 Hanna Mazur-Marzec1*
Hanna Mazur-Marzec1* Łukasz Grabowski2
Łukasz Grabowski2 Alicja Węgrzyn3
Alicja Węgrzyn3 Agata Błaszczyk1
Agata Błaszczyk1 Marta Cegłowska4*Przemysław Dąbek5Momina Farooq1
Marta Cegłowska4*Przemysław Dąbek5Momina Farooq1 Ewa Górecka5
Ewa Górecka5 Agata Jurczak-Kurek6Anna-Karina Kaczorowska7
Agata Jurczak-Kurek6Anna-Karina Kaczorowska7 Tadeusz Kaczorowski8
Tadeusz Kaczorowski8 Marija Kataržytė9
Marija Kataržytė9 Robert Konkel1
Robert Konkel1 Ewa Kotlarska10
Ewa Kotlarska10 Donata Overlingė9Waldemar Surosz11
Donata Overlingė9Waldemar Surosz11 Anna Toruńska-Sitarz1Semko Walat1Monika Zielenkiewicz2
Anna Toruńska-Sitarz1Semko Walat1Monika Zielenkiewicz2 Grzegorz Węgrzyn2
Grzegorz Węgrzyn2- 1Department of Marine Biology and Biotechnology, University of Gdańsk, Gdynia, Poland
- 2Department of Molecular Biology, University of Gdańsk, Gdańsk, Poland
- 3University Center for Applied and Interdisciplinary Research, University of Gdańsk, Gdańsk, Poland
- 4Department of Marine Chemistry and Biochemistry, Institute of Oceanology, Polish Academy of Sciences, Sopot, Poland
- 5Institute of Marine and Environmental Sciences, University of Szczecin, Szczecin, Poland
- 6Department of Evolutionary Genetics and Biosystematics, University of Gdańsk, Gdańsk, Poland
- 7Collection of Plasmids and Microorganisms, University of Gdańsk, Gdansk, Poland
- 8Laboratory of Extremophiles Biology, Department of Microbiology, University of Gdańsk, Gdańsk, Poland
- 9Marine Research Institute, Klaipeda University, Klaipeda, Lithuania
- 10Department of Genetics and Marine Biotechnology, Institute of Oceanology, Polish Academy of Sciences, Sopot, Poland
- 11Department of Phycology, University of Gdańsk, Gdynia, Poland
Marine microorganisms are increasingly recognized as valuable sources of bioactive compounds and enzymes with diverse applications in biotechnology. Despite its relatively low overall biodiversity, the Baltic Sea harbours a variety of diatoms, dinoflagellates, bacteria (including cyanobacteria), fungi, and bacteriophages with notable biotechnological potential. These organisms produce metabolites with promising application in environmental remediation or as components of pharmaceuticals, nutraceuticals, cosmetics and biomaterials. Enzymes produced by Baltic Sea bacteria catalyse reactions of industrial relevance, while bacteriophages may provide novel tools for pathogen control in aquaculture or serve as sources of genes encoding for valuable enzymes. Although advances in high-throughput genomics and metabolomics have accelerated marine biodiscovery, Baltic Sea microorganisms remain largely understudied and underexploited by industry. This review synthesizes current knowledge on the biotechnological potential of the Baltic Sea microorganisms and highlights opportunities to bridge the gap between basic research and commercial application, particularly in the context of international frameworks such as the Nagoya Protocol.
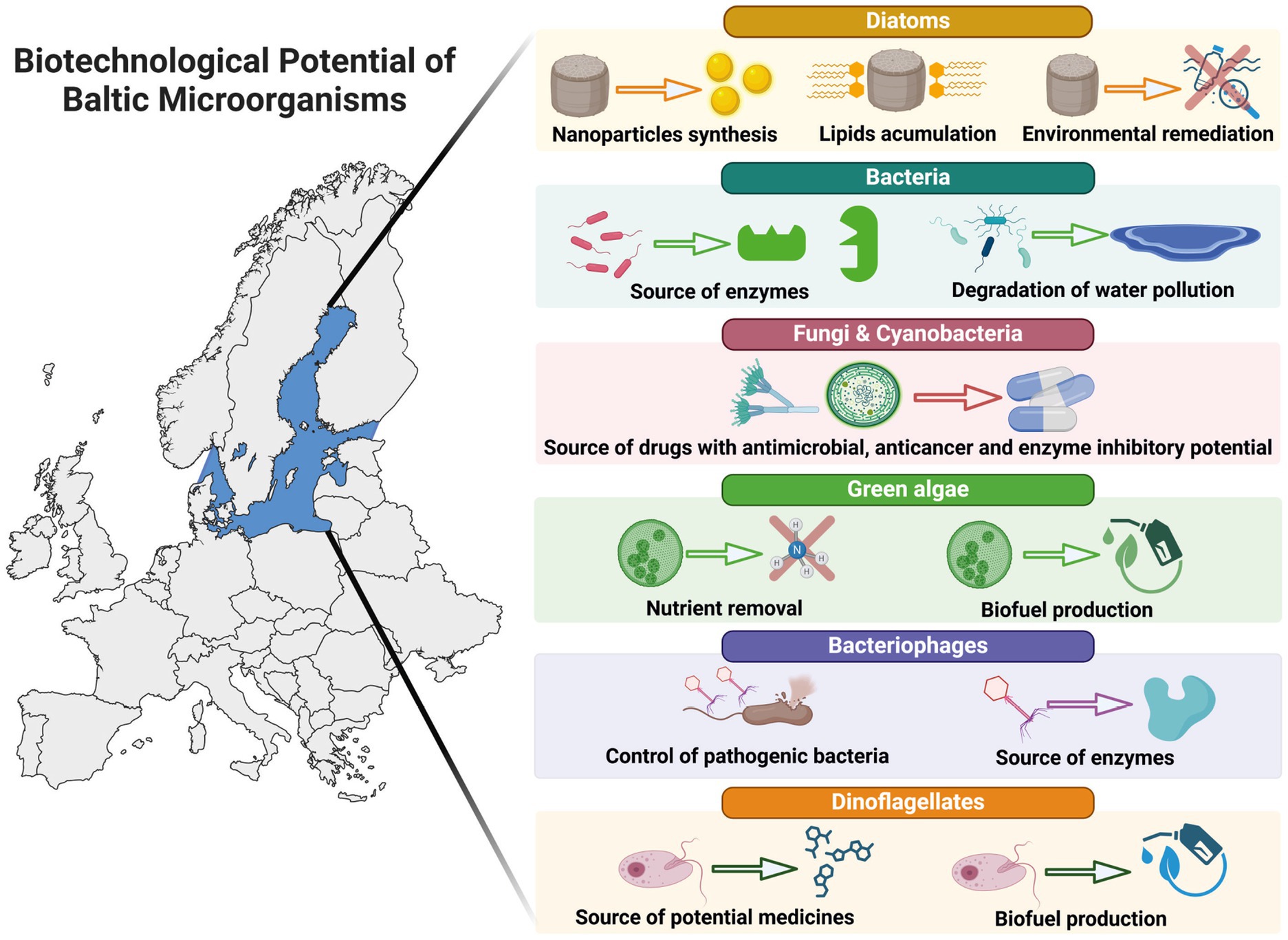
Graphical Abstract.
1 Introduction
Marine biological resources encompass living organisms, genetic resources, and their products (metabolites), all of which are key components of the marine ecosystem, influencing its diversity, structure, and functioning. Beyond their ecological roles and importance as essential elements of the human diet, people have explored the seas’ natural wealth for various purposes since ancient times. In recent decades, research into the potential of marine organisms as a basis for new, innovative products with significant economic and social implications has grown considerably. This trend is also evident in the Baltic Sea region, where numerous initiatives have been launched to promote the development of marine biotechnology (Supplementary Tables S1–S3). In this area of biotechnology, microorganisms play a particularly pivotal role. The biotechnological potential of marine ecosystems is primarily assessed based on two key factors: the high biological diversity of organisms, which provides a rich and varied source of material for exploration, and the unusual environmental conditions that drive distinctive metabolic processes and increase the likelihood of discovering novel bioactive compounds with unique properties.
The Baltic Sea is commonly regarded as an ecosystem characterised by low biological diversity. However, despite these limitation, the microbial community within this brackish waterbody (excluding viruses) was estimated to encompass over one million (Ojaveer et al., 2010). Microorganisms inhabiting this environment are exposed to pronounced and dynamic gradients in salinity, temperature, and nutrient availability, which impose strong selective pressure. These challenging conditions drive the evolution of specialized physiological adaptation, frequently resulting in the development of unique metabolic pathways and the biosynthesis of structurally and functionally novel biomolecules that are uncommon in microorganisms from more stable ecosystems.
This study aims underscore the underappreciated biotechnological potential of the Baltic Sea microorganisms. Their metabolites and bioactive compounds, developed into low and high added value products, may find applications across a variety of sectors, including pharmaceutical industry, industrial biotechnology or environmental remediation. Importantly, the functional diversity within different microbial taxa suggests that each group may provide different contribution to the development of marine biotechnology sector in the region.
2 Low-added value products from the Baltic microorganisms
Value-added products are products that, through processing, acquire additional qualities that contribute to their higher market value compared to the raw materials used to make them. In marine biotechnology, biofuel, food and feed products are defined as low-value, high-volume, and low-risk products (Rotter et al., 2020).
During the last few decades, Baltic Sea microalgae, including diatoms, green algae, cyanobacteria, and dinoflagellates, have gained interest as a potential source of lipids for biofuel production. Most of the research conducted in this area has focused on optimizing microalgae cultivation conditions in order to increase lipid content in cells (Schwenk et al., 2013; Steinhoff et al., 2014; Klin et al., 2020; Klin et al., 2023; Spilling et al., 2021). Nutrient limitation, especially nitrogen, was found to have a beneficial effect on lipid content, but resulted in a decrease in microalgal biomass production (Schwenk et al., 2013; Ratomski and Hawrot-Paw, 2021). Promising results were observed in co-adapted polycultures (microalgae dominated by green algae, protozoans, and bacteria) cultivated under nitrogen limitation and diurnal temperature fluctuations (Mattsson et al., 2019). Under these conditions, the lipid content increased while the biomass remained unaffected. Additionally, the incorporation of elicitors, such as chitosan, into the culture of Chlorella pyrenoidosa from the Baltic Sea led to a significant increase in lipid content within a short period (Spilling, 2011). An alternative approach was proposed by Pechsiri and Gröndahl (2022), who suggested that microalgae harvested during extensive and massive blooms of Nodularia spumigena in the Baltic Sea could be utilized as a source of biomass for biogas and biofertilizer production.
Due to their ability to uptake nutrients (nitrogen, phosphorus, organic, and inorganic carbon), microalgae have gained significant attention as a component of wastewater treatment processes (Plöhn et al., 2021). In this context, the use of Baltic Sea microalgae (green algae and cyanobacteria) for nutrient removal and biofuel production has been investigated in a couple of studies (Lynch et al., 2015; Jämsä et al., 2017). Wastewater treatment by the green alga Scenedsmus sp. UHCC0027 resulted in high nutrient removal rates (100% of ammonium and 90% of phosphate). The high efficiency of this process was achieved when an increased CO2 level was applied. However, the regulatory levels for nitrogen and phosphorus removal in treated water were unmet (Lynch et al., 2015). The same strain was used in a subsequent study, where at temperatures between 7 and 13 °C, an effective nutrient removal was recorded with a simultaneous increase in the total lipid content. However, due to the high content of polyunsaturated fatty acids, which are undesirable components of biodiesel, the algal biomass did not meet current standards set by the European Union and the United States (Jämsä et al., 2017).
Furthermore, the Baltic Sea microalge demonstrated effective CO2 capture (20–60%) from flue gas (Olofsson et al., 2015; Lindehoff et al., 2024). The results showed both high productivity and the excellent quality of microalgal biomass harvested from these cultures. Additionally, selected strains of Baltic cyanobacteria were found to produce significant amounts of hydrogen gas (H2), which is considered a clean energy carrier and a promising alternative to fossil fuels (Khetkorn et al., 2017; Allahverdiyeva et al., 2010).
Few studies have addressed the economic evaluation of the commercial application of Baltic microalgae as a source of low-value products. Pechsiri and Gröndahl (2022) determined the energy return on investment (EROI = total energy output/total primary energy input), as an indicator of environmental and economic performance, for the biomass harvesting process of surface accumulated cyanobacteria from the Gotland basin and its conversion to biogas and biofertilizers. They found that depending on biomass harvesting and transport methods, as well as the type of the obtained products, harvesting toxic Nodularia spumigena during blooms from the Baltic Sea could be both environmentally and economically beneficial (Pechsiri and Gröndahl, 2022).
Overall, Baltic Sea microalgae gained academic interest as potential sources of low-value products; however, none have been introduced to the market. The most common challenge lies in scaling up the systems and developing cost-effective outdoor bioreactors (Allahverdiyeva et al., 2010; Steinhoff et al., 2014; Olofsson et al., 2015; Hawrot-Paw and Ratomski, 2024). In particular, the biomass harvesting and dewatering processes remain challenging. Potential solutions for simple and low-cost flocculation and dewatering techniques in closed bioreactor systems using Baltic Sea diatoms and green algae as surrogate species were proposed by Spilling (2011). Future progress will depend on the integration of technological innovations with economic and environmental assessments, paving the way for sustainable large-scale applications within the bioeconomy.
3 Baltic Sea diatoms as a source of high-value products
Diatoms are an incredibly diverse group of microorganisms which possess significant biotechnological potential (Figure 1). Their exoskeleton, known as frustule, primarily composed of silica (SiO2) exhibits intricate ornamentation patterns with diverse shapes and sizes. The silica frustule is durable, transparent, and resistant to degradation (Mishra et al., 2017; Roychoudhury et al., 2022a; Saoud et al., 2022; Zobi, 2022). The frustules, with a large surface area, and high adsorption capacity, represent promising and eco-friendly ‘silica resource’ offering unique properties and significant potential for application in material engineering, nanotechnology, catalysis, and environmental remediation (Golubeva et al., 2023). Moreover, the combination of various metabolic capabilities allows diatoms to uptake, transport, and metabolize metal ions, facilitating the formation of metal nanoparticles both inside and outside their cells. The exact mechanisms of metal nanoparticle synthesis in diatoms are still an area of active research, and the specific enzymes, biomolecules, and cellular processes involved may differ between diatom species and metal ions (Roychoudhury et al., 2022a).
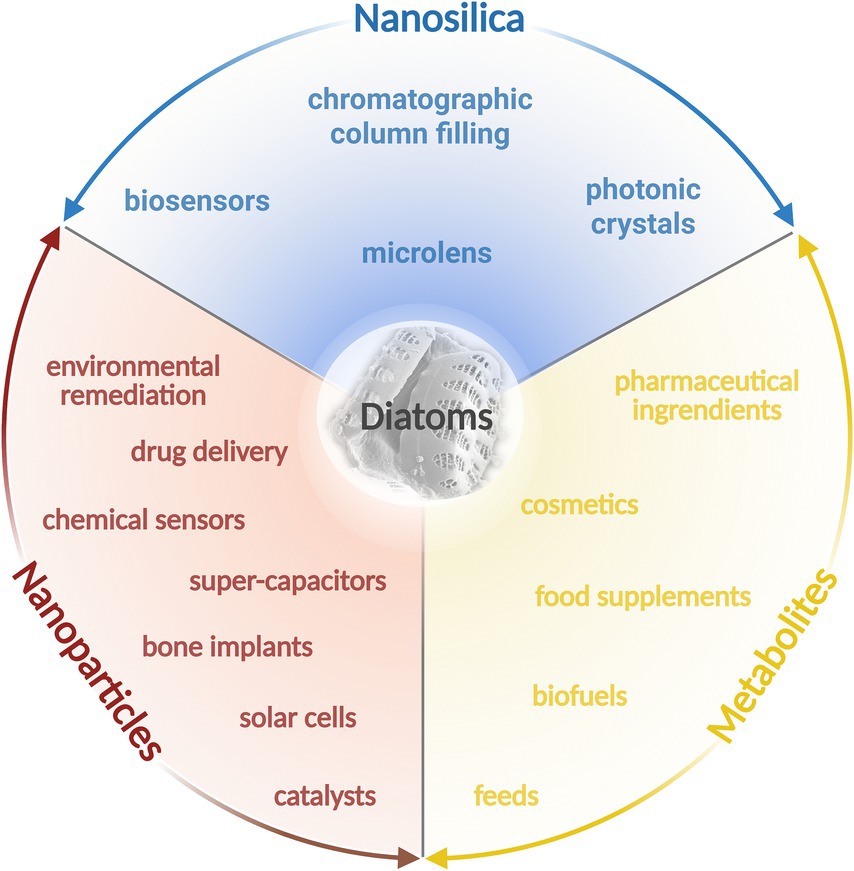
Figure 1. Baltic diatoms and their biotechnological potential. The figure was created using BioRender.com.
Recently, diatoms from the Baltic Sea have been used in several studies as a source of high-value compounds, including nanosilica, in the fabrication of metal-biosilica complexes or fatty acids. Two species Gedaniella mutabilis and G. flavovirens, isolated from the Gulf of Gdańsk and Resko Przymorskie Lake, respectively, when treated with a 9 mM silver nitrate solution, were effective in synthesizing Ag-SiO2 flower-shaped nanoparticles, with a diameter of 100–500 nm (Roychoudhury et al., 2021). These nanohybrids exhibited autofluorescence properties in the blue light region (excitation ~450–490 nm), and their catalytic activity in the degradation of methylene blue, a dye commonly used in industry, was also demonstrated.
Pseudostaurosira trainorii has recently become the Baltic Sea model species in applied research. It has been utilized in the biosynthesis of spindle-shaped iron-oxide nanoparticles (IONPs) (Roychoudhury et al., 2022b). In this work, IONPs diatom bioreagent exhibited catalytic properties in the degradation of nitrophenol after 3 h of reaction. In another study, P. trainorii was used to synthesize diatom-based metal nanoparticles (Sprynskyy et al., 2021). The authors conducted a photocatalysis using biosilica doped with palladium(II) chloride nanoparticles to degrade methyl orange in aqueous solution under UV radiation. The degradation efficiency rate reached more than 98% after 75 min. Furthermore, Brzozowska et al. (2022) assessed the optical and physicochemical properties of the latter species by metabolic insertion of titanium ions into the culture medium, achieving a titanium content of more than 8.5% wt. in the diatom frustules. As a result, a reinforced, three-dimensional biosilica enriched with titanium ions was obtained, exhibiting promising semiconductor and optoelectronic properties. The material was characterised by four types of photoluminescence activity in ultraviolet, narrow blue, green, and red-light regions (Brzozowska et al., 2022). The same species was also used in a study by Nowak et al. (2020), where P. trainorii biomass served as a source of biosilica and carbon with electrochemical properties. After pyrolyzing the biomass at the temperature of 600 °C, it was found that the carbonized material had a specific discharge capacity of 460 mAh/g−1, demonstrating that diatom biosilica can become an electroactive material, potentially valuable for applications such as lithium-ion batteries (Nowak et al., 2020).
Diatoms are also well known for their ability to accumulate lipids within their cells, particularly triacylglycerols (TAGs), as storage compounds (Leyland et al., 2020; Yi et al., 2017). These lipids make diatoms, especially those from epiphytic and benthic communities, a primary food source for micrograzers and higher trophic-level organisms in the world’s most productive coastal areas (Cox et al., 2020). Diatoms can adjust their lipid biosynthesis in response to various environmental stressors, such as nutrient and light limitation (d'Ippolito et al., 2015; Hildebrand et al., 2012; Sayanova et al., 2017; Singha et al., 2022). They are also a storehouse for a wide range of bioactive compounds, including pigments, and polyunsaturated fatty acids (PUFAs), such as omega-3 (Bayu et al., 2020; Nieri et al., 2023). Due to antioxidant, antibacterial, anticancer and other activities, these compounds have potential applications in the pharmaceutical and nutraceutical biotechnological sectors as a starting material for the development of biomaterials, drugs, dietary supplements, and functional foods (Mal et al., 2021; Marzec et al., 2022; Sharma et al., 2021).
With a growing demand for eco-friendly alternative energy sources and nutraceutical components, more research is being conducted on the topic of cold-water microalgae with high lipid yield potential. A detailed study of two diatom species commonly found in the Baltic Sea waters, Chaetoceros wighamii Brightwell and Thalassiosira baltica (Grunow) Ostenfeld, revealed a significant variation in their fatty acids (FA) profiles depending on temperature, light, and nutrient limitation (Spilling et al., 2021). Both diatoms were confirmed to contain eicosapentaenoic acis (EPA), but the concentration of this PUFA in T. baltica was generally high (with a maximum of approximately 30 mg EPA/g of ash-free dry weight under Si-limitation) when compared to other diatom species C. wighamii.
The effect of light intensity on EPA production was studied by Spilling et al. (2021). The authors found that the fraction of EPA was lower under high light conditions, but no correlation was observed between overall FA content and light intensity. The most significant factors affecting the FA profile in these diatoms were the growth phase and nutrient limitation pattern. A similar conclusion regarding the relationship between FA profile and growth phase was reached in another study that explored factors influencing lipid content in Baltic diatoms and other brackish and marine microalgae (Schwenk et al., 2013). It was found that FA content increased by an average of 2.6 times as cultures transitioned from the exponential to the stationary phase. The collected data strongly support the hypothesis that the growth phase, regardless of nutrient limitation, is a significant factor affecting the total FA content in diatom cells.
The challenges that the cultivation of diatom for biomass in the Baltic Sea region may face include the variation in seasonal environmental conditions. While outdoor culturing systems are most efficient in areas with a warm, mild climate, temperature and light intensity fluctuations could affect cultivated diatoms’ growth and fatty acid content. This disadvantage is worth considering, as the amount and quality of the yield from diatom cultivation systems are essential for the competitiveness of the final product (Gilmour, 2019). The simulation of seasonal variation in culture conditions by Cheregi et al. (2021) shows that the same species cannot be grown throughout the year; instead, a seasonal species rotation should be applied, using local species that reach maximum growth at temperatures matching the seasonal conditions. An example of the advantages of large-scale diatom cultivation is the ability to combine biomass growth with sustainable methods for removing waste gases. A screening study of the most promising temperate climate diatom species, T. baltica and C. wighamii (Spilling, 2011), demonstrated the potential for diatom cultivation combined with CO2 fixation from industrial exhaust gases, such as those from geothermal power plant fumes. The source of CO2 did not affect growth efficiency when compared to pure, commercial CO2. However, the potential impact of pollutants, such as SO2 and other toxic gases, present in CO2 sources on diatom growth remains to be investigated in further research. Such studies may provide further insights into the sustainability and viability of this cultivation method.
In conclusion, Baltic diatoms can represent a significant and sustainable source of high-value products due to their diverse metabolic capabilities, efficient lipid accumulation, and ability to synthesize metal nanoparticles. With ongoing research into optimizing cultivation conditions and extraction methods, diatoms could play a pivotal role in advancing biotechnological applications in materials science, environmental remediation, and the pharmaceutical sector.
4 Dinoflagellates phycotoxins as potential pharmaceuticals
Phycotoxins produced by dinoflagellates cause significant damage to the global fishing industry and aquaculture. These toxins are also responsible for various human diseases (Karlson et al., 2021). Despite their harmful effects, some of these metabolites have been investigated as potential candidates for developing high-value products (Cho et al., 2020). The Baltic population of Alexandrium ostenfeldii produces saxitoxins (STXs), the alkaloids with guanidine group (Figure 2), responsible for paralytic shellfish poisoning (PSP) (Salgado et al., 2015).
