
 Abdullah S. Ahmad1
Abdullah S. Ahmad1
- 1Department of Anesthesiology, College of Medicine, University of Florida, Gainesville, FL, USA
- 2Departments of Neurology, Psychiatry, and Neuroscience, College of Medicine, University of Florida, Gainesville, FL, USA
Each year, approximately 795,000 people experience a new or recurrent stroke. Of all strokes, 84% are ischemic, 13% are intracerebral hemorrhage (ICH) strokes, and 3% are subarachnoid hemorrhage strokes. Despite the decreased incidence of ischemic stroke, there has been no change in the incidence of hemorrhagic stroke in the last decade. ICH is a devastating disease 37–38% of patients between the ages of 45 and 64 die within 30 days. In an effort to prevent ischemic and hemorrhagic strokes we and others have been studying the role of prostaglandins and their receptors. Prostaglandins are bioactive lipids derived from the metabolism of arachidonic acid. They sustain homeostatic functions and mediate pathogenic mechanisms, including the inflammatory response. Most prostaglandins are produced from specific enzymes and act upon cells via distinct G-protein coupled receptors. The presence of multiple prostaglandin receptors cross-reactivity and coupling to different signal transduction pathways allow differentiated cells to respond to prostaglandins in a unique manner. Due to the number of prostaglandin receptors, prostaglandin-dependent signaling can function either to promote neuronal survival or injury following acute excitotoxicity, hypoxia, and stress induced by ICH. To better understand the mechanisms of neuronal survival and neurotoxicity mediated by prostaglandin receptors, it is essential to understand downstream signaling. Several groups including ours have discovered unique roles for prostaglandin receptors in rodent models of ischemic stroke, excitotoxicity, and Alzheimer disease, highlighting the emerging role of prostaglandin receptor signaling in hemorrhagic stroke with a focus on cyclic-adenosine monophosphate and calcium (Ca2+) signaling. We review current ICH data and discuss future directions notably on prostaglandin receptors, which may lead to the development of unique therapeutic targets against hemorrhagic stroke and brain injuries alike.
Introduction
Stroke is a leading cause of long-term disability and accounts for one of every 18 deaths in the United States (Roger et al., 2012). More than 795,000 people experience a new or recurrent stroke in the United States each year and 10–15% experience a hemorrhagic stroke (Thom et al., 2006; Roger et al., 2012). Although the incidence of intracerebral hemorrhage (ICH) is lower than that of ischemic stroke, the mortality and disability rates are greater. It is thought that many of the deleterious effects of ICH are due to the release of blood, increased intracranial pressure, and ischemic damage to the surrounding brain tissue (Gong et al., 2001). Current measures to reduce mortality and increase functional recovery include early diagnosis, blood pressure management, hypothermia, and surgical removal of blood and clots (Adeoye and Broderick, 2010; Kollmar et al., 2010). Due to the complexity of ICH, the development of effective interventions has been challenging.
In experimental models of ICH, increased neuronal loss has been correlated with the development of localized collection of blood outside the blood vessels called hematomas (Fernandes et al., 2000). Hematomas are made up of red blood cells, in turn consisting of hemoglobin, a complex metalloprotein containing four heme molecules whose iron atoms temporarily bind oxygen molecules. When released from red blood cells, heme-iron molecules contribute to the expansion of ICH-induced brain damage (Macdonald and Weir, 1991; Hua et al., 2000). Due to the toxic role of red blood cells in ICH, we and others have been determining whether intracranial injection of blood mimics many of the hallmarks of ICH (Wang et al., 2008). Over the past years, we have been interested in the regulation of the pro-oxidant heme by the catalytic heme oxygenase enzymes and the impact of this process on ischemic stroke (Doré et al., 1999; Li et al., 2009; Zeynalov et al., 2009) and ICH (Wang et al., 2006; Wang and Doré, 2007a, 2008). Based on the importance of heme toxicity, we are interested in determining the role of blood components in hemorrhagic stroke (Namiranian et al., 2005; Wang and Doré, 2007b).
Inflammation is the immune system’s response to infection and injury, but if persistent, it can also lead to the loss of cellular and organ function. For example, the inflammatory milieu following a hemorrhagic stroke is integral to the development of secondary injury; important components of this have traditionally included the so-called “pro-inflammatory” prostaglandins (Wang and Doré, 2007b). Despite previous work on experimental ICH by others, the role of prostaglandin receptors in ICH remains to be explored. Prostaglandins are a large family of lipids enzymatically derived from arachidonic acid by the cyclooxygenase enzymes COX-1 and -2. Prostaglandins function by activating corresponding prostaglandin receptors (DP1–2, EP1–4, FP, IP, and TP). The diversity of the receptors allows prostaglandins to act on an array of cells with a wide variety of effects. In this review, we discuss the regulation of prostaglandin receptors through selective pharmacologic ligands or genetic deletion with regard to stroke with special emphasis on ICH.
Pathophysiology and Clinical Features of ICH
Intracerebral hemorrhage is a medical emergency that requires rapid diagnosis and management to minimize neuronal loss and deterioration after ICH. Vomiting, severe headaches, increased systolic blood pressure, and coma are common symptoms of ICH and confirmation the ICH incidence is essentially achieved by neuroimaging techniques (Sarrafzadeh et al., 2003; Goldstein and Simel, 2005).
Vasoconstriction and Blood Pressure
Reversible cerebral vasoconstriction is characterized by severe headaches with or without seizures and focal neurological deficits, and constriction of cerebral arteries following occurs in 7% of ICH patients, and 22% of SAH patients (Ba et al., 2012). Increase in cerebral vasoconstriction directly affects blood pressure. Elevated blood pressure (BP) at admission has been found to predict worse outcomes; early intensive BP management reduces the risk of hematoma expansion (Anderson et al., 2010). Continuous intravenous infusion of BP lowering drugs aggressively reduces systolic BP (Manawadu et al., 2010). Prostaglandin E2 (PGE2), which is synthesized in the vasculature of the brain (within neuron and glia cells), acts as a powerful endogenous pyrogenic mediator of the preoptic area by activating specific receptors (Nakamura et al., 2009). Due to elevated BP, there is an increased risk of hemorrhage in the brain. As blood leaks and collects into a hematoma, pressure builds on nearby brain tissue, reducing vital blood flow, and killing brain cells.
Intracranial Pressure and Edema
Blood components such as serum may be connected to the pathogenesis of secondary brain injury after ICH (Wang et al., 2011). Inflammatory agents released from hematomas can cause the breakdown of blood-brain barrier, which is a proposed mechanism of edema (Wagner et al., 1998). Uncontrolled inflammation contributes to brain edema and prostacyclin PGI2 has been reported to contribute by augmenting the permeability of capillaries (Masada et al., 2001; Xi et al., 2001, 2002; Bentzer and Grande, 2004). Following experimental lipopolysaccharide (LPS)-induced edema, low-dose infusion of PGI2 caused further increase in intracranial pressure (Gardenfors et al., 2004). Recently, in a mouse model of subarachnoid hemorrhage (SAH), cerebral edema was reduced following the inhibition of inducible cyclooxygenase-2 (Ayer et al., 2011). In addition to inflammatory components, iron-mediated free radical damage can also contribute to secondary damage after ICH. For example, iron released from heme after the breakdown of hemoglobin accumulates in the parenchyma and has been linked to cerebral edema; thus, removal of the hematoma reduces edema formation (Wagner et al., 1999; Huang et al., 2002; Qing et al., 2009; Zhao et al., 2011). Due to the significant impact of inflammatory components like prostaglandins following ICH, we propose that prostaglandin receptors may be ideal targets to fight against ICH-induced brain edema.
Inflammation
The major inflammatory cells that are activated and accumulate within the brain after ICH are blood-derived leukocytes, macrophages, and resident microglia. In addition, the infiltration of short-lived neutrophils following ICH contributes to blood vessel disruption, blood-brain barrier degradation, axon damage, and the glia responses that evolve after ICH (Moxon-Emre and Schlichter, 2011). Following ischemic stroke, infiltrating neutrophils play a role in exacerbating inflammation by up-regulating matrix metalloproteinase (MMP; Justicia et al., 2003). For example, a correlation between MMP-9 activity and hemorrhagic transformation of the ischemic lesion has been reported (Heo et al., 1999; Lapchak et al., 2000; Sumii and Lo, 2002; Gautier et al., 2009).
In addition to their effect on inflammation, microglial cells are increasingly being studied as the cell type responsible for the resolution of hematomas formed after ICH (Zhao et al., 2007). Data suggest that microglia activation occurs very early after the onset of ICH and persists for weeks (Hickenbottom et al., 1999; Xue and Del Bigio, 2000). However, uncontrolled microglia activation can also play a role in secondary damage following ICH (Wasserman et al., 2008). In addition to being expressed in neurons, prostaglandin receptors are expressed in microglia, astrocytes, and endothelial cells (Caggiano and Kraig, 1999; Takemiya et al., 2011). Microglia predominately express EP1 and EP2 receptors, reactive astrocytes express EP1 and DP1 receptors and endothelia cells express all four EP (EP1–4) receptors (Mohri et al., 2007; Carlson et al., 2009; Taniguchi et al., 2011a). As well as expressing receptors, microglia and astrocytes are also a source of calcium-mediated PGE2 in the brain (Sanzgiri et al., 1999; Zonta et al., 2003; Anrather et al., 2011; Font-Nieves et al., 2012).
Also, due to the increased permeability of the blood-brain barrier following ICH, blood components, and plasma proteins enter the brain, initiating an exacerbated inflammatory response involving glial activation, release of cytokines, chemokines, and formation of reactive oxygen species, together resulting in the breakdown of brain tissue and repair (Peeling et al., 1998; Mayne et al., 2001a,b). For example, in a rat model of ICH induced by double autologous intrastriatal blood injection, elevated levels of interleukin-1β and tumor necrosis factor-α was observed at 3 and 24 h after injection (Mayne et al., 2001b).
Growing knowledge of the pathophysiology of ICH has led to the exploration of neuroprotective strategies aiming to improve its outcomes through reduction of secondary pathological processes. Work by us and others have previously shown that prostaglandins are important agents released in response to ischemic stroke and their affect is prostaglandin receptor-dependent. Despite some recent data by our group showing that when deleted, EP1 receptor may exacerbate brain injury in the early hours after ICH by regulating microglial phagocytosis, further studies of the role prostaglandin receptors (EP1–4) on the function of cells such as microglia and neurotrophils after ICH is still needed. More information on the general role of various cell types modulating inflammation after ICH can be found in a previous review from our group (Wang and Doré, 2007b).
Seizures
Seizures related to ICH occur in approximately 11% of patients, although those related to ischemic stroke appear in approximately 9% of patients (De Reuck, 2007). Post-stroke seizures are generally classified as early- and late-onset seizures, with most early onset seizures occurring at onset or within the first 24 h of ICH (Takasugi et al., 1985). Early onset seizures and increased risk of epilepsy are associated with hemorrhagic stroke (Burn et al., 1997; Berges et al., 2000; De Herdt et al., 2011). Using animal models of seizure, data suggest that hemolysis and deposition of iron rich compounds may play a significant role (Willmore and Triggs, 1991). It is believed that in iron-induced experimental epilepsy models, seizure activity is mediated by free radicals and membrane lipid peroxidation (Jyoti et al., 2009). The role of pro-inflammatory molecules is also likely to contribute to ICH-related seizures.
In experimental models of kainic acid-induced seizures, COX-2 and PGE2 levels are increased in neuronal and non-neuronal cells and blockade of COX-2 and/or PGE2 reduced cell death (Takemiya et al., 2006). Vascular endothelial cells in the brain produce PGE2 in response to excitotoxicity (Takemiya et al., 2010). When blocked, PGE2 receptors EP1, EP3, and EP4 and activation of EP2 receptors have anticonvulsant and neuroprotective properties in different rodent seizure models (Oliveira et al., 2008; Fischborn et al., 2010; Takemiya et al., 2011; Rehni and Singh, 2012). Nevertheless, in a mouse model of pilocarpine-induced status epilepticus consecutive inhibition of EP2 receptors after termination of seizures with antiepileptic drugs significantly reduced hippocampal neuronal injury (Jiang et al., 2012). Furthermore prostaglandin PGF2α and its FP receptor have been implicated in kainic acid-induced seizures (Kim et al., 2008). Consequently, it is likely that the production of prostaglandins following ICH may contribute to the incidence and severity of the seizures.
Role of Prostaglandin Receptors in ICH
Prostaglandins are generated from arachidonic acid, a 20-carbon unsaturated fatty acid, which is released from the bilipid layers of the plasma membrane by the action of three different phospholipases A2: secreted (sPLA2α), and cytosolic calcium-dependent (sPLA2α) and calcium independent (iPLA2α) and metabolized to PGH2 by cyclooxygenase enzymes, COX-1 and COX-2 (Kishimoto et al., 2010). PGH2 is the common substrate for a series of specific isomerase and synthase enzymes that produce prostaglandins. Prostaglandins PGE2, PGI2, PGD2, PGF2α, and TxA2 are generated by the action of their respective synthases: PGE synthase (cytosolic; cPGES-1, and membrane-associated; mPGES-1 and mPGES-2), prostaglandin I synthase (PGIS), prostaglandin D synthase (hematopoietic-type: H-PGDS, and lipocalin-type: L-PGDS), prostaglandin F synthase (PGFS), and thromboxane synthase (TxS). It should be noted that mPGES-1 was found to be significantly increased in the AD brains as compared to age matched controls (Chaudhry et al., 2008, 2010). Once synthesized, prostaglandins can be further metabolized or they can act on their specific G-protein coupled receptors.
Prostaglandin Receptor Expression (Message and Protein)
Elegant work was done to measure the respective affinity of the various radioactive-labeled prostaglandin ligands to their respective receptors by expressing each of the receptors in CHO cells and performing traditional ligand binding assays (Kiriyama et al., 1997). Furthermore, specific binding of PGD2, PGE2, and PGF2α were found in rat and human brain tissue (Watanabe et al., 1985). In postmortem human brains of normal subjects, [3H]-PGD2 and [3H]-PGE2 bindings were highest in the hypothalamus, amygdala, and hippocampus followed by cerebellar nuclei and the thalamus. In addition [3H]-PGD2 was found in nucleus accumbens and cerebral cortex. [3H]-PGF2α binding was most abundant in the amygdala, cingulate cortex, cerebellar medulla, hippocampus, nucleus accumbens, midbrain, and hypothalamus (Watanabe et al., 1985). Similar regional distribution of PGE2 binding sites were detected in the rat brain (Matsumura et al., 1992).
PGE2-EP Receptor Expression
PGE2-EP2 receptors are expressed throughout the PNS and CNS, for example, at the mRNA level, the EP1 receptor is mainly expressed in the thalamus and hypothalamus, and have been studied in neurons and microglia under different pathologic conditions (Batshake et al., 1995; Caggiano and Kraig, 1999; Kawano et al., 2006). EP2 receptor mRNA are expressed mostly in astrocytes and neurons of the cerebral cortex, striatum, hippocampus, and CA1 pyramidal neurons (Zhang and Rivest, 1999). The mRNA expressions of EP3 receptor are most abundant in the olfactory system, hippocampus, and subcortical telencephalic structures in the septal region and amygdala of the brain (Sugimoto et al., 1994; Ek et al., 2000; Nakamura et al., 2000). EP3 receptor mRNA are expressed mostly in subcortical-hypothalamic regions of the brain owing to their role in thermoregulation (Vasilache et al., 2007). The EP3 receptor has multiple splice variants differing in their C-terminal tails. In human tissue, nine mRNA and eight isoforms (EP3I, EP3II, EP3III, EP3IV, EP3V, EP3IV, EP3e, and EP3f) have been described (Kotani et al., 1995, 1997; Schmid et al., 1995). EP3 receptor variants are mostly expressed in clusters of multiple isoforms; for example, the human uterus expresses mRNA for EP3V and EP3VI receptor isoforms, whereas in primary keratinocytes, EP3I, EP3II, and EP3IV splice variants are expressed (Kotani et al., 2000). Similar to humans, mice also have EP3 receptor alternative splicing that are well characterized EP3 isoforms (α, β, and Γ) and contain carboxyl tails of 30, 26, and 29 amino acids. EP4 receptor mRNA are expressed in the forebrain, hypothalamus, lower brainstem, and at lower levels in endothelial cells (Narumiya et al., 1999; Zhang and Rivest, 1999; Li et al., 2008). Messenger RNA of the EP1–4 receptors was detected both in the hippocampus and in the neocortex. All four immunoreactive EP receptors appeared to be detected in neurons and were also present in astrocytes, though perhaps at weaker levels (Zhu et al., 2005).
