 Andrew W. Buxton
Andrew W. Buxton Frank L. Jalufka
Frank L. Jalufka Margaret E. Hruska
Margaret E. Hruska Jack R. Kubaney
Jack R. Kubaney Dylan A. McCreedy
Dylan A. McCreedy- 1Department of Biomedical Engineering, Texas A&M University, College Station, TX, United States
- 2Department of Biology, Texas A&M University, College Station, TX, United States
- 3Department of Public Health, Texas A&M University, College Station, TX, United States
Tissue clearing and 3D imaging have emerged as powerful techniques to assess the cellular and tissue-level architecture of the spinal cord. With the rapidly increasing variety and complexity of optical tissue clearing techniques, there is a critical need for optimization and streamlining of tissue-specific protocols, particularly when dealing with injury or disease states. We evaluated and combined multiple organic solvent-based techniques to develop sciDISCO: a spinal cord injury-optimized DISCO tissue clearing protocol. sciDISCO allows for the robust clearing, labeling, and 3D imaging of the intact spinal cord, as well as clearing around and through the lesion site formed after contusive spinal cord injury. In addition, we have identified alternatives for hazardous chemicals commonly used in organic solvent-based clearing including dichloromethane and dibenzyl ether. In this study, we demonstrate the compatibility of sciDISCO with multiple different labeling techniques to provide robust analysis of unique neuronal populations and morphologies in addition to cellular and tissue-level changes occurring following spinal cord injury.
Introduction
As the critical mediator of sensory and motor communication between the body and the brain, the spinal cord is a high priority area for neurological research. The spinal cord is composed of many phenotypically and morphologically distinct neuronal and glial populations as well as axon tracts from supraspinal, propriospinal, and peripheral neurons (Hayashi et al., 2018; Asboth et al., 2018; Wang et al., 2018). Disease and mechanical injury to spinal neuron populations or the myelinated axon tracts can disrupt signals coming from or going to the brain, leading to motor and sensory dysfunctions (Garcia-Ramirez et al., 2021; Shen et al., 2021). Understanding how spinal neuron populations transmit and modify motor, sensory, and autonomic signals in the intact spinal cord, as well as the cellular and circuit changes that occur in disease and injury, is critical for the development of new treatments for individuals with SCI (Izmailov et al., 2017; Naqvi et al., 2020; Courtine et al., 2008).
Historically, study of neuronal populations and tracts within the spinal cord has been achieved using mechanical sectioning techniques utilizing bladed instruments such as vibratomes or cryostats. While the data collected from physical tissue sections has proven highly valuable in our understanding of spinal cord circuits and function, the process can be prone to tissue deformations and is very labor-intensive. Recent technological advancements have been made to section and image spinal cord tissue in a more efficient manner (Fiederling et al., 2021), but are still prone to the limitations of mechanical sectioning including tearing or distortions of tissue sections (Oh et al., 2014; Ragan et al., 2012). In contrast with the historical sectioning techniques, the emergence of optical tissue clearing has provided a fast and efficient method of imaging whole, intact tissues using virtual or optical tissue sections that can then be rapidly assembled back into a 3D image (Ariel, 2017; Azaripour et al., 2016; Berke et al., 2016; Frenkel et al., 2023; Matryba et al., 2019; Richardson et al., 2021). With tissue clearing, cells and tissue components can be visualized in their native 3D configuration with minimal tissue loss (Rocha et al., 2019; Susaki et al., 2014). Over the past decade, many different forms of optical tissue clearing protocols have emerged.
Tissue clearing protocols have been commonly categorized based on their mechanism of action or properties of the solvents utilized. Categories of tissue clearing protocols include the aqueous-based methods, such as CLARITY (Chung et al., 2013), CUBIC (Susaki et al., 2015), and PACT (Yang et al., 2014; McCreedy et al., 2021; Treweek et al., 2015), which utilize aqueous detergents to delipidate tissues. The other major category of tissue clearing protocols are hydrophobic or organic solvent-based methods, such as 3DISCO (Ertürk et al., 2012; Becker et al., 2012), iDISCO+ (Renier et al., 2014; Renier et al., 2016), uDISCO (Pan et al., 2016), fDISCO (Qi et al., 2019), Ethanol-ECi (Masselink et al., 2019; Saritas et al., 2019), and PEGASOS (Jing et al., 2018), which utilize organic solvents to delipidate and match the refractive indices of tissues. Collectively, the various tissue clearing protocols provide an essential tool for the 3D imaging of many different tissues. However, the abundance of protocols and inherent differences between each method often results in the need for further optimization to achieve successful tissue clearing.
