 Rosanne Aparecida Capanema Ribeiro1†
Rosanne Aparecida Capanema Ribeiro1† Micaela Guidotti-Takeuchi2
Micaela Guidotti-Takeuchi2 Carolyne Ferreira Dumont1
Carolyne Ferreira Dumont1 Ana Beatriz Garcez Buiatte1Bárbara de Araújo Brum1†Thais Jansen Martins1†Luana Munique Sousa Ramos3†Wendell Guerra3†
Ana Beatriz Garcez Buiatte1Bárbara de Araújo Brum1†Thais Jansen Martins1†Luana Munique Sousa Ramos3†Wendell Guerra3† Richard Costa Polveiro1
Richard Costa Polveiro1 Roberta Torres de Melo1*
Roberta Torres de Melo1* Daise Aparecida Rossi1
Daise Aparecida Rossi1- 1Laboratório de Biotecnologia Animal Aplicada, Faculdade de Medicina Veterinária, Universidade Federal de Uberlândia, Uberlândia, Brazil
- 2Instituto Federal de Educação, Ciência e Tecnologia Farroupilha, Santa Maria, Brazil
- 3Instituto de Química, Universidade Federal de Uberlândia, Uberlândia, Brazil
The high prevalence of infections caused by contaminated food, coupled with growing antimicrobial resistance, especially through horizontal gene transfer, is a challenge for public health worldwide. It is possible that this situation is intensified in the presence of by-products from animal product processing industries. In view of this, we investigated the horizontal transfer of the blaTEM gene from S. Heidelberg to E. coli J53 AzR, in the absence and presence of whey (WH) and chicken juice (CJ) in: (i) liquid medium for 3 h under agitation; (ii) solid medium overnight; (iii) liquid medium overnight and the influence of the copper(II) complex Lu54 in mitigating this transfer. The first protocol showed the highest relative conjugation frequency (RCF) of 2.23% in the absence of supplements and increased by three and four orders of magnitude in the presence of CJ and WH and was selected for treatment with Lu54. In solid/overnight, there were RCFs of less than 1%, while the liquid/overnight medium showed RCFs higher than the first protocol only in WH. The presence of WH acidified the medium, which resulted in higher RCF. Lu 54 reduced RCF from 2.2 to 0.3%, 8.2 to 1.7% and 6.2 to 0.9%, respectively, for the tests without by-products and with WH and CJ. In addition, the genomes were sequenced to map the blaTEM gene and β-lactamase families in transconjugants. The results showed that three plasmids containing blaTEM were detected in the controls and the same gene was not identified in the treatments, suggesting plasmid loss induced by the copper(II) complex (Lu54). The results prove that WH and CJ increase the frequency of conjugation in liquid media, and the Lu54 complex is a promising alternative to mitigate conjugation and, consequently, the spread of antimicrobial resistance, especially in milk and meat processing industries.
1 Introduction
Salmonella Heidelberg and Escherichia coli are pathogens of worldwide importance, frequently related to foodborne outbreaks (Elhariri et al., 2020; CDC, 2022). The clinical manifestations associated with the consumption of food contaminated by these bacteria are especially characterized by enteric symptoms, but depending on the host–parasite relationship, complications can be observed, even fatal ones (EFSA, 2020; Abu Daher et al., 2019).
In fact, epidemiological data from infants in the United States between 1968 and 2015 revealed that S. Heidelberg accounted for 29% (1954/6767) of bacteremia and 18% (65/371) of meningitis cases in this age group, with hospitalization rates exceeding 70% and case fatality rates reaching 4% in meningitis (Schmidt et al., 2023). In Australia, a multistate outbreak in 2018–2019 across five jurisdictions resulted in a hospitalization rate of 36% (22/59), highlighting the moderate severity and invasiveness of the strain involved (Kerr et al., 2022), and a prolonged outbreaks linked to backyard poultry, approximately 33% of patients with available clinical data required hospitalization, highlighting the severity of these infections (CDC, 2024). More recent evidence from a prolonged outbreak associated with poultry in the United States (2013–2014) indicated that S. Heidelberg was responsible for 634 infections, with hospitalization and invasive disease rates of 38 and 15%, respectively. RNA sequencing of nine isolates from the outbreak revealed increased heat tolerance, enhanced biofilm formation, and transcriptional priming of multidrug efflux and virulence genes, indicating an enhanced ability to survive processing stress and cause disease (Etter et al., 2019). Together, these findings highlight the significant health impact caused by S. Heidelberg in vulnerable groups.
In addition to the health hazards and economic losses associated with infectious diseases due to the considerable morbidity and mortality rates, there is growing concern about an additional problem, the increase in antimicrobial resistance (AMR), which makes treatment and control more difficult (Lamichhane et al., 2024). Although AMR occurs naturally in bacteria, some factors can aggravate the problem and contribute to its spread. These include the inappropriate use of antimicrobials, poor regulation and oversight of the use of veterinary and human antibiotics, which exert selection pressure on resistant strains, extending even to other antimicrobials not involved in the initial challenge (Reygaert, 2018; Murray et al., 2022; MAPA, 2022). As a consequence, data presented by the US Centers for Disease Control and Prevention (2019) show that more than 2.8 million antibiotic-resistant infections occur in the United States each year, resulting in more than 35,000 deaths (CDC, 2019).
One of the antibiotics widely used to treat various infections by Gram-positive and Gram-negative bacteria is the beta-lactam ampicillin, which acts by inhibiting the synthesis of the bacterial cell wall, causing it to weaken and cell lysis (NCBI, 2022). Due to indiscriminate use, AMR to this drug is growing, especially due to the presence of β-lactam resistance genes, such as bla(TEM; Elhariri et al., 2020; Srichumporn et al., 2022; Delmani et al., 2021).
Among the factors that contribute to the spread of AMR is horizontal gene transfer, in which there is temporary or permanent acquisition of genetic material from another microorganism. Transfer can occur through transformation, transduction and conjugation (Reygaert, 2018). Conjugation involves the transfer of plasmids through direct contact between a donor and recipient bacterium, and is considered the major mechanism of horizontal gene transfer (HGT; Neil et al., 2021). This transfer begins in the cytoplasm, where the relaxosome binds to the transfer origin (oriT) present in the DNA and then the coupling protein (T4CP) guides the relaxosome-oriT complex to the type IV T4SS secretion system present in the membrane, through which the DNA strand leaves the donor bacterium and passes to the recipient via the pili (Ilangovan et al., 2015; Lee and Grossman, 2007).
Plasmids containing AMR genes can spread rapidly between different bacterial strains, species or genera, which leads to the global dissemination of various genes responsible for resistance to different classes of drugs (Von Wintersdorff et al., 2016; Huddleston, 2014). Gene transfer can also endow the recipient bacterium with new metabolic capabilities, facilitating its adaptation to diverse ecological niches (Sultan et al., 2018).
Different situations can influence the frequency of conjugation, such as cell density, solid or liquid substrate, temperature, presence of nutrients, among others (Alderliesten et al., 2020; Shintani et al., 2019), but the strategies for sharing genes between bacteria are still little known in the food production chain. This prompts questions about the influence of by-products present in animal product processors, such as chicken juice and whey, on bacterial conjugation rates. It is known that chicken juice stimulates multiplication and is capable of positively interfering with bacterial virulence mechanisms, such as biofilm formation (Melo et al., 2017), but its action is not known when it comes to gene transfer. There are also no studies in the literature investigating whether whey can exert any influence on this phenomenon.
As a way of mitigating the dangers of resistance to available antimicrobials, it is important to look for new alternatives, and among the options are copper complexes, which have demonstrated antimicrobial properties (Krasnovskaya et al., 2020; Zalevskaya and Gur’eva, 2021). Ott and Mellata (2024) demonstrated that short-chain fatty acids inhibit the conjugation of resistant plasmids between Escherichia coli in vitro and in chicken tissue explants, suggesting applicability in animal processing environments. Another study showed that extracts of natural products and pure compounds reduce the conjugation of Enterobacterales plasmids, including plasmids encoding broad-spectrum β-lactamases or carbapenemases (“IncK,” “IncFII,” etc.), although the magnitude of the reduction is still modest (Alav et al., 2024). In addition, Jia et al. (2024) identified that azidothymidine (AZT), a drug used as a reverse transcriptase inhibitor for HIV, has a broad inhibitory effect on the transfer of plasmids with antibiotic resistance genes (ARG) between clinically relevant species, acting on energy metabolism, bacterial secretion system, and membrane potential; this finding points to synthetic alternatives with potential for practical use.
In this context of searching for innovative compounds to mitigate the spread of resistance, copper stands out. Copper is an essential element, it acting as a cofactor in different enzymatic reactions, energy metabolism, respiration, cellular DNA synthesis, it as a powerful antioxidant that has demonstrated, for example, efficacy in inhibiting S. typhimurium when in synergism with other antibiotics, which makes its use in the development of antimicrobial drugs interesting (Stern, 2010; Monteiro et al., 2022). Regardin the mechanism of action, copper complexes promote the generation of reactive oxygen species (ROS), disrupt enzyme activity, induce DNA damage, and compromise cell membrane integrity (Ngece et al., 2025). These effects explain why copper have emerged as promising candidates, since factors such as the nature of the ligands, coordination geometry, lipophilicity, and electron donor systems significantly modulate their antimicrobial potency and selectivity (Salah et al., 2021; Ngece et al., 2025). In this scenario, the copper(II) complex Lu54 (Takeuchi et al., in press) emerges as a relevant proposal, as it can promote greater selectivity, act as a conjugation inhibitor, and maintain activity against resistant strains.
We aimed to evaluate the influence of chicken juice and whey on the transfer of the blaTEM gene between Salmonella Heidelberg and E. coli J53 AzR under different conditions, such as solid and liquid media at different incubation times, and to determine the effectiveness of the copper(II) complex Lu54 [Cu(Clmp)(mTFpy)](PF6)2 in mitigating recombination rates by conjugation.