IP, DP, FP, and TP Receptor Expression
IP receptor mRNA is expressed in many tissue types including spleen, thymus, aorta, coronary, pulmonary, and cerebral arteries, kidney, and in neuronal cell bodies (Oida et al., 1995). DP1 receptor mRNA is expressed in the cerebral cortex, hippocampal pyramidal layers, dentate gyrus, thalamus, choroid plexus, and leptomeninges (Oida et al., 1997). The mRNA expression of the FP receptor has been previously demonstrated in mouse brains and in brain synaptosomes of newborn pigs and in human eye tissue (Li et al., 1993; Kitanaka et al., 1994; Liang et al., 2008). FP receptor mRNA is highly expressed in rodent whole brain (www.brain-map.org; Kitanaka et al., 1994). The FP receptor is also expressed in neuronal and astrocyte cultures (Kitanaka et al., 1991, 1993; Gotoh et al., 1994). In the brain, astrocytes, oligodendrocytes, and white matter of the striatum express TP receptor mRNA (Borg et al., 1994; Kitanaka et al., 1995, 1996; Honma et al., 2006). Like the expression patterns of prostaglandin receptors, when activated, the signaling cascades that follow are also varied.
Figure 2 illustrates the overview of prostaglandin receptor-mediated signaling. Depending on a given prostaglandin receptor, we and others have found that global deletion of specific prostaglandin receptors can greatly influence stroke outcomes (Ahmad et al., 2005, 2006a, 2008, 2010a; Saleem et al., 2007a, 2009a,b,c). However, the mechanisms that mediate these outcomes following ICH have not been fully elucidated. The prostaglandin receptors can be grouped according to the downstream signaling pathways they activate.
Prostaglandin Receptors and Signaling
Prostaglandins exert cellular affects through their specific receptors: PGE2 receptors EP1, EP2, EP3, and EP4; PGD2 receptors DP1 and DP2; PGF2α receptor FP; PGI2 receptor IP; and TxA2 receptor TP (Narumiya et al., 1982, 1991, 1999; Ogorochi et al., 1984; Narumiya and Toda, 1985; Ito et al., 1989; Namba et al., 1992, 1994; Honda et al., 1993; Irie et al., 1993; Watabe et al., 1993; Hirata et al., 1994a, 1996; Katsuyama et al., 1994, 1995; Hasegawa et al., 1996; Ishikawa et al., 1996; Kiriyama et al., 1997; Kobayashi and Narumiya, 2002; Woodward et al., 2011). Alternative splicing of the C-terminal has generated additional isoforms of human TP (TPα, TPβ), FP (and some relatively rare splice variants, not receptor subtypes), and eight EP3 receptor isoforms (Narumiya and Fitzgerald, 2001). Both human and mouse prostaglandin receptors signal via either Gαs (EP2, EP4, IP, and DP1 receptors) to increase intracellular levels of cyclic-adenosine monophosphate (cAMP), or Gαq (EP1, EP3, and FP) to increase intracellular levels of calcium or Gαs-proteins (EP3, DP2, and TP receptors) to increase or decrease intracellular levels of cAMP and calcium. Table 2 depicts these prostaglandin receptors and their signaling and binding properties.
In addition to cAMP-PKA dependent second messenger signaling, which activates G-protein, G-protein activation also occurs via a PKA-independent mechanism. An example of protein activated by cAMP but PKA-independent include the exchange protein directly activated by cAMP (Epac1 and 2). Epac1 and 2 function as guanine nucleotide exchange factors for the small G-protein Rap. Epac proteins are expressed throughout the body, with Epac1 specifically abundant in blood vessels, kidney, adipose tissue, CNS, ovary, and uterus, whereas Epac2 is mostly expressed in the CNS, adrenal gland, and pancreas (de Rooij et al., 1998; Kawasaki et al., 1998). Due to the abundance of GPCRs that mediate cAMP signaling, Epac proteins have many biological functions. For example, Epac regulates electrically evoked Ca2+ transients in response to β-adrenergic receptors, Epac2 activation induces exocytosis in human β cells, increasing the number of exocytic sites on the plasma membrane via its effects on Ca2+ signaling, and effects neuronal function following activation by potentiating the postsynaptic excitation currents (Kang et al., 2003; Kaneko and Takahashi, 2004; Wang et al., 2005; Cheung et al., 2006; Gekel and Neher, 2008). Furthermore, together with PKA, Epac also contributes to the regulation of neuronal differentiation, neurite outgrowth, and axon generation, implicating an Epac role in the development and maintenance of the nervous system (Christensen et al., 2003; Kiermayer et al., 2005; Shi et al., 2006; Murray and Shewan, 2008). In addition and perhaps relevant to the ICH-pathology, Epac proteins have been implicated in vascular function.
In vivo administration of a Epac agonist (007) inhibited vascular endothelial growth factor-induced dye leakage from mouse dermal blood vessels (Fukuhara et al., 2005). Also, 007 induced secretion of von Willebrand factor-containing Weibel–Palade bodies, which may further contribute to the regulation of vasculature homeostasis by Epac1 (Rondaij et al., 2004). Epac proteins have also been implicated in inflammation, where cAMP signaling directly controls inflammation by regulating leukocyte-mediated immune responses (Lorenowicz et al., 2007). Epac proteins are expressed in leukocytes and following the activation of Epac1 regulate the monocytes adhesion and chemotaxis (Lorenowicz et al., 2006). Epac1 also induces pathogen-mediated production of pro-inflammatory cytokines and chemokines (Gerlo et al., 2010). Due to recent reports on the role of Epacs in the brain and neurons, we hypothesis that Epacs may function to minimize neuronal death involved in ICH. Such Epac-mediated neuroprotection may occur by potentiating PKA-independent activation of the small G-protein Rap by cAMP. Mechanisms of Epac-dependent neuroprotection remain to be studied in correlation to prostaglandin receptor activation. Protein homology between human and mouse prostaglandin receptors are at best 88% (i.e., human FP receptor vs. mouse FP receptor) with little homology between receptor subtypes. A phylogenetic tree and table of the amino acid homology (% identity) between human and mouse prostaglandin receptors is illustrated in Table 1 and Figure 1, respectively. Due to the mechanistic nature of prostaglandin receptors, evolutionary relationships among human and mouse prostaglandin receptors can be broadly grouped into two signaling pathways: essentially leading to cAMP or calcium signaling cascade. Table 2 and Figure 2 illustrate signal transduction mechanisms activated when prostaglandin receptor specific agonist bind. Prostaglandin receptors have been grouped into specific G-protein-dependent cAMP and/or Ca2+ signaling. Prostaglandin receptors are expressed throughout the peripheral and central nervous system and therefore play an important role in the physiologic response following injury.
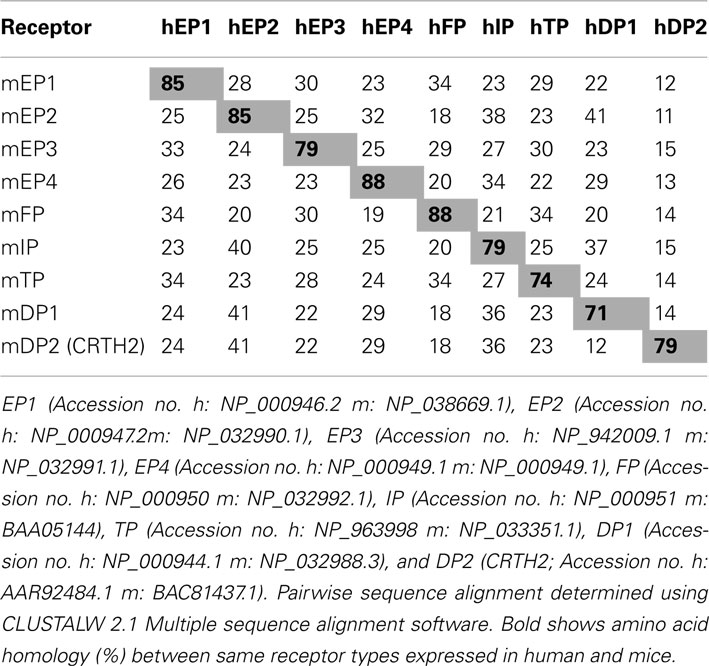
Table 1. Amino acid homology (% identity) between human (h) and mouse (m) prostaglandin receptors.
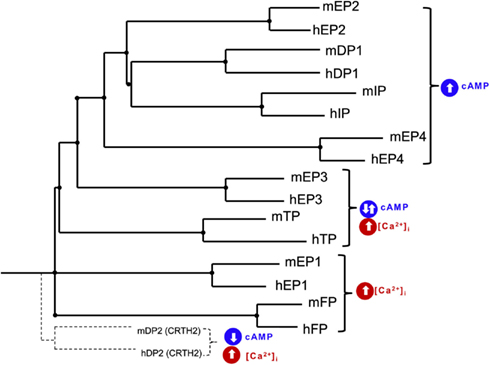
Figure 1. Phylogenetic tree of human (h) and mouse (m) EP1, EP2, EP3, EP4, FP, IP, TP, DP1, and DP2 (CRTH2) receptors. Tree was constructed using pairwise sequence alignment determined using CLUSTALW 2.1 Multiple sequence alignment software and presented as NJplot. Evolutionally conserved prostaglandin receptors can be grouped into signaling pathways: cAMP stimulating and calcium signaling.

Table 2. Structural, signal transduction, and agonist binding properties of mouse prostaglandin receptors.
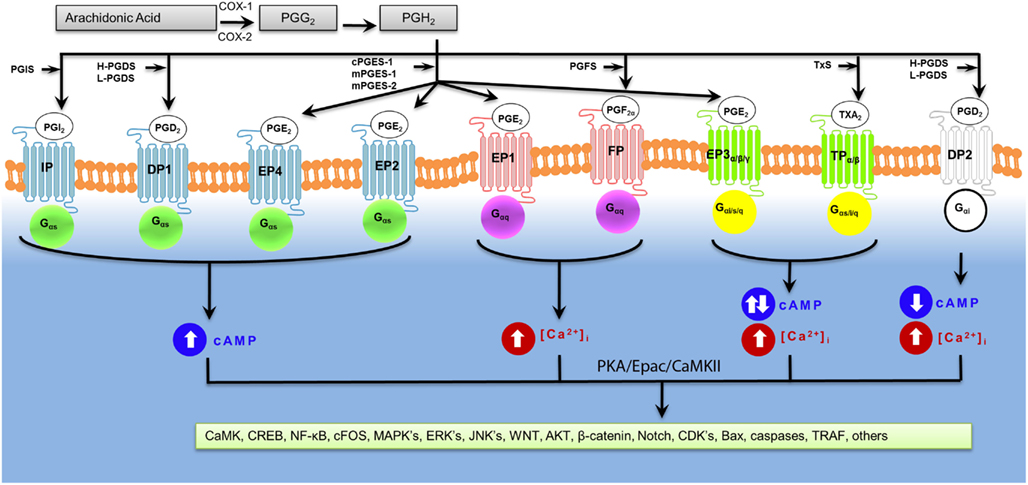
Figure 2. Signal transduction mechanisms of activated prostaglandin receptors. Prostaglandin receptors have been grouped into specific G-protein-dependent cAMP and/or Ca2+ signaling.
Prostaglandin Receptors (IP, DP1, EP2, EP4) Activating cAMP Pathway
PGI2-IP Receptor
Prostacyclin is a primary prostaglandin produced by endothelial cells and plays an important role in vascular homeostasis (Vane and Botting, 1995). Through the activation of its IP receptor, PGI2 induces vasodilation, inhibits platelet aggregation at high concentrations, and proliferates smooth muscle cells via Gαs-proteins, and increased cAMP levels (Moncada et al., 1977a,b; Falcetti et al., 2010). Due to their tendency for post-translational modifications, the IP receptors are capable of coupling to other signal transduction pathways via Gαq-protein dependent phosphoinositide turnover and Gαi-protein dependent inhibition of cAMP (Katsuyama et al., 1994; Hebert et al., 1998; Miggin et al., 2003). IP receptor-mediated vasodilation may also be modulated by the co-activation of EP3 receptors that couple to Gαi-proteins and therefore further research on role of PGI2-IP receptor following stroke is required (Orie and Clapp, 2011).
The role of PGI2 in response to neuronal injury or toxicity was measured using rat cortical neuron cultures subjected to hypoxia and glutamate toxicity (Cazevieille et al., 1993, 1994). Following transient ischemia in mice, PGIS and PGI2 levels are increased in neurons, macrophages, and microglial cells of the brain (Fang et al., 2006). Increased levels of PGIS and PGI2 provided neuroprotection against ischemia indicated by a reduction in infarct volume following adenovirus mediated overexpression of PGIS 72 h after ischemia (Fang et al., 2006). The role of IP receptors in stroke was not determined until recently.
We have also demonstrated the neuroprotective properties of the IP receptor in mouse models of transient middle cerebral artery occlusion (tMCAO) and permanent distal MCAO (pMCAO; Saleem et al., 2010). IP receptor knockout mice showed increased infarct volumes and neurological deficit scores compared with wildtype mice, and pretreatment with a selective IP receptor agonist (beraprost) reduced infarct volumes and deficit scores, confirming the neuroprotective role of IP receptor activation (Saleem et al., 2010). In addition, our group also found that when deleted, IP receptors aggravated hippocampal neuronal loss after bilateral common carotid artery occlusion in mice (Wei et al., 2008). The neuroprotective role of these CNS-specific IP receptor ligands was also demonstrated in monkeys subjected to MCAO (Cui et al., 2006).
Some reports suggest the existence of tissue specific subtypes of IP receptors. It is noteworthy that the activation of IP receptor (IP2) in the CNS prevented oxygen-induced neuronal death and protected CA1 pyramidal neurons against ischemia in gerbils (Takechi et al., 1996; Satoh et al., 1999; Watanabe et al., 1999). In rats subjected to MCAO, the effects of an IP receptor ligand (15-deoxy-TIC) designed to bind the IP2 receptor was found to be significantly neuroprotective when compared with the peripheral type IP receptor ligand, iloprost methylester, administered 24 h after ischemia (Takamatsu et al., 2002).
From evidence supporting the neuroprotective role of IP receptors, we hypothesize that activation of IP receptor could also serve to protect from ICH. The effects of increased IP receptor-dependent activation of cAMP signaling following ICH remain to be fully elucidated. Inversely, because of the vasodilatory properties of PGI2, it is possible that the activation of IP receptors would also potentially exacerbate the hemorrhagic transformation and hemorrhagic damage.
PGD2-DP1 Receptor
PGD2 has both peripheral and central physiologic effects (Whittle et al., 1983; Narumiya and Toda, 1985; Casteleijn et al., 1988; Sturzebecher et al., 1989; Darius et al., 1994; Matsugi et al., 1995; Matsuoka et al., 2000; Angeli et al., 2004). Activated DP1 receptor leads to the activation of the cAMP/PKA pathway and is one of the main cellular mechanism through which DP1 receptors exert their neuroprotective effects (Liang et al., 2005). Activation of the DP1 receptor stimulates adenylyl cyclase, leading to increased levels of cAMP, and decreased platelet aggregation (Hata and Breyer, 2004). PGD2 levels are significantly increased under pathologic conditions; however, the effects of PGD2 remain dependent on the cell type (Hata and Breyer, 2004; Hatoum et al., 2005). For example, PGD2 inhibited TNF-α-induced migration of Langerhans cells, which are involved in cutaneous inflammatory responses by decreasing the infiltration of T helper cells (Angeli et al., 2004). The anti-inflammatory properties of PGD2 are primarily associated with DP1 receptors. For example, DP1 receptor activation resulted in decreased allergic and asthma response (Matsuoka et al., 2000; Hammad et al., 2003; Angeli et al., 2004).
