presence of heat-inactivated E. faecium or P. aeruginosa does not change the morphology of VE-cadherin intercellular junctions compared to the untreated sample (CT). Delocalization of VE-cadherin from the cell membrane to the cell interior leads to increased permeability resulting from gaps formation between endothelial cells (Lee and Slutsky, 2010). In the presence of inactivated bacteria, delocalization of VE-cadherin was observed. The addition of CSA-13 reduced this effect, which indicates “sealing” of the HUVEC cell monolayer.
4 Discussion
Sepsis is caused by a selected population of microorganisms and is accompanied by a complex pathophysiological mechanism (Jarczak et al., 2021). It is a serious clinical problem associated with high mortality, especially among patients hospitalized in intensive care units and with weakened immunity (Sakr et al., 2018). The elimination of pathogens causing sepsis in the hospital environment is often hampered by their emerging resistance to most known classes of antibiotics (Sharma et al., 2024). To improve the outcomes of sepsis treatment, it is necessary to develop new antibacterial drugs with pleiotropic effects (Marques et al., 2023; Vincent, 2022).
Ceragenins have a broad spectrum of antimicrobial activity that includes bacteria, fungi and some viruses (Prasad et al., 2021). Overall, our results are largely consistent with studies previously conducted by Bozkurt-Guzel et al. (2014). Their study assessed, among other things, the activity of CSA-13 against 60 carbapenem-resistant A. baumannii strains isolated from the blood of patients with bacteremia. The MIC value for all tested isolates was in the range of 1–16 mg/mL (Bozkurt-Guzel et al., 2014). Another study confirmed the antimicrobial activities of: CSA-8, CSA-13, CSA-44, CSA-90, CSA-131, CSA-138, CSA-142, CSA-144 against MDR Gram (−) bacteria isolated from blood, including A. baumannii, K. pneumoniae, E. coli and P. aeruginosa (Yilmaz et al., 2023). Our research confirms the high activity of the tested ceragenins CSA-13, CSA-44 and CSA-131 against G (−) and G (+) bacteria. Comparable to colistin and vancomycin, ceragenins showed higher antimicrobial activity against the tested strains compared to the antimicrobial peptides LL-37, WLBU2, and VFR12. In the study presented here, among the collected clinical isolates, the most numerous were E. coli n = 17, P. aeruginosa n = 14 and E. faecium n = 11. Increasing resistance to colistin, called the “drug of last resort,” has been observed in A. baumannii (Qadri et al., 2025), E. coli (El-Mokhtar et al., 2021), P. aeruginosa (Narimisa et al., 2024), K. pneumoniae (Uzairue et al., 2022), as well as the nephrotoxic effect of colistin (Ordooei Javan et al., 2015), or the emergence of strains resistant to vancomycin (Iqbal et al., 2024), constitutes a serious public health problem and the need to search for new medicinal compounds. In this context, it is worth emphasizing our recent animal studies, which showed high activity of ceragenins against E. coli causing cystitis in mice, which did not show a toxic effect of ceragenins as a result of histopathological examination of the kidneys in the control group (Wnorowska et al., 2022).
An additional advantage of using ceragenins is their chemical nature, which makes them resistant to the action of proteolytic enzymes, which distinguishes them from natural and synthetic antimicrobial peptides (Czarnowski et al., 2024). Indeed, AMPs such as human cathelicidin (LL-37) may lose their antimicrobial activity when exposed to various proteolytical enzymes in plasma (Yang et al., 2024). However, according to previous reports, plasma might weaken the antibacterial effect of CSA-13 (Leszczyńska et al., 2011) and some antibacterial peptides due to their potential interaction with lipoproteins (Ciornei et al., 2005). In our study we observed high stability of CSA-13 in human plasma compared to CSA-44 and CSA-131 when tested against E. coli isolates, where the MIC was in the range of 0.25–8 μg/mL. This range of activity against E. coli is in good agreement with previous studies in which uropathogenic E. coli bacteria show sensitivity to ceragenin in the range of 1–8 μg/mL (Wnorowska et al., 2022). On this basis, it can be concluded that CSA-13 may be particularly important in the treatment of infections caused by E. coli.
