 Myriam Kratou1*
Myriam Kratou1* Hanène Belkahia1
Hanène Belkahia1 Rachid Selmi1,2
Rachid Selmi1,2 Meriam Ben Abdallah1
Meriam Ben Abdallah1 Ghassan Tayh1
Ghassan Tayh1 Lilia Messadi1
Lilia Messadi1 Mourad Ben Said1,3*
Mourad Ben Said1,3*- 1Laboratory of Microbiology, National School of Veterinary Medicine of Sidi Thabet, University of Manouba, Manouba, Tunisia
- 2Ministry of National Defense, General Directorate of Military Health, Veterinary Service, Tunis, Tunisia
- 3Department of Basic Sciences, Higher Institute of Biotechnology of Sidi Thabet, University of Manouba, Manouba, Tunisia
Introduction: Ticks and their associated spotted fever group Rickettsia (SFGR) represent an emerging zoonotic risk in Tunisia, where data on tick species distribution and pathogen prevalence remain limited. This study specifically aimed to investigate the diversity and phylogeny of Ixodes and Rhipicephalus tick species and to identify and genetically characterize their associated SFGR species in northwestern Tunisia.
Methods: Tick sampling was conducted over a five-month period, from November 2022 to March 2023, in the Jouza district, Beja Governorate, northern Tunisia. A total of 236 ticks were collected both from vegetation using flag-dragging and manually from a red fox (Vulpes vulpes) carcass encountered opportunistically in the field. Tick species were morphologically identified and confirmed by Sanger sequencing of the mitochondrial 16S rRNA gene. Rickettsia detection was performed by nested PCR targeting the ompB gene, followed by species-level identification through sequencing of ompA and gltA partial sequences. Phylogenetic analyses were conducted to assess genetic relationships. Additionally, chi-square tests were used to assess differences in infection rates between tick species, life stages, and collection sources.
Results: Ticks belonged to the Ixodes ricinus complex (Ix. ricinus and Ix. inopinatus), Ix. hexagonus, and the Rhipicephalus sanguineus sensu lato complex (Rh. sanguineus sensu strict and Rh. rutilus) have been identified. Twelve and ten genotypes were recorded from 45 and 59 partial 16S rRNA mitochondrial sequences isolated from Ix. ricinus and Ix. inopinatus, respectively. Additionally, one genotype was recorded from five Rh. rutilus specimens, and four genotypes were detected among 23 Rh. sanguineus (temperate lineage) individuals. Overall, 52.1% of ticks tested positive for Rickettsia spp., with significantly higher infection rates in Ix. ricinus (75.6%, p < 0.001) and Ix. inopinatus (67.4%, p < 0.01) compared to the Rhipicephalus group (37.2%) and Ix. hexagonus, which tested negative. Three Rickettsia species were identified: R. monacensis and R. helvetica in the Ix. ricinus complex (including one co-infection), and R. massiliae exclusively in Rhipicephalus ticks. Moreover, phylogenetic analysis revealed that our tick isolates and associated Rickettsia spp. from questing ticks and the red fox clustered primarily with those from other North African and Southern European countries, suggesting trans-Mediterranean strain circulation and a potential link to wildlife reservoirs.
Conclusion: These findings enhance our understanding of tick and SFGR diversity in Tunisia and underscores the zoonotic risks from co-circulating Rickettsia species in shared environments. The high infection rates in Ix. ricinus and Ix. inopinatus call for improved national tick surveillance. Public health implications include the need to consider tick-borne rickettsioses in unexplained febrile illness diagnoses. Future studies should assess infection status in hosts, explore seasonal tick dynamics, and evaluate environmental factors affecting Rickettsia transmission.
1 Introduction
Ticks are obligate hematophagous ectoparasites and prominent vectors of a wide range of pathogens affecting human and animal health globally (Nováková and Šmajs, 2019). Environmental changes including climate shifts, land-use transformation, and altered wildlife dynamics have facilitated the geographic expansion of ticks and their associated pathogens, particularly in the Mediterranean basin (Estrada-Peña and de la Fuente, 2014; Martina et al., 2017; Sonenshine, 2018). Among tick families, Ixodidae is the most epidemiologically relevant, owing to its abundance, ecological versatility, and broad host range. These characteristics position it centrally within the One Health framework (Tsatsaris et al., 2016).
Within this context, tick-borne zoonoses, especially vector-borne rickettsioses (VBRs), have gained increasing public health attention (Onyiche et al., 2022). VBRs are caused by obligate intracellular Gram-negative bacteria of the genus Rickettsia, which are primarily transmitted by arthropods, with hard ticks (Acari: Ixodidae) acting as both vectors and reservoirs (Parola et al., 2005, 2013). In addition, over 27 of the 30 recognized Rickettsia species are associated with hard ticks, and the majority belong to the spotted fever group (SFG), including R. rickettsii and R. conorii, the causative agents of Rocky Mountain spotted fever and Mediterranean spotted fever, respectively (Parola et al., 2005). These pathogens exhibit diverse vector-host associations and pathogenic profiles, resulting in variable clinical outcomes (Kernif et al., 2012). In Africa, at least 17 Rickettsia species have been identified across several tick genera, including Hyalomma, Amblyomma, Rhipicephalus, and Ixodes, underscoring the ecological complexity and zoonotic potential of SFG rickettsioses (Parola et al., 2013). These bacteria are primarily maintained in tick populations through vertical transmission (transovarial and transstadial), although horizontal transmission via vertebrate hosts also plays a critical role (Hauck et al., 2020; Laukaitis and Macaluso, 2021). However, some Rickettsia spp. negatively impact tick fitness by reducing fecundity and reproductive success (Nieri-Bastos et al., 2013; Krawczak et al., 2016; Gerardi et al., 2019). In addition, tick distribution is strongly influenced by abiotic factors, particularly temperature and humidity, which in turn affect their pathogen transmission potential (Onyiche et al., 2022).
Despite growing awareness, the epidemiological landscape of SFG Rickettsia in North Africa remains insufficiently documented. In Tunisia, Rickettsia infections have been documented for over a century, beginning with R. conorii, the causative agent of Mediterranean spotted fever, first reported in 1910 (Znazen et al., 2013). Nonetheless, more recent molecular studies have revealed a broader range of circulating species, including R. helvetica, R. africae, R. aeschlimannii, and R. massiliae, primarily detected in Hyalomma and Rhipicephalus ticks from domestic animals in central, southern and northern Tunisia (Demoncheaux et al., 2012; Khrouf et al., 2013; Selmi et al., 2019; Belkahia et al., 2021; Kratou et al., 2023). In this context, attention is increasingly turning to the role of wildlife hosts in pathogen maintenance and transmission. Case in point, red foxes (Vulpes vulpes) are becoming increasingly recognized as important hosts in the ecology of tick-borne pathogens, due to their wide distribution, synanthropic behavior, and frequent infestation by diverse tick species (Hornok et al., 2017; Chisu et al., 2014). As potential bridge hosts between wildlife and peri-domestic environments, they serve as valuable sentinels for monitoring the circulation of zoonotic agents such as Rickettsia (Marié et al., 2011; Liu et al., 2021; Selmi et al., 2025). However, in Tunisia, data on the tick fauna associated with red foxes and their infection status remain scarce, limiting our understanding of their role in pathogen maintenance and transmission.
Furthermore, the northwestern region of Tunisia, particularly the Jouza district in the Beja Governorate, provides a humid, forested habitat that supports diverse tick populations, including the sympatric occurrence of Ixodes ricinus and Ixodes inopinatus, both competent vectors of zoonotic pathogens (Zhioua et al., 1994; Sarih et al., 2003; Younsi et al., 2005; Selmi et al., 2024). In parallel, Rhipicephalus sanguineus sensu lato is the most widespread tick species in the Mediterranean basin and a primary vector of R. conorii, the causative agent of Mediterranean spotted fever (Kim, 2022). Within this complex, Rh. sanguineus sensu stricto and Rh. rutilus are key vectors of several zoonotic pathogens, including R. conorii, R. massiliae, R. felis, Ehrlichia canis, and Coxiella burnetii (Dantas-Torres, 2010; Khrouf et al., 2013; Šlapeta et al., 2023; Senbill et al., 2024).
Despite increasing reports of Rickettsia species in Tunisia, data on the genetic diversity and infection status of Ixodes and Rhipicephalus ticks with spotted fever group (SFG) Rickettsia remain limited. Given the ecological importance of both questing and host-associated ticks in pathogen transmission, further investigation is warranted, particularly regarding wildlife hosts such as the red fox (V. vulpes) as potential reservoirs. This study is an exploratory investigation focused on a single location in a sub-humid zone in northern Tunisia, known for its high infestation primarily by Ixodes ticks. In particular, we aimed to characterize the species composition, genetic diversity, and phylogenetic relationships of Ixodes and Rhipicephalus ticks collected from vegetation and an accidentally found red fox during sampling, and their associated SFG Rickettsia spp. Additionally, infection prevalence was assessed across tick species, life stages, and collection sources. We hypothesized that both questing and host-associated ticks in this region harbor a genetically diverse pool of SFG Rickettsia, with red foxes potentially playing a role in their maintenance and transmission within wildlife-associated cycles.
2 Materials and methods
2.1 Study regions, tick collection and morphological identification
Between November 2022 and March 2023, a total of 236 ticks were collected from a single site in the Jouza district, Beja Governorate, northwestern Tunisia, characterized by a humid and subhumid bioclimatic stage favorable to the presence of Ixodes and Rhipicephalus species (Figure 1). Tick sampling targeted this site specifically due to its known ecological suitability for these genera. Ticks were collected from both vegetation and a red fox (V. vulpes). Vegetation sampling was performed using the standard flag-dragging technique, involving sweeping a white flannel cloth over the vegetation to capture questing ticks. Ticks from the red fox were collected manually. The fox, found dead by chance on a rural road in the Jouza region, was a single adult individual that appeared to have died recently. Remarkably, it was heavily infested with Ixodes ticks, an original and noteworthy finding as investigations on red fox tick infestations in this region have not previously been conducted. Ticks were carefully removed using forceps from preferred attachment sites on the fox's body, including the ears, neck, udder area, and external genitalia. Since only this single carcass was sampled, the risk of resampling the same animal was eliminated. All collected ticks were placed in sterile, labeled tubes and preserved individually in 70% ethanol, then stored at −20 °C until further analysis. Morphological identification was performed using standard taxonomic keys (Walker et al., 2005; Estrada-Peña et al., 2017), allowing classification by species, life stage, and sex.
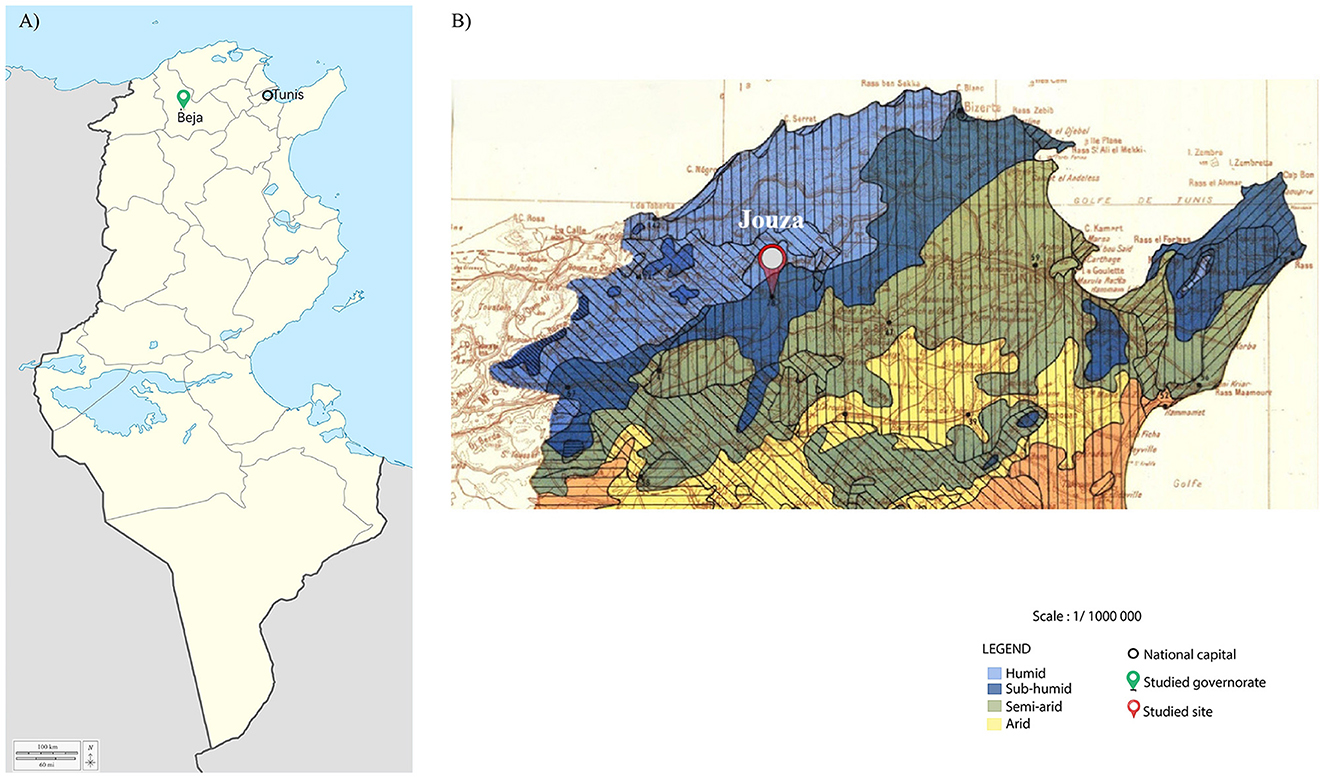
Figure 1. Maps of the study area in Tunisia. (A) Map of Tunisia divided by governorates, showing the location of the Beja governorate marked with a green pin and the national capital, Tunis, indicated by a black circle. (B) Zoomed-in map of northwestern Tunisia based on bioclimatic stages, showing the specific sampling site in the Jouza district marked with a red pin. National borders are outlined in black.
