 Chrisse Ngari
Chrisse Ngari Valérie Poulet
Valérie Poulet Giuseppe Percoco
Giuseppe Percoco Laurent Peno-Mazzarino
Laurent Peno-Mazzarino Damien Seyer
Damien Seyer- 1Laboratoires Clarins, Pontoise, France
- 2Eurofins BIO-EC, Longjumeau, France
- 3ERRMECe Laboratory, CY Cergy-Paris Université, Cergy-Pontoise, France
Introduction: The skin microbiome is crucial for skin health and homeostasis. It contributes to immune defense, promotes epidermal differentiation, and supports a robust skin barrier function. Despite its importance, few studies have used model skin systems to examine how simple bacterial communities interact and how they impact the skin.
Methods: We studied the interaction between a skin commensal, Staphylococcus epidermidis, and a pathogen, Staphylococcus aureus, by performing inoculations and co-inoculations on Reconstructed Human Epidermis (RHE) models maintained under classical humid conditions (>90% relative humidity) or dry conditions (<25% RH). In parallel, inoculations were conducted on human skin explants using a novel culture setup preserving physiological humidity levels (40–60% RH). Bacterial attachment was assessed 4 h post-inoculation. At 24 h, histology was examined, three natural moisturizing factors (NMFs) were quantified. Filaggrin (FLG) and ceramide levels were analyzed to assess the skin barrier function.
Results: In contrast to previous findings at 24 h, co-inoculation increased S. epidermidis and S. aureus attachment in RHEs, but only under humid conditions. Only RHE maintained in dry conditions and skin explant revealed an effect of co-inoculation on filaggrin (FLG), yet an increase in RHEs and a decrease in skin explants. In both RHEs maintained in dry conditions and skin explants, NMF levels were consistently reduced following co-inoculation. In RHEs and skin explants, inoculation with S. aureus alone also lowered NMFs, with co-inoculation further amplifying this effect. Finally, ceramide levels increased similarly across both inoculations and co-inoculation.
Discussion: Revealing unexpected early interactions between S. epidermidis, S. aureus, and the skin, our results could suggest that co-inoculation may trigger a synergetic disruption of the barrier function. Alternatively, it could imply that co-inoculation might reinforce the epidermal barrier via a S. aureus-mediated stimulation of the protective functions of S. epidermidis. Further studies are needed to confirm these effects and determine whether they are strain-specific or more broadly applicable.
1 Introduction
As a continuous organ covering the entire body, the skin serves as a critical barrier between the internal and external environments. This barrier function primarily relies on the stratum corneum (SC), the outermost skin layer, which fulfills physical, chemical, and microbial protective functions (Lefèvre-Utile et al., 2021; de Szalay and Wertz, 2023). The distinctive structure and composition of the SC, consisting of terminally differentiated keratinocytes – the corneocytes – embedded in a highly organized lipidic lamellar matrix, largely account for this protective mechanism (Elias, 1983; Baroni et al., 2012).
Filaggrin (FLG), a major constituent of corneocytes, is pivotal to the barrier function (Drislane and Irvine, 2020). Initially present as pro-FLG, enzymatic processing leads to FLG that aggregates with keratin filaments (Briot et al., 2020; Quiroz et al., 2020). This network binds to the keratinocyte membrane, resulting in its collapse and the flattened corneocyte shape that contributes to the physical strength of the external skin layer. As maturation proceeds, FLG undergoes fragmentation into hygroscopic amino acids and their metabolites, the natural moisturizing factors (NMFs), which are essential for SC hydration (Rawlings and Harding, 2004). Proper hydration ensures corneocyte maturation, desquamation, and SC homeostasis (Rawlings and Harding, 2004). It is under the tight control of internal factors and external relative humidity, both of which regulate FLG degradation into NMFs (Cau et al., 2017). Skin hydration is also critical for SC flexibility and contributes to skin suppleness. It also has important cosmetic properties, dry skin being rough, scaly, flaky, and itchy.
In addition to these endogenous mechanisms, the skin microbiome, comprising bacteria, fungi, viruses, and microeukaryotes, has emerged as a key player in skin health and homeostasis (Flowers and Grice, 2020). It is involved in skin defense and immune responses. Bacteria such as Staphylococcus epidermidis, one of the most common skin commensals, can produce toxins that, along with antimicrobial skin peptides, selectively kill pathogens such as Staphylococcus aureus (Cogen et al., 2010a, 2010b). The microbiome also stimulates genes from the epidermal differentiation complex, promoting epidermal differentiation and participating in the skin barrier function (Meisel et al., 2018). Furthermore, S. epidermidis increases the skin’s lipid content, thereby limiting water evaporation and improving skin hydration (Nodake et al., 2015). S. epidermidis also secretes lipases that hydrolyze sebum and can ferment glycerol. Both processes produce organic fatty acids, especially lactic acid, which help maintain acidic conditions (Nodake et al., 2015). These acidic conditions contribute to the inhibition of pathogenic bacteria, and lactic acid helps improve skin hydration. Conversely, a bacterium such as S. aureus downregulates FLG expression, thereby affecting skin hydration. Moreover, FLG deficiency favors S. aureus colonization, leading to a vicious cycle (Kim and Lim, 2021).
While numerous studies have evaluated the diversity of the skin microbiome, very few have investigated inter-species interactions. An early work used immobilized keratinocytes and simplified commensal-pathogen community models to demonstrate that commensals restrict pathogen growth and biofilm formation, thus limiting damages to keratinocytes (Jordana-Lluch et al., 2020). Although reconstructed human epidermis (RHE) models reflect the skin and its epidermal barrier, only a few studies have analyzed single-species colonization (Rademacher et al., 2018; Ochlich et al., 2023). A single work took advantage of RHE to investigate a commensal-pathogen community (Kohda et al., 2021), showing that S. epidermidis restricts S. aureus colonization, limits S. aureus-induced cytotoxicity and secretion of proinflammatory cytokines, also preventing S. aureus from entering deep cell layers.
While RHE is a model of choice, it suffers some limitations. In particular, the SC present an altered lipid composition and organization, resulting in a weakened barrier function (Thakoersing et al., 2012). While skin explants do not suffer these limitations, there are maintained under a minimum of 90% relative humidity and 5% carbon dioxide, which are far from physiological conditions. In addition, they are maintained entirely at 37 °C, not presenting the 32–37 °C temperature gradient that exist in vivo and that is important for a normal hydration of the SC as well as for the proper proliferation and differentiation of keratinocytes (Spencer et al., 1975; Borowiec et al., 2013). To circumvent these limitations, a new culture system, the Perfex setup, was recently developed, enabling to maintain skin explants in conditions very close to physiological conditions (Peno-Mazzarino et al., 2024).
Thus, to investigate the interaction between S. epidermidis, S. aureus, and the skin, we used RHEs that were maintained under the “classical” humid conditions (37 °C, 5% CO2, >90% relative humidity), also assessing the effects of dry conditions closer to “normal” conditions (37 °C, 5% CO2, <25% relative humidity). We also took advantage of the new Perfex setup that enable maintaining skin explants under physiological conditions (32 °C, 0.04% CO2 and 40–60% relative humidity). In all these models, we evaluated bacteria attachment and analyzed hydration and barrier function-related markers in RHEs upon S. epidermidis or S. aureus inoculation and S. epidermidis - S. aureus co-inoculation.
2 Materials and methods
2.1 Bacterial strains and growth conditions
Staphylococcus epidermidis and Staphylococcus aureus strains producing easily distinguishable colonies in terms of size and color, were selected. S. epidermidis was either the strain Fussel 14990™ (RHE experiments) or CIP 107777 (Perfex skin explant experiments). Despite some differences, both strains are classical model strains of S. epidermidis commensals. For the inoculation of both RHEs and skin explants, the Staphylococcus aureus subsp. aureus ATCC 6538™ was used. All strains were stored at −80 °C and subcultured at 37 °C on Tryptone Soy Agar plates or in Tryptone Soy Broth.
