 María Cruz Juárez-Aragón1†
María Cruz Juárez-Aragón1† Jerjes R. Pantoja-Irys2*
Jerjes R. Pantoja-Irys2* Edilia de la Rosa-Manzano1,3*Lorena Garrido-Olvera3Hugo Mujica-Sánchez2
Edilia de la Rosa-Manzano1,3*Lorena Garrido-Olvera3Hugo Mujica-Sánchez2 Carlos Rafael Trejo-De León2
Carlos Rafael Trejo-De León2 Alejandra Vázquez-Lobo4
Alejandra Vázquez-Lobo4- 1Instituto de Ecología Aplicada, Universidad Autónoma de Tamaulipas, Ciudad Victoria, Mexico
- 2Corporación Ambiental de México, Monterrey, Mexico
- 3Facultad de Ingeniería y Ciencias, Universidad Autónoma de Tamaulipas, Ciudad Victoria, Mexico
- 4Centro de Investigación en Biodiversidad y Conservación, Universidad Autónoma del Estado de Morelos, Cuernavaca, Mexico
Introduction: Bacterial communities are fundamental to the functionality of thermal springs where they engage in essential processes such as the oxidation of sulfur, reduction of nitrates, carbon fixation, production of unique metabolites, and stabilization of microbial trophic networks. Northeastern Mexico presents a diverse array of thermal springs located within tropical karst systems situated among folded mountains and ancient inactive karstic regions. The geological complexity of these environments indicates a substantial potential for microbiome diversity; however, the composition and functional dynamics of microbial communities in these springs have not been thoroughly investigated.
Methods: This study involved the collection of water samples from six hot springs, to characterize the planktonic microbiome using advanced metagenomic sequencing techniques. Additionally, we examined the relationship between microbial composition and physicochemical parameters.
Results: Our analysis identified a total of 425 microbial species, which included 409 bacterial species, 13 eukaryotic organisms, and 3 archaeal taxa. The Ojo Caliente and Mainero Azufroso springs displayed the highest microbial diversity, whereas the Balneario El Bañito and Taninul springs exhibited the lowest. The Phyum Pseudomonadota was the predominant across the majority of springs, while Campylobacterota and Chlorobiota were specifically identified in the less diverse Balneario El Bañito and Taninul springs, respectively. A total of 30 indicator species were identified, predominantly in El Bañito and Potrero Prieto springs, emphasizing the distinctiveness of their microbial environments. Moreover, we found that electrical conductivity and bicarbonate concentration had a significant impact on the structure of this microbial communities.
Discussion: This study highlights the ecological importance of these unique ecosystems in northeastern Mexico, with the Mainero Azufroso and Ojo Caliente springs identified as reservoirs of high microbial diversity.
1 Introduction
Geothermal systems represent extreme environmental conditions conducive to the establishment of microbial communities, encompassing both prokaryotic and eukaryotic organisms. These microorganisms assume critical ecological roles, such as regulating biogeochemical cycles (Sorokin et al., 2014; Martínez-Espinosa, 2020) and fostering interspecific metabolic interactions (Weiland-Bräuer, 2021). In these unique environments, microorganisms have developed a range of adaptive strategies to withstand harsh conditions, which may include acidic or alkaline pH, high salinity, elevated pressure, and extreme temperature variations (Merino et al., 2019; Ortega-Villar et al., 2024). Such adaptations have led to the specialization and formation of distinct taxonomic groups such as mesophiles, acidophiles, alkaliphiles, and thermophiles (Von Hegner, 2020; Sriaporn et al., 2023).
Key physicochemical parameters such as temperature, pH, nutrient availability, oxygen concentration, and the presence of heavy metals significantly influence microbial distribution in hot springs (Cho et al., 2016). The interplay of these factors shapes microbial community composition, constrains species diversity and affects metabolic and biochemical functions. For instance, deviations in pH from optimal levels can drastically impair mesophilic growth (Madigan et al., 2021), whereas thermophiles exhibit resilience across acidic or alkaline environments contingent on environmental stability (Martínez, 2024). Mesophilic microorganisms generally thrive under moderate conditions, typically around 37 °C, while thermophiles are adapted to elevated temperatures yet may be sensitive to extreme pH ranges, notably between 5 and 9 (Kruglikov and Xia, 2024). Acidophilic bacteria such as Acidithiobacillus ferrooxidans and Leptospirillum spp. predominately inhabit acidic environments due to their proficiency in low pH conditions and ability to facilitate mineral oxidation processes (Aliyu et al., 2024). Conversely, alkaline springs favor species such as Bacillus alcalophilus and halophilic archaea from the genus Natronobacterium, which exhibit strong adaptations to high-pH conditions.
Electrical conductivity, a metric reflecting interactions among dissolved minerals in water, emerges as a crucial determinant of microbial community stability and diversity. This parameter holds particular significance in thermal environments where the concentration of compounds such as sulfates and carbonates directly influences microbial community structures (Dong et al., 2022). Additionally, oxygen availability serves as a vital factor in microbial development, distinguishing between aerobic microorganisms that require elevated oxygen levels for metabolism, and anaerobic organisms that employ alternative electron acceptors like sulfates. Microaerophilic microbes, meanwhile, can thrive under low-oxygen conditions, demonstrating adaptability to environments with limited oxygen concentrations (Valcheva et al., 2020). Notably, further research is required to elucidate how the interaction between these gradients, local geochemical characteristics, isotopic compositions, and oxygen levels shapes microbiome diversity within geothermal systems, offering significant implications for biotechnological and ecological applications.
The hot springs in northeastern Mexico emerge from the northeastern front of the Sierra Madre Oriental (Figure 1), a NW-SE-oriented mountain belt characterized by elongated, narrow ridges serving as recharge zones for regional aquifers. These aquifers develop within tropical karst systems shaped by folded and faulted mountains in the south, and inactive karst formations in the north (Espinasa-Pereña and Nieto-Torres, 2015; Pantoja-Irys et al., 2022). Within this region, various hot springs exist, emitting hydrogen sulfide vapors that give rise to unique aquatic ecosystems marked by sulfur, gypsum, calcite or halite precipitation nearby. However, knowledge regarding the diversity patterns and taxonomic composition of the microbiomes in these geothermal springs remains limited (Castelán-Sánchez et al., 2020; Prieto-Barajas et al., 2017).
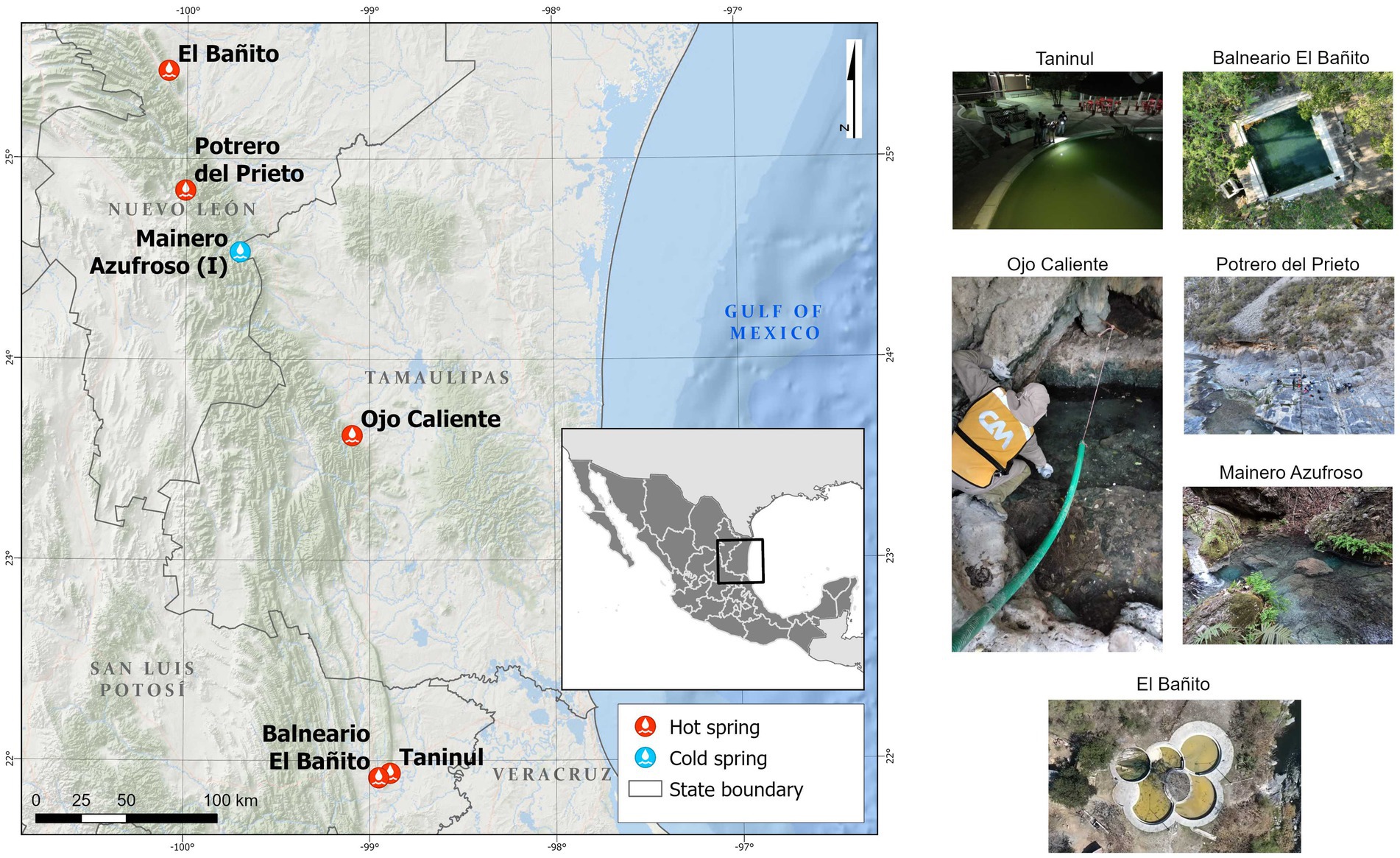
Figure 1. Location of hot springs in Northeastern Mexico. Potrero del Prieto, Balneario El Bañito, Taninul, El Bañito, Mainero Azufroso, and Ojo Caliente.
In this study, 16S rRNA gene amplicon sequencing was employed to: (1) analyze the diversity and taxonomic composition of microbial communities in thermal waters from six springs in northeastern Mexico; (2) evaluate the effects of geochemical variables on microbial community composition; and (3) identify microbial indicator species based on their ecological significance. Addressing these questions is paramount for advancing our understanding of microbial adaptability within geothermal ecosystems. The findings of this study aim to provide an integrated perspective on how microbiomes adjust to geothermal environments, establishing a baseline for the sustainable exploration and potential utilization of these invaluable resources.