2.2 Extraction of total DNA and amplification of tick DNA
Each tick specimen was washed three times in sterile water with gentle vortexing (10 s each), briefly rinsed in 70% ethanol, air-dried, and then individually homogenized using an automated TissueLyser LT system (Qiagen, Hilden, Germany). Genomic DNA was extracted using the DNeasy Tissue Kit (Qiagen), following the manufacturer's protocol, and stored at −20 °C until further use. To evaluate the efficiency of DNA extraction, PCR amplification of the mitochondrial 16S rRNA gene was performed using tick-specific primers TQ16S+1F and TQ16S-2R, as described by (Black and Piesman 1994) (Table 1). Each PCR run included both positive and negative controls to ensure the reliability and specificity of amplification.
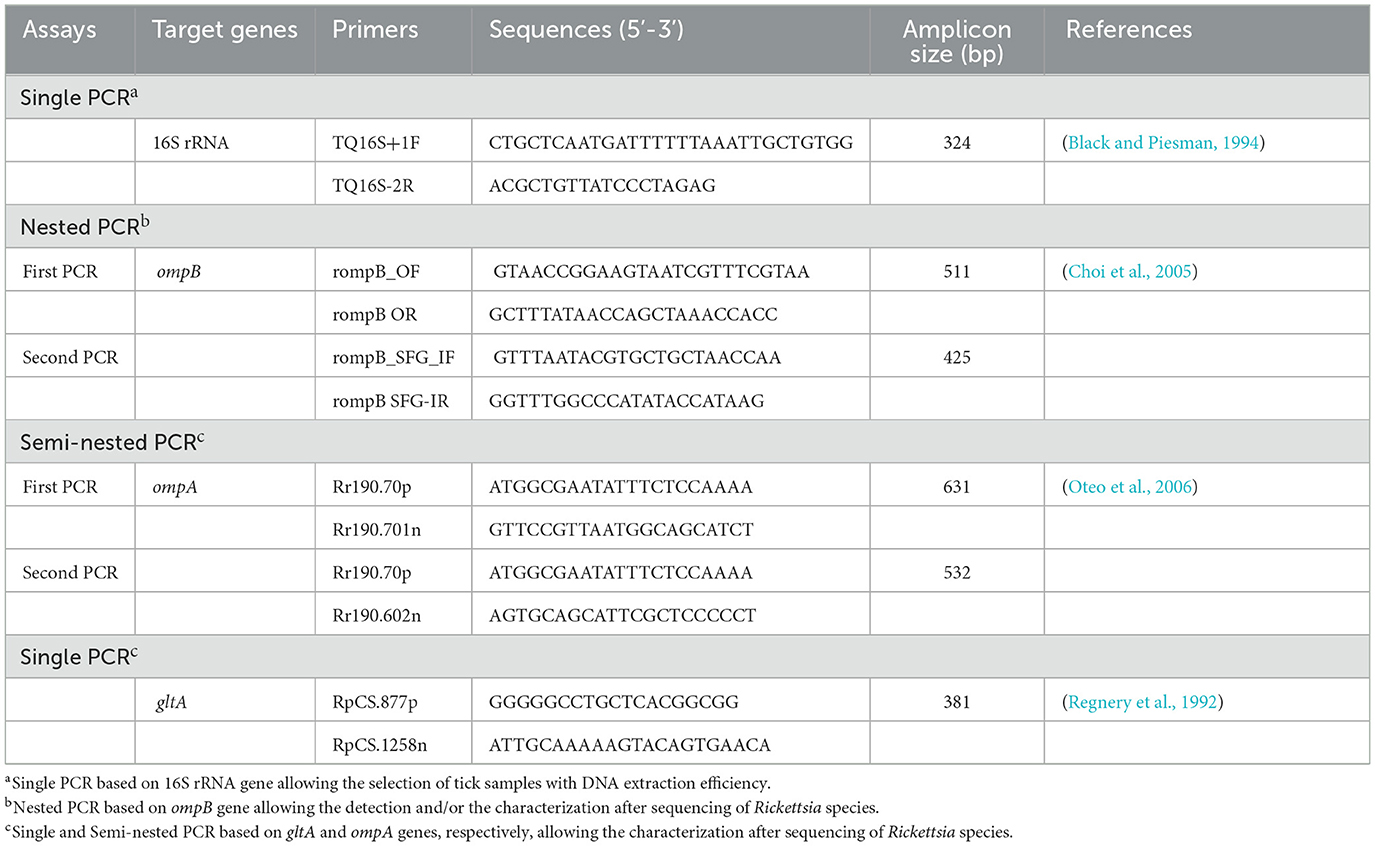
Table 1. Primers used for the identification and/or genetic characterization of Rickettsia species infecting ticks collected in this study.
2.3 Identification of Rickettsia species
Initially, a nested PCR was conducted to amplify a 425 bp fragment of the rickettsial outer membrane protein B (ompB) gene from tick DNA samples, in order to detect Rickettsia species. Subsequently, single PCRs were used for species-level characterization amplifying gene fragments encoding the outer membrane protein A (ompA) and citrate synthase protein (gltA), measuring 532 bp and 381 bp, respectively. All PCR assays were performed using an automated DNA thermal cycler, following the thermal cycling protocols established by (Oteo et al. 2006) and (Regnery et al. 1992). PCR reactions were carried out in a total volume of 50 μL, containing 0.125 U/μL of Taq DNA polymerase (Biobasic Inc., Markham, Canada), 1x PCR buffer, 0.2 mM of dNTPs, 1.5 mM of MgCl2, 3 μL of genomic DNA (50–150 ng) for the first PCR, and 1 μL for the nested PCR. Additionally, 0.5 μM of primers and autoclaved water were included. Each PCR run included both positive and negative controls to ensure the reliability and specificity of amplification. PCR products were subsequently analyzed using electrophoresis on 1.5% agarose gels stained with ethidium bromide, allowing visualization under UV transillumination.
2.4 Statistical analysis
Confidence intervals (CIs) for prevalence rates were calculated at the 95% confidence level. To investigate the association between tick-related factors, including species and gender, on the molecular prevalence of Rickettsia species, either a chi-square test or Fisher's exact test were conducted. These analyses were performed using Epi Info 6.01 (CDC, Atlanta, GA, USA), with a significance threshold set at 0.05.
2.5 DNA sequencing, alignment and phylogenetic analysis
Positive PCR products obtained from the amplification of mitochondrial 16S rRNA, ompB, ompA and gltA partial sequences of Rickettsia spp. were purified using the GF-1 Ambi Clean kit (Vivantis, Oceanside, CA, USA) following the manufacturer's protocol. The resulting purified DNA amplicons were sequenced in both directions, utilizing the same primers employed in the mitochondrial 16S rRNA ompA and gltA single PCRs, as well as the secondary PCR from the nested amplifications for the ompB partial sequences. Sequencing was conducted using the Big Dye Terminator Cycle Sequencing Kit (Applied Biosystems, Foster City, CA, USA) alongside an ABI3730XL automated DNA sequencer (Macrogen Europe, Amsterdam, The Netherlands). Chromatograms were analyzed using Chromas v.2.01. Raw sequences were obtained from both forward and reverse strands to ensure maximum accuracy. Complementary strands of each sequenced product were manually assembled using DNAMAN software v.5.2.2. (Lynnon Biosoft, Que., Canada). Additionally, overlapping regions were identified following the automated removal of primer sequences. The nucleotide sequences from the mitochondrial 16S rRNA marker, as well as the ompB, ompA, and gltA genes of Rickettsia spp., were utilized to calculate genotype diversity (Gd), nucleotide diversity (Pi) and the average number of nucleotide differences (k) using DnaSP v 5.10. Sequence similarities were determined using the CLUSTAL W v.1.81 method after performing multiple sequence alignments. A BLAST analysis was conducted to evaluate the similarity levels with previously reported sequences in GenBank (http://blast.ncbi.nlm.nih.gov/). Genetic distances among the operational taxonomic units were calculated using the maximum composite likelihood method through DNAMAN software, which were then utilized to construct Maximum Likelihood trees (Tamura and Nei, 1993). A bootstrapping process with 1,000 iterations was employed to assess the statistical support for the internal branches of these trees (Felsenstein and Sinauer, 2008).
3 Results
3.1 Morphological and molecular identification of ticks and phylogenetic analysis
3.1.1 Efficiency of DNA isolation and distribution of collected ticks
A total of 236 ticks (160 females, 52 males, and 24 nymphs) were collected from a single site in the Jouza district, Beja, Tunisia (Figure 1), comprising 167 ticks sampled from vegetation and 69 ticks removed from a single red fox found at this location. Genomic DNA was extracted from all specimens, followed by PCR amplification targeting the mitochondrial 16S rRNA, which yielded successful amplification in 100% of the samples, confirming the efficiency of the DNA isolation method. Morphological identification, based on the diagnostic keys of (Walker et al. 2005) and (Estrada-Peña et al. 2017), classified nymph ticks (n = 24) as belonging to the Ixodes genus, and adult ticks into two complexes: Ix. ricinus complex (n = 94) and Rh. sanguineus sensu lato (s.l.) complex (n = 78). Molecular identification through sequencing of all Ixodes PCR products and BLAST analysis of partial 16S rRNA sequences revealed that, within the Ix. ricinus complex, Ix. inopinatus (n = 89) and Ix. ricinus (n = 34) were identified, while all nymphs (n = 24) were classified as Ix. hexagonus (Table 2). Additionally, for the Rh. sanguineus s.l. complex, sequencing of specimens positive for Rickettsia spp. and subsequent BLAST analysis identified Rh. sanguineus (temperate lineage) (n = 24) and Rh. rutilus (n = 5) (Table 2). To prevent and monitor potential contamination, particularly important for nested PCR protocols, all reactions were conducted in separate areas with unidirectional workflow, and negative controls were systematically included in each PCR run (Supplementary Figure S1A).
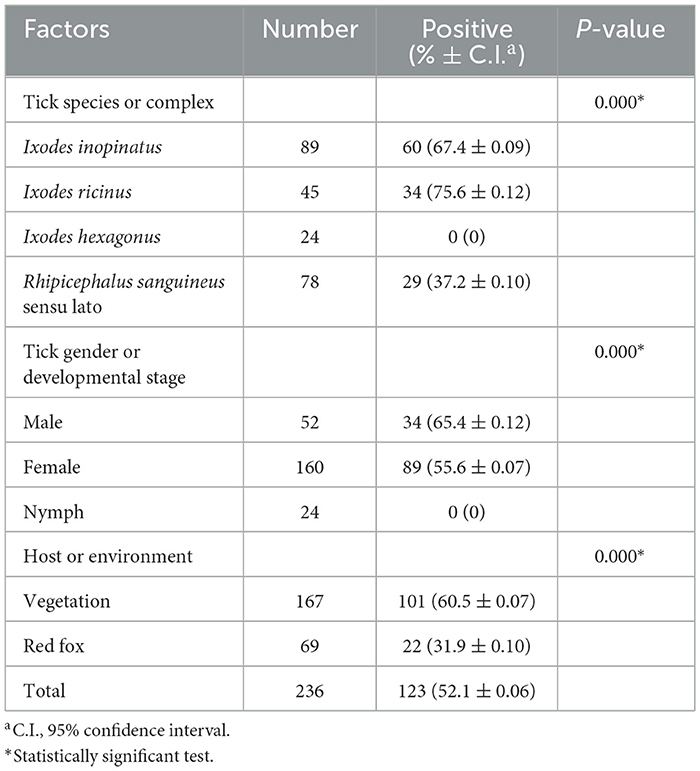
Table 2. Molecular prevalence results of Rickettsia spp. infecting Ixodes spp. and Rhipicephalus sanguineus sensu lato complex ticks according to several risk factors.
3.1.2 Genotyping and phylogenetic analysis of selected tick specimens
Partial sequence analysis of the mitochondrial 16S rRNA revealed genotypic variation among the collected tick specimens, with distinct genotypes differing by at least one nucleotide mutation in the mitochondrial 16S rRNA.
3.1.2.1 Genetic diversity and phylogeny of Ixodes spp. isolates
Ixodes ricinus specimens, collected from both vegetation and V. vulpes in the Jouza district (Beja, Tunisia), exhibited substantial genetic diversity. A total of 12 distinct genotypes were identified (Ixric16SG1–Ixric16SG12) (Table 3 and Supplementary File 1). Genotypic diversity (Gd) was estimated at 0.539, with a GC content of 51.1%. Nucleotide diversity (Pi) was 0.00224, and the mean number of pairwise nucleotide difference (k) was 0.614 (Table 4). Phylogenetic tree revealed that genotypes such as Ixric16SG2 and Ixric16SG6 were detected in vegetation and V. vulpes (Figure 2). Conversely, Ixric16SG1, Ixric16SG4, Ixric16SG7, Ixric16SG9, Ixric16SG10, and Ixric16SG11 were exclusively associated with questing ticks, and Ixric16SG3, Ixric16SG5, Ixric16SG8, Ixric16SG12 to V. vulpes (Figure 2). Notably, Ixric16SG1 and Ixric16SG11 clustered with an Ix. ricinus isolate from Homo sapiens in China (GenBank accession number: OK484994), while Ixric16SG12 was detected solely in V. vulpes, clustering within the principal Ix. ricinus lineage (Figure 2).