2.2 Reconstituted human epidermis (RHE)
For RHEs, primary human keratinocytes from an adult donor were cultured on 0.6 cm2 porous polycarbonate inserts (Millipore, #PIHP01250) before being exposed to the air-liquid interface under either classical humid conditions (37 °C, 5% CO2, >90% relative humidity) or dry conditions (37 °C, 5% CO2, <25% relative humidity). These humid or dry conditions were maintained thorough the experiment.
RHE were cultured for 8 d in optimized EpiLife® culture medium (Gibco, #MEPI500CA), supplemented with the Human Keratinocyte Growth Supplement kit (Gibco, #S0015) and penicillin/streptomycin (Sigma, #P433). The medium was renewed daily and, on day 8, replaced with antibiotic- and hydrocortisone-free EpiLife® medium. After 24 h (day 9), RHEs were inoculated and used for experiments.
Inoculation and subsequent analysis were conducted on three different RHEs from the same batch per condition and time point.
2.3 Perfex skin explants
A skin explant was obtained from an elective abdominoplasty performed on a 28-year-old Caucasian woman. Its procurement strictly complied Articles L.1245–2 and L.1211–1 of the French Public Health Code. Written informed consent was obtained from the donor.
After removal of the hypodermis, explant fragments were maintained using the Perfex setup (Peno-Mazzarino et al., 2024). Briefly, the dermal side was perfused with buffered, temperature-controlled BEM medium (BIO-EC’s Explant Medium), while the epidermal surface was exposed to “normal” conditions (32 °C, 0.04% CO2 and 40–60% relative humidity). Explants were inoculated after 3 d and used for experiments.
Inoculations were conducted on two different skin explants from the same donor per condition and time point, each being divided in three to conduct subsequent analysis in triplicates.
2.4 Bacterial inoculation and incubation
Overnight liquid bacterial cultures, in early stationary phases, were washed and resuspended in PBS buffer. For mono-inoculations, 1.5×105 CFU/mm2 were deposited onto the surface of RHEs or skin explants. In the case of the strain mixture, both strains were applied at a 1:1 ratio, each at 1.5×105 CFU/mm2.
S. epidermidis strain Fussel (14990™) and/or S. aureus subsp. aureus Rosenbach were used to inoculate RHEs. S. epidermidis CIP 4.83 and/or S. aureus strain ATCC 10577 were used to inoculate Perfex skin explants. Inoculum size was confirmed by plate count. Application of PBS buffer alone served as a negative control.
Inoculated RHE were incubated in dry (37 °C, 5% CO2, <25% relative humidity) or humid (37 °C, 5% CO2, >90% relative humidity) conditions. Perfex skin explants were incubated under “normal” conditions (32 °C, 0.04% CO2 and 40–60% relative humidity). The incubation was stopped after four and 24 h. Prior to further analysis, non-adherent bacteria were removed by four consecutive PBS buffer washes.
2.5 Bacterial numeration
RHEs were homogenized using the Percellys® system (Bertin Technologies). The surface of skin explants was gently scraped with a scalpel blade while washed with PBS. Serial dilutions of RHE homogenates and skin surface washings were plated on Tryptone Soy Agar. Colony-forming units (CFUs) were numerated after overnight incubation at 37 °C. S. epidermidis and S. aureus colonies were identified based on the very different colony morphology of the strains used, confirmed, in part of the experiments, using CHROMagar™ S. aureus.
Numeration results were obtained from a single bacterial enumeration performed 4 h after inoculation on three independent RHEs or on three fragments of the same skin explant 4 h after inoculation.
2.6 Histological analysis of RHE
Twenty-four hours after inoculation, RHEs and skin explants were fixed in formaldehyde, dehydrated in ethanol, and embedded in paraffin. Five-micrometer-thick cross-sections were stained with hematoxylin and eosin (RHEs) or with Masson’s trichrome staining (skin explants). Histology was evaluated using the Olympus SLIDEVIEW™ VS200 slide scanner and analyzed with the OlyVIA software (Olympus). Alternatively, a Leica DMLB or an Olympus BX63 microscope equipped with a DP Olympus digital camera were used. Analysis was conducted using the CellSens software (Olympus). A minimum of three representative images per replicate were assessed.
2.7 Analysis of NMFs
Among the various NMFs derived from FLG degradation, analyses focused on serine, pyrrolidone carboxylic acid (PCA), and urocanic acid. Analytes were extracted from RHEs using an aqueous extraction.
NMFs were quantified using an UltiMate™ 3,000 liquid chromatography system (Thermo Scientific, CA, USA) coupled with an ISQ EM mass detector (Thermo Scientific, CA, USA). Mobile phases [M1: water + 0.1% formic acid] and [M2: methanol + 0.1% formic acid] were eluted at a 0.4 mL/min flow rate. The injection volume was 20 μL, and the column temperature was maintained at 40 °C. For MS detection, Positive Electrospray Ionization (ESI+) was used. Data were analyzed using the Chromeleon™ 7.2 software (Thermo Fisher Scientific, MA, USA).
Results were obtained from measurements (conducted 24 h after bacterial inoculation) on three independent RHEs (each with duplicate technical repeats) or on three fragments of the same skin explant (each with triplicate technical repeats).
2.8 Analysis of pro-filaggrin and filaggrin proteins from RHEs
RHE or skin explant in RIPA buffer (Sigma, #R0278) supplemented with a protease inhibitor cocktail (Roche, #5892791001) were homogenized using the Precellys® system (Bertin Technologies). Following centrifugation, the supernatants were analyzed using the Wes™ capillary electrophoresis system (ProteinSimple). Proteins were separated by molecular weight, immobilized via UV, and total protein quantified using a specific dye. Filaggrin (FLG) monomers (50 kDa) were detected using a mouse anti-FLG primary antibody (Santa Cruz, #sc-66192) and a goat anti-mouse peroxidase-conjugated secondary antibody (ProteinSimple, #DM-002).
Quantifications were performed 24 h after bacterial inoculation on three independent RHEs, with each measurement carried out in duplicate technical repeats.
2.9 Immunohistochemical analysis of filaggrin, loricrin, and ceramides from skin explants
Formalin-fixed, paraffin-embedded skin explant sections were used for immunostaining. FLG was detected with a mouse monoclonal primary antibody (Santa Cruz, #sc-66192) and an Alexa Fluor™ 488-conjugated goat anti-mouse secondary antibody (Invitrogen, #A-11001). Ceramides were detected using a monoclonal anti-ceramide antibody (Glycobiotech, #MAB_0013). Detection relied on a biotin/streptavidin amplification system, revealed with the Vector® VIP peroxidase substrate (Vector Laboratories, #SK-4600). After image acquisition with an Olympus BX63 microscope equipped with an Olympus DP camera, signal intensity was quantified using the CellSens software (Olympus).
Immunohistochemical labeling was quantified from at least three representative fields in cross-sections of three fragments of the same skin explant.
2.10 Statistical analysis
All results are expressed as mean ± standard deviation. Normal data distribution was assessed using the Shapiro–Wilk tests (α = 0.01). Statistical comparisons were performed using Student’s t-tests or one-way ANOVA, followed by pairwise Tukey HSD tests. p-values below 0.05 were considered statistically significant, and those between 0.1 and 0.5 at the significance limit.
3 Results
3.1 Colonization of RHEs and skin explants by bacteria does not affect their morphology
As a preliminary step, we used histological staining to ascertain that bacteria colonization had no observable impact on RHEs or skin explants morphology (Figure 1).