2 Materials and methods
chicken juice (CJ) and whey (WH). chicken juice was obtained as described by Brown et al. (2014), with modifications. Chicken exudate was collected from frozen commercial chicken thawed at room temperature for approximately 24 h. Subsequently, the chicken juice was centrifuged at least three times at 14,000 rpm for 10 min at 4 °C to eliminate large particles, and the supernatant was sterilized by filtration through a membrane with a pore size of 0.22 μm (Kasvi®, Paraná, Brazil). Similarly, whey collected from Minas frescal cheese was prepared and sterilized. Both supplements were stored at −20 °C.
Copper(II) complex Lu54. The copper(II) complex of general formula [Cu(clmp)(mtfpy)](PF6)2 (clmp = 4-chloro-N-(pyridine-2-methylene)aniline and mtfpy = 4′-(4-methylphenyl)-2,2′:6′,2″-terpyridine), herein designated Lu54, was produced and fully characterized at the Institute of Chemistry of the Federal University of Uberlândia. In this complex, the Cu(II) ions adopt a square pyramidal geometry, involving the three nitrogen atoms of the mTFpy ligand and the two nitrogen atoms of Clmp. Furthermore, in the solid state, two PF6− anions act as counter ions (Takeuchi et al., n.d.).
Determination of the target concentration of Lu54. The minimal inhibitory concentration (MIC) of the copper(II) complex Lu54 was determined using the broth microdilution method (EUCAST, 2022). Briefly, concentrations of 500, 250, 125, 62.5 and 31.25 μg/mL were initially tested. The bacterial suspensions were prepared at a concentration corresponding to 0.5 on the MacFarland scale, inoculated into microplates containing Mueller-Hinton broth (Kasvi®, Paraná, Brazil) with the copper complex at different concentrations, and then incubated at 37 °C for 24 h. For the negative control, culture medium without added bacteria was used; and for the positive control, culture medium with added S. Heidelberg and E. coli J53 bacteria without added complex was used.
The wells containing concentrations lower than the MIC were quantified using the plate count method in order to verify the highest concentration of Lu54 that did not exert bactericidal activity against the Salmonella Heidelberg and E. coli strains. This was due to the need to select the highest concentration that would allow the cells to survive and thus assess its effect on conjugation.
All the tests were carried out in three replicates and three repetitions.
Bacterial conjugation. Three different conjugation protocols were tested and the one with the highest conjugation frequency was selected for the Lu54 copper complex tests.
In all protocols, the donor strain used was Salmonella enterica Heidelberg (SH), which has sexual pili and the plasmid blaTEM gene, which codes for resistance to the β-lactam class of antimicrobials, such as ampicillin. S. Heidelberg was previously isolated from broiler litter in 2018, characterized by MELO et al. (2021), and deposited in the strain bank of the Molecular Epidemiology Laboratory of the Faculty of Veterinary Medicine of the Federal University of Uberlândia. The J53AzR (J53) strain of E. coli, which is resistant to sodium azide and does not have the blaTEM gene, was used as a recipient and is sensitive to ampicillin.
The SH and J53 strains were reactivated in brain and heart infusion pre-enrichment broth (BHI, Sigma-Aldrich®, Missouri, United States) at 37 °C for a minimum period of 6 h, until visual growth is observed due to turbidity and then individually replated on Tripitic Soy Agar (TSA - Biokar®, Allone, France), incubated at 37 °C for 18 h (Mo et al., 2017).
The conjugation protocols tested followed the methodologies proposed by Rodriguez & Fernandez (Rodriguez-Grande and Fernandez-Lopez, 2020), with adaptations. Briefly, approximately five colonies from each strain (donor and recipient) were selected and inoculated separately into sterile Falcon tubes containing 3 mL of Luria Bertani broth (LB-Neogen®, Michigan, USA) and incubated at 37 °C on an orbital shaker rotating at 60 rpm for approximately 3 h. After incubation, the absorbance (O. D.600 nm) was measured on a microplate reader (Perlong DNM-9602, Nanjing, China) to check whether the exponential phase had been reached (O. D.600 nm = 0.4 to 0.5).
The cultures of the donor and recipient strains were individually subjected to serial decimal dilution (10−1 to 10−8) on surface MacConkey agar (MC, Biokar®, Allone, France), enriched with 90 μg/mL ampicillin for SH growth and 160 μg/mL sodium azide for J53, in order to phenotypically express their resistance. The plates were incubated at 37 °C for 24 h, when the number of colonies was counted.
For conjugation protocol 1, SH and J53 were mixed (1:4 ratio) in a final volume of 1 mL. In parallel, the same process was carried out, but adding 5% of the WH or CJ (50 μL) to 190 μL of the donor strain and 760 μL of the recipient strain previously grown in LB. Finally, the tubes containing the cocultures, with and without the addition of WH and CJ, were incubated at 37 °C with slow agitation (20 rpm) for 3 h.
Conjugation protocol 2 used solid medium. Briefly, 0.22 micrometer cellulose ester membranes were placed on LB agar distributed in six-well cell culture plates. Next, 200 μL of the coculture previously obtained in conjugation 1, with and without the addition of 5% WH or CJ were added to the membrane. The plates were incubated at 37 °C overnight.
For conjugation protocol 3, the procedure was initially identical to conjugation protocol 1 and after 3 h of conjugation, a volume of 400 μL of cocultures, with and without the addition of WH or CJ, was transferred to 1,600 μL of LB broth in a sterile flask in order to renew the culture medium, followed by incubation at 37 °C overnight without stirring. The same procedure was carried out with LB medium supplemented with 5% WH or CJ, maintaining a 1:4 ratio.
All tests were carried out in triplicate and in three technical replicates.
Identification of transconjugants. The frequency of transfer of resistance to β-lactams was investigated by determining the acquisition of the blaTEM gene by phenotypic and molecular methods.
For phenotypic confirmation, after the conjugation assays, the cultures were subjected to serial decimal dilution (10−1to 10−8), sown on the surface of MacConkey agar (Kasvi®, Paraná, Brazil) supplemented with 90 μg/mL ampicillin and 160 μg/mL sodium azide (MCT), and incubated at 37 °C for 48 h.
In the case of conjugation 2, which used solid medium, phenotypic identification took place after submerging the membrane in 20 mL of saline solution to resuspend the cell mass, as well as serial dilution and seeding on MCT plates, followed by the procedures described above. The selection of the transconjugants was determined by the growth of colonies on the MCT plates, since only the strains of E. coli J53 AzR that have acquired the resistance gene blaTEM show resistance to both antimicrobials, in addition to the inhibition of growth of S. Heidelberg due to the presence of sodium azide.
Lu54 complex in conjugation frequency. After carrying out the three different conjugation protocols, the one with the highest conjugation frequencies was selected for treatment with 100 μL of the Lu54 complex. The concentration used was the pre-established one, lower than the MIC, in which we obtained no statistical difference with the controls, so as not to exert bactericidal activity.
Conjugation frequency. The efficiency of conjugation was measured by calculating the relative frequency of conjugation by the recipient (RCF), which is the ratio of the number of transconjugating colonies at the end of the conjugation process to the number of recipients at the start of the process, counted by the average number of repetitions of each treatment. Plates were selected for dilutions with growth between 25 and 250 colonies (Taniwaki et al., 2017). Calculations were carried out separately for conjugations with and without the addition of WH and CJ, and also for those treated with the Lu54 copper complex.
Visualization in Scanning Electron Microscopy (SEM). The ultrastructure of conjugation was viewed by scanning microscopy, using a modified method (Brown et al., 2014) considering the growth conditions in Luria Bertani broth (LB-Neogen®) and conjugation at 37 °C for 3 h/20 rpm. After conjugation, samples were fixed on 1-cm2 (PU) slides (Habasit Cleandrive TM, Reinach, CH) and fixed with 2.5% glutaraldehyde and 2.5% paraformaldehyde in 0.1 M PBS buffer (pH 7.4) at 4 °C overnight. Subsequently, the fixative was removed, and the samples were washed with PBS buffer. The samples were post-fixed for 2 h with 1% osmium tetroxide and washed three times with PBS buffer. The dehydrated series was applied with ethanol solutions (30, 40, 50, 60, 70, 80, and 90% and then three times at 100%) for 20 min for each step. The samples were dried in a CPD (critical drying point) system (Leica EM CPD300; Leica, Wien, Austria) using liquid carbon dioxide as a transition fluid, coated with a 20-nm gold layer (SCD 050, Baltec), and exposed on a Zeiss Supra 55 FEG SEM operating at 5 kV. Images were captured with standard ZEISS SmartSEM.
pH evaluation. The pH assessment was carried out using a previously calibrated MS mPA210 pH meter (Tecnopon, São Paulo, Brazil) and took place in the liquid conjugation media (protocols 1 and 3), with the first measurement (0 h) carried out immediately after the donor and recipient were joined; the second (3 h), after conjugation 1 had finished; and the third the following day (overnight), after conjugation 3 had finished. All the procedures were carried out with and without the addition of WH and CJ.
The tests were carried out in triplicate and two technical replicates.
Whole genome sequencing (WGS) and pre-processing of the raw data. DNA extracted from E. coli using a commercial extraction kit (Wizard Genomic DNA Purification Kit—Promega) and the methodology described above was sent for whole-genome sequencing. Six samples were selected: three DNA samples from bacteria not treated with Lu54, one with added WH, one with CJ and one without WH and CJ; and three treated with Lu54, one with added WH, one with CJ and one without WH and CJ. The genomic DNA was sequenced using the MinION platform from Oxford Nanopore Technologies (ONT) with the Rapid Sequencing gDNA-Barcoding kit (SQK- RBK004) in a run time of 72 h. To prepare the libraries, 5–10 ng of DNA were used per sample (3 μL volume). The raw sequencing data was subjected to an initial quality assessment to check the size distribution and coverage of the reads, followed by the selection of reads with high integrity for the subsequent assembly stages. Reads were filtered by quality thresholds (Phred scores: Q > 10, Q > 15 and Q > 20) using Filtlong v0.2.1 (Wick, 2021a, b). The quality metrics and summaries were generated with NanoPlot v1.41.0 (Coster, 2018), and the removal of adapters was carried out with Porechop v0.2.4 (Wick et al., 2017).