In whole blood, the antimicrobial activity of CSA-13 is lower compared to PBS, and a higher dose and longer incubation time are required to observe its effect. We noticed particular bactericidal effectiveness when using CSA-13 at a concentration of ≥ 20 μg/mL against E. coli after 3 h of incubation. Blood reduces the pharmacological activity of some amphipathic drugs due to their interaction with blood cells membranes and/or blood plasma components. These findings may open a new avenue of research to understand the pharmacokinetics of ceragenin. Despite these limitations, it is worth noting that CSA-13 at this dose is non-toxic to human erythrocytes and maintains its antimicrobial properties. A limitation of our study was the small group of subjects. In the future, the group of blood donors for whom blood counts will be performed should be increased, as individual differences may affect the obtained results. Individual differences in blood counts impact the risk of infection, the course of disease, and response to treatment. Our study should be considered as a preliminary study evaluating the effects of ceragenin in the presence of whole blood and its components. Additionally, the future, studies using this experimental model should be carried out on a wider group of microorganisms.
Blood contains various immune components cells and molecules, such as white blood cells (WBCs), platelets (PLTs), complement components, and antimicrobial peptides, which have antimicrobial functions (Taha et al., 2019). In our study, Escherichia coli grows in the control blood sample, which proves that blood itself does not inhibit the growth of this pathogen. With increasing CSA-13 concentration and increasing incubation time in blood, we observe a decrease in CFU/mL of this bacterium, but we assume that the extension of incubation time may correlate with a reduction in the therapeutic dose of CSA-13.
An alternative to increasing the bioavailability of CSA-13 after intravenous administration may be the use of ceragenin-based nanosystems. According to previous studies, in the presence of body fluids: blood plasma, serum, urine, cerebrospinal fluid, abdominal fluid, sputum, dental plaque (Niemirowicz et al., 2017), ear wax (Prasad et al., 2021), ceragenins and ceragenin-based nanosystems demonstrated strong antimicrobial activity.
An interesting direction of research is the combination of ceragenins with conventional antibiotics in order to reduce the therapeutic dose. Recently, the activity of CSA-44 and CSA-131 against blood clinical isolates of G (−) bacteria in combination with classical antibiotics was reported (Yilmaz et al., 2023). This study highlights that CSA-44 and CSA-131, alone or in combination with meropenem or ceftazidime + avibactam, should be considered when developing new alternative treatments for infections caused by antibiotic-resistant bacterial strains. Another study confirmed the effectiveness of the combination of CSA-13 with colistin, tobramycin and ciprofloxacin (Bozkurt-Guzel et al., 2014).
As a result of infection, the host’s reaction is impaired, which is associated with an excessive pro-inflammatory response (Liu et al., 2022). Therefore, we decided to check whether ceragenins can reduce the inflammatory response. In our previous study, 30 μM CSA-13 and LL-37 reduced IL-8 release by 50% in A549 cells infected with P. aeruginosa Xen5 (Bucki et al., 2015). In another study, CSA-13 and CSA-131 effectively reduced the inflammatory response associated with Gardnerella vaginalis infection. Reduced levels of MIP-1α/β, IL-1α and IL-1β IL-10 was observed (Wnorowska et al., 2024). Our results suggest activation of immune system cells upon blood exposure to CSA-13, as evidenced by the increased levels of MIP-1α and MIP-1β in the CSA-13 + P. aeruginosa sample. MIP-1α is an inflammatory protein of macrophages, secreted by cells of the immune system: monocytes, T and B lymphocytes, neutrophils, dendritic cells and NK cells (Bhavsar et al., 2015). In a study by Leszczyńska et al. (2012), CSA-13 was found to induce IL-8 release, which we confirmed here. Proinflammatory cytokine IL-8, has a chemotactic effect on neutrophils and stimulates phagocytosis. Enhanced phagocytosis allows for the restoration of homeostasis which remains disturbed in sepsis (Hortová-Kohoutková et al., 2020). Based on the available experimental data, it can be concluded that CSA-13 has immunomodulatory properties, but further studies are needed to understand its molecular basis. In the future, cytokine levels should be also measured after longer incubation times. It is also worthwhile to test a lower dose of CSA-13.