2 Materials and methods
2.1 Description of low and medium enthalpy springs
This study focuses on six selected hot springs, located along the northeastern foothills of the Sierra Madre Oriental (SMO) in the Mexican states of Nuevo León, Tamaulipas, and San Luis Potosí, Mexico (Figure 1). The springs examined include: Potrero del Prieto (PP), El Bañito (EB), Ojo Caliente (OC), Taninul (TA), Balneario El Bañito (BEB), and Mainero Azufroso (MA). Notably, Mainero Azufroso, although exhibiting a cooler temperature similar to that of the adjacent stream, releases hydrogen sulfide vapors that contribute to the formation of unique aquatic ecosystems characterized by the presence of “green mats” and localized precipitation of sulfur, gypsum, calcite, or halite, warranting its inclusion in this study.
The PP hot spring is situated near the Prieta Linda waterfall and the town of El Potrero del Prieto de Arriba, nestled between the Iturbide anticline and the El Mezquital syncline in the Sierra El Baño, on the bed of the Cabezones River, at an elevation of 1,229 meters above sea level (masl). This spring is recognized as the highest hot spring in the SMO. The closest meteorological station -19073 Galeana- (Servicio Meteorológico Nacional (Mexico), 2021), reports an annual precipitation of 361.8 mm, and the region is classified as having a dry semi-warm climate (http://www.inegi.org.mx/temas/climatologia/). This hot spring emerges from the Lower Tamaulipas Formation of the Lower Cretaceous and currently has no designated use.
The EB hot spring is situated at the central part of the anticline of the sierra Cerro de La Silla, near the Rodriguez Gómez dam and the La Chueca creek at an elevation of 405 masl. The closest meteorological station -19069 La Boca- (SMN, 2021), reports an annual precipitation of 1001 mm, and the region is classified as having a semi-warm climate with summer rain (www.inegi.org.mx/temas/climatologia/). This hot spring emerges from the La Casita Formation of the Upper Jurassic and currently is used for recreational purposes.
The OC hot spring is located at an altitude of 364 masl, at the base of the El Platanillo mountain range in the Sierra El Filo. The nearest meteorological station, -28218 La Boca- (Servicio Meteorológico Nacional (Mexico), 2021), documents an average annual precipitation of 743 mm, with the area experiencing a temperate subhumid climate (http://www.inegi.org.mx/temas/climatologia/). This spring originates in the San Felipe Formation of the Upper Cretaceous and is situated on private property, currently utilized for livestock.
The TA hot spring is well-regarded locally for its medicinal and recreational applications, possibly dating back to pre-Hispanic times. It is part of a hotel complex and is located in the Sierra El Abra-Tanchipa, at the foot of the El Abra mountain range, emerging from the Cretaceous El Abra Formation at an elevation of 64 masl. The nearest meteorological station -3145 El Choy- (Servicio Meteorológico Nacional (Mexico), 2021) indicates an annual precipitation of 1165.4 mm, with the region characterized by a warm subhumid climate (http://www.inegi.org.mx/temas/climatologia/).
The BEB hot spring is currently used for recreational purposes within the municipality of Ciudad Valles. It is situated on a gently sloping hilltop, at 55 masl. According to the Ciudad Valles -24012- meteorological station (Servicio Meteorológico Nacional (Mexico), 2021) annual precipitation is reported at 1241.2 mm. This spring emerges from the San Felipe Formation, existing within a warm subhumid climate (http://www.inegi.org.mx/temas/climatologia/).
The MA spring rises at 715 masl, along the bed of an intermittent stream in the Sierra La Guitarra, at the base of the San Manuel Mountain range. The Villa Mainero −3735- meteorological station (Servicio Meteorológico Nacional (Mexico), 2021), records an annual precipitation of 993.8 mm, and the area is characterized by a temperate subhumid climate (http://www.inegi.org.mx/temas/climatologia/). This spring originates from the Taraises Formation of the Lower Cretaceous and currently lacks a specific use.
2.2 Physicochemical variables
At each spring location, a comprehensive assessment was conducted involving the measurement of 11 physicochemical variables: temperature, dissolved oxygen, electrical conductivity, salinity, oxidation–reduction potential, pH, turbidity, alkalinity, OH−, CO₃2−, and HCO₃−. The methodologies employed for the precise measurement of each variable are detailed in Pantoja-Irys et al. (2025).
2.3 Sample collection
Water samples were systematically collected directly from the spring source to minimize any influence from external water flows and to maintain the integrity of the native microbial communities. At each designated sampling site, five 1-liter replicates were collected using sterile plastic bottles. Upon filling, the bottles were promptly placed into a cooler to sustain a low temperature and inhibit microbial growth prior to the filtration process.
Filtration was conducted to effectively isolate and concentrate the microorganisms present in the water samples. This was achieved using sterile cellulose ester membranes with pore sizes of 0.45 μm, which are capable of retaining bacteria and other microbial cells. The filtration equipment consisted of a filtration funnel, a Kitazato flask connected to a vacuum pump, and sterilized membrane filters. During the assembly of the equipment, the membranes were meticulously positioned in the funnel using sterile gloves and dissecting forceps to prevent any risk of contamination.
The funnel was securely attached to the Kitazato flask, which was connected to a vacuum pump to establish negative pressure. Subsequently, the membranes were transferred to sterile 10 mL Falcon tubes, appropriately labeled and stored at 4 °C to preserve the DNA until subsequent analysis.
The filtered samples were shipped to MR DNA Laboratory (Shallowater, Texas, United States) for comprehensive microbial community profiling utilizing 16S rRNA gene amplicon sequencing. DNA was extracted using proprietary MR DNA protocols specifically optimized for environmental samples. The V4 region of the 16S rRNA gene was amplified using the universal primer pair 515F (GTGYCAGCMGCCGCGGTAA) and 806R (GGACTACNVGGGTWTCTAAT). PCR amplification was conductedperformed in a single-step reaction using the HotStarTaq Plus Master Mix Kit (Qiagen, USA) withemploying the following thermocycling conditions: an initial denaturation at 95 °C for 5 min; 30 cycles of denaturation at 95 °C for 30 s, annealing at 53 °C for 40 s, and extension at 72 °C for 1 min; followed by a final extension at 72 °C for 10 min. The PCR products were visualized on 2% agarose gels, pooled in equimolar ratios based on concentration and molecular weight, and purified using calibrated SPRI (Solid Phase Reversible Immobilization) beads. Sequencing was performed on the Illumina NovaSeq 6000 platform using paired-end chemistry (2 × 250 bp) according to the manufacturer’s protocols.
The sequence data were processed the MR DNA proprietary bioinformatics pipeline and QIIME2 v2023.2 (Bolyen et al., 2019). Paired-end reads were joined, and sequences shorter than 150 bp or containing ambiguous base calls were removed. Primer sequences were trimmed using Cutadapt, and reads were quality-filtered using a maximum expected error threshold of 1.0. Unique sequences were dereplicated and denoised using the UNOISE3 algorithm to generate amplicon sequence variants (ASVs), also referred to as zero-radius operational taxonomic units (zOTUs). Chimeras were detected and removed with UCHIME (Edgar et al., 2011). Taxonomic classification of ASVs was performed using BLASTn against a curated version of the NCBI nucleotide database (Edgar, 2010). The final outputs included absolute abundance tables and relative abundance matrices at various taxonomic ranks from phylum to species, as well as zOTU-to-sample mapping files (Edgar, 2016).
2.4 Data analysis
In order to assess species diversity within six hot spring communities, species richness and diversity were estimated using Hill numbers of order q = 0, 1 and 2. Hill numbers offer a comprehensive framework for quantifying diversity based on the effective number of species, with variations contingent on the parameter q. This parameter allows for an adjustment in the weighting of species abundances, effectively reflecting diverse aspects of community structure and enabling comparisons across samples with distinct dominance patterns (Jost, 2006; Moreno et al., 2011). Specifically, species richness (q = 0) quantifies the total number of species, without regard to their abundances. Shannon diversity (q = 1) provides a balanced estimate that incorporates both richness and evenness, thereby moderately weighting species according to their relative abundances. In contrast, Simpson diversity (q = 2) places greater emphasis on the most abundant species, consequently reducing the influence of rare species and effectively highlighting patterns of dominance within the community.
Inventory completeness was standardized through the use of sample coverage (Ĉn) facilitating a meaningful comparison of spring diversity, across various communities and ensuring that all were analyzed at consistent level of sampling completeness (Chao and Jost, 2012; Chao and Hsieh, 2016). Effective diversity estimates and sample coverage were derived utilizing the “iNEXT” function from the iNEXT package in R (https://www.r-project.org/). Statistical comparisons were conducted on the 95% confidence intervals of the Hill numbers, where significant differences were inferred if the 95% confidence intervals did not overlap.
Additionally, spring communities were classified based on species composition using a cluster dendrogram constructed using the Bray–Curtis dissimilarity index and Ward’s agglomeration method. Given the sensitivity of the Bray–Curtis index to species abundances, bacterial species abundances were log-transformed (x + 1) prior to analysis to mitigate this influence and to achieve a balanced representation of both common and rare species. The analysis was performed using the “hclust” function from the vegan package in R.
A Principal Component Analysis (PCA) was performed to identify the physicochemical variables associated with the variance among community groups observed in the studied springs. Prior to conducting the analysis, we examined the correlations among variables, identifying pairs with high correlation coefficients (> 0.85). Representative variables were selected and subsequently log-transformed, with the exception of pH, which is inherently expressed on a logarithmic scale. The PCA facilitated a visualization of the influence of environmental variables on the spring groups, allowing us to and to discern key environmental gradients. The analysis was executed using the “rda” function from the vegan package in R, resulting in a two-dimensional ordination plot. In this plot, community groups are represented as points, while physicochemical variables are denoted as vectors (arrows). The length and direction of each vector indicate the magnitude and direction of influence exerted by the respective variable. Groups situated near the terminus of the vectors exhibit a strong association with corresponding environmental gradients, whereas those positioned closer to the origin exhibit reduced influence from the measured variables.
Additionally, we estimated the indicator values of species within the community groups to identify the most robust and ecologically relevant indicator species for each group. To enhance the reliability of this analysis, we filtered the dataset used in the clustering analysis to include only 168 species, which collectively represented 95% of the total abundance. This approach allowed us to concentrate on species that significantly contribute to community structure. The indicator value for each species was calculated using the method proposed by Dufrêne and Legendre (1997) known as IndVal. This method quantifies both the specificity and fidelity of each species to a particular habitat, where specificity denotes the exclusivity of a species to a given group, and fidelity refers to the frequency of occurrence within that group. Indicator values were calculated independently for each taxon and expressed as percentages, yielding a robust metric for assessing species associations with various spring types. The analysis was conducted using the statistical software PAST version 4.17 (https://www.nhm.uio.no/english/research/resources/past/).