Genome assembly, quality check and annotation of plasmids and blaTEM gene. The Raven 1.8.0 tool (Vaser and Šikić, 2021) was used to assemble the genomes using the de novo methodology. QUAST v.5.0.2 (Mikheenko et al., 2018) and CheckM v.1.1.3 (Parks et al., 2015) were used to assess the quality of the genomes. These tools were used to ensure that the genomes had >90% completeness, < 5% contamination,< 200 contigs and N50 > 40,000 bp. To confirm that the genomes belonged to the Escherichia coli species, we calculated the genome-wide average nucleotide identity (ANI) with the reference genome (ASM584v2—GenBank accession GCA_000005845.2) using FastANI v.1.32 (Jain et al., 2018). We used the recommended threshold of ≥95% ANI to delimit the boundaries between species (Jain et al., 2018). We used the mob-recon v.3.1.0 tool from MOB-suite (Robertson and Nash, 2018) to identify and characterize the plasmids.
ABRicate v.1.0.01 was used as a screening tool to align the sequenced genomes and plasmid sequences to the CARD database (Alcock et al., 2023) to identify the presence and location of the blaTEM gene, using coverage values >60% and identity >80%. The Resistance Gene Identifier (RGI; Alcock et al., 2023) was then used to confirm the presence of the blaTEM gene and β-lactamase families in the sequenced genomes. The parameters adopted included the DIAMOND aligner and the include_loose flag, and genes with identity values >80% were considered positive. The plasmid sequences were annotated using Bakta (Schwengers et al., 2021), and the GenBank files generated were used as input for Clinker (Gilchrist and Chooi, 2021), to compare sequences and locate the bla(TEM) gene.
Statistical analysis of the results. Statistical analysis of the results. The data was tabulated, log-transformed and subjected to normality analysis. One way ANOVA with a 5% of significance level was used to compare the tests and possible differences in the values obtained in Traditional/No supplement, Whey or chicken juice. The T-test was used to compare groups with and without Lu54 treatment. All analyses were carried out using the GraphPad Prism 8.0 program (GraphPad Software Inc., San Diego, CA, United States).
3 Results
Conjugation frequency—different methodologies. The counts of donor, recipient and transconjugant bacteria in all these repetitions showed no statistical difference between them (p < 0.05—one way ANOVA), thus guaranteeing the quantitative maintenance of each test.
When comparing the results obtained in the three protocols (GRAPH 1), we found that conjugation 1 was the most effective, since the conjugation frequency (RCF) was 2.23%, to the detriment of 0.79 and 1.98% identified in methods 2 and 3, respectively. At the same time, we found that technique 1 was also more promising in the presence of WH and CJ, whose percentages of RCF were equivalent to 8.16 and 6.22%, respectively. The exception was method 3, which was the most effective in the presence of WH (10.16%).
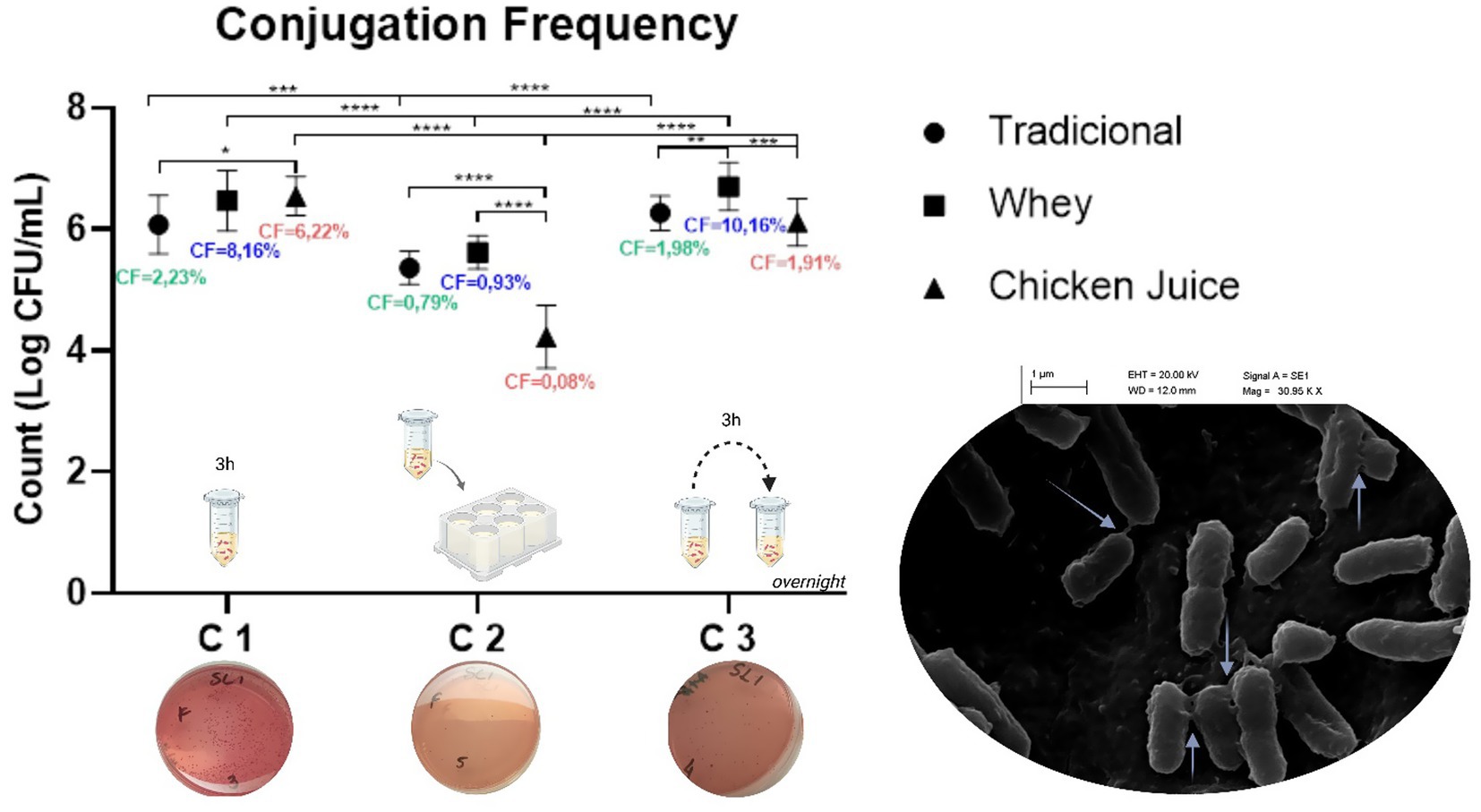
GRAPH 1. Conjugation frequencies in the different methodologies. C1: Conjugation 1, C2: Conjugation 2 and C3: Conjugation 3. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 (one way ANOVA). The graph mentions the methodology used in each type of conjugation, as well as the phenotypic demonstration of the presence of E. coli transconjugants on MacConckey plates. The scanning electron microscopy (SEM) image of bacterial conjugation between Salmonella Heidelberg and Escherichia coli (on the right) demonstrates multiple pili (arrows) for horizontal gene transfer (image obtained at 30,000 × and 40,000 × magnifications).
Statistical analysis revealed significant differences in all three conjugation groups. In Conjugation 1, there was a difference between the traditional (T) and CJ groups (p = 0.0436). In Conjugation 2, significant differences were found between T and CJ (p < 0.0001) and between WH and CJ (p < 0.0001). For Conjugation 3, differences were observed between T and SL (p = 0.0057) and between SL and CJ (p = 0.00070). Inter-conjugation comparisons were also conducted. Significant differences were observed between Conjugation 1 and 2 for the T, SL, and CJ groups (p = 0.0001, p < 0.0001, and p < 0.0001, respectively). Likewise, Conjugations 2 and 3 showed significant differences across all three groups (T, SL, and CJ), with all p-values less than 0.0001. No significant statistical difference was found between Conjugations 1 and 3.
We found that WH increased gene recombination rates, especially in liquid media, in conjugations 1 and 3, with values equivalent to four and five orders of magnitude, respectively. CJ, on the other hand, increased the rates by three orders of magnitude for conjugation in liquid with 3 h of incubation.
It is worth considering that the incubation time in the liquid conjugation media influenced the pH indices, especially in the presence of WH (GRAPH 2). There was a clear reduction in the pH of the medium with WH, varying from an average pH of 7.4 in the Traditional (media without CJ and WH) and CJ tests to 5.2 in WH (p < 0.0001, one way ANOVA) after 3 h and from 8.32 in the Traditional and CJ tests to 6.5 when supplemented with WH (p = 0.0005, one way ANOVA) in overnight incubation.