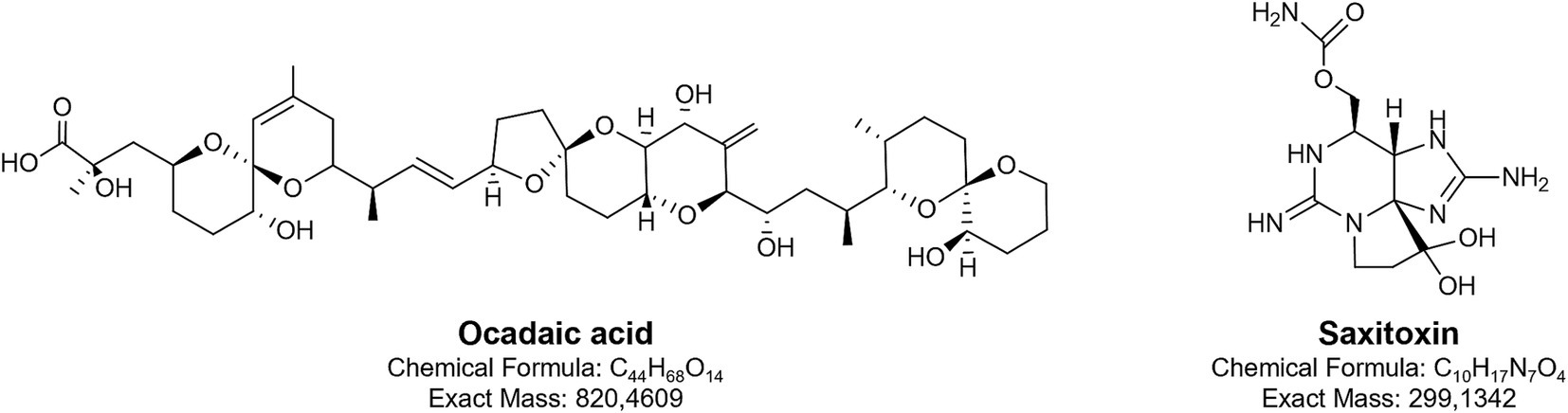
Figure 2. Chemical structures of two toxins produced by Baltic dinoflagellates: okadaic acid and saxitoxin.
Over 60 structural variants of saxitoxins have been identified, though in Alexandrium ostenfeldii from the Baltic region, only saxitoxin (STX) with a carbamoyl group and two decarbamoyl gonyautoxins, GTX-3 and GTX-2, have been detected. Among these, the neurotoxic activity of the carbamoyl analogues is the most potent. STXs selectively bind to voltage-gated sodium channels (Navs), particularly at site 1 on the α-subunit, and physically block the pore of the channels. This prevents sodium ion influx during membrane depolarization, thereby inhibiting the initiation and propagation of action potentials in excitable cells, such as neurons and muscle fibres (Wiese et al., 2010; Stevens et al., 2011; Shen et al., 2018). The blockade of Nav-mediated sodium currents leads to the interruption of neuronal signalling and neuromuscular transmission, which ultimately results in paralysis. Owing to this specific interaction with Nav channels, STXs have been explored for potential applications as local anaesthetics or anti-wrinkle agents, where controlled inhibition of nerve conduction is desirable (Kohane et al., 2000; Epstein-Barash et al., 2009). The drug-like properties of saxitoxins, particularly the decarbamoyl analogues, have been confirmed by Flores-Holguín et al. (2024) using the SwissADME web tool. In the 1950s and 1960s, due to its acute toxicity, saxitoxin was also investigated as a potential chemical weapon (Sierra and Martínez-Álvarez, 2020).
Okadaic acid (OA) and dinophysiotoxin DTX-1 (methylated form of OA) produced in the Baltic by Dynophysis species are polyether fatty acids (Figure 2) and represent the most widely occurring toxins associated with diarrhetic shellfish poisoning (DSP). OA and DTX-1 exert their biological activity primarily through potent inhibition of serine/threonine-specific protein phosphatases PP1 and PP2A by binding to their catalytic subunits. This inhibition disrupts the balance between phosphorylation and dephosphorylation in cells, leading to abnormal hyperphosphorylation of numerous intracellular proteins. As a result, the function of many signalling and regulatory pathways becomes impaired (Valdiglesias et al., 2013; Lee et al., 2016). The downstream effects of this molecular disruption include gastrointestinal symptoms such as diarrhoea, vomiting, and abdominal pain. Furthermore, OA has been shown to induce both cytotoxic (Valdiglesias et al., 2011) and neurotoxic effects (Kamat et al., 2013). Because of its precise and potent inhibition of PP1 and PP2A, OA is widely used as a research tool to study intracellular signalling pathways, particularly those involving reversible phosphorylation. It has been instrumental in elucidating the role of phosphatases in processes such as cell cycle regulation, apoptosis, and neurodegeneration, including mechanisms implicated in Alzheimer’s disease (Cruz et al., 2013; Kamat et al., 2013). Furthermore, the immunoregulatory potential of OA has been demonstrated (López et al., 2011). In experiments involving the mouse T lymphocyte cell line EL-4, exposure to OA led to down regulation of the T cell receptor complex. A similar effect was observed with yessotoxin, another polyether toxin produced by dinoflagellates, although its mechanism of action differed (López et al., 2011).
Biotechnological applications of dinoflagellate metabolites, including toxins associated with PSP and DSP, have been extensively reviewed in several studies (Camacho et al., 2007; Assunção et al., 2017; Cho et al., 2020). A compilation of patents highlights the diverse therapeutic potential of the toxins (Camacho et al., 2007; Assunção et al., 2017). However, it has been noted that the limited quantities of available metabolites, in relation to the amounts required for preclinical studies, pose a significant challenge to advancing research on their therapeutic potential. The production of these metabolites in bioreactors, is insufficient for commercialization, primarily due to the slow growth rate of dinoflagellates. Furthermore, genetic modifications to enhance the efficiency of production processes are more complex than in microorganisms with smaller genomes. The number of sequenced dinoflagellate genomes is also notably limited in comparison to other microbial species (Lin, 2024). Moreover, chemical synthesis of these compounds is challenging, involving numerous steps, which significantly impact both the cost and availability of the product. Addressing these challenges will likely pave the way for developing at least some of dinoflagellate metabolites into viable therapeutic agents.
5 Baltic cyanobacteria a treasure trove of bioactive compounds
Summer cyanobacteria blooms are among the most problematic issues of the Baltic Sea. They generate high biomass and disrupt the functioning of the ecosystem (Karjalainen et al., 2007). Regardless of their negative environmental impact, cyanobacteria are a valuable source of compounds that can serve as a basis for developing high-value products (Luesch et al., 2025; Baunach et al., 2024; Demay et al., 2019). A particularly prominent example in this context is dolastatin, which has served as a lead compound for the development of several anticancer agents (Luesch et al., 2025). However, as its 40-year development pathway illustrates, the process of translating a biologically active compound into a marketable pharmaceutical remains both time-consuming and resource-intensive. Thus far, the biotechnological and pharmaceutical potential of Baltic cyanobacteria has been explored using filamentous forms from the order Nostocales, such as Nodularia, Anabaena, and Nostoc (Fewer et al., 2009; Jokela et al., 2012; Mazur-Marzec et al., 2018; Fewer et al., 2021; Konkel et al., 2023a,b). These microorganisms produce secondary metabolites with antifungal, antiviral, anticancer, and enzyme-inhibiting activities.
Antifungal cyanometabolites such as hassallidins, hassallidin-like balticidins, puwainaphycins, and anabaenolysins are cyclic lipopeptides (Fewer et al., 2021). They contain a peptide ring and a fatty acid chain, and each of these structural elements has specific characteristics for a given class of lipopeptides (Figure 3). Hassallidins have been identified in various filamentous cyanobacteria, including the Baltic strains of Anabaena (Bui et al., 2014; Vestola et al., 2014). These compounds contain a peptide ring with eight amino acids linked to a saturated fatty acid chain (C14-18) through the exocyclic residue (Bui et al., 2014; Vestola et al., 2014). In addition to chlorine (in balticidins), different types and numbers of monosaccharides have been identified in the structure of hassallidins. Puwainaphycins, another class of cyclic lipopeptides produced by cyanobacteria (Vašíček et al., 2020), were detected in the benthic Nodularia harveyana strain UHCC-0300 (Saurav et al., 2022). Puwainaphycins (and structurally homologous minutissamides) are composed of nine amino acids and a β-amino-α-hydroxy-γ-methyl fatty acid incorporated into the ring (Gregson et al., 1992; Vašíček et al., 2020; Saurav et al., 2022). The unbranched fatty acid part (C10-18) of the molecule can be substituted by chlorine, hydroxyl, or oxo functional groups. Anabaenolysins were discovered in Anabaena strains from the Gulf of Finland (Jokela et al., 2012; Oftedal et al., 2012). In the structure of these lipopeptides, four amino acids, including the unusual 2-(3-amino-5-oxytetrahydrofuran-2-yl)-2-hydroxyacetic acid, and fatty acid chain (C18) with different degrees of unsaturation are present (Jokela et al., 2012; Oftedal et al., 2012).
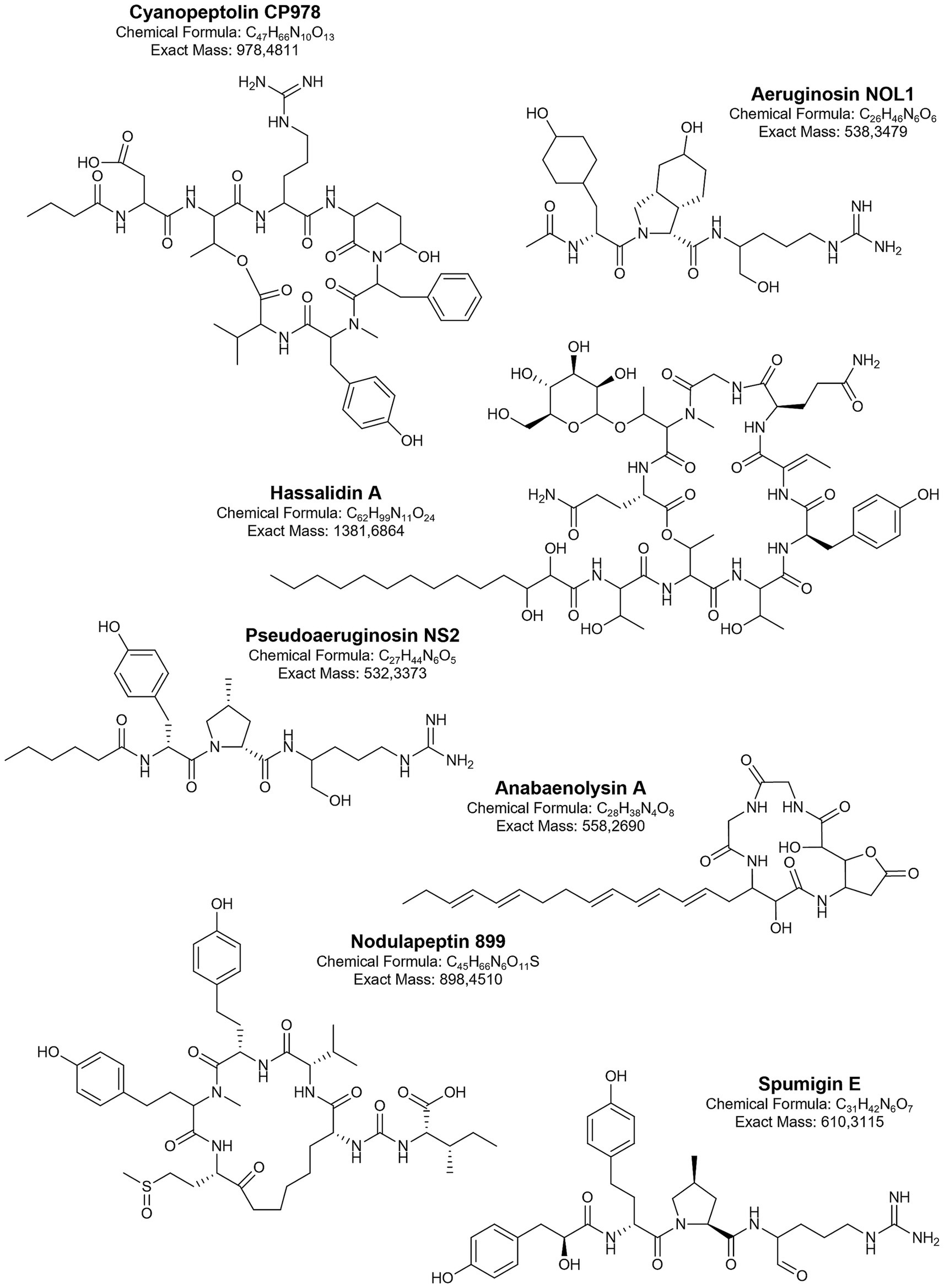
Figure 3. Biologically active metabolites produced by Baltic cyanobacteria.
All these lipopeptides exhibit activity against human pathogenic fungi (Neuhof et al., 2005; Bui et al., 2014; Vestola et al., 2014; Shishido et al., 2015; Humisto et al., 2019; Hájek et al., 2021). They act by disintegrating sterol-containing cell surface membranes (Jokela et al., 2012; Oftedal et al., 2012; Hrouzek et al., 2012; Humisto et al., 2019). The effects clearly depend on the structure of the lipophilic part of the molecules, particularly the length of the fatty acid chain and the type and position of the substituted functional group (Hájek et al., 2021). For example, puwainaphycins with longer fatty acid chains exhibit more potent antifungal properties compared to compounds with shorter chains, while hydroxylation of part of the lipopeptides significantly reduces their activity (Hrouzek et al., 2012; Hájek et al., 2021; Saurav et al., 2022). In addition to antifungal activity, cyanobacterial lipopeptides also cause lysis, necrotic changes, and death of various eukaryotic cells through interaction with sterol-containing membranes (Hrouzek et al., 2012; Jokela et al., 2012; Oftedal et al., 2012; Humisto et al., 2019). Since the cytotoxicity of cyanobacterial lipopeptides has also been observed in healthy cells, their biotechnological application would require structural modifications to eliminate the undesirable effects while preserving antifungal activity.
Serine protease inhibitors are another important class of bioactive metabolites detected in Baltic cyanobacteria. Serine proteases are involved in many essential metabolic pathways and are found in both prokaryotic and eukaryotic cells. Disruption of their balanced activity can lead to various pathologies, including cancer, inflammation, cardiovascular disease, and neurodegenerative disorders (Patel, 2017; Harish and Uppuluri, 2018; Singh et al., 2023). All of this makes serine proteases important targets in the drug discovery process.
The activity against serine proteases is demonstrated by several classes of linear and cyclic peptides produced by Baltic cyanobacteria. These include non-ribosomally synthesised spumigins, aeruginosins, pseudoaeruginosins, anabaenopeptins, cyanopeptolins, and nostocyclopeptides. Different structural variants of linear spumigins, aeruginosins, and pseudoaeruginosins have been identified in many Baltic strains of N. spumigena (Fewer et al., 2009; Mazur-Marzec et al., 2012; Liu et al., 2015; Mazur-Marzec et al., 2016), Nostoc spp. from the Gulf of Finland (Heinilä et al., 2022), and Aphanizomenon sp. strain KUCC C2 from the Curonian Lagoon (Overlingė et al., 2024a). All these tetrapeptides contain arginine (Arg) or its mimetics at the C-terminal position. The presence and form of Arg have a significant impact on the biological activity of these peptides (Liu et al., 2023). At the N-terminal part of aeruginosins and pseudoaeruginosins, a short fatty acid chain is present. In spumigins and varlaxins, this position is occupied by hydroxyphenyl lactic acid (Hpla) or 2-(4-hydroxyphenyl) acetic acid (Hpaa), respectively. In spumigins and pseudoaeruginosins, the position adjacent to Arg contains proline or methylproline (MePro), while in aeruginosins and varlaxins, a 2-carboxy-6-hydroxyoctahydroindole (Choi) is present (Figure 3).
Although these peptides are common components of Baltic cyanobacterial cells, only a few have been isolated and tested in enzyme inhibition assays. Spumigin E (Fewer et al., 2009), varlaxins (1046A and 1022A) (Heinilä et al., 2022), and pseudoaeruginosin NL2 (Liu et al., 2015) inhibited porcine or/and human trypsins with IC50 in the nanomolar range, while aeruginosin AER525 was most active against thrombin (IC50 = 0.59 μM), it also inhibited trypsin and carboxypeptidase A (Overlingė et al., 2024b). A comparison of the biological activity of peptides from the described classes (Liu et al., 2023) indicates that those produced by Baltic strains are among the strongest inhibitors of trypsin and thrombin. The significance of serine protease inhibitors is emphasized by the growing body of research on their potential use in cancer treatment. For example, trypsin-3 inhibitors have been shown to reduce metastasis in prostate, breast, and ovarian cancers (Sananes et al., 2023). The link between trypsin inhibition and anticancer activity has also been observed for suomilide from the Baltic N. sphaerocarpa UHCC 0038 (Ahmed et al., 2021). This compound, which shares some structural similarities with aeruginosins (Figure 3), was previously identified in N. spumigena strains AV1 and HKVV (Fujii et al., 1997). Suomilide isolated from N. sphaerocarpa UHCC 0038 inhibited the activity of recombinant human trypsins (1, 2, and 3) at nanomolar concentrations (4.7–104.0 nM) and, at 3.3 μM, also inhibited the invasion of aggressive metastatic PC-3 M prostate cancer cells (trypsin-3) (Ahmed et al., 2021).
Cyanopeptolins are another class of serine protease inhibitors commonly found in cyanobacteria from various taxonomic groups (Weckesser et al., 1996; Rounge et al., 2007; Jones et al., 2021). In the Baltic Sea, their presence has been detected in Nostoc edaphicum CCNP1411 (93 structural variants) (Mazur-Marzec et al., 2018; Konkel et al., 2023a) and in bloom samples from the Curonian Lagoon (Overlingė et al., 2021). The unique feature of cyanopeptolins is the presence of 3-amino-6-hydroxy-2-piperidone (Ahp) in a six-amino acid peptide ring cyclized by an ester bond (Figure 3). They also have a side chain composed of 1–2 amino acids and a short fatty acid (Weckesser et al., 1996; Konkel et al., 2023a). Cyanopeptolins isolated from the Baltic N. edaphicum CCNP1411 inhibited the activity of trypsin, chymotrypsin, and elastase in the low μM range. The activity against these enzymes was selective and depended on the type of amino acid in the second position, between conserved threonine and Ahp (Figure 3). Although all of these depsipeptides were active against chymotrypsin, those containing Arg in this position also inhibited trypsin, while elastase was only inhibited by cyanopeptolins with leucine (Konkel et al., 2023a). Interestingly, two of the leucine-containing cyanopeptolins and CP978 (with Arg) also showed cytotoxic effects against human cervical cancer cells (HeLa). Cyanopeptolin CP978 appears to be one of the most interesting peptides produced by Baltic cyanobacteria in terms of potential applications. In addition to inhibiting the activity of serine proteases, it also protects human cells from SARS-CoV-2 infection (Konkel et al., 2023b). CP978 interacts with the virion by binding directly to the coronaviral S protein. This action caused a significant decline in SARS-CoV-2 replication in A549ACE2/™PRSS2 cells (adenocarcinoma human alveolar basal epithelial cells overexpressing ACE2 and TMPRSS2) and in primary human airway epithelial cells (HAE) (Konkel et al., 2023b). In the viral infection assay, the determined IC50 was 82 nM, classifying CP978 as one of the most potent natural products inhibiting SARS-CoV-2 infection (Wang et al., 2022). The antiviral activities of other cyanometabolites (e.g., lectins, sulphated polysaccharides) have also been described, but their presence in Baltic cyanobacteria has never been explored.
Similarly to cyanobacteria from other aquatic environments (Monteiro et al., 2021; Zervou et al., 2021), the Baltic strains produce numerous variants of cyclic hexapeptides classified as anabaenopeptins. These peptides have been identified in N. spumigena strains (Rouhiainen et al., 2010; Mazur-Marzec et al., 2012; Mazur-Marzec et al., 2016), N. edaphicum CCNP1411 (Konkel et al., 2022), picocyanobacterium Coelosphaeriaceae 06S067 (Häggqvist et al., 2016), Aphanizomenon sp. KUCC C1 (Overlingė et al., 2024a), and in bloom samples dominated by N. spumigena, Aphanizomenon sp., and Dolichospermum sp. (Spoof et al., 2015; Overlingė et al., 2021). A characteristic feature of this class of compounds is the cyclic pentapeptide ring, with a conserved D-lysine linked to the exocyclic amino acid through the ureido bond (Figure 3). Anabaenopeptins identified in Baltic cyanobacteria inhibit protein phosphatase and carboxypeptidase A (IC50 in the nanomolar range) (Spoof et al., 2015; Häggqvist et al., 2016); their activity against elastase has also been documented (Konkel et al., 2022). The potency of the effects on tested enzymes depends mainly on the type of amino acid in the exocyclic position. The pharmaceutical potential of anabaenopeptins (from Planktothrix and Nostoc) with Arg in this position has been well documented by Schreuder et al. (2016). These peptides, as strong inhibitors of thrombin-activable fibrinolysis (TAFIa) (IC50 = 1.5–16 nM), could be important agents in the treatment and prevention of thrombosis. Although structural variants of anabaenopeptins with Arg in the exocyclic position are produced by Baltic cyanobacteria (Häggqvist et al., 2016; Overlingė et al., 2021; Overlingė et al., 2024a), their activity against TAFIa has never been tested.
In addition to the extensive body of research on isolated and identified cyanometabolites, bioassays were performed on extracts and fractions from Baltic cyanobacterial (Mazur-Marzec et al., 2015; Felczykowska et al., 2015; Szubert et al., 2018; Cegłowska et al., 2022; Overlingė et al., 2024a). The results of these studies demonstrate that many biologically active metabolites produced by Baltic cyanobacteria remain to be discovered. Notably, several already known cyanometabolites described here exhibit strong antifungal, antiviral, anticancer, and enzyme-inhibitory activities. However, none of them has progressed beyond the stage of basic research. One of the limitations appears to be the insufficient yield of these metabolites from biomass, which hinders efforts to conduct more advanced studies.
6 Biotechnological potential of other Baltic bacteria
In addition to cyanobacteria, other bacterial species possess significant biotechnological potential due to their diverse capabilities, which can be applied across various sectors of human activity. These microorganisms exhibit a range of functions, from the degradation of uncommon organic compounds and the detection of mutagenic substances to the production of potentially valuable enzymes, antimicrobial agents, and the synthesis of nutrients using H2O and CO2 as substrates (Ferrer et al., 2019; Barzkar et al., 2024). Bacteria exhibiting such characteristics are also found in the Baltic Sea (Figure 4). Among them, Actinomycetes have emerged as a predominant group of marine bacteria with significant biotechnological relevance (Jagannathan et al., 2021; Ngamcharungchit et al., 2023). The general role of marine bacteria in advancing biotechnological innovations has been reviewed recently in several papers (Ferrer et al., 2019; Jagannathan et al., 2021; Ngamcharungchit et al., 2023; Barzkar et al., 2024). Thus, in this chapter, we will specifically focus on the biotechnological potential of Baltic bacteria (excluding cyanobacteria, which were addressed in the preceding chapter).