Endothelial dysfunction, which accompanies sepsis (Joffre et al., 2020), is a crucial determinant of sepsis outcome; therefore, compounds are being sought that will enhance the integrity of this barrier. We used heat-inactivated bacteria to assess endothelial cell permeability. The observed changes are most likely related to immune signaling. In our studies, in addition to its antimicrobial properties, CSA-13 regulates the permeability of endothelial monolayers. We hypothesize that CSA-13 may stabilize the junctions between endothelial cells and V-cadherin and/or modulate inflammatory mediators. Future studies should investigate the mechanism by which CSA regulates endothelial cell permeability. In addition to its antimicrobial properties, CSA-13 regulates the permeability of endothelial monolayers. Another important aspect in the search for new therapeutic agents is the low toxicity of CSAs. The biocompatibility of CSA-13, CSA-44 and CSA-131 has been tested in some previous studies (Paprocka et al., 2022; Skłodowski et al., 2021). Our results confirm low cytotoxicity at bactericidal doses. Toxic doses are several times higher than the MIC values. It is also worth emphasizing that ceragenin-induced host cell damage can be controlled by using Pluronic F-127 (Paprocka et al., 2021), which reduces the toxicity of CSA-13 and does not affect its bactericidal activity (Bucki et al., 2015).
In vivo, CSA-13 demonstrates antibacterial efficacy in an animal model of peritoneal infection (Bucki et al., 2015) in a mouse model of urinary tract infections (Wnorowska et al., 2022) in bacterial vaginosis in mice (Wnorowska et al., 2024). To date, there are no data assessing the effect of ceragenin on an animal model of sepsis.
5 Conclusion
This study demonstrates the potential of ceragenins, especially CSA-13, as possible agents for developing new therapies for patients with bloodstream infections. Low hemolytic activity at bactericidal concentrations, high antibacterial activity maintained in the presence of plasma and protective function for endothelial cells encourage further evaluation of the potential of this substance in the treatment of sepsis, which is associated with infection and accompanied by endothelial dysfunction. In the future, the focus should be on understanding the molecular mechanism associated with the immunomodulatory function of ceragenins as well as assessing the potential of ceragenins in vivo using animal models of sepsis in order to assess the efficacy and safety of these substances from the perspective of potential clinical trials.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
The studies involving humans were approved by Bioethics Committee of the Collegium Medicum of the Jan Kochanowski University in Kielce. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AM: Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing, Conceptualization. PP: Investigation, Methodology, Software, Visualization, Writing – review & editing. ŁS: Investigation, Methodology, Writing – review & editing. AL: Software, Writing – review & editing. KF: Validation, Writing – review & editing. GK: Software, Validation, Visualization, Writing – review & editing. JS: Software, Writing – review & editing. SO: Validation, Writing – review & editing. BD: Conceptualization, Data curation, Investigation, Methodology, Resources, Supervision, Writing – review & editing. TD: Validation, Funding acquisition, Writing – review & editing. EP: Methodology, Project administration, Writing – review & editing. PBS: Resources, Supervision, Writing – review & editing. RB: Conceptualization, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was financially supported by the Medical University of Białystok (B.SUB.23.325 to TD).
Acknowledgments
Some parts of the figures were prepared using Biorender.io software.
Conflict of interest
PBS is a paid consultant for N8 Medical, Inc. Other authors declare no competing interests.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aghapour, Z., Gholizadeh, P., Ganbarov, K., Bialvaei, A. Z., Mahmood, S. S., Tanomand, A., et al. (2019). Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infect. Drug Resist. 12, 965–975. doi: 10.2147/IDR.S199844
Bhavsar, I., Miller, C. S., and Al-Sabbagh, M. (2015). “Macrophage inflammatory Protein-1 alpha (MIP-1 alpha)/CCL3: as a biomarker” in General methods in biomarker research and their applications. eds. V. R. Preedy and V. B. Patel (Dordrecht: Springer Netherlands), 223–249.