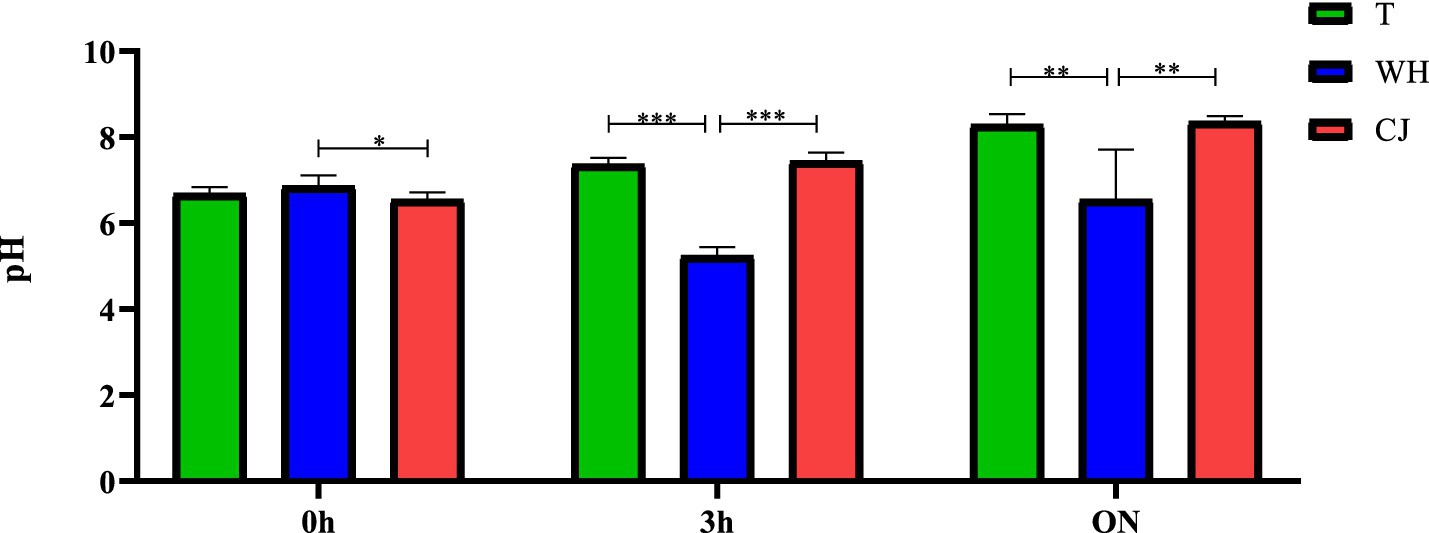
GRAPH 2. pH variation of the conjugation media in liquid at the different incubation times. T = traditional, with Luria Bertani (LB). WH = LB + 5% whey. CJ = LB + 5% chicken juice. 0 h: Immediately after donor and recipient were joined (start of conjugation). 3 h: End of conjugation 1. ON = overnight (End of conjugation 3). *p < 0.05, **p < 0.01, ***p < 0.001 (one way ANOVA). Different uppercase letters in the same row indicate statistical difference by ANOVA test (p < 0.05); different lowercase letters in the same column indicate statistical difference by ANOVA test (p < 0.05); T, Traditional/No supplement; WH, Whey; CJ, chicken juice. Conjugation 1: Liquid medium, incubated at 37 °C, for 3 h, 20 rpm; Conjugation 2: Solid medium, incubated at 37 °C, overnight, without stirring; Conjugation 3: Liquid medium, incubated at 37 °C, overnight, without stirring.
Table 1 shows the average counts (log CFU/mL) obtained in the conjugation protocols in all the tests. We found that in conjugation 1 we obtained an average increase of 0.4 and 0.5 log CFU/mL (p = 0.0130) when in the presence of WH and CJ. In methods 2 and 3, the increase was only identified in the presence of WH and was equivalent to averages of 0.3 and 0.4 log UFC/mL (p = 0.0444 and p > 0.05), respectively. In general, conjugation 2 was statistically the least effective in the production of transconjugants both in the traditional test with LB and with the supplementation of WH and CJ (p < 0.0001).
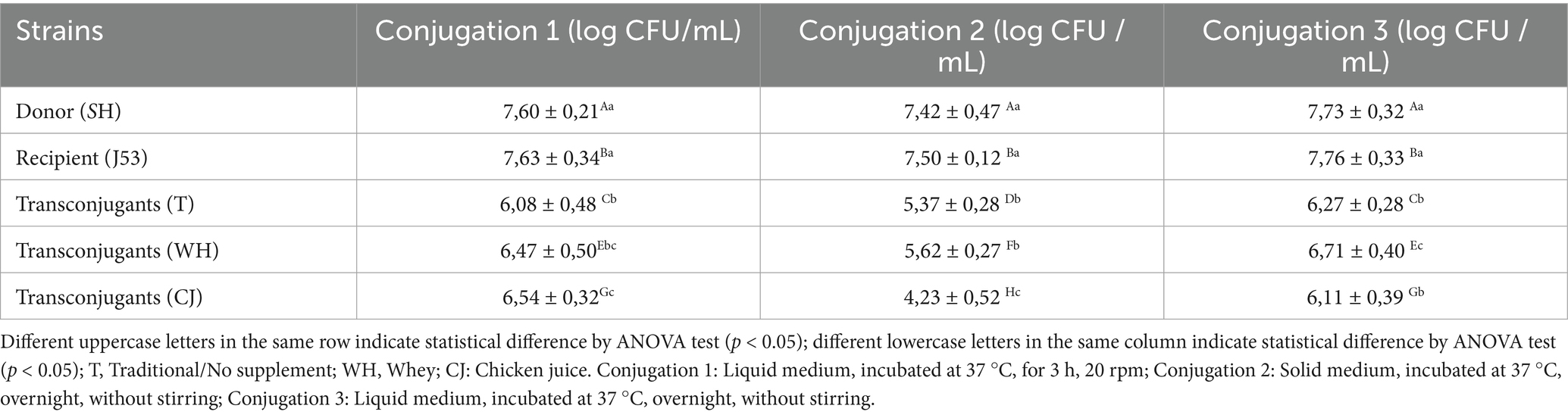
Table 1. Mean count (Log CFU/mL) of donor, recipient, and transconjugant bacteria and conjugation frequency obtained in the three conjugation protocols, with and without the addition of 5% whey and chicken juice.
Treatment with the Lu54 copper complex. The tests to determine the best concentration of the Lu54 copper complex allowed us to identify that the minimum inhibitory concentration (MIC) was equivalent to 250 ug/mL, while the concentration of 125 ug/mL was the most effective for conjugation, since there was no significant difference in reducing the count of E. coli (p = 0.42) or Salmonella Heidelberg (p = 0.46), and thus did not interfere with the results of the conjugations. At this concentration, we had a count of 6.71 ± 0.30 log UFC/mL in the control and 6.84 ± 0.37 log UFC/mL in the test with Lu 54 in E. coli, while in the test with S. Heidelberg we had 6.81 ± 0.58 log UFC/mL and 7.00 ± 0.52 log UFC/mL in the control and test, respectively.
Conjugation method 1 was selected for the tests with Lu54 due to its effectiveness in producing transconjugants. The use of this copper complex showed efficiency in controlling the conjugation process since there was a reduction in RCF from 2.2 to 0.3% in the traditional test, from 8.2 to 1.7% in conjugation with WH and from 6.2 to 0.9% in the presence of CJ, equivalent to a 7.3, 4.8 and 6.9-fold reduction, respectively. The values found also showed a statistical difference in transconjugant counts from 6.1 to 5.2 log UFC/mL in traditional conjugation, 6.5 to 6.0 log UFC/mL in the presence of WH and 6.5 to 5.6 log UFC/mL with CJ supplementation (GRAPH 3).
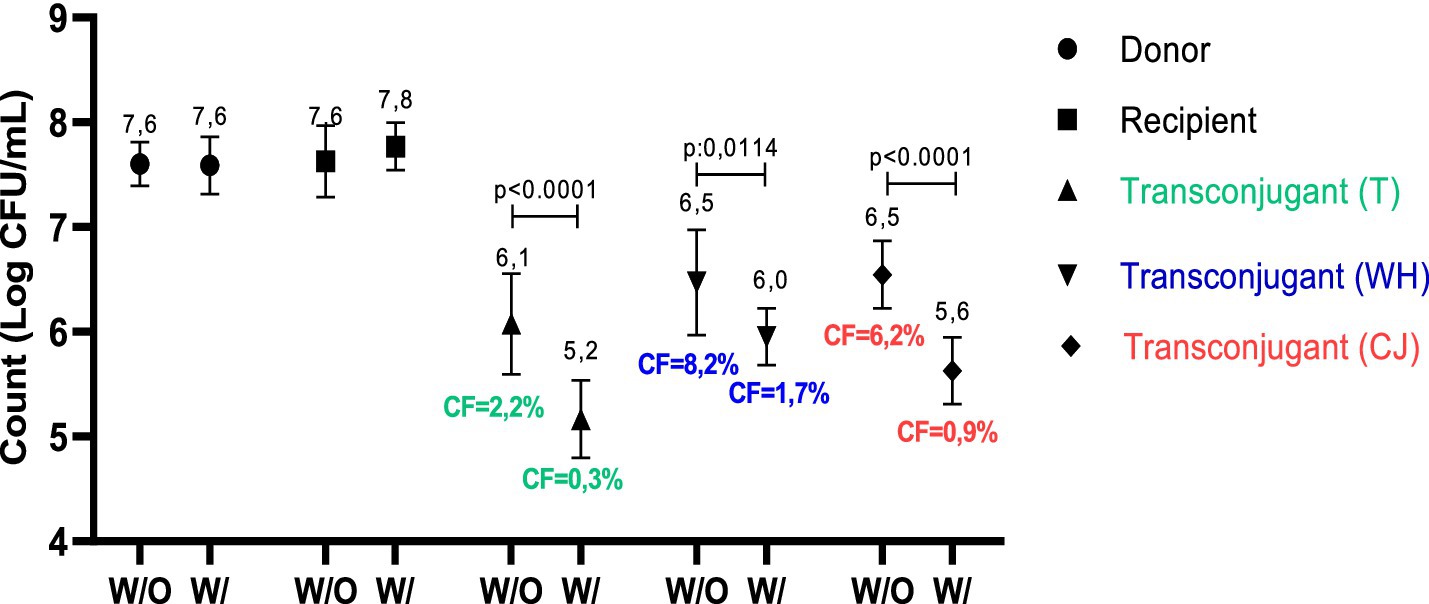
GRAPH 3. Comparison of counts and conjugation frequency obtained in conjugates with and without treatment with 125 μg/mL Lu54 [Cu(Clmp)(mTFpy)](PF6)2. W/O, Without Lu54 treatment; W/, With Lu54 treatment; T, traditional; WH: whey; CJ, chicken juice. CF, Conjugation frequency in percent. p < 0.0001 and p = 0.0114, statistical difference by T-test when comparing with and without Lu54 treatment.