3 Results
3.1 Physicochemical environment
The TA hot spring exhibited the highest temperature and turbidity among the studied springs, while the PP spring demonstrated the highest salinity levels. Although no significant thermal anomaly was identified in the MA spring, it recorded the highest concentration of dissolved oxygen (Table 1). All springs maintained a neutral pH, and generally exhibited high alkalinity, predominantly in the form of bicarbonate, along with a negative redox potential, with the exception of the OC spring (Table 1).
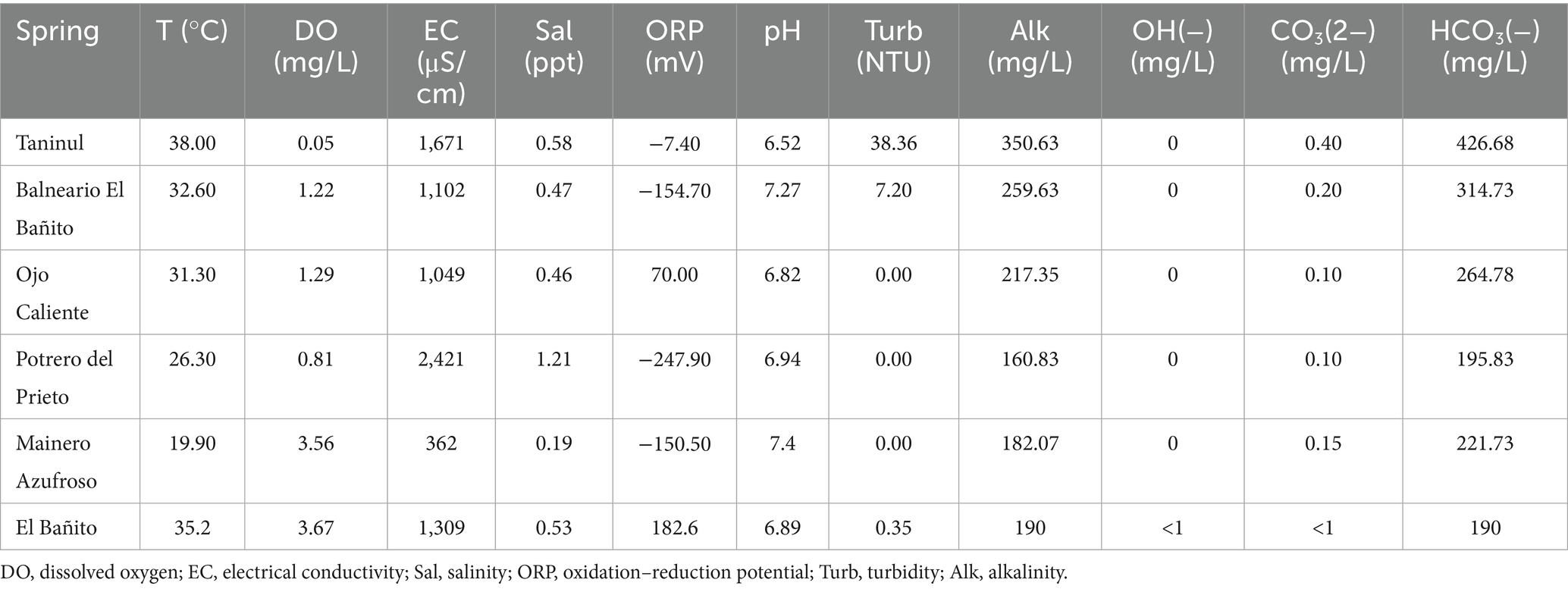
Table 1. Characterization of the physicochemical variables of hot springs in Northeastern Mexico.
3.2 Taxonomic composition
The microbiome of the six hot springs comprised 425 species, with the majority (409) attributed to the domain Bacteria, alongside 13 to Eukarya, and 3 to Archaea. The bacterial dataset encompassed 177,136 counts across the six springs, distributed among 31 phyla, 43 classes, 73 orders, 138 families, 245 genera, and 409 species. Notably, the highest bacterial counts were recorded in the TA, BEB, and PP springs, with 32,718, 31,580, and 30,921 reads, respectively. Conversely, the MA, the OC, and the EB springs exhibited the lowest counts, with 26,301, 27,803, and 27,813 reads, respectively.
The bacterial communities within the MA and the OC springs displayed the highest taxonomic richness across all taxonomic levels, from phylum to species. In contrast, the TA and the BEB springs exhibited the lowest overall taxonomic representation. While the BEB and the PP springs contained a greater number of sequence-assigned species compared to the TA spring, both communities exhibited lower richness at the phylum level, indicating a more constrained phylogenetic breadth. It is noteworthy that despite a similar number of species in the MA and OC springs, the MA spring was characterized by a higher richness at the phylum level, suggesting a broader phylogenetic diversity (Table 2).
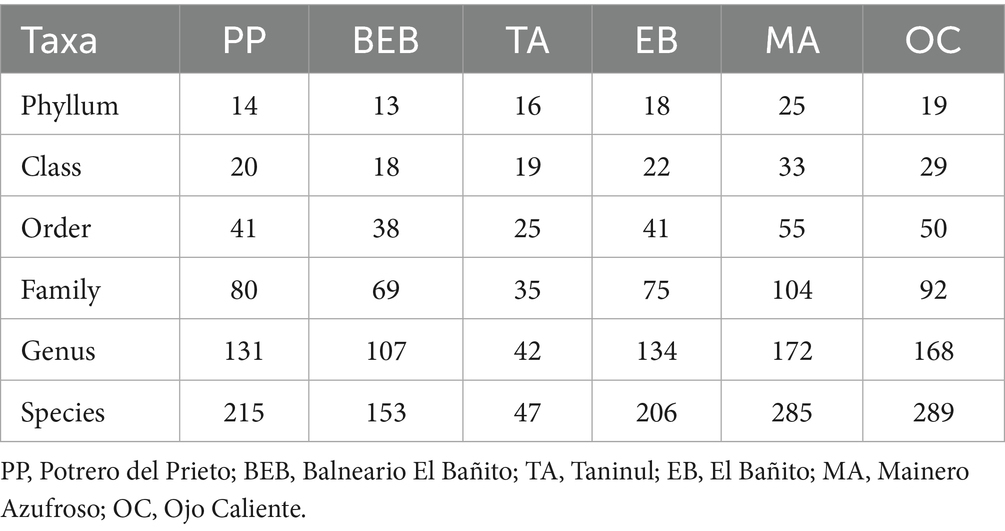
Table 2. Taxonomic composition of bacterial communities recorded in six hot springs from Northeastern Mexico.
The phylum Pseudomonadota was found to be the most abundant across all springs with the exception of the BEB and the TA springs, where Campylobacterota and Chlorobiota, respectively, dominated. Additionally, Bacteroidota ranked as the second most abundant phylum within spring EB (Figure 2).
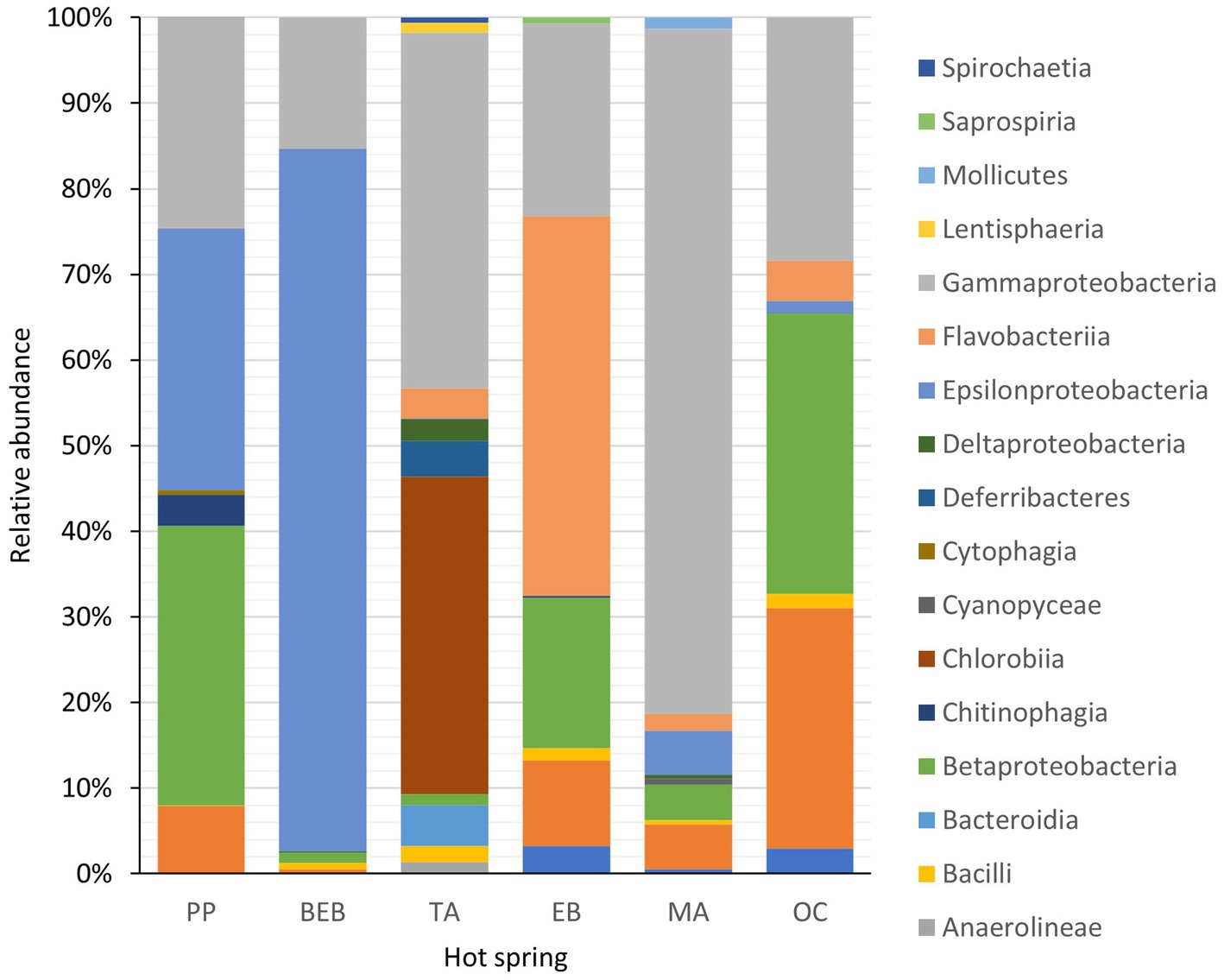
Figure 2. Relative abundance of 17 bacterial phyla recorded in six hot springs located in Northeastern Mexico. PP, Potrero del Prieto; BEB, Balneario El Bañito; TA, Taninul; EB, El Bañito; MA, Mainero Azufroso; OC, Ojo Caliente.