In the brain, until recently DP1 receptor related studies were limited to investigations of sleep induction, modulation of body temperature, olfactory function, hormone release, nociception, eye movement, and neuromodulation (Eguchi et al., 1999; Urade and Hayaishi, 1999; Mizoguchi et al., 2001; Hayaishi, 2002; Obal and Krueger, 2003; Angeli et al., 2004; Hata and Breyer, 2004; Gelir et al., 2005; Koch et al., 2005). In addition to expression and function in the brain, the neuroprotective effects of the DP1 receptor was revealed following activation by PGD2 protected hippocampal cultures derived from E18 rat embryos against glutamate toxicity (Liang et al., 2005). In a mouse model of cerebral ischemia, we and others have shown that PGD2 mediates neuroprotection via the DP1 receptors (Liang et al., 2005; Saleem et al., 2007c). We have also shown that NMDA-induced excitotoxicity and cerebral ischemic brain damage were significantly attenuated by the DP1 receptor-selective agonist BW245C (Ahmad et al., 2010a).
Although the potential neuroprotective effect of DP1 receptors has been reported, the signaling cascade leading to this neuroprotective effect remains unclear. Based on the reports from us and others, it is evident that activation of DP1 receptors leads to increased levels of cAMP/PKA. However, more experiments are required to fully elucidate the role of DP1 receptors in ICH. Based on the role of DP1 receptors in activating the cAMP/PKA pathway, we expect that these receptors could be novel endogenous targets in attenuating ICH. However, the vasodilatory effect of PGD2 and DP1 could also pose a higher risk of hemorrhage. Nevertheless, use of pharmacologic and genetic approaches targeting DP1 receptors could provide better insight and might help in protecting the brain from devastating hemorrhagic conditions.
PGE2-EP2 Receptor
Among various effects of PGE2 many of them have been attributed to its known capacity to stimulate cAMP via EP2 receptors and selective agonist also increase cAMP levels in a concentration-dependent manner with the same potency as PGE2 (Narumiya et al., 1999). EP2 receptor-selective agonists increase the cAMP levels in a concentration-dependent manner with the same potency as PGE2 (Choi et al., 2001). PGE2-stimulated cAMP formation has been shown to be blocked not only by an EP1/EP2 receptor antagonist, AH6809, but also by an inhibitor of adenylyl cyclase, SQ22536 (Fiebich et al., 2001).
In the brain, activation of EP2 receptors by PGE2 is involved in long-term synaptic plasticity and cognitive function, where mice deficient in EP2 receptors showed impaired hippocampal synaptogenesis (Sang et al., 2005; Yang et al., 2009). Therefore it is no surprise that brain injuries can affect the expression and function of EP2 receptors. For example, EP2 receptor expression increased in CA1 pyramidal neurons following cerebral ischemia in rats (Choi et al., 2006). However, does this increase in expression of EP2 receptors following brain injury result from its neuroprotective properties when activated? In support of this question, our group and others have shown EP2 receptor-mediated neuroprotection when subjected to glutamate receptor-mediated toxicity and MCAO and that EP2−/− mice has increased damage (Liu et al., 2005; Ahmad et al., 2006b). In a mouse model of focal cerebral ischemia, we have shown that butaprost, an EP2 receptor-selective agonist, provided dose-dependent neuroprotection, whereas deletion of EP2 receptors aggravated ischemic brain damage (Ahmad et al., 2010b). Neuroprotection mediated by EP2 receptor activation occurs by a PKA-dependent mechanism, as demonstrated, against oxidative stress and excitotoxicity (Echeverria et al., 2005).
Additional studies support the mechanism by which PGE2 affords neuroprotection through EP2 receptor-associated increases in cAMP, followed by a PKA-dependent pathway (Araki et al., 2000; McCullough et al., 2004; Jiang et al., 2010). In an attempt to discover neuroprotective agents, EP2 receptor agonist conferred neuroprotection in an NMDA receptor-induced excitotoxicity model, strongly reinforcing the notion that EP2 receptor activation by endogenous PGE2 in a cell-injury setting is neuroprotective (Jiang et al., 2010). These findings and others suggest the cAMP/PKA pathway is the site of neuroprotection following EP2 receptor activation (Araki et al., 2000; McCullough et al., 2004; Liu et al., 2005). In addition to EP2 receptor-mediated PKA-dependent neuroprotection, cAMP activation independent of PKA may also afford neuroprotection. However, recent studies have reported that PGE2 activation of the EP2 receptor had anti-proliferative effects in human gingival fibroblasts and this seems to be mediated by the EP2-cAMP-Epac pathway (Weinberg et al., 2009). Research on cortical neurons showed increased apoptosis through the expression of the Bcl-2 interacting membrane protein Bim and may therefore warrant further research on how Epac regulation may be used as a strategy for the treatment of ICH (Suzuki et al., 2010).
In contrast to previous studies demonstrating the neuroprotective effects of EP2 receptor activation, a recent study showed that when blocked using a selective small molecule EP2 receptor antagonist also significantly reduced neuronal injury in the hippocampus when administered in mice beginning 1 h after termination of pilocarpine-induced status epilepticus (Jiang et al., 2012). This study provides unique insight into the effects of EP2 receptor blockade and may also prove to be a therapeutic strategy for inflammation-related brain injuries such as ICH. Identification of the specific PGE2 receptor subtypes involved in acute brain injury following ICH will provide the building blocks for future studies aimed at generating new tools for possible therapeutic interventions. Similarly to EP2 receptors, EP4 receptors have been found to activate the cAMP/PKA pathway; however, EP2 and EP4 receptors have significant differences.
PGE2-EP4 Receptor
Like the EP2 receptor, most of the effects PGE2 mediated activation of EP4 receptor is due to cAMP signaling; however, there is a subtle difference between their abilities to stimulate cAMP. Using COS cells expressing both EP2 and EP4 receptors, PGE2 stimulation produced ∼11- and ∼8-fold increase in cAMP, respectively (Honda et al., 1993; Regan et al., 1994). EP4 receptor-mediated weaker coupling to Gαs-proteins suggests cAMP plays a less important role in EP4 receptor signaling compared to EP2 receptors (Fujino et al., 2002, 2005). Regardless of the differences in the formation of intracellular cAMP, EP4 receptor had a greater affinity for PGE2 than the EP2 receptor (Fujino et al., 2003). Compared to EP2, EP4 also has a longer C-terminus, which has been implicated in agonist-induced desensitization and internalization (Nishigaki et al., 1996; Bastepe and Ashby, 1999; Desai et al., 2000). Despite the low levels of intracellular cAMP, when activated following injury, the EP4 receptor has proven to have neuroprotective properties.
It is well documented that EP4 receptor activation has anti-inflammatory effects (Narumiya, 2009; Shi et al., 2010; Tang et al., 2012). Also, in the same study, conditional inactivation of EP4 receptor in neurons and endothelial cells increased infarct size in mice subjected to MCAO. In addition to its neuroprotective effects, EP4 receptor activation blocked LPS-induced pro-inflammatory gene expression in the brain tissue of mice. Additionally, an in vivo study by the same group, demonstrated that conditional deletion of EP4 receptor in macrophages and microglia increased lipid peroxidation and pro-inflammatory gene expression in the brain (Shi et al., 2010). We have shown that β-amyloid-induced (Aβ42) toxicity is minimized by EP4 receptor activation (Echeverria et al., 2005). Similarly, following acute striatal excitotoxicity, activation of EP4 receptor protected the brain (Ahmad et al., 2005).
In addition to their role in neuroprotection, endothelial cells expressing EP4 receptor are involved in vasodilation due to the direct activation of endothelial NOS, thus their role in the relaxation of smooth muscles (Dumont et al., 1999). Because EP4 receptor signaling is more complex (involving not only Gαs-protein mediated increase in cAMP but also coupling to pertussis toxin-sensitive Gαi-proteins and β-arrestin mediated effects), the role of EP4 receptors in ICH may be more dynamic than in ischemic stroke (Penn et al., 2001; Fujino and Regan, 2006).
Due to the role of EP4 receptors in vasodilation, we hypothesize that activation of EP4 receptors may play an important role in the control of cerebral blood flow, and thus may represent a novel target for the prevention or treatment of cerebral ischemia. In ICH, the endothelium and cerebral vasculature are compromised by potential brain injury with the expansion of blood away from the region of hematoma. To date, the exploration of EP4 receptors as a target to protect or rescue neurons from blood components after ICH remains to be studied. As mentioned, the effects of PGE2 are vast and beyond the scope of this review; however, the varied effects of the PGE2 may be attributed to the receptors onto which it binds and activates. For example, PGE2 when bound to its receptors cannot only stimulate cAMP signaling but also calcium signaling by binding to EP1 and FP receptors.
Prostaglandin Receptors (EP1 and FP) Activating Calcium Pathway
PGE2-EP1 Receptor
When activated, EP1 receptors couple to Gαq-proteins, resulting in increased phosphatidyl inositol hydrolysis and elevation of intracellular Ca2+. EP1 receptor activation by PGE2 leads to increased Ca2+ signaling and subsequent neurotoxicity (Kandasamy and Hunt, 1990; Kawano et al., 2006). EP1 receptor-mediated neuronal toxicity was found to be normalized to basal levels following either blockade or deletion of EP1 receptors, and improved function of the Na+/Ca2+ exchanger (Kawano et al., 2006). Glial-derived neurotrophic factor (GDNF) therapy has been shown to be beneficial in treating Parkinson’s disease when EP1 receptor is blocked or ablated, resulting in a 60% enhancement in GDNF, which suggests that selective inhibition of EP1 receptor signaling might be a means to augment GDNF secretion in diseased regions of the brain (Li et al., 2012).
Due to decreased calcium signaling, pharmacological blockade of EP1 receptor could make good therapeutic targets against brain injury (Gendron et al., 2005). Therefore, using EP1−/− mice and selective EP1 receptor antagonist, we and others have found that EP1 receptors contribute to excitotoxicity following focal cerebral ischemia (Ahmad et al., 2006a, 2008; Saleem et al., 2007b; Abe et al., 2009). Using a neonate model of hypoxic-ischemia encephalopathy, EP1 receptor blockade with a selective antagonist, SC-51089, and co-activation of EP2–4 receptors, cerebral injury was reduced 24 h after injury (Taniguchi et al., 2011b). The role of Ca2+ signaling in neuronal activity and death following ICH is unknown; however, current data suggests that increased Ca2+ signaling contributes to increased cerebral vasospasm commonly seen in SAH (Kikkawa et al., 2010; Koide et al., 2011). Neuroprotection following the blockade of EP1 receptor has been found to involve the PTEN/AKT survival pathway following ischemic stroke (Zhou et al., 2008; Abe et al., 2009). Relating the blockade of EP1 receptor to neuroprotection in vivo may be complicated by the role of various cells types; for example, neuroprotection via the blockade of EP1 receptor was reduced by the presence of microglia in NMDA stimulated neuron-glial cultures (Carlson et al., 2009). The role of EP1 receptor in ICH is unknown. However, one study showed increased expression of EP1 receptors following hemorrhage in splenic macrophages from male mice (Stapleton et al., 2004).
Data accumulating thus far from ischemic brain injuries and recent data from our group on ICH suggests that activation of EP1 receptor might lead to different outcomes in different ICH models. Models of ICH at different time points, using both EP1 receptor knockout and pharmacological approaches to elucidate the role of EP1 receptor would increase our in vivo knowledge of EP1 receptors in ICH. Therefore, we recommend further detailed investigations into the potentialities of EP1 receptor as a therapeutic target.
PGF2α-FP Receptor
The amino acid sequence of FP receptor has a high sequence homology with that of the PGE2-EP1 receptor, and shares the same phylogenetic branch with EP1 receptor (Toh et al., 1995). Despite the abundance of arachidonic acid in the brain, the function of its metabolite, PGF2α, is poorly understood. However, PGF2α is known to play a significant role in the initiation of parturition, renal function, control of cerebral blood flow, and intraocular pressure principally by an increase in uveoscleral outflow of aqueous humor, autoregulation in newborn piglets, contraction of arteries, and myocardial dysfunction (Chemtob et al., 1990; Sugimoto et al., 1997; Takayama et al., 2005; Jovanovic et al., 2006; Hao and Breyer, 2007). Pathological conditions in humans influence PGF2α levels in cerebrospinal fluid, where elevated levels of PGF2α were measured following epilepsy, meningtitis, brain injury, and stroke (Wolfe and Mamer, 1975; Kostic et al., 1984). Interestingly, elevated levels of PGF2α in the CSF of patients following acute cerebral ischemia did not correlate with the degree of neurological deficit (Kostic et al., 1984).
Activation of FP receptor initiates several events, including stimulation of the phospholipase C/IP3R/Ca2+ signaling pathway (Heaslip and Sickels, 1989; Abramovitz et al., 1994; Knock et al., 2005). FP receptor activation causes the release of Inositol 1,4,5-triphosphate and diacylglycerol in turn activating the calcium-calmodulin-CaMK-II pathway, which may be associated with FP receptor-mediated excitotoxicity following transient focal brain ischemia (Narumiya et al., 1999; Saleem et al., 2009a). Using FP−/− mice we have shown that FP receptor is involved in the enhancement of cerebral ischemia and excitotoxic brain injury and may mitigate the effects of ischemic stroke brain injury (Saleem et al., 2009a). Recently, we have also demonstrated that FP receptor blockade and knockout protects against ischemic stroke in mice and oxygen-glucose deprivation-induced cell death in slice cultures (Kim et al., 2012).
Although FP receptor has been implicated in ischemic stroke, the roles of PGF2α and FP receptor have not been studied in the pathogenesis of ICH. Because FP receptor regulates [Ca2+]i levels, we propose that the activation subsequent to injury contributes to excitotoxicity and hemorrhagic brain damage. Therefore, blockade of FP receptor would be beneficial for the treatment of ischemic stroke.
Prostaglandin Receptors (EP3, TP, DP2) Activating Both cAMP and/or Calcium Pathway
PGE2-EP3 Receptor
Because of its coupling to several G-proteins the EP3 receptor has various biological properties. EP3 is important in a number of physiological functions including vasoconstriction of the pulmonary arteries, growth inhibition of keratinocytes, and inhibition of aromatase activity in breast fibroblasts (Qian et al., 1994; Konger et al., 1998, 2005; Richards and Brueggemeier, 2003). The EP3 receptor has multiple splice variants that differ in both their C-terminal tails and signaling pathways (Kotani et al., 1995). The activation of the human EP3 subtypes EP3I, EP3II, EP3III, EP3IV, EP3e, and EP3f isoforms have been shown to inhibit cAMP production, whereas stimulation of EP3I, EP3II, and EP3III increase IP3/[Ca2+]i (Kotani et al., 1995; Schmid et al., 1995). The EP3 receptor isoforms expressed in mice (α, β, and Γ) contain carboxyl tails of 30, 26, and 29 amino acids that modulate signal transduction (Irie et al., 1994). In this context, EP3α and EP3β receptors couple to Gαi-proteins and inhibit cAMP, whereas the EP3γ couples to Gαs-proteins in addition to Gαi-proteins and evokes cAMP production (Sugimoto et al., 1993). In neurological diseases such as stroke, the function of EP3 receptor so far is not firmly defined; this may be in part due to the variants of the EP3 receptor subtypes.
When activated with selective EP3 receptor agonist ONO-AE-248, our group recorded a dose-dependent increase in infarct volume after MCAO in mice (Ahmad et al., 2007). Recently, the proposed mechanism of EP3 receptor-mediated neuronal death in stroke has been implicated by the glutamate-dependent increase in mPGES-1 activity, which in turn increases EP3 receptor activation and activation of Rho and/or Gαi proteins signaling (Ikeda-Matsuo et al., 2010). In contrast to EP3 activation, we have shown that the deletion of EP3 receptor results in decreased infarct volume 48 h after ischemia (Saleem et al., 2009b). Another group showed that deletion of the EP3 receptor did not alter infarct volume or behavior 24 h after ischemia (Li et al., 2008). The discrepancy in results could be due to differences in time points post-MCAO used to measure infarct volume and the age of the mice used in both studies. Also, recently it has been shown that when EP3 receptors are deleted, damage is done to the blood-brain barrier, activation of microglia and infiltration of neutrophils into the ischemic cortex are reduced, and the underlying neuroprotection mediated by EP3 receptor deletion may be due to decreased inflammation and apoptotic signaling (Ikeda-Matsuo et al., 2011).