The NCBI accession numbers for the deposited genome sequence data are provided in the Data Availability Statement. Whole genome sequencing (WGS). The six sequenced genomes had between 2 and 5 contigs (mean 3.83), with a total size between 4706500pb and 4778038pb (mean 4743274.67pb), completeness between 96.12 and 97.57% (mean 96.85%) and contamination between 0.38 and 0.69% (mean 0.53%), within the quality metrics (Supplementary Table 1). The species was confirmed, with the genomes showing >99% average nucleotide identity when compared to the reference genome.
The blaTEM-181 gene was identified in three genomes by Abricate, which were not treated with Lu54. RGI analysis identified blaTEM variants (blaTEM-1, blaTEM-91, blaTEM-98, blaTEM-144, blaTEM-186, blaTEM-219, blaTEM-232). The pipeline confirmed the results, and separated the transconjugants into two distinct clusters based on the presence of Lu54, showing that the transfer of blaTEM occurred only in the untreated samples, while the beta lactamase ampC was identified in all the transconjugants (Supplementary Table 2).
Fourteen plasmids were identified in the sequenced genomes, with each genome having between one and three plasmids (Supplementary Table 3). The blaTEM gene was identified in three plasmids, which were in the genomes of control E. coli (without CJ, WH and Lu54), control + CJ and control + WH. These plasmids were annotated with Bakta (Schwengers et al., 2021), and compared using clinker (Gilchrist and Chooi, 2021), to visualize blaTEM (Figure 1).
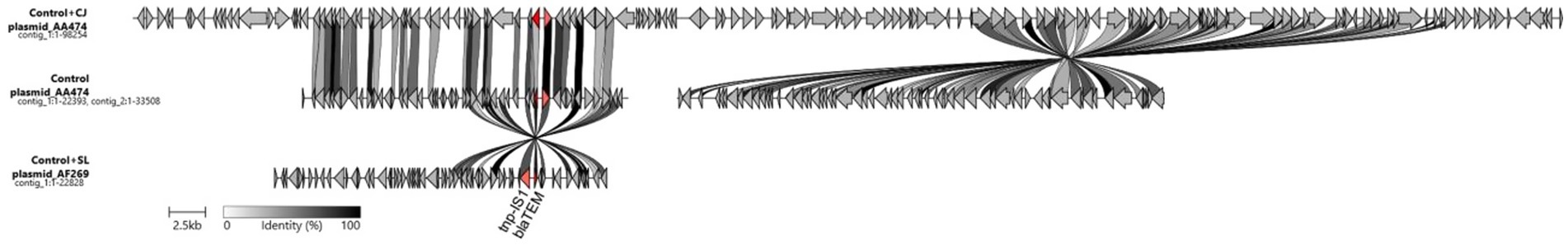
Figure 1. Comparison of plasmid contigs in which the blaTEM gene was identified. Control = control without the addition of Lu54; CJ = chicken juice; SL = whey; Tnp-IS1 = transposable element flanking the bla(TEM) gene. The colors of the bars linking the genes are based on the percentage of identity between the genes.
4 Discussion
The increase in antimicrobial resistance (AMR) has become a global public health problem, which makes it urgent to search for more in-depth knowledge about the mechanisms that affect and stimulate this phenomenon, as well as the search for new control alternatives (Frieri et al., 2017). In this context, conjugation is the most common form of recombination and dissemination of AMR in bacteria and is caused by the transfer of a gene from the donor bacterium to the recipient through direct contact (Neil et al., 2021). Transfer by conjugation can occur at different times, in different media and on different surfaces, such as the intestinal microbiota (Neil et al., 2021), soil (Tecon et al., 2018), aquatic conditions (Arias-Andres et al., 2018) and microbial biofilms (Águila-Arcos et al., 2017). There is no defined methodology for determining resistance gene transfer by conjugation, which prompted this investigation into the differences in blaTEM transfer in both liquid (at different times) and solid media.
Conjugation in liquid media probably favors greater exposure and immediate contact between the bacteria, which enhances the process, as we identified in method 1, which was the most efficient. This method also differed in that it used agitation, which, in addition to aeration, can improve access to nutrients and increase the frequency of collisions cell, as shown by Headd and Bradford (2018), who also observed that, in general, there was a higher frequency of conjugation in agitated cultures than in non-agitated cultures, although vigorous agitation can cause the conjugative pili to break. However, maintaining these bacteria overnight and removing moderate agitation (conjugation 3) can reduce the conjugation frequency (CF) due to extrinsic and intrinsic oscillations resulting from the maintenance medium, metabolism and bacterial physiology, such as stress caused by increased osmolarity, reduced nutritional availability and changes in the pH of the medium (Saliu et al., 2019; Kohyama and Suzuki, 2019; van Wonterghem et al., 2022).
We found that the average conjugation rates in solid/overnight media were lower than 1% and therefore lower than the other methods. By fixing the bacteria in a space/membrane, conjugation is restricted to neighboring cells and cell motility is reduced when compared to liquid media, which can hinder the frequency of gene exchanges. In addition, in relation to the incubation period, our results corroborate previous studies which found significant reductions in CF in experiments with a contact time of between 16 and 24 h compared to those with a time of less than 4 h (Alderliesten et al., 2020; Sakuda et al., 2018).
In addition to the differences between the times and media used, it is important to establish the interference that the presence of supplements can cause in the conjugation process. This study is one of the first to demonstrate the influence of the presence of whey and chicken juice - by- products routinely found in animal product processing industries - on bacterial conjugation.
In Brazil, this issue is even more relevant, considering that agriculture is one of the most important economic activities in the country, which ranks third in the world in meat production and second largest exporter, in addition to significant milk production (EMBRAPA, 2021). This role highlights the need to produce innocuous foods and to better understand the influence of by-products commonly present in the meat and cheese processing industries on the transmission and consequent spread of resistance genes.
In this context, WH and CJ were shown to enhance the transfer of the blaTEM gene by conjugation in liquid media. Both of the by-products used in this study are rich in nutrients, such as carbohydrates, lipids and proteins (Melo et al., 2017; Prazeres et al., 2012) which are widely used by bacteria for their metabolism (Kim and Gadd, 2008; Alderliesten et al., 2020), which certainly provides greater multiplication, survival and conditions to allow these microrganisms to express their potential. However, other circumstances can also influence the action of bacteria, such as incubation time, cell density, species used, the ratio between donor and recipient, temperature and changes in pH (Alderliesten et al., 2020).
Pallares-Vega et al. (2021) found that temperatures of 25 °C or less and low nutrient media can reduce the number of conjugative events. A similar situation occurs when there is an increase in the density of donors in relation to recipients (Chatterjee et al., 2013). The fact that the donor and recipient bacteria belong to the same species, when in a 1:1 ratio, both in liquid and filter media, can lead to an increase in the frequency of plasmid transfer. The same study also observed that, although the frequency of transfer was similar at both three and 16 h, the trend was lower in the longer period (Sakuda et al., 2018).
With regard to the influence of pH on the conjugation process, our study made it clear that reducing it to values between 5.2 and 6.5 in the presence of WH intensified the transfer of the blaTEM gene, while alkalinization caused lower production of transconjugants. In broth containing pH 5.0, at a temperature of 37 °C, the microorganisms obtain partial resistance to the acid in 5 mins and complete resistance in 10 min, which does not occur at a pH higher than 6.5. This habituation requires the synthesis of proteins and, when compared to organisms grown at pH 7.0, those grown at pH 5.0 have greater quantities of proteins, especially cytoplasmic proteins, which may be involved in protecting the cell against acid damage or in DNA repair (Raja et al., 1991). On the other hand, pH close to 8.0 can characterize a condition of alkaline stress, which in a study conducted by Cortés et al. (2016) represented a 36% decrease in the specific growth rates of E. coli when compared to those at pH 7.2. In addition, Van Wonterghem et al. (2022) pointed out that lowering the pH (from 7.6–8.0 to ~7.4) can boost conjugation. They also point out that pH dependence has been observed in the transfer of some plasmids, with pHs of around 6.0–7.5 being considered optimal for conjugation.
Also in this regard, the fact that the transfer of the blaTEM gene in liquid media containing whey is at least three times greater than in its absence is even more worrying, since dairy farms use antimicrobial treatments constantly, which can lead to microorganisms present in milk showing high levels of resistance. WH is abundantly present in many dairy products and it is also possible to infer that raw or pasteurized milk may exert the same tendency to increase conjugation rates. In agreement with this concern, a study conducted in China analyzing 350 raw milk samples from dairy farms found E. coli in 23.1% of the samples, with 33.3% of the isolates classified as multidrug resistant (MDR). Antimicrobial susceptibility testing revealed resistance to 92.9% of the 14 antibiotics tested, except meropenem, and notably, the blaTEM gene was present in 92.6% of the MDR isolates (Gao et al., 2025).
In addition, the use of raw milk for cheese making is a common practice, and the lack of heat treatment allows a greater number of bacteria, including those from dairy animals and the milking environment, to remain viable, which consequently amplifies the presence of resistance genes and increases the risk of horizontal gene transfer (Tóth et al., 2020; Marques et al., 2020). Supporting this, a study on small farms in Brazil detected pathogenic, commensal, and ceftriaxone-resistant (ESBL) E. coli at various points in milk collection and cheese production, including pathogenic strains of EPEC, STEC, and ExPEC. Where ExPEC or potentially ExPEC carried the blaTEM gene. Many isolates were resistant to antimicrobials such as nalidixic acid, ampicillin, kanamycin, streptomycin, sulfisoxazole and tetracycline. Genetic analysis revealed the presence of identical virulence genes at different stages of production within the same farm (Ribeiro et al., 2024).