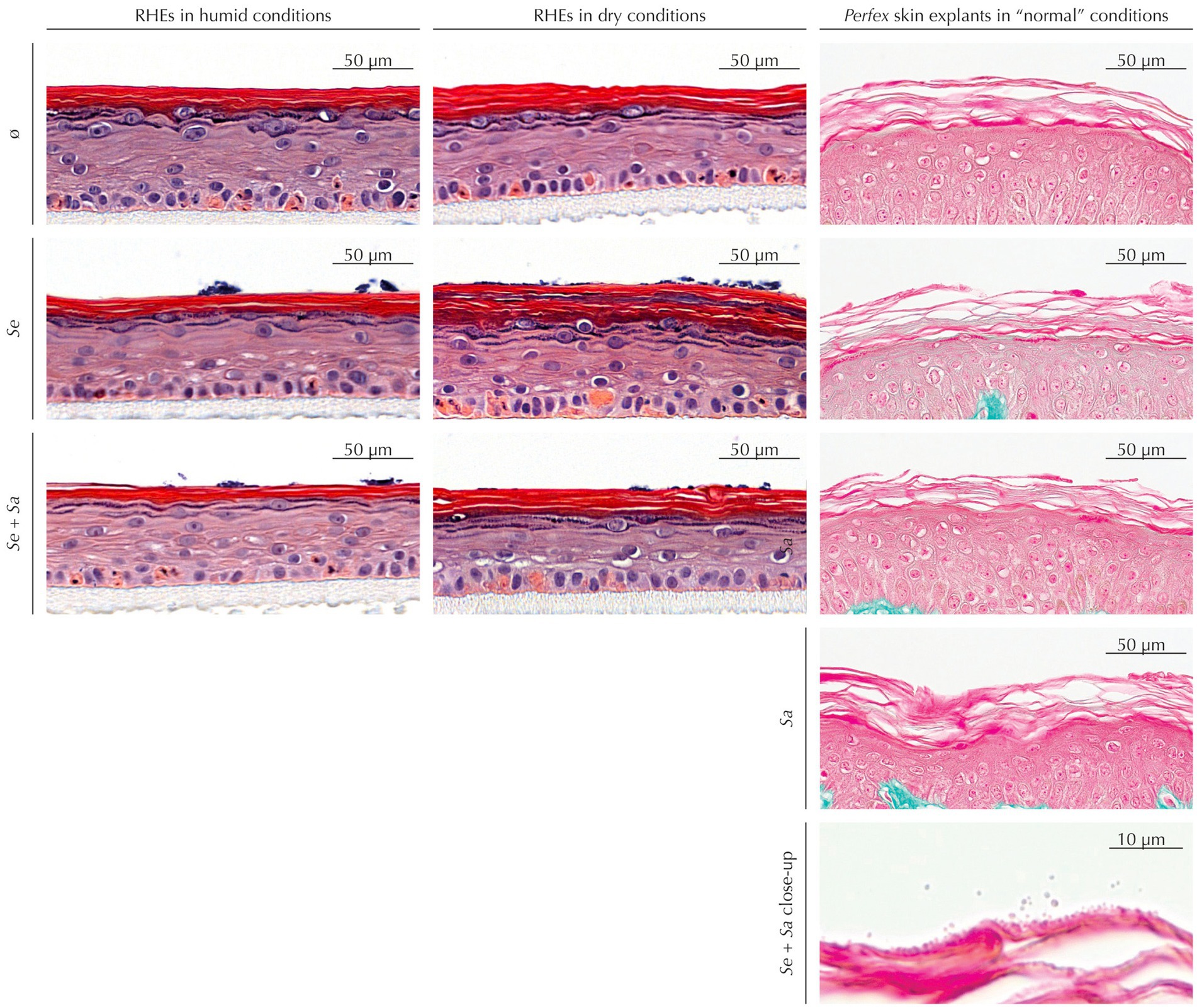
Figure 1. Histological staining of RHEs (hematoxylin and eosin staining) and Perfex skin explants (Masson’s trichrome staining) uninoculated or inoculated by S. epidermidis and/or S. aureus.
Uninoculated RHEs maintained in dry conditions exhibited the expected SC increased thickness compared to those subjected to humid conditions. This thickening was also observed in inoculated RHEs – either by S. epidermidis alone or by the S. epidermidis and S. aureus mixture. Regardless of humidity, colonization did not induce any histological alterations, although bacterial colonies were visible on the SC surface.
Similarly, skin explants showed no histological differences between inoculated and uninoculated samples, whatever the inoculum. While Masson’s trichrome staining provided limited visualization of bacteria at low magnification, higher magnifications revealed bacterial presence in the outermost SC layers.
3.2 Adhesion of bacteria
Bacterial adhesion was assessed by quantifying attached bacteria 4 h post-inoculation.
On RHEs (Figure 2A), humid or dry conditions had no effect on the adhesion efficiency of either S. epidermidis or S. aureus when the two strains were inoculated individually (p = 0.567 and p = 0.634, respectively). However, S. aureus consistently adhered more effectively than S. epidermidis (9.3% of the S. aureus from the inoculum adhered to the RHE versus 2.1% of S. epidermidis, p = 0.037 in humid conditions; 11.13% versus 2.7%, respectively, p = 0.044 in dry conditions). Under humid conditions, co-inoculation modestly increased adhesion for both species compared to single-species inoculation (S. epidermidis: +38%; S. aureus: +39%) and the increase is at the significance limit (S. epidermidis: p = 0.088; S. aureus: p = 0.066). while a similar trend seems to exist in dry conditions, differences are not significant (S. epidermidis: p = 0.739; S. aureus: p = 0.901).
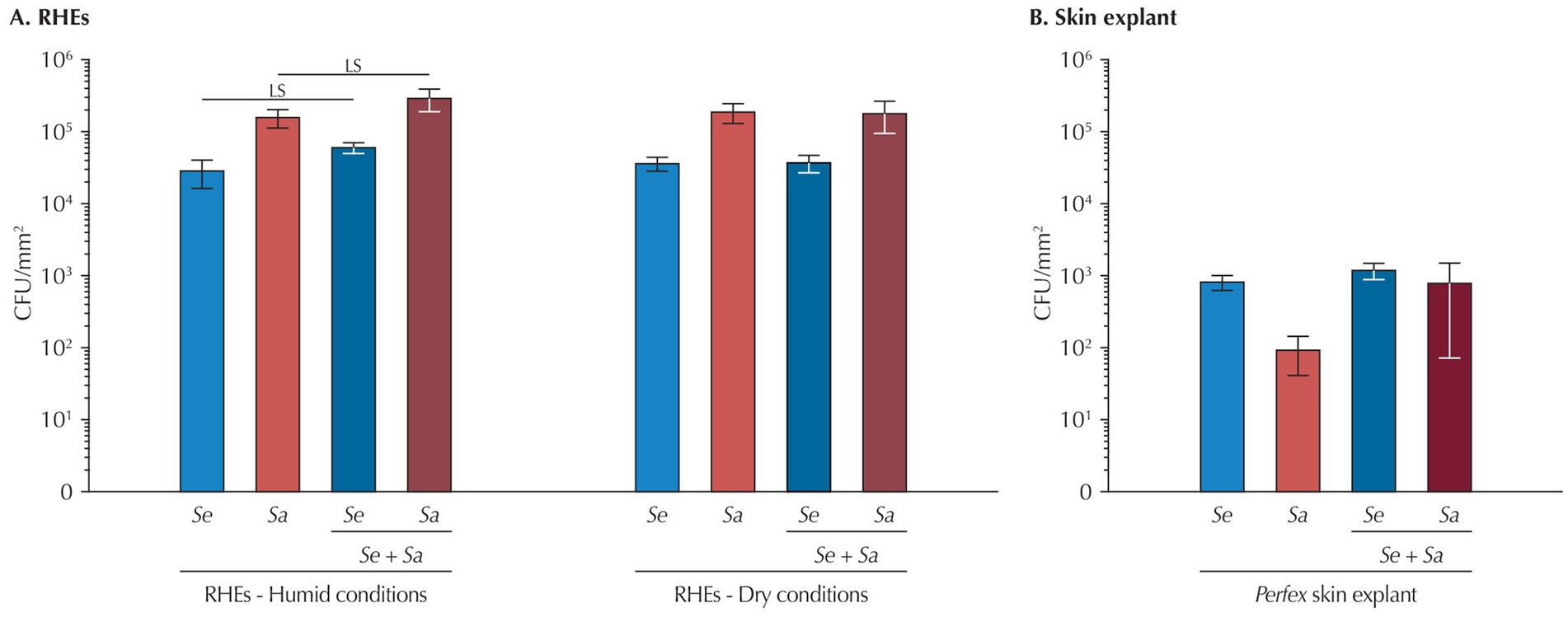
Figure 2. Bacteria attachment 4 h after inoculation (A) to RHEs and (B) to Perfex skin explants. Results are expressed as mean ± standard deviation from a single bacterial count performed on three different RHEs or on three fragments of the same skin explant. Statistical differences are reported with LS: p < 0.10 and <0.05.