Bozkurt-Guzel, C., Hacioglu, M., and Savage, P. B. (2018). Investigation of the in vitro antifungal and antibiofilm activities of ceragenins CSA-8, CSA-13, CSA-44, CSA-131, and CSA-138 against Candida species. Diagn. Microbiol. Infect. Dis. 91, 324–330. doi: 10.1016/j.diagmicrobio.2018.03.014
Bozkurt-Guzel, C., Savage, P. B., Akcali, A., and Ozbek-Celik, B. (2014). Potential synergy activity of the novel ceragenin, CSA-13, against carbapenem-resistant Acinetobacter baumannii strains isolated from bacteremia patients. Biomed. Res. Int. 2014:710273. doi: 10.1155/2014/710273
Bucki, R., Niemirowicz, K., Wnorowska, U., Byfield Fitzroy, J., Piktel, E., Wątek, M., et al. (2015). Bactericidal activity of ceragenin CSA-13 in cell culture and in an animal model of peritoneal infection. Antimicrob. Agents Chemother. 59, 6274–6282. doi: 10.1128/AAC.00653-15
Byfield, F. J., Wen, Q., Leszczynska, K., Kulakowska, A., Namiot, Z., Janmey, P. A., et al. (2011). Cathelicidin LL-37 peptide regulates endothelial cell stiffness and endothelial barrier permeability. Am. J. Physiol. Cell Physiol. 300, C105–C112. doi: 10.1152/ajpcell.00158.2010
Chmielewska, S. J., Skłodowski, K., Piktel, E., Suprewicz, Ł., Fiedoruk, K., Daniluk, T., et al. (2020). NDM-1 Carbapenemase-producing Enterobacteriaceae are highly susceptible to Ceragenins CSA-13, CSA-44, and CSA-131. Infect. Drug Resist. 13, 3277–3294. doi: 10.2147/IDR.S261579
Ciornei, C. D., Sigurdardóttir, T., Schmidtchen, A., and Bodelsson, M. (2005). Antimicrobial and chemoattractant activity, lipopolysaccharide neutralization, cytotoxicity, and inhibition by serum of analogs of human cathelicidin LL-37. Antimicrob. Agents Chemother. 49, 2845–2850. doi: 10.1128/AAC.49.7.2845-2850.2005
Czarnowski, M., Wnorowska, U., Łuckiewicz, M., Dargiewicz, E., Spałek, J., Okła, S., et al. (2024). Natural antimicrobial peptides and their synthetic analogues for effective Oral microflora control and Oral infection treatment—the role of Ceragenins in the development of new therapeutic methods. Pharmaceuticals 17:1725. doi: 10.3390/ph17121725
de Souza, I. L. A., Cappellano, P., Ferreira, D. B., Bergamasco, M. D., das Chagas Neto, T. C., Kerbauy, F. R., et al. (2024). Carbapenem-resistant Klebsiella pneumoniae bloodstream infections in haematological malignances and hematopoietic stem cell transplantation: clinical impact of combination therapy in a 10-year Brazilian cohort. PLoS One 19:e0297161. doi: 10.1371/journal.pone.0297161
Deslouches, B., Islam, K., Craigo, J. K., Paranjape, S. M., Montelaro, R. C., and Mietzner, T. A. (2005). Activity of the de novo engineered antimicrobial peptide WLBU2 against Pseudomonas aeruginosa in human serum and whole blood: implications for systemic applications. Antimicrob. Agents Chemother. 49, 3208–3216. doi: 10.1128/AAC.49.8.3208-3216.2005
Ding, B., Guan, Q., Walsh, J. P., Boswell, J. S., Winter, T. W., Winter, E. S., et al. (2002). Correlation of the antibacterial activities of cationic peptide antibiotics and cationic steroid antibiotics. J. Med. Chem. 45, 663–669. doi: 10.1021/jm0105070
Dolmatova, E. V., Wang, K., Mandavilli, R., and Griendling, K. K. (2021). The effects of sepsis on endothelium and clinical implications. Cardiovasc. Res. 117, 60–73. doi: 10.1093/cvr/cvaa070
El-Mokhtar, M. A., Daef, E., Mohamed Hussein, A. A. R., Hashem, M. K., and Hassan, H. M. (2021). Emergence of nosocomial pneumonia caused by Colistin-resistant Escherichia coli in patients admitted to chest intensive care unit. Antibiotics 10:226. doi: 10.3390/antibiotics10030226
Epand, R. M., Epand, R. F., and Savage, P. B. (2008). Ceragenins (cationic steroid compounds), a novel class of antimicrobial agents. Drug News Perspect. 21, 307–311. doi: 10.1358/dnp.2008.21.6.1246829
EUCAST. Routine and extended internal quality control for MIC determination and disk diffusion as recommended by EUCAST Version 13.2, valid from 2023-06-29. The European Committee on Antimicrobial Susceptibility Testing (EUCAST). (2023).
EUCAST. Reading guide for broth microdilution 2024: Version 5.0. The European Committee on Antimicrobial Susceptibility Testing (EUCAST). (2024).