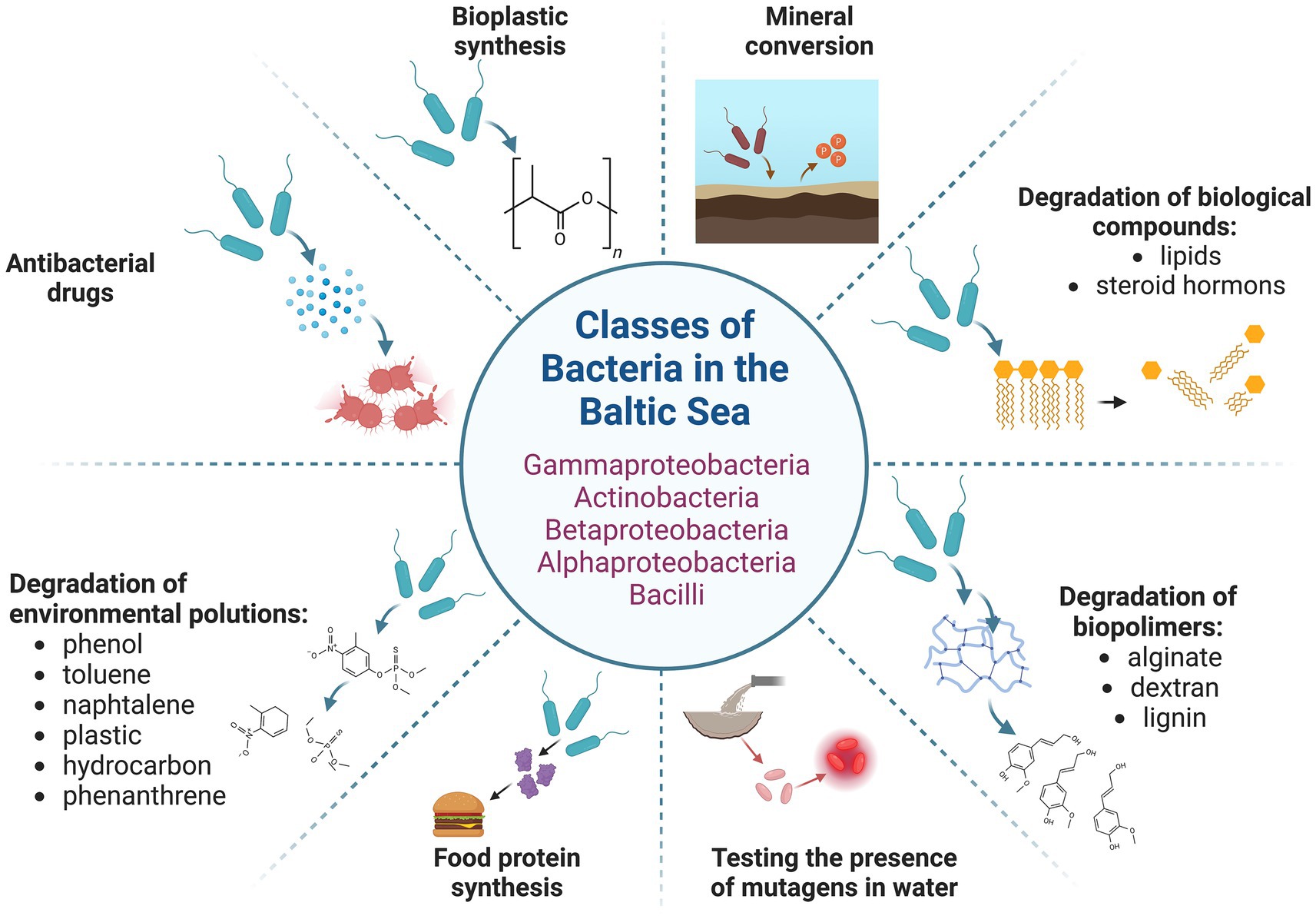
Figure 4. Baltic bacteria and their potential biotechnological application. The figure was created using BioRender.com.
6.1 Enzymes from Baltic bacteria
When evaluating the biotechnological potential of microorganisms, one of the most apparent applications is the isolation and use of various enzymes that catalyse reactions of technological significance. Indeed, marine bacteria may encode enzymes of atypical activities or characteristics other than those found in species from more extensively explored environments, such as those from the Entrobacteriaceae family. These enzymes might be of interest due to the unique environmental conditions under which they function.
Early research led to the cloning and characterisation of a gene encoding a unique hemolysin from Vibrio pommerensis from the Baltic Sea (Jores et al., 2003). Hemolysins are of pharmaceutical interest, particularly as potential anticancer enzymes, and may also be utilized in developing biosensors (Panchal et al., 2002). Another group of biotechnologically relevant enzymes are lipases. A gene encoding a lipase active at temperatures as low as 5–10 °C has been cloned with the gene identified in a metagenomic library of Baltic Sea sediment bacteria. Thus, the specific bacterium containing this gene remains unidentified (Hårdeman and Sjöling, 2007). Interestingly, microalgae residing in the Baltic Sea are known to contain relatively high levels of lipids (Dolganyuk et al., 2022), suggesting that bacteria in the same habitat may have evolved to utilize these compounds by producing effective lipases. Another metagenomic analysis revealed that bacteria living in Baltic waters harbour numerous genes coding for transposases (Vigil-Stenman et al., 2017), enzymes responsible for transposition of mobile genetic elements in cells, which hold significant biotechnological potential in genetic engineering.
The chemical degradation of complex organic compounds can be challenging, yet these molecules could serve as valuable sources of molecular monomers. Examples include alginate and dextran, which can be hydrolysed into oligo- or monosaccharides through enzymatic processes. Marine bacteria represent a promising source of alginate lyases (Barzkar et al., 2022a) and dextranases (Barzkar et al., 2022b). Bacterial species that produce these enzymes have been found in the Baltic Sea (Barzkar et al., 2022a,b). Another example are fatty acids and sialic acid degrading enzymes. In this context, a cold-active bacterium isolated from Baltic Sea ice, Shewanella glacialimarina, with an interesting fatty acid profile and the ability to metabolize sialic acid (Qasim et al., 2021), could represent a promising step toward the biotechnological application of enzymes derived from this organism.
6.2 Potential direct use of bacteria to conduct specific degradation processes
Although enzymes encoded by bacteria can be highly effective under laboratory conditions or as components of pharmaceuticals, the degradation of specific compounds in the environment is typically more efficient when living bacteria are employed rather than isolated enzymes.
Numerous studies have described examples of bacteria capable of degrading a wide range of compounds, including xenobiotics. These microorganisms could assist in the remediation of chemical pollution in the environment. For example, Pseudomonas spp. and Exiguobacterium sp., isolated from a phenanthrene-enriched, Baltic Sea sediments, revealed the ability to degrade the pollutant (Edlund and Jansson, 2008). Specifically, phenanthrene concentration was decreased from 3.7 μg/mL to 0.8 μg/mL in such sediments, during the incubation period of 60 days, due to the action of these bacteria (Edlund and Jansson, 2008). Isolations of bacteria degrading phenol and toluene from the Baltic Sea have also been reported; the degradation capabilities were assumed on the basis of the presence of genes encoding specific enzymes, phenol hydroxylases and catechol 2,3-dioxygenases (Vedler et al., 2013) and toluene monooxygenase and toluene/xylene monooxygenase (Heinaru et al., 2016). These strains belong to the Limnobacter genus and Pseudomonas stutzeri species, respectively, and were proposed as potential biocontrol agents for the remediation of phenol and toluene contamination in marine waters. In some cases, the degradation of specific compounds may require the involvement of bacterial communities rather than single species. Such a phenomenon was observed in the degradation of nonylphenol and butyltin by bacteria present in bottom sediments from the Baltic Sea, particularly those from the Gulf of Finland (Kuzikova et al., 2022). Namely, the sediments were enriched with nonylphenol and/or butyltin, up to 300 mg/kg and 95 mg/kg, respectively, and upon incubation for 240 days with bacterial communities consisting with species belonging mainly to genera Halothiobacillus, Geothrix, Methanosarcina, Dyella, Parvibaculum, Pseudomonas, and Proteiniclasticum, and order Rhizobiales, a total removal of monobutyltin was observed, with the nonylphenol residual rate estimated for 40% (Kuzikova et al., 2022). In fact, the Gulf of Finland seems to be particularly rich in xenobiotic-degrading bacteria, as demonstrated by the isolation of the Delftia tsuruhatensis strain, which is effective in degrading naphthalene, as demonstrated by the observation of the 43% degree of naphthalene degradation during about 24 h incubation of this bacterium in a mineral medium containing naphthalene as the only carbon source (Sazonova et al., 2023).
Bacteria that oxidize petroleum-derived pollutants from accidents related to the transportation or processing of petroleum may either exist as free-living organisms in marine waters and sediments or colonize the digestive tracts of fish, as demonstrated in recent studies on samples from the Baltic Sea (Polyak et al., 2020). Majority of the petroleum hydrocarbon-degrading bacteria from this sea belong to Actinobacteria and Betaproteobactera (Reunamo et al., 2013). It is essential to highlight that the structure of microbial communities can change significantly in the presence of crude oil, shale oil or diesel fuel, as demonstrated by studies on the coastal waters of the Baltic Sea (Viggor et al., 2013). Genomic studies, indicate that Baltic bacteria possess considerable potential for oil degradation, as evidenced by the abundance of alkB genes encoding alkane hydroxylases (Viggor et al., 2015). This conclusion was further supported by more comprehensive metagenomic analyses (Miettinen et al., 2019). Notably, recent studies on microbial genomes derived from Baltic Sea samples have shown that the metabolic potential of marine bacteria is closely linked to both the size of their genome and abiotic environmental factors (Rodríguez-Gijón et al., 2023).
Another group of pollutants includes pharmaceuticals, which can enter marine waters through their use and subsequent waste disposal (Wagil et al., 2015; Lindim et al., 2017). Among these, there are hormones, especially those belonging to the steroid group. Baltic bacteria capable of degrading steroid compounds were isolated, offering potential biotechnological applications for removing this kind of contaminants from natural marine environments. Examples of such steroid-degrading bacteria include strains from the Buttiauxella genus, which were demonstrated to be able to utilize cholesterol, oestradiol and testosterone as a carbon source when growing in a minimal medium (Zhang et al., 2011). In the case of Vibrio, genes encoding enzymes responsible for steroid degradation have been identified (Sang et al., 2012).
6.3 Putative drugs produced by Baltic bacteria
Various Baltic bacterial strains produce bioactive compounds with potential pharmaceutical significance. One such example is the Streptomyces champavatii strain, which is capable of producing a cyclic octapeptide called champacyclin, known for its antibacterial properties (Pesic et al., 2013).
The synthesis of various antimicrobial compounds has been observed in several Baltic bacterial strains associated with eukaryotic organisms. The proposed biological significance of this symbiosis lies in the protection of the eukaryotic host against other microorganisms, while the host, in turn, provides a favourable microenvironment for the bacterial symbionts. Notable examples of antimicrobial compound-producing Baltic bacteria associated with higher organisms include strains living in close proximity to bryozoan species, Marinomonas, Shewanella, and Vibrio (Heindl et al., 2010; Heindl et al., 2012). Additionally, bacteria associated with the brown alga Laminaria saccharina represent multiple species within the Alphaproteobacteria, Betaproteobacteria, Gammaproteobacteria, Bacteroidetes, Firmicutes, and Actinobacteria phyla (Wiese et al., 2009). Further examples include bacteria associated with the soft coral Alcyonium digitatum, classified as Bacillus methylotrophicus and Bacillus amyloliquefaciens (Pham et al., 2016), as well as Actinobacteria associated with marine sponges (Schneemann et al., 2010; Kunz et al., 2014). Among these groups, Actinobacteria appear to be the most abundant bacterial group in the Baltic Sea, comprising approximately 22–27% of bacterial communities in this environment (Holmfeldt et al., 2009; Jiang et al., 2011). These bacteria are known to produce a wide range of bioactive molecules of significant biotechnological relevance.
6.4 Baltic bacteria as metabolizers of minerals, (bio)plastic and lignin, and producers of potential nutrient proteins
Research has shown that bacteria isolated from Baltic bottom sediment contribute to both the corrosion and decomposition of submerged objects and structures, including shipwrecks, sunken objects, underwater pipelines. According to Cybulska et al. (2020), sulfate-reducing bacteria are the most abundant microorganisms involved in these processes and play a key role in microbial corrosion. In contrast, iron-converting bacteria have been found to be considerably less abundant. Moreover, sulfate-reducing bacteria, such as members of the family Desulfomicobiaceae identified in the Baltic Sea, have recently been proposed as potential agents responsible for the release of phosphorus from sediments (Cakmak et al., 2024). This is of particular biotechnological interest, as phosphorus is a critical element in eutrophication processes that enhance biological production.
Plastic is one of the most prevalent environmental contaminants, found in virtually all ecosystems, including marine waters and sediments. Consequently, its degradation is of significant environmental interest. Bacteria capable of growing on and potentially degrading plastic particles have been isolated from the Baltic Sea. Notable examples include Rubripirellula spp. (Kallscheuer et al., 2020), as well as various strains belonging predominantly to the phyla Actinobacteria and Gammaproteobacteria (Scales et al., 2021; Eronen-Rasimus et al., 2022). Conversely, the biotechnological production of biodegradable and environmentally friendly plastic represent a promising alternative (Aransiola et al., 2025; Multisanti et al., 2025). In this context, it is noteworthy that cold-adapted bacteria from the Baltic Sea, capable of producing polyhydroxyalkanoates, have been identified (Pärnänen et al., 2015). These biopolymers exhibit physicochemical properties similar to conventional plastics, and are therefore considered environmentally friendly materials. Although the specific bacterial producers were not identified, genetic analyses revealed the presence of phaC genes, which encode polyhydroxyalkanoate synthases (Pärnänen et al., 2015).
Lignin is a complex heteropolymer composed of alkyl and aromatic moieties. It is one of the most abundant biopolymers, occurring in the plant cell wall. Pseudomonas sp. strains capable of utilizing lignin-related compounds, including ferulate, p-coumarate, benzoate, and vanillin, have been isolated and characterised (Ravi et al., 2018). Depending on the growth medium, these bacteria excreted various usuful metabolites such as vanillyl alcohol, cis,cis-muconate, catechol, vanillate, and 4-hydroxybenzoate (Ravi et al., 2018).
The production of food using green technologies has emerged as a significant trend. Although plant cultivation remains an effective method of biomass production, it requires large areas of arable land and intensive irrigation, which presents increasing challenge due to limited water resources. Therefore, alternative biotechnological approaches are highly desirable. One promising strategy involves partial replacement of plant- and animal-derived proteins in the human diet with microbial proteins. Recently, a strain SoF1 of Xanthobacter sp. was isolated from the Baltic Sea (Klinzing et al., 2024). This autotrophic microorganism utilizes carbon dioxide as its sole carbon source and produces a protein-rich biomass that can be processed into a powder suitable for incorporation into various food products. Importantly, safety assessments have shown no evidence of genotoxicity or mutagenicity in the tested samples of this bacterial protein (Klinzing et al., 2024).
6.5 Assessing mutagenicity of marine water samples based on tests employing marine bacteria
Mutagenic pollution is another significant issue in the Baltic Sea and monitoring of this ecosystem for mutagenic pollutants is of critical importance. While numerous tests are available for the rapid assessment of mutagenicity in environmental samples, they rely on bacteria that do not thrive in marine waters, such as Salmonella enterica (Biran et al., 2010; Rueff et al., 2019; Dantas et al., 2020). Therefore, to enable effective and direct testing of the mutagenicity of marine water samples, an alternative assay was developed using the marine bacterium Vibrio harveyi, a species found in various marine habitats, including the Baltic Sea. Genetically modified V. harveyi strains were engineered to exhibit a high frequency of neomycin-resistant cells, which form colonies on nutrient agar containing antibiotics in the presence of low concentrations of mutagenic compounds (Czyz et al., 2000). Comparative studies demonstrated that this assay can detect lower concentrations of chemical mutagens than the traditional Ames test (Czyz et al., 2002). Furthermore, the V. harveyi assay was shown to be effective in detecting mutagenic pollution in Baltic Sea waters (Czyz et al., 2003). Subsequent studies led to the optimisation of assay conditions, further expanding its applicability. Another mutagenicity test employing the marine bacterium V. harveyi was also developed to use the bioluminescence phenomenon, characteristic of this species. Dark mutants of V. harveyi, bearing a point mutation in luxE, one of the genes responsible for light emission of bacterial cells, regained the ability to luminescence in the presence of mutagenic compounds due to the appearance of reverse mutations (Podgórska and Wegrzyn, 2006; Cheć et al., 2006).
Mutagenicity assays of biotechnological relevance, initially developed and optimized using marine bacteria, have since been applied in studies involving diverse marine samples, including water (Podgórska et al., 2007a), sediments (Podgórska et al., 2007b), and extracts from marine plants (Podgórska et al., 2008). A notable example includes the analysis of seawater from mussel farms, where the Vibrio harveyi bioluminescence assay was identified as “an optimal and rapid method for monitoring the water quality of mussel farms and as a tool for controlling the risks of pollution on mussel production and its safety for human consumption” (Ruiz et al., 2013). In another application, the assay was employed to assess the mutagenic, genotoxic, and cytotoxic activities of surface water samples from a wetland ecosystem in India (Kaur et al., 2014). Collectively, these studies underscore the broader applicability of biotechnological tools developed using marine bacteria, highlighting their potential for environmental monitoring and risk assessment across geographically and ecologically distinct regions.
In summary, Baltic Sea bacteria exhibit diverse biotechnological capacities that extend far beyond Cyanobacteria. Reported applications include the production of enzymes with unique properties, the degradation of complex organic pollutants, and the biosynthesis of bioactive compounds with pharmaceutical potential. Several taxa, particularly Actinomycetes and Vibrio species, stand out as prolific producers of bioactive molecules.
Other Baltic bacteria have been implicated in mineral cycling, plastic degradation, as well as bioplastic and microbial protein production. In addition, Baltic isolates have served as the basis for innovative environmental monitoring tools, such as mutagenicity assays employing genetically engineered and/or luminescent Vibrio harveyi strains, which offer higher sensitivity than conventional methods.
7 Biotechnological potential of Baltic fungi
The Baltic Sea harbours a variety of fungal species (Hassett et al., 2020; Tibell et al., 2020). However, compared to other microorganisms, knowledge of their true taxonomic diversity and environmental significance remains limited (Utermann et al., 2020a,b; Mazur-Marzec et al., 2024). These organisms have attracted the attention of researchers and biotechnologists due to the potential applications of their bioactive secondary products. In Baltic fungi, especially those living in symbiosis with aquatic animals and macrophytes or living in sediments, antimicrobial, anticancer, antiviral, and other activities have been observed (Figure 5).
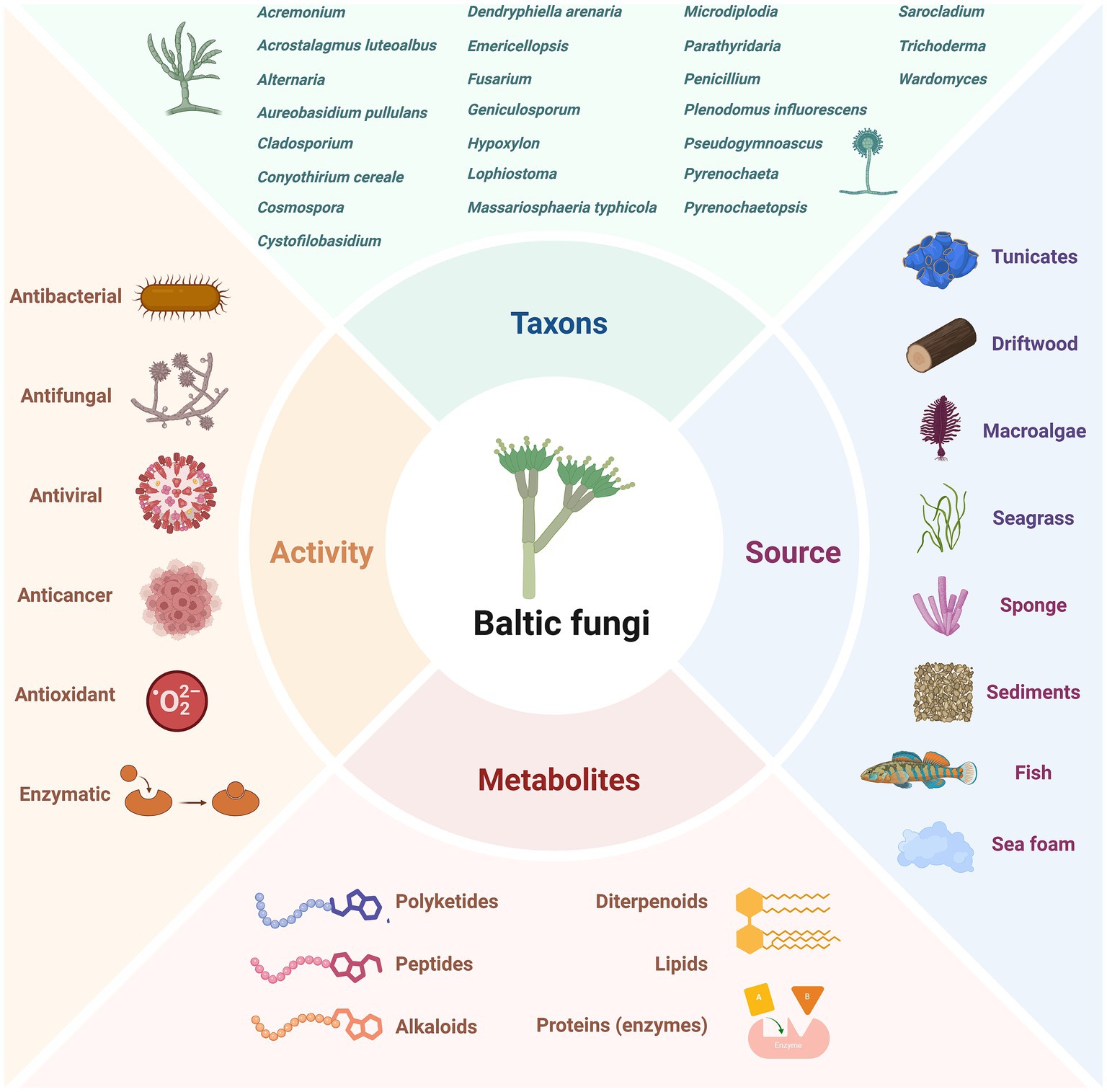
Figure 5. Diversity of investigated Baltic Sea fungi, their sources, produced metabolites and observed activities. The figure was created using BioRender.com.
In Penicillium chrysogenum strains from various sources, the presence of sorbicillin-derived alkaloids as well as sorbicillactones, was documented. This class of metabolites is also produced by the Baltic Sea fungi (Bringmann et al., 2007). The strain KIPB33 isolated from the Baltic sponge Callopora aurita showed a higher yield of sorbicillactone A (Figure 6) than other P. chrysogenum strains. Sorbicillactone A demonstrated highly selective activity against murine leukemic lymphoblast L5178y cells while exhibiting minimal cytotoxicity to healthy cells. To supply sufficient material for preclinical investigations, an efficient process for its large-scale biotechnological production was developed (Bringmann et al., 2007). Lindgomycin is another bioactive metabolite produced by sponge-associated Baltic fungi (Wu et al., 2015). Lindgomycin from Massariosphaeria typhicola displayed antimicrobial activity against different Staphylococcus strains.
