 Emil Varghese1,2*
Emil Varghese1,2* Sarayu Krishnamoorthy1,2
Sarayu Krishnamoorthy1,2 T. K. Hredhya1Kiran Kumari3,4B. K. Bhattacharya5S. S. Kundu6Jonali Goswami6Shweta Yadav4Rama Shanker Verma7R. Ravikrishna8
T. K. Hredhya1Kiran Kumari3,4B. K. Bhattacharya5S. S. Kundu6Jonali Goswami6Shweta Yadav4Rama Shanker Verma7R. Ravikrishna8 Sachin S. Gunthe1,2*
Sachin S. Gunthe1,2*- 1EE Division, Department of Civil Engineering, Indian Institute of Technology Madras, Chennai, India
- 2Center for Atmospheric and Climate Sciences, Indian Institute of Technology Madras, Chennai, India
- 3Department of Environmental Sciences, Central University of Jammu, Samba, India
- 4Department of Environmental Sciences, Government Degree College, Billawar, India
- 5Space Applications Centre, Indian Space Research Organization, Ahmedabad, India
- 6North Eastern Space Applications Centre, Umiam, Meghalaya, India
- 7Department of Biotechnology, Indian Institute of Technology Madras, Chennai, India
- 8Department of Chemical Engineering, Indian Institute of Technology Madras, Chennai, India
Particle size is one of the important characteristics of bioaerosols that influences their fate and transport. This study investigates the characterization and size-resolved fungal bioaerosol diversity at an agricultural field in northern India during the winter season. The size-resolved bioaerosol samples were collected using a Micro-Orifice Uniform Deposition Impactor (MOUDI) in two phases: the onset of winter (phase 1) and the end of winter (phase 2), and Next-Generation Sequencing (NGS) was used to identify bioaerosol diversity up to the species level. Ascomycota was the predominant phylum in both phases, with Aspergillus penicillioides being the dominant species in phase 1 and Mycosphaerella tassiana in phase 2. The size range 1–1.8 μm exhibited higher diversity (H =3.7 and 2.0 in phases 1 and 2, respectively) and evenness (Eh = 0.9 and 0.5 in phases 1 and 2, respectively), while the 5.6–10 μm range has the highest dominance (D = 0.4 and 0.5 in phases 1 and 2, respectively). A total of 189 Operational Taxonomic Units (OTUs) were identified in phase 1 and 128 OTUs in phase 2, classified into ‘pathogenic’ and ‘beneficial/useful’ categories to study their role in ecosystem-health interactions. Species such as Aspergillus flavus, Aspergillus niger, Macrophomina phaseolina, and Penicillium citrinum were identified as affecting multiple crop hosts, highlighting the potential for multiple crop yield loss. The size range 1–8-3.2 μm had the highest number of species in phase 1, while 3.2–5.6 and 5.6–10 μm were predominant in phase 2 for plant pathogens. The 1.8–3.2 μm range also had the highest number of potential human pathogens in both phases. This study is the first to explain fungal bioaerosol diversity in an agricultural field based on size and its role in ecosystem-health interaction. These findings emphasize the importance of monitoring and managing fungal bioaerosols to protect agricultural productivity and human health.
1 Introduction
Fungi, well-known pathogenic microbes, comprise several yeast species, mushrooms, molds, and more (Hawksworth and Lücking, 2017; Taylor et al., 2014; Woo et al., 2018). It is estimated that the annual emission rate of fungal bioaerosols (such as spores and their various structural segments) from various surfaces and substrates ranges from 28 to 50 Tg a−1 (Buée et al., 2009; Elbert et al., 2007; Fröhlich-Nowoisky et al., 2009; Heald and Spracklen, 2009; Tedersoo et al., 2014). During its course in the atmosphere, fungal bioaerosols pose a significant negative impact on agriculture, ecosystem, and human health on local, regional, and global scales, by the dispersion of genetic material, causing a serious threat to humans, animals, and plants, leading to lethal infectious diseases and allergies (Varghese et al., 2024; Krishnamoorthy et al., 2020; Priyamvada et al., 2017a, 2017b; Valsan et al., 2016; Yadav et al., 2020; Fisher et al., 2012; Fröhlich-Nowoisky et al., 2016 and references therein). The ability of fungi to survive independently and the rising number of diseases caused by them have attracted the researcher’s attention to study their pathogenic effects on crops, which hampers a country’s food security (Fisher et al., 2012; Varghese et al., 2024). Also, several fungal plant diseases have been reported worldwide that could cause even 100% crop losses (Després et al., 2012). Concurrently, various fungal propagules and their toxins present in the bioaerosols have been repeatedly reported to cause a wide variety of human infections (Brown et al., 2012; Fröhlich-Nowoisky et al., 2016; Goudarzi et al., 2017; Jaenicke, 2005; Krishnamoorthy et al., 2020; Laumbach and Kipen, 2005; Priyamvada et al., 2017b).
Researchers worldwide have highlighted the emergence of human pathogenic fungal species, attributing this phenomenon to environmental stressors experienced by the fungi. These stresses, alongside the fungi’s ability to develop resistance to commonly used fungicides, have resulted in fungal species that are increasingly pathogenic. These fungi can overcome both host defence mechanisms and the drugs used for treatment (Fisher et al., 2012; Rokas, 2022). This growing threat has been emphasized by Pfaller (2012), who warned of the significant risks posed by these emerging fungal pathogens, especially to high-risk patients such as those undergoing treatment, immunocompromised individuals, and patients on immunosuppressive therapies. The most common fungal allergens belong to Aspergillus, Cladosporium, Alternaria, and Penicillium (Priyamvada et al., 2017a,b; Fennelly et al., 2017). Hence, to detect and monitor the infectious diseases caused by microbes, two data sources, ‘ProMED’ (the Program for Monitoring Emerging Diseases) and ‘HealthMap’, have been developed globally (Fisher et al., 2012). It has been reported that the most significant number of animal-infecting and plant-infecting fungal alerts have been reported in the United States and the Indian subcontinent, respectively. Over India, during the last two decades, approximately 48% of the alerts reported by ProMED were of fungal related diseases which affected nearly 39 varieties of crops, and Puccinia striiformis and Pyricularia oryzae affecting wheat and rice hosts, respectively, are the species that were reported multiple times (Yadav et al., 2020).
Many attempts have been made worldwide to address the aerosolization properties, dispersion, deposition, and the adverse implications caused by the fungal propagules on the ecosystem health and climate (Calhim et al., 2018; Elbert et al., 2007; Fröhlich-Nowoisky et al., 2009; Krishnamoorthy et al., 2020; Priyamvada et al., 2017a, 2017b; Valsan et al., 2016; Woo et al., 2018). The particle size, one of the most important characteristics of the fungal bioaerosols, plays a vital role in fungal fate and transport, deposition in the respiratory system, settling and deposition on the Earth’s surface, resuspension to air, penetration into buildings, and pathogenicity potential to cause diseases in plants (Gat et al., 2021; Tanaka et al., 2020; Thomas, 2013; Wang and Lin, 2012; Yamamoto et al., 2012, 2014). Fungi have unique morphological characteristics, which result in effective aerosolisation and long-distance transport of the spores (Yadav et al., 2020). Fröhlich-Nowoisky et al. (2009) have reported that more human pathogens and allergens are found in the fine fraction (< 3 μm) and plant pathogens in the coarse fraction. Therefore, the size and shape-dependent understanding and behavior of the fungal bioaerosols will not only help to delineate their impacts from other types of bioaerosols but also improve our understanding of the specificity of the role of fungal bioaerosols in ecosystem health (Wang and Lin, 2012).
As the threat caused by fungi to plant and animal biodiversity has been increasing in recent years, several studies are now being carried out worldwide to understand the pathogenic virulence of fungal bioaerosols on crops, plants, animals, insects, and human health (Calhim et al., 2018; Elbert et al., 2007; Fisher et al., 2012; Fröhlich-Nowoisky et al., 2009; Krishnamoorthy et al., 2020; Priyamvada et al., 2017a, 2017b; Valsan et al., 2016; Woo et al., 2018). Advanced studies involving next-generation sequencing have replaced the culture-dependent studies to explore the finer details of the pathogenicity of the bioaerosols (Baldrian et al., 2022; Nilsson et al., 2019; Palmero et al., 2011; Shelton et al., 2002; Tedersoo et al., 2014; Woo et al., 2018). Various tools like FUNGuild have also been introduced to taxonomically parse fungal OTUs by ecological guild independent of sequencing platform or analysis pipeline (Nguyen et al., 2016).
In this study, we investigate the size-resolved diversity of the fungal bioaerosols present in an agricultural field in north India, during the winter season. Further, their ‘pathogenic’ and ‘beneficial/useful’ roles on the plants, humans, and the environment have been studied in detail by analyzing and reviewing the available literature, emphasizing their impact on ecosystem health based on size-resolved diversity. Our analysis generates new insights about the size-resolved diversity of bioaerosols over an agricultural field and their ecosystem-health implications.
2 Materials and methods
2.1 Study site and bioaerosol sampling
The study site is a wheat crop field (Gurdaspur, Punjab; 32°2′21” N and 75°23′11″E) in India, which is surrounded by horticultural fields growing vegetables and fruits. The size-resolved air samples were collected on glass microfiber filter papers (47 mm diameter, Whatman grade GF/C) using a 10-stage Micro-Orifice Uniform Deposition Impactor MOUDI II 120R, TSI Inc., USA. MOUDI has an inlet with a nominal cut-off of 18 μm and ten different stages for collecting uniformly deposited particles with a nominal cut-off of 10, 5.6, 3.2, 1.8, 1.0, 0.56, 0.32, 0.18, 0.10, and 0.056 μm when operated at a flow rate of 30 lpm. The inlet of the impactor was positioned at a height of 2 meters above ground level, corresponding to the typical human breathing level. The sampling was carried out for 70 h each during the winter season in India in two phases - phase 1 to cover the onset of the winter season (initial two-leaf stage of wheat crop; December 2019) and phase 2 to cover the end of the winter season (harvest period of wheat crop; March 2020). During the winter season, daytime temperatures average around 10 °C, while nighttime temperatures drop to a chilly 2 °C. The prevailing wind direction was predominantly northwesterly, bringing in continental air masses that were dry and characterized by low relative humidity - conditions favorable for the long-distance transport and atmospheric persistence of dust particles and bioaerosols. Another set of samples was collected using a 2-stage sampler as described by Valsan et al. (2015) on the nucleopore membrane filters of 25 mm diameter with a pore size of 0.2 and 5 μm to observe the morphological details of the bioaerosols.