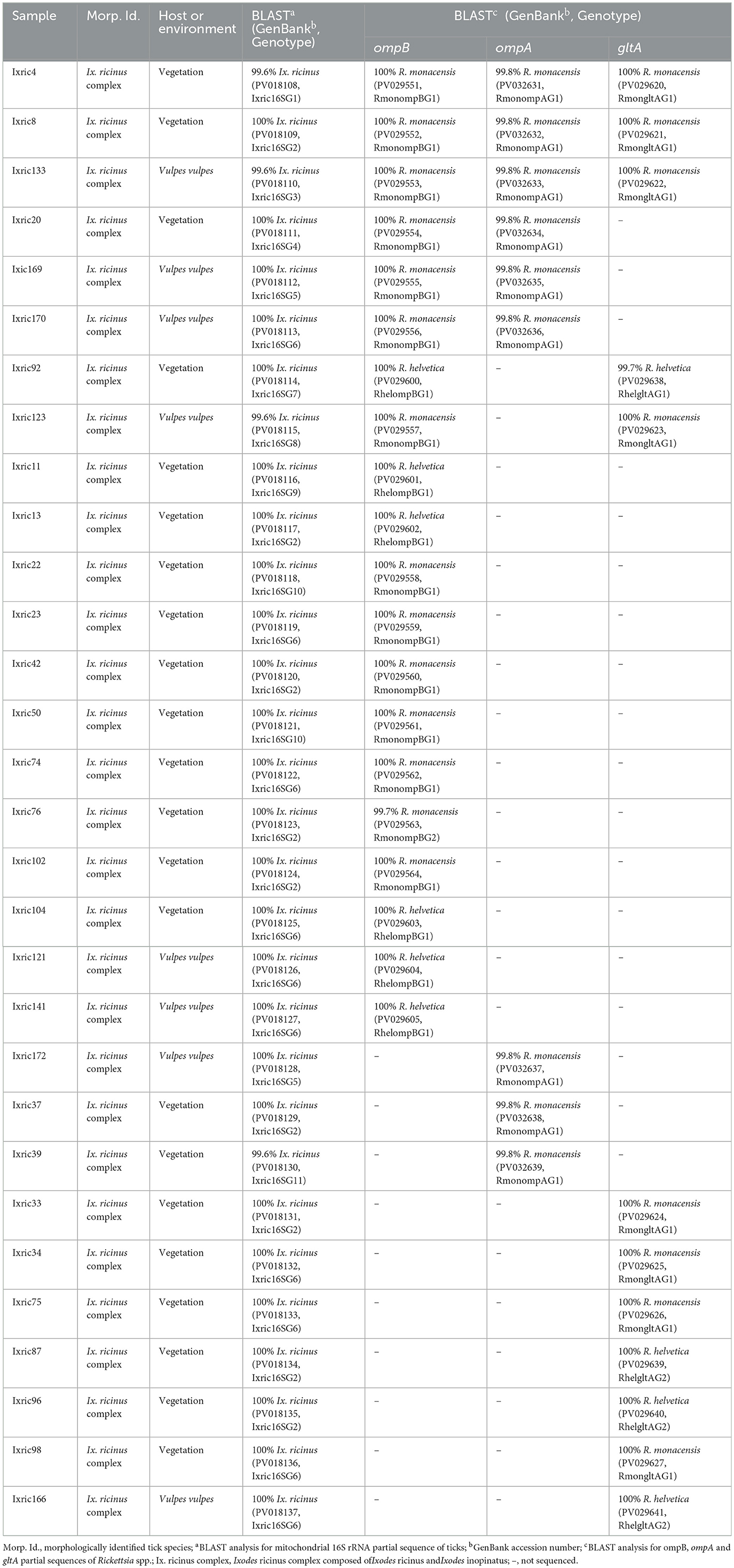
Table 3. Designation and information on the origins and genotypes of Tunisian isolates of Rickettsia spp. isolated from Ix. ricinus ticks collected from vegetation and a red fox (Vulpes vulpes) in Jouza district (Beja governorate, Tunisia).
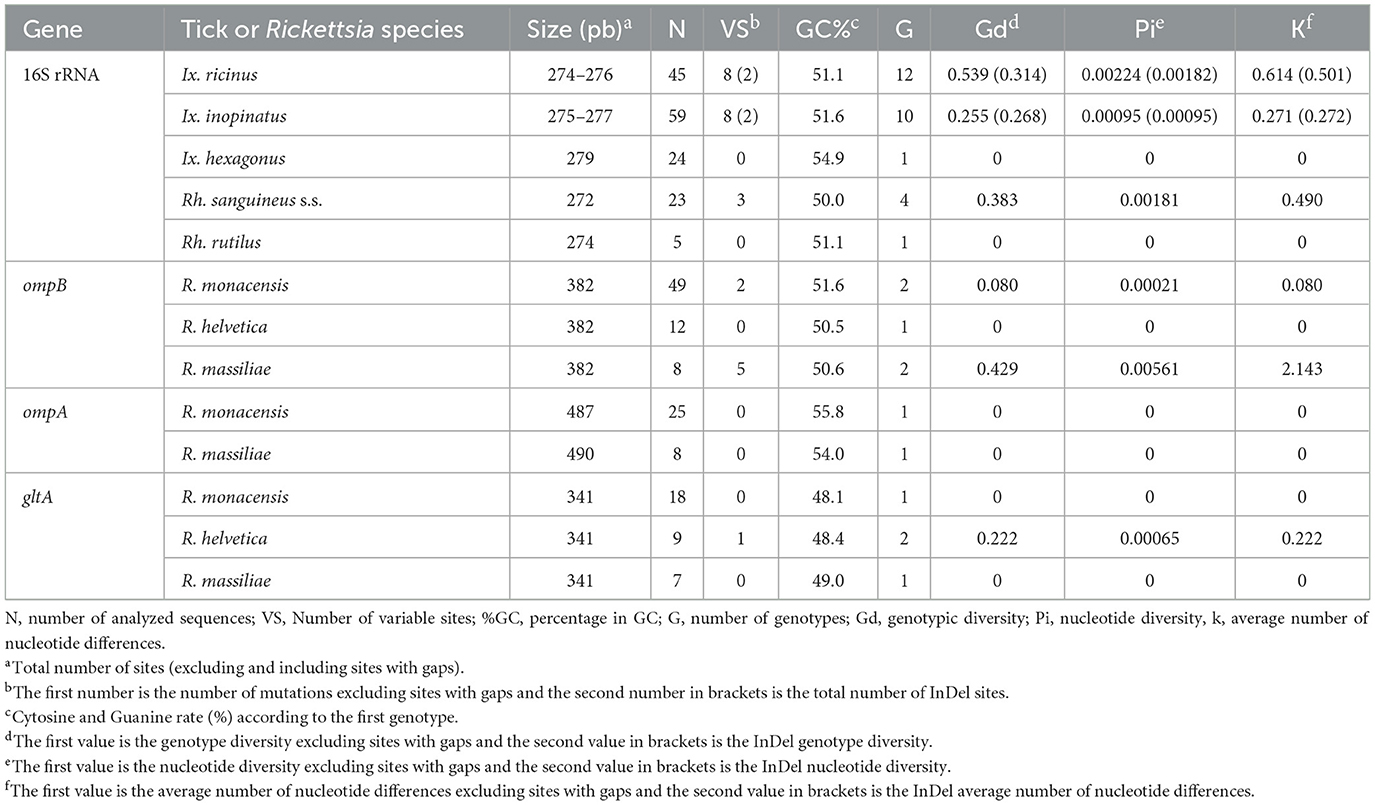
Table 4. Genetic diversity found within mitochondrial 16S rRNA partial sequences isolated from selected ticks and ompB, ompA and gltA partial sequences isolated from Rickettsia spp. infecting ticks.
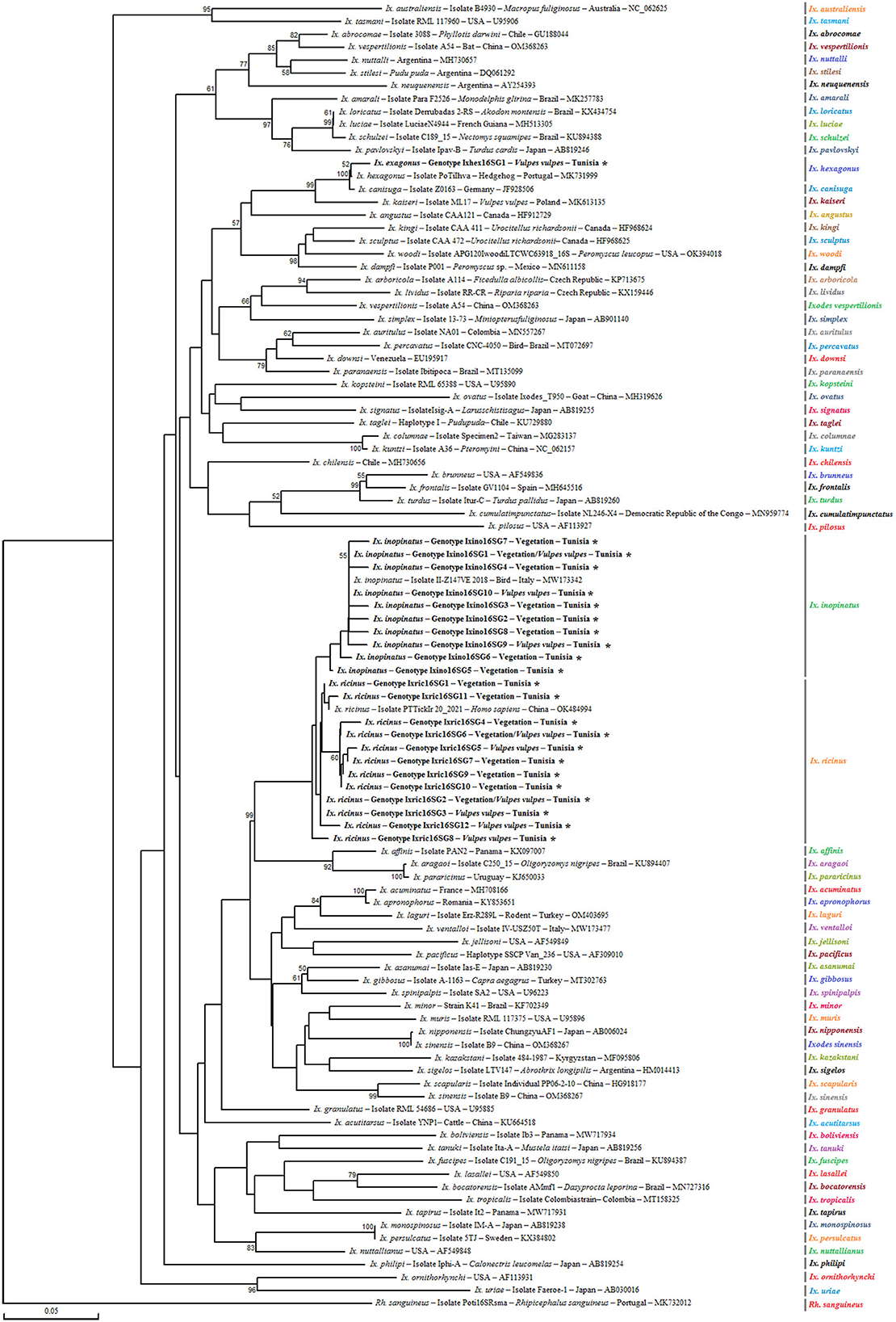
Figure 2. Phylogenetic analysis of partial (320 bp) mitochondrial 16S rRNA sequences obtained from Ixodes ricinus complex tick specimens and reference sequences from other Ixodes species available in GenBank, using the Neighbor-Joining method. Bootstrap values based on 1,000 iterations are shown at the nodes (only values above 50% are displayed). Information on host, strain, isolate or clone, country of origin, and GenBank accession number is provided. Sequences from the Ixodes ricinus complex generated in this study are shown in bold and marked with an asterisk. A partial mitochondrial 16S rRNA sequence from Rhipicephalus sanguineus was included as an outgroup.
On the other hand, for Ix. inopinatus, 10 genotypes (Ixino16SG1–Ixino16SG10) were identified (Table 5 and Supplementary File 2). Genotypic diversity (Gd) was 0.255, with a GC content of 51.6%. Nucleotide diversity (Pi) was 0.0095, with a mean pairwise nucleotide difference (k) of 0.271 (Table 4). Phylogenetic analysis confirmed that all identified genotypes belonged to the Ix. inopinatus cluster (Figure 2). With the exception of the two genotypes Ixino16SG5 and Ixino16SG6, which clustered closely, all other genotypes are part of a relatively homogeneous sub-cluster that is phylogenetically close to the Ix. inopinatus isolate found in a bird in Italy (GenBank accession number: MW173342) (Figure 2). Furthermore, genetic diversity analysis of Ix. hexagonus using DnaSP v5.10.01 identified a single genotype (Ixhex16SG1) in all 24 nymphs collected from V. vulpes (Supplementary File 3). This genotype exhibited a GC content of 54.9% and shared 98.9% sequence similarity with the PoTiIhva isolate from a Portuguese hedgehog (GenBank accession number: MK731999) (Table 4 and Figure 2).
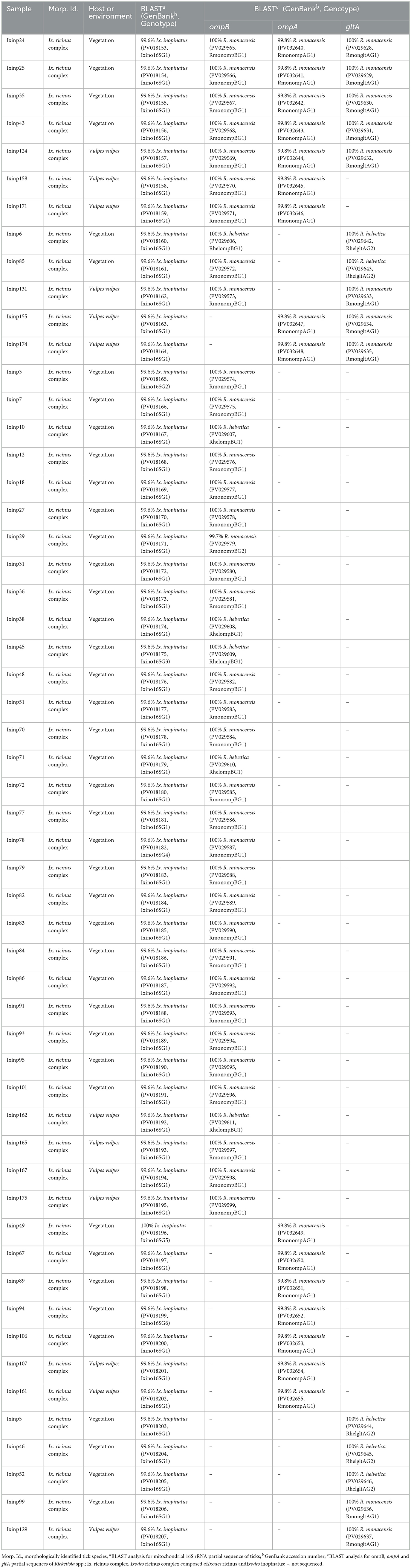
Table 5. Designation and information on the origins and genotypes of Tunisian isolates of Rickettsia spp. isolated from Ix. inopinatus ticks collected from vegetation and a red fox (Vulpes vulpes) in Jouza district (Beja governorate, Tunisia).