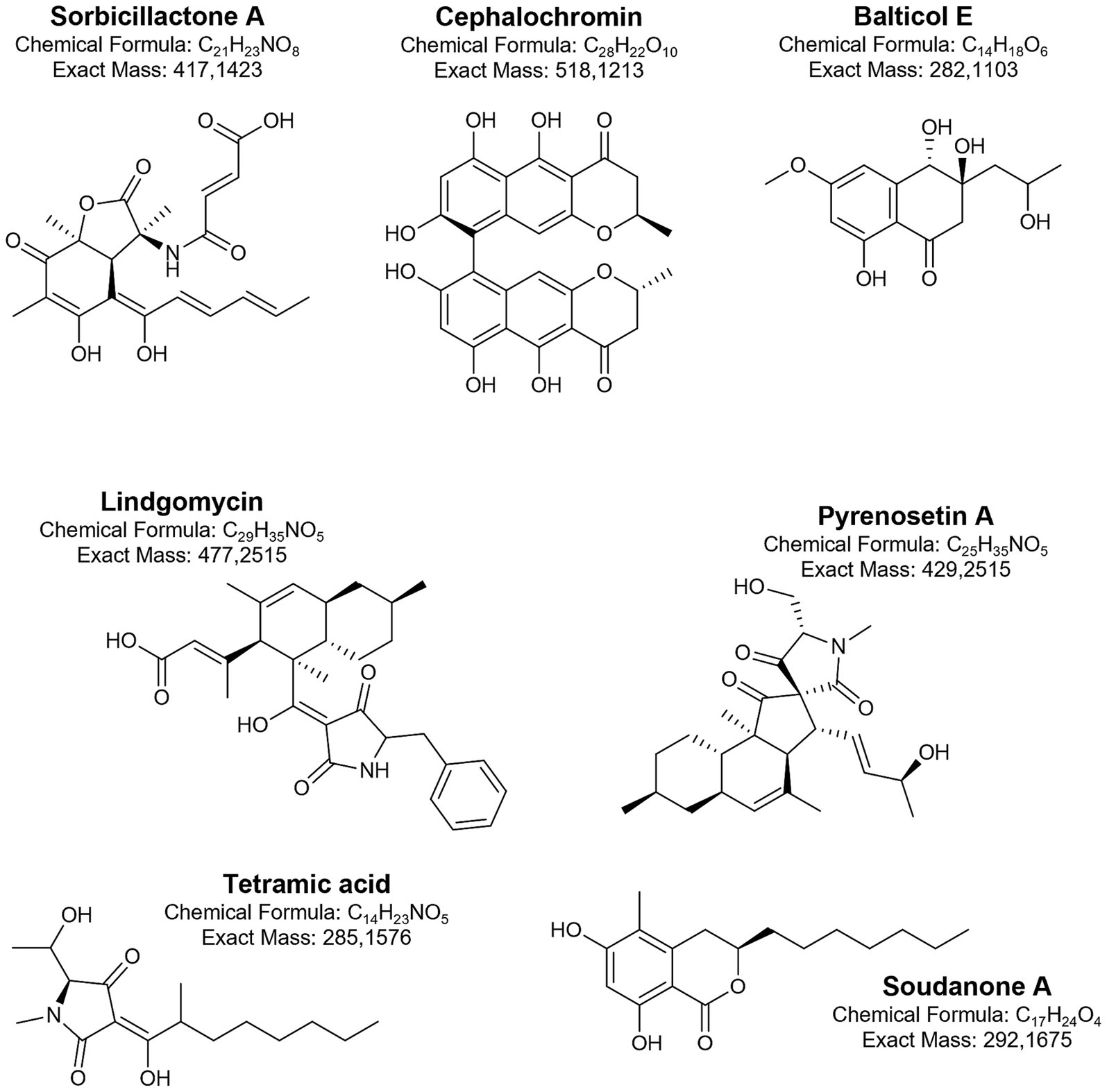
Figure 6. Biologically active metabolites produced by Baltic fungi.
Fungal isolates from the Baltic samples (water, sediment, seafoam, and plant debris) revealed the ability to interact with co-cultured phytopathogenic bacteria and fungi. In these cultures, interspecies communication was triggered, and an increase in chemical diversity was observed (Oppong-Danquah et al., 2018). The identified metabolites were classified as terpenes, alkaloids, peptides, and polyketides. In further studies, the large-scale co-cultivation of Cosmospora sp. with the phytopathogen Magnaporthe oryzae supplied sufficient biomass for the isolation and identification of several classes of metabolites, including pseudoanguillosporins A and B, naphtho-γ-pyrones, cephalochromin, and ustilaginoidin G, and ergosterol. Among other compounds, three known soudanones A, E, and D and two new soudanones variants, soudanones H and I, were detected (Figure 6) (Oppong-Danquah et al., 2022). Of these, soudanones E and D showed antimicrobial activity against M. oryzae and Phytophthora infestans, while pseudoanguillosporin A was active against Pseudomonas syringae, Xanthomonas campestris, M. oryzae, and P. infestans. In another co-culture experiment with sediment-derived fungi, Plenodomus influorescens and the phytopathogenic Pyrenochaeta nobilis, the anti-phytopathogenic activity of cephalochromin against X. campestris (IC₅₀ = 0.9 μg/mL) and P. infestans (IC₅₀ = 1.7 μg/mL) was revealed (Oppong-Danquah et al., 2020a). Moreover, fungi (Penicillium sp.) with activity against plant and human pathogens were also isolated from sea foam (Oppong-Danquah et al., 2020b). The most potent effects of extract from Penicillium strains were recorded in the case of M. oryzae and Candida albicans (IC₅₀ = 2.2 μg/mL and 6.3 μg/mL, respectively).
Another group of Baltic fungi, endophytes, primarily isolated from macroalgae, also gained attention for their promising bioactive properties. Tetramic acid sch210972 (Figure 6) produced by the endophytic ascomycete Microdiplodia sp. isolated from the green alga Enteromorpha sp. inhibits human leucocyte elastase (HLE) with an IC50 = 1.04 μg/mL, and shows moderate effects on the growth of Bacillus megaterium (Neumann et al., 2009). Generally, marine fungi, mainly from the genera Penicillium, Cladosporium, and Aspergillus, provide the majority of the known natural tetramic acid compounds (Jiang et al., 2020). The pharmacological potential of these compounds is related to their wide range of activities, including cytotoxic, antibacterial, and antifungal effects. From other endophytes, Coniothyrium cereale and Wardomyces anomalus OS4T3–2-1, cytotoxic phenalenones (Elsebai et al., 2011; Elsebai et al., 2012; Elsebai et al., 2016) and xanthone derivatives (Abdel-Lateff et al., 2003) were isolated.
From the brown algae Fucus vesiculosus, two endophytic fungal strains, Pyrenochaetopsis sp. FVE-001 and FVE-087, with anticancer activity, were isolated (Fan et al., 2020a,b). These strains were found to produce new decalinoylspirotetramic acid derivatives, pyrenosetins A-F (Figure 6), and the known decalin derivative—phomasetin. Pyrenosetins A and B demonstrated significant inhibitory activity against malignant melanoma A-375 cells, with IC₅₀ values of 2.8 and 6.3 μM, respectively (Fan et al., 2020a). Pyrenosetin D was toxic toward A-375 and HaCaT cells with IC50 values of 77.5 and 39.3 μM (Fan et al., 2020b). Pyrenosetin E inhibited the growth of A-375 with IC50 = 40.9 μM (Fan et al., 2022). Geniculosporium sp. 6,580, associated with the red algae Polysiphonia from the Baltic Sea, was cultured to yield 11 new botryane sesquiterpenoids (Krohn et al., 2005). Some of these compounds exhibited mild herbicidal, antifungal, and antibacterial activities. Their structures differed from known botryanes in substitution patterns and altered oxidation sites, alkylation, and unsaturation.
Extracts from fungal strains (mainly Penicillium species) isolated from the leaf and the root rhizosphere of Baltic eelgrass Zostera marina, showed strong anti-quorum sensing activity. They also displayed antimicrobial or anti-biofilm activity against Gram-negative environmental marine and human pathogens (Petersen et al., 2019). In later research (Tasdemir et al., 2024), the fungal (and bacterial) isolates from the microbiome of Z. marina were found to possess a wide spectrum of antipathogenic properties. The most active fungi, Cladosporium halotolerans and Fusarium sp., were linked to decaying Z. marina leaf surface. In the fungi, diketopiperazines, epipolythiodioxopiperazines, polyketides, diterpenoids, peptides, naphthoquinones, polyol lipids, and peptaibols were detected (Tasdemir et al., 2024). The analysis of three Fucus species with varying levels of fouling revealed that the least fouled F. distichus subsp. evanescens harbors abundant Mucor and Alternaria, known for producing antimicrobial and antifouling compounds, which may contribute to its fouling resistance (Oppong-Danquah et al., 2023).
In Baltic fungi collected from driftwood, six new naphthalenone derivatives (balticols A-F) (Figure 6), along with the known metabolite altechromone A, were isolated (Shushni et al., 2009; Kolařík et al., 2017). Among these, balticol E demonstrated the strongest antiviral activity against Herpes simplex virus type I (IC₅₀ 0.01 μg/mL). Balticolid, a novel class of 12-membered macrolide, active against antiHSV-1, was also identified (Shushni et al., 2011). Intrigued by its biological activity and interesting molecular architecture, an efficient approach to total balticolid synthesis from the known and cost-effective starting material was reported (Sudhakar et al., 2019). However, even if many in vitro tests with the application of these compounds were performed and showed promising results, limited data from in vivo experiments are available to date (Yi et al., 2020). Another fungus isolated from driftwood, Lophiostoma, produces oxasetin, an antibacterial polyketide that demonstrated in vitro activity against Vibrio anguillarum, Flexibacter maritimus, and Pseudomonas anguilliseptica (Shushni et al., 2013).
Some Baltic Sea fungi have been identified as degraders of shipwrecks (Björdal, 2012). Fungi involved in wood degradation produce cellulases, hemicellulases, and other enzymes capable of breaking down plant cell walls (Andlar et al., 2018). These enzymes are useful for producing biofuels, in the pulp and paper industry, and for converting agricultural waste into valuable products. Fucodainase was purified from the obligate marine fungus Dendryphiella arenaria TM94, isolated from Baltic Sea sand, and showed optimal activity at pH 6 and 50 °C (Wu et al., 2003). Fucoidanases can be involved in the production of sulfated fucooligosaccharides, compounds with a wide range of biological activities and pharmaceutical potential (Barzkar et al., 2023).
Fungi have been also studied for their ability to degrade and detoxify pollutants in marine environments. Research in the Baltic Sea suggests that the fungal communities within this ecosystem hold considerable potential for the bioremediation of petroleum hydrocarbons and other environmental pollutants (Al-Nasrawi, 2012; Miettinen et al., 2019). For example, diesel oil spills in the surface waters of the Baltic Sea increased the relative abundance of fungi from the phylum Ascomycota. The use of Baltic fungi as part of a hydrophobic sorbent for oil removal from water has also been tested (Paulauskiene et al., 2023).
Exposure experiments with polyethylene and polystyrene conducted in the Baltic Sea, with wood as a control surface, revealed that microplastic-associated fungal communities were distinct from those in the surrounding water and on wood (as a natural substrate) (Kettner et al., 2017). Understanding how fungal communities form and thrive on microplastics can provide valuable insights for developing methods for plastic waste degradation and advancing biofilm engineering.
Based on the studies discussed here, it can be concluded that fungi, as one of the least explored groups of microorganisms in the Baltic Sea, offer an opportunity to introduce innovative solutions that could both improve environmental conditions and contribute to better human health outcomes.
Baltic fungi represent an underexplored resource for biotechnology, offering diverse bioactive metabolites with pharmacological potential, industrially relevant enzymes, and promising applications in bioremediation and antifouling. While large-scale production, co-cultivation, and total synthesis demonstrate methodological feasibility, most findings remain limited to in vitro studies. Future research should prioritise in vivo validation, broader taxonomic and ecological exploration, and the systematic integration of biodiversity, functional traits, and metabolite profiling to unlock their full biotechnological potential.
8 Baltic viruses and their biotechnological potential
Reports on viruses from the Baltic Sea are relatively scarce. Among viruses infecting eukaryotic organisms, those specific to algae are most abundant, however, viruses pathogenic to insects, fish and seals have also been documented. The presence of viruses specific to pigs and humans strongly suggests the occurence of biological contaminants originating from urban or industrial environments. Nevertheless, to our knowledge, there are no reports on the biotechnological application of Baltic viruses specific to eukaryotes. Conversely, bacteriophages (or shortly – phages) are highly abundant viruses in the Baltic Sea (Mazur-Marzec et al., 2024). Their significant potential for biotechnological application is discussed below.
One of the most prominent biotechnological application of phages is their use in combating pathogenic bacteria affecting various organisms (Figure 7). In the Baltic Sea, phages infecting members of the Flavobacteriaceae family are among the most abundant. Some of these bacteria are dangerous pathogens of marine animals, particularly fish (Nilsson et al., 2019). Therefore, bacteriophages could serve as effective agents for controlling such pathogens, especially in marine fish aquaculture systems (Daniel et al., 2021). Notably, phages specific to Cellulophaga baltica (Holmfeldt et al., 2007), Flavobacterium spp. (Nilsson et al., 2020; Daniel et al., 2021), and species from the genus Rheinheimera (Nilsson et al., 2019; Hoetzinger et al., 2021) have been identified in the Baltic Sea. In the addition to their potential use in aquaculture, bacteriophages may also help control bacterial infections in wild marine organisms. For example, the Baltic jellyfish Aurelia aurita is susceptible to infections caused by a variety of bacterial pathogens. Phages isolated from Baltic Sea water were shown to infect and efficiently lyse bacterial colonizers of jellyfish, including strains belonging to the genera Citrobacter, Escherichia, Shigella and Staphylococcus (Stante et al., 2024).
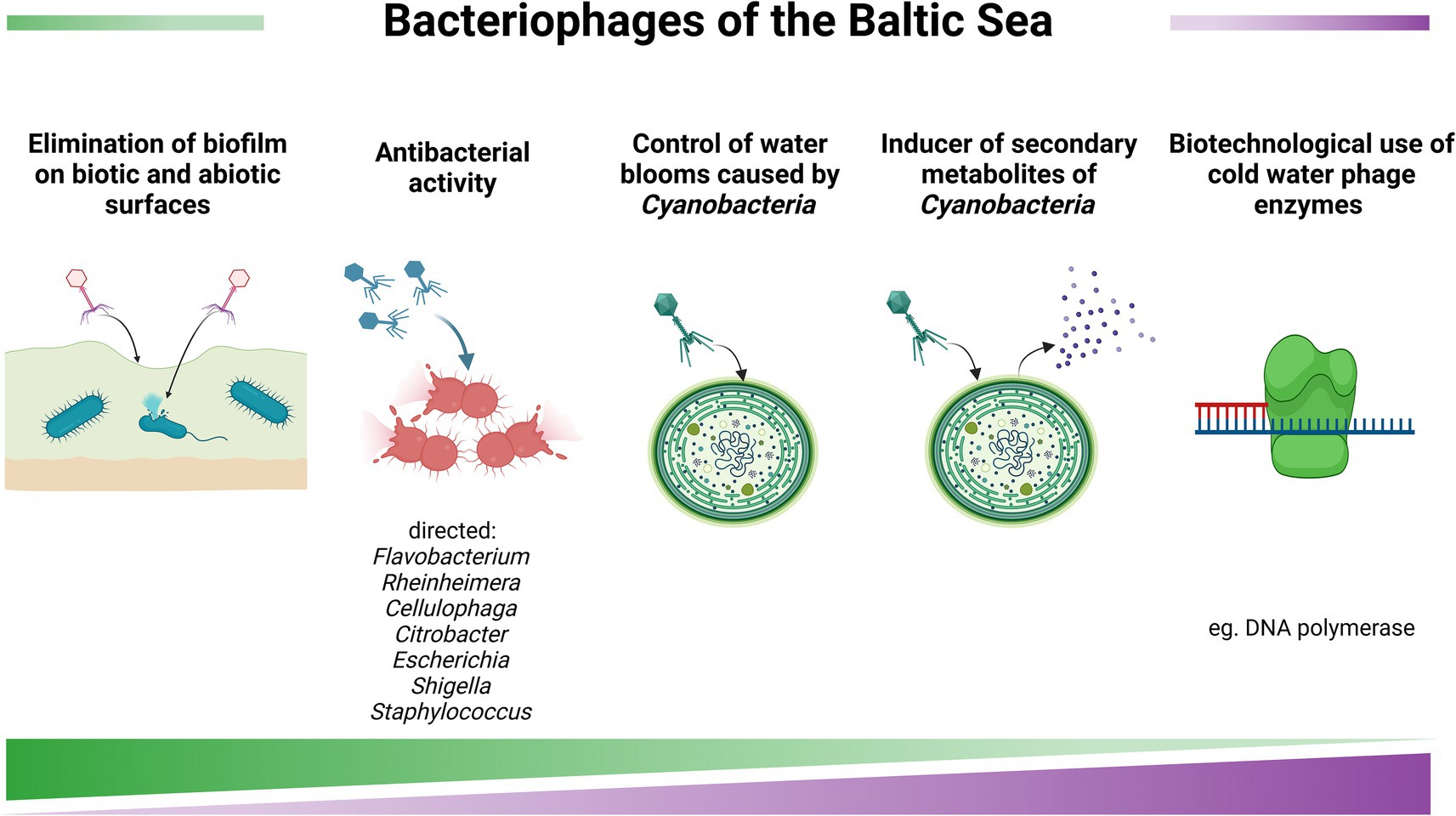
Figure 7. Baltic viruses and their biotechnological potential. The figure was created using BioRender.com.
Although cyanobacteria are not direct pathogens to animals and humans, they can cause blooms and produce toxic compounds (Karlson et al., 2021). Therefore, it is potentially possible to use bacteriophages to control cyanobacterial blooms using specific cyanophages that would eliminate specific species of these microorganisms. Indeed, cyanophages were isolated from the Baltic Sea and characterised (Šulčius et al., 2015; Šulčius et al., 2019). As documented in previous section of this work, certain metabolites produced by cyanobacteria may have potential medical significance (Grabowski et al., 2024). However, large-scale production of such compounds is problematic, as the conditions that stimulate their synthesis are largely unknown. It was demonstrated that infecting cyanobacterial cells with cyanophages can enhance the production of various metabolites, including nodularin, anabaenopeptins, aeruginosins, and spumigins (Šulčius et al., 2018). Therefore, controlled cyanophage infection could be considered a potential method for stimulating the synthesis of such compounds.
Considering the antibacterial potential of bacteriophages, one of the interesting approaches may involve the use of phages with long tails in their virions. Such phages may prove particularly effective in combating biofilm-forming bacteria. Indeed, phages with these characteristics have been isolated from the Baltic Sea (Jakubowska-Deredas et al., 2012).
A fascinating group of viruses consists of phages isolated from the Baltic Sea ice. These phages infect psychrophilic hosts from the genera Flavobacterium and Shewanella and are capable of effectively replication within their host cells at temperatures as low as 3–4 °C (Luhtanen et al., 2014; Senčilo et al., 2015). Interestingly, the genomes of these psychrophilic bacteriophages encode various enzymes with potential biotechnological applications, including DNA polymerases, helicases, primases, DNA ligases, RNase H, and proteins involved in genetic recombination, such as homologues of NinB and NinG (Senčilo et al., 2015). Decreasing temperatures of various processes of in vitro DNA manipulation would be beneficial in genetic engineering. Thus, cloning corresponding genes from genomes of Baltic bacteriophages and overproduction and purification of their products might be of biotechnological significance.
Although research on viruses in the Baltic Sea is still less extensive than on other Baltic microorganisms, the available findings already highlight their considerable promise for biotechnology. Among them, bacteriophages are the most prominent group, with demonstrated potential to control bacterial pathogens relevant to aquaculture and marine ecosystems.
Cyanophages isolated from the Baltic Sea not only offer possibilities for mitigating harmful cyanobacterial blooms, but may also stimulate the production of valuable bioactive metabolites. Particularly intriguing are phages from Baltic Sea ice that thrive in low-temperature environments and encode enzymes, which could become powerful tools for genetic engineering and molecular biology under cold conditions.
9 Culture collections
Culture collections play a crucial role in preserving both living organisms and their genetic resources. They also provide organisms and their products for various areas of human activity, contributing to advancements in research, medicine and bioindustry (Garibay et al., 2024).
Effective management of these collections requires standardized protocols for cultivating organisms, detailed characterisation, and authentication of cultures, as well as the involvement of well-trained personnel. Furthermore, information about the source, taxonomic position, and ecological relationships of the preserved organisms should be accessible. In particular, if available, genomic data, are essential for identifying strains with biotechnological potential (Garibay et al., 2024).
In the Baltic countries, at least 79 culture collections of microorganisms have been identified, (Supplementary Table S3). In the living resources cathegory, microalgae (more than 100,000 isolates) and bacteria (more than 200,000 isolates) are the most abundant. However, it is challenging to estimate the contribution of Baltic Sea microorganisms in these collections, primarily due to insufficient characterisation and documentation of deposited strains. Another significant issue is the limited availability of online searchable catalogues, which hinders the broader utilization of the resources. Holdings of many research institutions remain uncatalogued, and thus, maintained microbial strains are used exclusively by local researchers. Improved documentation and cataloguing of these resources could significantly enhance their visibility, accessibility and value to the scientific community (Kurtzman and Labeda, 2009).
10 Genetic resources of Baltic microorganisms
There is a paradigm shift from traditional biology to bioinformatics that is revolutionizing exploitable biology driven by a rapid development of DNA sequencing and amplification technologies, annotation, proteome analysis, and phenotypic inventorying, resulting in the establishment of huge databases that can be mined in order to generate useful knowledge (Bull et al., 2000). Currently, sequencing technologies constitute a cost-effective and time-saving tool for sequencing microorganisms and samples that may harbor genes with potential biotechnological applications. Particularly, there is a remarkable increase in the number of microbial genomes retrieved from marine systems (e.g., Chen et al., 2024). There are numerous sequence databases, with the most well-known GenBank database,1 maintained by the National Center for Biotechnology Information (NCBI; a part of the National Institutes of Health in the United States) (Sayers et al., 2025), as part of the International Nucleotide Sequence Database Collaboration (INSDC). These developments are particularly relevant for microbial research, as microbial genomes and metagenomes are frequently used and redistributed via online databases, effectively decoupling physical access to genetic materials from downstream utilisation of sequence data. Consequently, the increasing availability of genomic data has raised important questions regarding access, benefit-sharing, and the governance of so-called digital sequence information (DSI). This issue has become a focal point in negotiations under the Convention on Biological Diversity (CBD) and its Nagoya Protocol, particularly during the adoption of the Kunming–Montreal Global Biodiversity Framework (GBF) in December 2022, which recognised the need for a multilateral mechanism to ensure the fair and equitable sharing of benefits arising from the use of DSI on genetic resources (CBD, 2022). The concrete design of this mechanism, including decisions on traceability, provenance metadata, and obligations for users and databases, remains under active development.