2.2 DNA isolation and sequencing
The fungal DNA was isolated from each filter paper using the ZR Quick-DNA™ Fungal/Bacterial Miniprep Kit (Zymo Research, USA). The Internal Transcribed Region (ITR) of the extracted DNA was amplified using the primers forward - GCATCGATGAAGAACGCAGC and reverse – TCCTCCGCTTATTGATATGC using the PCR (Polymerase chain reaction) plate of the thermocycler (SureCycler 8,800, Agilent Technologies Inc., USA). The PCR amplicons were then sequenced using the Next-Generation Sequencing (NGS) method in the Illumina MiSeq platform 2 × 300 bp (forward and reverse sequencing) chemistry to generate ≈1 lakh reads per sample. The DNA sequences obtained from the Illumina MiSeq platform were analyzed using the open-source bioinformatics pipeline QIIME™ 2 (Quantitative Insights into Microbial Ecology) (Caporaso et al., 2010) and the Operational Taxonomic Units (OTUs) were picked using the UCLUST algorithm from the UNITE database (version 7.2). The detailed DNA isolation, NGS, and analysis protocol is mentioned in Varghese et al. (2024).
2.3 Biodiversity analysis
The size-resolved fungal diversity of the OTUs identified at the species level is visualized in terms of alpha and beta diversity. The alpha diversity (diversity within the ecosystem) is observed in terms of various parameters such as species richness, relative abundance, Shannon-Wiener diversity index (H) (Yadav et al., 2020), Shannon’s evenness of equitability (Eh), and Simpson’s dominance index (D). The calculated alpha diversity indices are plotted using Python libraries such as “pandas,” “seaborn,” “numpy,” and “matplotlib” in the open-source web application Jupyter Notebook (V.6.0.3). The beta diversity (the diversity between the various stages) is expressed using the Principal Coordinate Analysis (PCoA). For the obtained OTUs, the eigenvalues for the PCoA are estimated with Bray-Curtis distance, and the values are aligned in a two-dimensional space using the packages “vegan” and “ape” of the R programming language [version 4.1.0 (2021-05-18)].
2.4 Morphological characterization using SEM analysis
The filter paper samples collected for morphological characterization were cut into 1 cm2 pieces, sputter-coated with gold, and imaged using a Hitachi S-4800 Scanning Electron Microscope (SEM) equipped with EDX/EDS at the Chemical Engineering Department, Indian Institute of Technology Madras, Chennai, India. Based on the EDX results, particles with a relative contribution of Carbon (C), Oxygen (O), and Nitrogen (N) greater than 90% were classified as bioaerosols (Valsan et al., 2015), and the images were captured.
3 Results
3.1 Diversity of fungal bioaerosols
During the DNA sequencing, fungal bioaerosols were identified in five size ranges: 1–1.8 μm (Stage 5 of MOUDI), 1.8–3.2 μm (Stage 4 of MOUDI), 3.2–5.6 μm (Stage 3 of MOUDI), 5.6–10 μm (Stage 2 of MOUDI), and 10–18 μm (Stage 1 of MOUDI). The cumulative number of DNA sequences observed from the samples was 684,827 in phase 1 (onset of winter) and 501,672 in phase 2 (end of winter). In phase 1, 24.2% of the total DNA sequences were identified up to the species level, identifying 189 species-level OTUs. In phase 2, 21.8% of the DNA sequences were identified up to the species level, resulting in 128 OTUs. In phase 1, Aspergillus penicillioides was the most dominant species, constituting 37.1% of the species-level OTUs, followed by Aspergillus flavus at 8.7%. Other significant species observed in phase 1 included Bipolaris melinidis (5.9%), Schizophyllum commune (5.6%), Curvularia lunata (5.4%), Rhizopus arrhizus (3.9%), Ustilaginoidea virens (3.6%), Periconia digitata (3.0%), Coprinopsis laanii (2.4%), and Curvularia hawaiiensis (2.3%). In phase 2, Mycosphaerella tassiana predominated, making up 66.4% of the sequences, followed by Aspergillus penicillioides (9.6%), Tilletiopsis washingtonensis (7.9%), Aspergillus flavus (2.3%), Ruinenia clavata (2.1%), Blumeria graminis (2.0%), and Curvularia lunata (1.1%).
The species-level identified OTUs belonged to five major phyla: Ascomycota, Basidiomycota, Mucoromycota, Mortierellomycota, and Chytridiomycota. In both phases, Ascomycota was predominant, with 81.4% in phase 1 and 86.0% in phase 2. This was followed by Basidiomycota (14.5% in phase 1 and 13.8% in phase 2), Mucoromycota (4.02% in Phase 1 and 0.13% in phase 2), Mortierellomycota (0.012% in phase 1 and 0.008% in phase 2), and Chytridiomycota (0.001% in both phase 1 and phase 2). The family-level taxonomic classification of the species-level identified OTUs comprised 104 families in phase 1 and 70 families in phase 2. In phase 1, Aspergillaceae dominated, constituting 52.3% of the species-level OTUs, and in phase 2, Mycosphaerellaceae was predominant, making up 66.4%. Figure 1 shows the SEM images of the fungal bioaerosols collected from the agricultural field during phases 1 and 2. Based on morphology, spores from Ascomycota and Basidiomycota were identified.

Figure 1. SEM images confirming the presence of fungal bioaerosols covering a wide size range, explaining the broad fungal size distribution.
In phase 1, out of the 104 family OTUs, 21 OTUs were observed in all five size ranges, and in Phase 2, 23 out of 70 family-level OTUs were present in all five size ranges. The highest number of family-level OTUs was observed in the size range of 1.8–3.2 μm in both phases (77 OTUs in phase 1 and 45 OTUs in phase 2), followed by 3.2–5.6 μm in phase 1 (61 OTUs). In phase 2, the size ranges 3.2–5.6 μm and 5.6–10 μm had 44 OTUs each. Table 1 shows the size-resolved diversity of species-level OTUs, expressed as diversity indices. The phase 1 has a higher diversity (H = 2.7) than phase 2 (H = 1.5). This was due to the higher richness and abundance in phase 1 (189 OTUs and 165,493 DNA sequences) compared to phase 2 (128 OTUs and 109,161 DNA sequences). The evenness is higher in phase 1 (Eh = 0.5), and dominance is higher in phase 2 (D = 0.5). The size range 1–1.8 μm has higher diversity (H = 3.7 and 2.0 in phases 1 and 2, respectively) and evenness (Eh = 0.9 and 0.5 in phases 1 and 2, respectively) in both the phases, while the size range 5.6–10 μm has the highest dominance (D = 0.4 and 0.5 in phases 1 and 2, respectively).
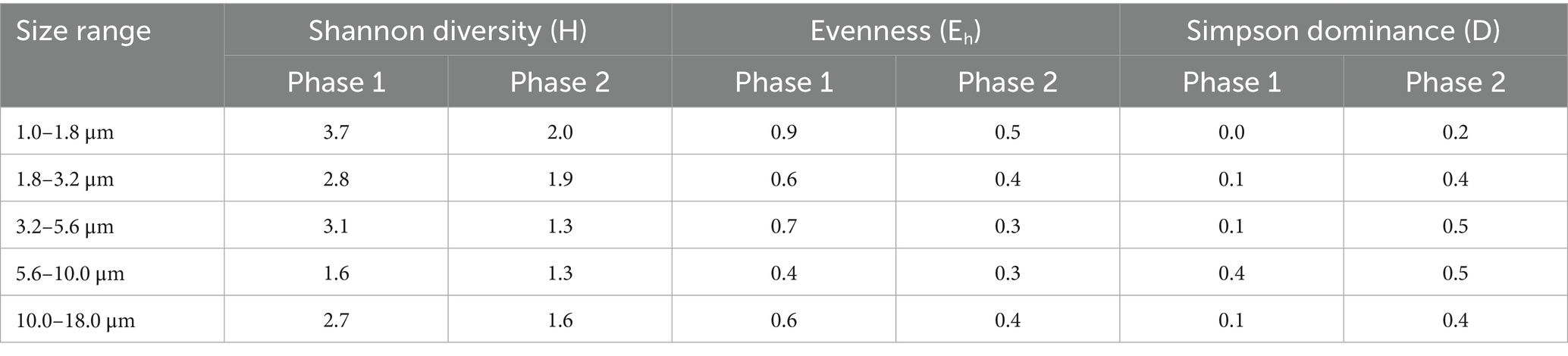
Table 1. Size-resolved diversity of fungal bioaerosols, identified at the species level, during two phases.
3.2 Fungal bioaerosol characterization and their size-resolved diversity
The species-level identified fungal OTUs are classified into two categories: ‘pathogenic’ and ‘beneficial/useful’ based on their ecosystem-health implications. Supplementary Tables S1, S2 list various species identified as ‘pathogenic’ and ‘beneficial/useful’, respectively, based on the literature review. ‘Pathogenic’ refers to the fungal species that cause disease in various hosts such as crops, plants, insects, nematodes and humans, while the ‘beneficial/useful’ refers to the fungal species beneficial in biotechnology, industry, medicine, as well as saprophytic/environmental strains or edible mushrooms. The following sections discuss the size-resolved diversity of these categories.
3.2.1 Size-resolved diversity of crop pathogens
Figure 2 illustrates the size-resolved diversity of various crop pathogens, classified into cereals, pulses, cash crops, fruits, vegetables, and spices based on their potential hosts. We observed 21 potential cereal pathogens, 7 pulse pathogens, 6 cash crop pathogens, 15 fruit pathogens, 7 vegetable pathogens, and 2 spice pathogens (Figure 2A). The number of DNA sequences for cereals was the least in the 1–1.8 μm size range in both phases. Pulses showed a high number of species in the 5.6–10 μm size range in phase 1 and 3.2–5.6 μm in phase 2. Cash crop species were mainly observed in size ranges greater than 1.8 μm in phase 1 and greater than 3.2 μm in phase 2. For fruits, 15 species were identified, with 12 species in the 1.8–3.2 μm size range and 10 species in the 3.2–5.6 μm size range. Vegetables had the maximum number of species in the 5.6–10 μm size range in phase 1, while the 3.2–5.6 μm size range dominated in phase 2. Both species of spices pathogens were present in the 5.6–10 μm size range in phase 1. Figure 2B shows that cereals exhibited the highest diversity, high evenness, and low dominance among crop pathogens. In phase 1, cereal pathogens showed high diversity in the 1–1.8 μm size range (H=1.9), while in phase 2, the 5.6–10 μm and 10–18 μm size ranges were highly diverse (H=1.7). Pulse pathogens had low diversity in both phases, with the maximum diversity observed in the 10–18 μm size range (H=0.3). Cash crops pathogens displayed moderate diversity and evenness across all size ranges. Fruits pathogens showed unique intra-community structures in each size range during phase 1, but similar diversity indices across multiple size ranges in phase 2. Vegetable pathogens had highly variable diversity, with the highest diversity in the 3.2–5.6 μm size range (H=1.3) in phase 1 and (H=0.9) in phase 2. Spice pathogens exhibited relatively low diversity in both phases due to the presence of only a few species identified in this category.