3.1.2.2 Genetic diversity and phylogeny within the Rh sanguineus s.l. complex
Analysis of the partial 16S rRNA sequence of Rh. sanguineus s.l. specimens collected from vegetation in Jouza identified four genotypes, designated as Rhsan16SG1-Rhsan16SG4 (Table 6 and Supplementary File 4). Genotypic diversity (Gd) was 0.421, with a GC content of 50%. Nucleotide diversity (Pi) was 0.0089, and k was 0.512 (Table 4). Interestingly, a single genotype (Rhrut16SG1) was identified in five Rh. rutilus specimens (Tables 4, 6 and Supplementary File 4). Phylogenetic analysis revealed that the group of Rh. sanguineus s.l. comprises four tick species within the Rhipicephalus genus, namely Rh. sanguineus sensu strito, Rh. rutilus, Rh. secundus, and Rh. turanicus (Figure 3). Rh. sanguineus s.s. cluster was composed of two sub-clusters supported by bootstrap values of 100% (Figure 3). All our genotypes of Rh. sanguineus s.s. were classified within sub-cluster 1, along with Haplotype 7 of Rh. sanguineus s.s. infecting a dog in Portugal (GenBank accession number: KY216136) (Figure 3). Rh. rutilus cluster was composed of three sub-clusters, with a bootstrap value of 93% between sub-cluster 1 and sub-cluster 2, and 87% between sub-cluster 2 and sub-cluster 3 (Figure 3). The genotype identified in this study of Rh. rutilus was present in sub-cluster 1, along with the isolate R02G20B of Rh. rutilus from Greece (GenBank accession number: PQ002091) (Figure 3).
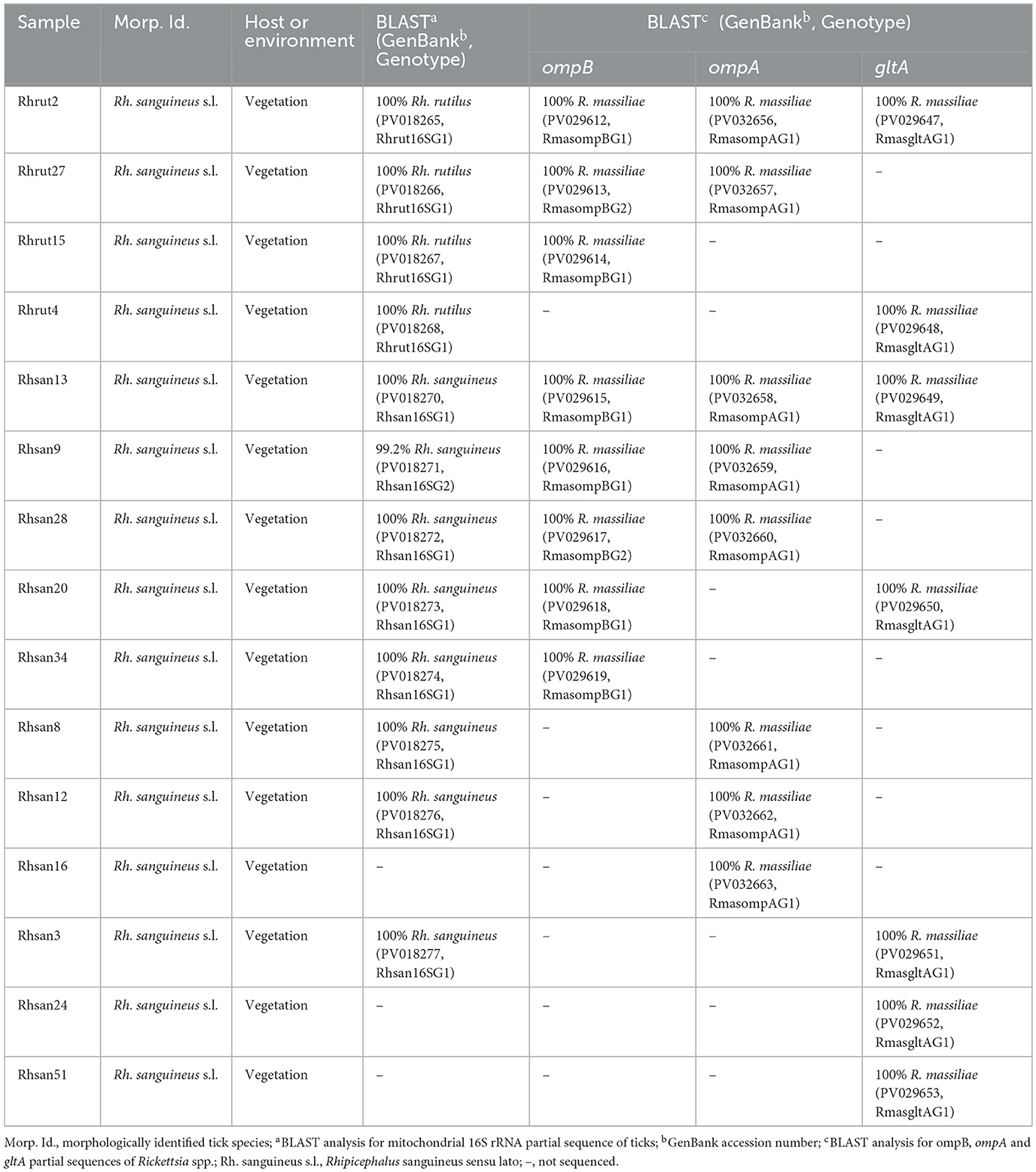
Table 6. Designation and information on the origins and genotypes of Tunisian isolates of Rickettsia spp. isolated from Rh. rutilus and Rh. sanguineus ticks collected from vegetation in Jouza district (Beja governorate, Tunisia).
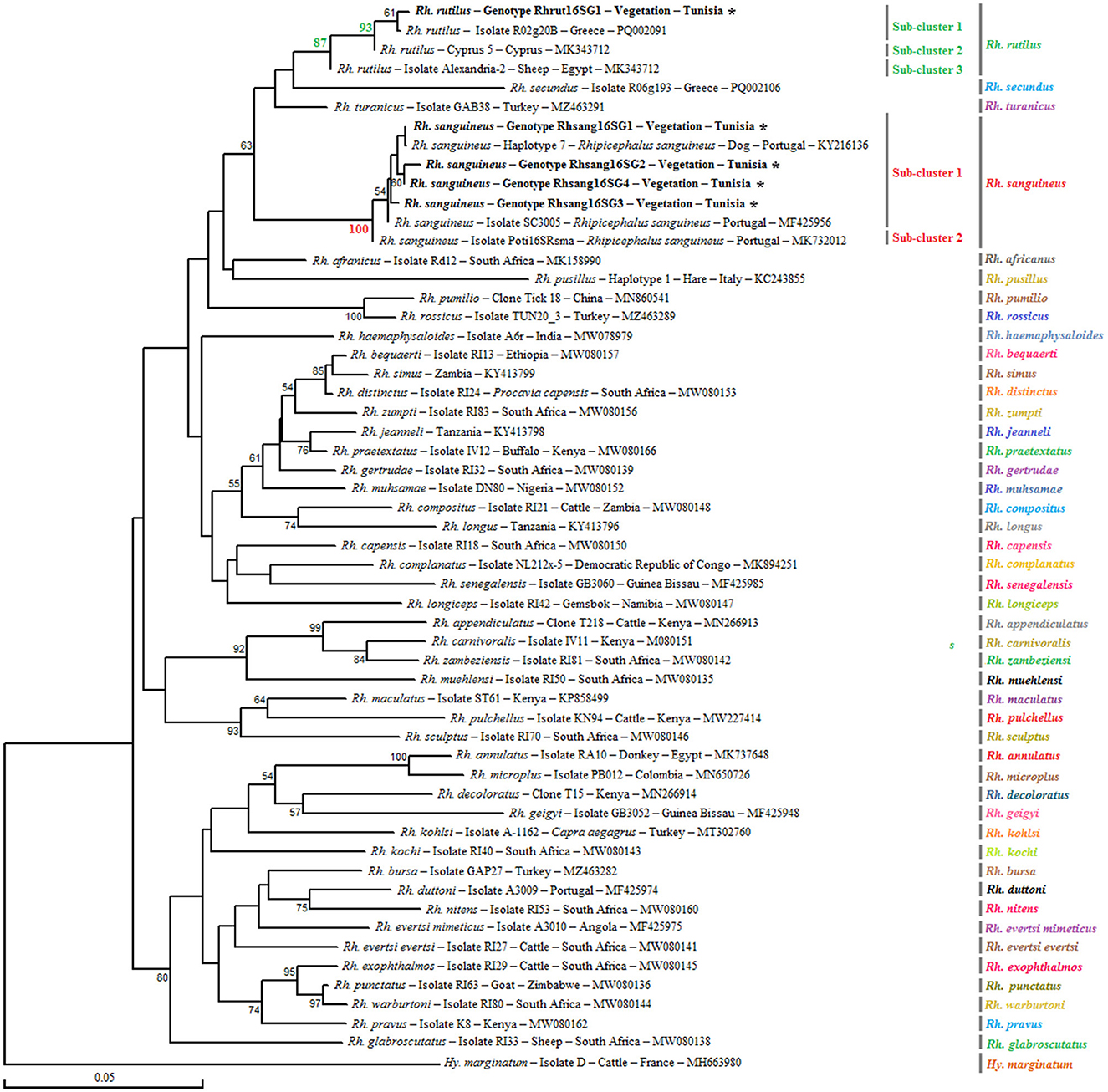
Figure 3. Phylogenetic analysis of partial (320 bp) mitochondrial 16S rRNA sequences obtained from tick specimens identified as Rhipicephalus sanguineus sensu lato and reference sequences from other Rhipicephalus species available in GenBank, using the Neighbor-Joining method. Bootstrap values based on 1,000 iterations are shown at the nodes (only values above 50% are displayed). Information on host, strain, isolate or clone, country of origin, and GenBank accession number is provided. Sequences of Rh. sanguineus s.l. generated in this study are shown in bold and marked with an asterisk. A partial mitochondrial 16S rRNA sequence from Hyalomma marginatum was included as an outgroup.
3.2 Molecular prevalence of Rickettsia spp.
PCR amplification targeting ompB revealed an overall Rickettsia spp. infection rate of 52.1% (123/236) (Table 2). Infection rates were comparable between the two tick species of the Ix. ricinus complex, with Ix. inopinatus at 67.4% and Ix. ricinus at 75.6% (Table 2). In contrast, the Rh. sanguineus s.l. complex exhibited a lower infection rate of 37.2% (Table 2). Notably, all Ix. hexagonus samples tested negative for Rickettsia sp. (0%) (Table 2). These differences were statistically significant (p < 0.001; Table 2). Additionally, infection prevalence varied by gender and developmental stage, with rates of 65.4% in males, 55.6% in females, and 0% in nymphs, showing a statistically significant difference (p < 0.001; Table 2). Furthermore, distinct infection rates were observed based on collection source, with statistically significant differences (p < 0.001; Table 2). Infection rates included 60.5% (101 out of 167) in vegetation, 31.9% (22 out of 69) in the red fox host (Table 2).
3.3 Rickettsia species identification
To identify and genetically characterize the detected Rickettsia species, at least one of the three targeted genes (ompB, ompA, and gltA) was partially sequenced for the 123 Rickettsia-positive tick samples selected for analysis (Ix. inopinatus, n = 60; Ix. ricinus, n = 30; Rh. sanguineus s.l., n = 29) (Table 7). All 123 partial sequences were successfully obtained and deposited in GenBank under the accession numbers PV029551–PV029619 for ompB, PV032631–PV032663 for ompA, and PV029620–PV029653 for gltA. Three Rickettsia species were identified in ticks positive for Rickettsia spp. selected for genetic analysis namely R. monacensis, R. helvetica, and R. massiliae (Table 7). Furthermore, based on the analysis of the three genes, coinfection by R. monacensis and R. helvetica was reported in one specimen of Ix. inopinatus tick (Ixinp85) (Table 7). Furthermore, to validate the specificity and reliability of the PCR assays used for Rickettsia detection, amplification products of the ompB (425 bp), ompA (532 bp), and gltA (381 bp) genes were analyzed by electrophoresis on 1.5% agarose gels stained with ethidium bromide. Each PCR run included both positive controls (DNA from confirmed Rickettsia spp.) and negative controls (nuclease-free water). Clear, specific bands of the expected sizes were observed in positive controls and sample lanes, while no amplification was detected in the negative controls, confirming the absence of contamination and the efficiency of the assays (Supplementary Figures S1B–S1D).
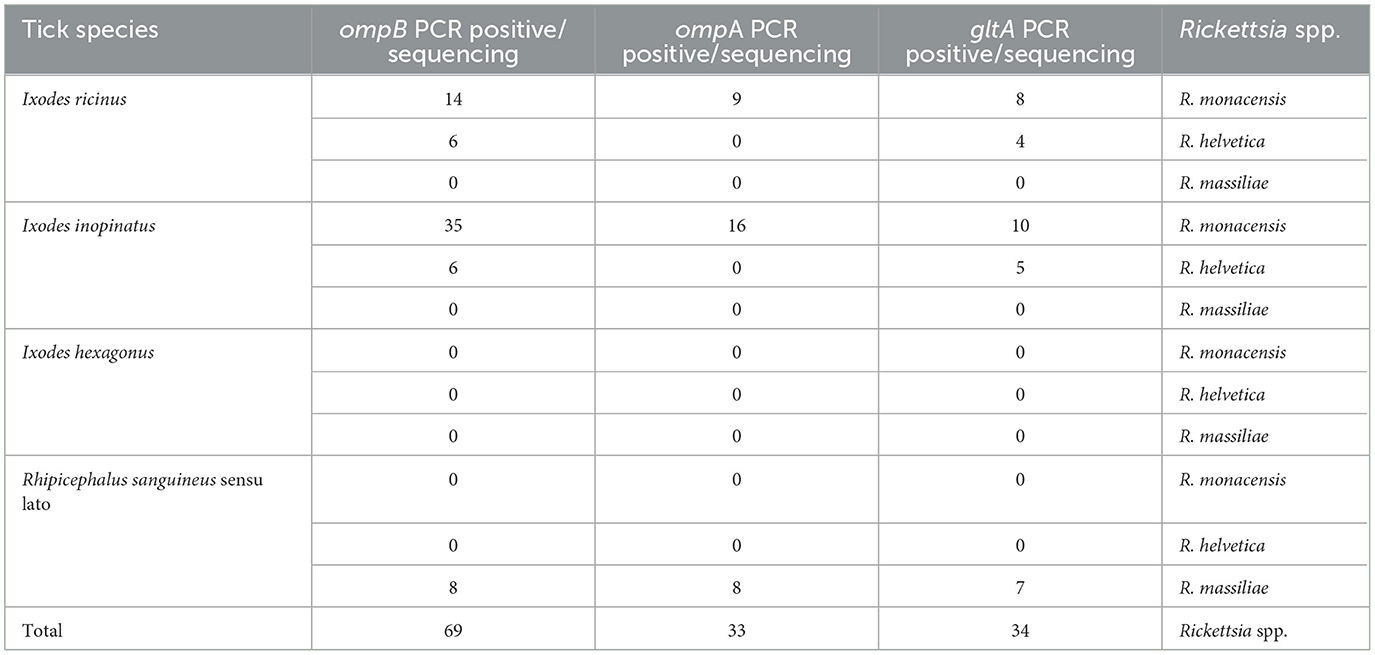
Table 7. Rickettsia species identified by sequencing of partial ompB, ompA and gltA gene sequences infecting Ixodes and Rhipicephalus sanguineus sensu lato ticks.