In this study, we sought to develop an optical tissue clearing protocol specifically designed for studying the intact and injured spinal cord. We aimed to address several limitations commonly encountered with optical clearing of the spinal cord including the need to remove the meninges due to tissue swelling, poor clearing of the extracellular matrix (ECM)-rich lesion site following contusive spinal cord injury, curvature of the spinal cord that hinders efficient imaging, lengthy protocol times, and use of hazardous chemicals. By combining multiple organic solvent-based methods, including iDISCO+ and Ethanol-ECi, we developed sciDISCO (spinal cord injury-optimized DISCO). The sciDISCO tissue clearing protocol has a shorter tissue processing time than iDISCO+ and other aqueous-based protocols, allows for the immunolabeling and clearing of the meningeal layer around the spinal cord, clears through the dense ECM that forms in the lesion site after contusive spinal cord injury (SCI), straightens the spinal cord to reduce imaging time and data size, replaces dichloromethane and dibenzyl ether with less hazardous alternatives, and allows for 3D visualization and imaging of the entire spinal cord using lightsheet microscopy. We demonstrate the utility of sciDISCO for visualizing lesion formation and neuronal loss, quantification of immune cell accumulation, imaging of the lymphatic system in the meninges of the spinal cord, and morphological analysis of sparsely-labeled neuronal populations. We believe that sciDISCO provides a robust and time efficient method for optically clearing the spinal cord that will accelerate research on normal spinal cord function as well as SCI and other spinal cord afflictions.
Materials and equipment
Delipidation
(1) 5 mL conicals (Axygen©, SCT-5ML).
(2) Methanol (Fisher Chemicals, A452-1).
(3) Dichloromethane (DCM, AcroSeal©, 326850010).
• Alternative: Benzotrifluoride (BTF, Thermo Scientific™, AAB2134030).
(4) 10X phosphate-buffered saline (Corning®, 20-031-CV).
(5) Nutating rocker—LabDoctor™ Mini Nutating Rocker (Ward’s Science™, H3D1020).
(6) Tween-20 (Thermo Scientific™, J20605-AP).
(7) Heparin (Sigma-Aldrich®, H3393-100KU).
(8) Sodium Azide (Millipore Sigma, S2002).
Immunofluorescent labeling
(1) Normal Donkey Serum (Millipore Sigma, D9663).
(2) Dimethyl Sulfoxide (Fischer Bioreagents, BP231-1).
(3) 0.45 μm Syringe Filter Unit (Millex-HP, SLHPR33RS).
(4) 10 mL syringe (Fisherbrand™, 14955458).
(5) 1.5 mL microcentrifuge tubes (Denville, C2170).
(6) 15 mL conicals (Falcon™, 352196).
(7) Incubator—Ward’s® Mini Digital Incubator (Ward’s Science™, 470230-608).
(8) Primary antibodies—Rabbit anti-NeuN (1:500, Millipore Sigma, ABN78), Chicken anti-NeuN (1:500, Millipore Sigma, ABN91), Rabbit anti-Lyve1 (1:500, Abcam, ab14917), Goat anti-mCherry/RFP (1:500, SicGen, AB0040-200), Goat anti-S100a8/MRP8 (1:1000, R&D Systems, AF3059) Rabbit anti-V5 (1:100, Bethyl Laboratories, A190-120A).
(9) Secondary antibodies (1:200)—Donkey anti-Rabbit IgG AlexaFluor® 488 (Invitrogen, A21206), Donkey anti-Rabbit IgG AlexaFluor® 647 (Invitrogen, A32795), Donkey anti-Chicken IgG AlexaFluor® 488 (Jackson, 703-545-155), Donkey anti-Goat IgG AlexaFluor® 647 (Invitrogen, A32849).
Agarose mounting and refractive index matching
(1) Sodium phosphate, monobasic, monohydrate (Millipore Sigma, S9638).
(2) Sodium phosphate, dibasic, anhydrous (Millipore Sigma, 567550).
(3) Agarose, low-melting temperature (Millipore Sigma, A9045).
(4) Heat block with 1.5 mL adaptor.
(5) Ethyl Cinnamate (Sigma-Aldrich, 112372-100G).
• Alternative for Dibenzyl Ether.
Methods
Animals
Adult (2–6 months) male and female C57Bl/6N (Charles Rivers), Chx10-Cre (obtained from Dr. Steven Crone) (Azim et al., 2014; Romer et al., 2017), Confetti (The Jackson Laboratory, Gt(ROSA)26Sortm1(CAG-Brainbow2.1)Cle) (Livet et al., 2007) and MORF3 (The Jackson Laboratory, Gt(ROSA)26Sortm3(CAG-smfp-V5*)Xwy) (Veldman et al., 2020; Viswanathan et al., 2015) were used for this study. Chx10-Cre mice were maintained on a C57Bl/6 J background and bred with Confetti or MORF3 mice to obtain sparse labeling of V2a interneurons. All studies were approved by the Institutional Animal Care and Use Committee at Texas A&M University.
Spinal cord injury
Adult male and female mice were anesthetized with 2% isoflurane before a laminectomy was performed at the T9 vertebrae level to expose the thoracic spinal cord. A 60 kDyne contusion injury was delivered to the spinal cord with a 1 s dwell time using the Infinite Horizons impactor (Precision Systems and Instrumentation). Surrounding musculature was sutured closed and the skin was closed with wound clips. Manual bladder expression and antibiotic/saline subcutaneous injections were performed twice daily until the spinal cords were collected.