The taxonomic diversity observed in the investigated ecosystems revealed several dominant classes, notably Gammaproteobacteria, Betaproteobacteria, Epsilonproteobacteria, Chlorobiia, Flavobacteriia, and Alphaproteobacteria. Epsilonproteobacteria was particularly prevalent in the BEB spring, representing 80.2% of the total microbial abundance. Similarly, Gammaproteobacteria showed significant abundance in the MA spring, accounting for 71.5% of the total, while Flavobacteriia was the predominant group in the EB spring, contributing 40% to the overall abundance. It is noteworthy that Chlorobiia was exclusively identified in the TA spring, where it constituted 36.3% of the total microbial population.
At the order level, key representative groups included Campylobacterales in both the PP and BEB springs; Chromatiales and Chlorobiales in the TA spring; Flavobacteriales in the EB spring; Chromatiales again in the MA spring. In the OC spring, the dominant orders were and Burkholderiales, Moraxellales, and Rhodobacterales in the OC spring.
A detailed analysis of representative families and genera across the springs reveals that in the PP spring, Arcobacteraceae and Chromobacteriaceae were predominant, with the genera Halarcobacter and Vogesella; the BEB spring was characterized by Sulfurovaceae and Thiovulaceae, with Sulfurovum and Sulfuricurvum identified; the TA spring exhibited Halothiobacillaceae and Chlorobiaceae, featuring Thiofaba and Chlorobaculum; in the EB spring, Flavobacteriaceae, particularly Flavobacterium was prevalent; while the MA spring showcased, Halothiobacillaceae, represented by Thiofaba; finally, in the OC spring was marked by Comamonadaceae and Moraxellaceae, including Limnohabitans and Acinetobacter.
3.3 Effective diversity profile
The sample coverage across all six springs surpassed 99%, with values ranging from 0.9985 to 0.9997; (Table 3). Notably, species richness (q = 0) demonstrated significant variation among springs. Springs OC and MA displayed the highest levels of bacterial richness compared to the other sites. While no statistically significant differences in species richness were detected between the PP and the EB springs, both sites exhibited greater richness compared to the BEB and the TA springs. The TA spring recorded the lowest level of richness (Table 3; Figure 3).
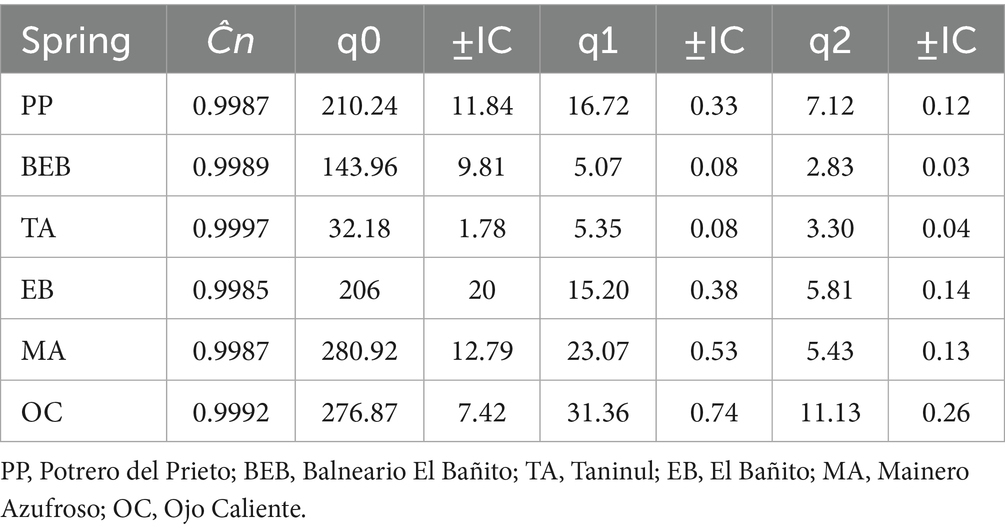
Table 3. Sample coverage (Ĉn), species richness (q0), common species (q1), and dominant species (q2) with their confidence intervals (±CI) for bacterial communities recorded in six hot springs from Northeastern Mexico.
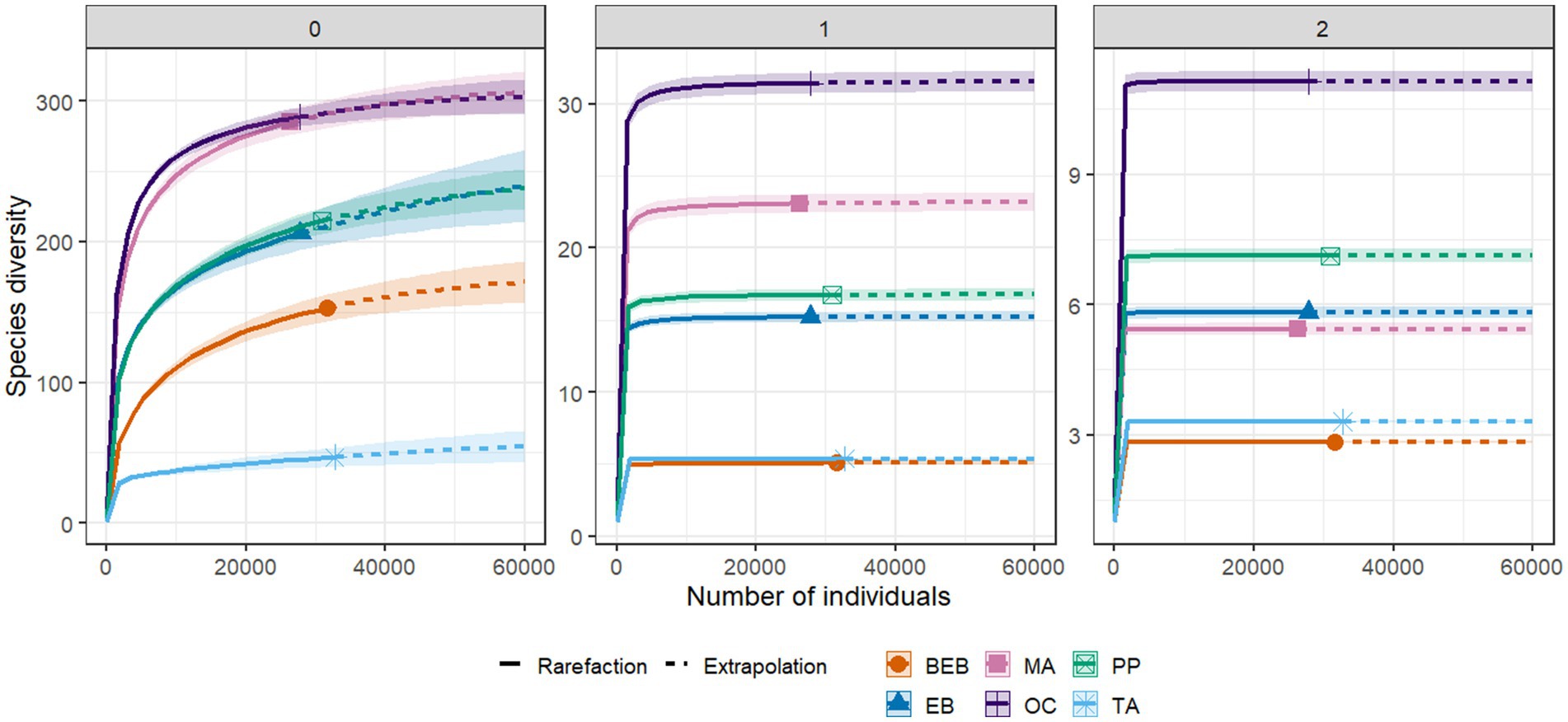
Figure 3. Diversity profile of bacterial communities in six hot springs from Northeastern Mexico. 0: species richness; 1: Shannon diversity; 2: inverse Simpson diversity. PP, Potrero del Prieto; BEB, Balneario El Bañito; TA, Taninul; EB, El Bañito; MA, Mainero Azufroso; OC, Ojo Caliente.
In terms of Shannon diversity (q = 1), the OC spring exhibited the highest value among all studied springs, with a q1 = 31.36. This was followed, in descending order, by the MA, the PP, and the EB springs (Table 3; Figure 3). In contrast, the BEB and the TA springs, which demonstrated similar diversity values, recorded the lowest levels of diversity, indicating a reduced evenness in species abundance. For inverse Simpson diversity (q = 2), the OC spring again represented the apex of diversity, achieving a q2 = 11.13, while the BEB spring marked the lowest at q2 = 2.83, suggesting a pronounced dominance of a select few species. The PP, the EB, and the MA springs displayed moderately high diversity values, reflecting varied degrees of evenness in species distribution (Table 3). Notably, despite the TA spring’s relatively low species richness, its abundances were distributed more evenly than those observed in the BEB spring (Table 3).
3.4 Community classification
The classification of springs based on species composition revealed five distinct groups (Figure 4). Group I was exclusively comprised of the TA spring, while the BEB, EB, and PP springs formed separate clusters, designated as Groups II, III, and IV, respectively. The OC and MA springs were classified together in Group V, exhibiting the highest similarity in bacterial community composition (51%).