Gudmundsson, G. H., Agerberth, B., Odeberg, J., Bergman, T., Olsson, B., and Salcedo, R. (1996). The human gene FALL39 and processing of the cathelin precursor to the antibacterial peptide LL-37 in granulocytes. Eur. J. Biochem. 238, 325–332. doi: 10.1111/j.1432-1033.1996.0325z.x
Harris, E. S., and Nelson, W. J. (2010). VE-cadherin: at the front, center, and sides of endothelial cell organization and function. Curr. Opin. Cell Biol. 22, 651–658. doi: 10.1016/j.ceb.2010.07.006
Hashemi, M. M., Rovig, J., Holden, B. S., Taylor, M. F., Weber, S., Wilson, J., et al. (2018). Ceragenins are active against drug-resistant Candida auris clinical isolates in planktonic and biofilm forms. J. Antimicrob. Chemother. 73, 1537–1545. doi: 10.1093/jac/dky085
Hoppens, M. A., Wheeler, Z. E., Qureshi, A. T., Hogan, K., Wright, A., Stanley, G. G., et al. (2014). Maghemite, silver, ceragenin conjugate particles for selective binding and contrast of bacteria. J. Colloid Interface Sci. 413, 167–174. doi: 10.1016/j.jcis.2013.09.016
Hortová-Kohoutková, M., Tidu, F., De Zuani, M., Šrámek, V., Helán, M., and Frič, J. (2020). Phagocytosis-inflammation crosstalk in Sepsis: new avenues for therapeutic intervention. Shock 54, 606–614. doi: 10.1097/SHK.0000000000001541
Iqbal, F., Alocious, A., Joy, S. C., Stanly, E. A. R., Rajesh, V., Unnikrishnan, M. K., et al. (2024). Vancomycin-resistant enterococci: a rising challenge to global health. Clin. Epidemiol. Glob. Health 28:101663. doi: 10.1016/j.cegh.2024.101663
Jarczak, D., Kluge, S., and Nierhaus, A. (2021). Sepsis—pathophysiology and therapeutic concepts. Front. Med. 8:8. doi: 10.3389/fmed.2021.628302
Joffre, J., Hellman, J., Ince, C., and Ait-Oufella, H. (2020). Endothelial responses in Sepsis. Am. J. Respir. Crit. Care Med. 202, 361–370. doi: 10.1164/rccm.201910-1911TR
Karasinski, M., Wnorowska, U., Durnas, B., Krol, G., Daniluk, T., Sklodowski, K., et al. (2023). Ceragenins and Ceragenin-based Core-Shell Nanosystems as new antibacterial agents against gram-negative rods causing nosocomial infections. Pathogens 12:1346. doi: 10.3390/pathogens12111346
Karasiński, M., Wnorowska, U., Daniluk, T., Deptuła, P., Łuckiewicz, M., Paprocka, P., et al. (2024). Investigating the Effectiveness of Ceragenins against Acinetobacter baumannii to Develop New Antimicrobial and Anti-Adhesive Strategies. Int. J. Mol. Sci. 25:7036. Available at: https://www.mdpi.com/1422-0067/25/13/7036
Kasetty, G., Papareddy, P., Kalle, M., Rydengård, V., Mörgelin, M., Albiger, B., et al. (2011). Structure-activity studies and therapeutic potential of host defense peptides of human thrombin. Antimicrob. Agents Chemother. 55, 2880–2890. doi: 10.1128/AAC.01515-10
Lai, X. Z., Feng, Y., Pollard, J., Chin, J. N., Rybak, M. J., Bucki, R., et al. (2008). Ceragenins: cholic acid-based mimics of antimicrobial peptides. Acc. Chem. Res. 41, 1233–1240. doi: 10.1021/ar700270t
Latorre, M. C., Perez-Granda, M. J., Savage, P. B., Alonso, B., Martin-Rabadan, P., Samaniego, R., et al. (2021). Endotracheal tubes coated with a broad-spectrum antibacterial ceragenin reduce bacterial biofilm in an in vitro bench top model. J. Antimicrob. Chemother. 76, 1168–1173. doi: 10.1093/jac/dkab019
Lee, W. L., and Slutsky, A. S. (2010). Sepsis and endothelial permeability. N. Engl. J. Med. 363, 689–691. doi: 10.1056/NEJMcibr1007320