In the NCBI genome database, a total of 2,871,411 genomes are available, of which, 2,314,807 are annotated, and 39,268 are reference genomes.2 Due to a huge number of records, of which a large part lacked information on the isolation locations of the organism, we used the BacDive database to search for the genomes of Baltic Sea bacteria3 (Schober et al., 2025). The search with advanced options, using the geographical location ‘Baltic Sea’, yielded 70 results; however, genomic information was available only for 31 strains (Supplementary Table S4). Among 8,967 genomes assigned to the Cyanobacteriota/Melainabacteria group, there were 6231/2216/84/436 genomes assembled to levels of contig/scaffold/chromosome/complete genome. 8/1/14 genomes (scaffold/chromosome/complete genome level) belong to Baltic organisms.4 These genomes are not a comprehensive representation of the Cyanobacteria phylum: 14 represents the Nostocales order, 6 Synechococcales order (including one metagenome-assembled genome, MAG,) and one from the order Pleurocapsales. Also, two MAGs were obtained from unculturable cyanobacteria. The primary objectives of those genomic studies were (1) to identify gene clusters associated with bioactive metabolite production and, more broadly, to determine the biosynthetic potential of Baltic cyanobacteria, (2) to verify their taxonomy, (3) to determine adaptation to variable salinity conditions, (4) to identify host-microbiota interactions in holobiont (Voss et al., 2013; Teikari et al., 2018; Teikari et al., 2019; Österholm et al., 2020; Ahmed et al., 2021; Bonthond et al., 2021; Dreher et al., 2021; Heinilä et al., 2022). Baltic Cyanobacteria with available genomes are listed in Supplementary Table S5.
Among 29,056 genomes assigned to the Archaea kingdom there were 16,139/12058/98/761 genomes assembled to levels of contig/scaffold/chromosome/complete genome. To narrow down the search criteria, we took into account only representative genomes, with RefSeq category (738 genomes in total; with 252/118/126/360 genomes assembled to levels of contig/scaffold/chromosome/complete genome). Also, we checked among 2,604 MAGs, with 1477/450/24/653 assembled to levels of contig/scaffold/chromosome/complete genome. Unfortunately, none of them belong to Baltic organisms.5
Regarding the biodiversity of Baltic bacteriophages, the NCBI Database reports 199 bacteriophages isolated from various regions of the Baltic Sea with sequenced genomes.6 Among these, sixteen phages are classified as unassigned members of the Caudoviricetes class (Supplementary Table S6), while four phages are small, tailless members of the Microviridae family, characterised by ssDNA genomes of approximately 6 kb in size. The remaining 179 phages are classified into 20 genera within the Caudoviricetes class.
As reported by Šulčius and Holmfeldt (2016), phages that infect Cellulophaga baltica (formerly known as Flavobacterium aceae) belong to the following genera: Akihdevirus, Bacelvirus, Baltivirus, Callevirus, Cbastvirus, Cebadecemvirus, Cellubavirus, Helsingorvirus, and Lightbulbvirus. These phages possess dsDNA genomes ranging from 38.5 kb to 145.9 kb (Holmfeldt et al., 2007; Šulčius and Holmfeldt, 2016). The largest collection of phages infecting Rheinheimera sp. was created by Nilsson et al. (2019), comprising 54 phages from the Barbavirus genus, with genomes ranging from 80.2 kb to 84.6 kb.
The majority of Baltic phages with sequenced genomes (over 100) infect Flavobacterium spp., and more than half of these belong to the Elemovirus genus, with genomes around 60 kb. Other abundant genera include Muminvirus (genomes of 37.5 kb to 38.5 kb) and Lillamyvirus (genomes of 37.5 kb to 39 kb) (Nilsson et al., 2020; Hoetzinger et al., 2021). The largest genomes among Baltic Flavobacterium phages (~166 kb) belong to the Immutovirus genus (Hoetzinger et al., 2021).
The only cyanophage from the Baltic Sea with a sequenced genome in the NCBI database is the vB_AphaS-CL131 virus, which infects Aphanizomenon flos-aquae (Šulčius et al., 2015; Šulčius et al., 2019). Recently, one of phages isolated from Kiejl Fjord infects Staphylococcus sp. and is currently the only example of a Baltic virus specific to Bacillota (Stante et al., 2024). Among the unassigned members of the Caudoviricetes class, Shewanella viruses were also identified. Interestingly, the isolation and cultivation of ice bacteria and cold-active phages from the Baltic Sea ice were reported (Luhtanen et al., 2014; Senčilo et al., 2015).
11 Rules of access to microbial resources of the Baltic Sea in the context of the Nagoya protocol
11.1 International law
Researchers and organizations (institutions, companies, etc.) conducting research and accessing microbial resources must adhere to international and national regulations. Many countries require compliance with these regulations to achieve intellectual property protection. A lack of awareness or non-compliance may jeopardize their research and innovation outcomes (Schneider et al., 2022). Moreover, there has been some debate about whether publishers should mandate documentation of compliance with the Nagoya Protocol for manuscript submissions dealing with genetic resources (Kumar, 2018).
Researchers need to be aware that, as reaffirmed by the Convention on Biological Diversity (CBD),7 adopted in 1992 at the Rio Earth Summit, States have sovereign rights over their biological resources within their jurisdiction. In the case of coastal countries, the right to explore, exploit, conserve and manage natural resources also includes marine waters, the seabed and its underground of the country’s territorial waters, and extends to the exclusive economic zone, up to 200 nautical miles from the baseline as stated by the 1982 United Nations Convention on the Law of the Sea.8 This means that each state has the authority to regulate and manage its biological resources in accordance with its laws and policies. This includes controlling access to these resources, setting conditions for their use, and establishing mechanisms for sharing the benefits derived from their utilization. Anyone seeking to access genetic resources from a country of origin must obtain prior informed consent (PIC) from the relevant authorities (unless the country has specified otherwise) and establish mutually agreed terms (MAT) for their use (Figure 8).
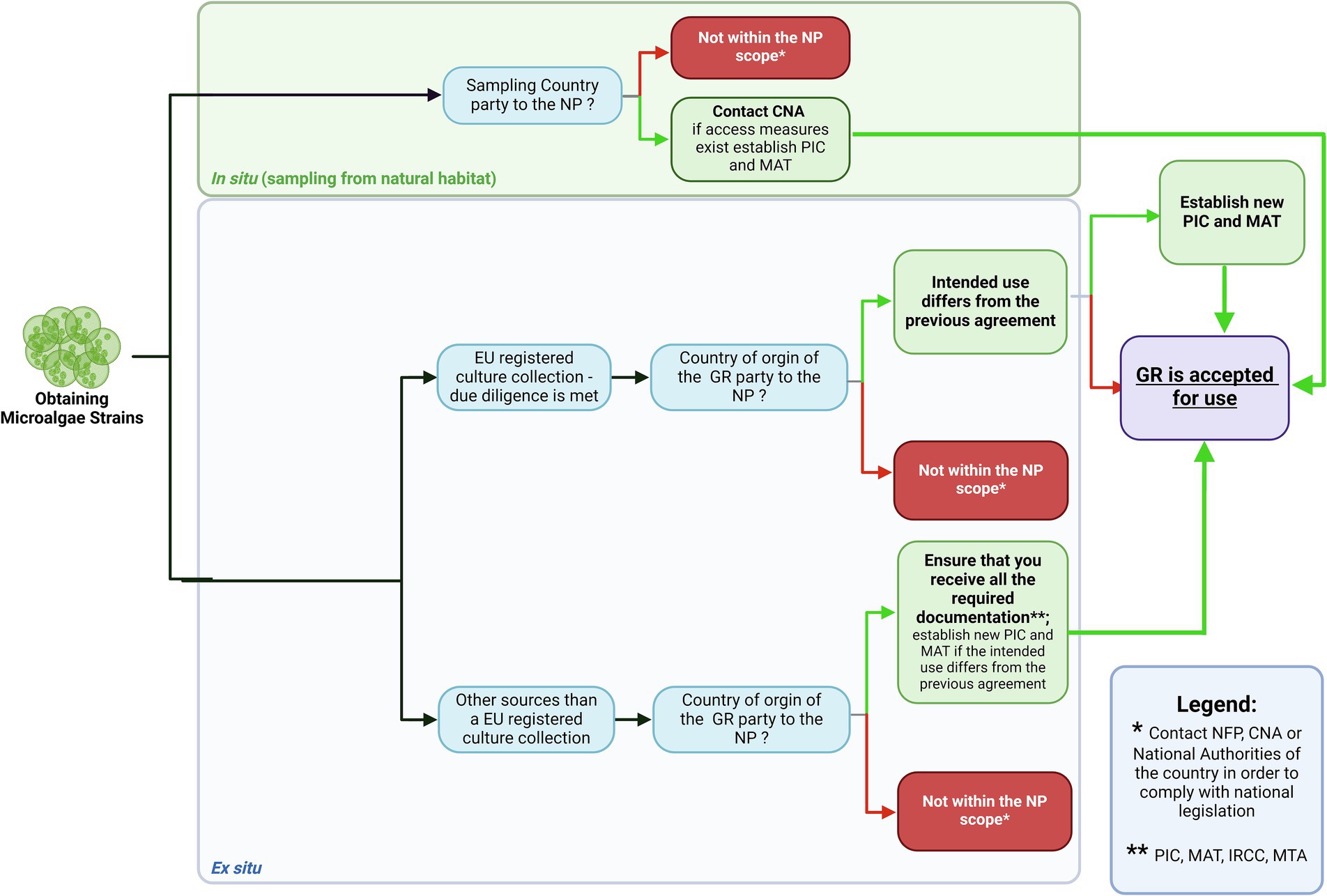
Figure 8. Decision framework proposed by Martins et al. (2020) presenting the steps necessary to obtain access to genetic resources (GR) according to Nagoya Protocol (NP). The figure was created using BioRender.com.
Based on these principles, another international agreement, the Nagoya Protocol on Access to Genetic Resources and the Fair and Equitable Sharing of Benefits Arising from their Utilization,9 was adopted in 2010.
Under this regulation, commonly referred to as ABS regulation (Access and Benefit Sharing), all countries party to the NP are required to designate (i) an ABS National Focal Point (NFP) responsible for providing information on how to obtain the PIC and establish MAT, and (ii) Competent National Authorities (CNA), which grant access to genetic resources (Figure 8). Additionally, countries must make their legislative, administrative, and policy measures related to ABS available through a searchable database called the ‘ABS Clearing House.’10 This website should be a starting point for anyone seeking access to bioresources. However, it is important to note that even countries that have not signed or implemented the NP may have national legislation governing access to genetic resources and ABS.
While a detailed overview of the NP is beyond the scope of this paper, it is worth mentioning that the NP broadly defines genetic resources (GR). It includes (i) any material of plant, animal, microbial or other (non-human) origin containing functional units of heredity that can be explored through research and utilized in development, thus having actual or potential value, as well as (ii) their derivatives—naturally occurring biochemical compounds resulting from genetic expression or metabolism, such as proteins (e.g., enzymes), lipids, RNA, or secondary metabolites.
Utilization refers to the use, exploration, research and/or development of GR. It is important to note that this term encompasses basic research, and many research activities—regardless of whether they have a commercial component—fall within the scope of ABS regulation, as they generate new knowledge and may, in the future, lead to the development of new commercial products. While some national ABS laws differentiate between non-commercial and commercial use (e.g., with varying benefit-sharing requirements), it is a common misconception that NP applies solely to commercial research. Another misunderstanding is that benefit-sharing is limited to monetary benefits. In fact, the NP also foresees non-monetary benefits, such as data sharing, joint publications, project partnerships, education, training, and shared reference collections-practices with a long-standing tradition in academia. In addition to the CBD and the Nagoya Protocol, it is also worth mentioning recent legal developments concerning marine biological diversity in areas beyond national jurisdiction (BBNJ). These regions, often referred to as the “high seas,” lie outside the sovereignty of individual States and until recently lacked a comprehensive legal framework for access and benefit-sharing of marine genetic resources. In June 2023, the United Nations adopted the Agreement under the United Nations Convention on the Law of the Sea on the conservation and sustainable use of marine biological diversity of areas beyond national jurisdiction (the “BBNJ Treaty”), introducing rules for access, use, and fair sharing of benefits arising from such resources.11 The BBNJ Treaty will enter into force on 17 January 2026, 120 days after the 60th instrument of ratification is deposited. To ensure coherent implementation in Europe, the European Commission has proposed a Directive of the European Parliament and of the Council on the conservation and sustainable use of marine biological diversity of areas beyond national jurisdiction, designed to transpose the BBNJ Treaty into EU law and extend ABS principles into the high seas governance regime.12
11.2 The Baltic Sea case
The Baltic Sea, surrounded by nine countries: Denmark, Germany, Poland, Lithuania, Latvia, Estonia, Finland, Sweden and Russia, is a confined sea where opposite coasts are never more than 400 nautical miles apart, and most of the maritime boundaries have been settled (Franckx, 2021). Depending on their geographic origin, Baltic microbial resources fall under the jurisdiction of either of the bordering countries. While all Baltic-bordering countries are Parties to the CBD (at the time of writing), only five—Denmark, Estonia, Finland, Germany and Sweden are Parties to the NP (Supplemntary Table S7).
However, since the European Union (EU) ratified the protocol on May 16th 2014 (EU, 2014), it became a Party to the NP, meaning that all EU member states, as well as users of GR and associated traditional knowledge (aTK) within the EU, must comply with EU regulations on ABS (EU ABS, 2014). The main objectives of these two adopted documents, the ABS Regulation (EU No 511/2014) and the Implementing Regulation (EU, 2015), are to prevent the utilization of the GR or aTK in the Union that were not accessed in accordance with the requirements of NP, to promote fair and equitable benefit-sharing, and to ensure legal certainty regarding the use of these resources. Researchers within the EU who use genetic resources are now legally obliged to follow specific procedures, including obtaining permits, maintaining records, and submitting due diligence declarations. An exemplary decision framework proposed by Martins et al. (2020) presents some initial steps necessary to obtain microalgae GR.
To assist users in navigating the complexities of ABS law, the European Commission published a guidance document (Guidance Document, 2021). While not legally binding, it serves as a useful reference for clarifying many of the intricate issues arising from ABS regulations.
However, the decision to regulate access to their GR under the NP remains within the competence of individual Member States. Fortunately, for those wishing to utilize Baltic genetic resources, some countries bordering the Baltic Sea have chosen not to assert sovereign rights over their genetic resources under the NP, thus reducing administrative burdens. This means that, in practice, these resources are freely accessible, and there is no requirement to establish Prior Informed Consent or negotiate Mutually Agreed Terms for their utilization.
Nevertheless, users who obtain microbial resources in situ may need to comply with other national legislation, such as those related to protected areas, which should be considered before sampling the country’s sovereign genetic resources. An overview of access and benefit-sharing policies in nine Baltic coastal countries, along with their NP status, is summarised in Supplementary Table S7. The relevant information for each country can be found on the ‘ABS Clearing House’ website,13 while details on access rules are available in the countries’ Interim National Reports on the Implementation of the Nagoya Protocol.
Russia, the only non-EU country in the Baltic Sea, is not a Party to NP; thus, the utilisation of GR originating in Russia is outside the scope of the EU ABS Regulation. However, the EU recommends that users of such resources should comply with the country’s national legislation or regulatory requirements and respect any mutually agreed terms entered into.
11.3 Obtaining the Baltic genetic resources ex-situ
The NP and EU ABS regulations also apply to genetic resources (e.g., strains, specimens) from culture collections held in scientific institutions. Within the EU, users accessing material from these collections (access of GR) are required to seek, obtain, and maintain the relevant information (such as PIC, MAT), transfer it to the subsequent users (exercising due diligence), and submit a due diligence declaration at a designated checkpoint (either at the research funding stage or upon the final development of a product), as specified in the Implementing Regulation (EU) 2015/1866 (Article 7). To facilitate compliance and reduce administrative requirements, the European Commission (EC) has established a voluntary Register of Collections within the Union. Collections are granted registered status upon verification by the relevant national competent authority that they meet the criteria specified in Regulation (EU) No 2015/1866. Users accessing genetic resources from the registered collection are deemed to have exercised due diligence in obtaining all necessary information. At the time of writing, three collections have been recognised, as recognized through a Commission Decision.14
Accessing genetic resources from collections originating in countries that have chosen not to impose access regulations does not create ABS obligations on the user. Since most Baltic-bordering countries allow grants of free access to their GR, culture collections in the region can provide these resources without the required PIC and MAT.
12 Future perspectives
Despite the discovery of many promising avenues for the biotechnological application of Baltic Sea microorganisms, the successful commercialisation of their potential depends on addressing several key challenges. These include the limited amount of biomass obtained from organisms collected from the environment or grown in laboratory cultures, insufficient quantities of isolated compounds for extensive studies, and suboptimal physicochemical or biological properties of bioactive compounds. Additional obstacles include difficulties in securing long-term funding for basic research and limited interest from the industrial sector in developing novel bioproducts.
Today, modern scientific tools offer powerful solutions to these early-stage challenges. Artificial intelligence (AI)-based technologies and comprehensive natural product databases are playing an increasingly important role in accelerating the discovery pipeline. However, to date, these tools have been used to a limited extent in studies of biotechnological potential of Baltic microorganisms. Meanwhile, by applying suitable AI algorithms, it is now possible to conduct high-throughput screening of large datasets, enabling rapid identification of compounds with properties optimised for specific applications. These tools also allow in silico prediction of biological activity and pharmacokinetic properties, significantly reducing the time and resources needed in the discovery phase. Furthermore, systems biology approaches enhance our understanding of microbial metabolism, physiology, and the genetic basis of their biosynthetic capabilities. In parallel, advances in synthetic biology and chemical synthesis offer the means to produce natural products with improved characteristics, in quantities sufficient to support both in-depth research and commercialisation. The integration of these modern tools not only streamlines the development of new natural products, but also increases the likelihood of selecting compounds with high application potential. Nevertheless, the ultimate success of such endeavours depends on close collaboration between the scientific community and industry stakeholders. Strengthening these partnerships will be essential to unlocking the full biotechnological value of microorganisms from the Baltic Sea and other ecosystems.
Author contributions
HM-M: Supervision, Conceptualization, Writing – original draft, Writing – review & editing. ŁG: Writing – original draft, Visualization, Writing – review & editing. AW: Writing – review & editing, Writing – original draft, Conceptualization. AB: Writing – original draft, Writing – review & editing. MC: Writing – original draft, Visualization, Writing – review & editing. PD: Writing – review & editing, Writing – original draft. MF: Writing – review & editing. EG: Writing – original draft, Visualization, Writing – review & editing. AJ-K: Writing – review & editing, Writing – original draft, Visualization. A-KK: Conceptualization, Writing – review & editing, Writing – original draft, Visualization. TK: Writing – original draft, Writing – review & editing. MK: Writing – review & editing, Writing – original draft. RK: Writing – review & editing, Writing – original draft, Visualization. EK: Writing – review & editing, Visualization, Writing – original draft. DO: Writing – original draft, Writing – review & editing. WS: Writing – review & editing, Conceptualization, Writing – original draft. AT-S: Writing – original draft, Writing – review & editing. SW: Writing – review & editing. MZ: Writing – review & editing. GW: Conceptualization, Writing – review & editing, Supervision, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The work was a part of the CYANOCRAB project supported by the National Science Centre (Poland) within project grant no. 2022/45/B/NZ9/02024, and the project S-A-UEI-23-9 supported by the Lithuanian Science Council and the Ministry of Education, Science and Sports of the Republic of Lithuania.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1682611/full#supplementary-material
Footnotes
1. ^National Center for Biotechnology Information. https://ncbi.nlm.nih.gov/ (accessed 28 February 2025).
2. ^National Center for Biotechnology Information – Genome Datasets. https://ncbi.nlm.nih.gov/datasets/genome/ (accessed 28 February 2025).
3. ^The Bacterial Diversity Database BacDive. https://bacdive.dsmz.de/ (accessed 28 February 2025).
4. ^National Center for Biotechnology Information – Cyanobacteriota/Melainabacteria Genomes. https://ncbi.nlm.nih.gov/datasets/genome/#!/prokaryotes/ (accessed 28 February 2025).
5. ^National Center for Biotechnology Information – Cyanobacteriota/Melainabacteria Genomes. https://ncbi.nlm.nih.gov/datasets/genome/#!/prokaryotes/ (accessed 28 February 2025).
6. ^National Center for Biotechnology Information – Genome list. https://ncbi.nlm.nih.gov/ (accessed 28 February 2025).
7. ^The convention on biological diversity. https://cbd.int/doc/legal/cbd-en.pdf (accessed 6 June 2023).
8. ^United Nations convention on the law of the sea. https://un.org/depts/los/convention_agreements/texts/unclos/unclos_e.pdf (accessed 6 June 2023).
9. ^The Nagoya Protocol on access to genetic resources and fair and equitable sharing of benefits arising from their utilization. https://www.cbd.int/abs/doc/protocol/nagoya-protocol-en.pdf (accessed 6 June 2023).
10. ^Access and Benefit Sharing Clearing House. https://absch.cbd.int/en/ (accessed 6 June 2023).
11. ^United Nations rules for access, use, and fair sharing of benefits from marine biological diversity of areas beyond national jurisdiction. https://www.un.org/bbnj (accessed 1 October 2025).
12. ^Proposal for a Directive of the European Parliament and of the Council on the conservation and sustainable use of marine biological diversity of areas beyond national jurisdiction (Document 52025PC0173). https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=celex:52025PC0173 (accessed 1 October 2025).
13. ^Access and Benefit Sharing Clearing House. https://absch.cbd.int/en/ (accessed 6 June 2023).
14. ^European Commission Decision on the collections recognition. https://environment.ec.europa.eu/topics/nature-and-biodiversity/sharing-natures-genetic-resources_en (accessed 6 June 2023).