3.4 Genotyping and phylogenetic analysis
For the three genes analyzed, we identified distinct genotypes, each defined by at least one nucleotide difference.
3.4.1 Rickettsia spp. ompB partial sequences
The sequencing of ompB partial sequence (382 bp) was performed on 69 tick samples belonging to Ix. ricinus (n = 20), Ix. inopinatus (n = 41), and Rh. sanguineus s.l. (n = 8) (Table 7). The BLAST analysis revealed the presence of R. monacensis in 14 Ix. ricinus and 35 Ix. inopinatus specimens, and R. helvetica in 6 Ix. ricinus and 6 Ix. inopinatus specimens (Tables 5, 7). In addition, all Rh. sanguineus s.l. ticks were positive for R. massiliae (Table 7).
The genetic diversity analysis carried out using DnaSP version 5.10.01 software on a 382 bp of the ompB gene made it possible to identify two genotypes for R. monacensis, named RmonompBG1 and RmonompBG2, both found in Ix. ricinus (14 specimens) and Ix. inopinatus (35 specimens), with genetic diversity equal to 0.080. The GC rate was 51.6%. The nucleotide diversity (Pi) and average number of nucleotide differences (k) were estimated, respectively, at 0.00021 and 0.080 by noting the presence of two mutational positions between the two different revealed genotypes, sharing 99.74% nucleotide similarity (Table 4). RmonompBG1 and RmonompBG2 genotypes clustered with an isolate from Ix. ricinus in Romania (GenBank accession number: JX631117) and Haemaphysalis punctata in Russia (GenBank accession number: KU961543) (Figure 4).
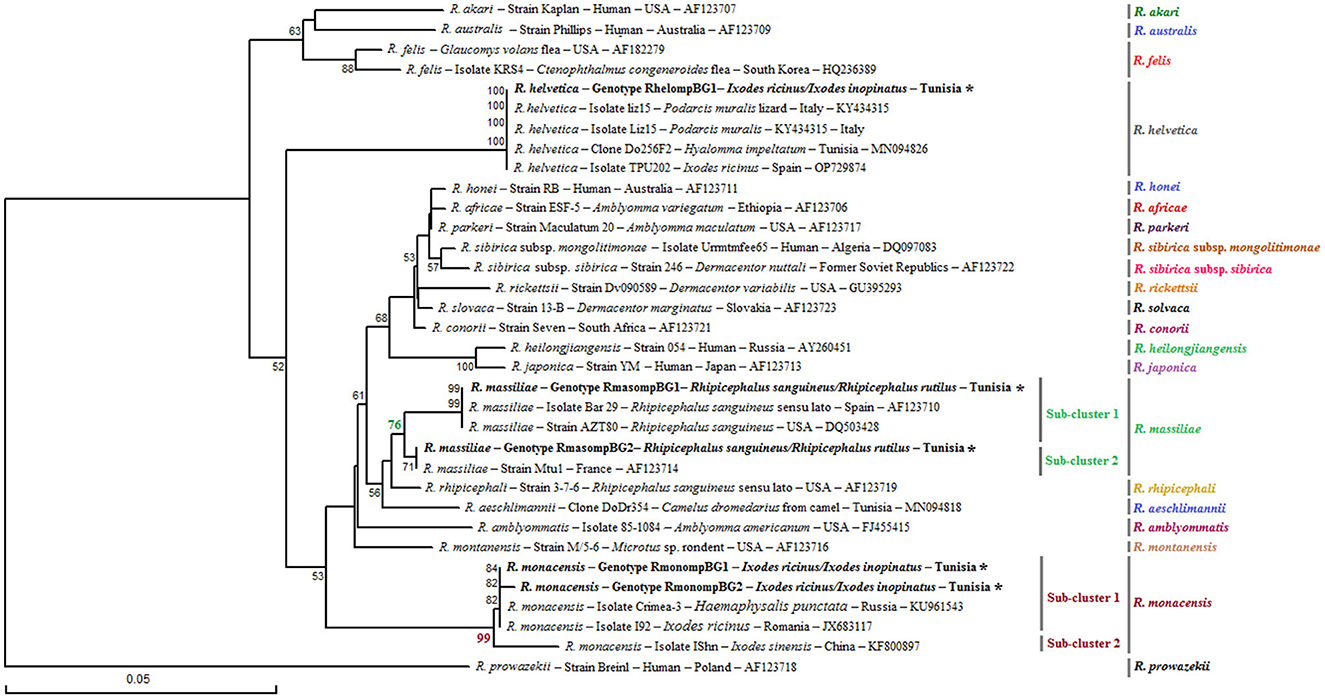
Figure 4. Phylogenetic tree of Rickettsia species based on partial ompB sequences (382 bp) of Rickettsia spp. obtained in this study, along with selected representative sequences of the genus Rickettsia. Bootstrap values from 1,000 replicates are shown above the branches (only values above 50% are displayed). Partial ompB sequences representing the different Rickettsia genotypes identified in this study are shown in bold and marked with an asterisk. Information on the host or vector, genotype, strain or isolate name, country of origin, and GenBank accession number is provided. A partial ompB sequence from Rickettsia prowazekii was included as an outgroup.
The sequence alignment of R. helvetica revealed a single genotype named RhelompBG1 isolated from specimens belonging to Ix. ricinus and Ix. inopinatus (Tables 5, 6). This genotype was found identical to three previously reported ompB isolates: clone Do256F2 from a Hy. impeltatum tick in Tunisia (GenBank accession number: MN094826), isolate TPU202 from an Ix. ricinus tick in Spain (GenBank accession number: OP729874), and isolate liz15 detected in Podarcis muralis from Italy (GenBank accession number: KY434315) (Figure 4).
Additionally, the alignment of partial sequences belonging to R. massiliae made it possible to select two genotypes named RmasompBG1 and RmasompBG2 infecting six Rh. sanguineus s.l. specimens, with genetic diversity equal to 0.429. The GC rate was 50.6%. The nucleotide diversity (Pi) and average number of nucleotide differences (k) were estimated, respectively, at 0.00561 and 2.143 by noting the presence of two mutational positions between the two different revealed genotypes, sharing 98.69% nucleotide similarity (Tables 4, 7). The genotype RmasompBG1, identified in six Rh. sanguineus s.l. specimens was found identical to isolates from Rh. sanguineus s.l. in Spain (GenBank accession number: AF123710) and the USA (GenBank accession number: DQ503428). However, the second genotype RmasompBG2, detected in a single Rh. sanguineus specimen, was found identical to a pathogenic R. massiliae isolate previously reported in France (GenBank accession number: AF123714) (Figure 4).
Phylogenetic analysis, based on the alignment of ompB genotypes with reference sequences from GenBank, revealed distinct clusters corresponding to the three identified Rickettsia species (Figure 4). The R. monacensis cluster comprised two subclusters with a node robustness equal to 99%. The first subcluster included genotypes RmonompBG1 and RmonompBG2, which grouped with isolates from Ix. ricinus in Romania (GenBank accession number: JX631117) and Haemaphysalis punctata in Russia (GenBank accession number: KU961543). The second subcluster consisted solely of isolate IShn from Ix. sinensis in China (GenBank accession number: KF800897) (Figure 4). R. helvetica genotype RhelompBG1 formed a single cluster with previously reported isolates from Hy. impeltatum in Tunisia (GenBank accession number: MN094826), Ix. ricinus in Spain (GenBank accession number: OP729874), and Podarcis muralis in Italy (GenBank accession number: KY434315) (Figure 4). Besides, the R. massiliae cluster was divided into two closely related sub-clusters with a node robustness equal to 76%. Genotype RmasompBG1 grouped with isolates from Rh. sanguineus s.l. in Spain (GenBank accession number: AF123710) and the USA (GenBank accession number: DQ503428), while genotype RmasompBG2 clustered with the pathogenic strain Mtu1 from France (GenBank accession number: AF123714) (Figure 4).
3.4.2 Rickettsia spp. ompA partial sequences
The analysis of ompA partial sequences confirmed the presence of R. monacensis and R. Massiliae (Tables 5, 6). The results showed that 9 Ix. ricinus and 16 Ix. inopinatus ticks tested positive for R. monacensis. However, 8 specimens of Rh. sanguineus were found positive for R. massiliae (Tables 5–7).
The alignment of sequences isolated for R. monacensis allowed us to select a single genotype named RmonompAG1 infecting 9 Ix. ricinus and 16 Ix. inopinatus. This genotype was found to be identical to isolates from Rattus rattus in Tunisia (GenBank accession number: MW978774) and Rh. sanguineus s.l in Spain (GenBank accession number: ON859981) (Figure 5). Furthermore, the sequences belonging to R. massiliae infecting eight Rh. sanguineus s.l. tick specimens allowed us to select a single genotype named RmasompAG1. The BLAST analysis showed that this genotype was identical to three previously reported isolates from Hy. marginatum in Tunisia (GenBank accession number: OQ123676), Homo sapiens in Spain (GenBank accession number: PP552102) and Ornithodoros lahorensis in China (GenBank accession number: OM475667) (Figure 5).
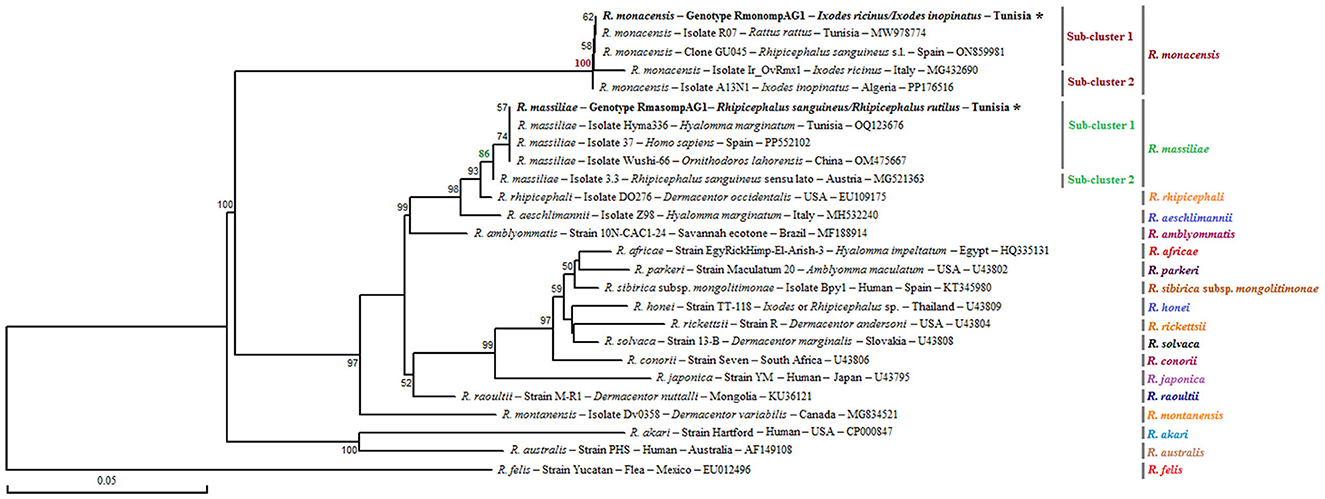
Figure 5. Neighbor-Joining tree based on the alignment of partial ompA sequences (490 bp), showing newly obtained sequences from ticks collected in vegetation. Bootstrap values from 1,000 replicates are shown at each node (only values above 50% are displayed). Rickettsia spp. genotypes identified in this study are shown in bold and marked with an asterisk. Information on the host or vector, genotype, strain or isolate name, country of origin, and GenBank accession number is provided. A partial ompA sequence from Rickettsia felis was included as an outgroup.
Phylogenetic analysis of the ompA gene, based on the alignment of our Tunisian genotypes with reference sequences from GenBank, revealed several distinct clusters (Figure 5). The R. monacensis cluster was divided into two sub-clusters with a node robustness equal to 100%. The genotype RmonompAG1 identified in this study grouped within the first sub-cluster, alongside isolates from Rattus rattus in Tunisia (isolate07; GenBank accession number: MW978774) and from Rh. sanguineus s.l. in Spain (clone GU045; GenBank accession number: ON859981) (Figure 5). The second sub-cluster comprised isolates Ir_OVrmx1 from Ix. ricinus in Italy (GenBank accession number: MG432690) and A13N1 from Ix. inopinatus in Algeria (GenBank accession number: PP176516) (Figure 5). On the other hand, the R. massiliae cluster also formed two subclusters with a node robustness equal to 86%. The genotype identified in this study (RmasompAG1) clustered within the first sub-cluster, together with isolates from Hy. marginatum in Tunisia (GenBank accession number: OQ123676), a human isolate from Spain (GenBank accession number: PP552102), and Ornithodoros lahorensis in China (GenBank accession number: OM475667) (Figure 5).