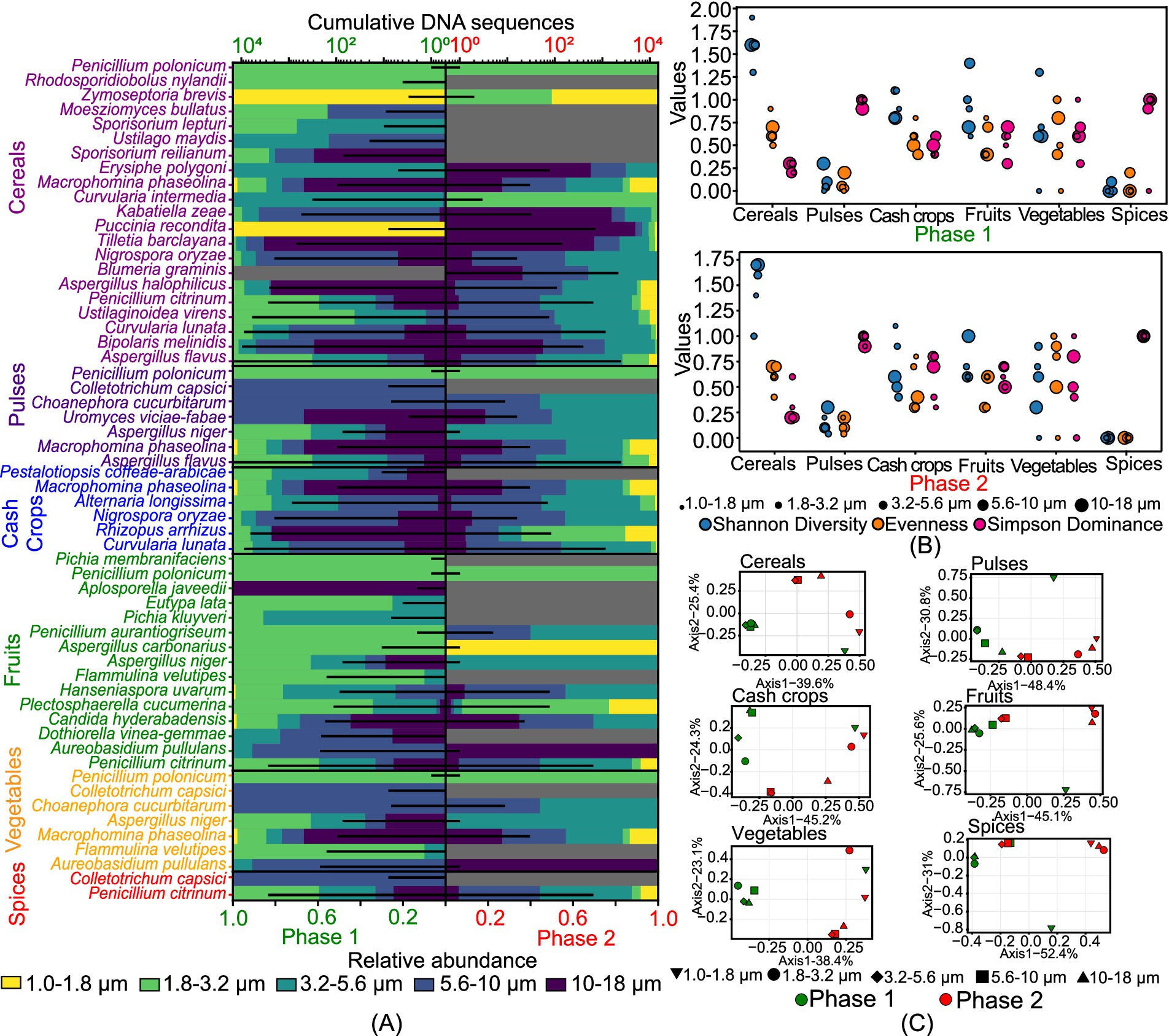
Figure 2. Size-resolved diversity of species identified as crop pathogens during phase 1 and phase 2: (A) cumulative DNA sequences and the size-resolved relative abundance; (B) size-resolved diversity explained through the various diversity indices like Shannon diversity index, evenness, and Simpson’s dominance; and (C) inter-size range diversity.
Figure 2C presents the Principal Coordinate Analysis (PCoA), revealing distinct inter-community structures among pathogenic fungi across different crop categories. For cereal pathogens, phase 1 showed less correlation in the 1–1.8 μm size range, while phase 2 had overlapping communities in the 3.2–5.6 μm and 5.6–10 μm ranges. Pulse pathogens exhibited overlapping species in the 1.8–3.2 μm and 3.2–5.6 μm ranges in phase 1, with unique compositions in the 1.0–1.8 μm range. In phase 2, unique compositions were specific to each size range, with similar communities in the 1.0–1.8 μm, 1.8–3.2 μm, and 10–18 μm ranges. Cash crop pathogens had size-specific compositions, except for overlapping communities in the 5.6–10 μm and 10–18 μm ranges in phase 1, and 3.2–5.6 μm and 5.6–10 μm ranges in phase 2. Fruit pathogens showed similar structures across all size ranges in phase 1, except for 10–18 μm, and overlapping species in the 3.2–5.6 μm and 5.6–10 μm ranges in phase 2. Vegetable pathogens had similar populations across all size ranges in phase 1 but diverse populations with overlapping species in the 3.2–5.6 μm and 5.6–10 μm ranges in phase 2. Spice pathogens exhibited distinct compositions in the 5.6–10 μm range in phase 1 and similar structures in the 3.2–5.6 μm and 5.6–10 μm ranges in phase 2.
3.2.2 Size-resolved diversity of plant pathogens
Figure 3 identifies 38 plant pathogens (excluding crops), with Bipolaris melinidis being the most predominant in phase 1 and Mycosphaerella tassiana dominant in both phases. In phase 1, 24 species were present in the 1.8–3.2 μm size range, while in phase 2, 17 species each were observed in the 3.2–5.6 μm and 5.6–10 μm size ranges, indicating a shift from smaller to larger size ranges. The Shannon diversity index (H) shows highly variable diversity in phase 1 compared to phase 2, with moderate evenness (Figure. 3B). Figure 3C illustrates the diverse nature of plant pathogens across different size ranges in both phases, except for overlapping communities in the 3.2–5.6 μm and 5.6–10 μm ranges in phase 2. The 1.8–3.2 μm size range in phase 1 and the 1–1.8 μm size range in phase 2 were grouped, indicating similar population structures.
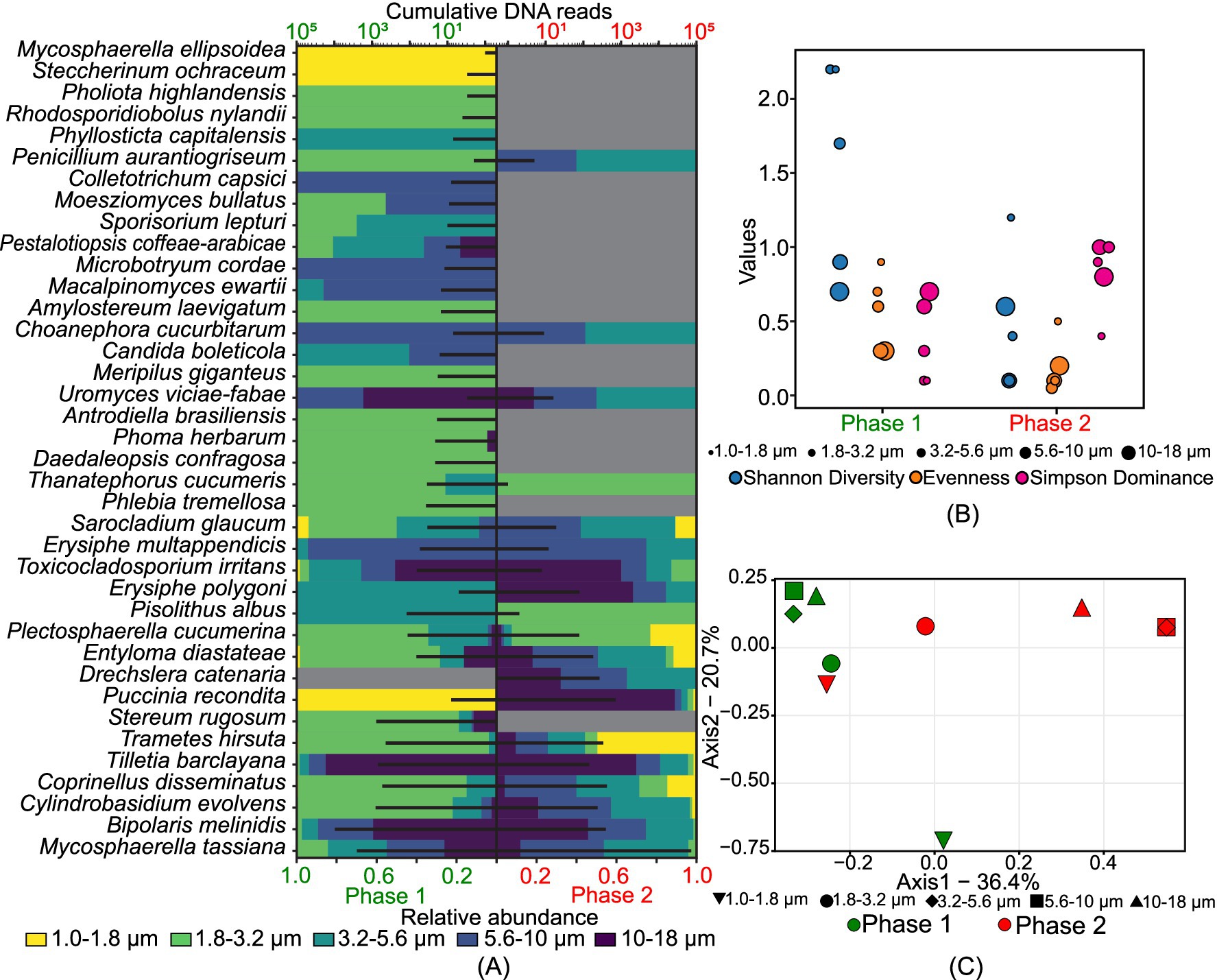
Figure 3. Size-resolved diversity of species identified as plant pathogens during phase 1 and phase 2: (A) cumulative DNA sequences and the size-resolved relative abundance; (B) size-resolved diversity explained through the various diversity indices like Shannon diversity index, evenness, and Simpson’s dominance; and (C) inter-size range diversity.