No current study has correlated the activation of EP3 receptors in ICH. Three isoforms of the EP3 receptor are expressed in mice; consequently, the EP3 receptor role in ICH may prove to be different than its role in ischemic stroke because different cell types may not only express different levels of EP3 receptors, but their functions may be different after ICH. Furthermore, due to the differences between isoforms, it is possible that the dose and time lapse in administration of EP3 receptor drug treatments post-stroke could regulate the final outcomes. Therefore, use of conditional EP3 receptor transgenic animals or a therapeutically selective agonist/antagonist for these EP3 receptor isoforms could be beneficial in discerning the role that each of these isoforms plays following ICH.
TxA2-TP Receptor
TxA2-TP receptor activity is coupled with Gαs-, Gαq-, Gαi-, and Gα12/13/15/16-proteins (Shenker et al., 1991; Laugwitz et al., 1996; Muck et al., 1998). These G-proteins in turn regulate several effectors including phospholipase C, adenylyl cyclase, cAMP, guanine nucleotide exchange factor of the small G-protein Rho, and intracellular calcium (Kozasa et al., 1998). In humans there are two isoforms of TP receptors, TPα (placental/platelet) and TPβ (endothelial), which account for some of the differences in intracellular signaling after activation of TP receptors. These isoforms differ in length and sequence at the C-terminal distal to the last amino acid (Arg328) and are expressed in most tissues including platelets, placenta, vascular smooth muscle, brain, small intestine, and thymus (Colman, 1991; Hirata et al., 1991; Namba et al., 1992; Ushikubi et al., 1993; Raychowdhury et al., 1994; Miggin and Kinsella, 1998). Despite the relatively limited physiological role of TPβ receptors, their activation results in pertussis toxin-sensitive inhibition of cAMP (Namba et al., 1992; Hirata et al., 1996). In mice, no TP receptor isoforms exist; therefore, TP receptors in mice are considered to be similar to human TPα (74% identical, see Table 2 and Figure 1) and when activated, mediate the increase in cAMP following I-BOP (a TP receptor agonist) treatment.
Due to the isoforms, TP receptor signaling in humans varies with tissue type. For example, in platelets, TP receptor coupling to Gαq-proteins results in increased [Ca2+]i (Hirata et al., 1994b). In addition, platelet activation may also occur due to theGα12/13-protein pathways (Offermanns et al., 1996). In the CNS, hippocampal TP receptor plays a functional role in both neuronal excitability and synaptic transmission (Schwartz-Bloom et al., 1996). Activation of presynaptic TP receptor results in increased glutamate release, and postsynaptic activation of TP receptor results in the inhibition of synaptic transmission, suggesting that TP receptor plays different roles based on localization (Hsu and Kan, 1996). Moreover, activation of TP receptor dose-dependently suppressed whole-cell Ca2+ currents in rat CA1 neurons (Hsu et al., 1996).
TP receptors play an important role in the development of cerebral ischemia and mediate vascular proliferation and contraction (De Clerck and Janssen, 1990). Following SAH, mRNA and protein levels of TP receptors were elevated in cerebral arteries and smooth muscle cells, respectively (Ansar et al., 2010). Elevated levels of TxA2 in CSF have been reported in patients after SAH and ICH (Pickard et al., 1994; van Kooten et al., 1999b). TxA2 mediated activation of TP receptors expressed on platelets surrounding the region of the brain (cortex and striatum) has been linked to exacerbating ICH injury (Kong et al., 1991; van Kooten et al., 1999a; Saloheimo et al., 2005; Yalcin et al., 2005). Due to the expression and distribution of TP receptor in the brain, TP receptors could be considered an ideal therapeutic target for the treatment of ICH. Platelet activation is often a reflection of vascular risk factors, diffused atherosclerotic lesions, or the extent of damage caused by stroke. Terutroban, a selective TP receptor antagonist (Ki = 0.82 nM) used as an antithrombotic agent, could also be used to prevent atherothrombosis and ischemic stroke (Chamorro, 2009). Using a spontaneous hypertensive stroke-prone rat model, terutroban increased the survival rate by reducing systemic inflammation, thus promoting the TP receptor antagonists therapeutic intervention for stroke as a possible antithrombotic agent in human compared to aspirin alone (Gelosa et al., 2010). However, in a recent human study, no significant differences in outcomes were recorded between terutroban and aspirin in patients with cerebral ischemia (Bousser et al., 2011). Several receptors are upregulated following ischemia and the mulifactorial nature of ischemia may explain the lack of effect of many substances tested in clinical trials. These accumulating data warrant further examination of the therapeutic potential of TP receptors and their selective ligands to fully elucidate their role in ICH.
PGD2-DP2 (or CRTH2) Receptor
One of two PGD2 receptors, DP2 receptor, is the most recently discovered. DP2 receptor was initially cloned as an orphan receptor and later identified as T helper cell type 2 cells (Th2), specific surface PGD2 receptor (Nagata et al., 1999a,b; Hirai et al., 2001). The DP2 receptor is distinguished by their similarity to chemoattractant receptors and thus, they are also known as chemoattractant receptor-homologous molecules expressed on Th2 cells (CRTH2; Monneret et al., 2001). The CRTH2 receptors are also known as DP2 receptor based on their similar binding affinity to that of DP1 toward PGD2, although the DP1 and DP2 receptors do not share structural homology (Hirai et al., 2001). Similar to many chemoattractant receptors, the DP2 receptor is coupled to Gαi-proteins, leading to the inhibition of cAMP and increase in Ca2+ in a variety of cell types (Sawyer et al., 2002). Given the role of the DP2 receptor in inflammatory diseases such as asthma, blocking of DP2 receptor represents a novel therapeutic approach for the treatment of such conditions.
Despite its expression in the spinal cord and its anti-inflammatory effect as an activator of peroxisome proliferator-activated receptors, little is known about the role of DP2 receptors after stroke (Genovese et al., 2008; Grill et al., 2008; Morgenweck et al., 2010). Based on the role of the DP2 receptor in cAMP inhibition and Ca2+ activation, we hypothesize that activation of this receptor could lead to aggravated brain damage and its inhibition could lead to better functional and anatomical outcomes after ICH. Moreover, blockade of this receptor could result in more availability of PGD2 to DP1 receptor, which could then lead to increased cAMP level and subsequent neuroprotection as we have previously reported (Saleem et al., 2007c; Ahmad et al., 2010a). We hypothesize that blockade of DP2 receptor following ICH will result in greater injury based on the increased injury we observed with DP1−/− mice following ischemic stroke and acute NMDA-induced excitotoxicity due to similar binding affinity to DP1 receptor, DP2 receptors would have little therapeutic potential in ICH (Ahmad et al., 2010a).
Conclusion
Intracerebral hemorrhage accounts for 13% of all strokes in the United States each year. Following ICH, extravasated blood accumulates and compresses the surrounding brain tissue and treatment is primarily supportive with poor clinical outcome. To improve the outcome of ICH patients, a better understanding of the pathogenesis of ICH-induced brain injury is needed. We and others have identified prostaglandin receptors as potential therapeutic agents against ischemic stroke. Prostaglandins and their receptors regulate many physiological, inflammatory, and immunological processes. In the CNS, the role of prostaglandin receptors has been essentially first elucidated by gene-deletion studies and evidence suggests these receptors are a therapeutic target for the treatment of stroke. However, the therapeutic potential of prostaglandin receptors in ICH remains to be elucidated. Understanding the integration of a network of prostaglandin receptor signaling would improve our knowledge of these putative therapeutic targets for the effective treatment of neurological diseases. Evidence supports that prostaglandin receptors play a cooperative and/or sequential role in ICH-mediated inflammation and neurotoxicity. Intracellular signaling pathways activated by prostaglandin receptors in ICH are likely to be different than in ischemic stroke. However, we hypothesize that evolutionary regulated prostaglandin receptor signaling will be conserved and therefore this group of receptors, found to have a given role in ischemia may also have a unique role in hemorrhagic stroke. However it remains to be determined whether the effect of these receptors will be similar. Also, identification of a multitude of intracellular protein interactions with prostaglandin receptors may prove to regulate their neuronal expression, selectivity toward ligands, and crosstalk with cytokines, chemokines, and neurotransmitters. Studying these interactions and the cell-specific functions will help map a detailed network of signaling that would be necessary for an injury-induced cellular response. The discovery of a prostaglandin receptor specific intracellular response may enable us to navigate a complex pathway leading to the discovery of beneficial drugs for the treatment of hemorrhagic stroke.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors would like to thank all lab members for insightful comments used for the review. This work is supported in part by the National Institute of Health research grants (NS046400, AT005246, AG022971; Sylvain Doré) and University of Florida, Howard Hughes Medical Institute (HHMI) Science for Life Award (Chase Chambers).
Abbreviations
15-deoxy-TIC, 15-deoxy-(16-m-tolyl)-17,18,19,20 tetranorisocarbacyclin methyl ester; AKT, serine/threonine-specific protein kinase family; BP, blood pressure; BW245C, (4S)-(3-[(3R,S)-3-cyclohexyl-3-hydroxypropyl]-2,5-dioxo)-4-imidazolidineheptanoic acid; BWA868C, 3-[(2-cyclohexyl-2-hydroxyethyl)amino]-2,5-dioxo-1-(phenylmethyl)-4-imidazolidineheptanoic acid; CA1 pyramidal neurons, Cornu Ammonis-1 pyramidal neurons; CaMK, Ca2+/calmodulin-dependent protein kinase; cAMP, cyclic-adenosine monophosphate; COX, cyclooxygenase; CRTH2, chemoattractant receptor expressed Th2 lymphocytes; CSF, cerebrospinal fluid; Epac, exchange protein directly activated by cAMP; Gαi-protein, guanine nucleotide-binding protein (inhibitory); Gαq-protein, guanine nucleotide-binding protein (stimulatory/inhibitory); Gαs-protein, guanine nucleotide-binding protein (stimulatory); I-BOP, (Z)-7-[(1S,4R,5R,6R)-5-[(E,3R)-3-hydroxy-4-(4-iodophenoxy)but-1-enyl]-7-oxabicyclo[2.2.1]heptan-6-yl]hept-5-enoic acid; ICH, intracerebral hemorrhage; ICP, intracranial pressure; IP3R, inositol trisphosphate receptor; MCAO, middle cerebral artery occlusion; MK-0524, 2-[(3R)-4-[(4-chlorophenyl)methyl]-7-fluoro-5-methylsulfonyl-2,3-dihydro-1H-cyclopenta[b]indol-3-yl]acetic acid; NMDA, N-methyl-D-aspartic acid; NOS, nitric oxide synthase; ONO-AE-248, (Z)-7-[(1R,2R,3R)-3-methoxy-2-[(E,3S)-3-methoxyoct-1-enyl]-5-oxocyclopentyl]hept-5-enoic acid; PGDS, prostaglandin D synthase; PGES, prostaglandin E synthase; PGFS, prostaglandin F synthase; PGIS, prostaglandin I synthase; PKA, protein kinase A; PTEN, phosphatase and tensin homolog; SAH, subarachnoid hemorrhage; TS-022, 4-[(1R,2S,3R,5R)-5-Chloro-2-((S)-3-cyclohexyl-3-hydroxyprop-1-ynyl)-3-hydroxycyclopentyl]butylthio acetic acid monohydrate; TxS, thromboxane synthase.
References
Abe, T., Kunz, A., Shimamura, M., Zhou, P., Anrather, J., and Iadecola, C. (2009). The neuroprotective effect of prostaglandin E2 EP1 receptor inhibition has a wide therapeutic window, is sustained in time and is not sexually dimorphic. J. Cereb. Blood Flow Metab. 29, 66–72.
Abramovitz, M., Boie, Y., Nguyen, T., Rushmore, T. H., Bayne, M. A., Metters, K. M., et al. (1994). Cloning and expression of a cDNA for the human prostanoid FP receptor. J. Biol. Chem. 269, 2632–2636.
Adeoye, O., and Broderick, J. P. (2010). Advances in the management of intracerebral hemorrhage. Nat. Rev. Neurol. 6, 593–601.
Ahmad, A. S., Ahmad, M., De Brum-Fernandes, A. J., and Doré, S. (2005). Prostaglandin EP4 receptor agonist protects against acute neurotoxicity. Brain Res. 1066, 71–77.
Ahmad, A. S., Ahmad, M., Maruyama, T., Narumiya, S., and Doré, S. (2010a). Prostaglandin D2 DP1 receptor is beneficial in ischemic stroke and in acute exicitotoxicity in young and old mice. Age (Dordr.) 32, 271–282.
Ahmad, M., Saleem, S., Shah, Z., Maruyama, T., Narumiya, S., and Doré, S. (2010b). The PGE2 EP2 receptor and its selective activation are beneficial against ischemic stroke. Exp. Transl. Stroke Med. 2, 12.
Ahmad, A. S., Kim, Y. T., Ahmad, M., Maruyama, T., and Doré, S. (2008). Selective blockade of PGE2 EP1 receptor protects brain against experimental ischemia and excitotoxicity, and hippocampal slice cultures against oxygen-glucose deprivation. Neurotox. Res. 14, 343–351.
Ahmad, A. S., Saleem, S., Ahmad, M., and Doré, S. (2006a). Prostaglandin EP1 receptor contributes to excitotoxicity and focal ischemic brain damage. Toxicol. Sci. 89, 265–270.
Ahmad, A. S., Zhuang, H., Echeverria, V., and Doré, S. (2006b). Stimulation of prostaglandin EP2 receptors prevents NMDA-induced excitotoxicity. J. Neurotrauma 23, 1895–1903.
Ahmad, M., Ahmad, A. S., Zhuang, H., Maruyama, T., Narumiya, S., and Doré, S. (2007). Stimulation of prostaglandin E2-EP3 receptors exacerbates stroke and excitotoxic injury. J. Neuroimmunol. 184, 172–179.
Anderson, C. S., Huang, Y., Arima, H., Heeley, E., Skulina, C., Parsons, M. W., et al. (2010). Effects of early intensive blood pressure-lowering treatment on the growth of hematoma and perihematomal edema in acute intracerebral hemorrhage: the Intensive Blood Pressure Reduction in Acute Cerebral Haemorrhage Trial (INTERACT). Stroke 41, 307–312.
Angeli, V., Staumont, D., Charbonnier, A. S., Hammad, H., Gosset, P., Pichavant, M., et al. (2004). Activation of the D prostanoid receptor 1 regulates immune and skin allergic responses. J. Immunol. 172, 3822–3829.
Anrather, J., Gallo, E. F., Kawano, T., Orio, M., Abe, T., Gooden, C., et al. (2011). Purinergic signaling induces cyclooxygenase-1-dependent prostanoid synthesis in microglia: roles in the outcome of excitotoxic brain injury. PLoS ONE 6, e25916. doi:10.1371/journal.pone.0025916
Ansar, S., Larsen, C., Maddahi, A., and Edvinsson, L. (2010). Subarachnoid hemorrhage induces enhanced expression of thromboxane A2 receptors in rat cerebral arteries. Brain Res. 1316, 163–172.
Araki, H., Ukawa, H., Sugawa, Y., Yagi, K., Suzuki, K., and Takeuchi, K. (2000). The roles of prostaglandin E receptor subtypes in the cytoprotective action of prostaglandin E2 in rat stomach. Aliment. Pharmacol. Ther. 14(Suppl 1), 116–124.
Ayer, R., Jadhav, V., Sugawara, T., and Zhang, J. H. (2011). The neuroprotective effects of cyclooxygenase-2 inhibition in a mouse model of aneurysmal subarachnoid hemorrhage. Acta Neurochir. Suppl. 111, 145–149.
Ba, F., Giuliani, F., Camicioli, R., and Saqqur, M. (2012). A reversible cerebral vasoconstriction syndrome. BMJ Case Rep.
Bastepe, M., and Ashby, B. (1999). Identification of a region of the C-terminal domain involved in short-term desensitization of the prostaglandin EP4 receptor. Br. J. Pharmacol. 126, 365–371.
Batshake, B., Nilsson, C., and Sundelin, J. (1995). Molecular characterization of the mouse prostanoid EP1 receptor gene. Eur. J. Biochem. 231, 809–814.