In skin explants (Figure 2B), adhesion efficiency at 4 h was approximately 100-fold lower than in RHEs (0.039% for S. epidermidis and 0.003% for S. aureus), with, contrary to what was observed on RHEs, S. epidermidis exhibiting better attachment. No differences were detected between single and co-inoculations for either bacterium.
3.3 Quantification of natural moisturizing factors (NMFs)
To better understand the impact of bacterial colonization on skin hydration, we quantified three components from the NMFs – serine, pyrrolidone carboxylic acid (PCA), and urocanic acid (Figure 3).
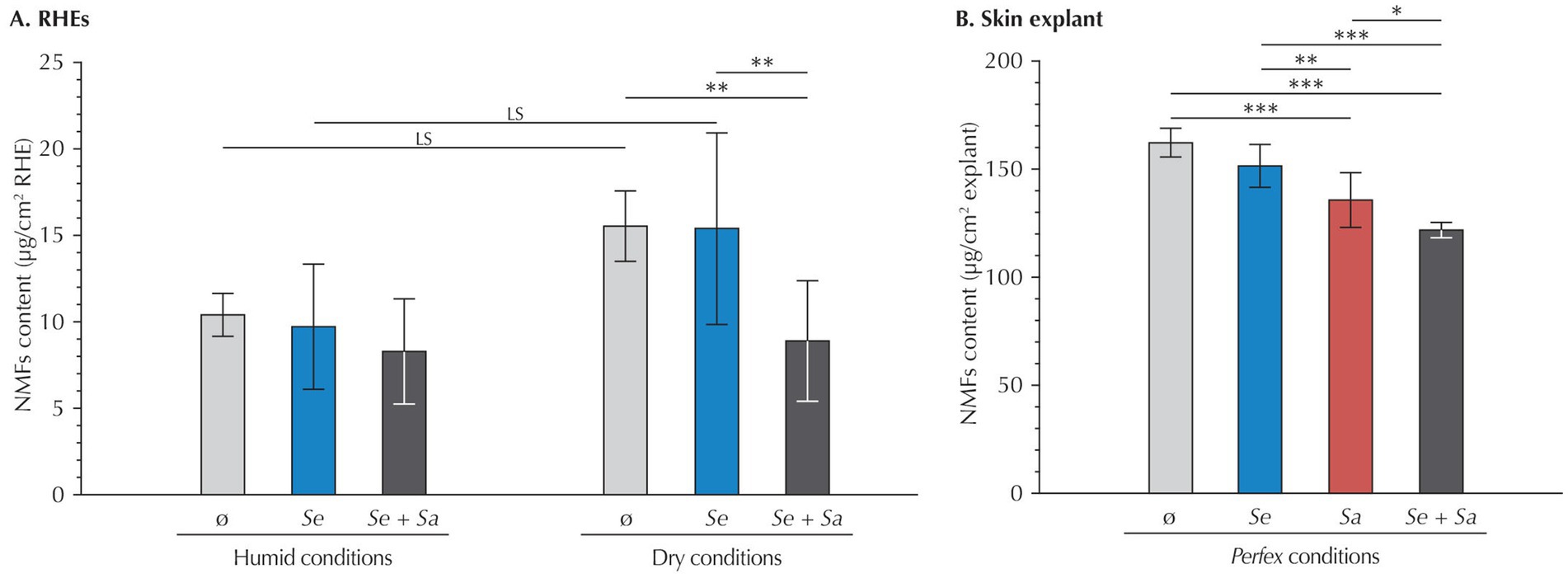
Figure 3. NMF content (serine, PCA and urocanic acid) in (A) RHEs and (B) Perfex skin explants. Results are expressed as mean ± standard deviation from measurements performed 24 h after inoculation on three different RHEs (each with duplicate technical repeats) or on three fragments of the same skin explant (each with triplicate technical repeats). Statistical differences are reported with LS: p < 0.1, *: p < 0.05, **: p < 0.01, and ***: p < 0.001.
In RHEs maintained under humid conditions, quantification of the three NMFs revealed no significant variation, regardless of whether RHEs were uninoculated or inoculated with S. epidermidis, S. aureus, or both. This absence of variation applied both to the total amount of the three NMF and to each individual compound (data not shown).
Under dry conditions, uninoculated RHEs showed a trend toward increased total NMF levels compared to those maintained under humid conditions (+54.7%, p = 0.092), although this difference was not statistically significant. At the individual compound level, increases remained non-significant or at the limit of significance (serine: p = 0.602; PCA: p = 0.100; urocanic acid: p = 0.116).
Besides, in RHEs subjected to dry conditions, inoculation with S. epidermidis alone did not alter total NMF levels compared to uninoculated controls (p = 1.000). However, co-inoculation with both bacterial strains significantly decreased the total content of the three NMFs (−42.7%, p = 0.025), restoring levels comparable to those observed under humid conditions (p = 1.000). This decrease was primarily driven by a reduction in PCA (−68.7%, p < 0.001), while changes in serine and urocanic acid levels were not statistically significant (−20.2%, p = 0.873 and −32.5%, p = 0.486, respectively).
Considering skin explants, inoculation with S. epidermidis alone had no significant impact on the total levels of the three NMFs compared to uninoculated samples (p = 0.110). In contrast, inoculation with S. aureus alone led to a 16% reduction in the total content of the three NMFs (p < 0.001), driven by significant decreases in all three components (−11% for serine, −19% for PCA, and −19% for urocanic acid; p < 0.001 for all). Co-inoculation produced an even greater reduction in the total levels of the three NMFs (−25%, p < 0.001), with consistent decreases across all components (−23% for serine, −18% for PCA, and −32% for urocanic acid; p < 0.001 for all). This reduction was also significant when compared to explants inoculated with S. epidermidis alone.
3.4 Assessment of the skin barrier function
To further investigate the effects of the microbiome model system on RHEs and skin explants, we next assessed skin barrier integrity by quantifying filaggrin (FLG) in RHEs and both FLG and ceramides in skin explants (Figure 4). As quantification methods differed between models, results were normalized to those of the respective uninoculated controls.