3.2.3 Size-resolved diversity of insects and nematodes pathogens and human pathogens
A greater diversity of insects and nematodes pathogens was observed in phase 1 compared to phase 2. Figures 4A,B,E shows the size-resolved diversity of insects and nematode pathogens. Specifically, the size ranges of 1.8–3.2 μm and 3.2–5.6 μm in phase 1 had more species, with 7 and 6 species, respectively. In phase 2, the size range of 5.6–10 μm had the most species, with 3. Diversity was higher in all size ranges during Phase 1, while phase 2 had lower diversity. Evenness values, which indicate how evenly species are distributed, were higher in Phase 1 (0.1 to 0.9) compared to phase 2 (0.05 to 0.7). Dominance, or the presence of a few species being very common, was higher in phase 2, reaching a maximum value of 1. Principal Coordinate Analysis (PCoA) showed distinct population structures in both phases, except for the size ranges of 3.2–5.6 μm and 5.6–10 μm, which had overlapping communities. This indicates that phase 1 had a higher diversity of size-specific species than phase 2, despite some overlap in certain size ranges.
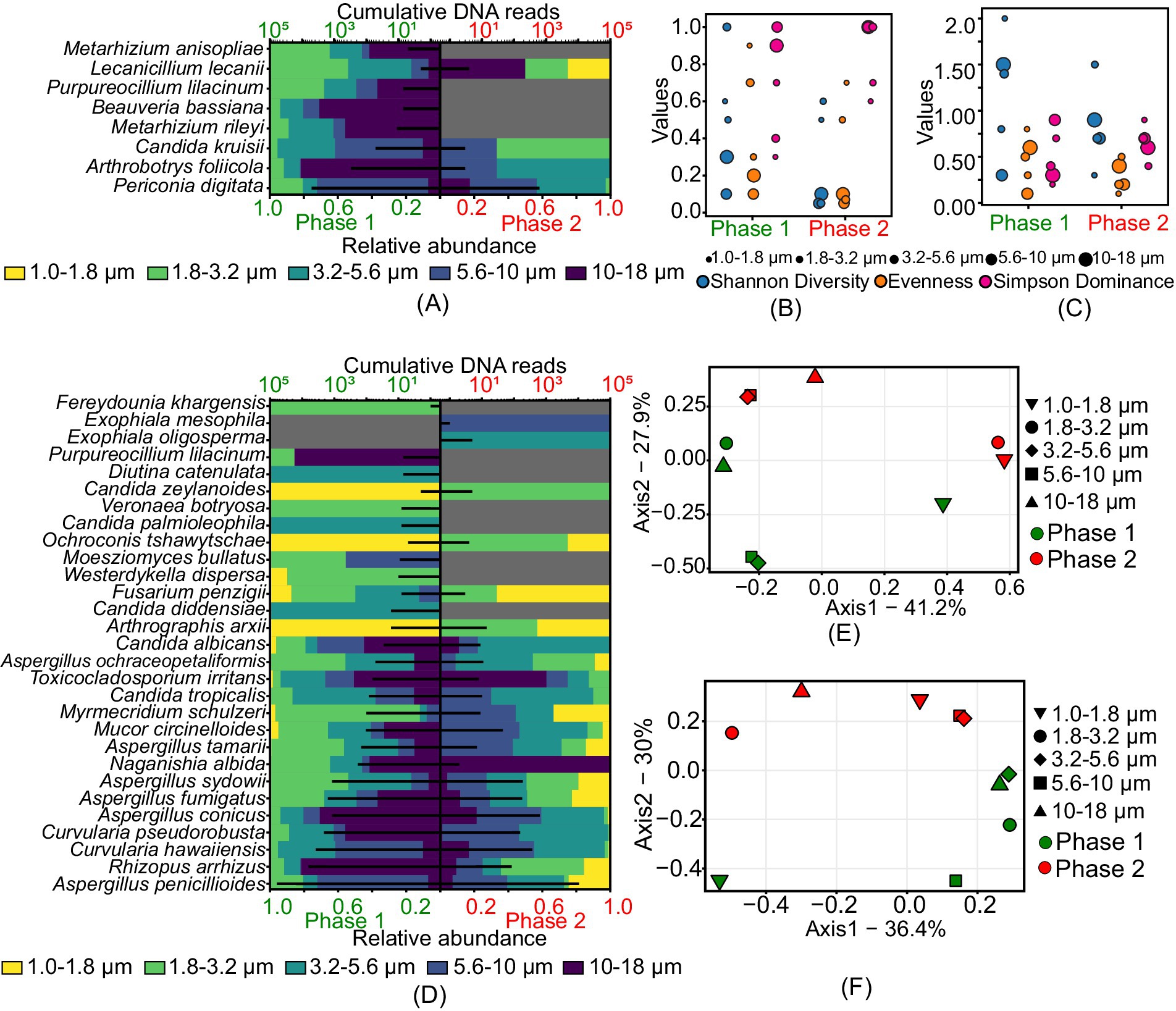
Figure 4. Size-resolved diversity of species identified as insects and nematodes and human pathogens during phase 1 and phase 2: (A) cumulative DNA sequences and the size-resolved relative abundance of insects and nematodes pathogens; (B) size-resolved diversity of insects and nematodes pathogens explained through the various diversity indices like Shannon diversity index, evenness, and Simpson’s dominance; (C) size-resolved diversity of human pathogens explained through the various diversity indices like Shannon diversity index, evenness, and Simpson’s dominance; (D) cumulative DNA sequences and the size-resolved relative abundance of human pathogens; (E) inter-size range diversity of insects and nematodes pathogens; and (F) inter-size range diversity of human pathogens.
Figures 4D,C,F illustrates the type and size-resolved diversity of human pathogens. Aspergillus penicillioides was common in both phases. Unique to phase 1 were species like Candida diddensiae, Candida palmioleophila, Diutina catenulata, Purpureocillium lilacinum, Veronaea botryosa, and Westerdykella dispersa, while Exophiala mesophila and Exophiala oligosperma were specific to phase 2. The size range of 1.8–3.2 μm had the highest number of species in both phases, with a total of 21 species in phase 1 and 16 in phase 2. In phase 1, the Shannon diversity index (H) ranged from 0.3 to 2, evenness from 0.1 to 0.8, and dominance (D) from 0.2 to 0.9. In phase 2, the diversity index (H) ranged from 0.3 to 1.5, evenness from 0.1 to 0.5, and dominance (D) from 0.4 to 0.9. Principal Coordinates Analysis (PCoA) showed highly diverse communities across all size ranges in both phases, except for overlapping structures in the 3.2–5.6 μm and 5.6–10 μm ranges in phase 2. This analysis indicates that phase 1 had a higher diversity of size-specific species than phase 2.
3.2.4 Size-resolved diversity of beneficial/useful fungi
The study identified 176 beneficial fungal species, including 77 saprophytic/environmental strains, 20 biotechnological and industrial strains, 8 medicinal fungi, and 5 edible mushrooms (Supplementary Figure S1). Among the saprophytic/environmental strains, Aspergillus penicillioides was the most dominant species in both sampling phases, followed by Coprinopsis laani in phase 1 and Tilletiopsis washingtonensis in phase 2. Figure 5 represents the size-resolved diversity of beneficial/useful fungi. In phase 1, the size ranges of 1.8–3.2 μm and 3.2–5.6 μm had the highest number of species, with 48 and 41 OTUs, respectively. In phase 2, the 1.0–1.8 μm range contained the most species (29), followed closely by the 1.8–3.2 μm and 3.2–5.6 μm ranges, each with 27 species. This indicates a shift in the distribution of species across different size ranges between the two phases.
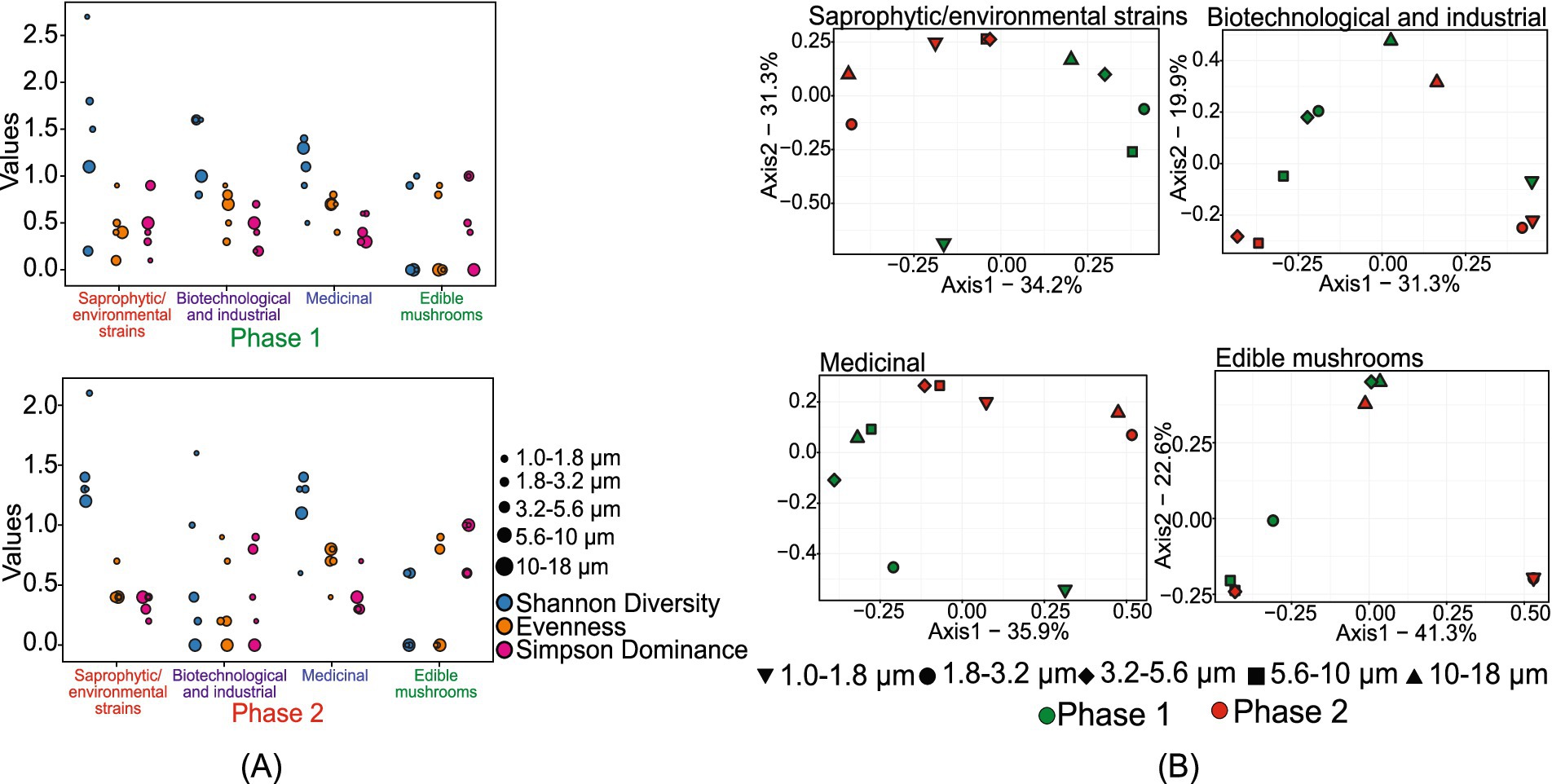
Figure 5. Size-resolved diversity of species identified as beneficial/useful species during phase 1 and phase 2: (A) size-resolved diversity explained through the various diversity indices like Shannon diversity index, evenness, and Simpson’s dominance; and (B) inter-size range diversity.