Bentzer, P., and Grande, P. O. (2004). Low-dose prostacyclin restores an increased protein permeability after trauma in cat skeletal muscle. J. Trauma 56, 385–392.
Berges, S., Moulin, T., Berger, E., Tatu, L., Sablot, D., Challier, B., et al. (2000). Seizures and epilepsy following strokes: recurrence factors. Eur. Neurol. 43, 3–8.
Borg, C., Lim, C. T., Yeomans, D. C., Dieter, J. P., Komiotis, D., Anderson, E. G., et al. (1994). Purification of rat brain, rabbit aorta, and human platelet thromboxane A2/prostaglandin H2 receptors by immunoaffinity chromatography employing anti-peptide and anti-receptor antibodies. J. Biol. Chem. 269, 6109–6116.
Bousser, M. G., Amarenco, P., Chamorro, A., Fisher, M., Ford, I., Fox, K. M., et al. (2011). Terutroban versus aspirin in patients with cerebral ischaemic events (PERFORM): a randomised, double-blind, parallel-group trial. Lancet 377, 2013–2022.
Burn, J., Dennis, M., Bamford, J., Sandercock, P., Wade, D., and Warlow, C. (1997). Epileptic seizures after a first stroke: the Oxfordshire Community Stroke Project. BMJ 315, 1582–1587.
Caggiano, A. O., and Kraig, R. P. (1999). Prostaglandin E receptor subtypes in cultured rat microglia and their role in reducing lipopolysaccharide-induced interleukin-1beta production. J. Neurochem. 72, 565–575.
Carlson, N. G., Rojas, M. A., Black, J. D., Redd, J. W., Hille, J., Hill, K. E., et al. (2009). Microglial inhibition of neuroprotection by antagonists of the EP1 prostaglandin E2 receptor. J. Neuroinflammation 6, 5.
Casteleijn, E., Kuiper, J., van Rooij, H. C., Kamps, J. A., Koster, J. F., and van Berkel, T. J. (1988). Prostaglandin D2 mediates the stimulation of glycogenolysis in the liver by phorbol ester. Biochem. J. 250, 77–80.
Cazevieille, C., Muller, A., and Bonne, C. (1993). Prostacyclin (PGI2) protects rat cortical neurons in culture against hypoxia/reoxygenation and glutamate-induced injury. Neurosci. Lett. 160, 106–108.
Cazevieille, C., Muller, A., Meynier, F., Dutrait, N., and Bonne, C. (1994). Protection by prostaglandins from glutamate toxicity in cortical neurons. Neurochem. Int. 24, 395–398.
Chamorro, A. (2009). TP receptor antagonism: a new concept in atherothrombosis and stroke prevention. Cerebrovasc. Dis. 27(Suppl 3), 20–27.
Chaudhry, U., Zhuang, H., Crain, B. J., and Doré, S. (2008). Elevated microsomal prostaglandin-E synthase-1 in Alzheimer’s disease. Alzheimers Dement. 4, 6–13.
Chaudhry, U., Zhuang, H., and Doré, S. (2010). Microsomal prostaglandin E synthase-2: cellular distribution and expression in Alzheimer’s disease. Exp. Neurol. 223, 359–365.
Chemtob, S., Beharry, K., Rex, J., Varma, D. R., and Aranda, J. V. (1990). Prostanoids determine the range of cerebral blood flow autoregulation of newborn piglets. Stroke 21, 777–784.
Cheung, U., Atwood, H. L., and Zucker, R. S. (2006). Presynaptic effectors contributing to cAMP-induced synaptic potentiation in Drosophila. J. Neurobiol. 66, 273–280.
Choi, J. S., Kim, H. Y., Chun, M. H., Chung, J. W., and Lee, M. Y. (2006). Expression of prostaglandin E2 receptor subtypes, EP2 and EP4, in the rat hippocampus after cerebral ischemia and ischemic tolerance. Cell Tissue Res. 324, 203–211.
Choi, S. Y., Choi, B. H., Suh, B. C., Chae, H. D., Kim, J. S., Shin, M. J., et al. (2001). Potentiation of PGE(2)-mediated cAMP production during neuronal differentiation of human neuroblastoma SK-N-BE(2)C cells. J. Neurochem. 79, 303–310.
Christensen, A. E., Selheim, F., de Rooij, J., Dremier, S., Schwede, F., Dao, K. K., et al. (2003). cAMP analog mapping of Epac1 and cAMP kinase. Discriminating analogs demonstrate that Epac and cAMP kinase act synergistically to promote PC-12 cell neurite extension. J. Biol. Chem. 278, 35394–35402.
Cui, Y., Takamatsu, H., Kakiuchi, T., Ohba, H., Kataoka, Y., Yokoyama, C., et al. (2006). Neuroprotection by a central nervous system-type prostacyclin receptor ligand demonstrated in monkeys subjected to middle cerebral artery occlusion and reperfusion: a positron emission tomography study. Stroke 37, 2830–2836.
Darius, H., Michael-Hepp, J., Thierauch, K. H., and Fisch, A. (1994). Inhibition of human platelets and polymorphonuclear neutrophils by the potent and metabolically stable prostaglandin D2 analog ZK 118.182. Eur. J. Pharmacol. 258, 207–213.
De Clerck, F., and Janssen, P. A. (1990). 5-Hydroxytryptamine and thromboxane A2 in ischaemic heart disease. Blood Coagul. Fibrinolysis 1, 201–209.
De Herdt, V., Dumont, F., Henon, H., Derambure, P., Vonck, K., Leys, D., et al. (2011). Early seizures in intracerebral hemorrhage: incidence, associated factors, and outcome. Neurology 77, 1794–1800.
de Rooij, J., Zwartkruis, F. J., Verheijen, M. H., Cool, R. H., Nijman, S. M., Wittinghofer, A., et al. (1998). Epac is a Rap1 guanine-nucleotide-exchange factor directly activated by cyclic AMP. Nature 396, 474–477.
Desai, S., April, H., Nwaneshiudu, C., and Ashby, B. (2000). Comparison of agonist-induced internalization of the human EP2 and EP4 prostaglandin receptors: role of the carboxyl terminus in EP4 receptor sequestration. Mol. Pharmacol. 58, 1279–1286.
Doré, S., Sampei, K., Goto, S., Alkayed, N. J., Guastella, D., Blackshaw, S., et al. (1999). Heme oxygenase-2 is neuroprotective in cerebral ischemia. Mol. Med. 5, 656–663.
Dumont, I., Hou, X., Hardy, P., Peri, K. G., Beauchamp, M., Najarian, T., et al. (1999). Developmental regulation of endothelial nitric oxide synthase in cerebral vessels of newborn pig by prostaglandin E(2). J. Pharmacol. Exp. Ther. 291, 627–633.
Echeverria, V., Clerman, A., and Doré, S. (2005). Stimulation of PGE2 receptors EP2 and EP4 protects cultured neurons against oxidative stress and cell death following β-amyloid exposure. Eur. J. Neurosci. 22, 2199–2206.
Eguchi, N., Minami, T., Shirafuji, N., Kanaoka, Y., Tanaka, T., Nagata, A., et al. (1999). Lack of tactile pain (allodynia) in lipocalin-type prostaglandin D synthase-deficient mice. Proc. Natl. Acad. Sci. U.S.A. 96, 726–730.
Ek, M., Arias, C., Sawchenko, P., and Ericsson-Dahlstrand, A. (2000). Distribution of the EP3 prostaglandin E(2) receptor subtype in the rat brain: relationship to sites of interleukin-1-induced cellular responsiveness. J. Comp. Neurol. 428, 5–20.
Falcetti, E., Hall, S. M., Phillips, P. G., Patel, J., Morrell, N. W., Haworth, S. G., et al. (2010). Smooth muscle proliferation and role of the prostacyclin (IP) receptor in idiopathic pulmonary arterial hypertension. Am. J. Respir. Crit. Care Med. 182, 1161–1170.
Fang, Y. C., Wu, J. S., Chen, J. J., Cheung, W. M., Tseng, P. H., Tam, K. B., et al. (2006). Induction of prostacyclin/PGI2 synthase expression after cerebral ischemia-reperfusion. J. Cereb. Blood Flow Metab. 26, 491–501.
Fernandes, H. M., Gregson, B., Siddique, S., and Mendelow, A. D. (2000). Surgery in intracerebral hemorrhage. The uncertainty continues. Stroke 31, 2511–2516.
Fiebich, B. L., Schleicher, S., Spleiss, O., Czygan, M., and Hull, M. (2001). Mechanisms of prostaglandin E2-induced interleukin-6 release in astrocytes: possible involvement of EP4-like receptors, p38 mitogen-activated protein kinase and protein kinase C. J. Neurochem. 79, 950–958.
Fischborn, S. V., Soerensen, J., and Potschka, H. (2010). Targeting the prostaglandin E2 EP1 receptor and cyclooxygenase-2 in the amygdala kindling model in mice. Epilepsy Res. 91, 57–65.
Font-Nieves, M., Sans-Fons, M. G., Gorina, R., Bonfill-Teixidor, E., Salas-Perdomo, A., Marquez-Kisinousky, L., et al. (2012). Induction of COX-2 enzyme and down-regulation of COX-1 expression by lipopolysaccharide (LPS) control prostaglandin E2 production in astrocytes. J. Biol. Chem. 287, 6454–6468.
Fujino, H., and Regan, J. W. (2006). EP(4) prostanoid receptor coupling to a pertussis toxin-sensitive inhibitory G protein. Mol. Pharmacol. 69, 5–10.
Fujino, H., Salvi, S., and Regan, J. W. (2005). Differential regulation of phosphorylation of the cAMP response element-binding protein after activation of EP2 and EP4 prostanoid receptors by prostaglandin E2. Mol. Pharmacol. 68, 251–259.
Fujino, H., West, K. A., and Regan, J. W. (2002). Phosphorylation of glycogen synthase kinase-3 and stimulation of T-cell factor signaling following activation of EP2 and EP4 prostanoid receptors by prostaglandin E2. J. Biol. Chem. 277, 2614–2619.
Fujino, H., Xu, W., and Regan, J. W. (2003). Prostaglandin E2 induced functional expression of early growth response factor-1 by EP4, but not EP2, prostanoid receptors via the phosphatidylinositol 3-kinase and extracellular signal-regulated kinases. J. Biol. Chem. 278, 12151–12156.
Fukuhara, S., Sakurai, A., Sano, H., Yamagishi, A., Somekawa, S., Takakura, N., et al. (2005). Cyclic AMP potentiates vascular endothelial cadherin-mediated cell-cell contact to enhance endothelial barrier function through an Epac-Rap1 signaling pathway. Mol. Cell. Biol. 25, 136–146.
Gardenfors, F., Nilsson, A., Ungerstedt, U., and Nordstrom, C. H. (2004). Adverse biochemical and physiological effects of prostacyclin in experimental brain oedema. Acta Anaesthesiol. Scand. 48, 1316–1321.
Gautier, S., Ouk, T., Petrault, O., Caron, J., and Bordet, R. (2009). Neutrophils contribute to intracerebral haemorrhages after treatment with recombinant tissue plasminogen activator following cerebral ischaemia. Br. J. Pharmacol. 156, 673–679.
Gekel, I., and Neher, E. (2008). Application of an Epac activator enhances neurotransmitter release at excitatory central synapses. J. Neurosci. 28, 7991–8002.
Gelir, E., Arslan, S. O., Sayan, H., and Pinar, L. (2005). Effect of rapid-eye-movement sleep deprivation on rat hypothalamic prostaglandins. Prostaglandins Leukot. Essent. Fatty Acids 73, 391–396.
Gelosa, P., Ballerio, R., Banfi, C., Nobili, E., Gianella, A., Pignieri, A., et al. (2010). Terutroban, a thromboxane/prostaglandin endoperoxide receptor antagonist, increases survival in stroke-prone rats by preventing systemic inflammation and endothelial dysfunction: comparison with aspirin and rosuvastatin. J. Pharmacol. Exp. Ther. 334, 199–205.
Gendron, T. F., Brunette, E., Tauskela, J. S., and Morley, P. (2005). The dual role of prostaglandin E(2) in excitotoxicity and preconditioning-induced neuroprotection. Eur. J. Pharmacol. 517, 17–27.
Genovese, T., Esposito, E., Mazzon, E., Di Paola, R., Muia, C., Meli, R., et al. (2008). Effect of cyclopentanone prostaglandin 15-deoxy-delta12,14PGJ2 on early functional recovery from experimental spinal cord injury. Shock 30, 142–152.
Gerlo, S., Verdood, P., and Kooijman, R. (2010). Modulation of cytokine production by cyclic adenosine monophosphate analogs in human leukocytes. J. Interferon Cytokine Res. 30, 883–891.
Gong, C., Boulis, N., Qian, J., Turner, D. E., Hoff, J. T., and Keep, R. F. (2001). Intracerebral hemorrhage-induced neuronal death. Neurosurgery 48, 875–882; discussion 882–873.
Gotoh, M., Kitanaka, J., Hirasawa, Y., Kondo, K., and Baba, A. (1994). Desensitization of prostaglandin F2 alpha receptor-mediated phosphoinositide hydrolysis in cultured rat astrocytes. Neurochem. Res. 19, 679–685.
Grill, M., Heinemann, A., Hoefler, G., Peskar, B. A., and Schuligoi, R. (2008). Effect of endotoxin treatment on the expression and localization of spinal cyclooxygenase, prostaglandin synthases, and PGD2 receptors. J. Neurochem. 104, 1345–1357.
Hammad, H., De Heer, H. J., Soullie, T., Hoogsteden, H. C., Trottein, F., and Lambrecht, B. N. (2003). Prostaglandin D2 inhibits airway dendritic cell migration and function in steady state conditions by selective activation of the D prostanoid receptor 1. J. Immunol. 171, 3936–3940.
Hao, C. M., and Breyer, M. D. (2007). Physiologic and pathophysiologic roles of lipid mediators in the kidney. Kidney Int. 71, 1105–1115.
Hasegawa, H., Negishi, M., and Ichikawa, A. (1996). Two isoforms of the prostaglandin E receptor EP3 subtype different in agonist-independent constitutive activity. J. Biol. Chem. 271, 1857–1860.
Hata, A. N., and Breyer, R. M. (2004). Pharmacology and signaling of prostaglandin receptors: multiple roles in inflammation and immune modulation. Pharmacol. Ther. 103, 147–166.
Hatoum, O. A., Gauthier, K. M., Binion, D. G., Miura, H., Telford, G., Otterson, M. F., et al. (2005). Novel mechanism of vasodilation in inflammatory bowel disease. Arterioscler. Thromb. Vasc. Biol. 25, 2355–2361.
Hayaishi, O. (2002). Molecular genetic studies on sleep-wake regulation, with special emphasis on the prostaglandin D2 system. J. Appl. Physiol. 92, 863–868.
Heaslip, R. J., and Sickels, B. D. (1989). Evidence that prostaglandins can contract the rat aorta via a novel protein kinase C-dependent mechanism. J. Pharmacol. Exp. Ther. 250, 44–51.
Hebert, R. L., O’Connor, T., Neville, C., Burns, K. D., Laneuville, O., and Peterson, L. N. (1998). Prostanoid signaling, localization, and expression of IP receptors in rat thick ascending limb cells. Am. J. Physiol. 275, F904–F914.
Heo, J. H., Lucero, J., Abumiya, T., Koziol, J. A., Copeland, B. R., and Del Zoppo, G. J. (1999). Matrix metalloproteinases increase very early during experimental focal cerebral ischemia. J. Cereb. Blood Flow Metab. 19, 624–633.
Hickenbottom, S. L., Grotta, J. C., Strong, R., Denner, L. A., and Aronowski, J. (1999). Nuclear factor-kappaB and cell death after experimental intracerebral hemorrhage in rats. Stroke 30, 2472–2477; discussion 2477–2478.
Hirai, H., Tanaka, K., Yoshie, O., Ogawa, K., Kenmotsu, K., Takamori, Y., et al. (2001). Prostaglandin D2 selectively induces chemotaxis in T helper type 2 cells, eosinophils, and basophils via seven-transmembrane receptor CRTH2. J. Exp. Med. 193, 255–261.