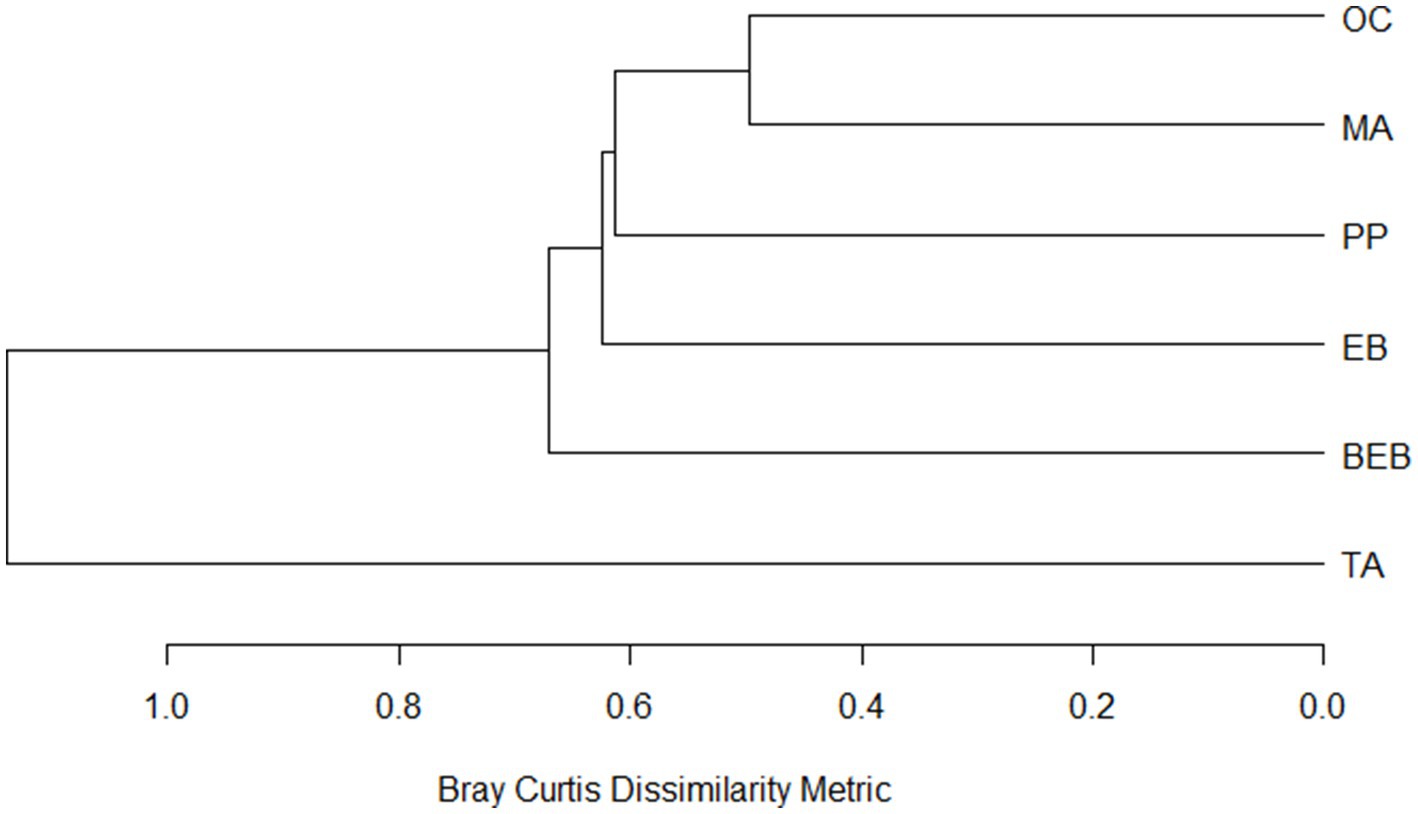
Figure 4. Dissimilarity dendrogram of bacterial communities from six hot springs in Northeastern Mexico. PP, Potrero del Prieto; BEB, Balneario El Bañito; TA, Taninul; EB, El Bañito; MA, Mainero Azufroso; OC, Ojo Caliente.
3.5 Community groups and their relationship to physicochemical properties
The Principal Component Analysis (PCA) revealed that the first two components accounted for 92.2% of the total variation in the dataset (Figure 5). The first principal component (PC1) accounted for 73.9% of the variability and was significantly influenced by electrical conductivity (EC) and bicarbonates (HCO₃−). The second principal component (PC2) accounted for 18.3% of the variation, with dissolved oxygen (DO) and pH contributing to its high values. Groundwater temperature (T°gw) had a moderate influence on PC2, although to a lesser extent compared to the other key other variables (Figure 5).
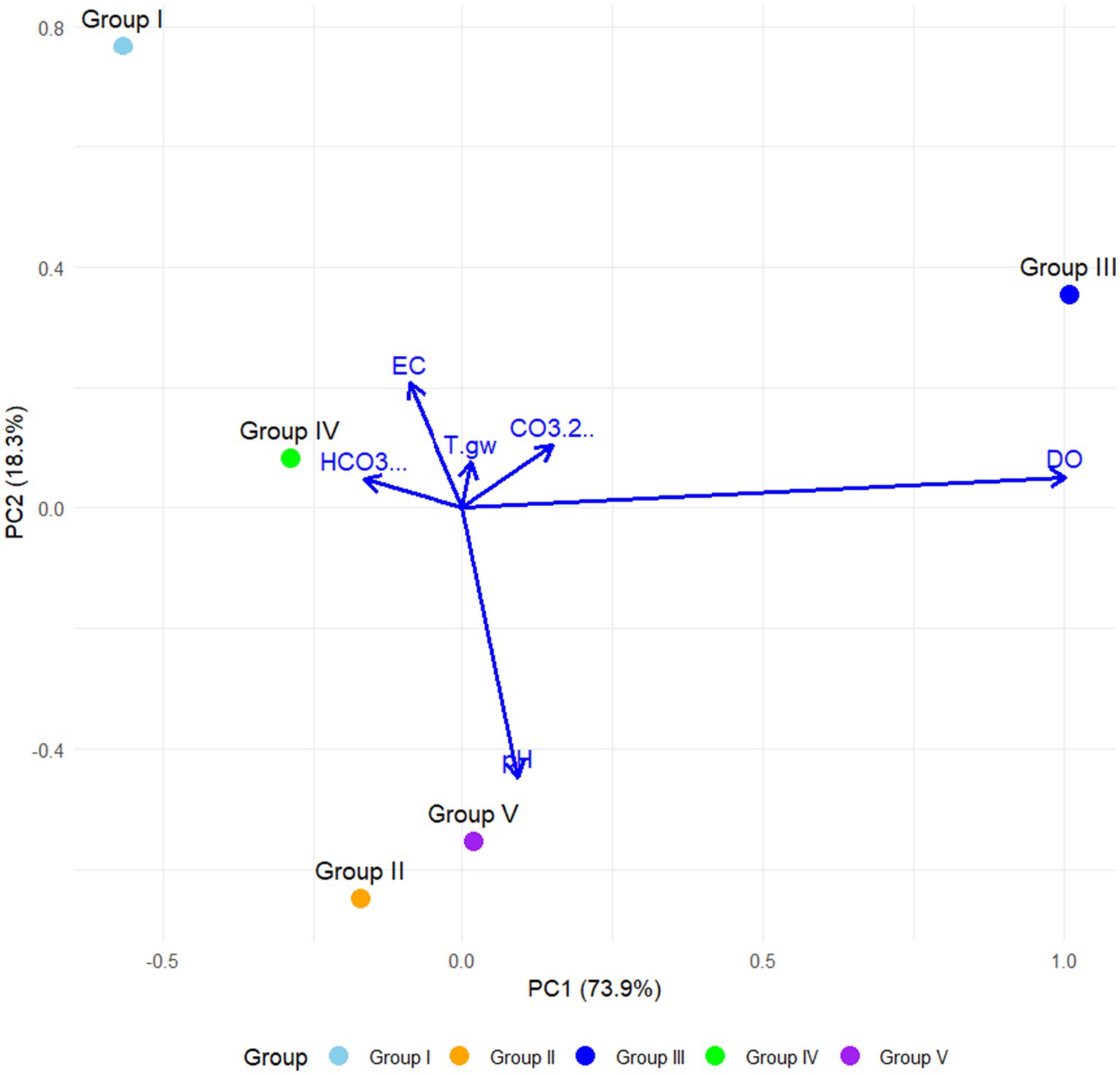
Figure 5. Two-dimensional principal component analysis (PCA) plot showing the physicochemical variables associated with community groups of sampled hot springs from Northeastern Mexico.
Group I demonstrated a strong association with elevated levels of HCO₃− and EC along PC1. Group II was positioned near the center of the plot, indicating intermediate values across the majority of physicochemical variables. Conversely, Group III exhibited a pronounced correlation with pH and DO, suggesting conditions characterized by higher oxygen availability and a neutral to basic pH. Group IV displayed some relationship with T°gw, although no other physicochemical variable appeared to exert a dominant influence. Lastly, Group V was located near the origin, indicating more balanced or less differentiated physicochemical characteristics across the measured variables (Figure 5).
3.6 Indicator species
A total of 98 species were identified as having indicator potential (IndVal ≥ 50%), with 30 species demonstrating statistical significance as indicators for the various groups (Table 4). Notably, species classified as indicators in each spring exhibited over 97% homology with existing sequences.
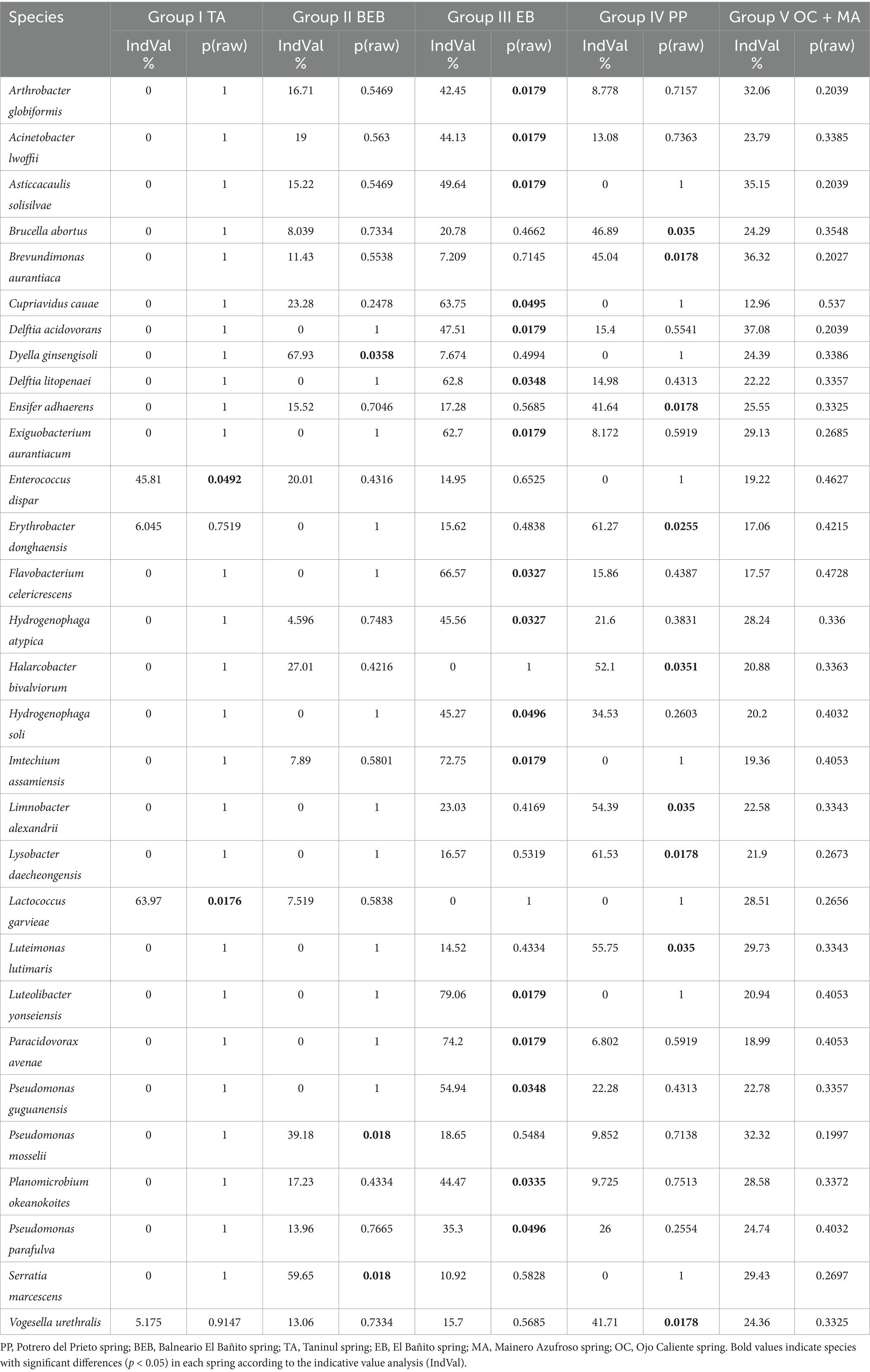
Table 4. Indicator value (IndVal %) and statistical significance (p(raw)) of 30 indicator species in four bacterial community groups from Northeastern Mexico.