At the same time, CJ also proved to enhance the process of gene exchange by conjugation, just as it had already intensified other bacterial perpetuation processes, such as the formation of biofilms. Melo et al. (2017) found that supplementation with CJ allowed viable cells to be maintained and strong biofilms to form, supporting the idea that this by-product promotes the survival and permanence of microorganisms in the production chain and can play an important role in food safety.
In addition, it is necessary to consider that the influence of these by-products on gene transfer can be considered a unique health problem, since, if not properly treated in the industry, the disposal of this waste in the environment further amplifies the spread of AMR in an environmental context, since animals, soil and aquatic organisms also contribute to the origin, maintenance and propagation of this phenomenon (Berendonk et al., 2015). In view of the above, it can be seen that horizontal gene transfer and antimicrobial resistance are global public health problems that can be exacerbated in conditions where animal products are processed. The emergence of antimicrobial resistance has led to difficulties in providing effective therapies and a lack of successful industrial hygiene prevention measures, which requires the development of new alternatives that can also prevent AMR (Frieri et al., 2017). In this way, we present an option for controlling the blaTEM transfer process using a copper complex.
In general, the antimicrobial potential of copper can occur through the generation of reactive oxygen species (ROS), which compromise proteins and DNA, or through oxidative damage to phospholipids in the bacterial membrane. In addition, these metals can interfere with the functioning of enzymes by competing with or displacing other ions needed in the active site or by binding covalently to functional groups in the active site, thus preventing their function and maintaining bacterial viability (Palm et al., 2022; Zhang et al., 2019).
In addition, the use of copper complexes can reduce or promote conjugation events. We found that the use of Lu54 was able to reduce the transfer of the blaTEM gene by conjugation at least 4.8 times, both in the presence and absence of supplements. These results were confirmed by whole genome sequencing analysis, where plasmids containing the blaTEM gene were identified only in control samples with or without supplements, but not in the presence of Lu54. Our results are corroborated by Buberg et al. (2020), who investigated the influence of copper on the conjugative transfer of a plasmid carrying the blaCMY-2 gene isolated from extended-spectrum betalactamase (ESBL)-producing E. coli from retail chicken meat, and found that there was at least a 40% reduction in transfer when compared to the control. They justified the reduction by the presence of copper, which is capable of causing disturbances in the function of the donor’s conjugative machinery by reducing the gene expression of traB and nikB, two important genes responsible for the ability to mobilize and transfer plasmids from one strain to another. However, these results contrast with a study that used E. coli K-12 as the donor, carrying the RP4 plasmid, which is resistant to kanamycin, ampicillin and tetracycline, and Pseudomonas putida KT2440 as the recipient. The use of Cu2+ led to greater production of ROS, expression of proteins involved in oxidative stress and increased permeability of the donor and recipient membranes, which promoted greater gene transfer by conjugation (Zhang et al., 2019). These differences are probably due to different metal concentrations and compositions, differences in experimental conditions, bacterial species and associated plasmids, as well as genes and proteins involved in conjugation.
WGS-supported analysis proved effective in differentiating the mobilome and families of β-lactamases, as well as highlighting the selective impact of Lu54 on mobile elements. The reduction in CF and the disappearance of blaTEM after treatment suggest that copper interferes with both the transfer machinery and the stability of plasmids, confirming its potential as a sanitary barrier in industrial environments. Our results revealed that by-products of animal origin favor the spread of blaTEM, while the Lu54 complex acts to reduce this spread and reorganize the plasmid resistome.
The difference in the blaTEM gene variants identified by Abricate and RGI demonstrates the greater specificity of the latter, which discriminated several variants based on specific mutations associated with distinct resistance profiles. In contrast, Abricate grouped these highly similar sequences under a single reference (blaTEM-181)The discrepancy between variants observed in RGI may be related to two main factors: (i) the emergence of variants during the experimental treatments with CJ and WH, or (ii) technical limitations inherent in the quality of the sequenced genomes, since it has already been shown that the quality of the data and the assembly method significantly interfere with the accurate identification of point mutations (Zankari et al., 2017). Despite these differences, both methods were concordant in identifying the blaTEM gene, confirming the presence of this family of β-lactamases in isolates not treated with Lu54.
From the results, it was concluded that the presence of whey and chicken juice, which mimic industrial conditions, is capable of stimulating the transfer of the blaTEM gene by conjugation, and the methodology using liquid medium, a few hours of incubation and agitation is the most effective for this event to occur. Furthermore, the use of the [Cu(Clmp)(mTFpy)](PF6)2 complex has proved to be a promising alternative for controlling gene exchange and mitigating antimicrobial resistance, especially in industrial environments where animal products are processed.
Data availability statement
Genome sequence data of the six E. coli isolates sequenced in this study are available in the National Center for Biotechnology Information Assembly/Genome database under BioProject accession number PRJNA1344634, biosamples SAMN52649862, SAMN52649863, SAMN52649864, SAMN52649865, SAMN52649866, SAMN52649867.
Author contributions
RR: Writing – review & editing, Methodology, Writing – original draft, Investigation, Conceptualization, Data curation. MG-T: Writing – original draft, Formal analysis, Methodology, Data curation, Validation. CD: Writing – review & editing, Methodology, Investigation, Formal analysis. AB: Formal analysis, Writing – review & editing, Software. BA: Methodology, Investigation, Writing – review & editing. TM: Writing – review & editing, Methodology. LR: Writing – review & editing, Resources. WG: Writing – review & editing, Resources. RP: Formal analysis, Writing – review & editing, Data curation. RM: Writing – review & editing, Conceptualization, Project administration, Supervision. DR: Supervision, Funding acquisition, Project administration, Writing – original draft, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was carried out with the support of the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior-Brasil (CAPES)-Funding Code 001. We would also like to thank CNPq-Funding Codes 405296/2021–5 and 303963/2024–7 for their financial support for this study.
Acknowledgments
We would like to thank the CAPES, CNPq and FAPEMIG.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1676649/full#supplementary-material
Footnotes
References
Abu Daher, G., Aoun, B., Jaafar, F., Khafaja, S., and Sanjad, S. (2019). Norovirus: a novel etiologic agent in hemolytic uremic syndrome in an infant. BMC Nephrol. 20, 1–4. doi: 10.1186/S12882-019-1427-6/FIGURES/1
Águila-Arcos, S., Álvarez-Rodríguez, I., Garaiyurrebaso, O., Garbisu, C., Grohmann, E., and Alkorta, I. (2017). Biofilm-forming clinical Staphylococcus isolates harbor horizontal transfer and antibiotic resistance genes. Front. Microbiol. 8:289347. doi: 10.3389/FMICB.2017.02018/BIBTEX
Alav, I., Pordelkhaki, P., Rodriguez-Navarro, J., Neo, O., Kessler, C., and Awodipe, R. J. (2024). Natural products from food sources can alter the spread of antimicrobial resistance plasmids in Enterobacterales. Microbiology 170:001496. doi: 10.1099/mic.0.001496
Alcock, B. P., Huynh, W., Chalil, R., Smith, K. W., Raphenya, A. R., Wlodarski, M. A., et al. (2023). CARD 2023: expanded curation, support for machine learning, and resistome prediction at the comprehensive antibiotic resistance database. Nucleic Acids Res. 51, D690–D699. doi: 10.1093/NAR/GKAC920
Alderliesten, J. B., Duxbury, S. J. N., Zwart, M. P., de Visser, J. A. G. M., Stegeman, A., and Fischer, E. A. J. (2020). Effect of donor-recipient relatedness on the plasmid conjugation frequency: a meta-analysis. BMC Microbiol. 20:135. doi: 10.1186/S12866-020-01825-4
Arias-Andres, M., Klümper, U., Rojas-Jimenez, K., and Grossart, H. P. (2018). Microplastic pollution increases gene exchange in aquatic ecosystems. Environ. Pollut. 237, 253–261. doi: 10.1016/J.ENVPOL.2018.02.058
Berendonk, T. U., Manaia, C. M., Merlin, C., Fatta-Kassinos, D., Cytryn, E., Walsh, F., et al. (2015). Tackling antibiotic resistance: the environmental framework. Nat. Rev. Microbiol. 13, 310–317. doi: 10.1038/nrmicro3439
Brown, H. L., Reuter, M., Salt, L. J., Cross, K. L., Betts, R. P., and van Vliet, A. H. M. (2014). Chicken juice enhances surface attachment and biofilm formation of Campylobacter jejuni. Appl. Environ. Microbiol. 80, 7053–7060. doi: 10.1128/AEM.02614-14
Buberg, M. L., Witsø, I. L., L’Abée-Lund, T. M., and Wasteson, Y. (2020). Zinc and copper reduce conjugative transfer of resistance plasmids from extended-spectrum beta-lactamase-producing Escherichia coli. Microb. Drug Resist. 26, 842–849. doi: 10.1089/MDR.2019.0388
CDC (2022). About Escherichia coli Infection | E. coli infection. Available online at: https://www.cdc.gov/ecoli/about/index.html (Accessed on July 28, 2025)
CDC (2019). Antibiotic Resistance Threats in the United States. Atlanta, GA: U.S. Department of Health and Human Services, CDC. doi: 10.15620/cdc:82532
Chatterjee, A., Cook, L. C. C., Shu, C. C., Chen, Y., Manias, D. A., Ramkrishna, D., et al. (2013). Antagonistic self-sensing and mate-sensing signaling controls antibiotic-resistance transfer. Proc. Natl. Acad. Sci. USA 110, 7086–7090. doi: 10.1073/PNAS.1212256110
Cortés, J. T., Flores, N., Bolívar, F., Lara, A. R., and Ramírez, O. T. (2016). Physiological effects of pH gradients on Escherichia coli during plasmid DNA production. Biotechnol. Bioeng. 113, 598–611. doi: 10.1002/BIT.25817
Coster, W. (2018). GitHub - wdecoster/NanoPlot: plotting scripts for long read sequencing data. NanoPlot. 34:2666–2669.