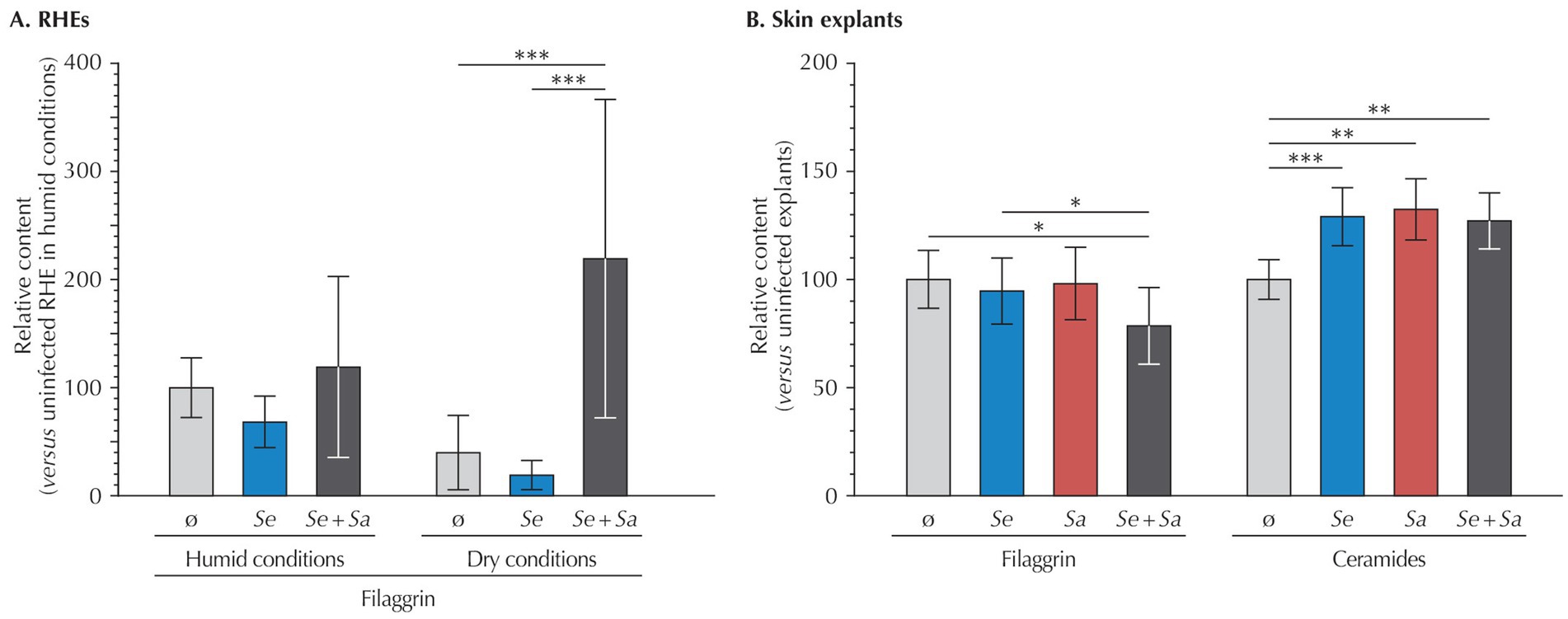
Figure 4. Relative quantification of filaggrin and ceramides in (A) RHEs (filaggrin only) and (B) Perfex skin explants, measured 24 h after bacterial inoculation. Results are expressed as mean ± standard deviation compared to the uninoculated control. For RHEs, quantifications were performed on three different RHEs (each with duplicate technical repeats). For skin explants, immunohistochemical labeling was quantified by analyzing at least three representative fields from cross-sections of three fragments of the same skin explant. Statistical differences are reported with *: p < 0.05, **: p < 0.01, and ***: p < 0.001.
In RHEs maintained under humid conditions, bacterial inoculation by S. epidermidis or S. epidermidis and S. aureus had no significant effect on FLG expression compared to the uninoculated control. A comparison between uninoculated RHEs under humid and dry conditions revealed a non-significant decrease in FLG levels in the dry condition (p = 0.559). However, under dry conditions, co-inoculation with S. epidermidis and S. aureus significantly increased FLG expression, showing a 5.5-fold rise compared to uninoculated RHEs (p = 0.007), while inoculation by S. epidermidis had no impact (p = 0.996).
In contrast, skin explants exhibited a 21% decrease in FLG upon co-inoculation by S. epidermidis and S. aureus compared to uninoculated explants (p = 0.022), whereas bacteria alone had no effect (p = 0.328 for S. epidermidis and p = 0.783 for S. aureus).
Further investigating the barrier function of skin explants, we quantified ceramides, the most abundant lipid in corneocytes, the terminally differentiated cells forming the SC. Compared to uninoculated explants, inoculation by either bacterial strain alone or their co-inoculation resulted in higher ceramide levels (S. epidermidis: +29%, p < 0.001; S. aureus: +32%, p = 0.001, and +27%, p = 0.001 for the co-inoculation).
4 Discussion
Kohda et al. (2021) used RHEs to analyze the interaction of a skin commensal (S. epidermidis) and a pathogen (S. aureus). As generally accepted, they showed that 48 h after co-inoculation, S. epidermidis reduces S. aureus colonization compared to inoculation by S. aureus alone, without promoting its own growth. Performing a similar experiment, but focusing on an earlier time point, 4 h post-inoculation, we observed different results. Under the “classical” humid conditions, co-inoculation might enhance the adhesion of both S. epidermidis and S. aureus, an effect that is not significant when RHEs are maintained under dry conditions or in a more physiological model – basal-side-perfused skin explants maintained under standard environmental conditions. Furthermore, bacterial adhesion was approximately two orders of magnitude lower in skin explants than in RHEs.
Several factors could explain the discrepancies between our results and those of Kohda et al. (2021). A first consideration is the bacterial strains. Although we used different S. epidermidis strains for RHE and skin explant inoculations, the strain used in the skin explant experiments was the same as that employed by Kohda et al. (2021). Despite differences in origin and biofilm-related traits, the two strains are similar in the sense that they represent classical reference commensals and are generally regarded as presenting “balanced,” physiologically relevant models of beneficial and pathogenic skin colonization. A substantial difference lies in the S. aureus strains. The S. aureus subsp. aureus ATCC 6538™ strain we used is a slightly virulent but sensitive reference strain, representing a model of skin colonization and biofilm formation. By contrast, Kohda et al. (2021) used the ATCC BAA-1717™ strain, which is epidemic, highly virulent, and methicillin-resistant. It is generally considered as a robust pathological and infectious model. In addition to strain choice, the S. epidermidis-S. aureus inoculation ratio also differs between studies. This ratio is important, as the ability of S. epidermidis to limit S. aureus colonization is more effective when present in 100-fold excess compared to a 10-fold excess (Kohda et al., 2021). However, the outcome of a 1:1 co-inoculation ratio, as tested in our study, was not evaluated by Kohda et al. (2021).
Another important difference concerns the skin models. Although both studies used RHEs, they likely differed substantially. One key factor is the origin of the keratinocytes RHEs are derived from, neonatal foreskin versus adult skin in our case. This difference impacts RHEs’ architecture and the expression of various markers (Nedachi et al., 2023; Bajsert et al., 2024). Moreover, RHEs from both studies can only be from different donors which also affects their characteristics (Lange et al., 2016). Therefore, they could be significant variations in colonization dynamic as this dynamic vary between individuals (Kowalski et al., 2025). The lack of biological replicates in both studies – RHEs from multiple batches/donors and skin explants from a single donor in our case – further limits the generalizability and comparability of results. Finaly, important differences exist between RHEs and skin explants, particularly considering their barrier function properties (Schäfer-Korting et al., 2008; Nakamura et al., 2018), which further complicates direct comparisons across model systems.
While these limitations are important to consider, the difference observed between both studies could also arise from the different time points analyzed. Knowledges about the precise kinetic and the early events of skin colonization by both bacterial species remains limited. Skin colonization is preceded by an initial attachment phase, mediated by various bacterial components such as adhesins – among which the Accumulation-Associated Protein (Aap) in S. epidermidis and its ortholog, the S. aureus Surface protein G (SasG) in S. aureus – as well as various cell wall-anchored proteins binding the skin matrix – the largest family consisting of the Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSCRAMMs), which are shared across both species (Paharik and Horswill, 2016; Josse et al., 2017; Severn and Horswill, 2023). The high degree of conservation of these adhesion molecules might explain the little to no differences observed, particularly given that many regulatory mechanisms active at later stages of colonization (24–48 h) are not yet functional at 4 hours. For instance, suppression of S. aureus by S. epidermidis through the quorum-sensing Accessory Gene Regulator (Agr) system depends on the accumulation of Auto-Inducing Peptideq (AIPs), which requires rather high bacterial densities (Le and Otto, 2015; Glatthardt et al., 2024), densities probably not reached at early time points. Similarly, the synthesis of antimicrobial peptides or the induction of host immune responses also requires more than 4 h to become effective (Pastar et al., 2020; Bier and Schittek, 2021). Biofilm formation and disruption, another key factor in bacterial persistence, is unlikely to be involved at this stage, as mature biofilms generally form after 24–48 h (Oliveira et al., 2007; Severn and Horswill, 2023).