Hirata, M., Hayashi, Y., Ushikubi, F., Yokota, Y., Kageyama, R., Nakanishi, S., et al. (1991). Cloning and expression of cDNA for a human thromboxane A2 receptor. Nature 349, 617–620.
Hirata, M., Kakizuka, A., Aizawa, M., Ushikubi, F., and Narumiya, S. (1994a). Molecular characterization of a mouse prostaglandin D receptor and functional expression of the cloned gene. Proc. Natl. Acad. Sci. U.S.A. 91, 11192–11196.
Hirata, T., Kakizuka, A., Ushikubi, F., Fuse, I., Okuma, M., and Narumiya, S. (1994b). Arg60 to Leu mutation of the human thromboxane A2 receptor in a dominantly inherited bleeding disorder. J. Clin. Invest. 94, 1662–1667.
Hirata, T., Ushikubi, F., Kakizuka, A., Okuma, M., and Narumiya, S. (1996). Two thromboxane A2 receptor isoforms in human platelets. Opposite coupling to adenylyl cyclase with different sensitivity to Arg60 to Leu mutation. J. Clin. Invest. 97, 949–956.
Honda, A., Sugimoto, Y., Namba, T., Watabe, A., Irie, A., Negishi, M., et al. (1993). Cloning and expression of a cDNA for mouse prostaglandin E receptor EP2 subtype. J. Biol. Chem. 268, 7759–7762.
Honma, S., Saika, M., Ohkubo, S., Kurose, H., and Nakahata, N. (2006). Thromboxane A2 receptor-mediated G12/13-dependent glial morphological change. Eur. J. Pharmacol. 545, 100–108.
Hsu, K. S., Huang, C. C., Kan, W. M., and Gean, P. W. (1996). TXA2 agonists inhibit high-voltage-activated calcium channels in rat hippocampal CA1 neurons. Am. J. Physiol. 271, C1269–C1277.
Hsu, K. S., and Kan, W. M. (1996). Thromboxane A2 agonist modulation of excitatory synaptic transmission in the rat hippocampal slice. Br. J. Pharmacol. 118, 2220–2227.
Hua, Y., Xi, G., Keep, R. F., and Hoff, J. T. (2000). Complement activation in the brain after experimental intracerebral hemorrhage. J. Neurosurg. 92, 1016–1022.
Huang, F. P., Xi, G., Keep, R. F., Hua, Y., Nemoianu, A., and Hoff, J. T. (2002). Brain edema after experimental intracerebral hemorrhage: role of hemoglobin degradation products. J. Neurosurg. 96, 287–293.
Ikeda-Matsuo, Y., Tanji, H., Narumiya, S., and Sasaki, Y. (2011). Inhibition of prostaglandin E2 EP3 receptors improves stroke injury via anti-inflammatory and anti-apoptotic mechanisms. J. Neuroimmunol. 238, 34–43.
Ikeda-Matsuo, Y., Tanji, H., Ota, A., Hirayama, Y., Uematsu, S., Akira, S., et al. (2010). Microsomal prostaglandin E synthase-1 contributes to ischaemic excitotoxicity through prostaglandin E2 EP3 receptors. Br. J. Pharmacol. 160, 847–859.
Irie, A., Sugimoto, Y., Namba, T., Asano, T., Ichikawa, A., and Negishi, M. (1994). The C-terminus of the prostaglandin-E-receptor EP3 subtype is essential for activation of GTP-binding protein. Eur. J. Biochem. 224, 161–166.
Irie, A., Sugimoto, Y., Namba, T., Harazono, A., Honda, A., Watabe, A., et al. (1993). Third isoform of the prostaglandin-E-receptor EP3 subtype with different C-terminal tail coupling to both stimulation and inhibition of adenylate cyclase. Eur. J. Biochem. 217, 313–318.
Ishikawa, T. O., Tamai, Y., Rochelle, J. M., Hirata, M., Namba, T., Sugimoto, Y., et al. (1996). Mapping of the genes encoding mouse prostaglandin D, E, and F and prostacyclin receptors. Genomics 32, 285–288.
Ito, S., Narumiya, S., and Hayaishi, O. (1989). Prostaglandin D2: a biochemical perspective. Prostaglandins Leukot. Essent. Fatty Acids 37, 219–234.
Jiang, J., Ganesh, T., Du, Y., Quan, Y., Serrano, G., Qui, M., et al. (2012). Small molecule antagonist reveals seizure-induced mediation of neuronal injury by prostaglandin E2 receptor subtype EP2. Proc. Natl. Acad. Sci. U.S.A. 109, 3149–3154.
Jiang, J., Ganesh, T., Du, Y., Thepchatri, P., Rojas, A., Lewis, I., et al. (2010). Neuroprotection by selective allosteric potentiators of the EP2 prostaglandin receptor. Proc. Natl. Acad. Sci. U.S.A. 107, 2307–2312.
Jovanovic, N., Pavlovic, M., Mircevski, V., Du, Q., and Jovanovic, A. (2006). An unexpected negative inotropic effect of prostaglandin F2α in the rat heart. Prostaglandins Other Lipid Mediat. 80, 110–119.
Justicia, C., Panes, J., Sole, S., Cervera, A., Deulofeu, R., Chamorro, A., et al. (2003). Neutrophil infiltration increases matrix metalloproteinase-9 in the ischemic brain after occlusion/reperfusion of the middle cerebral artery in rats. J. Cereb. Blood Flow Metab. 23, 1430–1440.
Jyoti, A., Sethi, P., and Sharma, D. (2009). Curcumin protects against electrobehavioral progression of seizures in the iron-induced experimental model of epileptogenesis. Epilepsy Behav. 14, 300–308.
Kandasamy, S. B., and Hunt, W. A. (1990). Arachidonic acid and prostaglandins enhance potassium-stimulated calcium influx into rat brain synaptosomes. Neuropharmacology 29, 825–829.
Kaneko, M., and Takahashi, T. (2004). Presynaptic mechanism underlying cAMP-dependent synaptic potentiation. J. Neurosci. 24, 5202–5208.
Kang, G., Joseph, J. W., Chepurny, O. G., Monaco, M., Wheeler, M. B., Bos, J. L., et al. (2003). Epac-selective cAMP analog 8-pCPT-2′-O-Me-cAMP as a stimulus for Ca2+-induced Ca2+ release and exocytosis in pancreatic beta-cells. J. Biol. Chem. 278, 8279–8285.
Katsuyama, M., Nishigaki, N., Sugimoto, Y., Morimoto, K., Negishi, M., Narumiya, S., et al. (1995). The mouse prostaglandin E receptor EP2 subtype: cloning, expression, and northern blot analysis. FEBS Lett. 372, 151–156.
Katsuyama, M., Sugimoto, Y., Namba, T., Irie, A., Negishi, M., Narumiya, S., et al. (1994). Cloning and expression of a cDNA for the human prostacyclin receptor. FEBS Lett. 344, 74–78.
Kawano, T., Anrather, J., Zhou, P., Park, L., Wang, G., Frys, K. A., et al. (2006). Prostaglandin E2 EP1 receptors: downstream effectors of COX-2 neurotoxicity. Nat. Med. 12, 225–229.
Kawasaki, H., Springett, G. M., Mochizuki, N., Toki, S., Nakaya, M., Matsuda, M., et al. (1998). A family of cAMP-binding proteins that directly activate Rap1. Science 282, 2275–2279.
Kiermayer, S., Biondi, R. M., Imig, J., Plotz, G., Haupenthal, J., Zeuzem, S., et al. (2005). Epac activation converts cAMP from a proliferative into a differentiation signal in PC12 cells. Mol. Biol. Cell 16, 5639–5648.
Kikkawa, Y., Kameda, K., Hirano, M., Sasaki, T., and Hirano, K. (2010). Impaired feedback regulation of the receptor activity and the myofilament Ca2+ sensitivity contributes to increased vascular reactiveness after subarachnoid hemorrhage. J. Cereb. Blood Flow Metab. 30, 1637–1650.
Kim, H. J., Chung, J. I., Lee, S. H., Jung, Y. S., Moon, C. H., and Baik, E. J. (2008). Involvement of endogenous prostaglandin F2alpha on kainic acid-induced seizure activity through FP receptor: the mechanism of proconvulsant effects of COX-2 inhibitors. Brain Res. 1193, 153–161.
Kim, Y. T., Moon, S. K., Maruyama, T., Narumiya, S., and Doré, S. (2012). Prostaglandin FP receptor inhibitor reduces ischemic brain damage and neurotoxicity. Neurobiol. Dis. 48, 58–65.
Kiriyama, M., Ushikubi, F., Kobayashi, T., Hirata, M., Sugimoto, Y., and Narumiya, S. (1997). Ligand binding specificities of the eight types and subtypes of the mouse prostanoid receptors expressed in Chinese hamster ovary cells. Br. J. Pharmacol. 122, 217–224.
Kishimoto, K., Li, R. C., Zhang, J., Klaus, J. A., Kibler, K. K., Doré, S., et al. (2010). Cytosolic phospholipase A2 alpha amplifies early cyclooxygenase-2 expression, oxidative stress and MAP kinase phosphorylation after cerebral ischemia in mice. J. Neuroinflammation 7, 42.
Kitanaka, J., Hashimoto, H., Sugimoto, Y., Sawada, M., Negishi, M., Suzumura, A., et al. (1995). cDNA cloning of a thromboxane A2 receptor from rat astrocytes. Biochim. Biophys. Acta 1265, 220–223.
Kitanaka, J., Hasimoto, H., Sugimoto, Y., Negishi, M., Aino, H., Gotoh, M., et al. (1994). Cloning and expression of a cDNA for rat prostaglandin F2α receptor. Prostaglandins 48, 31–41.
Kitanaka, J., Ishibashi, T., and Baba, A. (1993). Phloretin as an antagonist of prostaglandin F2 alpha receptor in cultured rat astrocytes. J. Neurochem. 60, 704–708.
Kitanaka, J., Onoe, H., and Baba, A. (1991). Astrocytes possess prostaglandin F2 alpha receptors coupled to phospholipase C. Biochem. Biophys. Res. Commun. 178, 946–952.
Kitanaka, J., Takuma, K., Kondo, K., and Baba, A. (1996). Prostanoid receptor-mediated calcium signaling in cultured rat astrocytes. Jpn. J. Pharmacol. 71, 85–87.
Knock, G. A., De Silva, A. S., Snetkov, V. A., Siow, R., Thomas, G. D., Shiraishi, M., et al. (2005). Modulation of PGF2α- and hypoxia-induced contraction of rat intrapulmonary artery by p38 MAPK inhibition: a nitric oxide-dependent mechanism. Am. J. Physiol. Lung Cell Mol. Physiol. 289, L1039–L1048.
Kobayashi, T., and Narumiya, S. (2002). Function of prostanoid receptors: studies on knockout mice. Prostaglandins Other Lipid Mediat. 68–69, 557–573.
Koch, K. A., Wessale, J. L., Moreland, R., Reinhart, G. A., and Cox, B. F. (2005). Effects of BW245C, a prostaglandin DP receptor agonist, on systemic and regional haemodynamics in the anaesthetized rat. Clin. Exp. Pharmacol. Physiol. 32, 931–935.
Koide, M., Nystoriak, M. A., Brayden, J. E., and Wellman, G. C. (2011). Impact of subarachnoid hemorrhage on local and global calcium signaling in cerebral artery myocytes. Acta Neurochir. Suppl. 110, 145–150.
Kollmar, R., Staykov, D., Dorfler, A., Schellinger, P. D., Schwab, S., and Bardutzky, J. (2010). Hypothermia reduces perihemorrhagic edema after intracerebral hemorrhage. Stroke 41, 1684–1689.
Kong, D. L., Prough, D. S., Whitley, J. M., Taylor, C., Vines, S., Deal, D. D., et al. (1991). Hemorrhage and intracranial hypertension in combination increase cerebral production of thromboxane A2. Crit. Care Med. 19, 532–538.
Konger, R. L., Brouxhon, S., Partillo, S., Vanbuskirk, J., and Pentland, A. P. (2005). The EP3 receptor stimulates ceramide and diacylglycerol release and inhibits growth of primary keratinocytes. Exp. Dermatol. 14, 914–922.
Konger, R. L., Malaviya, R., and Pentland, A. P. (1998). Growth regulation of primary human keratinocytes by prostaglandin E receptor EP2 and EP3 subtypes. Biochim. Biophys. Acta 1401, 221–234.
Kostic, V. S., Djuricic, B. M., and Mrsulja, B. B. (1984). Cerebrospinal fluid prostaglandin F2 alpha in stroke patients: no relationship to the degree of neurological deficit. Eur. Neurol. 23, 291–295.
Kotani, M., Tanaka, I., Ogawa, Y., Suganami, T., Matsumoto, T., Muro, S., et al. (2000). Multiple signal transduction pathways through two prostaglandin E receptor EP3 subtype isoforms expressed in human uterus. J. Clin. Endocrinol. Metab. 85, 4315–4322.
Kotani, M., Tanaka, I., Ogawa, Y., Usui, T., Mori, K., Ichikawa, A., et al. (1995). Molecular cloning and expression of multiple isoforms of human prostaglandin E receptor EP3 subtype generated by alternative messenger RNA splicing: multiple second messenger systems and tissue-specific distributions. Mol. Pharmacol. 48, 869–879.
Kotani, M., Tanaka, I., Ogawa, Y., Usui, T., Tamura, N., Mori, K., et al. (1997). Structural organization of the human prostaglandin EP3 receptor subtype gene (PTGER3). Genomics 40, 425–434.
Kozasa, T., Jiang, X., Hart, M. J., Sternweis, P. M., Singer, W. D., Gilman, A. G., et al. (1998). p115 RhoGEF, a GTPase activating protein for Galpha12 and Galpha13. Science 280, 2109–2111.
Lapchak, P. A., Chapman, D. F., and Zivin, J. A. (2000). Metalloproteinase inhibition reduces thrombolytic (tissue plasminogen activator)-induced hemorrhage after thromboembolic stroke. Stroke 31, 3034–3040.
Laugwitz, K. L., Allgeier, A., Offermanns, S., Spicher, K., Van Sande, J., Dumont, J. E., et al. (1996). The human thyrotropin receptor: a heptahelical receptor capable of stimulating members of all four G protein families. Proc. Natl. Acad. Sci. U.S.A. 93, 116–120.
Li, D. Y., Varma, D. R., Chatterjee, T. K., Fernandez, H., Abran, D., and Chemtob, S. (1993). Fewer PGE2 and PGF2 alpha receptors in brain synaptosomes of newborn than of adult pigs. J. Pharmacol. Exp. Ther. 267, 1292–1297.
Li, J., Liang, X., Wang, Q., Breyer, R. M., McCullough, L., and Andreasson, K. (2008). Misoprostol, an anti-ulcer agent and PGE2 receptor agonist, protects against cerebral ischemia. Neurosci. Lett. 438, 210–215.
Li, R. C., Saleem, S., Zhen, G., Cao, W., Zhuang, H., Lee, J., et al. (2009). Heme-hemopexin complex attenuates neuronal cell death and stroke damage. J. Cereb. Blood Flow Metab. 29, 953–964.
Li, X., Cudaback, E., Breyer, R. M., Montine, K. S., Keene, C. D., and Montine, T. J. (2012). Eicosanoid receptor subtype-mediated opposing regulation of TLR-stimulated expression of astrocyte glial-derived neurotrophic factor. FASEB J. 26, 3075–3083.
Liang, X., Wu, L., Hand, T., and Andreasson, K. (2005). Prostaglandin D2 mediates neuronal protection via the DP1 receptor. J. Neurochem. 92, 477–486.
Liang, Y., Woodward, D. F., Guzman, V. M., Li, C., Scott, D. F., Wang, J. W., et al. (2008). Identification and pharmacological characterization of the prostaglandin FP receptor and FP receptor variant complexes. Br. J. Pharmacol. 154, 1079–1093.
Liu, D., Wu, L., Breyer, R., Mattson, M. P., and Andreasson, K. (2005). Neuroprotection by the PGE2 EP2 receptor in permanent focal cerebral ischemia. Ann. Neurol. 57, 758–761.
Lorenowicz, M. J., Fernandez-Borja, M., and Hordijk, P. L. (2007). cAMP signaling in leukocyte transendothelial migration. Arterioscler. Thromb. Vasc. Biol. 27, 1014–1022.