References
Abdel-Lateff, A., Klemke, C., König, G. M., and Wright, A. D. (2003). Two new xanthone derivatives from the algicolous marine fungus Wardomyces anomalus. J. Nat. Prod. 66, 706–708. doi: 10.1021/np020518b
Ahmed, M. N., Wahlsten, M., Jokela, J., Nees, M., Stenman, U. H., Alvarenga, D. O., et al. (2021). Potent inhibitor of human trypsins from the aeruginosin family of natural products. ACS Chem. Biol. 16, 2537–2546. doi: 10.1021/acschembio.1c00611
Allahverdiyeva, Y., Leino, H., Saari, L., Fewer, D. P., Shunmugam, S., Sivonen, K., et al. (2010). Screening for biohydrogen production by cyanobacteria isolated from the Baltic Sea and Finnish lakes. Int. J. Hydrog. Energy 35, 1117–1127. doi: 10.1016/j.ijhydene.2009.12.030
Al-Nasrawi, H. (2012). Biodegradation of crude oil by fungi isolated from Gulf of Mexico. J. Bioremed. Biodegr. 3, 147–152. doi: 10.4172/2155-6199.1000147
Andlar, M., Rezić, T., Marđetko, N., Kracher, D., Ludwig, R., and Šantek, B. (2018). Lignocellulose degradation: an overview of fungi and fungal enzymes involved in lignocellulose degradation. ELS 18, 768–778. doi: 10.1002/elsc.201800039
Aransiola, S. A., Victor-Ekwebelem, M. O., Daza, B. X., Oladoye, P. O., Alli, Y. A., Bamisaye, A., et al. (2025). Micro- and nano-plastics pollution in the marine environment: progresses, drawbacks and future guidelines. Chemosphere 374:144211. doi: 10.1016/j.chemosphere.2025.144211
Assunção, J., Guedes, A. C., and Malcata, F. X. (2017). Biotechnological and pharmacological applications of biotoxins and other bioactive molecules from dinoflagellates. Mar. Drugs 15:393. doi: 10.3390/md15120393
Barzkar, N., Babich, O., Das, R., Sukhikh, S., Tamadoni Jahromi, S., and Sohail, M. (2022b). Marine bacterial dextranases: fundamentals and applications. Molecules 27:5533. doi: 10.3390/molecules27175533
Barzkar, N., Rungsardthong, V., Tamadoni Jahromi, S., Laraib, Q., Das, R., Babich, O., et al. (2023). A recent update on fucoidonase: source, isolation methods and its enzymatic activity. Front. Mar. Sci. 10:1129982. doi: 10.3389/fmars.2023.1129982
Barzkar, N., Sheng, R., Sohail, M., Jahromi, S. T., Babich, O., Sukhikh, S., et al. (2022a). Alginate lyases from marine bacteria: an enzyme ocean for sustainable future. Molecules 27:3375. doi: 10.3390/molecules27113375
Barzkar, N., Sukhikh, S., and Babich, O. (2024). Study of marine microorganism metabolites: new resources for bioactive natural products. Front. Microbiol. 14:1285902. doi: 10.3389/fmicb.2023.1285902
Baunach, M., Guljamow, A., Miguel-Gordo, M., and Dittmann, E. (2024). Harnessing the potential: advances in cyanobacterial natural product research and biotechnology. Nat. Prod. Rep. 41, 347–369. doi: 10.1039/d3np00045a
Bayu, A., Rachman, A., Noerdjito, D. R., Putra, M. Y., and Widayatno, W. B. (2020). High-value chemicals from marine diatoms: a biorefinery approach. IOP Conf. Ser. Earth Environ. Sci. 460:012012. doi: 10.1088/1755-1315/460/1/012012
Biran, A., Yagur-Kroll, S., Pedahzur, R., Buchinger, S., Reifferscheid, G., Ben-Yoav, H., et al. (2010). Bacterial genotoxicity bioreporters. Microb. Biotechnol. 3, 412–427. doi: 10.1111/j.1751-7915.2009.00160.x
Björdal, C. G. (2012). Evaluation of microbial degradation of shipwrecks in the Baltic Sea. Int. Biodeterior. Biodegrad. 70, 126–140. doi: 10.1016/j.ibiod.2012.01.012
Bonthond, G., Shalygin, S., Bayer, T., and Weinberger, F. (2021). Draft genome and description of Waterburya agarophytonicola gen. Nov. sp. nov. (Pleurocapsales, Cyanobacteria): a seaweed symbiont. Antonie Van Leeuwenhoek 114, 2189–2203. doi: 10.1007/s10482-021-01672-x
Bringmann, G., Gulder, T. A., Lang, G., Schmitt, S., Stöhr, R., Wiese, J., et al. (2007). Large-scale biotechnological production of the antileukemic marine natural product sorbicillactone A. Mar. Drugs 5, 23–30. doi: 10.3390/md502023
Brzozowska, W., Sprynskyy, M., Wojtczak, I., Dąbek, P., Markuszewski, M. J., Witkowski, A., et al. (2022). Metabolically doping of 3D diatomaceous biosilica with titanium. Materials 15:5210. doi: 10.3390/ma15155210
Bui, T. H., Wray, V., Nimtz, M., Fossen, T., Preisitsch, M., Schröder, G., et al. (2014). Balticidins A-D, antifungal hassallidin-like lipopeptides from the Baltic Sea cyanobacterium Anabaena cylindrica Bio33. J. Nat. Prod. 77, 1287–1296. doi: 10.1021/np401020a
Bull, A. T., Ward, A. C., and Goodfellow, M. (2000). Search and discovery strategies for biotechnology: the paradigm shift. Microbiol. Mol. Biol. Rev. 64, 573–606. doi: 10.1128/MMBR.64.3.573-606.2000
Cakmak, E. K., Chen, C., Cuartero, M., and Cetecioglu, Z. (2024). How to develop a bio-based phosphorus mining strategy for eutrophic marine sediments: unlocking native microbial processes for anaerobic phosphorus release. Chemosphere 358:142188. doi: 10.1016/j.chemosphere.2024.142188
Camacho, F. G., Rodríguez, J. G., Mirón, A. S., García, M. C., Belarbi, E. H., Chisti, Y., et al. (2007). Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 25, 176–194. doi: 10.1016/j.biotechadv.2006.11.008
CBD (2022). CBD/COP/DEC/15/9: Digital sequence information on genetic resources. Decision adopted by the conference of the parties to the convention on biological diversity.
Cegłowska, M., Szubert, K., Grygier, B., Lenart, M., Plewka, J., Milewska, A., et al. (2022). Pseudanabaena galeata CCNP1313 - biological activity and peptides production. Toxins. 14:330. doi: 10.3390/toxins14050330
Cheć, E., Podgórska, B., and Wegrzyn, G. (2006). Direct addition of cultures of tester bacteria into semi-permeable membrane devices (SPMDs) as a modified procedure for preliminary detection of mutagenic pollution of the marine environment by use of microbiological mutagenicity assays. Mutat. Res. 611, 17–24. doi: 10.1016/j.mrgentox.2006.06.031
Chen, J., Jia, Y., Sun, Y., Liu, K., Zhou, C., Liu, C., et al. (2024). Global marine microbial diversity and its potential in bioprospecting. Nature. 633:371379. doi: 10.1038/s41586-024-07891-2
Cheregi, O., Engelbrektsson, J., Andersson, M. X., Strömberg, N., Ekendahl, S., Godhe, A., et al. (2021). Marine microalgae for outdoor biomass production - a laboratory study simulating seasonal light and temperature for the west coast of Sweden. Physiol. Plant. 173, 543–554. doi: 10.1111/ppl.13412
Cho, K., Heo, J., Han, J., Hong, H. D., Jeon, H., Hwang, H. J., et al. (2020). Industrial applications of dinoflagellate phycotoxins based on their modes of action: a review. Toxin 12:805. doi: 10.3390/toxins12120805
Cox, T. E., Cebrian, J., Tabor, M., West, L., and Krause, J. W. (2020). Do diatoms dominate benthic production in shallow systems? L&O 5, 425–434. doi: 10.1002/lol2.10167
Cruz, P. G., Norte, M., Creus, A. H., Fernández, J. J., and Daranas, A. H. (2013). Self-association of okadaic acid: structural and pharmacological significance. Mar. Drugs 11, 1866–1877. doi: 10.3390/md11061866
Cybulska, K., Łońska, E., and Fabisiak, J. (2020). Bacterial benthic community composition in the Baltic Sea in selected chemical and conventional weapons dump sites affected by munition corrosion. Sci. Total Environ. 709:136112. doi: 10.1016/j.scitotenv.2019.136112
Czyz, A., Jasiecki, J., Bogdan, A., Szpilewska, H., and Wegrzyn, G. (2000). Genetically modified Vibrio harveyi strains as potential bioindicators of mutagenic pollution of marine environments. Appl. Environ. Microbiol. 66, 599–605. doi: 10.1128/AEM.66.2.599-605.2000
Czyz, A., Kowalska, W., and Wegrzyn, G. (2003). Vibrio harveyi mutagenicity assay as a preliminary test for detection of mutagenic pollution of marine water. Bull. Environ. Contam. Toxicol. 70, 1065–1070. doi: 10.1007/s00128-003-0091-4
Czyz, A., Szpilewska, H., Dutkiewicz, R., Kowalska, W., Biniewska-Godlewska, A., and Wegrzyn, G. (2002). Comparison of the Ames test and a newly developed assay for detection of mutagenic pollution of marine environments. Mutat. Res. 519, 67–74. doi: 10.1016/s1383-5718(02)00112-2
Daniel, C., Johanna, J., Krister, S., Lone, M., Inger, D., Tom, W., et al. (2021). Genome-informed approach to identify genetic determinants of Flavobacterium psychrophilum phage susceptibility. Environ. Microbiol. 23, 4185–4199. doi: 10.1111/1462-2920.15593
Dantas, F. G. D. S., Castilho, P. F., Almeida-Apolonio, A. A., Araújo, R. P., and Oliveira, K. M. P. (2020). Mutagenic potential of medicinal plants evaluated by the Ames Salmonella/microsome assay: a systematic review. Mutat. Res. Rev. Mutat. Res. 786:108338. doi: 10.1016/j.mrrev.2020.108338
Demay, J., Bernard, C., Reinhardt, A., and Marie, B. (2019). Natural products from cyanobacteria: focus on beneficial activities. Mar. Drugs 17:320. doi: 10.3390/md17060320
d'Ippolito, G., Sardo, A., Paris, D., Vella, F. M., Adelfi, M. G., Botte, P., et al. (2015). Potential of lipid metabolism in marine diatoms for biofuel production. Biotechnol. Biofuels 8:28. doi: 10.1186/s13068-015-0212-4
Dolganyuk, V., Andreeva, A., Sukhikh, S., Kashirskikh, E., Prosekov, A., Ivanova, S., et al. (2022). Study of the physicochemical and biological properties of the lipid complex of marine microalgae isolated from the coastal areas of the eastern water area of the Baltic Sea. Molecules 27:5871. doi: 10.3390/molecules27185871
Dreher, T. W., Davis, E. W., and Mueller, R. S. (2021). Complete genomes derived by directly sequencing freshwater bloom populations emphasize the significance of the genus level ADA clade within the Nostocales. Harmful Algae 103:102005. doi: 10.1016/j.hal.2021.102005
Edlund, A., and Jansson, J. K. (2008). Use of bromodeoxyuridine immunocapture to identify psychrotolerant phenanthrene-degrading bacteria in phenanthrene-enriched polluted Baltic Sea sediments. FEMS Microbiol. Ecol. 65, 513–525. doi: 10.1111/j.1574-6941.2008.00513.x
Elsebai, M. F., Ghabbour, H. A., and Mehiri, M. (2016). Unusual nitrogenous phenalenone derivatives from the marine-derived fungus Coniothyrium cereale. Molecules 21:178. doi: 10.3390/molecules21020178
Elsebai, M. F., Kehraus, S., Lindequist, U., Sasse, F., Shaaban, S., Gütschow, M., et al. (2011). Antimicrobial phenalenone derivatives from the marine-derived fungus Coniothyrium cereale. Org. Biomol. Chem. 9, 802–808. doi: 10.1039/c0ob00625d
Elsebai, M. F., Nazir, M., Kehraus, S., Egereva, E., Ioset, K. N., Marcourt, L., et al. (2012). Polyketide skeletons from the marine alga-derived fungus Coniothyrium cereale. Eur JOC 31, 6197–6203. doi: 10.1002/ejoc.201200700
Epstein-Barash, H., Shichor, I., Kwon, A. H., Hall, S., Lawlor, M. W., Langer, R., et al. (2009). Prolonged duration local anesthesia with minimal toxicity. Proc. Natl. Acad. Sci. USA 106, 7125–7130. doi: 10.1073/pnas.0900598106
Eronen-Rasimus, E. L., Näkki, P. P., and Kaartokallio, H. P. (2022). Degradation rates and bacterial community compositions vary among commonly used bioplastic materials in a brackish marine environment. Environ. Sci. Technol. 56, 15760–15769. doi: 10.1021/acs.est.2c06280
EU (2014) 2014/283/EU: Council decision of 14 April 2014 on the conclusion, on behalf of the European Union, of the Nagoya protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilization to the convention on biological diversity text with EEA relevance, ELI. OJEU. L150/231.
EU (2015) Commission implementing regulation (EU) 2015/1866 of 13 October 2015 laying down detailed rules for the implementation of regulation (EU) no 511/2014 of the European Parliament and of the council as regards the register of collections, monitoring user compliance and best practices. OJEU. L275/4.
EU ABS (2014) EU ABS regulation-regulation (EU) no 511/2014 of the European parliament and of the council of 16 April 2014 on compliance measures for users from the Nagoya protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilization in the union. OJEU. L150/59.
Fan, B., Dewapriya, P., Li, F., Blümel, M., and Tasdemir, D. (2020a). Pyrenosetins a-c, new decalinoylspirotetramic acid derivatives isolated by bioactivity-based molecular networking from the seaweed-derived fungus Pyrenochaetopsis sp. fve-001. Mar. Drugs 18:47. doi: 10.3390/md18010047
Fan, B., Dewapriya, P., Li, F., Grauso, L., Blümel, M., Mangoni, A., et al. (2020b). Pyrenosetin D, a new pentacyclic decalinoyltetramic acid derivative from the algicolous fungus Pyrenochaetopsis sp. FVE-087. Mar. Drugs 18:281. doi: 10.3390/md18060281
Fan, B., Grauso, L., Li, F., Scarpato, S., Mangoni, A., and Tasdemir, D. (2022). Application of feature-based molecular networking for comparative metabolomics and targeted isolation of stereoisomers from algicolous fungi. Mar. Drugs 20:210. doi: 10.3390/md20030210
Felczykowska, A., Pawlik, A., Mazur-Marzec, H., Toruńska-Sitarz, A., Narajczyk, M., Richert, M., et al. (2015). Selective inhibition of cancer cells' proliferation by compounds included in extracts from Baltic Sea cyanobacteria. Toxicon 108, 1–10. doi: 10.1016/j.toxicon.2015.09.030
Ferrer, M., Méndez-García, C., Bargiela, R., Chow, J., Alonso, S., García-Moyano, A., et al. (2019). Decoding the ocean's microbiological secrets for marine enzyme biodiscovery. FEMS Microbiol. Lett. 366:fny285. doi: 10.1093/femsle/fny285
Fewer, D. P., Jokela, J., Heinilä, L., Aesoy, R., Sivonen, K., Galica, T., et al. (2021). Chemical diversity and cellular effects of antifungal cyclic lipopeptides from cyanobacteria. Physiol. Plant. 173, 639–650. doi: 10.1111/ppl.13484
Fewer, D. P., Jokela, J., Rouhiainen, L., Wahlsten, M., Koskenniemi, K., Stal, L. J., et al. (2009). The non-ribosomal assembly and frequent occurrence of the protease inhibitors spumigins in the bloom-forming cyanobacterium Nodularia spumigena. Mol. Microbiol. 73, 924–937. doi: 10.1111/j.1365-2958.2009.06816.x
Flores-Holguín, N., Salas-Leiva, J. S., Núñez-Vázquez, E. J., Tovar-Ramírez, D., and Glossman-Mitnik, D. (2024). Marine toxins as pharmaceutical treasure troves: a focus on saxitoxin derivatives from a computational point of view. Molecules 29:275. doi: 10.3390/molecules29010275
Franckx, E. (2021). Maritime delimitation in the Baltic Sea: an update. TransNav: international journal on marine navigation and safety of sea. Transportation 15, 885–891. doi: 10.12716/1001.15.04.22
Fujii, K., Sivonen, K., Adachi, K., Noguchi, K., Shimizu, Y., Sano, H., et al. (1997). Comparative study of toxic and non-toxic cyanobacterial products: a novel glycoside, suomilide, from non-toxic Nodularia spumigena HKVV. Tetrahedron Lett. 38, 5529–5532. doi: 10.1016/S0040-4039(97)01193-3
Garibay, R. A., Gomez-Godinez, L. J., Gomez, J. M. R., and Zaldivar-Lopez, H. A. (2024) The role of microbial culture collections in the development of microbial inoculants New insights, trends, and challenges in the development and applications of microbial inoculants in agriculture S. Los Santos VillalobosDe 131–141 Academic Press London, UK
Gilmour, D. J. (2019). Microalgae for biofuel production. Adv. Appl. Microbiol. 109, 1–30. doi: 10.1016/bs.aambs.2019.10.001
Golubeva, A., Roychoudhury, P., Dąbek, P., Pryshchepa, O., Pomastowski, P., Pałczyńska, J., et al. (2023). Removal of the basic and diazo dyes from aqueous solution by the frustules of Halamphora cf. salinicola (Bacillariophyta). Mar. Drugs 21:312. doi: 10.3390/md21050312
Grabowski, Ł., Wiśniewska, K., Żabińska, M., Konarzewska, M., Zielenkiewicz, M., Rintz, E., et al. (2024). Cyanobacteria and their metabolites - can they be helpful in the fight against pathogenic microbes? Blue Biotechnol. 1:4. doi: 10.1186/s44315-024-00003-9
Gregson, J. M., Chen, J.-L., Patterson, G. M. L., and Moore, R. E. (1992). Structures of puwainaphycins A-E. Tetrahedron 48, 3727–3734. doi: 10.1016/S0040-4020(01)92264-1
Guidance Document (2021) Guidance document on the scope of application and core obligations of regulation (EU) no 511/2014 of the European Parliament and of the council on the compliance measures for users from the Nagoya protocol on access to genetic resources and the fair and equitable sharing of benefits arising from their utilisation in the union 2021/C 13/01. OJEU. C13/1.
Häggqvist, K., Toruńska-Sitarz, A., Błaszczyk, A., Mazur-Marzec, H., and Meriluoto, J. (2016). Morphologic, phylogenetic and chemical characterization of a brackish colonial picocyanobacterium (Coelosphaeriaceae) with bioactive properties. Toxins 8:108. doi: 10.3390/toxins8040108
Hájek, J., Bieringer, S., Voráčová, K., Macho, M., Saurav, K., Delawská, K., et al. (2021). Semi-synthetic puwainaphycin/minutissamide cyclic lipopeptides with improved antifungal activity and limited cytotoxicity. RSC Adv. 11, 30873–30886. doi: 10.1039/d1ra04882a
Hårdeman, F., and Sjöling, S. (2007). Metagenomic approach for the isolation of a novel low-temperature-active lipase from uncultured bacteria of marine sediment. FEMS Microbiol. Ecol. 59, 524–534. doi: 10.1111/j.1574-6941.2006.00206.x
Harish, B. S., and Uppuluri, K. B. (2018). Microbial serine protease inhibitors and their therapeutic applications. Int. J. Biol. Macromol. 107, 1373–1387. doi: 10.1016/j.ijbiomac.2017.09.115
Hassett, B. T., Vonnahme, T. R., Peng, X., Jones, E. G., and Heuzé, C. (2020). Global diversity and geography of planktonic marine fungi. Bot. Mar. 63, 121–139. doi: 10.1515/bot-2018-0113
Hawrot-Paw, M., and Ratomski, P. (2024). Efficient production of microalgal biomass-step by step to industrial scale. Energies 17:944. doi: 10.3390/en17040944
Heinaru, E., Naanuri, E., Grünbach, M., Jõesaar, M., and Heinaru, A. (2016). Functional redundancy in phenol and toluene degradation in Pseudomonas stutzeri strains isolated from the Baltic Sea. Gene 589, 90–98. doi: 10.1016/j.gene.2016.05.022
Heindl, H., Thiel, V., Wiese, J., and Imhoff, J. F. (2012). Bacterial isolates from the bryozoan Membranipora membranacea: influence of culture media on isolation and antimicrobial activity. Int. Microbiol. 15, 17–32. doi: 10.2436/20.1501.01.155
Heindl, H., Wiese, J., Thiel, V., and Imhoff, J. F. (2010). Phylogenetic diversity and antimicrobial activities of bryozoan-associated bacteria isolated from Mediterranean and Baltic Sea habitats. Syst. Appl. Microbiol. 33, 94–104. doi: 10.1016/j.syapm.2009.12.002
Heinilä, L. M. P., Jokela, J., Ahmed, M. N., Wahlsten, M., Kumar, S., Hrouzek, P., et al. (2022). Discovery of varlaxins, new aeruginosin-type inhibitors of human trypsins. Org. Biomol. Chem. 20, 2681–2692. doi: 10.1039/d1ob02454j
Hildebrand, M., Davis, A. K., Smith, S. R., Traller, J., and Abbriano, R. M. (2012). The place of diatoms in the biofuels industry. Biofuels 3, 221–240. doi: 10.4155/bfs.11.157
Hoetzinger, M., Nilsson, E., Arabi, R., Osbeck, C. M. G., Pontiller, B., Hutinet, G., et al. (2021). Dynamics of Baltic Sea phages driven by environmental changes. Environ. Microbiol. 23, 4576–4594. doi: 10.1111/1462-2920.15651
Holmfeldt, K., Dziallas, C., Titelman, J., Pohlmann, K., Grossart, H. P., and Riemann, L. (2009). Diversity and abundance of freshwater Actinobacteria along environmental gradients in the brackish northern Baltic Sea. Environ. Microbiol. 11, 2042–2054. doi: 10.1111/j.1462-2920.2009.01925.x
Holmfeldt, K., Middelboe, M., Nybroe, O., and Riemann, L. (2007). Large variabilities in host strain susceptibility and phage host range govern interactions between lytic marine phages and their Flavobacterium hosts. Appl. Environ. Microbiol. 73, 6730–6739. doi: 10.1128/AEM.01399-07
Hrouzek, P., Kuzma, M., Černý, J., Novák, P., Fišer, R., Simek, P., et al. (2012). The cyanobacterial cyclic lipopeptides puwainaphycins F/G are inducing necrosis via cell membrane permeabilization and subsequent unusual actin relocalization. Chem. Res. Toxicol. 25, 1203–1211. doi: 10.1021/tx300044t
Humisto, A., Jokela, J., Teigen, K., Wahlsten, M., Permi, P., Sivonen, K., et al. (2019). Characterization of the interaction of the antifungal and cytotoxic cyclic glycolipopeptide hassallidin with sterol-containing lipid membranes. Biochim. Biophys. Acta Biomembr. 1861, 1510–1521. doi: 10.1016/j.bbamem.2019.03.010
Jagannathan, S. V., Manemann, E. M., Rowe, S. E., Callender, M. C., and Soto, W. (2021). Marine actinomycetes, new sources of botechnological products. Mar. Drugs 19:365. doi: 10.3390/md19070365
Jakubowska-Deredas, M., Jurczak-Kurek, A., Richert, M., Łoś, M., Narajczyk, M., and Wróbel, B. (2012). Diversity of tailed phages in Baltic Sea sediment: large number of siphoviruses with extremely long tails. Res. Microbiol. 163, 292–296. doi: 10.1016/j.resmic.2012.02.002
Jämsä, M., Lynch, F., Santana-Sánchez, A., Laaksonen, P., Zaitsev, G., Solovchenko, A., et al. (2017). Nutrient removal and biodiesel feedstock potential of green alga UHCC00027 grown in municipal wastewater under Nordic conditions. Algal Res. 26, 65–73. doi: 10.1016/j.algal.2017.06.019
Jiang, Y., Cao, Y., Wang, Q., and Jin, R. (2011). Diversity of cultured actinomycete in the Baltic Sea. Wei Sheng Wu Xue Bao 51, 1461–1467.