3.4.3 Rickettsia spp. gltA partial sequences
Sequencing of a 341 bp fragment of the gltA gene, which corresponds to the 381 bp amplified sequence excluding the forward and reverse primer sequences, revealed infections with R. monacensis, R. helvetica and R. massiliae (Tables 1–7). BLAST analysis confirmed the infection of 8 Ix. ricinus and 10 Ix. inopinatus with R. monacensis (Tables 5, 6). In addition, 4 Ix. ricinus and 5 Ix. inopinatus ticks tested positive for R. helvetica (Tables 5, 6). However, two Rh. rutilus ticks were found to be infected with R. massiliae along with 5 Rh. sanguineus tick specimens (Table 7). Sequence alignment of R. monacensis revealed a single genotype named RmongltAG1 infecting 8 and 10 specimens belonging, respectively, to Ix. ricinus and Ix. inopinatus (Tables 5, 6). This genotype was found to be identical to the R. monacensis isolate IIR28 infecting one Ix. ricinus from Algeria (GenBank accession number: PP315976) (Figure 6). In addition, the sequence alignment of R. helvetica isolates identified two genotypes named RhelgltAG1 and RhelgltAG2, infecting 4 Ix. ricinus and 5 Ix. inopinatus specimens, with genetic diversity equal to 0.222 (Tables 4–6). The GC content of the gltA sequences was 48.4%. The nucleotide diversity (Pi) and average number of nucleotide differences (k) were estimated at 0.00065 and 0.222, respectively, by noting the presence of two mutational positions between the two different revealed genotypes, sharing 99.71% nucleotide similarity (Table 4). These genotypes shared 99.71% and 100% genetic similarity, respectively, with an isolate KChR108-21 of R. helvetica from Ix. ricinus in Russia (GenBank accession number: PP431041). Alignment of R. massilae sequences revealed a single genotype named RmasgltAG1 identified in two Rh. rutilus and five Rh. sanguineus, respectively (Table 7). This genotype was found to be identical to the R. monacensis isolate IIR28 infecting one Ix. ricinus from Algeria (GenBank accession number: PP315976) (Figure 6).

Figure 6. Neighbor-Joining tree based on the alignment of partial gltA sequences (341 bp), showing newly obtained sequences from ticks collected in vegetation. Bootstrap values from 1,000 replicates are shown at each node (only values above 50% are displayed). Rickettsia spp. genotypes identified in this study are shown in bold and marked with an asterisk. Information on the host or vector, genotype, strain or isolate name, country of origin, and GenBank accession number is provided. A partial ompA sequence from Rickettsia prowazekii was included as an outgroup.
Phylogenetic analysis of the gltA gene revealed that the R. monacensis cluster comprised three sub-clusters with a node robustness equal to 83% (Figure 6). The genotype RmongltAG1 identified in this study grouped within the first sub-cluster, along with isolate lir28 from Ix. ricinus in Algeria (GenBank accession number: PP315976) (Figure 6). For R. helvetica, both genotypes RhelgltAG1 and RmongltAG2 clustered within the first sub-cluster with an isolate from Ix. ricinus in Russia (GenBank accession number: PP431041) (Figure 6). Additionally, the R. massiliae cluster was homogeneous, with a single genotype (RmasgltAG1) detected in this study. This genotype clustered with several previously reported R. massiliae isolates from Rh. sanguineus s.l specimens collected from several countries, as well as with an isolate from Hyalomma asiaticum (GenBank accession numbers: MH064442, KU498299, and MF098406) (Figure 6).
4 Discussion
Hard ticks (Acari: Ixodidae) are among the most significant arthropod vectors of Rickettsia spp. (Estrada-Peña et al., 2021). Several species within the genera Hyalomma, Amblyomma, Rhipicephalus, Ixodes, Haemaphysalis, and Dermacentor have been confirmed as competent vectors of these bacteria (Ammerman et al., 2004; Parola et al., 2005; Fournier and Raoult, 2019). Yet, in Tunisia, and more broadly across North Africa, data on the circulation of Rickettsia remain surprisingly limited. The present study contributes to filling this gap by conducting a descriptive investigation at a single location in Jouza district, Beja governorate, northwestern Tunisia. Ticks were collected from both vegetation and a red fox (V. vulpes) and their associated Rickettsia species were molecularly characterized.
Tick identification based on both morphological and molecular criteria confirmed the presence of five species: Ix. inopinatus, Ix. ricinus, Ix. hexagonus, Rh. sanguineus (temperate lineage), and Rh. rutilus. The detection of Ix. ricinus and Ix. inopinatus supports previous studies reporting their sympatric distribution in humid, forested regions of northern Tunisia (Younsi et al., 2019; Elati et al., 2022). The >99% identity of our sequences with European references confirms strong genetic relatedness between North African and European populations, likely reflecting shared Palearctic biogeographic patterns (Noureddine et al., 2011). Interestingly, Ix. inopinatus, initially described in southwestern Europe, has been increasingly detected across North Africa. In Algeria, (Mechouk et al. 2022) documented its wide habitat range, and (Daněk et al. 2024) identified all 149 Ixodes specimens in northeastern Algeria as Ix. inopinatus, suggesting ecological dominance or superior adaptation relative to Ix. ricinus. These findings raise important, yet unaddressed, ecological questions in the Tunisian context, particularly regarding niche partitioning, host preference, and how climate or land-use changes might be reshaping local species dynamics (Rosà et al., 2018; Horak et al., 2022; Hoch et al., 2024). Adding to this complexity, our phylogenetic analyses revealed substantial intra-specific variability in Ixodes spp., with 12 genotypes in Ix. ricinus and 10 in Ix. inopinatus. The presence of such high diversity in a single locality suggests multiple coexisting lineages shaped by host availability, microhabitat variation, or historical dispersal (Noureddine et al., 2011; Remesar et al., 2019). This is consistent with studies across the Mediterranean and Europe where similar patterns were interpreted as evidence of local structuring and possible cryptic speciation (Elati et al., 2022). In North Africa, the sympatric occurrence of genetically divergent Ixodes haplotypes may also signal introgression events or emerging species complexes, topics that call for deeper population-level resolution using high-throughput genomic tools.
Our study also marks the first molecular confirmation of Ix. hexagonus in Tunisia. Known for its nidicolous behavior and preference for hedgehogs, it also parasitizes a broad range of mammals, including foxes, mustelids, and domestic pets (Balti et al., 2021; Valentyna and Beata, 2024). Its detection on a red fox in Beja may reflect either an overlooked endemic presence or a subtle range expansion, masked by its endophilic lifestyle and under-sampling of wild hosts. In Algeria, this species was previously found on hedgehogs and dogs (Mechouk et al., 2022), and its widespread European distribution from the UK to Germany attests to its ecological plasticity (Abdullah et al., 2016; Cull et al., 2018; Kahl et al., 2022). In addition, the low mitochondrial 16S rRNA diversity observed in our specimens, all nymphs, may be linked to collection bias (single host), developmental constraints, or restricted dispersal associated with its lifestyle (Hornok et al., 2017). Furthermore, the high GC content and slow evolutionary rate of the 16S marker, especially in immature stages, could mask finer-scale variation (Boore, 1999; Estrada-Peña et al., 2017). Regarding the Rh. sanguineus s.l. complex, we confirmed the presence of both Rh. sanguineus (temperate lineage) and Rh. rutilus, consistent with recent findings in Tunisia and surrounding regions (Elati et al., 2018; Jomli et al., 2025; Senbill et al., 2024). Our phylogenetic analysis revealed that Rh. sanguineus clustered within sub-cluster 1 of Rh. sanguineus sensu stricto, while Rh. rutilus formed a distinct clade comprising three sub-clusters. Moreover, our genotype grouped with a Greek isolate (Ligda et al., 2025), reinforcing the unresolved taxonomy and complex genetic structure within the Rhipicephalus genus (Dantas-Torres et al., 2017; Estrada-Peña et al., 2017). This presence of host-specific or ecologically distinct genotypes suggests adaptive divergence possibly driven by geographic isolation or selective pressures which are mechanisms well recognized in tick evolution, but understudied in the Maghreb.
Infection prevalence by Rickettsia spp. reached 52.1% across all collected ticks. Among them, Ix. ricinus showed the highest infection rate (75.6%), comparable to levels reported in Germany (52.5%) and Poland (42.3%) (May and Strube, 2014; Dyczko et al., 2024). Ix. inopinatus also showed a high prevalence (67.4%), in agreement with findings from Algeria (81.9%) and Italy (31.2%) (Daněk et al., 2024). These results confirm the important role of Ixodes species in pathogen transmission in Tunisia and suggest that Ix. inopinatus, though less studied, might be as epidemiologically relevant as Ix. ricinus. On the other hand, Ix. hexagonus tested negative, possibly due to the nymphal stage of the specimens, which may limit pathogen acquisition and maintenance. This aligns with prior studies showing lower infection rates in immature ticks (Hartelt et al., 2004; Parola et al., 2005; Socolovschi et al., 2009). Additional sampling of adults is thus essential to clarify this species' potential role. Moreover, ticks of the Rh. sanguineus s.l. complex exhibited an overall infection rate of 37.2%, comparable to rates previously reported in Rhipicephalus ticks infesting small ruminants and cattle in Tunisia reported, respectively, by (Belkahia et al. 2021) and (Kratou et al. 2023). The detection of multiple genotypes in different host and environmental contexts suggests microevolutionary differentiation and possibly varying vectorial capacities (Gómez-Díaz et al., 2010; Jia et al., 2020; Ring et al., 2022).
Through sequencing of ompB, ompA, and gltA gene fragments, we identified R. monacensis, R. helvetica, and R. massiliae. Rickettsia monacensis was the most frequently detected species in Ixodes ticks from both vegetation and the red fox. This observation reinforces the role of Ix. ricinus and Ix. inopinatus as key vectors of these zoonotic agents, in line with previous reports from Europe and North Africa (Daněk et al., 2024). These results are consistent with European data, where R. monacensis predominates in southern regions such Bavaria (Germany) and Luxembourg, where R. helvetica predominated in Ix. ricinus populations (Reye et al., 2010; Schorn et al., 2011). In contrast, in southern and eastern Europe, R. monacensis was more commonly detected. For instance, studies from Spain have shown that the majority of Ix. ricinus specimens collected from vegetation or mammals were infected with R. monacensis, while R. helvetica was rarely identified (Márquez, 2008). A similar pattern was reported in Turkey, where R. monacensis showed significantly higher infection rates than R. helvetica (Gargili et al., 2012). These geographic differences in Rickettsia prevalence underscore the influence of environmental, ecological, and host-related factors on pathogen circulation within Ixodes populations. The high sequence similarity of our R. monacensis isolates with strains from Algeria, Spain, and even Rattus rattus in Tunisia suggests possible trans-Mediterranean circulation, perhaps facilitated by migratory birds or anthropogenic movements (Wasfi et al., 2021). Moreover, the detection of R. helvetica, especially in Ix. ricinus, and its close similarity to strains from the south of Tunisia, Spain, and Russia (Tomassone et al., 2017; Selmi et al., 2019; Rakov et al., 2024), reinforces its status as a widespread zoonotic agent in temperate zones. Several small mammal species and deer have been suggested as potential reservoirs (Špitalská et al., 2014), and the identification of R. helvetica in Tunisia raises questions about the local vertebrate hosts involved in its maintenance. On the other hand, R. massiliae was detected in Rh. sanguineus s.l. and Rh. rutilus, and sequences showed high similarity to isolates from France, Spain, and the United States (Roux and Raoult, 2000; Eremeeva et al., 2006), including strains previously reported from cattle, dogs, humans, and even Ornithodoros lahorensis in China (Vieira Lista et al., 2024). This confirms its broad host range and cosmopolitan distribution (Khrouf et al., 2013; Selmi et al., 2019; Belkahia et al., 2021; Kratou et al., 2023). Given the presence of pathogenic strains and the close proximity of these ticks to human and domestic animal populations, the potential public health risk cannot be overlooked. Given the detection of zoonotic Rickettsia species, including R. monacensis, R. helvetica, and R. massiliae, this study also highlights a tangible risk of pathogen spillover into human populations. These bacteria have been implicated in human febrile illnesses across Europe and the Mediterranean basin, with documented clinical cases associated with R. massiliae in France and the USA (Beati and Raoult, 1993; Eremeeva et al., 2006), R. monacensis in Spain and Germany (Márquez, 2008; Blanton, 2019), and R. helvetica in Switzerland and Sweden (Nilsson et al., 1999; Fournier, 2000). The close phylogenetic proximity of our sequences to those linked to human infections underscores the need to consider these pathogens in the differential diagnosis of febrile syndromes and to enhance integrated tick-borne disease surveillance in Tunisia under a One Health framework (Parola et al., 2013; Onyiche et al., 2022).
This study provides valuable baseline data on the diversity of Ixodes and Rhipicephalus ticks and their associated Rickettsia (spotted fever group) at an ecologically relevant site in Jouza, in the Beja Governorate, northwestern Tunisia, a region belonging to the subhumid bioclimatic zone favorable to these tick genera. However, several limitations should be noted. First, the focus on a single site limits the generalizability of the findings across Tunisia. This targeted descriptive approach was adopted as a pioneering study aimed at precisely characterizing local tick species and their Rickettsia diversity using molecular methods, including 16S rRNA sequencing of all tick samples and most amplicons of the three markers used to identify and characterize the revealed Rickettsia species. Future studies extending to multiple sites and diverse bioclimatic zones will be essential for providing broader epidemiological insights. Second, the host range sampled was primarily limited to ticks collected from vegetation, with the opportunistic inclusion of a single heavily infested red fox (V. vulpes). While this unexpected finding adds originality and highlights the potential role of wildlife in tick ecology, a more systematic inclusion of diverse wild and domestic hosts is needed to better understand host-pathogen interactions and reservoir dynamics. Third, some tick species, such as Rh. rutilus, were represented by relatively few specimens, which limits statistical power and phylogenetic resolution for these taxa. Increasing sample sizes for less common species in future research will strengthen these analyses. Fourth, molecular characterization relied on partial sequences of key genes (ompA, ompB, and gltA), providing robust initial insights into species and strain diversity. Nonetheless, whole-gene or whole-genome sequencing approaches would yield higher resolution of genetic variation and evolutionary relationships. Finally, this study focused exclusively on Rickettsia species, limiting the understanding of the full spectrum of tick-borne pathogens in the region. Incorporating screening for other important pathogens such as Borrelia and Anaplasma in future studies would provide a more comprehensive view of tick-borne disease ecology and co-infection patterns. Despite these limitations, this pioneering study offers critical baseline information and establishes a framework for more expansive epidemiological research across northern Tunisia.