Delmani, F.-A., Jaran, A. S., Masaadeh, H., and Zaki, O. (2021). Characterization of ampicillin resistant gene (blaTEM-1) isolated from E. coli in northern Jordan. Asian J. Biomed. Pharmaceutical Sci. 7, 11–5. Available online at: https://scholar.google.com/scholar?hl=pt-BR&as_sdt=0%2C5&q=Characterization+of+Ampicillin+resistant+gene+%28blaTEM-1%29+Isolated+from+E.coli+in+northern+Jordan.&btnG=.
EFSA. Salmonella | EFSA. (2020). Available online at: https://www.efsa.europa.eu/en/topics/topic/salmonella (Accessed on July 28, 2025)
Elhariri, M., Elhelw, R., Selim, S., Ibrahim, M., Hamza, D., and Hamza, E. (2020). Virulence and antibiotic resistance patterns of extended-spectrum beta-lactamase-producing Salmonella enterica serovar Heidelberg isolated from broiler chickens and poultry workers: a potential hazard. Foodborne Pathog. Dis. 17, 373–381. doi: 10.1089/FPD.2019.2719
EMBRAPA. O agro no brasil e no mundo: uma síntese do período de 2000 a 2020 Adalberto Aragão e Elisio Contini Embrapa SIRE. (2021.). Available online at: https://www.embrapa.br/documents/10180/62618376/O+AGRO+NO+BRASIL+E+NO+MUNDO.pdf/41e20155-5cd9-f4ad-7119-945e147396cb (Accessed on July 28, 2025)
Etter, A. J., West, A. M., Burnett, J. L., Wu, S. T., Veenhuizen, D. R., Ogas, R. A., et al. (2019). Salmonella enterica subsp. enterica serovar Heidelberg food isolates associated with a salmonellosis outbreak have enhanced stress tolerance capabilities. Appl. Environ. Microbiol. 85:e01065-19. doi: 10.1128/AEM.01065-19
EUCAST. EUCAST broth microdilution reading guide updated. (2022). Available online at: https://www.eucast.org/eucast_news/news_singleview?tx_ttnews%5Btt_news%5D=362&cHash=f2196d399c8618b7313ccb7f048f7af5 (Accessed on July 28, 2025)
Frieri, M., Kumar, K., and Boutin, A. (2017). Antibiotic resistance. J. Infect. Public Health 10, 369–378. doi: 10.1016/j.jiph.2016.08.007
Gao, J., Wu, Y., Ma, X., Xu, X., Tuerdi, A., Shao, W., et al. (2025). Prevalent and drug-resistant phenotypes and genotypes of Escherichia coli isolated from healthy cow's milk of large-scale dairy farms in China. Int. J. Mol. Sci. 26:454. doi: 10.3390/ijms26020454
Gilchrist, C. L. M., and Chooi, Y. H. (2021). Clinker & clustermap.Js: automatic generation of gene cluster comparison figures. Bioinformatics 37, 2473–2475. doi: 10.1093/BIOINFORMATICS/BTAB007
Headd, B., and Bradford, S. A. (2018). Physicochemical factors that favor conjugation of an antibiotic resistant plasmid in non-growing bacterial cultures in the absence and presence of antibiotics. Front. Microbiol. 9:396093. doi: 10.3389/FMICB.2018.02122/BIBTEX
Huddleston, J. R. (2014). Horizontal gene transfer in the human gastrointestinal tract: potential spread of antibiotic resistance genes. Infection Drug Resistance 7, 167–176. doi: 10.2147/IDR.S48820
Ilangovan, A., Connery, S., and Waksman, G. (2015). Structural biology of the gram-negative bacterial conjugation systems. Trends Microbiol. 23, 301–310. doi: 10.1016/j.tim.2015.02.012
Jain, C., Rodriguez-R, L. M., Phillippy, A. M., Konstantinidis, K. T., and Aluru, S. (2018). High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 9, 1–8. doi: 10.1038/S41467-018-07641-9
Jia, Y., Zheng, Z., Yang, B., Zhang, H., Wang, Z., and Liu, Y. (2024). A broad-Spectrum horizontal transfer inhibitor prevents transmission of plasmids carrying multiple antibiotic resistance genes. Transbound. Emerg. Dis. 2024:7063673. doi: 10.1155/2024/7063673
Kerr, D., Vohra, R., Chandra, M., Ballard, S. A., Martin, N., Lan, R., et al. (2022). A multijurisdictional outbreak of Salmonella enterica serovar Heidelberg, Australia, November 2018–May 2019. Emerg. Infect. Dis. 28, 53–61. doi: 10.3201/eid2801.211462
Kim, B. H., and Gadd, G. M. (2008). Bacterial Physiology and Metabolism. United States of America, New York: Cambridge University Press.
Kohyama, Y., and Suzuki, S. (2019). Conjugative gene transfer between nourished and starved cells of photobacterium damselae ssp. Damselae and escherichia coli. Microbes Environ. 34, 388–392. doi: 10.1264/JSME2.ME19099
Krasnovskaya, O., Naumov, A., Guk, D., Gorelkin, P., Erofeev, A., Beloglazkina, E., et al. (2020). Copper coordination compounds as biologically active agents. Int. J. Mol. Sci. 21:3965. doi: 10.3390/IJMS21113965
Lamichhane, B., Mawad, A. M. M., Saleh, M., Kelley, W. G., Harrington, P. J., Ii,, et al. (2024). Salmonellosis: an overview of epidemiology, pathogenesis, and innovative approaches to mitigate the antimicrobial resistant infections. Antibiotics 2024:76. doi: 10.3390/antibiotics13010076
Lee, C. A., and Grossman, A. D. (2007). Identification of the origin of transfer (oriT) and DNA relaxase required for conjugation of the integrative and conjugative element ICEBs1 of Bacillus subtilis. J. Bacteriol. 189, 7254–7261. doi: 10.1128/JB.00932-07
MAPA Resistência aos antimicrobianos — Ministério da Agricultura e Pecuária. (2022). Available online at: https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/insumos-pecuarios/resistencia-aos-antimicrobianos (Accessed on July 28, 2025)
Marques, K. H. D. S. R., Carvalho, J. B., Mesquita, A. A., Silva, J. A. G. E., Ferreira, S. V., Sousa, T. L., et al. (2020). Influência do tratamento térmico no rendimento e composição físico-química de queijos frescais / influence of thermal treatment in the yield and physical-chemical composition of fresh cheeses. Brazilian J. Develop. 6, 19446–19460. doi: 10.34117/BJDV6N4-203
Melo, R. T., Galvão, N. N., Guidotti-Takeuchi, M., Peres, P. A. B. M., Fonseca, B. B., Profeta, R., et al. (2021). Molecular characterization and survive abilities of Salmonella Heidelberg strains of poultry origin in Brazil. Front. Microbiol. 12:674147. doi: 10.3389/FMICB.2021.674147/XML
Melo, R. T., Mendonça, E. P., Monteiro, G. P., Siqueira, M. C., Pereira, C. B., Peres, P. A. B. M., et al. (2017). Intrinsic and extrinsic aspects on Campylobacter jejuni biofilms. Front. Microbiol. 8:268085. doi: 10.3389/FMICB.2017.01332/BIBTEX
Mikheenko, A., Prjibelski, A., Saveliev, V., Antipov, D., and Gurevich, A. (2018). Versatile genome assembly evaluation with QUAST-LG. Bioinformatics 34, i142–i150. doi: 10.1093/BIOINFORMATICS/BTY266
Monteiro, G. P., de Melo, R. T., Guidotti-Takeuchi, M., Dumont, C. F., Ribeiro, R. A. C., Guerra, W., et al. (2022). A ternary copper (II) complex with 4-fluorophenoxyacetic acid hydrazide in combination with antibiotics exhibits positive synergistic effect against Salmonella Typhimurium. Antibiotics 11:388. doi: 10.3390/ANTIBIOTICS11030388/S1
Mo, S. S., Sunde, M., Ilag, H. K., Langsrud, S., and Heird, E. (2017). Transfer potential of plasmids conferring extended-spectrum-cephalosporin resistance in Escherichia coli from poultry. Appl. Environ. Microbiol. 83, 654–671. doi: 10.1128/AEM.00654-17/SUPPL_FILE/ZAM999117885S1.PDF
Murray, C. J., Ikuta, K. S., and Sharara, F. (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399, 629–655. doi: 10.1016/S0140-6736(21)02724-0
NCBI. National Center for biotechnology information. Ampicillin | C16H19N3O4S | CID 6249 - PubChem. (2022). Available online at: https://pubchem.ncbi.nlm.nih.gov/compound/Ampicillin (Accessed on July 28, 2025)
Neil, K., Allard, N., and Rodrigue, S. (2021). Molecular Mechanisms Influencing Bacterial Conjugation in the Intestinal Microbiota. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.673260
Ngece, K., Khwaza, V., Paca, A. M., and Aderibigbe, B. A. (2025). The antimicrobial efficacy of copper complexes: a review. Antibiotics 14:516. doi: 10.3390/antibiotics14050516
Ott, L. C., and Mellata, M. (2024). Short-chain fatty acids inhibit bacterial plasmid transfer through conjugation in vitro and in ex vivo chicken tissue explants. Front. Microbiol. 15:1414401. doi: 10.3389/fmicb.2024.1414401
Pallares-Vega, R., Macedo, G., Brouwer, M. S. M., Hernandez Leal, L., van der Maas, P., van Loosdrecht, M. C. M., et al. (2021). Temperature and nutrient limitations decrease transfer of conjugative IncP-1 plasmid pKJK5 to wild Escherichia coli strains. Front. Microbiol. 12:656250. doi: 10.3389/FMICB.2021.656250/BIBTEX