Jiang, M., Chen, S., Li, J., and Liu, L. (2020). The biological and chemical diversity of tetramic acid compounds from marine-derived microorganisms. Mar. Drugs 18:114. doi: 10.3390/md18020114
Jokela, J., Oftedal, L., Herfindal, L., Permi, P., Wahlsten, M., Døskeland, S. O., et al. (2012). Anabaenolysins, novel cytolytic lipopeptides from benthic Anabaena cyanobacteria. PLoS One 7:e41222. doi: 10.1371/journal.pone.0041222
Jones, M. R., Pinto, E., Torres, M. A., Dörr, F., Mazur-Marzec, H., Szubert, K., et al. (2021). CyanoMetDB, a comprehensive public database of secondary metabolites from cyanobacteria. Water Res. 196:117017. doi: 10.1016/j.watres.2021.117017
Jores, J., Appel, B., and Lewin, A. (2003). Cloning and molecular characterization of a unique hemolysin gene of Vibrio pommerensis sp. nov.: development of a DNA probe for the detection of the hemolysin gene and its use in identification of related Vibrio spp. from the Baltic Sea. FEMS Microbiol. Lett. 229, 223–229. doi: 10.1016/S0378-1097(03)00843-7
Kallscheuer, N., Jogler, M., Wiegand, S., Peeters, S. H., Heuer, A., Boedeker, C., et al. (2020). Three novel Rubripirellula species isolated from plastic particles submerged in the Baltic Sea and the estuary of the river Warnow in northern Germany. Antonie Van Leeuwenhoek 113, 1767–1778. doi: 10.1007/s10482-019-01368-3
Kamat, P. K., Rai, S., and Nath, C. (2013). Okadaic acid induced neurotoxicity: an emerging tool to study Alzheimer's disease pathology. Neurotoxicology 37, 163–172. doi: 10.1016/j.neuro.2013.05.002
Karjalainen, M., Engström-Ost, J., Korpinen, S., Peltonen, H., Pääkkönen, J. P., Rönkkönen, S., et al. (2007). Ecosystem consequences of cyanobacteria in the northern Baltic Sea. Ambio 36, 195–202. doi: 10.1579/0044-7447(2007)36[195:ecocit]2.0.co;2
Karlson, B., Andersen, P., Arneborg, L., Cembella, A., Eikrem, W., John, U., et al. (2021). Harmful algal blooms and their effects in coastal seas of northern Europe. Harmful Algae 102:101989. doi: 10.1016/j.hal.2021.101989
Kaur, J., Chaudhary, A., Kaur, R., and Arora, S. (2014). Assessment of mutagenic, genotoxic, and cytotoxic potential of water samples of Harike wetland: a Ramsar site in India using different ex vivo biological systems. Ecotoxicology 23, 967–977. doi: 10.1007/s10646-014-1240-8
Kettner, M. T., Rojas-Jimenez, K., Oberbeckmann, S., Labrenz, M., and Grossart, H. P. (2017). Microplastics alter composition of fungal communities in aquatic ecosystems. Environ. Microbiol. 19, 4447–4459. doi: 10.1111/1462-2920.13891
Khetkorn, W., Rastogi, R. P., Incharoensakdi, A., Lindblad, P., Madamwar, D., Pandey, A., et al. (2017). Microalgal hydrogen production - a review. Bioresour. Technol. 243, 1194–1206. doi: 10.1016/j.biortech.2017.07.085
Klin, M., Pniewski, F., and Latała, A. (2020). Growth phase-dependent biochemical composition of green microalgae: theoretical considerations for biogas production. Bioresour. Technol. 303:122875. doi: 10.1016/j.biortech.2020.122875
Klin, M., Pniewski, F., and Latała, A. (2023). Multistage optimization of growth and physiological condition of brackish green microalgae with the use of natural waters. Aquaculture 562:738820. doi: 10.1016/j.aquaculture.2022.738820
Klinzing, K., Søndergaard, I. A., Chirom, T., Whitwell, J., Bisini, L., Marabottini, C., et al. (2024). In vitro genotoxicological evaluation of protein-rich powder derived from Xanthobacter sp. SoF1. J. Appl. Toxicol. 44, 1347–1360. doi: 10.1002/jat.4621
Kohane, D. S., Lu, N. T., Gökgöl-Kline, A. C., Shubina, M., Kuang, Y., Hall, S., et al. (2000). The local anesthetic properties and toxicity of saxitonin homologues for rat sciatic nerve block in vivo. Reg. Anesth. Pain Med. 25, 52–59. doi: 10.1016/s1098-7339(00)80011-5
Kolařík, M., Spakowicz, D. J., Gazis, R., Shaw, J., Kubátová, A., Nováková, A., et al. (2017). Biatriospora (Ascomycota: Pleosporales) is an ecologically diverse genus including facultative marine fungi and endophytes with biotechnological potential. Plant Syst. Evol. 303, 35–50. doi: 10.1007/s00606-016-1350-2
Konkel, R., Cegłowska, M., Szubert, K., Wieczerzak, E., Iliakopoulou, S., Kaloudis, T., et al. (2023a). Structural diversity and biological activity of cyanopeptolins produced by Nostoc edaphicum CCNP1411. Mar. Drugs 21:508. doi: 10.3390/md21100508
Konkel, R., Grabski, M., Cegłowska, M., Wieczerzak, E., Węgrzyn, G., and Mazur-Marzec, H. (2022). Anabaenopeptins from Nostoc edaphicum CCNP1411. Int. J. Environ. Res. Public Health 19:12346. doi: 10.3390/ijerph191912346
Konkel, R., Milewska, A., Do, N. D. T., Barreto Duran, E., Szczepanski, A., Plewka, J., et al. (2023b). Anti-SARS-CoV-2 activity of cyanopeptolins produced by Nostoc edaphicum CCNP1411. Antivir. Res. 219:105731. doi: 10.1016/j.antiviral.2023.105731
Krohn, K., Dai, J. Q., Flörke, U., Aust, H. J., Dräger, S., and Schulz, B. (2005). Botryane metabolites from the fungus Geniculosporium sp. isolated from the marine red alga Polysiphonia. J. Nat. Prod. 68, 400–405. doi: 10.1021/np0498206
Kumar, D. M. (2018). The Nagoya protocol: legal protections for genetic resources and ramifications for aquatic science. L&O Bull. 27, 31–35. doi: 10.1002/lob.10235
Kunz, A. L., Labes, A., Wiese, J., Bruhn, T., Bringmann, G., and Imhoff, J. F. (2014). Nature's lab for derivatization: new and revised structures of a variety of streptophenazines produced by a sponge-derived Streptomyces strain. Mar. Drugs 12, 1699–1714. doi: 10.3390/md12041699
Kurtzman, C. P., and Labeda, D. P. (2009). “Type culture collections and their databases” in Encyclopaedia of microbiology. ed. M. Schaechter. 3rd ed (Oxford, UK: Elsevier), 306–312.
Kuzikova, I., Andronov, E., Zaytseva, T., Metelkova, L., Zhakovskaya, Z., and Medvedeva, N. (2022). A microcosm approach for evaluating the microbial nonylphenol and butyltin biodegradation and bacterial community shifts in co-contaminated bottom sediments from the Gulf of Finland, the Baltic Sea. Environ. Sci. Pollut. Res. Int. 29, 69849–69860. doi: 10.1007/s11356-022-20751-8
Lee, T. C., Fong, F. L., Ho, K. C., and Lee, F. W. (2016). The mechanism of diarrhetic shellfish poisoning toxin production in Prorocentrum spp.: physiological and molecular perspectives. Toxins 8:272. doi: 10.3390/toxins8100272
Leyland, B., Boussiba, S., and Khozin-Goldberg, I. (2020). A review of diatom lipid droplets. Biol. 9:38. doi: 10.3390/biology9020038
Lin, S. (2024). A decade of dinoflagellate genomics illuminating an enigmatic eukaryote cell. BMC Genomics 25:932. doi: 10.1186/s12864-024-10847-5
Lindehoff, E., Mattsson, L., Olofsson, M., Svensson, F., Farnelid, H., and Legrand, C. (2024). Biomass performance and stability of 5-year outdoor microalgal cultivation for CO2 removal from cement flue gas. Bioresour. Technol. Rep. 25:101730. doi: 10.1016/j.biteb.2023.101730
Lindim, C., van Gils, J., Cousins, I. T., Kühne, R., Georgieva, D., Kutsarova, S., et al. (2017). Model-predicted occurrence of multiple pharmaceuticals in Swedish surface waters and their flushing to the Baltic Sea. Environ. Pollut. 223, 595–604. doi: 10.1016/j.envpol.2017.01.062
Liu, L., Budnjo, A., Jokela, J., Haug, B. E., Fewer, D. P., Wahlsten, M., et al. (2015). Pseudoaeruginosins, nonribosomal peptides in Nodularia spumigena. ACS Chem. Biol. 10, 725–733. doi: 10.1021/cb5004306
Liu, J., Zhang, M., Huang, Z., Fang, J., Wang, Z., Zhou, C., et al. (2023). Diversity, biosynthesis and bioactivity of aeruginosins, a family of cyanobacteria-derived nonribosomal linear tetrapeptides. Mar. Drugs 21:217. doi: 10.3390/md21040217
López, A. M., Rodríguez, J. J., Mirón, A. S., Camacho, F. G., and Grima, E. M. (2011). Immunoregulatory potential of marine algal toxins yessotoxin and okadaic acid in mouse T lymphocyte cell line EL-4. Toxicol. Lett. 207, 167–172. doi: 10.1016/j.toxlet.2011.09.007
Luesch, H., Ellis, E. K., Chen, Q. Y., and Ratnayake, R. (2025). Progress in the discovery and development of anticancer agents from marine cyanobacteria. Nat. Prod. Rep. 42, 208–256. doi: 10.1039/d4np00019f
Luhtanen, A.-M., Eronen-Rasimus, E., Kaartokallio, H., Rintala, J.-M., Autio, R., and Roine, E. (2014). Isolation and characterization of phage-host systems from the Baltic Sea ice. Extremophiles 18, 121–130. doi: 10.1007/s00792-013-0604-y
Lynch, F., Santana-Sánchez, A., Jämsä, M., Sivonen, K., Aro, E.-M., and Allahverdiyeva, Y. (2015). Screening native isolates of cyanobacteria and a green alga for integrated wastewater treatment, biomass accumulation and neutral lipid production. Algal Res. 11, 411–420. doi: 10.1016/j.algal.2015.05.015
Mal, N., Srivastava, K., Sharma, Y., Singh, M., Rao, K. M., Enamala, M. K., et al. (2021). Facets of diatom biology and their potential applications. Biomass Convers. Biorefinery 12, 1959–1975. doi: 10.1007/s13399-020-01155-5
Martins, J., Cruz, D., and Vasconcelos, V. (2020). The Nagoya protocol and its implications on the EU Atlantic area countries. J Mar Sci Eng 8:92. doi: 10.3390/jmse8020092
Marzec, M., Dąbek, P., Witkowski, A., Monedeiro, F., Pomastowski, P., Buszewski, B., et al. (2022). Lipid constituents of diatoms (Halamphora) as components for production of lipid nanoparticles. Pharmaceutics 14:1171. doi: 10.3390/pharmaceutics14061171
Mattsson, L., Lindehoff, E., Olofsson, M., and Legrand, C. (2019). Boosting algal lipids: diurnal shifts in temperature exceed the effects of nitrogen limitation. Eng. Rep. 1:e212067. doi: 10.1002/eng2.12067
Mazur-Marzec, H., Andersson, A. F., Błaszczyk, A., Dąbek, P., Górecka, E., Grabski, M., et al. (2024). Biodiversity of microorganisms in the Baltic Sea: the power of novel methods in the identification of marine microbes. FEMS Microbiol. Rev. 48:fuae024. doi: 10.1093/femsre/fuae024
Mazur-Marzec, H., Bertos-Fortis, M., Toruńska-Sitarz, A., Fidor, A., and Legrand, C. (2016). Chemical and genetic diversity of Nodularia spumigena from the Baltic Sea. Mar. Drugs 14:209. doi: 10.3390/md14110209
Mazur-Marzec, H., Błaszczyk, A., Felczykowska, A., Hohlfeld, N., Kobos, J., Toruńska-Sitarz, A., et al. (2015). Baltic cyanobacteria - a source of biologically active compounds. Eur. J. Phycol. 50, 343–360. doi: 10.1080/09670262.2015.1062563
Mazur-Marzec, H., Fidor, A., Cegłowska, M., Wieczerzak, E., Kropidłowska, M., Goua, M., et al. (2018). Cyanopeptolins with trypsin and chymotrypsin inhibitory activity from the cyanobacterium Nostoc edaphicum CCNP1411. Mar. Drugs 16:220. doi: 10.3390/md16070220
Mazur-Marzec, H., Kaczkowska, M. J., Blaszczyk, A., Akcaalan, R., Spoof, L., and Meriluoto, J. (2012). Diversity of peptides produced by Nodularia spumigena from various geographical regions. Mar. Drugs 11, 1–19. doi: 10.3390/md11010001
Miettinen, H., Bomberg, M., Nyyssönen, M., Reunamo, A., Jørgensen, K. S., and Vikman, M. (2019). Oil degradation potential of microbial communities in water and sediment of Baltic Sea coastal area. PLoS One 14:e0218834. doi: 10.1371/journal.pone.0218834
Mishra, M., Arukha, A. P., Bashir, T., Yadav, D., and Prasad, G. B. K. S. (2017). All new faces of diatoms: potential source of nanomaterials and beyond. Front. Microbiol. 8:1239. doi: 10.3389/fmicb.2017.01239
Monteiro, P. R., do Amaral, S. C., Siqueira, A. S., Xavier, L. P., and Santos, A. V. (2021). Anabaenopeptins: what we know so far. Toxins 13:522. doi: 10.3390/toxins13080522
Multisanti, C. R., Ferrara, S., Piccione, G., and Faggio, C. (2025). Plastics and their derivatives are impacting animal ecophysiology: A review. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 291:110149. doi: 10.1016/j.cbpc.2025.110149
Neuhof, T., Schmieder, P., Preussel, K., Dieckmann, R., Pham, H., Bartl, F., et al. (2005). Hassallidin A, a glycosylated lipopeptide with antifungal activity from the cyanobacterium Hassallia sp. J. Nat. Prod. 68, 695–700. doi: 10.1021/np049671r
Neumann, K., Kehraus, S., Gütschow, M., and König, G. M. (2009). Cytotoxic and HLE-inhibitory tetramic acid derivatives from marine-derived fungi. Nat. Prod. Commun. 4:1934578X0900400308. doi: 10.1177/1934578X0900400308
Ngamcharungchit, C., Chaimusik, N., Panbangred, W., Euanorasetr, J., and Intra, B. (2023). Bioactive metabolites from terrestrial and marine actinomycetes. Molecules 28:5915. doi: 10.3390/molecules28155915
Nieri, P., Carpi, S., Esposito, R., Costantini, M., and Zupo, V. (2023). Bioactive molecules from marine diatoms and their value for the nutraceutical industry. Nutrients 15:464. doi: 10.3390/nu15020464
Nilsson, E., Bayfield, O. W., Lundin, D., Antson, A. A., and Holmfeldt, K. (2020). Diversity and host interactions among virulent and temperate Baltic Sea Flavobacterium Phages. Viruses 12:158. doi: 10.3390/v12020158
Nilsson, E., Li, K., Fridlund, J., Šulčius, S., Bunse, C., Karlsson, C. M. G., et al. (2019). Genomic and seasonal variations among aquatic phages infecting the Baltic Sea Gammaproteobacterium Rheinheimera sp. strain BAL341. Appl. Environ. Microbiol. 85:e01003-19. doi: 10.1128/AEM.01003-19
Nowak, A. P., Sprynskyy, M., Wojtczak, I., Trzciński, K., Wysocka, J., Szkoda, M., et al. (2020). Diatoms biomass as a joint source of biosilica and carbon for lithium-ion battery anodes. Materials 13:1673. doi: 10.3390/ma13071673
Oftedal, L., Myhren, L., Jokela, J., Gausdal, G., Sivonen, K., Døskeland, S. O., et al. (2012). The lipopeptide toxins anabaenolysin A and B target biological membranes in a cholesterol-dependent manner. Biochim. Biophys. Acta 1818, 3000–3009. doi: 10.1016/j.bbamem.2012.07.015
Ojaveer, H., Jaanus, A., Mackenzie, B. R., Martin, G., Olenin, S., Radziejewska, T., et al. (2010). Status of biodiversity in the Baltic Sea. PLoS One 5:e12467. doi: 10.1371/journal.pone.0012467
Olofsson, M., Lindehoff, E., Frick, B., Svensson, F., and Legrand, C. (2015). Baltic Sea microalgae transform cement flue gas into valuable biomass. Algal Res. 11, 227–233. doi: 10.1016/j.algal.2015.07.001
Oppong-Danquah, E., Blümel, M., Scarpato, S., Mangoni, A., and Tasdemir, D. (2022). Induction of isochromanones by co-cultivation of the marine fungus Cosmospora sp. and the phytopathogen Magnaporthe oryzae. Int. J. Mol. Sci. 23:782. doi: 10.3390/ijms23020782
Oppong-Danquah, E., Blümel, M., and Tasdemir, D. (2023). Metabolomics and microbiomics insights into differential surface fouling of three macroalgal species of fucus (Fucales, Phaeophyceae) that co-exist in the German Baltic Sea. Mar. Drugs 21:595. doi: 10.3390/md21110595
Oppong-Danquah, E., Budnicka, P., Blümel, M., and Tasdemir, D. (2020a). Design of fungal co-cultivation based on comparative metabolomics and bioactivity for discovery of marine fungal agrochemicals. Mar. Drugs 18:73. doi: 10.3390/md18020073
Oppong-Danquah, E., Parrot, D., Blümel, M., Labes, A., and Tasdemir, D. (2018). Molecular networking-based metabolome and bioactivity analyses of marine-adapted fungi co-cultivated with phytopathogens. Front. Microbiol. 9:2072. doi: 10.3389/fmicb.2018.02072
Oppong-Danquah, E., Passaretti, C., Chianese, O., Blümel, M., and Tasdemir, D. (2020b). Mining the metabolome and the agricultural and pharmaceutical potential of sea foam-derived fungi. Mar. Drugs 18:128. doi: 10.3390/md18020128
Österholm, J., Popin, R. V., Fewer, D. P., and Sivonen, K. (2020). Phylogenomic analysis of secondary metabolism in the toxic cyanobacterial genera Anabaena, Dolichospermum and Aphanizomenon. Toxins (Basel) 12:248. doi: 10.3390/toxins12040248
Overlingė, D., Cegłowska, M., Konkel, R., and Mazur-Marzec, H. (2024b). Aeruginosin 525 (AER525) from cyanobacterium Aphanizomenon Sp. (KUCC C2): a new serine proteases inhibitor. Mar. Drugs 22:506. doi: 10.3390/md22110506
Overlingė, D., Toruńska-Sitarz, A., Cegłowska, M., Szubert, K., and Mazur-Marzec, H. (2024a). Phylogenetic and molecular characteristics of two Aphanizomenon strains from the Curonian lagoon, southeastern Baltic Sea and their biological activities. Sci. Rep. 14:24686. doi: 10.1038/s41598-024-76064-y
Overlingė, D., Toruńska-Sitarz, A., Kataržytė, M., Pilkaitytė, R., Gyraitė, G., and Mazur-Marzec, H. (2021). Characterization and diversity of microcystins produced by cyanobacteria from the Curonian lagoon (SE Baltic Sea). Toxins. 13:838. doi: 10.3390/toxins13120838
Panchal, R. G., Smart, M. L., Bowser, D. N., Williams, D. A., and Petrou, S. (2002). Pore-forming proteins and their application in biotechnology. Curr. Pharm. Biotechnol. 3, 99–115. doi: 10.2174/1389201023378418
Pärnänen, K., Karkman, A., Virta, M., Eronen-Rasimus, E., and Kaartokallio, H. (2015). Discovery of bacterial polyhydroxyalkanoate synthase (PhaC)-encoding genes from seasonal Baltic Sea ice and cold estuarine waters. Extremophiles 19, 197–206. doi: 10.1007/s00792-014-0699-9
Patel, S. (2017). A critical review on serine protease: key immune manipulator and pathology mediator. Allergol Immunopathol (Madr) 45, 579–591. doi: 10.1016/j.aller.2016.10.011
Paulauskiene, T., Uebe, J., Kryzevicius, Z., Katarzyte, M., Overlingė, D., and Shevchenko, L. (2023). Sorption and removal of petroleum hydrocarbons from brackish water by hydrophobic sorbents immobilized with fungi. J Mar Sci Eng 11:1283. doi: 10.3390/jmse11071283
Pechsiri, J. S., and Gröndahl, F. (2022). Assessing energy return on investment for harvest of wild Nodularia spumigena during blooms in the Baltic Sea. J. Ind. Ecol. 26, 1979–1991. doi: 10.1111/jiec.13170
Pesic, A., Baumann, H. I., Kleinschmidt, K., Ensle, P., Wiese, J., Süssmuth, R. D., et al. (2013). Champacyclin, a new cyclic octapeptide from Streptomyces strain C42 isolated from the Baltic Sea. Mar. Drugs 11, 4834–4857. doi: 10.3390/md11124834
Petersen, L. E., Marner, M., Labes, A., and Tasdemir, D. (2019). Rapid metabolome and bioactivity profiling of fungi associated with the leaf and rhizosphere of the Baltic seagrass Zostera marina. Mar. Drugs 17:419. doi: 10.3390/md17070419
Pham, T. M., Wiese, J., Wenzel-Storjohann, A., and Imhoff, J. F. (2016). Diversity and antimicrobial potential of bacterial isolates associated with the soft coral Alcyonium digitatum from the Baltic Sea. Antonie Van Leeuwenhoek 109, 105–119. doi: 10.1007/s10482-015-0613-1
Plöhn, M., Spain, O., Sirin, S., Silva, M., Escudero-Oñate, C., Ferrando-Climent, L., et al. (2021). Wastewater treatment by microalgae. Physiol. Plant. 173, 568–578. doi: 10.1111/ppl.13427
Podgórska, B., Królicka, A., Lojkowska, E., and Wegrzyn, G. (2008). Rapid detection of mutagens accumulated in plant tissues using a novel Vibrio harveyi mutagenicity assay. Ecotoxicol. Environ. Saf. 70, 231–235. doi: 10.1016/j.ecoenv.2007.04.004
Podgórska, B., Pazdro, K., Pempkowiak, J., and Wegrzyn, G. (2007a). The use of a novel Vibrio harveyi luminescence mutagenicity assay in testing marine water for the presence of mutagenic pollution. Mar. Pollut. Bull. 54, 808–814. doi: 10.1016/j.marpolbul.2006.12.015
Podgórska, B., Pazdro, K., and Wegrzyn, G. (2007b). The use of the Vibrio harveyi luminescence mutagenicity assay as a rapid test for preliminary assessment of mutagenic pollution of marine sediments. J. Appl. Genet. 48, 409–412. doi: 10.1007/BF03195241
Podgórska, B., and Wegrzyn, G. (2006). A modified Vibrio harveyi mutagenicity assay based on bioluminescence induction. Lett. Appl. Microbiol. 42, 578–582. doi: 10.1111/j.1472-765X.2006.01891.x
Polyak, Y. M., Demchuk, A. S., Sharov, A. N., Gubelit, Y. I., and Berezina, N. A. (2020). Hydrocarbon-oxidizing bacteria in the digestive system of fish as an indicator of coastal pollution. Dokl. Biol. Sci. 491, 71–74. doi: 10.1134/S001249662002009X
Qasim, M. S., Lampi, M., Heinonen, M. K., Garrido-Zabala, B., Bamford, D. H., Käkelä, R., et al. (2021). Cold-active Shewanella glacialimarina TZS-4T nov. features a temperature-dependent fatty acid profile and putative sialic acid metabolism. Front. Microbiol. 12:737641. doi: 10.3389/fmicb.2021.737641