5 Conclusion
This study confirms the presence of ticks belonging to the Ix. ricinus complex (Ix. ricinus and Ix. inopinatus), Ix. hexagonus, and the Rh. sanguineus s.l. complex (Rh. sanguineus (temperate lineage) and Rh. rutilus) in northwestern Tunisia. It highlights the presence of R. monacensis, R. helvetica, and R. massiliae in these ticks, emphasizing their role as vectors for spotted fever group rickettsiae. These findings enhance our understanding of the regional circulation of Rickettsia spp. in North Africa and underscore the need for ongoing epidemiological and molecular surveillance to evaluate their zoonotic potential. Such efforts are crucial for informing public health strategies and reducing the risk of tick-borne rickettsioses in the Mediterranean region.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary material.
Author contributions
MK: Conceptualization, Investigation, Writing – original draft, Writing – review & editing, Formal analysis, Data curation, Visualization, Methodology, Validation. HB: Validation, Methodology, Visualization, Writing – review & editing, Investigation. RS: Writing – review & editing, Visualization, Validation, Methodology, Investigation. MA: Writing – review & editing, Methodology, Investigation. GT: Writing – review & editing. LM: Writing – review & editing, Funding acquisition. MB: Visualization, Validation, Supervision, Writing – review & editing, Conceptualization, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the research laboratory “Laboratory of Epidemiology of Zoonotic Infections in Herbivores in Tunisia” (LR02AGR03), as well as by the following research projects funded by the Ministry of Higher Education and Scientific Research of Tunisia: “Screening and Molecular Characterization of Pathogenic and Zoonotic Bacteria of Medical and Economic Interest in Cattle and Camel Ticks in Tunisia” (19PEJC07-22), and “Study of the Bacterial Microbiota in Ticks with a Medical and Economic Impact in Tunisia: Contribution to the Control of Vector-Borne Bacterial Diseases” (P2ES2020-D4P1).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1644524/full#supplementary-material
References
Abdullah, S., Helps, C., Tasker, S., Newbury, H., and Wall, R. (2016). Ticks infesting domestic dogs in the UK: a large-scale surveillance programme. Parasit. Vectors 9:391. doi: 10.1186/s13071-016-1673-4
Ammerman, N. C., Swanson, K. I., Anderson, J. M., Schwartz, T. R., Seaberg, E. C., Glass, G. E., et al. (2004). Spotted-fever group Rickettsia in Dermacentor variabilis, Maryland. Emerg. Infect. Dis. 10, 1478–1481. doi: 10.3201/eid1008.030882
Balti, G., Galon, C., Derghal, M., Souguir, H., Guerbouj, S., Rhim, A., et al. (2021). Atelerix algirus, the North African Hedgehog: suitable wild host for infected ticks and fleas and reservoir of vector-borne pathogens in Tunisia. Pathogens 10:953. doi: 10.3390/pathogens10080953
Beati, L., and Raoult, D. (1993). NOTES: Rickettsia massiliae sp. nov., a New Spotted Fever Group Rickettsia. Int. J. Syst. Bacteriol. 43, 839–840. doi: 10.1099/00207713-43-4-839
Belkahia, H., Selmi, R., Sayed Zamiti, Monia Daaloul-Jedidi, Messadi, L., and Said, M. B. (2021). Zoonotic Rickettsia species in small ruminant ticks from Tunisia. Front. Vet. Sci. 8:676896. doi: 10.3389/fvets.2021.676896
Black, W. C., and Piesman, J. (1994). Phylogeny of hard- and soft-tick taxa (Acari: Ixodida) based on mitochondrial 16S rDNA sequences. Proc. Natl. Acad. Sci. U.S.A. 91, 10034–10038. doi: 10.1073/pnas.91.21.10034
Blanton, L. S. (2019). The rickettsioses. Infect. Dis. Clin. North Am. 33, 213–229. doi: 10.1016/j.idc.2018.10.010
Boore, J. L. (1999). Animal mitochondrial genomes. Nucleic Acids Res. 27, 1767–1780. doi: 10.1093/nar/27.8.1767
Chisu, V., Masala, G., Foxi, C., Socolovschi, C., Raoult, D., and Parola, P. (2014). Rickettsia conorii israelensis in Rhipicephalus sanguineus ticks, Sardinia, Italy. Ticks Tick Borne Dis. 5, 446–8. doi: 10.1016/j.ttbdis.2014.02.003
Choi, Y. J., Jang, W. J., Kim, J. H., Ryu, J. S., Lee, S. H., Park, K. H., et al. (2005). Spotted fever group and typhus group rickettsioses in humans, South Korea. Emerg. Infect. Dis. 11, 237–244. doi: 10.3201/eid1102.040603
Cull, B., Pietzsch, M. E., Hansford, K. M., Gillingham, E. L., and Medlock, J. M. (2018). Surveillance of British ticks: an overview of species records, host associations, and new records of Ixodes ricinus distribution. Ticks and Tick-Borne Dis. 9, 605–614. doi: 10.1016/j.ttbdis.2018.01.011
Daněk, O., Hrbatová, A., Volfová, K., Ševčíková, S., Lesiczka, P., Nováková, M., et al. (2024). Italian peninsula as a hybridization zone of Ixodes inopinatus and I. ricinus and the prevalence of tick-borne pathogens in I. inopinatus, I. ricinus, and their hybrids. Parasit. Vectors 17:196. doi: 10.1186/s13071-024-06271-z
Dantas-Torres, F. (2010). Biology and ecology of the brown dog tick, Rhipicephalus Sanguineus Parasit. Vectors 3:26. doi: 10.1186/1756-3305-3-26
Dantas-Torres, F., Maia, C., Latrofa, M. S., Annoscia, G., Cardoso, L., and Otranto, D. (2017). Genetic characterization of Rhipicephalus sanguineus (sensu lato) ticks from dogs in Portugal. Parasit. Vectors 10:133. doi: 10.1186/s13071-017-2072-1
Demoncheaux, J.-P., Socolovschi, C., Davoust, B., Haddad, S., Raoult, D., and Parola, P. (2012). First detection of Rickettsia aeschlimannii in Hyalomma dromedarii ticks from Tunisia. Ticks and Tick-Borne Dis. 3, 398–402. doi: 10.1016/j.ttbdis.2012.10.003
Dyczko, D., Błażej, P., and Kiewra, D. (2024). The influence of forest habitat type on Ixodes ricinus infections with Rickettsia spp. in south-western Poland. Curr. Res. Parasitol. Vector Borne Dis. 6:100200. doi: 10.1016/j.crpvbd.2024.100200
Elati, K., Hamdi, D., Jdidi, M., and Mourad Rekik, Gharbi, M. (2018). Differences in tick infestation of Tunisian sheep breeds. Vet. Parasitol. Reg. Stud. Reports 13, 50–54. doi: 10.1016/j.vprsr.2018.03.011
Elati, K., Khbou, M. K., Kahl, O., Mwacharo, J. M., El Shamaa, K., Rekik, M., et al. (2022). Preliminary study on the seasonal questing of Ixodes ricinus group ticks in Ain Draham forest (north-western Tunisia) with analyses of their phylogenetic diversity. Vet. Parasitol. Reg. Stud. Reports 36:100786. doi: 10.1016/j.vprsr.2022.100786
Eremeeva, M. E., Bosserman, E., Demma, L. J., Zambrano, M. L., Blau, D. M., and Dasch, G. A. (2006). Isolation and identification of Rickettsia massiliae from Rhipicephalus sanguineus ticks collected in Arizona. Appl. Environ. Microbiol. 72, 5569–5577. doi: 10.1128/AEM.00122-06
Estrada-Peña, A., Binder, L. C., Nava, S., Szabó, M. P. J., and Labruna, M. B. (2021). Exploring the ecological and evolutionary relationships between Rickettsia and hard ticks in the Neotropical region. Ticks Tick-Borne Dis. :101754. doi: 10.1016/j.ttbdis.2021.101754
Estrada-Peña, A., and de la Fuente, J. (2014). The ecology of ticks and epidemiology of tick-borne viral diseases. Antiviral Research 108, 104–128. doi: 10.1016/j.antiviral.2014.05.016
Estrada-Peña, A., Mihalca, A. D., and Petney, T. N. (eds)., (2017). Ticks of Europe and North Africa. Cham: Springer International Publishing doi: 10.1007/978-3-319-63760-0
Felsenstein, J., and Sinauer Associates (2008). Inferring phylogenies. Sunderland, Mass.: Sinauer Associates, Inc., [Ca ], Cop.
Fournier, P.-E. (2000). Evidence of infection in humans with Rickettsia helvetica in Eastern France. Emerg. Infect. Dis. 6, 389–392. doi: 10.3201/eid0604.000412
Fournier, P.-E., and Raoult, D. (2019). Tick-Borne Spotted Fever Rickettsioses. Amsterdam: Elsevier. 587–593. doi: 10.1016/B978-0-323-55512-8.00069-7
Gargili, A., Palomar, A. M., Midilli, K., Portillo, A., Kar, S., and Oteo, J. A. (2012). Rickettsia species in ticks removed from humans in Istanbul, Turkey. Vector Borne Zoonotic. Dis. 12, 938–41. doi: 10.1089/vbz.2012.0996
Gerardi, M., Ramírez-Hernández, A., Binder, L. C., Felipe, Gregori, F., and Marcelo, B. L. (2019). Comparative susceptibility of different populations of Amblyomma sculptum to Rickettsia rickettsii. Front. Physiol. 10:653. doi: 10.3389/fphys.2019.00653
Gómez-Díaz, E., Doherty, P. F. Jr, Duneau, D., and McCoy, K. D. (2010). Cryptic vector divergence masks vector-specific patterns of infection: an example from the marine cycle of Lyme borreliosis. Evol. Appl. 3, 391–401. doi: 10.1111/j.1752-4571.2010.00127.x
Hartelt, K., Oehme, R., Frank, H., Brockmann, S. O., Hassler, D., and Kimmig, P. (2004). Pathogens and symbionts in ticks: prevalence of Anaplasma phagocytophilum (Ehrlichia sp.), Wolbachia sp., Rickettsia sp., and Babesia sp. in Southern Germany. Int. J. Med. Microbiol. 293, 86–92. doi: 10.1016/S1433-1128(04)80013-5
Hauck, D., Jordan, D., Springer, A., Schunack, B., Pachnicke, S., Fingerle, V., et al. (2020). Transovarial transmission of Borrelia spp., Rickettsia spp. and Anaplasma phagocytophilum in Ixodes ricinus under field conditions extrapolated from DNA detection in questing larvae. Parasit. Vectors 13:176. doi: 10.1186/s13071-020-04049-7
Hoch, T., Aurélien, M., Jacquot, M., Phrutsamon, W., Beugnet, F., Bournez, L., et al. (2024). Seasonality of host-seeking Ixodes ricinus nymph abundance in relation to climate. Peer Commun. J. 4:e2. doi: 10.24072/pcjournal.355
Horak, I. G., Junker, K., and Krasnov, B. R. (2022). Similarity in ixodid tick communities harboured by wildlife and livestock in the Albany Thicket Biome of South Africa. Parasitology 149, 667–674. doi: 10.1017/S0031182022000129
Hornok, S, Mulvihill, M., Szoke, K., Gönczi, E., Sulyok, K. M., Gyuranecz, M., et al. (2017). Impact of a freeway on the dispersal of ticks and Ixodes ricinus-borne pathogens: forested resting areas may become Lyme disease hotspots. Acta Vet. Hung. 65, 242–252. doi: 10.1556/004.2017.024
Jia, N., Wang, J., Shi, W., Du, L., Sun, Y., Zhan, W., et al. (2020). Large-scale comparative analyses of tick genomes elucidate their genetic diversity and vector capacities. Cell 182, 1328-1340.e13. doi: 10.1016/j.cell.2020.07.023
Jomli, A., Sebai, E., Amairia, S., Zamiti, S., Mhadhbi, M., Dhibi, M., et al. (2025). Multi-marker analysis reveals an extensive genetic diversity in Rhipicephalus sanguineus (temperate lineage) and the first identification of Rhipicephalus rutilus in Tunisia. Acta Trop. 267:107685. doi: 10.1016/j.actatropica.2025.107685
Kahl, O., Bulling, I., and Chitimia-Dobler, L. (2022). Some new findings on the endophilic vector tick Ixodes hexagonus in Germany. Ticks Tick-Borne Dis. 13:101954. doi: 10.1016/j.ttbdis.2022.101954
Kernif, T., Socolovschi, C., Bitam, I., Raoult, D., and Parola, P. (2012). Vector-Borne Rickettsioses in North Africa. Infect. Dis. Clin. North Am. 26, 455–478. doi: 10.1016/j.idc.2012.03.007
Khrouf, F., M'Ghirbi, Y., Znazen, A., Ben Jemaa, M., Hammami, A., and Bouattour, A. (2013). Detection of Rickettsia in Rhipicephalus sanguineus ticks and Ctenocephalides felis fleas from Southeastern Tunisia by reverse line blot assay. J. Clin. Microbiol. 52, 268–274. doi: 10.1128/JCM.01925-13
Kim, H. K. (2022). Rickettsia-host-tick interactions: knowledge advances and gaps. Infect. Immun. 90:e0062121. doi: 10.1128/iai.00621-21
Kratou, M., Belkahia, H., Selmi, R., Andolsi, R., Dhibi, M., Mhadhbi, M., et al. (2023). Diversity and phylogeny of cattle ixodid ticks and associated spotted fever group Rickettsia spp. in Tunisia. Pathogens 12, 552–552. doi: 10.3390/pathogens12040552
Krawczak, F. S., Agostinho, W. C., Polo, G., Moraes-Filho, J., and Labruna, M. B. (2016). Comparative evaluation of Amblyomma ovale ticks infected and noninfected by Rickettsia sp. strain Atlantic rainforest, the agent of an emerging rickettsiosis in Brazil. Ticks Tick-Borne Dis. 7, 502–507. doi: 10.1016/j.ttbdis.2016.02.007