Interestingly, under humid conditions (>90% relative humidity), S. epidermidis - S. aureus co-inoculation of RHEs led to enhanced bacterial attachment compared to single-species inoculation. This effect was not observed upon co-inoculation of RHEs maintained under dry conditions (<25% relative humidity) or in skin explants (40–60% relative humidity). The possible increased attachment observed upon co-inoculation suggests a potential synergistic interaction between the two bacterial species, specific to humid conditions. A direct effect of high humidity on bacteria could be possible as it positively influences bacterial metabolism and growth (Blanco et al., 2017; Qiu et al., 2022). It could also modulate host skin properties known to affect skin colonization (Bojar and Holland, 2002). However, these effects would be expected to apply equally to single and co-inoculated conditions, which is not the case. Still, S. epidermidis isolates from healthy skin modulate skin barrier function and immune responses via the Aryl hydrocarbon Receptor (AhR) pathway, but only under hydrated, conditions (Landemaine et al., 2023). In contrast, isolates from atopic dermatitis or dry skin lack these capacities, suggesting that a humid environment may be required for S. epidermidis to exert protective functions, support interspecies communication, and contribute to stable biofilm formation that could also include S. aureus.
Another notable observation is the 100-fold difference in bacterial attachment between RHE and skin explants. Although different methods were used to recover attached bacteria, quantification of unattached bacteria in explant cultures indicated that approximately half of the initial inoculum could be accounted for (data not shown), underscoring the lower attachment efficiency in this model. RHEs maintained in humid or dry conditions, are cultured at the air-liquid interface in microwell plates, allowing partial submersion and potential dynamic exposure to bacteria. In contrast, skin explants are positioned over a reservoir, exposing only their basal side to culture medium, with the stratum corneum exposed to the external environment. Inoculation in this context occurs under static conditions, resulting in less efficient bacterial deposition (Weidenmaier et al., 2018). Besides, as previously mentioned, the differences between the two models, particularly in their outermost layers, are substantial (Thakoersing et al., 2012). RHEs possess a non-desquamating stratum corneum with altered lipid content, reduced barrier function, and increased permeability. Skin explants, by contrast, maintain a fully differentiated stratum corneum, with native lipid organization and robust barrier properties. These features likely contribute to the lower bacterial adhesion observed in explants, with the reduced barrier function of RHEs possibly facilitating bacterial access to adhesion targets.
In contrast to the differential adhesion observed under humid conditions, changes in skin barrier components such as FLG and NMFs were primarily detected under dry conditions. Consistent with previous studies, dry conditions in uninoculated RHEs led to thickening of the stratum corneum and increased NMF levels (Cau et al., 2017), although the increase we observed was only at the limit of statistical significance. Besides, we also observed a non-significant decrease in FLG, potentially reflecting its degradation into NMFs.
Interestingly, only co-inoculation of RHEs maintained under dry conditions resulted in an increase in FLG levels and a concurrent reduction in NMFs, restoring values similar to those observed for RHEs maintained under humid conditions. Although S. aureus-only inoculations were not included in this series, skin explant results showed that S. aureus alone decreased NMF levels, with further reductions upon co-inoculation. Moreover, co-inoculated explants exhibited reduced FLG levels, unlike RHEs, suggesting opposite responses in the two models. Notably, both single and co-inoculation increased ceramide levels in explants.
All these results are difficult to fully explain. S. epidermidis has been reported to reinforce the skin barrier function by stimulating ceramide synthesis (Zheng et al., 2022), while S. aureus is known to impair epidermal barrier integrity and disrupt lipid metabolism (Nakatsuji et al., 2016; Kim et al., 2023). This makes the observed ceramide increase upon S. aureus inoculation somewhat unexpected. One possibility is that S. aureus-induced activation of keratinocyte stress pathways triggers compensatory upregulation of lipid synthesis, as part of a homeostatic response to barrier disruption (Kim et al., 2014). Alternatively, inflammatory cytokines induced by S. aureus (e.g., IL-1 family members, TNF-α) may indirectly stimulate ceramide production, as has been reported in epithelial stress models (Hänel et al., 2013; Feingold and Elias, 2024). Furthermore, S. epidermidis activation of AhR signaling may promote FLG synthesis (Rademacher et al., 2019; Furue, 2020), but studies on its impact on FLG expression remain inconsistent, with reports of no effect or decreased expression (Ochlich et al., 2023; Landemaine et al., 2023). In our experiments, single-species inoculation did not significantly affect FLG in either model. While our conditions were similar to those used by Ochlich et al. (2023), who observed decreased FLG in RHEs inoculated with S. epidermidis, our results showed only a non-significant reduction. This divergence might reflect context-dependent differences in AhR signaling outcomes, which can vary with ligand repertoire and local cytokine milieu (Denison and Faber, 2017). The situation is similarly complex for S. aureus. While it thrives on dry, barrier-compromised skin, its effect on FLG expression is poorly established (van Drongelen et al., 2014). Infection of isolated keratinocytes leads to increased protease activity and FLG degradation (Williams et al., 2017), suggesting that protease-mediated processing could underlie reductions in FLG. However, expression changes might be minimal, explaining our results in single-inoculation conditions. The only variation in FLG levels we observed were upon co-inoculation, with opposite results in RHEs and skin explants. While this opposite reaction may be due to the different bacterial strain used, it could also relate to the already discussed differences existing between the stratum corneum of RHEs and skin explants and the specificity of the effect of co-inoculation suggests a synergistic interaction between the two bacterial species. Potential mechanisms include competition for AhR ligands, convergence on cytokine signaling pathways, or altered protease activity resulting from interspecies crosstalk.
Notwithstanding the inconsistency in FLG expression, co-inoculation consistently led to decreased NMF levels in both RHEs and skin explants. Results from single-strain inoculations indicate that S. aureus contributes to this effect, and that co-inoculations exert a synergistic impact. Given that FLG is degraded into NMFs upon terminal differentiation of keratinocytes, the decrease in NMFs should have be accompanied by increased FLG, as was only evidenced in RHEs subjected to humid conditions. One hypothesis is that co-inoculation would trigger an immune reaction, with cytokine release reducing FLG expression, which might result in impaired barrier function and NMF loss. Alternatively, ceramide results could suggest that co-inoculation might improve the barrier function due to the stimulation of the S. epidermidis protective role by S. aureus, reducing the need for FLG synthesis and its degradation into NMFs.
Taken together, our study reveals unexpected interactions between S. epidermidis, S. aureus, and the skin. While co-inoculation may enhance bacterial attachment only under humid conditions, changes in skin barrier markers (FLG and NMFs) were observed only under dry conditions. Further studies are needed to confirm these findings and determine whether they are strain-dependent or not. Testing RHEs from different batches and various donors, as well as evaluating skin samples from various subjects will also be essential to assess whether results are generalizable. Investigating the early stages of bacterial colonization and their effects on skin biology will be essential, particularly in light of growing interest in the use of commensal bacteria as cosmetic agents or alternative therapies for skin diseases. These findings should also be considered in the broader context of skin diseases such as atopic dermatitis, in which S. aureus colonization is strongly shaped by the quorum-sensing mechanisms, antimicrobial peptide responses, and biofilm formation (Di Domenico et al., 2019; Okamoto et al., 2025). Assessing how the dysregulation of these pathways in AD not only facilitates S. aureus persistence but also amplifies inflammation, further highlights the clinical relevance of assessing the early events of host–microbe interactions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the studies involving humans because Part of the experiments were conducted on reconstructed human epidermis. The rest of the experiments were conducted on surgical discards which procurement strictly complied Articles L.1245-2 and L.1211-1 of the French Public Health Code. Written informed consent was obtained from the donor. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from A skin explant was obtained from an elective abdominoplasty. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements.
Author contributions
CN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Writing – original draft, Writing – review & editing. VP: Conceptualization, Investigation, Supervision, Validation, Writing – review & editing. GP: Data curation, Formal analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing. LP-M: Supervision, Validation, Writing – review & editing. DS: Data curation, Formal analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The present study was funded by Laboratoires Clarins.
Acknowledgments
The authors wish to thank Dr. Jérôme Dellacasagrande, QIMA Life Sciences, for his technical support. They also wish to thank Caroline Durand, Delhia Romain, and Cynthia Delpy-Fays for their help with the Perfex experiments. Finally, they are thankful to Dr. Philippe Crouzet, Estium-Concept, for providing scientific writing services.