Leszczyńska, K., Namiot, D., Byfield, F. J., Cruz, K., Żendzian-Piotrowska, M., Fein, D. E., et al. (2012). Antibacterial activity of the human host defence peptide LL-37 and selected synthetic cationic lipids against bacteria associated with oral and upper respiratory tract infections. J. Antimicrob. Chemother. 68, 610–618. doi: 10.1093/jac/dks434
Leszczyńska, K., Namiot, A., Cruz, K., Byfield, F. J., Won, E., Mendez, G., et al. (2011). Potential of ceragenin CSA-13 and its mixture with pluronic F-127 as treatment of topical bacterial infections. J. Appl. Microbiol. 110, 229–238. doi: 10.1111/j.1365-2672.2010.04874.x
Liu, D., Huang, S.-Y., Sun, J.-H., Zhang, H.-C., Cai, Q.-L., Gao, C., et al. (2022). Sepsis-induced immunosuppression: mechanisms, diagnosis and current treatment options. Mil. Med. Res. 9:56. doi: 10.1186/s40779-022-00422-y
Marques, A., Torre, C., Pinto, R., Sepodes, B., and Rocha, J. (2023). Treatment advances in Sepsis and septic shock: modulating pro- and anti-inflammatory mechanisms. J. Clin. Med. 12:2892. doi: 10.3390/jcm12082892
Mills, R. J., Boyling, A., Cheng, T. L., Peacock, L., Savage, P. B., Tagil, M., et al. (2020). CSA-90 reduces periprosthetic joint infection in a novel rat model challenged with local and systemic Staphylococcus aureus. J. Orthop. Res. 38, 2065–2073. doi: 10.1002/jor.24618
Nandi, A., Yadav, R., and Singh, A. (2022). Phage derived lytic peptides, a secret weapon against Acinetobacter baumannii-an in silico approach. Front. Med. (Lausanne) 9:1047752. doi: 10.3389/fmed.2022.1047752
Narimisa, N., Keshtkar, A., Dadgar-Zankbar, L., Bostanghadiri, N., Far, Y. R., Shahroodian, S., et al. (2024). Prevalence of colistin resistance in clinical isolates of Pseudomonas aeruginosa: a systematic review and meta-analysis. Front. Microbiol. 15:1477836. doi: 10.3389/fmicb.2024.1477836
Niemirowicz, K., Durnaś, B., Tokajuk, G., Piktel, E., Michalak, G., Gu, X., et al. (2017). Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci. Rep. 7:4610. doi: 10.1038/s41598-017-04653-1
Olekson, M. A., You, T., Savage, P. B., and Leung, K. P. (2017). Antimicrobial ceragenins inhibit biofilms and affect mammalian cell viability and migration in vitro. FEBS Open Bio 7, 953–967. doi: 10.1002/2211-5463.12235
Ordooei Javan, A., Shokouhi, S., and Sahraei, Z. (2015). A review on colistin nephrotoxicity. Eur. J. Clin. Pharmacol. 71, 801–810. doi: 10.1007/s00228-015-1865-4
Papadimitriou-Olivgeris, M., Jacot, D., and Guery, B. (2022). How to manage Pseudomonas aeruginosa infections. Adv. Exp. Med. Biol. 1386, 425–445. doi: 10.1007/978-3-031-08491-1_16
Paprocka, P., Durnaś, B., Mańkowska, A., Skłodowski, K., Król, G., Zakrzewska, M., et al. (2021). New β-lactam antibiotics and Ceragenins - a study to assess their potential in treatment of infections caused by multidrug-resistant strains of Pseudomonas aeruginosa. Infect. Drug Resist. 14, 5681–5698. doi: 10.2147/IDR.S338827
Paprocka, P., Mańkowska, A., Skłodowski, K., Król, G., Wollny, T., Lesiak, A., et al. (2022). Bactericidal activity of Ceragenin in combination with Ceftazidime, levofloxacin, co-Trimoxazole, and Colistin against the opportunistic pathogen Stenotrophomonas maltophilia. Pathogens 11:621. doi: 10.3390/pathogens11060621
Pedrozo-Peñafiel, M., Gutierrez-Beleño, L., Mendoza, C. A. D., Freire-Júnior, F. L., Lima, M. A., Teixeira, T., et al. (2025). In vitro and ex vivo biocompatibility, biomolecular interactions, and characterization of graphene quantum dots and its glutathione-modified variant for qualitative cell imaging. ACS Omega 10, 16194–16206. doi: 10.1021/acsomega.4c10014