Laukaitis, H. J., and Macaluso, K. R. (2021). Unpacking the intricacies of Rickettsia-vector interactions. Trends Parasitol. 37, 734–746. doi: 10.1016/j.pt.2021.05.008
Ligda, P., Šlapeta, J., Saratsis, A., Kantzoura, V., Chong, J., and Sotiraki, S. (2025). Redefining Rhipicephalus sanguineus (sensu lato) species complex in Greece focusing on the mitogenome of Rhipicephalus secundus. Curr. Res. Parasitol. Vector-Borne Dis. 7:100231. doi: 10.1016/j.crpvbd.2024.100231
Liu, G., Zhao, S., Tan, W., Sándor Hornok, Yuan, W., Mi, L., et al. (2021). Rickettsiae in red fox (Vulpes vulpes), marbled polecat (Vormela peregusna) and their ticks in northwestern China. Parasit. Vectors 14:204. doi: 10.1186/s13071-021-04718-1
Marié, J., Davoust, B., Socolovschi, C., Oleg, M., Cédric Roqueplo, Jean-Claude Beaucournu, et al. (2011). Rickettsiae in arthropods collected from red foxes (Vulpes vulpes) in France. Comp. Immunol. Microbiol. Infect. Dis. 35, 59–62. doi: 10.1016/j.cimid.2011.10.001
Márquez, F. J. (2008). Spotted fever group Rickettsia in ticks from southeastern Spain natural parks. Exp. Appl. Acarol. 45, 185–194. doi: 10.1007/s10493-008-9181-7
Martina, B., Barzon, L., Pijlman, G. P., de, J., Rizzoli, A., Wammes, L. J., et al. (2017). Human to human transmission of arthropod-borne pathogens. Curr. Opin. Virol. 22, 13–21. doi: 10.1016/j.coviro.2016.11.005
May, K., and Strube, C. (2014). Prevalence of Rickettsiales (Anaplasma phagocytophilum and Rickettsia spp.) in hard ticks (Ixodes ricinus) in the city of Hamburg, Germany. Parasitol. Res. 113, 2169–2175. doi: 10.1007/s00436-014-3869-x
Mechouk, N., Mihalca, A. D., Deak, G., and Bouslama, Z. (2022). Synopsis of the ticks of Algeria with new hosts and localities records. Parasit. Vectors 15:302. doi: 10.1186/s13071-022-05424-2
Nieri-Bastos, F. A., Szabó, M. P. J., Pacheco, R. C., Soares, J. F., Soares, H. S., Moraes-Filho, J., et al. (2013). Comparative evaluation of infected and noninfected Amblyomma triste ticks with Rickettsia parkeri, the agent of an emerging rickettsiosis in the New World. BioMed Res. Int. 2013:402737. doi: 10.1155/2013/402737
Nilsson, K., Lindquist, O., and Påhlson, C. (1999). Association of Rickettsia helvetica with chronic perimyocarditis in sudden cardiac death. Lancet 354, 1169–1173. doi: 10.1016/S0140-6736(99)04093-3
Noureddine, R., Chauvin, A., and Plantard, O. (2011). Lack of genetic structure among Eurasian populations of the tick Ixodes ricinus contrasts with marked divergence from north-African populations. Int. J. Parasitol. 41, 183–192. doi: 10.1016/j.ijpara.2010.08.010
Nováková, M., and Šmajs, D. (2019). Rickettsial Endosymbionts of Ticks. IntechOpen eBooks. doi: 10.5772/intechopen.80767
Onyiche, T. E., Labruna, M. B., and Saito, T. B. (2022). Unraveling the epidemiological relationship between ticks and rickettsial infection in Africa. Front. Trop. Dis. 3:952024. doi: 10.3389/fitd.2022.952024
Oteo, J. A., Portillo, A., Santibáñez, S., Blanco, J. R., Pérez-Martínez, L., and Ibarra, V. (2006). Cluster of cases of human Rickettsia felis infection from Southern Europe (Spain) diagnosed by PCR. J. Clin. Microbiol. 44, 2669–2671. doi: 10.1128/JCM.00366-06
Parola, P., Paddock, C. D., and Raoult, D. (2005). Tick-Borne rickettsioses around the world: emerging diseases challenging old concepts. Clin. Microbiol. Rev. 18, 719–756. doi: 10.1128/CMR.18.4.719-756.2005
Parola, P., Paddock, C. D., Socolovschi, C., Labruna, M. B., Mediannikov, O., Kernif, T., et al. (2013). Update on tick-borne rickettsioses around the world: a geographic approach. Clin. Microbiol. Rev. 26, 657–702. doi: 10.1128/CMR.00032-13
Rakov, A. V., Chekanova, T. A., Petremgvdlishvili, K., Linnik, S. B., Batchaev, K. K., and Akimkin, V. G. (2024). The diversity of spotted fever group rickettsia found in ixodidae hard ticks removed from Humans in Karachay-Cherkessia, North Caucasus, Russia. Microorganisms 12, 1293–1293. doi: 10.3390/microorganisms12071293
Regnery, R. L., Olson, J. G., Perkins, B. A., and Bibb, W. (1992). Serological response to “Rochalimaea henselae” antigen in suspected cat-scratch disease. Lancet 339, 1443–1445. doi: 10.1016/0140-6736(92)92032-B
Remesar, S., Fernández, P. D., Venzal, J. M., Pérez-Creo, A., Prieto, A., Estrada-Peña, A., et al. (2019). Tick species diversity and population dynamics of Ixodes ricinus in Galicia (north-western Spain). Ticks Tick-Borne Dis. 10, 132–137. doi: 10.1016/j.ttbdis.2018.09.006
Reye, A. L., Hübschen, J. M., Sausy, A., and Muller, C. P. (2010). Prevalence and seasonality of tick-borne pathogens in questing Ixodes ricinus ticks from Luxembourg. Appl. Environ. Microbiol. 76, 2923–2931. doi: 10.1128/AEM.03061-09
Ring, K., Couper, L. I., Sapiro, A. L., Yarza, F., Yang, X. F., Clay, K., et al. (2022). Host blood meal identity modifies vector gene expression and competency. Mol. Ecol. 31, 2698–2711. doi: 10.1111/mec.16413
Rosà, R., Andreo, V., Tagliapietra, V., Baráková, I., Arnoldi, D., Hauffe, H., et al. (2018). Effect of climate and land use on the spatio-temporal variability of tick-borne bacteria in Europe. Int. J. Environ. Res. Public Health 15:732. doi: 10.3390/ijerph15040732
Roux, V., and Raoult, D. (2000). Phylogenetic analysis of members of the genus Rickettsia using the gene encoding the outer-membrane protein rOmpB (ompB). Int. J. Syst. Evol. Microbiol. 50, 1449–1455. doi: 10.1099/00207713-50-4-1449
Sarih, M', Jouda, F., Gern, L., and Postic, D. (2003). First isolation of Borrelia burgdorferi Sensu Lato from Ixodes ricinus ticks in Morocco. Vector-Borne Zoo. Dis. 3, 133–139. doi: 10.1089/153036603768395834
Schorn, S., Pfister, K., Reulen, H., Mahling, M., and Silaghi, C. (2011). Occurrence of Babesia spp., Rickettsia spp. and Bartonella spp. in Ixodes ricinus in Bavarian public parks, Germany. Parasit. Vectors 4:135. doi: 10.1186/1756-3305-4-135
Selmi, R., Abdi, K., Belkahia, H., Abdallah, M. B., Mamlouk, A., Kratou, M., et al. (2024). Detection and genetic identification of Borrelia lusitaniae in questing Ixodes inopinatus tick from Tunisia. Infect. Med. 3:100093. doi: 10.1016/j.imj.2024.100093
Selmi, R., Belkahia, H., Ben Abdallah, M., Mamlouk, A., Daaloul-Jedidi, M., Ben Said, M., et al. (2025). First identification of Brucella melitensis and Rickettsia monacensis in a red fox, vulpes vulpes. Comp. Immunol. Microbiol. Infect. Dis. 117:102315. doi: 10.1016/j.cimid.2025.102315
Selmi, R., Said, M., Ben Yahia, H., Abdelaali, H., and Messadi, L. (2019). Molecular epidemiology and phylogeny of spotted fever group Rickettsia in camels (Camelus dromedarius) and their infesting ticks from Tunisia. Transbound. Emerg. Dis. 67, 733–744. doi: 10.1111/tbed.13392
Senbill, H., Karawia, D., Zeb, J., Alyami, N. M., Almeer, R., Rahman, S., et al. (2024). Molecular screening and genetic diversity of tick-borne pathogens associated with dogs and livestock ticks in Egypt. PLoS Negl. Trop. Dis. 18, e0012185–e0012185. doi: 10.1371/journal.pntd.0012185
Šlapeta, J., Halliday, B., Dunlop, J. A., Nachum-Biala, Y., Salant, H., Ghodrati, S., et al. (2023). The “southeastern Europe” lineage of the brown dog tick Rhipicephalus sanguineus (sensu lato) identified as Rhipicephalus rutilus Koch, 1844: Comparison with holotype and generation of mitogenome reference from Israel. Curr. Res. Parasitol. Vector Borne Dis. 3:100118. doi: 10.1016/j.crpvbd.2023.100118
Socolovschi, C., Mediannikov, O., Raoult, D., and Parola, P. (2009). The relationship between spotted fever group Rickettsiae and Ixodid ticks. Vet. Res. 40:34. doi: 10.1051/vetres/2009017
Sonenshine, D. (2018). Range expansion of tick disease vectors in North America: implications for spread of tick-borne disease. Int. J. Environ. Res. Public Health.15:478. doi: 10.3390/ijerph15030478
Špitalská, E., Boldiš, V., Derdáková, M., Selyemová, D., and Rusnáková Taragelová, V. (2014). Rickettsial infection in Ixodes ricinus ticks in urban and natural habitats of Slovakia. Ticks and Tick-borne Diseases 5, 161–165. doi: 10.1016/j.ttbdis.2013.10.002
Tamura, K., and Nei, M. (1993). Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10, 512–26. doi: 10.1093/oxfordjournals.molbev.a040023
Tomassone, L., Ceballos, L. A., C. Ragagli, Martello, E., Rita de Sousa, Stella, M., et al. (2017). Importance of common wall lizards in the transmission dynamics of tick-borne pathogens in the Northern Apennine Mountains, Italy. Microb. Ecol. 74, 961–968. doi: 10.1007/s00248-017-0994-y
Tsatsaris, A., Chochlakis, D., Papadopoulos, B., Petsa, A., Georgalis, L., Angelakis, E., et al. (2016). Species composition, distribution, ecological preference and host association of ticks in Cyprus. Exp. Appl. Acarol. 70, 523–542. doi: 10.1007/s10493-016-0091-9
Valentyna, K., and Beata, W. (2024). Molecular identification of Borreliella species in Ixodes hexagonus ticks infesting hedgehogs (Erinaceus europaeus and E. roumanicus) in North-Western Poland. Int. J. Mol. Sci. 26:58. doi: 10.3390/ijms26010058
Vieira Lista, M. C., Vicente Santiago, M. B., Soto-López, J. D., García-Martín, J. M., Álamo-Sanz, R., Belhassen-García, M., et al. (2024). Identification of Rickettsia spp. in ticks removed from tick-bitten humans in Northwestern Spain. Insects 15:571. doi: 10.3390/insects15080571
Walker, A. R., Matthews, J., and Preston, P. M. (2005). The development of electronic keys for the identification of ticks. Int. J. Trop. Insect Sci. 25, 2–5. doi: 10.1079/IJT200546
Wasfi, F., Dachraoui, K., Cherni, S., Barhoumi, W., Slimane, T. B., Younsi, H., et al. (2021). Tick-borne encephalitis virus in Ixodes ricinus (Acari: Ixodidae) ticks, Tunisia. Ticks and Tick-Borne Dis. 12:101606. doi: 10.1016/j.ttbdis.2020.101606
Younsi, H., Fares, W., Cherni, S., Dachraoui, K., Barhoumi, W., Najjar, C., et al. (2019). Ixodes inopinatus and Ixodes ricinus (Acari: Ixodidae) Are Sympatric Ticks in North Africa. J. Med. Entomol. 57, 952–956. doi: 10.1093/jme/tjz216
Younsi, H., Sarih, M., Jouda, F., Godfroid, E., Gern, L., Bouattour, A., et al. (2005). Characterization of Borrelia lusitaniae isolates collected in Tunisia and Morocco. J. Clin. Microbiol. 43, 1587–1593. doi: 10.1128/JCM.43.4.1587-1593.2005
Zhioua, E., Aeschlimann, A., and Gern, L. (1994). Infection of field-collected ixodes ricinus (acari: ixodidae) larvae with Borrelia burgdorferi in Switzerland. J. Med. Entomol. 31, 763–766. doi: 10.1093/jmedent/31.5.763
Keywords: vector-borne rickettsioses, ticks, Ixodes genus, Rhipicephalus sanguineus sensu lato, molecular identification, genetic diversity, phylogeny, Tunisia
Citation: Kratou M, Belkahia H, Selmi R, Abdallah MB, Tayh G, Messadi L and Ben Said M (2025) Molecular identification, genotyping and phylogenetic analysis of Ixodes and Rhipicephalus ticks and their associated spotted fever group Rickettsia species from a single location in northern Tunisia. Front. Microbiol. 16:1644524. doi: 10.3389/fmicb.2025.1644524
Received: 10 June 2025; Accepted: 22 July 2025;
Published: 14 August 2025.
Edited by:
Hosny El-Adawy, Institut für Bakterielle Infektionen und Zoonosen, Friedrich Loeffler Institut, GermanyReviewed by:
Aman Ullah Khan, University of Veterinary and Animal Sciences, PakistanRachid Achek, University of Khemis Miliana, Algeria
Ibrahim Nabi, University of Medea, Algeria
Copyright © 2025 Kratou, Belkahia, Selmi, Abdallah, Tayh, Messadi and Ben Said. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Myriam Kratou, bWFyaWVtLmtyYXRvdUBob3RtYWlsLmNvbQ==; Mourad Ben Said, YmVuc2FpZG1vdXJhZDgzQHlhaG9vLmZy