Shannon diversity analysis further highlighted these variations. In phase 1, the 1.0–1.8 μm size range showed a highly diverse population with a 𝐻 value of 2.7. In phase 2, the highest diversity was observed in the 1.8–3.2 μm size range, with a 𝐻 value of 2.1. The diversity indices suggested that both phases exhibited high evenness and low dominance in the 1.0–1.8 μm and 1.8–3.2 μm size ranges, respectively. The unique population diversity within each size range showed distinct community structures without significant overlap, except in the 3.2–5.6 μm and 5.6–10 μm size ranges of phase 2.
For biotechnological and industrial fungi, phase 1 exhibited a high number of species in the 1.8–3.2 μm and 3.2–5.6 μm size ranges. In contrast, in phase 2, the 1.0–1.8 μm size range showed the highest number of species. Some species were uniquely observed in phase 1, including Beauveria bassiana, Kluyveromyces lactis, Metarhizium rileyi, Myceliophthora thermophila, Pichia kluyveri, Pichia membranifaciens, Pseudozyma hubeiensis, Thermomyces dupontii, Trichoderma reesei, and Virgaria nigra. The Shannon diversity index provided further insights into the fungal diversity observed in the samples. Phase 1 exhibited higher diversity compared to phase 2, with a maximum 𝐻 value of 1.6 at the size ranges of 1.0–1.8 μm, 1.8–3.2 μm, and 5.6–10 μm. In phase 2, the 1.0–1.8 μm size range also showed a maximum H value of 1.6. This higher diversity in phase 1 suggests a more varied fungal community during this period. Additionally, the maximum evenness value of 0.9 was observed in the 1.0–1.8 μm size range in both phases, indicating a well-distributed fungal population within these size ranges. High intercommunity diversity among the different size ranges in both phases indicated distinct and separate fungal communities within each size range, except for the 1.0–1.8 μm size range in phase 1 and the 1.0–1.8 μm and 1.8–3.2 μm size ranges in phase 2, which grouped together. This grouping suggests a nearly similar fungal community structure within these specific size ranges.
The size-fractioned assessment of medicinal fungi showed that the 1.8–3.2 μm size range in phase 1 samples contained the highest number of species. In phase 2, 7 species were observed in the 3.2–5.6 μm and 5.6–10 μm size ranges. The diversity analysis indicated that phase 2 exhibited higher intra-community diversity than phase 1 samples. The evenness of fungal distribution in both phases was similar, except for the 1.0–1.8 μm and 1.8–3.2 μm size ranges. The dominance (D) value was comparatively higher at the 1.0–1.8 μm size range in phase 2, with a value of 0.7. In phase 1, a dominance (D) of 0.6 was observed in the 1.0–1.8 μm and 1.8–3.2 μm size ranges, consistent with the observed diversity values.
The size-fractioned characterization of edible mushrooms indicated no observable sequences in the 10–18 μm size range in phase 1. The diversity analysis showed the dominance of a single species across various size ranges in both phases. Furthermore, the analysis suggested that the 1.8–3.2 μm size range of phase 1 exhibited a diverse community structure distinct from other size ranges. However, due to the limited number of OTUs observed in phase 2, the diversity characteristics for this phase cannot be reliably assessed as an accurate representation of the broader fungal community.
4 Discussion
Changes in the cell size and shape of microorganisms as they adapt to environmental conditions significantly impact the presence of fungal aerobiomes across a wide size range. We characterized fungal bioaerosols using SEM imaging and discovered a diverse range of fungal propagules within the 3–20 μm size range. These propagules include spores, fungal fragments, clusters of spores, mycelium, and spore-dust agglomerates (Krishnamoorthy et al., 2020; Lacey and Goodfellow, 1975; Tong and Bruce Lighthart, 2000). Our study also revealed that fungal OTUs was present in various size ranges varying from 1 to 18 μm, with the highest diversity observed in the 1–1.8 μm size range. This increased diversity in smaller size fractions may be attributed to the presence of smaller basidiospores in the fungal bioaerosols samples from the agricultural field.
We identified several crop pathogens that can cause epiphytic or endophytic infections, including foliar diseases, powdery mildew, cankers, and more. Presence of pathogens like Aspergillus flavus and Aspergillus niger indicate a strong probability of postharvest infections and other diseases in pulses. The presence of Uromyces viciae-fabae and Puccinia recondita, Basidiomycota members, suggests a risk of faba-bean rust and wheat rust, respectively, in the sampling site or nearby locations (Varghese et al., 2024). Rhizopus arrhizus is known for causing barn rot in tobacco (Chen et al., 2020). Flammulina velutipes can affect mulberry, Chinese hackberries, and persimmon trees (Fischer and Garcia, 2015). Choanephora cucurbitarum can cause fruit and blossom rot in cucurbits, infect okra, and cause rot in teasel gourd. Penicillium citrinum and Colletotrichum capsici – spices fungi – suggest potential opportunistic infections that could reduce yield (Ragavendran et al., 2019; Saxena et al., 2016). The literature review also revealed that the same fungal pathogens can affect multiple crops. For example, Aspergillus flavus can infect both cereals and pulses, while Aspergillus niger targets pulses, fruits, and vegetables. Other species, such as Macrophomina phaseolina and Penicillium citrinum, were found to infect multiple crop types, including cereals, pulses, cash crops, and spices. Similar observations were made by Couch et al. (2005) regarding the ability of Magnaporthe oryzae to infect multiple crops.
The presence of certain pathogenic fungal species indicated that the local plant species in the sampling site were susceptible to a variety of fungal infections, including white rot (Slippers et al., 2003), wood rot, leaf spots (Limtong et al., 2020), blight, melting out, root rot (Carlucci et al., 2012), rotten trunks and leaves (Novaković et al., 2018), stem and leaf rot (Alfenas et al., 2018; Pornsuriya et al., 2017), leaf blight (Saxena et al., 2016), smut disease (Kellner et al., 2011), powdery mildew (Cowger and Brown, 2019), jelly rot (Yeo et al., 2008), cankers, fruit rot, collar rot (Rivedal et al., 2020), leaf rust, damping off, wire stem, general plant disease, leaf disease, and opportunistic infections (Okolo et al., 2015; Stoll et al., 2005). These findings were consistent with previous studies by Anonymous (2017), Fisher et al. (2012), Savary et al. (2012), and Simion (2018), which reported that the phytopathogenic fungi significantly reduce global crop yields, contaminate livestock feed, and cause various plant infections.
Several fungal pathogens identified could play a crucial role in controlling insect pests, including Culex mosquitoes, nematodes, and malarial mosquitoes (Castillo Lopez et al., 2014; Davies et al., 2015; Jiang et al., 2019; Khan et al., 2012; McKinnon et al., 2018; Pedrini et al., 2013; Ragavendran et al., 2019). As highlighted by Brown et al. (2012), Fisher et al. (2012, 2018), and Rhodes (2019), certain fungal pathogens are capable of causing severe diseases, particularly in immunocompromised individuals. The potential health impacts are wide-ranging and include opportunistic infections (Bezerra et al., 2017; Rudramurthy et al., 2019), neonatal sepsis (Okolo et al., 2015), and nosocomial infections such as candidiasis (Kim and Singh, 2000). Additional severe health issues identified include candidemia, infections associated with intravenous catheters (Yamin et al., 2021), Hickman catheter-associated fungemia (Whitby et al., 1996), allergies (Gunasekaran, 2017), and infections in immunocompromised individuals (Benedict and Mody, 2016). Other conditions include nail infections, infections in patients with ketoacidosis, cutaneous lesions, and more severe conditions such as golden tongue, infections in transplant patients, and angio-invasive infections. These fungal pathogens also pose a risk to animals, which can, in turn, enhance the likelihood of human infections and potential epidemic outbreaks (Gnat et al., 2021; Köhler et al., 2015; WHO, 2018). The results of the size-resolved diversity analysis aligns with the findings of Fröhlich-Nowoisky et al. (2016), Guarnieri and Balmes (2014), Hofmann (2011), and Nazaroff (2016) who noted that human pathogenic fungal bioaerosols are predominantly within the size varying from 1.8 to 10 μm. The size varying from 1.8 to 5.6 μm are particularly significant for the diverse community structure of human pathogens (Guarnieri and Balmes, 2014; Krishnamoorthy et al., 2020).