Conflict of interest
CN and VP are full-time employees of Laboratoires Clarins.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bajsert, J., De Glas, V., Faway, E., Lambert de Rouvroit, C., Pérez-Aso, M., Cook, P. W., et al. (2024). Characterization of reconstructed human epidermis in a chemically-defined, Animal Origin-Free Cell Culture. JID Innov. 4:100298. doi: 10.1016/j.xjidi.2024.100298
Baroni, A., Buommino, E., De Gregorio, V., Ruocco, E., Ruocco, V., and Wolf, R. (2012). Structure and function of the epidermis related to barrier properties. Clin. Dermatol. 30, 257–262. doi: 10.1016/j.clindermatol.2011.08.007
Bier, K., and Schittek, B. (2021). Beneficial effects of coagulase-negative staphylococci on Staphylococcus aureus skin colonization. Exp. Dermatol. 30, 1442–1452. doi: 10.1111/exd.14381
Blanco, N., Perencevich, E., Li, S. S., Morgan, D. J., Pineles, L., Johnson, J. K., et al. (2017). Effect of meteorological factors and geographic location on methicillin-resistant Staphylococcus aureus and vancomycin-resistant enterococci colonization in the US. PLoS One 12:e0178254. doi: 10.1371/journal.pone.0178254
Bojar, R., and Holland, K. (2002). Review: the human cutaneous microflora and factors controlling colonisation. World J. Microbiol. Biotechnol. 18, 889–903. doi: 10.1023/A:1021271028979
Borowiec, A. S., Delcourt, P., Dewailly, E., and Bidaux, G. (2013). Optimal differentiation of in vitro keratinocytes requires multifactorial external control. PLoS One 8:e77507. doi: 10.1371/journal.pone.0077507
Briot, J., Simon, M., and Méchin, M. C. (2020). Deimination, intermediate filaments and associated proteins. Int. J. Mol. Sci. 21:8746. doi: 10.3390/ijms21228746
Cau, L., Pendaries, V., Lhuillier, E., Thompson, P. R., Serre, G., Takahara, H., et al. (2017). Lowering relative humidity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 86, 106–113. doi: 10.1016/j.jdermsci.2017.02.280
Cogen, A. L., Yamasaki, K., Muto, J., Sanchez, K. M., Crotty Alexander, L., Tanios, J., et al. (2010a). Staphylococcus epidermidis antimicrobial delta-toxin (phenol-soluble modulin-gamma) cooperates with host antimicrobial peptides to kill group a Streptococcus. PLoS One 5:e8557. doi: 10.1371/journal.pone.0008557
Cogen, A. L., Yamasaki, K., Sanchez, K. M., Dorschner, R. A., Lai, Y., MacLeod, D. T., et al. (2010b). Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J. Invest. Dermatol. 130, 192–200. doi: 10.1038/jid.2009.243
de Szalay, S., and Wertz, P. W. (2023). Protective barriers provided by the epidermis. Int. J. Mol. Sci. 24:3145. doi: 10.3390/ijms24043145
Denison, M. S., and Faber, S. C. (2017). And now for something completely different: diversity in ligand-dependent activation of ah receptor responses. Curr. Opin. Toxicol. 2, 124–131. doi: 10.1016/j.cotox.2017.01.006.28845473
Di Domenico, E. G., Cavallo, I., Capitanio, B., Ascenzioni, F., Pimpinelli, F., Morrone, A., et al. (2019). Staphylococcus aureus and the cutaneous microbiota biofilms in the pathogenesis of atopic dermatitis. Microorganisms 7:301. doi: 10.3390/microorganisms7090301
Drislane, C., and Irvine, A. D. (2020). The role of filaggrin in atopic dermatitis and allergic disease. Ann. Allergy Asthma Immunol. 124, 36–43. doi: 10.1016/j.anai.2019.10.008
Elias, P. M. (1983). Epidermal lipids, barrier function, and desquamation. J. Invest. Dermatol. 80, 44s–49s. doi: 10.1038/jid.1983.12
Feingold, K. R., and Elias, P. M. (2024). The role of ceramides in the disruption of the cutaneous permeability barrier, a common manifestation of skin disorders. J. Lipid Res. 65:100593. doi: 10.1016/j.jlr.2024.100593
Flowers, L., and Grice, E. A. (2020). The skin microbiota: balancing risk and reward. Cell Host Microbe 28, 190–200. doi: 10.1016/j.chom.2020.06.017
Furue, M. (2020). Regulation of Filaggrin, Loricrin, and Involucrin by IL-4, IL-13, IL-17A, IL-22, AHR, and NRF2: pathogenic implications in atopic dermatitis. Int. J. Mol. Sci. 21:5382. doi: 10.3390/ijms21155382
Glatthardt, T., Lima, R. D., de Mattos, R. M., and Ferreira, R. B. R. (2024). Microbe interactions within the skin microbiome. Antibiotics 13:49. doi: 10.3390/antibiotics13010049
Hänel, K. H., Cornelissen, C., Lüscher, B., and Baron, J. M. (2013). Cytokines and the skin barrier. Int. J. Mol. Sci. 14, 6720–6745. doi: 10.3390/ijms14046720
Jordana-Lluch, E., Garcia, V., Kingdon, A. D. H., Singh, N., Alexander, C., Williams, P., et al. (2020). Simple polymicrobial biofilm keratinocyte colonization model for exploring interactions between commensals, pathogens and antimicrobials. Front. Microbiol. 11:291. doi: 10.3389/fmicb.2020.00291
Josse, J., Laurent, F., and Diot, A. (2017). Staphylococcal adhesion and host cell invasion: fibronectin-binding and other mechanisms. Front. Microbiol. 8:2433. doi: 10.3389/fmicb.2017.02433
Kim, J., Kim, B. E., Berdyshev, E., Bronova, I., Bin, L., Bae, J., et al. (2023). Staphylococcus aureus causes aberrant epidermal lipid composition and skin barrier dysfunction. Allergy 78, 1292–1306. doi: 10.1111/all.15640
Kim, Y., and Lim, K. M. (2021). Skin barrier dysfunction and filaggrin. Arch. Pharm. Res. 44, 36–48. doi: 10.1007/s12272-021-01305-x
Kim, Y. I., Park, K., Kim, J. Y., Seo, H. S., Shin, K. O., Lee, Y. M., et al. (2014). An endoplasmic reticulum stress-initiated sphingolipid metabolite, ceramide-1-phosphate, regulates epithelial innate immunity by stimulating β-defensin production. Mol. Cell. Biol. 34, 4368–4378. doi: 10.1128/MCB.00599-14
Kohda, K., Li, X., Soga, N., Nagura, R., Duerna, T., Nakajima, S., et al. (2021). An in vitro mixed infection model with commensal and pathogenic staphylococci for the exploration of interspecific interactions and their impacts on skin physiology. Front. Cell. Infect. Microbiol. 11:712360. doi: 10.3389/fcimb.2021.712360
Kowalski, C. H., Nguyen, U. T., Lawhorn, S., Smith, T. J., Corrigan, R. M., Suh, W. S., et al. (2025). Skin mycobiota-mediated antagonism against Staphylococcus aureus through a modified fatty acid. Curr. Biol. 35, 2266–2281.e8. doi: 10.1016/j.cub.2025.03.055
Landemaine, L., Da Costa, G., Fissier, E., Francis, C., Morand, S., Verbeke, J., et al. (2023). Staphylococcus epidermidis isolates from atopic or healthy skin have opposite effect on skin cells: potential implication of the AHR pathway modulation. Front. Immunol. 14:1098160. doi: 10.3389/fimmu.2023.1098160
Lange, J., Weil, F., Riegler, C., Groeber, F., Rebhan, S., Kurdyn, S., et al. (2016). Interactions of donor sources and media influence the histo-morphological quality of full-thickness skin models. Biotechnol. J. 11, 1352–1361. doi: 10.1002/biot.201600360