Ratomski, P., and Hawrot-Paw, M. (2021). Influence of nutrient-stress conditions on Chlorella vulgaris biomass production and lipid content. Catalysts 11:573. doi: 10.3390/catal11050573
Ravi, K., García-Hidalgo, J., Nöbel, M., Gorwa-Grauslund, M. F., and Lidén, G. (2018). Biological conversion of aromatic monolignol compounds by a Pseudomonas isolate from sediments of the Baltic Sea. AMB Express 8:32. doi: 10.1186/s13568-018-0563-x
Reunamo, A., Riemann, L., Leskinen, P., and Jørgensen, K. S. (2013). Dominant petroleum hydrocarbon-degrading bacteria in the Archipelago Sea in south-West Finland (Baltic Sea) belong to different taxonomic groups than hydrocarbon degraders in the oceans. Mar. Pollut. Bull. 72, 174–180. doi: 10.1016/j.marpolbul.2013.04.006
Rodríguez-Gijón, A., Buck, M., Andersson, A. F., Izabel-Shen, D., Nascimento, F. J. A., and Garcia, S. L. (2023). Linking prokaryotic genome size variation to metabolic potential and environment. ISME Commun 3:25. doi: 10.1038/s43705-023-00231-x
Rotter, A., Bacu, A., Barbier, M., Bertoni, F., Bones, A. M., Cancela, M. L., et al. (2020). A new network for the advancement of marine biotechnology in Europe and beyond. Front. Mar. Sci. 7:278. doi: 10.3389/fmars.2020.00278
Rouhiainen, L., Jokela, J., Fewer, D. P., Urmann, M., and Sivonen, K. (2010). Two alternative starter modules for the non-ribosomal biosynthesis of specific anabaenopeptin variants in Anabaena (Cyanobacteria). Chem. Biol. 17, 265–273. doi: 10.1016/j.chembiol.2010.01.017
Rounge, T. B., Rohrlack, T., Tooming-Klunderud, A., Kristensen, T., and Jakobsen, K. S. (2007). Comparison of cyanopeptolin genes in Planktothrix, Microcystis, and Anabaena strains: evidence for independent evolution within each genus. Appl. Environ. Microbiol. 73, 7322–7330. doi: 10.1128/AEM.01475-07
Roychoudhury, P., Bose, R., Dąbek, P., and Witkowski, A. (2022a). Photonic nano−/microstructured diatom based biosilica in metal modification and removal - a review. Materials 15:6597. doi: 10.3390/ma15196597
Roychoudhury, P., Golubeva, A., Dąbek, P., Gloc, M., Dobrucka, R., Kurzydłowski, K., et al. (2021). Diatom mediated production of fluorescent flower shaped silver-silica nanohybrid. Materials 14:7284. doi: 10.3390/ma14237284
Roychoudhury, P., Golubeva, A., Dąbek, P., Pryshchepa, O., Sagandykova, G., Pomastowski, P., et al. (2022b). Study on biogenic spindle-shaped iron-oxide nanoparticles by Pseudostaurosira trainorii in field of laser desorption/ionization applications. Int. J. Mol. Sci. 23:11713. doi: 10.3390/ijms231911713
Rueff, J., Rodrigues, A. S., and Kranendonk, M. (2019). A personally guided tour on some of our data with the Ames assay - a tribute to Professor Bruce Ames. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 846:503094. doi: 10.1016/j.mrgentox.2019.503094
Ruiz, Y., Suárez, P., Alonso, A., Longo, E., and San, F. J. (2013). Mutagenicity test using Vibrio harveyi in the assessment of water quality from mussel farms. Water Res. 47, 2742–2756. doi: 10.1016/j.watres.2013.02.031
Salgado, P., Riobó, P., Rodríguez, F., Franco, J. M., and Bravo, I. (2015). Differences in the toxin profiles of Alexandrium ostenfeldii (Dinophyceae) strains isolated from different geographic origins: evidence of paralytic toxin, spirolide, and gymnodimine. Toxicon 103, 85–98. doi: 10.1016/j.toxicon.2015.06.015
Sananes, A., Cohen, I., Allon, I., Ben-David, O., Abu Shareb, R., Yegodayev, K. M., et al. (2023). Serine protease inhibitors decrease metastasis in prostate, breast, and ovarian cancers. Mol. Oncol. 17, 2337–2355. doi: 10.1002/1878-0261.13513
Sang, Y., Xiong, G., and Maser, E. (2012). Identification of a new steroid degrading bacterial strain H5 from the Baltic Sea and isolation of two estradiol inducible genes. J. Steroid Biochem. Mol. Biol. 129, 22–30. doi: 10.1016/j.jsbmb.2011.01.018
Saoud, H. A. A., Sprynskyy, M., Pashaei, R., Kawalec, M., Pomastowski, P., and Buszewski, B. (2022). Diatom biosilica: source, physical-chemical characterization, modification, and application. J. Sep. Sci. 45, 3362–3376. doi: 10.1002/jssc.202100981
Saurav, K., Caso, A., Urajová, P., Hrouzek, P., Esposito, G., Delawská, K., et al. (2022). Fatty acid substitutions modulate the cytotoxicity of puwainaphycins/minutissamides isolated from the Baltic Sea cyanobacterium Nodularia harveyana UHCC-0300. ACS Omega 7, 11818–11828. doi: 10.1021/acsomega.1c07160
Sayanova, O., Mimouni, V., Ulmann, L., Morant-Manceau, A., Pasquet, V., Schoefs, B., et al. (2017). Modulation of lipid biosynthesis by stress in diatoms. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 372:20160407. doi: 10.1098/rstb.2016.0407
Sayers, E. W., Beck, J., Bolton, E. E., Brister, J. R., Chan, J., Connor, R., et al. (2025). Database resources of the National Center for biotechnology information in 2025. Nucleic Acids Res. 53, D20–D29. doi: 10.1093/nar/gkae979
Sazonova, O. I., Ivanova, A. A., Delegan, Y. A., Streletskii, R. A., Vershinina, D. D., Sokolov, S. L., et al. (2023). Characterization and genomic analysis of the naphthalene-degrading Delftia tsuruhatensis ULwDis3 isolated from seawater. Microorganisms 11:1092. doi: 10.3390/microorganisms11041092
Scales, B. S., Cable, R. N., Duhaime, M. B., Gerdts, G., Fischer, F., Fischer, D., et al. (2021). Cross-hemisphere study reveals geographically ubiquitous, plastic-specific bacteria emerging from the rare and unexplored biosphere. mSphere 6:e0085120. doi: 10.1128/mSphere.00851-20
Schneemann, I., Nagel, K., Kajahn, I., Labes, A., Wiese, J., and Imhoff, J. F. (2010). Comprehensive investigation of marine Actinobacteria associated with the sponge Halichondria panicea. Appl. Environ. Microbiol. 76, 3702–3714. doi: 10.1128/AEM.00780-10
Schneider, X. T., Stroil, B. K., Tourapi, C., Rebours, C., Gaudêncio, S. P., Novoveska, L., et al. (2022). Responsible research and innovation framework, the Nagoya protocol and other european blue biotechnology strategies and regulations: gaps analysis and recommendations for increased knowledge in the marine biotechnology community. Mar. Drugs 20:290. doi: 10.3390/md20050290
Schober, I., Koblitz, J., Sardà Carbasse, J., Ebeling, C., Schmidt, M. L., Podstawka, A., et al. (2025). BacDive in 2025: the core database for prokaryotic strain data. Nucleic Acids Res. 53, D748–D756. doi: 10.1093/nar/gkae959
Schreuder, H., Liesum, A., Lönze, P., Stump, H., Hoffmann, H., Schiell, M., et al. (2016). Isolation, co-crystallization and structure-based characterization of anabaenopeptins as highly potent inhibitors of activated thrombin activatable fibrinolysis inhibitor (TAFIa). Sci. Rep. 6:32958. doi: 10.1038/srep32958
Schwenk, D., Seppälä, J., Spilling, K., Virkki, A., Tamminen, T., Oksman-Caldentey, K. M., et al. (2013). Lipid content in 19 brackish and marine microalgae: influence of growth phase, salinity and temperature. Aquat. Ecol. 47, 415–424. doi: 10.1007/s10452-013-9454-z
Senčilo, A., Luhtanen, A.-M., Saarijärvi, M., Bamford, D. H., and Roine, E. (2015). Cold-active bacteriophages from the Baltic Sea ice have diverse genomes and virus-host interactions: comparative genomics of the sea-ice phages. Environ. Microbiol. 17, 3628–3641. doi: 10.1111/1462-2920.12611
Sharma, N., Simon, D., Diaz-Garza, A., Fantino, E., Messaabi, A., Meddeb-Mouelhi, F., et al. (2021). Diatoms biotechnology: various industrial applications for a greener tomorrow. Front. Mar. Sci. 8:636613. doi: 10.3389/fmars.2021.636613
Shen, H., Li, Z., Jiang, Y., Pan, X., Wu, J., Cristofori-Armstrong, B., et al. (2018). Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 362:eaau2596. doi: 10.1126/science.aau2596
Shishido, T. K., Humisto, A., Jokela, J., Liu, L., Wahlsten, M., Tamrakar, A., et al. (2015). Antifungal compounds from cyanobacteria. Mar. Drugs 13, 2124–2140. doi: 10.3390/md13042124
Shushni, M. A. M., Azam, F., and Lindequist, U. (2013). Oxasetin from Lophiostoma sp. of the Baltic Sea: identification, in silico binding mode prediction and antibacterial evaluation against fish pathogenic bacteria. Nat. Prod. Commun. 8, 1223–1226. doi: 10.1177/1934578X130080090
Shushni, M. A. M., Mentel, R., Lindequist, U., and Jansen, R. (2009). Balticols A-F, new naphthalenone derivatives with antiviral activity, from an ascomycetous fungus. Chem. Biodivers. 6, 127–137. doi: 10.1002/cbdv.200800150
Shushni, M. A. M., Singh, R., Mentel, R., and Lindequist, U. (2011). Balticolid: a new 12-membered macrolide with antiviral activity from an ascomycetous fungus of marine origin. Mar. Drugs 9, 844–851. doi: 10.3390/md9050844
Sierra, M. A., and Martínez-Álvarez, R. (2020). Ricin and saxitoxin: two natural products that became chemical weapons. J. Chem. Educ. 97, 1707–1714. doi: 10.1021/acs.jchemed.9b00841
Singh, K., Gupta, J. K., Pathak, D., and Kumar, S. (2023). The use of enzyme inhibitors in drug discovery: current strategies and future prospects. Curr. Enzym. Inhib. 19, 157–166. doi: 10.2174/1573408019666230731113105
Singha, T. R., Singh, S., Chahal, G. S., Vamsi, G. K., and Goutam, U. (2022). Production of biofuel using diatoms: an overview. BFAIJ 14, 1194–1200.
Spilling, K. (2011). Optimizing lipid production by planktonic algae: LIPIDO. End of project report 2008–2010. Nordic Energy Research, 1–62. Available online at: https://www.nordicenergy.org/wp-content/uploads/2012/01/Lipido-Final-report.pdf.
Spilling, K., Seppälä, J., Schwenk, D., Rischer, H., and Tamminen, T. (2021). Variation in the fatty acid profiles of two cold water diatoms grown under different temperature, light, and nutrient regime. J. Appl. Phycol. 33, 1447–1455. doi: 10.1007/s10811-021-02380-9
Spoof, L., Błaszczyk, A., Meriluoto, J., Cegłowska, M., and Mazur-Marzec, H. (2015). Structures and activity of new anabaenopeptins produced by Baltic Sea cyanobacteria. Mar. Drugs 14:8. doi: 10.3390/md14010008
Sprynskyy, M., Szczyglewska, P., Wojtczak, I., Nowak, I., Witkowski, A., Buszewski, B., et al. (2021). Diatom biosilica doped with palladium(II) chloride nanoparticles as new efficient photocatalysts for methyl orange degradation. Int. J. Mol. Sci. 22:6734. doi: 10.3390/ijms22136734
Stante, M., Weiland-Bräuer, N., Repnik, U., Werner, A., Bramkamp, M., Chibani, C. M., et al. (2024). Four novel Caudoviricetes bacteriophages isolated from Baltic Sea water iinfect colonizers of Aurelia aurita. Viruses 15:1525. doi: 10.3390/v15071525
Steinhoff, F. S., Karlberg, M., Graeve, M., and Wulff, A. (2014). Cyanobacteria in Scandinavian coastal waters - a potential source for biofuels and fatty acids? Algal Res. 5, 42–51. doi: 10.1016/j.algal.2014.05.005
Stevens, M., Peigneur, S., and Tytgat, J. (2011). Neurotoxins and their binding areas on voltage-gated sodium channels. Front. Pharmacol. 2:71. doi: 10.3389/fphar.2011.00071
Sudhakar, D. G. S., Reddy, C. V. R., and Alapati, S. R. (2019). An efficient approach for the total synthesis of balticolid. TETL 60:151027. doi: 10.1016/j.tetlet.2019.151027
Šulčius, S., and Holmfeldt, K. (2016). Viruses of microorganisms in the Baltic Sea: current state of research and perspectives. Mar. Biol. Res. 12, 115–124. doi: 10.1080/17451000.2015.1118514
Šulčius, S., Mazur-Marzec, H., Vitonytė, I., Kvederavičiūtė, K., Kuznecova, J., Šimoliūnas, E., et al. (2018). Insights into cyanophage-mediated dynamics of nodularin and other non-ribosomal peptides in Nodularia spumigena. Harmful Algae 78, 69–74. doi: 10.1016/j.hal.2018.07.004
Šulčius, S., Šimoliūnas, E., Alzbutas, G., Gasiūnas, G., Jauniškis, V., Kuznecova, J., et al. (2019). Genomic characterization of cyanophage vB_AphaS-CL131 infecting filamentous diazotrophic cyanobacterium Aphanizomenon flos-aquae reveals novel insights into virus-bacterium interactions. Appl. Environ. Microbiol. 85, e01311–e01318. doi: 10.1128/AEM.01311-18
Šulčius, S., Šimoliūnas, E., Staniulis, J., Koreivienė, J., Baltrušis, P., Meškys, R., et al. (2015). Characterization of a lytic cyanophage that infects the bloom-forming cyanobacterium Aphanizomenon flos-aquae. FEMS Microbiol. Ecol. 91, 1–7. doi: 10.1093/femsec/fiu012
Szubert, K., Wiglusz, M., and Mazur-Marzec, H. (2018). Bioactive metabolites produced by Spirulina subsalsa from the Baltic Sea. Oceanologia 60, 245–255. doi: 10.1016/j.oceano.2017.11.003
Tasdemir, D., Scarpato, S., Utermann-Thüsing, C., Jensen, T., Blümel, M., Wenzel-Storjohann, A., et al. (2024). Epiphytic and endophytic microbiome of the seagrass Zostera marina: do they contribute to pathogen reduction in seawater? Sci. Total Environ. 908:168422. doi: 10.1016/j.scitotenv.2023.168422
Teikari, J. E., Hou, S., Wahlsten, M., Hess, W. R., and Sivonen, K. (2018). Comparative genomics of the Baltic Sea toxic cyanobacteria Nodularia spumigena UHCC 0039 and its response to varying salinity. Front. Microbiol. 9:356. doi: 10.3389/fmicb.2018.00356
Teikari, J. E., Popin, R. V., Hou, S., Wahlsten, M., Hess, W. R., and Sivonen, K. (2019). Insight into the genome and brackish water adaptation strategies of toxic and bloom-forming Baltic Sea Dolichospermum sp. UHCC 0315. Sci. Rep. 9:4888. doi: 10.1038/s41598-019-40883-1
Tibell, S., Tibell, L., Pang, K. L., Calabon, M., and Jones, E. G. (2020). Marine fungi of the Baltic Sea. Mycology 11, 195–213. doi: 10.1080/21501203.2020.1729886
Utermann, C., Echelmeyer, V. A., Blümel, M., and Tasdemir, D. (2020a). Culture-dependent microbiome of the Ciona intestinalis tunic: isolation, bioactivity profiling and untargeted metabolomics. Microorganisms 8:1732. doi: 10.3390/microorganisms8111732
Utermann, C., Echelmeyer, V. A., Oppong-Danquah, E., Blümel, M., and Tasdemir, D. (2020b). Diversity, bioactivity profiling and untargeted metabolomics of the cultivable gut microbiota of Ciona intestinalis. Mar. Drugs 19:6. doi: 10.3390/md19010006
Valdiglesias, V., Laffon, B., Pásaro, E., Cemeli, E., Anderson, D., and Méndez, J. (2011). Induction of oxidative DNA damage by the marine toxin okadaic acid depends on human cell type. Toxicon 57, 882–888. doi: 10.1016/j.toxicon.2011.03.005
Valdiglesias, V., Prego-Faraldo, M. V., Pásaro, E., Méndez, J., and Laffon, B. (2013). Okadaic acid: more than a diarrheic toxin. Mar. Drugs 11, 4328–4349. doi: 10.3390/md11114328
Vašíček, O., Hájek, J., Bláhová, L., Hrouzek, P., Babica, P., Kubala, L., et al. (2020). Cyanobacterial lipopeptides puwainaphycins and minutissamides induce disruptive and pro-inflammatory processes in Caco-2 human intestinal barrier model. Harmful Algae 96:101849. doi: 10.1016/j.hal.2020.101849
Vedler, E., Heinaru, E., Jutkina, J., Viggor, S., Koressaar, T., Remm, M., et al. (2013). Limnobacter spp. as newly detected phenol-degraders among Baltic Sea surface water bacteria characterised by comparative analysis of catabolic genes. Syst. Appl. Microbiol. 36, 525–532. doi: 10.1016/j.syapm.2013.07.004
Vestola, J., Shishido, T. K., Jokela, J., Fewer, D. P., Aitio, O., Permi, P., et al. (2014). Hassallidins, antifungal glycolipopeptides, are widespread among cyanobacteria and are the end-product of a nonribosomal pathway. Proc. Natl. Acad. Sci. USA 111, E1909–E1917. doi: 10.1073/pnas.1320913111
Viggor, S., Jõesaar, M., Vedler, E., Kiiker, R., Pärnpuu, L., and Heinaru, A. (2015). Occurrence of diverse alkane hydroxylase alkB genes in indigenous oil-degrading bacteria of Baltic Sea surface water. Mar. Pollut. Bull. 101, 507–516. doi: 10.1016/j.marpolbul.2015.10.064
Viggor, S., Juhanson, J., Jõesaar, M., Mitt, M., Truu, J., Vedler, E., et al. (2013). Dynamic changes in the structure of microbial communities in Baltic Sea coastal seawater microcosms modified by crude oil, shale oil or diesel fuel. Microbiol. Res. 168, 415–427. doi: 10.1016/j.micres.2013.02.006
Vigil-Stenman, T., Ininbergs, K., Bergman, B., and Ekman, M. (2017). High abundance and expression of transposases in bacteria from the Baltic Sea. ISME J. 11, 2611–2623. doi: 10.1038/ismej.2017.114
Voss, B., Bolhuis, H., Fewer, D. P., Kopf, M., Möke, F., Haas, F., et al. (2013). Insights into the physiology and ecology of the brackish-water-adapted cyanobacterium Nodularia spumigena CCY9414 based on a genome-transcriptome analysis. PLoS One 8:e60224. doi: 10.1371/journal.pone.0060224
Wagil, M., Białk-Bielińska, A., Puckowski, A., Wychodnik, K., Maszkowska, J., Mulkiewicz, E., et al. (2015). Toxicity of anthelmintic drugs (fenbendazole and flubendazole) to aquatic organisms. Environ. Sci. Pollut. Res. Int. 22, 2566–2573. doi: 10.1007/s11356-014-3497-0
Wang, Z., Wang, N., Yang, L., and Song, X. Q. (2022). Bioactive natural products in COVID-19 therapy. Front. Pharmacol. 13:926507. doi: 10.3389/fphar.2022.926507
Weckesser, J., Martin, C., and Jakobi, C. (1996). Cyanopeptolins, depsipeptides from cyanobacteria system. Appl. Microbiol. 19, 133–138. doi: 10.1016/S0723-2020(96)80038-5
Wiese, M., D'Agostino, P. M., Mihali, T. K., Moffitt, M. C., and Neilan, B. (2010). Neurotoxic alkaloids: saxitoxin and its analogs. Mar. Drugs 8, 2185–2211. doi: 10.3390/md8072185
Wiese, J., Thiel, V., Nagel, K., Staufenberger, T., and Imhoff, J. F. (2009). Diversity of antibiotic-active bacteria associated with the brown alga Laminaria saccharina from the Baltic Sea. Mar. Biotechnol. (N.Y.) 11, 287–300. doi: 10.1007/s10126-008-9143-4
Wu, B., Wiese, J., Labes, A., Kramer, A., Schmaljohann, R., and Imhoff, J. F. (2015). Lindgomycin, an unusual antibiotic polyketide from a marine fungus of the Lindgomycetaceae. Mar. Drugs 13, 4617–4632. doi: 10.3390/md13084617
Wu, K., Wu, Q. Q., Yan, X. J., Liu, B., Yang, B. H., Zhang, J., et al. (2003). Production of fucoidanase from marine fungus Dendryphiella arenaria TM94 by solid substrate fermentation using marine algae resources. In Marine biotechnology: An overview of leading field (ed. S. Colliec-Jouault and J. Guézennec, Fleurence, J., Y. Le Gal and P. Roy), pp. 122–133. Actes de colloques-IFREMER, Nantes.
Yi, M., Lin, S., Zhang, B., Jin, H., and Ding, L. (2020). Antiviral potential of natural products from marine microbes. Eur. J. Med. Chem. 207:112790. doi: 10.1016/j.ejmech.2020.112790
Yi, Z., Xu, M., Di, X., Brynjólfsson, S., and Fu, W. (2017). Exploring valuable lipids in diatoms. Front. Mar. Sci. 4:17. doi: 10.3389/fmars.2017.00017
Zervou, S. K., Kaloudis, T., Gkelis, S., Hiskia, A., and Mazur-Marzec, H. (2021). Anabaenopeptins from cyanobacteria in freshwater bodies of Greece. Toxins 14:4. doi: 10.3390/toxins14010004
Zhang, T., Xiong, G., and Maser, E. (2011). Characterization of the steroid degrading bacterium S19-1 from the Baltic Sea at Kiel, Germany. Chem. Biol. Interact. 191, 83–88. doi: 10.1016/j.cbi.2010.12.021
Keywords: added value products, bioactivity, microorganisms, cyanobacteria, bacteriophages, microalgae, fungi, blue biotechnology
Citation: Mazur-Marzec H, Grabowski Ł, Węgrzyn A, Błaszczyk A, Cegłowska M, Dąbek P, Farooq M, Górecka E, Jurczak-Kurek A, Kaczorowska A-K, Kaczorowski T, Kataržytė M, Konkel R, Kotlarska E, Overlingė D, Surosz W, Toruńska-Sitarz A, Walat S, Zielenkiewicz M and Węgrzyn G (2025) Unlocking the biotechnological potential of Baltic microorganisms. Front. Microbiol. 16:1682611. doi: 10.3389/fmicb.2025.1682611
Edited by:
Rosa María Martínez-Espinosa, University of Alicante, SpainReviewed by:
Abdul Rauf Shakoori, University of the Punjab, PakistanShangwei Zhang, Beijing Normal University, China
Copyright © 2025 Mazur-Marzec, Grabowski, Węgrzyn, Błaszczyk, Cegłowska, Dąbek, Farooq, Górecka, Jurczak-Kurek, Kaczorowska, Kaczorowski, Kataržytė, Konkel, Kotlarska, Overlingė, Surosz, Toruńska-Sitarz, Walat, Zielenkiewicz and Węgrzyn. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hanna Mazur-Marzec, aGFubmEubWF6dXItbWFyemVjQHVnLmVkdS5wbA==; Marta Cegłowska, bWNlZ2xvd3NrYUBpb3Bhbi5wbA==