The saprophytic/environmental strains included a diverse array of species, including aquatic fungi (Jooste et al., 1990), marine fungi (Wang et al., 2017), wood-decaying fungi, xerotolerant fungi (Hirooka et al., 2016), soil fungi, environmental yeasts (Li et al., 2021), rare environmental mushrooms, fungi that thrive on minerals and mineral-rich rocks (Jiang et al., 2018), extremotolerant fungi, weeping widow mushrooms (Roberts and Evans, 2011), and dung fungi and mushrooms. These environmental fungi and saprophytes are ubiquitous and play crucial roles in maintaining the carbon-nitrogen cycle, the decomposition of organic matter, and various other ecological processes and cycles (Dagenais and Keller, 2009). The unique population diversity within each size range, showing distinct community structures without significant overlap, except in the 3.2–5.6 μm and 5.6–10 μm size ranges of phase 2, suggests a less distinct separation of fungal communities in these specific size ranges during Phase 2, highlighting the diverse nature of saprophytic/environmental strains present in the environment.
Among the Biotechnologically and industrially important fungi, Penicillium polonicum stood out due to its extensive applications in producing penicillic acid, verrucosidin, patulin, anacine, 3-methoxyviridicatin, and glycopeptides (Valente et al., 2021). Penicillic acid is known for its antibiotic properties, while verrucosidin and patulin are mycotoxins with potential uses in biocontrol and pharmaceuticals. Anacine is an alkaloid with potential therapeutic applications, and 3-methoxyviridicatin and glycopeptides are compounds with antimicrobial properties that can be used in medicine and agriculture. Beauveria bassiana is known for its use as a bio-pesticide due to its pathogenicity against a wide range of insect pests. Kluyveromyces lactis is utilized in the dairy industry for the production of lactose-free milk and other dairy products. Metarhizium rileyi is another important bio-pesticide, especially effective against caterpillar pests. Myceliophthora thermophila and Thermomyces dupontii are thermophilic fungi used in the production of industrial enzymes that can withstand high temperatures, making them valuable in various industrial processes. Pichia kluyveri and Pichia membranifaciens are known for their roles in fermentation, particularly in the production of bioethanol and other biofuels. Trichoderma reesei is widely used in the production of cellulases and hemicellulases, enzymes essential for the biofuel industry. Virgaria nigra, although less well-known, has potential applications in bioremediation and the production of secondary metabolites with pharmaceutical properties.
In the case of medicinal fungi, Lenzites betulina, known for its anticancer and antimicrobial properties (Liu et al., 2014), was uniquely observed in phase 1. In contrast, Ganoderma lucidum, which has various medicinal benefits, including stabilizing blood glucose levels, modulating the immune system, providing hepatoprotection, and exhibiting bacteriostatic properties (Unlu et al., 2016), was specifically observed in phase 2. Among the medicinal fungi identified in the bioaerosols, Trametes versicolor stood out as a particularly potent strain due to its extensive medical and immunological applications. This species is known for its ability to activate the reticuloendothelial system, modulate cytokine production (specifically enhancing INF and IL-2), improve the viability of dendritic cells, promote T-cell maturation, enhance the activity of natural killer cells, and stimulate antibody production (Knežević et al., 2018). Furthermore, Trametes versicolor has demonstrated antitumor and anticancer effects, making it a significant finding in this study. Size-resolved diversity analysis revealed that the communities present in all size ranges exhibited unique, non-overlapping populations in medicinal fungi. This suggests that the medicinal fungi observed in the bioaerosols had a more diverse community structure than other functional categories of fungal bioaerosols.
5 Conclusion
In conclusion, this study provides the first comprehensive, size-resolved characterization of fungal bioaerosols over an agricultural ecosystem in India during the winter season, highlighting clear seasonal and size-dependent variations in diversity, abundance, and ecological function. The observed enrichment of fungal diversity in smaller particle size ranges (1–1.8 μm) suggests that fine fungal spores contribute significantly to atmospheric biodiversity and may influence both local and regional climate - ecosystem interactions. Moreover, the coexistence of pathogenic and beneficial fungal taxa across size fractions underscores the complex interplay between agricultural practices, environmental stressors, and fungal adaptation mechanisms driven by ongoing climatic changes.
The identification of multiple crop pathogens as well as human pathogenic fungal species concentrated in the 1.8–3.2 μm size range, signals a potential risk of airborne transmission and long-range dispersal across ecosystems. Such findings emphasize the urgent need for continuous, high-resolution bioaerosol monitoring and integrative surveillance strategies to mitigate the threats posed by fungal pathogens to both crop productivity and public health. Establishing systematic, size-resolved bioaerosol databases and linking them with climatic and genomic data will be critical for developing predictive models and effective policy frameworks aimed at safeguarding ecosystem stability and food security in a changing environment.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, SRR22012342, SRR22012343, SRR22012344, SRR22012345, SRR22012346, SRR22012347, SRR22012348, SRR22012349, SRR22012350, SRR22012351.
Author contributions
EV: Formal analysis, Methodology, Data curation, Writing – review & editing, Writing – original draft, Investigation, Visualization. SK: Data curation, Resources, Formal analysis, Writing – review & editing, Methodology, Investigation, Supervision. TH: Writing – review & editing, Methodology. KK: Methodology, Writing – review & editing. BB: Writing – review & editing. SSK: Writing – review & editing. JG: Writing – review & editing. SY: Writing – review & editing. RV: Writing – review & editing. RR: Methodology, Writing – review & editing, Formal analysis, Validation, Visualization. SG: Resources, Funding acquisition, Writing – review & editing, Project administration, Conceptualization, Supervision.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
Emil Varghese acknowledges the Ministry of Education (MoE) for awarding the Half-Time Research Assistantship. Sarayu Krishnamoorthy thanks the Council of Scientific and Industrial Research (CSIR) for awarding the CSIR-Senior Research Associateship. Kiran Kumari acknowledges the University Grants Commission (UGC) for awarding UGC NET-JRF (National Eligibility Test-Junior Research Fellowship). The authors are also thankful to the owner of the wheat crop field for supporting the campaign. The authors acknowledge the financial support from ISRO – IITM (Indian Space Research Organization – Indian Institute of Technology Madras) Space Technology Cell and Max Planck Partner Group on Bioaerosol Research at IITM. The authors also express gratitude to Eurofins Genomics, Bangalore, India, for the NGS analysis.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1648820/full#supplementary-material
References
Alfenas, R. F., Bonaldo, S. M., Fernandes, R. A. S., and Colares, M. R. N. (2018). First report of Choanephora cucurbitarum on Crotalaria spectabilis: a highly aggressive pathogen causing a flower and stem blight in Brazil. Plant Dis. 102:1456. doi: 10.1094/PDIS-10-17-1610-PDN
Baldrian, P., Vetrovsk’y, T., Lepinay, C., and Kohout, P. (2022). High-throughput sequencing view on the magnitude of global fungal diversity. Fungal Divers. 114, 539–547. doi: 10.1007/s13225-021-00472-y
Benedict, K., and Mody, R. K. (2016). Epidemiology of histoplasmosis outbreaks, United States, 1938-2013. Emerg. Infect. Dis. 22, 370–378. doi: 10.3201/eid2203.151117
Bezerra, J. D. P., Sandoval-Denis, M., Paiva, L. M., Silva, G. A., Groenewald, J. Z., Souza-Motta, C. M., et al. (2017). New endophytic Toxicocladosporium species from cacti in Brazil, and description of Neocladosporium gen. Nov. IMA Fungus 8, 77–97. doi: 10.5598/imafungus.2017.08.01.06
Brown, G. D., Denning, D. W., Gow, N. A. R., Levitz, S. M., Netea, M. G., and White, T. C. (2012). Hidden killers: human fungal infections. Sci. Transl. Med. 4:165rv13. doi: 10.1126/scitranslmed.3004404
Buée, M., Reich, M., Murat, C., Morin, E., Nilsson, R. H., Uroz, S., et al. (2009). 454 pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 184, 449–456. doi: 10.1111/j.1469-8137.2009.03003.x
Calhim, S., Halme, P., Petersen, J. H., Læssøe, T., Bässler, C., and Heilmann-Clausen, J. (2018). Fungal spore diversity reflects substrate-specific deposition challenges. Sci. Rep. 8:5356. doi: 10.1038/s41598-018-23292-8
Caporaso, J. G., Kuczynski, J., Stombaugh, J., Bittinger, K., Bushman, F. D., Costello, E. K., et al. (2010). Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 7, 335–336. doi: 10.1038/nmeth.f.303
Carlucci, A., Raimondo, M. L., Santos, J., and Phillips, A. J. L. (2012). Plectosphaerella species associated with root and collar rots of horticultural crops in southern Italy. Persoonia Mol. Phylog. Evolut. Fungi 28, 34–48. doi: 10.3767/003158512X638251
Castillo Lopez, Diana, Zhu-Salzman, Keyan, Ek-Ramos, Maria Julissa, and Sword, Gregory A. 2014. “The entomopathogenic fungal endophytes Purpureocillium lilacinum (formerly Paecilomyces lilacinus) and Beauveria bassiana negatively affect cotton aphid reproduction under both greenhouse and field conditions” edited by T. L. Wilkinson. PLoS One 9:e103891. doi: 10.1371/journal.pone.0103891
Chen, Q. L., Cai, L., Wang, H. C., Cai, L. T., Goodwin, P., Ma, J., et al. (2020). Fungal composition and diversity of the tobacco leaf Phyllosphere during curing of leaves. Front. Microbiol. 11:554051. doi: 10.3389/fmicb.2020.554051
Couch, B. C., Fudal, I., Lebrun, M.-H., Tharreau, D., Valent, B., van Kim, P., et al. (2005). Origins of host-specific populations of the blast pathogen Magnaporthe Oryzae in crop domestication with subsequent expansion of pandemic clones on Rice and weeds of Rice. Genetics 170, 613–630. doi: 10.1534/genetics.105.041780
Cowger, C., and Brown, J. K. M. (2019). Durability of quantitative resistance in crops: greater than we know? Annu. Rev. Phytopathol. 57, 253–277. doi: 10.1146/annurev-phyto-082718-100016
Dagenais, T. R. T., and Keller, N. P. (2009). Pathogenesis of Aspergillus Fumigatus in invasive aspergillosis. Clin. Microbiol. Rev. 22, 447–465. doi: 10.1128/CMR.00055-08