In Group I, 13 species showed indicator potential; however, only two species, Enterococcus dispar (IndVal = 45.81%, p = 0.0492) and Lactococcus garvieae (IndVal = 63.97%, p = 0.0176) reached statistical significance.
Group II comprised four species with indicator potential, three of which were statistically significant (Table 4). Among these, Dyella ginsengisoli (IndVal = 67.93%, p = 0.0358) and Serratia marcescens (IndVal = 59.65%, p = 0.018) were particularly noteworthy.
Group III exhibited the highest number of significant indicator species, with 21 species showing indicator potential, of which 16 were statistically significant (Table 4). Strongly associated species in this group included Luteolibacter yonseiensis (IndVal = 79.06%, p = 0.0179), Paracidovorax avenae (IndVal = 74.2%, p = 0.0179), and Imtechium assamiensis (IndVal = 72.74%, p = 0.0179).
Group IV revealed a considerable number of potential indicators, totaling 20 species, among which nine were statistically significant. Highlighted species included Lysobacter daecheongensis (IndVal = 61.53%, p = 0.0178), Erythrobacter donghaensis (IndVal = 61.27%, p = 0.0255), and Luteimonas lutimaris (IndVal = 55.75%, p = 0.035).
Lastly, Group V comprised 40 species with indicator potential; however, no species were identified as statistically significant indicators.
4 Discussion
This study presents the first comprehensive analysis of microbial diversity and community composition in six hot springs located along the eastern flank of the SMO in northeastern Mexico, a region distinguished by its geological complexity and unique environmental conditions. Utilizing 16S rRNA gene sequencing, we identified 425 microbial species, predominantly belonging to the domain Bacteria. These species are classified across 31 phyla, 43 classes, 73 orders, 138 families, 245 genera, and 409 species.
The investigation of these ecosystems is particularly valuable due to their distinctive physicochemical conditions and relatively undisturbed environments, contrasting sharply with other hydrothermal systems influenced by anthropogenic activity (López-Sandoval et al., 2016). In nearby locations, such as the Cuatro Ciénegas Basin in Coahuila, Mexico, 325 metagenome-assembled genomes have been characterized: 277 from Bacteria and 48 from Archaea, representing 40 phyla (32 bacterial and 8 archaeal). This microbial diversity is attributed to extreme environmental conditions, including high salinity, variable pH levels (ranging from 5 to 9.8), and a significant nutrient imbalance. These factors create numerous ecological niches that likely drive the endemism of microbial lineages and foster remarkable adaptations to oligotrophic conditions (Rodríguez-Cruz et al., 2024; Medina-Chávez et al., 2025). Consequently, the extensive microbial diversity documented in this study contributes to a deeper understanding of biogeographic patterns in geothermal environments and underscores their promising biotechnological potential, particularly in applications related to bioremediation and biodegradation.
While the observed microbial composition here shares certain similarities with other thermal systems studied both in Mexico and internationally, it also shows remarkable differences. For instance, in contrast to the geothermal springs in Araró, Michoacán, Mexico (Prieto-Barajas et al., 2017), where Firmicutes are predominant, or those in Chignahuapan, México (Castelán-Sánchez et al., 2020) which are dominated by Actinobacteria and Proteobacteria, our findings indicate that Pseudomonadota was the predominant phylum in most of the springs examined. Exceptions include the BEB and TA springs, where Campylobacterota and Chlorobionta, were more prevalent, respectively. This suggests that specific adaptations to local physicochemical conditions, such as elevated temperature and turbidity, distinctly differentiate these springs from others. The presence and relative abundance of bacterial phyla including Pseudomonadota, Campylobacterota, Chlorobiota, and Bacteroidota in geothermal springs appear closely associated with the extreme environmental conditions present.
Pseudomonadota is frequently documented in geothermal systems and exhibits remarkable adaptability across various environmental gradients. For instance, studies in Eritrea, northeastern Africa, identified genera such as Pseudomonas and Marinobacter associated with high sodium and calcium concentrations (Ghilamicael et al., 2017). Likewise, research in Julong, China, Pseudomonas accounted for 72% of the bacterial diversity in thermal water samples (Wang and Pecoraro, 2021). The prevalence of this genus in the MA spring may be attributed to the favorable salinity and pH conditions unique to this environment (Table 1).
Campylobacterota, encompasses chemolithotrophic bacteria that thrive in sulfur-rich environments characterized by moderate temperatures (Sun et al., 2023). Their notable abundance of this group in the BEB spring is consistent with findings from the eolian archipelago in Italy, where Arcobacteraceae predominated in spring sediments within the temperature range of 40 °C to 53 °C (Barosa et al., 2023).
Chlorobiota exclusively identified in the TA spring, is a significant discovery potentially linked to its distinct geochemical profile. These bacteria are anaerobic phototrophs typically inhabiting sulfur-rich, oxygen-poor geothermal springs, playing critical roles in carbon and sulfur cycling (Madigan et al., 2017). Chlorobiota includes anoxygenic phototrophs of the order Chlorobiales, which have been demonstrated to significantly contribute to carbon fixation in oligotrophic geothermal systems such as those in Odisha, India (Badhai et al., 2015).
Bacteroidota was also detected across several springs and is generally associated with organic matter-rich environments. In the EB spring, Flavobacteriales were predominant, comprising bacteria frequently reported in extreme environments characterized by high temperatures, variable pH, and mineralization. Notably, the genus Flavobacterium is recognized for its capability to degrade biopolymers such as cellulose, chitin, and proteins, thereby facilitating nutrient availability for other microbial taxa (Seo et al., 2024).
The 16S sequencing technique facilitated an in-depth profiling of microbial communities, effectively addressing the limitations inherent to traditional culture-based methods (Briggs et al., 2014). However, when compared to more advanced techniques such as shotgun metagenomics or PhyloChip analysis (e.g., Hamady et al., 2010), sequencing may underestimate the total microbial diversity by overlooking low-abundance or unculturable species. Remarkable, 55 bacterial taxa in the MA, 24 in the OC, and 21 in PP spring could not be classified even at the genus level, exhibiting less than 95% sequence homology with existing databases. This finding emphasizes the exceptional and largely undescribed microbial diversity in the northeastern Mexico, along with its potential for novel metabolic function.
The microbial diversity observed in the MA and OC springs was notably higher compared to the TA and the BEB springs which exhibited lower levels of diversity. This discrepancy may be attributed to temperature, which is a crucial determinant of microbial diversity. The TA hot spring, characterized by its high temperature (30 °C), turbidity, alkalinity, elevated HCO3 concentration, and low pH, demonstrated a high abundance of microbial life yet a markedly low diversity. An inverse relationship between temperature and microbial diversity has been documented in similar circumneutral to alkaline hot springs worldwide (Sharp et al., 2014; Chan et al., 2017; Narsing Rao et al., 2021). Our study highlights a clear ecological partitioning of microbial communities across the springs, influenced by localized environmental gradients and distinct geochemical profiles. Despite being situated within a common geological framework (geothermal reservoirs within evaporitic-carbonate rocks), each site maintained a taxonomically and functionally unique assemblage, highlighting the sensitivity of the microbiomes to subtle physicochemical variations.
While it is generally observed that higher temperatures correlate with reduced microbial diversity, exceptions do exist. The SK spring in Malaysia, for instance, displayed significant diversity despite elevated temperatures, likely attributed to site-specific physical conditions (Chan et al., 2017). The dominance of Gammaproteobacteria in the MA spring (71.5%) and Epsilonproteobacteria in the BEB spring (80.2%) further illustrates the significant impact of local physicochemical conditions on microbial structure. These findings align with previous studies that link Proteobacteria abundance to nutrient-rich, sulfurous and extreme environments (Hou et al., 2013; Castelán-Sánchez et al., 2020; Susanti et al., 2025).
Notably, the OC spring exhibited not only high species richness but also substantial evenness in species distribution, as evidenced by consistently high q1 (Shannon diversity) and q2 (inverse Simpson diversity) values. This pattern suggests the microbial community is characterized by a balanced distribution of species, with no single taxon or small group predominating. The relative evenness in species abundances indicates a diverse community structure. The OC, with its intermediate temperature (31 °C), may offer greater ecological niche availability or microhabitat diversity, thereby fostering bacterial coexistence and contributing to a more diverse and evenly structured community.
The biotechnological potential of microbial communities is significant, particularly with genera such as Flavobacterium and Acinetobacter which are recognized for their abilities to degrade organic compounds and exhibit resistance to heavy metals. These characteristics indicate promising applications in bioremediation, especially in environments contaminated with arsenic or high salinity levels (Abed and Koster, 2005).
The Principal Component Analysis (PCA) revealed substantial correlations between physicochemical parameters and the structure of microbial communities. Notably, electrical conductivity (EC) and bicarbonate (HCO₃−) levels have a marked influence on communities classified within Group I (the TA spring), while dissolved oxygen (DO) and pH were more intricately associated with Group III (the EB spring). Extreme pH conditions may favor the development of more specialized, less diverse communities (Guo et al., 2021), while high concentrations of salts and ions, including sodium, magnesium, chloride, and sulfate, enhance the prevalence of halotolerant or halophilic species.
Furthermore, elements such as arsenic, iron, and manganese have been observed to impose limitations on microbial diversity due to their toxic effects; however, certain bacteria have evolved resistance mechanisms to mitigate these challenges (Prieto-Barajas et al., 2017). Factors such as total organic carbon, nitrogen, and the C: N ratio are also pivotal in determining microbial growth efficiency and, consequently, diversity. This phenomenon has been documented in both the Araró region (Prieto-Barajas et al., 2017) and Malaysian springs (Chan et al., 2017), where phosphorus availability was a critical determinant of community structure. These findings indicate that local geochemical conditions play a regulatory role in shaping microbial diversity, as previously documented in other hydrothermal systems (Purcell et al., 2007). Variability in bacterial composition across different springs may be attributed to variations in salinity, temperature, and pH, which collectively influence the abundance of specific phyla such as Chlorobiota and Campylobacterota.