Lorenowicz, M. J., van Gils, J., De Boer, M., Hordijk, P. L., and Fernandez-Borja, M. (2006). Epac1-Rap1 signaling regulates monocyte adhesion and chemotaxis. J. Leukoc. Biol. 80, 1542–1552.
Macdonald, R. L., and Weir, B. K. (1991). A review of hemoglobin and the pathogenesis of cerebral vasospasm. Stroke 22, 971–982.
Manawadu, D., Jeerakathil, T., Roy, A., Orwaard-Wong, K., and Butcher, K. (2010). Blood pressure management in acute intracerebral haemorrhage guidelines are poorly implemented in clinical practice. Clin. Neurol. Neurosurg. 112, 858–864.
Masada, T., Hua, Y., Xi, G., Yang, G. Y., Hoff, J. T., and Keep, R. F. (2001). Attenuation of intracerebral hemorrhage and thrombin-induced brain edema by overexpression of interleukin-1 receptor antagonist. J. Neurosurg. 95, 680–686.
Matsugi, T., Kageyama, M., Nishimura, K., Giles, H., and Shirasawa, E. (1995). Selective prostaglandin D2 receptor stimulation elicits ocular hypotensive effects in rabbits and cats. Eur. J. Pharmacol. 275, 245–250.
Matsumura, K., Watanabe, Y., Imai-Matsumura, K., Connolly, M., Koyama, Y., and Onoe, H. (1992). Mapping of prostaglandin E2 binding sites in rat brain using quantitative autoradiography. Brain Res. 581, 292–298.
Matsuoka, T., Hirata, M., Tanaka, H., Takahashi, Y., Murata, T., Kabashima, K., et al. (2000). Prostaglandin D2 as a mediator of allergic asthma. Science 287, 2013–2017.
Mayne, M., Fotheringham, J., Yan, H. J., Power, C., Del Bigio, M. R., Peeling, J., et al. (2001a). Adenosine A2A receptor activation reduces proinflammatory events and decreases cell death following intracerebral hemorrhage. Ann. Neurol. 49, 727–735.
Mayne, M., Ni, W., Yan, H. J., Xue, M., Johnston, J. B., Del Bigio, M. R., et al. (2001b). Antisense oligodeoxynucleotide inhibition of tumor necrosis factor-alpha expression is neuroprotective after intracerebral hemorrhage. Stroke 32, 240–248.
McCullough, L., Wu, L., Haughey, N., Liang, X., Hand, T., Wang, Q., et al. (2004). Neuroprotective function of the PGE2 EP2 receptor in cerebral ischemia. J. Neurosci. 24, 257–268.
Miggin, S. M., and Kinsella, B. T. (1998). Expression and tissue distribution of the mRNAs encoding the human thromboxane A2 receptor (TP) alpha and beta isoforms. Biochim. Biophys. Acta 1425, 543–559.
Miggin, S. M., Lawler, O. A., and Kinsella, B. T. (2003). Palmitoylation of the human prostacyclin receptor. Functional implications of palmitoylation and isoprenylation. J. Biol. Chem. 278, 6947–6958.
Mizoguchi, A., Eguchi, N., Kimura, K., Kiyohara, Y., Qu, W. M., Huang, Z. L., et al. (2001). Dominant localization of prostaglandin D receptors on arachnoid trabecular cells in mouse basal forebrain and their involvement in the regulation of non-rapid eye movement sleep. Proc. Natl. Acad. Sci. U.S.A. 98, 11674–11679.
Mohri, I., Kadoyama, K., Kanekiyo, T., Sato, Y., Kagitani-Shimono, K., Saito, Y., et al. (2007). Hematopoietic prostaglandin D synthase and DP1 receptor are selectively upregulated in microglia and astrocytes within senile plaques from human patients and in a mouse model of Alzheimer disease. J. Neuropathol. Exp. Neurol. 66, 469–480.
Moncada, S., Herman, A. G., Higgs, E. A., and Vane, J. R. (1977a). Differential formation of prostacyclin (PGX or PGI2) by layers of the arterial wall. An explanation for the anti-thrombotic properties of vascular endothelium. Thromb Res 11, 323–344.
Moncada, S., Vane, J. R., and Whittle, B. J. (1977b). Relative potency of prostacyclin, prostaglandin E1 and D2 as inhibitors of platelet aggregation in several species [proceedings]. J. Physiol. (Lond.) 273, 2P–4P.
Monneret, G., Gravel, S., Diamond, M., Rokach, J., and Powell, W. S. (2001). Prostaglandin D2 is a potent chemoattractant for human eosinophils that acts via a novel DP receptor. Blood 98, 1942–1948.
Morgenweck, J., Abdel-Aleem, O. S., McNamara, K. C., Donahue, R. R., Badr, M. Z., and Taylor, B. K. (2010). Activation of peroxisome proliferator-activated receptor gamma in brain inhibits inflammatory pain, dorsal horn expression of Fos, and local edema. Neuropharmacology 58, 337–345.
Moxon-Emre, I., and Schlichter, L. C. (2011). Neutrophil depletion reduces blood-brain barrier breakdown, axon injury, and inflammation after intracerebral hemorrhage. J. Neuropathol. Exp. Neurol. 70, 218–235.
Muck, S., Weber, A. A., Meyer-Kirchrath, J., and Schror, K. (1998). The bovine thromboxane A2 receptor: molecular cloning, expression, and functional characterization. Naunyn Schmiedebergs Arch. Pharmacol. 357, 10–16.
Murray, A. J., and Shewan, D. A. (2008). Epac mediates cyclic AMP-dependent axon growth, guidance and regeneration. Mol. Cell. Neurosci. 38, 578–588.
Nagata, K., Hirai, H., Tanaka, K., Ogawa, K., Aso, T., Sugamura, K., et al. (1999a). CRTH2, an orphan receptor of T-helper-2-cells, is expressed on basophils and eosinophils and responds to mast cell-derived factor(s). FEBS Lett. 459, 195–199.
Nagata, K., Tanaka, K., Ogawa, K., Kemmotsu, K., Imai, T., Yoshie, O., et al. (1999b). Selective expression of a novel surface molecule by human Th2 cells in vivo. J. Immunol. 162, 1278–1286.
Nakamura, K., Kaneko, T., Yamashita, Y., Hasegawa, H., Katoh, H., and Negishi, M. (2000). Immunohistochemical localization of prostaglandin EP3 receptor in the rat nervous system. J. Comp. Neurol. 421, 543–569.
Nakamura, Y., Nakamura, K., and Morrison, S. F. (2009). Different populations of prostaglandin EP3 receptor-expressing preoptic neurons project to two fever-mediating sympathoexcitatory brain regions. Neuroscience 161, 614–620.
Namba, T., Oida, H., Sugimoto, Y., Kakizuka, A., Negishi, M., Ichikawa, A., et al. (1994). cDNA cloning of a mouse prostacyclin receptor. Multiple signaling pathways and expression in thymic medulla. J. Biol. Chem. 269, 9986–9992.
Namba, T., Sugimoto, Y., Hirata, M., Hayashi, Y., Honda, A., Watabe, A., et al. (1992). Mouse thromboxane A2 receptor: cDNA cloning, expression and northern blot analysis. Biochem. Biophys. Res. Commun. 184, 1197–1203.
Namiranian, K., Koehler, R. C., Sapirstein, A., and Doré, S. (2005). Stroke outcomes in mice lacking the genes for neuronal heme oxygenase-2 and nitric oxide synthase. Curr. Neurovasc. Res. 2, 23–27.
Narumiya, S. (2009). Prostanoids and inflammation: a new concept arising from receptor knockout mice. J. Mol. Med. (Berl.) 87, 1015–1022.
Narumiya, S., and Fitzgerald, G. A. (2001). Genetic and pharmacological analysis of prostanoid receptor function. J. Clin. Invest. 108, 25–30.
Narumiya, S., Ogorochi, T., Nakao, K., and Hayaishi, O. (1982). Prostaglandin D2 in rat brain, spinal cord and pituitary: basal level and regional distribution. Life Sci. 31, 2093–2103.
Narumiya, S., Sugimoto, Y., and Ushikubi, F. (1999). Prostanoid receptors: structures, properties, and functions. Physiol. Rev. 79, 1193–1226.
Narumiya, S., and Toda, N. (1985). Different responsiveness of prostaglandin D2-sensitive systems to prostaglandin D2 and its analogues. Br. J. Pharmacol. 85, 367–375.
Narumiya, S., Ushikubi, F., Nakajima, M., Hirata, M., and Okuma, M. (1991). Purification and characterization of the human platelet TXA2/PGH2 receptor. Adv. Prostaglandin Thromboxane Leukot. Res. 21A, 339–346.
Nishigaki, N., Negishi, M., and Ichikawa, A. (1996). Two Gs-coupled prostaglandin E receptor subtypes, EP2 and EP4, differ in desensitization and sensitivity to the metabolic inactivation of the agonist. Mol. Pharmacol. 50, 1031–1037.
Obal, F. Jr., and Krueger, J. M. (2003). Biochemical regulation of non-rapid-eye-movement sleep. Front. Biosci. 8, d520–d550.
Offermanns, S., Hu, Y. H., and Simon, M. I. (1996). Galpha12 and galpha13 are phosphorylated during platelet activation. J. Biol. Chem. 271, 26044–26048.
Ogorochi, T., Narumiya, S., Mizuno, N., Yamashita, K., Miyazaki, H., and Hayaishi, O. (1984). Regional distribution of prostaglandins D2, E2, and F2 alpha and related enzymes in postmortem human brain. J. Neurochem. 43, 71–82.
Oida, H., Hirata, M., Sugimoto, Y., Ushikubi, F., Ohishi, H., Mizuno, N., et al. (1997). Expression of messenger RNA for the prostaglandin D receptor in the leptomeninges of the mouse brain. FEBS Lett. 417, 53–56.
Oida, H., Namba, T., Sugimoto, Y., Ushikubi, F., Ohishi, H., Ichikawa, A., et al. (1995). In situ hybridization studies of prostacyclin receptor mRNA expression in various mouse organs. Br. J. Pharmacol. 116, 2828–2837.
Oliveira, M. S., Furian, A. F., Rambo, L. M., Ribeiro, L. R., Royes, L. F., Ferreira, J., et al. (2008). Modulation of pentylenetetrazol-induced seizures by prostaglandin E2 receptors. Neuroscience 152, 1110–1118.
Orie, N. N., and Clapp, L. H. (2011). Role of prostanoid IP and EP receptors in mediating vasorelaxant responses to PGI2 analogues in rat tail artery: Evidence for Gi/o modulation via EP3 receptors. Eur. J. Pharmacol. 654, 258–265.
Peeling, J., Yan, H. J., Chen, S. G., Campbell, M., and Del Bigio, M. R. (1998). Protective effects of free radical inhibitors in intracerebral hemorrhage in rat. Brain Res. 795, 63–70.
Penn, R. B., Pascual, R. M., Kim, Y. M., Mundell, S. J., Krymskaya, V. P., Panettieri, R. A. Jr., et al. (2001). Arrestin specificity for G protein-coupled receptors in human airway smooth muscle. J. Biol. Chem. 276, 32648–32656.
Pickard, J. D., Walker, V., Brandt, L., Zygmunt, S., and Smythe, J. (1994). Effect of intraventricular haemorrhage and rebleeding following subarachnoid haemorrhage on CSF eicosanoids. Acta Neurochir. (Wien) 129, 152–157.
Qian, Y. M., Jones, R. L., Chan, K. M., Stock, A. I., and Ho, J. K. (1994). Potent contractile actions of prostanoid EP3-receptor agonists on human isolated pulmonary artery. Br. J. Pharmacol. 113, 369–374.
Qing, W. G., Dong, Y. Q., Ping, T. Q., Lai, L. G., Fang, L. D., Min, H. W., et al. (2009). Brain edema after intracerebral hemorrhage in rats: the role of iron overload and aquaporin 4. J. Neurosurg. 110, 462–468.
Raychowdhury, M. K., Yukawa, M., Collins, L. J., McGrail, S. H., Kent, K. C., and Ware, J. A. (1994). Alternative splicing produces a divergent cytoplasmic tail in the human endothelial thromboxane A2 receptor. J. Biol. Chem. 269, 19256–19261.
Regan, J. W., Bailey, T. J., Pepperl, D. J., Pierce, K. L., Bogardus, A. M., Donello, J. E., et al. (1994). Cloning of a novel human prostaglandin receptor with characteristics of the pharmacologically defined EP2 subtype. Mol. Pharmacol. 46, 213–220.
Rehni, A. K., and Singh, T. G. (2012). Selenium induced anticonvulsant effect: a potential role of prostaglandin E(1) receptor activation linked mechanism. J. Trace Elem. Med. Biol. doi:10.1016/j.jtemb.2012.05.001
Richards, J. A., and Brueggemeier, R. W. (2003). Prostaglandin E2 regulates aromatase activity and expression in human adipose stromal cells via two distinct receptor subtypes. J. Clin. Endocrinol. Metab. 88, 2810–2816.
Roger, V. L., Go, A. S., Lloyd-Jones, D. M., Benjamin, E. J., Berry, J. D., Borden, W. B., et al. (2012). Heart disease and stroke statistics – 2012 update: a report from the American Heart Association. Circulation 125, e2–e220.
Rondaij, M. G., Sellink, E., Gijzen, K. A., Ten Klooster, J. P., Hordijk, P. L., van Mourik, J. A., et al. (2004). Small GTP-binding protein Ral is involved in cAMP-mediated release of von Willebrand factor from endothelial cells. Arterioscler. Thromb. Vasc. Biol. 24, 1315–1320.
Saleem, S., Ahmad, A. S., Maruyama, T., Narumiya, S., and Doré, S. (2009a). PGF(2alpha) FP receptor contributes to brain damage following transient focal brain ischemia. Neurotox. Res. 15, 62–70.
Saleem, S., Kim, Y. T., Maruyama, T., Narumiya, S., and Doré, S. (2009b). Reduced acute brain injury in PGE2 EP3 receptor-deficient mice after cerebral ischemia. J. Neuroimmunol. 208, 87–93.
Saleem, S., Shah, Z. A., Urade, Y., and Doré, S. (2009c). Lipocalin-prostaglandin D synthase is a critical beneficial factor in transient and permanent focal cerebral ischemia. Neuroscience 160, 248–254.
Saleem, S., Li, R., Wei, G., and Doré, S. (2007a). Effects of EP1 receptor on cerebral blood flow in the middle cerebral artery occlusion model of stroke in mice. J. Neurosci. Res. 85, 2433–2440.
Saleem, S., Li, R. C., Wei, G., and Doré, S. (2007b). Effects of EP1 receptor on cerebral blood flow in the middle cerebral artery occlusion model of stroke in mice. J. Neurosci. Res. 85, 2433–2440.
Saleem, S., Zhuang, H., De Brum-Fernandes, A. J., Maruyama, T., Narumiya, S., and Doré, S. (2007c). PGD2 DP1 receptor protects brain from ischemia-reperfusion injury. Eur. J. Neurosci. 26, 73–78.
Saleem, S., Shah, Z. A., Maruyama, T., Narumiya, S., and Doré, S. (2010). Neuroprotective properties of prostaglandin I2 IP receptor in focal cerebral ischemia. Neuroscience 170, 317–323.
Saloheimo, P., Juvela, S., Riutta, A., Pyhtinen, J., and Hillbom, M. (2005). Thromboxane and prostacyclin biosynthesis in patients with acute spontaneous intracerebral hemorrhage. Thromb. Res. 115, 367–373.
Sang, N., Zhang, J., Marcheselli, V., Bazan, N. G., and Chen, C. (2005). Postsynaptically synthesized prostaglandin E2 (PGE2) modulates hippocampal synaptic transmission via a presynaptic PGE2 EP2 receptor. J. Neurosci. 25, 9858–9870.
Sanzgiri, R. P., Araque, A., and Haydon, P. G. (1999). Prostaglandin E(2) stimulates glutamate receptor-dependent astrocyte neuromodulation in cultured hippocampal cells. J. Neurobiol. 41, 221–229.