Davies, J. M., Beggs, P. J., Medek, D. E., Newnham, R. M., Erbas, B., Thibaudon, M., et al. (2015). Trans-disciplinary research in synthesis of grass pollen aerobiology and its importance for respiratory health in Australasia. Sci. Total Environ. 534, 85–96. doi: 10.1016/j.scitotenv.2015.04.001
Després, V. R., Huffman, J. A., Burrows, S. M., Hoose, C., Safatov, A. S., Buryak, G., et al. (2012). Primary biological aerosol particles in the atmosphere: a review. Tellus Ser. B Chem. Phys. Meteorol. 64:15598. doi: 10.3402/tellusb.v64i0.15598
Elbert, W., Taylor, P. E., Andreae, M. O., and Pöschl, U. (2007). Contribution of fungi to primary biogenic aerosols in the atmosphere: wet and dry discharged spores, carbohydrates, and inorganic ions. Atmos. Chem. Phys. 7, 4569–4588. doi: 10.5194/acp-7-4569-2007
Fennelly, M. J., Sewell, G., Prentice, M. B., O’connor, D. J., and Sodeau, J. R. (2017). The use of real-time fluorescence instrumentation to monitor ambient primary biological aerosol particles (pbap). Atmos. 9:1. doi: 10.3390/atmos9010001
Fischer, M., and Garcia, V. G. (2015). An annotated checklist of European Basidiomycetes related to white rot of grapevine (Vitis vinifera). Phytopathol. Mediterr. 54, 281–298. doi: 10.14601/Phytopathol_Mediterr-16293
Fisher, M. C., Hawkins, N. J., Sanglard, D., and Gurr, S. J. (2018). Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 360, 739–742. doi: 10.1126/science.aap7999
Fisher, M. C., Henk, D. A., Briggs, C. J., Brownstein, J. S., Madoff, L. C., McCraw, S. L., et al. (2012). Emerging fungal threats to animal, plant and ecosystem health. Nature 484, 186–194. doi: 10.1038/nature10947
Fröhlich-Nowoisky, J., Kampf, C. J., Weber, B., Huffman, J. A., Pöhlker, C., Andreae, M. O., et al. (2016). Bioaerosols in the earth system: climate, health, and ecosystem interactions. Atmos. Res. 182, 346–376. doi: 10.1016/j.atmosres.2016.07.018
Fröhlich-Nowoisky, J., Pickersgill, D. A., Després, V. R., and Pöschl, U. (2009). High diversity of Fungi in air particulate matter. Proc. Natl. Acad. Sci. 106, 12814–12819. doi: 10.1073/pnas.0811003106
Gat, D., Reicher, N., Schechter, S., Alayof, M., Tarn, M. D., Wyld, B. V., et al. (2021). Size-resolved community structure of Bacteria and Fungi transported by dust in the Middle East. Front. Microbiol. 12, 1–16. doi: 10.3389/fmicb.2021.744117
Gnat, S., Łagowski, D., Nowakiewicz, A., and Dyląg, M. (2021). A global view on fungal infections in humans and animals: opportunistic infections and Microsporidioses. J. Appl. Microbiol. 131, 2095–2113. doi: 10.1111/jam.15032
Goudarzi, G., Soleimani, Z., Sadeghinejad, B., Alighardashi, M., Latifi, S. M., and Moradi, M. (2017). Visiting hours impact on indoor to outdoor ratio of fungi concentration at Golestan University Hospital in Ahvaz, Iran. Environ. Pollut. 6:62. doi: 10.5539/ep.v6n1p62
Guarnieri, M., and Balmes, J. R. (2014). Outdoor air pollution and asthma. Lancet 383, 1581–1592. doi: 10.1016/S0140-6736(14)60617-6
Gunasekaran, D. S. (2017). A rare case of Curvularia hawaiiensis in the ear following trauma. J. Med. Sci. Clin. Res. 5, 28154–28158. doi: 10.18535/jmscr/v5i9.131
Hawksworth, D. L., and Lücking, R. (2017). Fungal diversity revisited: 2.2 to 3.8 million species. Microbiol. Spectr. 5, 79–95. doi: 10.1128/microbiolspec.funk-0052-2016
Heald, C. L., and Spracklen, D. V. (2009). Atmospheric budget of primary biological aerosol particles from fungal spores. Geophys. Res. Lett. 36, 1–5. doi: 10.1029/2009GL037493
Hirooka, Y., Tanney, J. B., Nguyen, H. D. T., and Seifert, K. A. (2016). Xerotolerant Fungi in house dust: taxonomy of Spiromastix, Pseudospiromastix and Sigleria gen. Nov. in Spiromastigaceae (Onygenales, Eurotiomycetes). Mycologia 108, 135–156. doi: 10.3852/15-065
Hofmann, W. (2011). Modelling inhaled particle deposition in the human lung—a review. J. Aerosol Sci. 42, 693–724. doi: 10.1016/j.jaerosci.2011.05.007
Jaenicke, R. (2005). Abundance of cellular material and proteins in the atmosphere. Science 308, 73–73. doi: 10.1126/science.1106335
Jiang, W., Peng, Y., Ye, J., Wen, Y., Liu, G., and Xie, J. (2019). Effects of the entomopathogenic fungus Metarhizium anisopliae on the mortality and immune response of Locusta migratoria. Insects 11:36. doi: 10.3390/insects11010036
Jiang, X. Z., Zhong Dong, Y., Ruan, Y. M., and Wang, L. (2018). Three new species of Talaromyces sect. Talaromyces discovered from soil in China. Sci. Rep. 8, 1–11. doi: 10.1038/s41598-018-23370-x
Jooste, W. J., Roldan, A., Van Der Merwe, W. J. J., and Honrubia, M. (1990). Articulospora proliferata sp. nov., an aquatic hyphomycete from South Africa and Spain. Mycol. Res. 94, 947–951. doi: 10.1016/S0953-7562(09)81310-5
Kellner, R., Vollmeister, E., Feldbrügge, M., and Begerow, D. (2011). Interspecific sex in grass smuts and the genetic diversity of their pheromone-receptor system. PLoS Genet. 7:e1002436. doi: 10.1371/journal.pgen.1002436
Khan, Z., Ahmad, S., Al-Ghimlas, F., Al-Mutairi, S., Joseph, L., Chandy, R., et al. (2012). Purpureocillium Lilacinum as a cause of Cavitary pulmonary disease: a new clinical presentation and observations on atypical morphologic characteristics of the isolate. J. Clin. Microbiol. 50, 1800–1804. doi: 10.1128/JCM.00150-12
Kim, Y. S., and Singh, A. P. (2000). Icromorphological characteristics of wood biodegradation in wet environments: a review. IAWA J. 21, 135–155. doi: 10.1163/22941932-90000241
Knežević, A., Stajić, M., Sofrenić, I., Stanojković, T., Milovanović, I., Tešević, V., et al. (2018). Antioxidative, antifungal, cytotoxic and Antineurodegenerative activity of selected Trametes species from Serbia. PLoS One 13, 1–18. doi: 10.1371/journal.pone.0203064
Köhler, J. R., Casadevall, A., and Perfect, J. (2015). The Spectrum of Fungi that infects humans. Cold Spring Harb. Perspect. Med. 5:a019273. doi: 10.1101/cshperspect.a019273
Krishnamoorthy, S., Muthalagu, A., Priyamvada, H., Akkal, S., Valsan, A. E., Raghunathan, R., et al. (2020). On distinguishing the natural and human-induced sources of airborne pathogenic viable bioaerosols: characteristic assessment using advanced molecular analysis. SN Appl. Sci. 2. doi: 10.1007/s42452-020-2965-z
Lacey, J., and Goodfellow, M. (1975). A novel Actinomycete from sugar-cane bagasse: Saccharopolyspora Hirsuta gen. Et Sp. Nov. Microbiology 88, 75–85. doi: 10.1099/00221287-88-1-75
Laumbach, R. J., and Kipen, H. M. (2005). Bioaerosols and sick building syndrome: particles, inflammation, and allergy. Curr. Opin. Allergy Clin. Immunol. 5, 135–139. doi: 10.1097/01.all.0000162305.05105.d0
Li, Q., Li, L., Feng, H., Tu, W., Bao, Z., Xiong, C., et al. (2021). Characterization of the complete mitochondrial genome of basidiomycete yeast Hannaella oryzae: intron evolution, gene rearrangement, and its phylogeny. Front. Microbiol. 12, 1–11. doi: 10.3389/fmicb.2021.646567
Limtong, S., Into, P., and Attarat, P. (2020). Biocontrol of Rice seedling rot disease caused by Curvularia Lunata and Helminthosporium Oryzae by epiphytic yeasts from plant leaves. Microorganisms 8:647. doi: 10.3390/microorganisms8050647
Liu, K., Wang, J.-L., Zhao, L., and Wang, Q. (2014). Anticancer and antimicrobial activities and chemical composition of the birch Mazegill mushroom Lenzites Betulina (higher Basidiomycetes). Int. J. Med. Mushrooms 16, 327–337. doi: 10.1615/IntJMedMushrooms.v16.i4.30
McKinnon, A. C., Glare, T. R., Ridgway, H. J., Mendoza-Mendoza, A., Holyoake, A., Godsoe, W. K., et al. (2018). Detection of the entomopathogenic fungus Beauveria bassiana in the rhizosphere of wound-stressed Zea mays plants. Front. Microbiol. 9:1161. doi: 10.3389/fmicb.2018.01161
Nguyen, N. H., Song, Z., Bates, S. T., Branco, S., Tedersoo, L., Menke, J., et al. (2016). Funguild: an open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 20, 241–248. doi: 10.1016/j.funeco.2015.06.006
Nilsson, R. H., Anslan, S., Bahram, M., Wurzbacher, C., Baldrian, P., and Tedersoo, L. (2019). Mycobiome diversity: high-throughput sequencing and identification of fungi. Nat. Rev. Microbiol. 17, 95–109. doi: 10.1038/s41579-018-0116-y
Novaković, A., Karaman, M., Milovanović, I., Torbica, A., Tomić, J., Pejin, B., et al. (2018). Nutritional and phenolic profile of small edible fungal species Coprinellus disseminatus (Pers.) J.E. Lange 1938. Food Feed Res. 45, 119–128. doi: 10.5937/FFR1802119N
Okolo, O. M., Van Diepeningen, A. D., Toma, B., Nnadi, N. E., Ayanbimpe, M. G., Onyedibe, I. K., et al. (2015). First report of neonatal Sepsis due to Moesziomyces Bullatus in a preterm low-birth-weight infant. JMM Case Rep. 2, 1–4. doi: 10.1099/jmmcr.0.000011
Palmero, D., Rodr’ıguez, J. M., de Cara, M., Camacho, F., Iglesias, C., and Tello, J. C. (2011). Fungal microbiota from rain water and pathogenicity of fusarium species isolated from atmospheric dust and rainfall dust. J. Ind. Microbiol. Biotechnol. 38, 13–20. doi: 10.1007/s10295-010-0831-5
Pedrini, N., Ortiz-Urquiza, A., Huarte-Bonnet, C., Zhang, S., and Keyhani, N. O. (2013). Targeting of insect Epicuticular lipids by the Entomopathogenic fungus Beauveria Bassiana: hydrocarbon oxidation within the context of a host-pathogen interaction. Front. Microbiol. 4, 1–18. doi: 10.3389/fmicb.2013.00024
Pfaller, M. A. (2012). Antifungal drug resistance: mechanisms, epidemiology, and consequences for treatment. Am. J. Med. 125, S3–S13. doi: 10.1016/j.amjmed.2011.11.001
Pornsuriya, C., Chairin, T., Thaochan, N., and Sunpapao, A. (2017). Choanephora rot caused by Choanephora cucurbitarum on Brassica chinensis in Thailand. Aust Plant Dis Notes 12, 13–15. doi: 10.1007/s13314-017-0237-6
Priyamvada, H., Akila, M., Singh, R. K., Ravikrishna, R., Verma, R. S., Philip, L., et al. (2017a). Terrestrial macrofungal diversity from the tropical dry evergreen biome of southern India and its potential role in aerobiology. PLoS One 12, 1–21. doi: 10.1371/journal.pone.0169333
Priyamvada, H., Singh, R. K., Akila, M., Ravikrishna, R., Verma, R. S., and Gunthe, S. S. (2017b). Seasonal variation of the dominant allergenic fungal aerosols - one year study from southern Indian region. Sci. Rep. 7, 1–12. doi: 10.1038/s41598-017-11727-7
Ragavendran, C., Manigandan, V., Kamaraj, C., Balasubramani, G., Prakash, J. S., Perumal, P., et al. (2019). Larvicidal, histopathological, antibacterial activity of indigenous fungus Penicillium Sp. against Aedes Aegypti L and Culex Quinquefasciatus (say) (Diptera: Culicidae) and its acetylcholinesterase inhibition and toxicity assessment of zebrafish (Danio rerio). Front. Microbiol. 10, 1–17. doi: 10.3389/fmicb.2019.00427
Rhodes, J. (2019). Rapid worldwide emergence of pathogenic fungi. Cell Host Microbe 26, 12–14. doi: 10.1016/j.chom.2019.06.009
Rivedal, H. M., Stone, A. G., Severns, P. M., and Johnson, K. B. (2020). Characterization of the fungal community associated with root, crown, and vascular symptoms in an undiagnosed yield decline of winter squash. Phytobiomes J. 4, 178–192. doi: 10.1094/PBIOMES-11-18-0056-R
Roberts, P., and Evans, S. (2011). The book of Fungi. Chicago, Illinois: University of Chicago Press.