Indicator species analysis has identified taxa that exhibit ecological functions congruent with the physicochemical gradients present, including pH, electrical conductivity, bicarbonates, and dissolved oxygen. Notably, Sulfurovum lithotrophicum, a mesophilic, microaerophilic chemolithotroph, oxidizes reduced sulfur compounds such as thiosulfate and hydrogen sulfide through the Sox pathway and sulfuroquinone reductase (SQR). This species is typically found in springs characterized by elevated sulfate and hydrogen sulfide concentrations, thriving under such conditions, and playing a key role in sulfur cycling. Its detection in high-conductivity, sulfate-rich environments like BA spring corroborates its function as a bioindicator of sulfur oxidation (Wang et al., 2023), emphasizing the functional and dynamic diversity of these ecosystems.
Similarly, Hydrogenophaga species oxidize molecular hydrogen via hydrogenases, using oxygen or nitrate as terminal electron acceptors. This metabolic adaptation facilitates survival in microaerophilic or low redox environments (Howells et al., 2022). In the BA spring, the prevailing low redox potential and reduced dissolved oxygen create conductive conditions for this metabolic process. Moreover, several species exhibit tolerance to heavy metals such as chromium and arsenic, attributes that are increasingly leveraged in bioremediation and electrochemical reduction of Cr(VI) (Beretta et al., 2024). These functional traits highlight the significance of indicator species analysis in elucidating how hydrogen and sulfur cycling influence microbial communities and propose various environmental, energy, and biotechnological applications (Thai et al., 2023; Tang et al., 2024; Qattan, 2025).
In the MA spring it has been reported that some species of the genus Thioalkalimicrobium possess metabolic adaptations specialized for sulfur oxidation under alkaline conditions. These metabolic characteristics elucidate their prevalence in springs exhibiting high pH and carbonate concentrations therefore, reinforcing their role in the biogeochemical cycles. This observation aligns with literature that highlights their metabolic specialization in alkaline-sulfur environments (Sun et al., 2020; Whaley-Martin et al., 2023), potentially attributable to the mineralogical composition of the caverns at the sampled sites.
While 16S sequencing has provided essential insights into microbial diversity, the absence of functional analysis constrains our understanding of the metabolic roles within these communities. Future research endeavors should incorporate metagenomic and metatranscriptomic approaches to conduct a more comprehensive investigation of microbial functions and interactions. Furthermore, expanding seasonal sampling initiatives could elucidate temporal influences on microbial composition and ecosystem functionality.
In conclusion, this study demonstrates that the hot springs in northeastern Mexico host highly diverse microbial communities and represents the first comprehensive analysis of microbial diversity present in low-to medium-enthalpy hot springs, a geothermal region that remains largely unexplored in microbiological research. The presence of geochemical gradients, such as fluctuations in electrical conductivity, bicarbonates, sulfates, and hydrogen sulfide emissions, creates unique environmental niches, that foster specialized communities with essential metabolic functions vital for maintaining the ecological balance of these ecosystems. Moreover, possess significant potential for biotechnological applications. For example, genera associated with sulfur and hydrogen oxidation metabolic pathways, such as Sulfurovum and Hydrogenophaga, have demonstrated significant potential in bioremediation and detoxification processes involving sulfur compounds and heavy metals (Qattan, 2025). Additionally, the dominance of phyla such as Pseudomonadota and Chlorobiota indicates their adaptation to these extreme environments and underscores their ecological significance. These findings not only advance our understanding of microbial ecology in low-to medium-enthalpy geothermal systems, but they also emphasize the importance of conserving the microbiomes associated with these unique habitats. The observed high microbial diversity is intricately linked to the functional integrity of the springs, and any loss could jeopardize essential ecological processes, such as nutrient cycling and detoxification, while also potentially eradicating valuable genetic and metabolic resources. Therefore, safeguarding these microbial communities is crucial, not only for maintaining ecosystem stability, but also for preserving their biotechnological potential.
Data availability statement
The data presented in this study are deposited in the NCBI GenBank repository under BioProject accession number PRJNA1288561 (Microbiome at the springs of the Sierra Madre Oriental, Mexico). The BioProject includes the associated BioSamples with the following accession number ranges: SAMN49854367 to SAMN49854621, SAMN49854706 to SAMN49854883, SAMN49856174 to SAMN49856399, SAMN49856858 to SAMN49857210, SAMN49857745 to SAMN49858075, and SAMN49855763 to SAMN49855820. This Targeted Locus Study project has been deposited at DDBJ/EMBL/GenBank under the accession KJIH00000000. The version described in this paper is the first version, KJIH01000000.
Author contributions
MJ-A: Formal analysis, Writing – original draft, Methodology, Investigation. JP-I: Conceptualization, Methodology, Writing – original draft, Investigation.ER-M: Methodology, Investigation, Formal analysis, Writing – original draft, Conceptualization. LG-O: Methodology, Writing – review & editing. HM-S: Writing – review & editing, Methodology. CRT-L: Methodology, Writing – review & editing. AV-L: Writing – review & editing, Methodology, Formal analysis.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
The authors sincerely thank Instituto de Ecología Aplicada, Universidad Autónoma de Tamaulipas and CAM’s staff, who shared its technical knowledge with the authors of this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abed, R., and Koster, J. (2005). The direct of aerobic heterotrophic bacteria associated with cyanobacteria in the degradation of oil compounds. Int. Biodeterior. Biodegradation 55, 29–37. doi: 10.1016/j.ibiod.2004.07.001
Aliyu, G., Ezugworie, F., Onwosi, C., Nnamchi, C., Ekwealor, C., Igbokwe, V., et al. (2024). Multi-stress adaptive lifestyle of acidophiles enhances their robustness for biotechnological and environmental applications. Sci. Total Environ. 954:176190. doi: 10.1016/j.scitotenv.2024.176190
Badhai, J., Ghosh, T., and Das, S. (2015). Taxonomic and functional characteristics of microbial communities and their correlation with physicochemical properties of four geothermal springs in Odisha, India. Front. Microbiol. 6:1166. doi: 10.3389/fmicb.2015.01166
Barosa, B., Ferrillo, A., Selci, M., Giardina, M., Bastianoni, A., Correggia, M., et al. (2023). Mapping the microbial diversity associated with different geochemical regimes in the shallow-water hydrothermal vents of the Aeolian archipelago, Italy. Front. Microbiol. 14:1134114. doi: 10.3389/fmicb.2023.1134114
Beretta, G., Sangalli, M., Sezenna, E., Tofalos, A. E., Franzetti, A., and Saponaro, S. (2024). Reducción electroquímica microbiana de Cr (VI) en un sistema de flujo continuo en el suelo. Eval. Gest. Amb. Integr. 20, 2033–2049.
Bolyen, E., Rideout, J. R., Dillon, M. R., Bokulich, N. A., Abnet, C. C., Al-Ghalith, G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Briggs, B., Brodie, E., Tom, L., Dong, H., Jiang, H., Huang, Q., et al. (2014). Seasonal patterns in microbial communities inhabiting the hot springs of Tengchong, Yunnan Province China. Environ. Microbiol. 16, 1579–1591. doi: 10.1111/1462-2920.12311
Castelán-Sánchez, H., Meza-Rodríguez, P., Carrillo, E., Ríos-Vázquez, D., Liñan-Torres, A., Batista-García, R., et al. (2020). The microbial composition in circumneutral thermal springs from Chignahuapan, Puebla, Mexico reveals the presence of particular sulfur-oxidizing bacterial and viral communities. Microorganisms 8:1677. doi: 10.3390/microorganisms8111677
Chan, C., Chan, K., Ee, R., Hong, K., Urbieta, M., Donati, E., et al. (2017). Effects of physiochemical factors on prokaryotic biodiversity in Malaysian circumneutral hot springs. Front. Microbiol. 8:1252. doi: 10.3389/fmicb.2017.01252
Chao, A., and Hsieh, T. (2016) iNEXT (iNterpolation and EXTrapolation) Online: Software for Interpolation and Extrapolation of Species Diversity. Available online at: http://chao.stat.nthu.edu.tw/wordpress/software_download/.
Chao, A., and Jost, L. (2012). Coverage-based rarefaction and extrapolation: standardizing samples by completeness rather than size. Ecology 93, 2533–2547. doi: 10.1890/11-1952.1
Cho, S., Kimm MiHee, K., and Lee YoungOk, L. (2016). Effect of pH on soil bacterial diversity. J. Ecol. Environ. 40:10. doi: 10.1186/s41610-016-0004-1
Dong, H., Huang, L., Zhao, L., Zeng, Q., Liu, X., Sheng, Y., et al. (2022). A critical review of mineral–microbe interaction and co-evolution: mechanisms and applications. Natl. Sci. Rev. 9:nwac128. doi: 10.1093/nsr/nwac128
Dufrêne, M., and Legendre, P. (1997). Species assemblages and indicator species: the need for a flexible asymmetrical approach. Ecol. Monogr. 67, 345–366. doi: 10.1890/0012-9615(1997)067[0345,SAAIST]2.0.CO;2
Edgar, R. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Edgar, R. (2016). SINTAX: a simple non-Bayesian taxon-omy classifier for 16S and ITS sequences. Bio Rxiv. doi: 10.1101/074161
Edgar, R., Haas, B., Clemente, J., Quince, C., and Knight, R. (2011). Uchime improves sensitivity and speed of chimera detection. Bioinformatics 27, 2194–2200. doi: 10.1093/bioinformatics/btr381
Espinasa-Pereña, R., and Nieto-Torres, A. (2015). Análisis de la vulnerabilidad a fenómenos kársticos: México, Technical Report. Secretaría de Gobernación, Coordinación Nacional de Protección Civil, Centro Nacional de Prevención de Desastres.
Ghilamicael, A., Budambula, N., Anami, S., Mehari, T., and Boga, H. (2017). Evaluation of prokaryotic diversity of five hot springs in Eritrea. BMC Microbiol. 17:203. doi: 10.1186/s12866-017-1113-4
Guo, L., Wang, G., Sheng, Y., and Shi, Z. (2021). Hydrogeochemical constraints shape hot spring microbial community compositions: evidence from acidic, moderate-temperature springs and alkaline, high-temperature springs, southwestern Yunnan geothermal areas, China. J. Geophys. Res. Biogeosci. 126:e2020JG005868. doi: 10.1029/2020JG005868
Hamady, M., Lozupone, C., and Knight, R. (2010). Fast Uni Frac: facilitating high-throughput phylogenetic analyses of microbial communities including analysis of pyrosequencing and PhyloChip data. ISME J. 4, 17–27. doi: 10.1038/ismej.2009.97
Hou, W., Wang, S., Dong, H., Jiang, H., Briggs, B., Peacock, J., et al. (2013). A comprehensive census of microbial diversity in hot springs of Tengchong, Yunnan Province China using 16S rRNA gene pyrosequencing. PLoS One 8:e53350. doi: 10.1371/journal.pone.0053350
Howells, A. E., Leong, J. A., Ely, T., Santana, M., Robinson, K., Esquivel-Elizondo, S., et al. (2022). Energetically informed niche models of hydrogenotrophs detected in sediments of serpentinized fluids of the Samail ophiolite of Oman. J. Geophys. Res. Biogeosci. 127:e2021JG006317. doi: 10.1029/2021JG006317
Kruglikov, A., and Xia, X. (2024). Mesophiles vs. thermophiles: untangling the hot mess of intrinsically disordered proteins and growth temperature of bacteria. Int. J. Mol. Sci. 25:2000. doi: 10.3390/ijms25042000
López-Sandoval, O., Montejano, G., Carmona, J., Cantoral, E., and Becerra-Absalón, I. (2016). Diversidad algal de un ambiente extremo: el manantial geotermal Los Hervideros, México. Rev. Mex. Biodivers. 87, 1–9. doi: 10.1016/j.rmb.2016.01.004
Madigan, M., Martinko, J., and Parker, J. (2021). Brock biology of microorganisms. 16th Edn. London, United Kingdom: Pearson.