Le, K. Y., and Otto, M. (2015). Quorum-sensing regulation in staphylococci-an overview. Front. Microbiol. 6:1174. doi: 10.3389/fmicb.2015.01174
Lefèvre-Utile, A., Braun, C., Haftek, M., and Aubin, F. (2021). Five functional aspects of the epidermal barrier. Int. J. Mol. Sci. 22:11676. doi: 10.3390/ijms222111676
Meisel, J. S., Sfyroera, G., Bartow-McKenney, C., Gimblet, C., Bugayev, J., Horwinski, J., et al. (2018). Commensal microbiota modulate gene expression in the skin. Microbiome 6:20. doi: 10.1186/s40168-018-0404-9
Nakamura, M., Haarmann-Stemmann, T., Krutmann, J., and Morita, A. (2018). Alternative test models for skin ageing research. Exp. Dermatol. 27, 495–500. doi: 10.1111/exd.13519
Nakatsuji, T., Chen, T. H., Two, A. M., Chun, K. A., Narala, S., Geha, R. S., et al. (2016). Staphylococcus aureus exploits epidermal barrier defects in atopic dermatitis to trigger cytokine expression. J. Invest. Dermatol. 136, 2192–2200. doi: 10.1016/j.jid.2016.05.127
Nedachi, T., Bonod, C., Rorteau, J., Chinoune, W., Ishiuchi, Y., Hughes, S., et al. (2023). Chronological aging impacts abundance, function and microRNA content of extracellular vesicles produced by human epidermal keratinocytes. Aging 15, 12702–12722. doi: 10.18632/aging.205245
Nodake, Y., Matsumoto, S., Miura, R., Honda, H., Ishibashi, G., Matsumoto, S., et al. (2015). Pilot study on novel skin care method by augmentation with Staphylococcus epidermidis, an autologous skin microbe - a blinded randomized clinical trial. J. Dermatol. Sci. 79, 119–126. doi: 10.1016/j.jdermsci.2015.05.001
Ochlich, D., Rademacher, F., Drerup, K. A., Gläser, R., and Harder, J. (2023). The influence of the commensal skin bacterium Staphylococcus epidermidis on the epidermal barrier and inflammation: implications for atopic dermatitis. Exp. Dermatol. 32, 555–561. doi: 10.1111/exd.14727
Okamoto, H., Li, S., and Nakamura, Y. (2025). The role of skin dysbiosis and quorum sensing in atopic dermatitis. JID Innov. 5:100377. doi: 10.1016/j.xjidi.2025
Oliveira, M., Nunes, S. F., Carneiro, C., Bexiga, R., Bernardo, F., and Vilela, C. L. (2007). Time course of biofilm formation by Staphylococcus aureus and Staphylococcus epidermidis mastitis isolates. Vet. Microbiol. 124, 187–191. doi: 10.1016/j.vetmic.2007.04.016
Paharik, A. E., and Horswill, A. R. (2016). The staphylococcal biofilm: Adhesins, regulation, and host response. Microbiol. Spectr. 4:15. doi: 10.1128/microbiolspec.VMBF-0022-2015
Pastar, I., O'Neill, K., Padula, L., Head, C. R., Burgess, J. L., Chen, V., et al. (2020). Staphylococcus epidermidis boosts innate immune response by activation of gamma delta T cells and induction of perforin-2 in human skin. Front. Immunol. 11:550946. doi: 10.3389/fimmu.2020.550946
Peno-Mazzarino, L., Jeanneton, O., Scalvino, S. A., Percoco, G., Beauchef, G., Nizard, C., et al. (2024). A new ex vivo human skin model for the topographic and biological analysis of cosmetic formulas. Int. J. Cosmet. Sci. 47, 305–320. doi: 10.1111/ics.13027
Qiu, Y., Zhou, Y., Chang, Y., Liang, X., Zhang, H., Lin, X., et al. (2022). The effects of ventilation, humidity, and temperature on bacterial growth and bacterial genera distribution. Int. J. Environ. Res. Public Health 19:15345. doi: 10.3390/ijerph192215345
Quiroz, F. G., Fiore, V. F., Levorse, J., Polak, L., Wong, E., Pasolli, H. A., et al. (2020). Liquid-liquid phase separation drives skin barrier formation. Science 367:eaax9554. doi: 10.1126/science.aax9554
Rademacher, F., Simanski, M., Gläser, R., and Harder, J. (2018). Skin microbiota and human 3D skin models. Exp. Dermatol. 27, 489–494. doi: 10.1111/exd.13517
Rademacher, F., Simanski, M., Hesse, B., Dombrowsky, G., Vent, N., Gläser, R., et al. (2019). Staphylococcus epidermidis activates aryl hydrocarbon receptor signaling in human keratinocytes: implications for cutaneous defense. J. Innate Immun. 11, 125–135. doi: 10.1159/000492162
Rawlings, A. V., and Harding, C. R. (2004). Moisturization and skin barrier function. Dermatol. Ther. 17, 43–48. doi: 10.1111/j.1396-0296.2004.04s1005.x
Schäfer-Korting, M., Bock, U., Diembeck, W., Düsing, H. J., Gamer, A., Haltner-Ukomadu, E., et al. (2008). The use of reconstituted human epidermis for skin absorption testing: results of the validation study. Altern. Lab. Anim 36, 161–187. doi: 10.1177/026119290803600207
Severn, M. M., and Horswill, A. R. (2023). Staphylococcus epidermidis and its dual lifestyle in skin health and infection. Nat. Rev. Microbiol. 21, 97–111. doi: 10.1038/s41579-022-00780-3
Spencer, T. S., Linamen, C. E., Akers, W. A., and Jones, H. E. (1975). Temperature dependence of water content of stratum corneum. Br. J. Dermatol. 93, 159–164. doi: 10.1111/j.1365-2133.1975.tb06735.x
Thakoersing, V. S., Danso, M. O., Mulder, A., Gooris, G., El Ghalbzouri, A., and Bouwstra, J. A. (2012). Nature versus nurture: does human skin maintain its stratum corneum lipid properties in vitro? Exp. Dermatol. 21, 865–870. doi: 10.1111/exd.12031
van Drongelen, V., Haisma, E. M., Out-Luiting, J. J., Nibbering, P. H., and El Ghalbzouri, A. (2014). Reduced filaggrin expression is accompanied by increased Staphylococcus aureus colonization of epidermal skin models. Clin. Exp. Allergy 44, 1515–1524. doi: 10.1111/cea.12443
Weidenmaier, C., Kokai-Kun, J. F., Kulauzovic, E., Kohler, T., Thumm, G., Stoll, H., et al. (2018). Differential roles of sortase-anchored surface proteins and wall teichoic acid in Staphylococcus aureus nasal colonization. Int. J. Med. Microbiol. 298, 505–513. doi: 10.1016/j.ijmm.2007.11.006
Williams, M. R., Nakatsuji, T., Sanford, J. A., Vrbanac, A. F., and Gallo, R. L. (2017). Staphylococcus aureus induces increased serine protease activity in keratinocytes. J. Invest. Dermatol. 137, 377–384. doi: 10.1016/j.jid.2016.10.008
Keywords: skin microbiome, reconstructed human epidermis, skin explant, Staphylococcus epidermidis, Staphylococcus aureus, barrier function
Citation: Ngari C, Poulet V, Percoco G, Peno-Mazzarino L and Seyer D (2025) Early skin colonization by Staphylococcus epidermidis and Staphylococcus aureus reveals environment-dependent synergistic effects. Front. Microbiol. 16:1677214. doi: 10.3389/fmicb.2025.1677214
Edited by:
Enea Gino Di Domenico, San Gallicano Dermatological Institute IRCCS, ItalyReviewed by:
Flavia Costa, University of Colorado Anschutz Medical Campus, United StatesTej Pratap Singh, University of Pennsylvania, United States
Copyright © 2025 Ngari, Poulet, Percoco, Peno-Mazzarino and Seyer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chrisse Ngari, Y2hyaXNzZS5uZ2FyaUBjbGFyaW5zLmNvbQ==