Rokas, A. (2022). Evolution of the human pathogenic lifestyle in fungi. Nat. Microbiol. 7, 607–619. doi: 10.1038/s41564-022-01112-0
Rudramurthy, S. M., Paul, R. A., Chakrabarti, A., Mouton, J. W., and Meis, J. F. (2019). Invasive aspergillosis by aspergillus flavus: epidemiology, diagnosis, antifungal resistance, and management. J. Fungi 5:55. doi: 10.3390/jof5030055
Savary, S., Ficke, A., Aubertot, J.-N., and Hollier, C. (2012). Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 4, 519–537. doi: 10.1007/s12571-012-0200-5
Saxena, A., Raghuwanshi, R., Gupta, V. K., and Singh, H. B. (2016). Chilli anthracnose: the epidemiology and management. Front. Microbiol. 7:1527. doi: 10.3389/fmicb.2016.01527
Shelton, B. G., Kirkland, K. H., Flanders, W. D., and Morris, G. K. (2002). Profiles of airborne fungi in buildings and outdoor environments in the United States. Appl. Environ. Microbiol. 68, 1743–1753. doi: 10.1128/AEM.68.4.1743-1753.2002
Simion, V.-E. (2018). “Dairy cows health risk: mycotoxins” in Ruminants - the husbandry, economic and health aspects (London, United Kingdom: InTech).
Slippers, B., Coutinho, T. A., Wingfield, B. D., and Wingfield, M. J. (2003). A review of the genus Amylostereum and its association with woodwasps. S. Afr. J. Sci. 99, 70–74.
Stoll, M., Begerow, D., and Oberwinkler, F. (2005). Molecular phylogeny of Ustilago, Sporisorium, and related taxa based on combined analyses of RDNA sequences. Mycol. Res. 109, 342–356. doi: 10.1017/S0953756204002229
Tanaka, D., Fujiyoshi, S., Maruyama, F., Goto, M., Koyama, S., Kanatani, J. i., et al. (2020). Size resolved characteristics of urban and suburban bacterial bioaerosols in Japan as assessed by 16S rRNA amplicon sequencing. Sci. Rep. 10, 1–10. doi: 10.1038/s41598-020-68933-z
Taylor, D., Teresa, N., Jack, W., Niall, J., Chad, N., and Roger, W. (2014). A first comprehensive census of fungi in soil reveals both hyperdiversity and fine-scale niche partitioning. Ecol. Monogr. 84, 3–20. Available online at: http://www.jstor.org/stable/43187594
Tedersoo, L., Bahram, M., Põlme, S., Kõljalg, U., Yorou, N. S., Wijesundera, R., et al. (2014). Global diversity and geography of soil Fungi. Science 346:1256688. doi: 10.1126/science.1256688
Thomas, R. J. (2013). Particle size and pathogenicity in the respiratory tract. Virulence 4, 847–858. doi: 10.4161/viru.27172
Tong, Y., and Bruce, L. (2000). The annual bacterial particle concentration and size distribution in the ambient atmosphere in a rural area of the Willamette Valley, Oregon. Aerosol Sci. Technol. 32, 393–403.
Unlu, A., Erdinc, N., Onder, K., and Mustafa, O. (2016). Ganoderma lucidum (Reishi mushroom) and cancer. JBUON 21, 792–798. doi: 10.1080/027868200303533
Valente, S., Piombo, E., Schroeckh, V., Meloni, G. R., Heinekamp, T., Brakhage, A. A., et al. (2021). CRISPR-Cas9-based discovery of the verrucosidin biosynthesis gene cluster in Penicillium polonicum. Front. Microbiol. 12, 660871–660811. doi: 10.3389/fmicb.2021.660871
Valsan, A. E., Hema, P., Ravikrishna, R., Després, V. R., Biju, C. V., Sahu, L. K., et al. (2015). Morphological characteristics of bioaerosols from contrasting locations in southern tropical India – a case study. Atmos. Environ. 122, 321–331. doi: 10.1016/j.atmosenv.2015.09.071
Valsan, A. E., Ravikrishna, R., Biju, C. V., Pöhlker, C., Després, V. R., Huffman, J. A., et al. (2016). Fluorescent biological aerosol particle measurements at a tropical high-altitude site in southern India during the southwest monsoon season. Atmos. Chem. Phys. 16, 9805–9830. doi: 10.5194/acp-16-9805-2016
Varghese, E., Krishnamoorthy, S., Patel, A., Thazhekomat, H., Kumari, K., Bhattacharya, B. K., et al. (2024). Relationship between fungal bioaerosols and biotic stress on crops: a case study on wheat rust fungi. J. Plant Dis. Prot. 131, 823–833. doi: 10.1007/s41348-024-00868-3
Wang, L., and Lin, X. (2012). Morphogenesis in fungal pathogenicity: shape, size, and surface. PLoS Pathog. 8, 8–11. doi: 10.1371/journal.ppat.1003027
Wang, M. M., Shenoy, B. D., Li, W., and Cai, L. (2017). Molecular phylogeny of Neodevriesia, with two new species and several new combinations. Mycologia 109, 965–974. doi: 10.1080/00275514.2017.1415075
Whitby, S., Madu, E. C., and Bronze, M. S. (1996). Candida Zeylanoides infective endocarditis complicating infection with the human immunodeficiency virus. Am J Med Sci 312, 138–139. doi: 10.1016/S0002-9629(15)41781-1
WHO (2018) Mycotoxins, fact sheets. Available online at: https://www.who.int/news-room/fact-sheets/detail/mycotoxins (Accessed October 22, 2022).
Woo, C., An, C., Siyu, X., Yi, S. M., and Yamamoto, N. (2018). Taxonomic diversity of Fungi deposited from the atmosphere. ISME J. 12, 2051–2060. doi: 10.1038/s41396-018-0160-7
Yadav, S., Gettu, N., Swain, B., Kumari, K., Ojha, N., and Gunthe, S. S. (2020). Bioaerosol impact on crop health over India due to emerging fungal diseases (EFDs): An important missing link. Environ. Sci. Pollut. Res. 27, 12802–12829. doi: 10.1007/s11356-020-08059-x
Yamamoto, N., Bibby, K., Qian, J., Hospodsky, D., Rismani-Yazdi, H., Nazaroff, W. W., et al. (2012). Particle-size distributions and seasonal diversity of allergenic and pathogenic Fungi in outdoor air. ISME J. 6, 1801–1811. doi: 10.1038/ismej.2012.30
Yamamoto, N., Nazaroff, W. W., and Peccia, J. (2014). Assessing the aerodynamic diameters of taxon-specific fungal bioaerosols by quantitative PCR and next-generation DNA sequencing. J. Aerosol Sci. 78, 1–10. doi: 10.1016/j.jaerosci.2014.08.007
Yamin, D. H., Husin, A., and Harun, A. (2021). Risk factors of Candida Parapsilosis catheter-related bloodstream infection. Front. Public Health 9:631865. doi: 10.3389/fpubh.2021.631865
Keywords: fungal pathogens, environmental strains, useful fungi, medicinal fungi, next-generation sequencing
Citation: Varghese E, Krishnamoorthy S, Hredhya TK, Kumari K, Bhattacharya BK, Kundu SS, Goswami J, Yadav S, Verma RS, Ravikrishna R and Gunthe SS (2025) Size-resolved fungal bioaerosol diversity over an Indian agricultural field and their ecosystem-health implications. Front. Microbiol. 16:1648820. doi: 10.3389/fmicb.2025.1648820
Edited by:
Jacek Kozdrój, University of Rzeszow, PolandReviewed by:
Raghawendra Kumar, Agricultural Research Organization (ARO), IsraelCarolina Brunner Mendoza, National Autonomous University of Mexico, Mexico
Copyright © 2025 Varghese, Krishnamoorthy, Hredhya, Kumari, Bhattacharya, Kundu, Goswami, Yadav, Verma, Ravikrishna and Gunthe. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Emil Varghese, ZW1pbHZhcmdoZXNlOTE2QGdtYWlsLmNvbQ== Sachin S. Gunthe, cy5ndW50aGVAaWl0bS5hYy5pbg==