Madigan, M., Schaaf, N., and Sattley, W. (2017). “The Chlorobiaceae, Chloroflexaceae, and Heliobacteriaceae” in Modern topics in the phototrophic prokaryotes. ed. P. Hallenbeck (Cham: Springer).
Martínez, G. T. (2024). Aislamiento y caracterización de bacterias resistentes a altas temperaturas de las aguas termales San Francisco-Guayllabamba ubicadas en el cantón Chambo, provincia de Chimborazo. Dissertation/bachelor’s thesis. Riobamba: Escuela Superior Politécnica de Chimborazo.
Martínez-Espinosa, R. (2020). Microorganisms and their metabolic capabilities in the context of the biogeochemical nitrogen cycle at extreme environments. Int. J. Mol. Sci. 21:4228. doi: 10.3390/ijms21124228
Medina-Chávez, N. O., Rodríguez-Cruz, U. E., Souza, V., De la Torre-Zavala, S., and Travisano, M. (2025). Salty secrets of Halobacterium salinarum AD88: a new archaeal ecotype isolated from Cuatro Cienegas Basin. BMC Genomics 26:399. doi: 10.1186/s12864-025-11550-9
Merino, N., Aronson, H., Bojanova, D., Feyhl-Buska, J., Wong, M., Zhang, S., et al. (2019). Vivir en los extremos: extremófilos y los límites de la vida en un contexto planetario. Front. Microbiol. 10:1785. doi: 10.3389/fmicb.2019.01785
Moreno, C., Barragán, F., Pineda, E., and Pavón, N. (2011). Reanálisis de la diversidad alfa: alternativas para interpretar y comparar información sobre comunidades ecológicas. Rev. Mex. Biodivers. 82, 1249–1261. doi: 10.22201/ib.20078706e.2011.4.745
Narsing Rao, M. P., Dong, Z. Y., Luo, Z. H., Li, M. M., Liu, B. B., Guo, S. X., et al. (2021). Physicochemical and microbial diversity analyses of Indian Hot Springs. Front. Microbiol. 12:627200. doi: 10.3389/fmicb.2021.627200
Ortega-Villar, R., Escalante, A., Astudillo-Melgar, F., Lizárraga-Mendiola, L., Vázquez-Rodríguez, G., Hidalgo-Lara, M., et al. (2024). Isolation and characterization of thermophilic bacteria from a hot spring in the state of Hidalgo, Mexico, and geochemical analysis of the thermal water. Microorganisms 12:1066. doi: 10.3390/microorganisms12061066
Pantoja-Irys, J., de la Rosa-Manzano, E., Martínez-Ávalos, J., Guerra-Pérez, A., Mora-Olivo, A., Arellano-Méndez, L., et al. (2025). Diversity of plant communities surrounding the hot springs on the eastern flank of the Sierra Madre Oriental, northeastern Mexico. Biology 14:382. doi: 10.3390/biology14040382
Pantoja-Irys, J., Mujica-Sánchez, H., Arista-Cázares, L., Hernández-García, C., and Wagner, M. (2022). Environmental geology and isotopic evaluation of springs within the central part of the sierra Cerro de La Silla, northeastern México. J. S. Am. Earth Sci. 119:104017. doi: 10.1016/j.jsames.2022.104017
Prieto-Barajas, C., Alfaro-Cuevas, R., Valencia-Cantero, E., and Santoyo, G. (2017). Effect of seasonality and physicochemical parameters on bacterial communities in two hot spring microbial mats from Araró, Mexico. Rev. Mex. Biodivers. 88, 616–624. doi: 10.1016/j.rmb.2017.07.010
Purcell, D., Sompong, U., Yim, L., Barraclough, T., Peerapornpisal, Y., and Pointing, S. (2007). The effects of temperature, pH and sulphide on the community structure of hyperthermophilic streamers in hot springs of northern Thailand. FEMS Microbiol. Ecol. 60, 456–466. doi: 10.1111/J.1574-6941.2007.00302.X
Qattan, S. Y. (2025). Harnessing bacterial consortia for effective bioremediation: targeted removal of heavy metals, hydrocarbons, and persistent pollutants. Environ. Sci. Eur. 37:85. doi: 10.1186/s12302-025-01103-y
Rodríguez-Cruz, U. E., Castelán-Sánchez, H. G., Madrigal-Trejo, D., Eguiarte, L. E., and Souza, V. (2024). Uncovering novel bacterial and archaeal diversity: genomic insights from metagenome-assembled genomes in Cuatro Cienegas, Coahuila. Front. Microbiol. 15:1369263. doi: 10.3389/fmicb.2024.1369263
Seo, H., Kim, J., Lee, S., and Lee, S. (2024). The plant-associated Flavobacterium: a hidden helper for improving plant health. Plant Pathol. J. 40, 251–260. doi: 10.5423/PPJ.RW.01.2024.0019
Servicio Meteorológico Nacional (Mexico). (2021). National Meteorological Service [Internet]. Mexico City: National Water Commission (CONAGUA); [cited 2021]. Available from: https://smn.conagua.gob.mx/es/
Sharp, C. E., Brady, A. L., Sharp, G. H., Grasby, S. E., Stott, M. B., and Dunfield, P. F. (2014). Humboldt’s spa: microbial diversity is controlled by temperature in geothermal environments. ISME J. 8, 1166–1174. doi: 10.1038/ismej.2013.237
Sorokin, D., Berben, T., Melton, E., Overmars, L., Vavourakis, C., and Muyzer, G. (2014). Microbial diversity and biogeochemical cycling in soda lakes. Extremophiles 18, 791–809. doi: 10.1007/s00792-014-0670-9
Sriaporn, C., Campbell, K., Van Kranendonk, M., and Handley, K. (2023). Bacterial and archaeal community distributions and cosmopolitanism across physicochemically diverse hot springs. ISME Commun. 3:80. doi: 10.1038/s43705-023-00291-z
Sun, Q. L., Xu, K., Cao, L., Du, Z., Wang, M., and Sun, L. (2023). Nitrogen and sulfur cycling driven by Campylobacterota in the sediment–water interface of deep-sea cold seep: a case in the South China Sea. MBio 14:e00117-23. doi: 10.1128/mbio.00117-23
Sun, Q. L., Zhang, J., Wang, M. X., Cao, L., Du, Z. F., Sun, Y. Y., et al. (2020). High-throughput sequencing reveals a potentially novel Sulfurovum species dominating the microbial communities of the seawater–sediment interface of a deep-sea cold seep in South China Sea. Microorganisms 8:687. doi: 10.3390/microorganisms8050687
Susanti, R., Listiaji, P., Mukaromah, R. L., Nisa, F. F., Kenarni, N. R., and Jaya, A. F. (2025). Microbial Diversity in Hot Spring Soil Microbiome. Biosaintifika, 17, 80–90.
Tang, H., Xiang, G., Xiao, W., Yang, Z., and Zhao, B. (2024). Microbial mediated remediation of heavy metals toxicity: mechanisms and future prospects. Front. Plant Sci. 15:1420408. doi: 10.3389/fpls.2024.1420408
Thai, T. D., Lim, W., and Na, D. (2023). Synthetic bacteria for the detection and bioremediation of heavy metals. Front. Bioeng. Biotechnol. 11:1178680. doi: 10.3389/fbioe.2023.1178680
Valcheva, N., Ignatov, I., and Dinkov, G. (2020). Microbiological and physicochemical research of thermal spring and mountain spring waters in the district of Sliven, Bulgaria. J. Adv. Microbiol. 20, 9–17. doi: 10.9734/jamb/2020/v20i230213
Von Hegner, I. (2020). Extremophiles: a special or general case in the search for extra-terrestrial life? Extremophiles 24, 167–175. doi: 10.1007/s00792-019-01144-1
Wang, X., and Pecoraro, L. (2021). Diversity and co-occurrence patterns of fungal and bacterial communities from alkaline sediments and water of Julong high-altitude Hot Springs at Tianchi volcano, Northeast China. Biology 10:894. doi: 10.3390/biology10090894
Wang, J., Zheng, Q., Wang, S., Zeng, J., Yuan, Q., Zhong, Y., et al. (2023). Characterization of two novel chemolithoautotrophic bacteria of Sulfurovum from marine coastal environments and further comparative genomic analyses revealed species differentiation among deep-sea hydrothermal vent and non-vent origins. Front. Mar. Sci. 10:1222526. doi: 10.3389/fmars.2023.1222526
Weiland-Bräuer, N. (2021). Friends or foes—microbial interactions in nature. Biology 10:496. doi: 10.3390/biology10060496
Keywords: bacterial diversity, environmental factors, hot springs, Mexico, microbiome
Citation: Juárez-Aragón MC, Pantoja-Irys JR, de la Rosa-Manzano E, Garrido-Olvera L, Mujica-Sánchez H, Trejo-De León CR and Vázquez-Lobo A (2025) Microbiome diversity across physicochemical gradient in low-medium enthalpy springs at the Sierra Madre Oriental eastern flank, northeastern Mexico. Front. Microbiol. 16:1663000. doi: 10.3389/fmicb.2025.1663000
Edited by:
Hidetoshi Urakawa, Florida Gulf Coast University, United StatesReviewed by:
Susana De La Torre-Zavala, Autonomous University of Nuevo León, MexicoJorge Membrillo-Hernández, Monterrey Institute of Technology and Higher Education (ITESM), Mexico
Copyright © 2025 Juárez-Aragón, Pantoja-Irys, de la Rosa-Manzano, Garrido-Olvera, Mujica-Sánchez, Trejo-De León and Vázquez-Lobo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jerjes R. Pantoja-Irys, amVyamVzLnBhbnRvamFpQGFuYWh1YWMubXg=; Edilia de la Rosa-Manzano, ZXJtYW56YW5vQGRvY2VudGVzLnVhdC5lZHUubXg=
†ORCID: María Cruz Juárez-Aragón, https://orcid.org/0000-0002-1882-5479