Palm, M., Fransson, A., Hultén, J., Búcaro Stenman, K., Allouche, A., Chiang, O. E., et al. (2022). The effect of heavy metals on conjugation efficiency of an F-plasmid in Escherichia coli. Antibiotics 11:1123. doi: 10.3390/ANTIBIOTICS11081123/S1
Parks, D. H., Imelfort, M., Skennerton, C. T., Hugenholtz, P., and Tyson, G. W. (2015). CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 25, 1043–1055. doi: 10.1101/GR.186072.114
Prazeres, A. R., Carvalho, F., and Rivas, J. (2012). Cheese whey management: a review. J. Environ. Manag. 110, 48–68. doi: 10.1016/J.JENVMAN.2012.05.018
Raja, N., Goodson, M., Chui, W. C. M., Smith, D. G., and Rowbury, R. J. (1991). Habituation to acid in Escherichia coli: conditions for habituation and its effects on plasmid transfer. J. Appl. Bacteriol. 70, 59–65. doi: 10.1111/J.1365-2672.1991.TB03787.X
Reygaert, C. (2018). An overview of the antimicrobial resistance mechanisms of bacteria. AIMS Microbiol. 4, 482–501. doi: 10.3934/MICROBIOL.2018.3.482
Ribeiro, L. F., Rossi, G. A. M., Rafael Akira Sato,, de Andressa Souza Pollo,, Marita Vedovelli Cardozo,, Luiz Augusto do Amaral,, et al. (2024). Epidemiology, virulence and antimicrobial resistance of Escherichia coli isolated from small Brazilian farms producers of raw milk fresh cheese. Microorganisms 12:1739. doi: 10.3390/microorganisms12081739
Robertson, J., and Nash, J. H. E. (2018). MOB-suite: software tools for clustering, reconstruction and typing of plasmids from draft assemblies. Microbial Genomics 4:e000206. doi: 10.1099/MGEN.0.000206
Rodriguez-Grande, J., and Fernandez-Lopez, R. (2020). Measuring plasmid conjugation using antibiotic selection. Methods Mol. Biol. 2075, 93–98. doi: 10.1007/978-1-4939-9877-7_6
Sakuda, A., Suzuki-Minakuchi, C., Okada, K., and Nojiri, H. (2018). Conjugative selectivity of plasmids is affected by coexisting recipient candidates. MSphere 3, 10–1128. doi: 10.1128/MSPHERE.00490-18
Salah, I., Parkin, I. P., and Allan, E. (2021). Copper as an antimicrobial agent: recent advances. RSC Adv. 11, 18179–18186. doi: 10.1039/D1RA02149D
Saliu, E. M., Eitinger, M., Zentek, J., and Vahjen, W. (2019). Nutrition related stress factors reduce the transfer of extended-Spectrum Beta-lactamase resistance genes between an Escherichia coli donor and a Salmonella Typhimurium recipient in vitro. Biomolecules 9:324. doi: 10.3390/BIOM9080324
Schmidt, M. B., Geissler, A. L., Edwards, J. R., Cieslak, P. R., Griffin, P. M., and Mahon, B. E. (2023). Epidemiology of salmonellosis among infants in the United States, 1968–2015. Pediatr. Infect. Dis. J. 42, 553–559. doi: 10.1097/INF.0000000000003830
Schwengers, O., Jelonek, L., Dieckmann, M. A., Beyvers, S., Blom, J., and Goesmann, A. (2021). Bakta: rapid and standardized annotation of bacterial genomes via alignment-free sequence identification. Microbial Genomics 7:000685. doi: 10.1099/MGEN.0.000685
Shintani, M., Ohkuma, M., and Kimbara, K. (2019). High-resolution comparison of bacterial conjugation frequencies. J. Vis. Exp. 143:e57812. doi: 10.3791/57812
Srichumporn, W., Chaisowwong, W., Intanon, M., and Na-Lampang, K. (2022). Extended-spectrum beta-lactamase-producing Escherichia coli from pork in Muang district, Chiang Mai Province, Thailand. Vet. World 15, 2903–2909. doi: 10.14202/VETWORLD.2022.2903-2909
Stern, B. R. (2010). Essentiality and toxicity in copper health risk assessment: overview, update and regulatory considerations. J. Toxicol. Environ. Health A 73, 114–127. doi: 10.1080/15287390903337100
Sultan, I., Rahman, S., Jan, A. T., Siddiqui, M. T., Mondal, A. H., and Haq, Q. M. R. (2018). Antibiotics, resistome and resistance mechanisms: a bacterial perspective. Front. Microbiol. 9:377549. doi: 10.3389/FMICB.2018.02066/XML
Takeuchi, M. G., Ferreira, A. L. M., Ramos, L. M. S., Peixoto, J. L. M., Chueiri, M. C., Dumont, C. F., et al. (in press). Anti-Campylobacter activity of ternary copper(II) complexes with imine ligands and 4′-(4-methylphenyl)-2,2′:6′,2′′-terpyridine. ACS Omega.
Takeuchi, MG, Rossi, DA, and Ramos, LMS. Synthesis, design and in vitro activity of new copper compounds against Campylobacter spp.
Taniwaki, M. H., Abeilar, R., Gomes, R., and Okazaki, M. M. (2017). Neusely da Silva Valéria Christina Amstalden Junqueira Neliane Ferraz de Arruda Silveira. Manual de métodos de análise microbiológica de alimentos e água. Ed. Edgard Blücher Ltda. (Book) 5ª ed. – São Paulo. 560:il, Available online at: https://www.blucher.com.br
Tecon, R., Ebrahimi, A., Kleyer, H., Levi, S. E., and Or, D. (2018). Cell-to-cell bacterial interactions promoted by drier conditions on soil surfaces. Proc. Natl. Acad. Sci. USA 115, 9791–9796. doi: 10.1073/PNAS.1808274115
Tóth, A. G., Csabai, I., Krikó, E., Tőzsér, D., Maróti, G., Patai, Á. v., et al. (2020). Antimicrobial resistance genes in raw milk for human consumption. Sci. Rep. 10, 1–7. doi: 10.1038/s41598-020-63675-4
van Wonterghem, L., de Chiara, M., Liti, G., Warringer, J., Farewell, A., Verstraeten, N., et al. (2022). Genome-wide association study reveals host factors affecting conjugation in Escherichia coli. Microorganisms 10:608. doi: 10.3390/MICROORGANISMS10030608/S1
Vaser, R., and Šikić, M. (2021). Raven: a de novo genome assembler for long reads. BioRxiv 2020:07.242461. doi: 10.1101/2020.08.07.242461
von Wintersdorff, C. J. H., Penders, J., van Niekerk, J. M., Mills, N. D., Majumder, S., van Alphen, L. B., et al. (2016). Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 7:174871. doi: 10.3389/FMICB.2016.00173/XML
Wick, R. (2021a). GitHub - rrwick/Filtlong: quality filtering tool for long reads. Git Hub repository. Available online at: https://github.com/rrwick/Filtlong (Accessed on May 25, 2025).
Wick, R. (2021b). Porechop/porechop/porechop.Py at master · rrwick/Porechop · GitHub. Git hub repository. Available online at: https://github.com/rrwick/Porechop/blob/master/porechop/porechop.py (Accessed on May 25, 2025).
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Completing bacterial genome assemblies with multiplex MinION sequencing. Microbial Genomics, 3. doi: 10.1099/mgen.0.000132
Zalevskaya, O. A., and Gur’eva, Y. A. (2021). Recent studies on the antimicrobial activity of copper complexes. Russian J. Coordination Chemistry/Koordinatsionnaya Khimiya 47, 861–880. doi: 10.1134/S1070328421120046/METRICS
Zankari, E., Allesøe, R., Joensen, K. G., Cavaco, L. M., Lund, O., and Aarestrup, F. M. (2017). PointFinder: a novel web tool for WGS-based detection of antimicrobial resistance associated with chromosomal point mutations in bacterial pathogens. J. Antimicrob. Chemother. 72, 2764–2768. doi: 10.1093/JAC/DKX217
Keywords: bacterial conjugation, whey, chicken juice, copper(II) complex, blaTEM
Citation: Ribeiro RAC, Guidotti-Takeuchi M, Dumont CF, Buiatte ABG, de Araújo Brum B, Martins TJ, Ramos LMS, Guerra W, Polveiro RC, de Melo RT and Rossi DA (2025) Transfer of blaTEM gene between Salmonella and Escherichia coli under processing conditions of animal products: influence of a copper(II) complex. Front. Microbiol. 16:1676649. doi: 10.3389/fmicb.2025.1676649
Edited by:
Daniel F. M. Monte, North Carolina State University, United StatesReviewed by:
Isabella Cardeal Campos, São Paulo State University, BrazilSahr Ilyas, University of Karachi, Pakistan
Copyright © 2025 Ribeiro, Guidotti-Takeuchi, Dumont, Buiatte, de Araújo Brum, Martins, Ramos, Guerra, Polveiro, de Melo and Rossi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Roberta Torres de Melo, cm9iZXJ0YS1tZWxvQGhvdG1haWwuY29t
†ORCID: Rosanne Aparecida Capanema Ribeiro, orcid.org/0000-0002-7337-3284
Bárbara de Araújo Brum, orcid.org/0000-0001-8004-8247
Luana Munique Sousa Ramos, orcid.org/0000-0001-7411-069X
Wendell Guerra, orcid.org/0000-0001-6979-8557
Thais Jansen Martins, orcid.org/0009-0008-5288-617X