Pezzani, M. D., Arieti, F., Rajendran, N. B., Barana, B., Cappelli, E., De Rui, M. E., et al. (2024). Frequency of bloodstream infections caused by six key antibiotic-resistant pathogens for prioritization of research and discovery of new therapies in Europe: a systematic review. Clin. Microbiol. Infect. 30, S4–S13. doi: 10.1016/j.cmi.2023.10.019
Prasad, S. V., Piktel, E., Depciuch, J., Maximenko, A., Suprewicz, Ł., Daniluk, T., et al. (2021). Targeting Bacteria causing otitis media using Nanosystems containing nonspherical gold nanoparticles and Ceragenins. Nanomedicine 16, 2657–2678. doi: 10.2217/nnm-2021-0370
Qadri, M., Tariq, H., Mehmood, M. S., Saddique, M. N., and Sajjad, W. (2025). Mitigating the global spread of multidrug-resistant Acinetobacter buamanni in immunocompromised patients. Discov. Public Health 22:60. doi: 10.1186/s12982-025-00451-7
Sakr, Y., Jaschinski, U., Wittebole, X., Szakmany, T., Lipman, J., Ñamendys-Silva, S. A., et al. (2018). Sepsis in intensive care unit patients: worldwide data from the intensive care over nations audit. Open Forum Infect. Dis. 5:ofy 313. doi: 10.1093/ofid/ofy313
Sharma, S., Chauhan, A., Ranjan, A., Mathkor, D. M., Haque, S., Ramniwas, S., et al. (2024). Emerging challenges in antimicrobial resistance: implications for pathogenic microorganisms, novel antibiotics, and their impact on sustainability. Front. Microbiol. 15:1403168. doi: 10.3389/fmicb.2024.1403168
Skłodowski, K., Chmielewska, S. J., Depciuch, J., Deptuła, P., Piktel, E., Daniluk, T., et al. (2021). Ceragenin-coated non-spherical gold nanoparticles as novel Candidacidal agents. Pharmaceutics 13:1940. doi: 10.3390/pharmaceutics13111940
Spałek, J., Daniluk, T., Godlewski, A., Deptuła, P., Wnorowska, U., Ziembicka, D., et al. (2021). Assessment of Ceragenins in prevention of damage to voice prostheses caused by Candida biofilm formation. Pathogens 10:1371. doi: 10.3390/pathogens10111371
Suprewicz, L., Szczepanski, A., Lenart, M., Piktel, E., Fiedoruk, K., Barreto-Duran, E., et al. (2023). Ceragenins exhibit antiviral activity against SARS-CoV-2 by increasing the expression and release of type I interferons upon activation of the host's immune response. Antivir. Res. 217:105676. doi: 10.1016/j.antiviral.2023.105676
Suprewicz, Ł., Tran, K. A., Piktel, E., Fiedoruk, K., Janmey, P. A., Galie, P. A., et al. (2022). Recombinant human plasma gelsolin reverses increased permeability of the blood–brain barrier induced by the spike protein of the SARS-CoV-2 virus. J. Neuroinflammation 19:282. doi: 10.1186/s12974-022-02642-4
Taha, M., Kyluik-Price, D., Kumaran, D., Scott, M. D., Toyofuku, W., and Ramirez-Arcos, S. (2019). Bacterial survival in whole blood depends on plasma sensitivity and resistance to neutrophil killing. Transfusion 59, 3674–3682. doi: 10.1111/trf.15550
Uzairue, L. I., Rabaan, A. A., Adewumi, F. A., Okolie, O. J., Folorunso, J. B., Bakhrebah, M. A., et al. (2022). Global prevalence of Colistin resistance in Klebsiella pneumoniae from bloodstream infection: a systematic review and Meta-analysis. Pathogens 11:1092. doi: 10.3390/pathogens11101092
Vaara, M. (2009). New approaches in peptide antibiotics. Curr. Opin. Pharmacol. 9, 571–576. doi: 10.1016/j.coph.2009.08.002
Vincent, J. L. (2022). Current sepsis therapeutics. EBioMedicine 86:104318. doi: 10.1016/j.ebiom.2022.104318
Wang, Y., Agerberth, B., Lothgren, A., Almstedt, A., and Johansson, J. (1998). Apolipoprotein A-I binds and inhibits the human antibacterial/cytotoxic peptide LL-37. J. Biol. Chem. 273, 33115–33118. doi: 10.1074/jbc.273.50.33115