Sarrafzadeh, A., Haux, D., Sakowitz, O., Benndorf, G., Herzog, H., Kuechler, I., et al. (2003). Acute focal neurological deficits in aneurysmal subarachnoid hemorrhage: relation of clinical course, CT findings, and metabolite abnormalities monitored with bedside microdialysis. Stroke 34, 1382–1388.
Satoh, T., Ishikawa, Y., Kataoka, Y., Cui, Y., Yanase, H., Kato, K., et al. (1999). CNS-specific prostacyclin ligands as neuronal survival-promoting factors in the brain. Eur. J. Neurosci. 11, 3115–3124.
Sawyer, N., Cauchon, E., Chateauneuf, A., Cruz, R. P. G., Nicholson, D. W., Metters, K. M., et al. (2002). Molecular pharmacology of the human prostaglandin D2 receptor, CRTH2. Br. J. Pharmacol. 137, 1163–1172.
Schmid, A., Thierauch, K. H., Schleuning, W. D., and Dinter, H. (1995). Splice variants of the human EP3 receptor for prostaglandin E2. Eur. J. Biochem. 228, 23–30.
Schwartz-Bloom, R. D., Cook, T. A., and Yu, X. (1996). Inhibition of GABA-gated chloride channels in brain by the arachidonic acid metabolite, thromboxane A2. Neuropharmacology 35, 1347–1353.
Shenker, A., Goldsmith, P., Unson, C. G., and Spiegel, A. M. (1991). The G protein coupled to the thromboxane A2 receptor in human platelets is a member of the novel Gq family. J. Biol. Chem. 266, 9309–9313.
Shi, G. X., Rehmann, H., and Andres, D. A. (2006). A novel cyclic AMP-dependent Epac-Rit signaling pathway contributes to PACAP38-mediated neuronal differentiation. Mol. Cell. Biol. 26, 9136–9147.
Shi, J., Johansson, J., Woodling, N. S., Wang, Q., Montine, T. J., and Andreasson, K. (2010). The prostaglandin E2 E-prostanoid 4 receptor exerts anti-inflammatory effects in brain innate immunity. J. Immunol. 184, 7207–7218.
Stapleton, P. P., Strong, V. E., Freeman, T. A., Winter, J., Yan, Z., and Daly, J. M. (2004). Gender affects macrophage cytokine and prostaglandin E2 production and PGE2 receptor expression after trauma. J. Surg. Res. 122, 1–7.
Sturzebecher, S., Nieuweboer, B., Matthes, S., and Schillinger, E. (1989). Effects of PGD2, PGE1, and PGI2-analogues on PGDF-release and aggregation of human gelfiltered platelets. Prog. Clin. Biol. Res. 301, 365–369.
Sugimoto, Y., Negishi, M., Hayashi, Y., Namba, T., Honda, A., Watabe, A., et al. (1993). Two isoforms of the EP3 receptor with different carboxyl-terminal domains. Identical ligand binding properties and different coupling properties with Gi proteins. J. Biol. Chem. 268, 2712–2718.
Sugimoto, Y., Shigemoto, R., Namba, T., Negishi, M., Mizuno, N., Narumiya, S., et al. (1994). Distribution of the messenger RNA for the prostaglandin E receptor subtype EP3 in the mouse nervous system. Neuroscience 62, 919–928.
Sugimoto, Y., Yamasaki, A., Segi, E., Tsuboi, K., Aze, Y., Nishimura, T., et al. (1997). Failure of parturition in mice lacking the prostaglandin F receptor. Science 277, 681–683.
Sumii, T., and Lo, E. H. (2002). Involvement of matrix metalloproteinase in thrombolysis-associated hemorrhagic transformation after embolic focal ischemia in rats. Stroke 33, 831–836.
Suzuki, S., Yokoyama, U., Abe, T., Kiyonari, H., Yamashita, N., Kato, Y., et al. (2010). Differential roles of Epac in regulating cell death in neuronal and myocardial cells. J. Biol. Chem. 285, 24248–24259.
Takamatsu, H., Tsukada, H., Watanabe, Y., Cui, Y., Kataoka, Y., Hosoya, T., et al. (2002). Specific ligand for a central type prostacyclin receptor attenuates neuronal damage in a rat model of focal cerebral ischemia. Brain Res. 925, 176–182.
Takasugi, S., Ueda, S., and Matsumoto, K. (1985). Chronological changes in spontaneous intracerebral hematoma – an experimental and clinical study. Stroke 16, 651–658.
Takayama, K., Yuhki, K., Ono, K., Fujino, T., Hara, A., Yamada, T., et al. (2005). Thromboxane A2 and prostaglandin F2α mediate inflammatory tachycardia. Nat. Med. 11, 562–566.
Takechi, H., Matsumura, K., Watanabe, Y., Kato, K., Noyori, R., and Suzuki, M. (1996). A novel subtype of the prostacyclin receptor expressed in the central nervous system. J. Biol. Chem. 271, 5901–5906.
Takemiya, T., Maehara, M., Matsumura, K., Yasuda, S., Sugiura, H., and Yamagata, K. (2006). Prostaglandin E2 produced by late induced COX-2 stimulates hippocampal neuron loss after seizure in the CA3 region. Neurosci. Res. 56, 103–110.
Takemiya, T., Matsumura, K., Sugiura, H., Maehara, M., Yasuda, S., Uematsu, S., et al. (2010). Endothelial microsomal prostaglandin E synthase-1 exacerbates neuronal loss induced by kainate. J. Neurosci. Res. 88, 381–390.
Takemiya, T., Matsumura, K., Sugiura, H., Yasuda, S., Uematsu, S., Akira, S., et al. (2011). Endothelial microsomal prostaglandin E synthase-1 facilitates neurotoxicity by elevating astrocytic Ca2+ levels. Neurochem. Int. 58, 489–496.
Tang, E. H., Libby, P., Vanhoutte, P. M., and Xu, A. (2012). Anti-inflammation therapy by activation of prostaglandin EP4 receptor in cardiovascular and other inflammatory diseases. J. Cardiovasc. Pharmacol. 59, 116–123.
Taniguchi, H., Anacker, C., Suarez-Mier, G. B., Wang, Q., and Andreasson, K. (2011a). Function of prostaglandin E2 EP receptors in the acute outcome of rodent hypoxic ischemic encephalopathy. Neurosci. Lett. 504, 185–190.
Taniguchi, H., Anacker, C., Suarez-Mier, G. B., Wang, Q., and Andreasson, K. (2011b). Function of prostaglandin E(2) EP receptors in the acute outcome of rodent hypoxic ischemic encephalopathy. Neurosci. Lett. 504, 185–190.
Thom, T., Haase, N., Rosamond, W., Howard, V. J., Rumsfeld, J., Manolio, T., et al. (2006). Heart disease and stroke statistics – 2006 update: a report from the American Heart Association Statistics Committee and Stroke Statistics Subcommittee. Circulation 113, e85–e151.
Toh, H., Ichikawa, A., and Narumiya, S. (1995). Molecular evolution of receptors for eicosanoids. FEBS Lett. 361, 17–21.
Urade, Y., and Hayaishi, O. (1999). Prostaglandin D2 and sleep regulation. Biochim. Biophys. Acta 1436, 606–615.
Ushikubi, F., Aiba, Y., Nakamura, K., Namba, T., Hirata, M., Mazda, O., et al. (1993). Thromboxane A2 receptor is highly expressed in mouse immature thymocytes and mediates DNA fragmentation and apoptosis. J. Exp. Med. 178, 1825–1830.
van Kooten, F., Ciabattoni, G., Koudstaal, P. J., Dippel, D. W., and Patrono, C. (1999a). Increased platelet activation in the chronic phase after cerebral ischemia and intracerebral hemorrhage. Stroke 30, 546–549.
van Kooten, F., Ciabattoni, G., Koudstaal, P. J., Grobbee, D. E., Kluft, C., and Patrono, C. (1999b). Increased thromboxane biosynthesis is associated with poststroke dementia. Stroke 30, 1542–1547.
Vane, J. R., and Botting, R. M. (1995). Pharmacodynamic profile of prostacyclin. Am. J. Cardiol. 75, 3A–10A.
Vasilache, A. M., Andersson, J., and Nilsberth, C. (2007). Expression of PGE2 EP3 receptor subtypes in the mouse preoptic region. Neurosci. Lett. 423, 179–183.
Wagner, K. R., Xi, G., Hua, Y., Kleinholz, M., De Courten-Myers, G. M., and Myers, R. E. (1998). Early metabolic alterations in edematous perihematomal brain regions following experimental intracerebral hemorrhage. J. Neurosurg. 88, 1058–1065.
Wagner, K. R., Xi, G., Hua, Y., Zuccarello, M., De Courten-Myers, G. M., Broderick, J. P., et al. (1999). Ultra-early clot aspiration after lysis with tissue plasminogen activator in a porcine model of intracerebral hemorrhage: edema reduction and blood-brain barrier protection. J. Neurosurg. 90, 491–498.
Wang, H., Oestreich, E. A., Maekawa, N., Bullard, T. A., Vikstrom, K. L., Dirksen, R. T., et al. (2005). Phospholipase C epsilon modulates beta-adrenergic receptor-dependent cardiac contraction and inhibits cardiac hypertrophy. Circ. Res. 97, 1305–1313.
Wang, H. C., Lin, W. C., Lin, Y. J., Rau, C. S., Lee, T. H., Chang, W. N., et al. (2011). The association between serum adhesion molecules and outcome in acute spontaneous intracerebral hemorrhage. Crit. Care 15, R284.
Wang, J., and Doré, S. (2007a). Heme oxygenase-1 exacerbates early brain injury after intracerebral haemorrhage. Brain 130, 1643–1652.
Wang, J., and Doré, S. (2007b). Inflammation after intracerebral hemorrhage. J. Cereb. Blood Flow Metab. 27, 894–908.
Wang, J., and Doré, S. (2008). Heme oxygenase 2 deficiency increases brain swelling and inflammation after intracerebral hemorrhage. Neuroscience 155, 1133–1141.
Wang, J., Fields, J., and Doré, S. (2008). The development of an improved preclinical mouse model of intracerebral hemorrhage using double infusion of autologous whole blood. Brain Res. 1222, 214–221.
Wang, J., Zhuang, H., and Doré, S. (2006). Heme oxygenase 2 is neuroprotective against intracerebral hemorrhage. Neurobiol. Dis. 22, 473–476.
Wasserman, J. K., Yang, H., and Schlichter, L. C. (2008). Glial responses, neuron death and lesion resolution after intracerebral hemorrhage in young vs. aged rats. Eur. J. Neurosci. 28, 1316–1328.
Watabe, A., Sugimoto, Y., Honda, A., Irie, A., Namba, T., Negishi, M., et al. (1993). Cloning and expression of cDNA for a mouse EP1 subtype of prostaglandin E receptor. J. Biol. Chem. 268, 20175–20178.
Watanabe, Y., Matsumura, K., Takechi, H., Kato, K., Morii, H., Bjorkman, M., et al. (1999). A novel subtype of prostacyclin receptor in the central nervous system. J. Neurochem. 72, 2583–2592.
Watanabe, Y., Tokumoto, H., Yamashita, A., Narumiya, S., Mizuno, N., and Hayaishi, O. (1985). Specific bindings of prostaglandin D2, E2 and F2 alpha in postmortem human brain. Brain Res. 342, 110–116.
Wei, G., Kibler, K. K., Koehler, R. C., Maruyama, T., Narumiya, S., and Doré, S. (2008). Prostacyclin receptor deletion aggravates hippocampal neuronal loss after bilateral common carotid artery occlusion in mouse. Neuroscience 156, 1111–1117.
Weinberg, E., Zeldich, E., Weinreb, M. M., Moses, O., Nemcovsky, C., and Weinreb, M. (2009). Prostaglandin E2 inhibits the proliferation of human gingival fibroblasts via the EP2 receptor and Epac. J. Cell. Biochem. 108, 207–215.
Whittle, B. J., Moncada, S., Mullane, K., and Vane, J. R. (1983). Platelet and cardiovascular activity of the hydantoin BW245C, a potent prostaglandin analogue. Prostaglandins 25, 205–223.
Willmore, L. J., and Triggs, W. J. (1991). Iron-induced lipid peroxidation and brain injury responses. Int. J. Dev. Neurosci. 9, 175–180.
Wolfe, L. S., and Mamer, O. A. (1975). Measurement of prostaglandin F2alpha levels in human cerebrospinal fluid in normal and pathological conditions. Prostaglandins 9, 183–192.
Woodward, D. F., Jones, R. L., and Narumiya, S. (2011). International Union of Basic and Clinical Pharmacology. LXXXIII: classification of prostanoid receptors, updating 15 years of progress. Pharmacol. Rev. 63, 471–538.
Xi, G., Hua, Y., Bhasin, R. R., Ennis, S. R., Keep, R. F., and Hoff, J. T. (2001). Mechanisms of edema formation after intracerebral hemorrhage: effects of extravasated red blood cells on blood flow and blood-brain barrier integrity. Stroke 32, 2932–2938.
Xi, G., Hua, Y., Keep, R. F., Younger, J. G., and Hoff, J. T. (2002). Brain edema after intracerebral hemorrhage: the effects of systemic complement depletion. Acta Neurochir. Suppl. 81, 253–256.
Xue, M., and Del Bigio, M. R. (2000). Intracerebral injection of autologous whole blood in rats: time course of inflammation and cell death. Neurosci. Lett. 283, 230–232.
Yalcin, M., Cavun, S., Yilmaz, M. S., Cengiz, F., and Savci, V. (2005). Involvement of brain thromboxane A in hypotension induced by haemorrhage in rats. Clin. Exp. Pharmacol. Physiol. 32, 960–967.
Yang, H., Zhang, J., Breyer, R. M., and Chen, C. (2009). Altered hippocampal long-term synaptic plasticity in mice deficient in the PGE2 EP2 receptor. J. Neurochem. 108, 295–304.
Zeynalov, E., Shah, Z. A., Li, R. C., and Doré, S. (2009). Heme oxygenase 1 is associated with ischemic preconditioning-induced protection against brain ischemia. Neurobiol. Dis. 35, 264–269.
Zhang, J., and Rivest, S. (1999). Distribution, regulation and colocalization of the genes encoding the EP2- and EP4-PGE2 receptors in the rat brain and neuronal responses to systemic inflammation. Eur. J. Neurosci. 11, 2651–2668.
Zhao, F., Song, S., Liu, W., Keep, R. F., Xi, G., and Hua, Y. (2011). Red blood cell lysis and brain tissue-type transglutaminase upregulation in a hippocampal model of intracerebral hemorrhage. Acta Neurochir. Suppl. 111, 101–105.
Zhao, X., Sun, G., Zhang, J., Strong, R., Song, W., Gonzales, N., et al. (2007). Hematoma resolution as a target for intracerebral hemorrhage treatment: Role for peroxisome proliferator-activated receptor gamma in microglia/macrophages. Ann. Neurol. 61, 352–362.
Zhou, P., Qian, L., Chou, T., and Iadecola, C. (2008). Neuroprotection by PGE2 receptor EP1 inhibition involves the PTEN/AKT pathway. Neurobiol. Dis. 29, 543–551.
Zhu, P., Genc, A., Zhang, X., Zhang, J., Bazan, N. G., and Chen, C. (2005). Heterogeneous expression and regulation of hippocampal prostaglandin E2 receptors. J. Neurosci. Res. 81, 817–826.
Keywords: brain, stroke, inflammation, GPCR, therapy, neurodegenerative diseases
Citation: Mohan S, Ahmad AS, Glushakov AV, Chambers C and Doré S (2012) Putative role of prostaglandin receptor in intracerebral hemorrhage. Front. Neur. 3:145. doi: 10.3389/fneur.2012.00145
Received: 09 July 2012; Accepted: 30 September 2012;
Published online: 22 October 2012.
Edited by:
Steven M. Greenberg, Harvard, USAReviewed by:
Edward C. Jauch, Medical University of South Carolina, USAGuohua Xi, University of Michigan, USA
Copyright: © 2012 Mohan, Ahmad, Glushakov, Chambers and Doré. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Sylvain Doré, Center for Translational Research in Neurodegenerative Disease, College of Medicine, University of Florida, 1275 Center Drive, Biomedical Science Building J493, Gainesville, FL 32610, USA. e-mail:c2RvcmVAdWZsLmVkdQ==