Wang, Y., Johansson, J., Agerberth, B., Jornvall, H., and Griffiths, W. J. (2004). The antimicrobial peptide LL-37 binds to the human plasma protein apolipoprotein A-I. Rapid Commun. Mass Spectrom. 18, 588–589. doi: 10.1002/rcm.1361
Wang, T., Liu, H., Huang, H., Weng, Y., and Wang, X. (2024). Colistin monotherapy or combination for the treatment of bloodstream infection caused by Klebsiella pneumoniae: a systematic review and meta-analysis. BMC Infect. Dis. 24:161. doi: 10.1186/s12879-024-09024-6
Wnorowska, U., Fiedoruk, K., Piktel, E., Prasad, S. V., Sulik, M., Janion, M., et al. (2020). Nanoantibiotics containing membrane-active human cathelicidin LL-37 or synthetic ceragenins attached to the surface of magnetic nanoparticles as novel and innovative therapeutic tools: current status and potential future applications. J. Nanobiotechnology 18:3. doi: 10.1186/s12951-019-0566-z
Wnorowska, U., Lysik, D., Piktel, E., Zakrzewska, M., Okla, S., Lesiak, A., et al. (2024). Ceragenin-mediated disruption of Pseudomonas aeruginosa biofilms. PLoS One 19:e0298112. doi: 10.1371/journal.pone.0298112
Wnorowska, U., Piktel, E., Daniluk, T., Paprocka, P., Savage, P. B., Durnaś, B., et al. (2024). Ceragenins prevent the development of murine vaginal infection caused by Gardnerella vaginalis. Pharmaceuticals 17:1445. doi: 10.3390/ph17111445
Wnorowska, U., Piktel, E., Deptuła, P., Wollny, T., Król, G., Głuszek, K., et al. (2022). Ceragenin CSA-13 displays high antibacterial efficiency in a mouse model of urinary tract infection. Sci. Rep. 12:19164. doi: 10.1038/s41598-022-23281-y
Yang, S., Liu, F., Leng, Y., Zhang, M., Zhang, L., Wang, X., et al. (2024). Development of Xanthoangelol-derived compounds with membrane-disrupting effects against gram-positive Bacteria. Antibiotics 13:744. doi: 10.3390/antibiotics13080744
Yang, Z. R., Qin, H., Fan, J. W., Du, K., Qi, L., Hou, D., et al. (2024). Acidity-activated aggregation and accumulation of self-complementary zwitterionic peptide-decorated gold nanoparticles for photothermal biofilm eradication. J. Colloid Interface Sci. 663, 1074–1086. doi: 10.1016/j.jcis.2024.02.018
Yilmaz, F. N., Öksüz, L., Demir, E. S., Döşler, S., Savage, P. B., and Güzel, Ç. B. (2023). Efficacy of Ceragenins alone and in combinations with antibiotics against multidrug-resistant gram negative pathogens from bloodstream infections. Curr. Microbiol. 80:327. doi: 10.1007/s00284-023-03443-5
Keywords: blood infections, antimicrobial peptides, ceragenins, cytokines, permeability, endothelial cell
Citation: Mańkowska A, Paprocka P, Suprewicz Ł, Lesiak A, Fortunka K, Król G, Spałek J, Okła S, Durnaś B, Daniluk T, Piktel E, Savage PB and Bucki R (2025) Bactericidal activity of ceragenin CSA-13, CSA-44 and CSA-131 against bacteria causing bloodstream infections. Front. Microbiol. 16:1640392. doi: 10.3389/fmicb.2025.1640392
Edited by:
Octavio Luiz Franco, Catholic University of Brasilia (UCB), BrazilReviewed by:
Nelson Gomes de Oliveira, University of Brasilia, BrazilCristiane dos Santos, Universidade Federal de Goiás, Brazil
Danieli Fernanda Buccini, Dom Bosco Catholic University, Brazil
Cagka Bozkurt Guzel, Istanbul University, Türkiye
Copyright © 2025 Mańkowska, Paprocka, Suprewicz, Lesiak, Fortunka, Król, Spałek, Okła, Durnaś, Daniluk, Piktel, Savage and Bucki. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Robert Bucki, YnVja2lyb2JlcnRAZ21haWwuY29t