Transcardial perfusion and fixation
Transcardial perfusion was performed as described previously (McCreedy et al., 2021). Mice are deeply anesthetized with a lethal dose of 2.5% Avertin (0.02 mL/g body weight; Sigma) before being perfused with 25 mL of ice-cold 1X phosphate buffered saline (PBS). Following the perfusion with PBS, animals were then perfused with 25 mL of ice-cold 4% paraformaldehyde in PBS for fixation. After fixation, the spinal column was removed and placed in ice-cold 4% paraformaldehyde overnight (12–24 h) for post-fixing. Following post-fixation, the spinal column was washed with 1X PBS. Using fine tweezers and microscissors, the spinal cord was carefully removed from the spinal column as previously described (McCreedy et al., 2021) and was stored at 4°C in PBS and 0.01% (w/v) sodium azide until ready for clearing.
3D printing of spinal cord straightener and agarose mounting chamber
The spinal cord straightener device and strap were 3D printed using natural polypropylene filament (Recreus PP3D) on a Prusa MK3S + printer with a layer height of 0.10 mm. Polypropylene filament was chosen due to its chemical resistance to the organic solvent clearing solutions. Packing tape was placed on the printer bed to increase adhesion of the polypropylene filament during printing. The agarose mounting chamber was 3D printed using polylactic acid (PLA) filament (Amazon Basics PLA) on a Prusa MK3S + with a layer height of 0.10 mm. 3D printing files are available in supplementary material.
Passive CLARITY and iDISCO+ tissue clearing
To compare our tissue clearing protocol to current aqueous and organic based clearing protocols, we performed PACT and iDISCO+ tissue clearing as previously described (Yang et al., 2014; McCreedy et al., 2021; Treweek et al., 2015; Renier et al., 2016; Jalufka et al., 2022) on spinal cords from adult male and female wild-type (WT) mice.
sciDISCO tissue clearing
Delipidation
Purpose: The delipidation step removes light scattering lipids from the tissue.
1. Dehydration of the spinal cord.
1.1. Transfer the dissected spinal cord from the 1X PBS solution into the 3D-printed spinal cord polypropylene straightener and secure it with the 3D-printed polypropylene strap. The spinal cord will remain in the straightener throughout the delipidation and rehydration process.
1.2. Prepare the dehydration series in 5 mL conicals of methanol and ddH2O: 20, 40, 60, 80, and 100% MeOH/ddH2O (v/v).
1.3. Transfer the spinal cord to the 20% MeOH/ddH2O solution, cap the conical tightly to prevent leaks, and rock on a nutating rocker for 1 h at room temperature. After the hour, transfer to the 40% MeOH/ddH2O solution and rock for another hour. Repeat throughout the entire dehydration series, rocking the spinal cord in each solution for 1 h at room temperature. After the final 100% MeOH/ddH2O solution, transfer the spinal cord to a new conical containing 5 mL of 100% MeOH and rock for 1 h at room temperature to ensure complete removal of water from the sample.
1.4. Prepare 5 mL of 66% DCM/ 33% MeOH (v/v) solution in a 5 mL conical. BTF can be used instead of DCM. Use glass stripettes and polypropylene tubes when working with DCM or BTF.
1.5. Transfer the spinal cord to the prepared DCM/MeOH solution and incubate overnight (12–24 h) while gently rocking on the nutating rocker at room temperature. BTF can be used instead of DCM.
2. Rehydration of the spinal cord.
2.1. Transfer the spinal cord from the 66% DCM/ 33% MeOH solution to a 5 mL conical filled with 100% methanol. Incubate the spinal cord in 100% MeOH for 30 min with gentle rocking, then replace the MeOH with 5 mL of fresh 100% MeOH and incubate for an additional 30 min with gentle rocking at room temperature.
2.2. Prepare the rehydration series (MeOH/H2O) in 5 mL conicals with the following percentages of methanol in ddH2O: 80, 60, 40, and 20% MeOH/ddH2O (v/v).
2.3. Rehydrate the spinal cord sample by placing the spinal cord in the rehydration series starting with the 80% MeOH/ddH2O solution for 1 h rocking on a nutating rocker at room temperature. After the hour, transfer the tissue through the remainder of the series, 1 h in each solution, rocking on a nutating rocker at room temperature.
2.4. Prepare 1x PBS.
2.4.1. Combine 100 mL of 10X PBS with 900 mL of ddH2O and stir.
2.5. Transfer 5 mL of the prepared PBS solution to the 5 mL conical. Place the spinal cord sample in this PBS solution for 30 min with gentle rocking at room temperature.
2.6. Prepare 1 L of PBS/Tween-20 with Heparin (PTwH).
2.6.1. Combine 100 mL of 10X PBS, 2 mL Tween-20, and 1 mL of 10 mg/mL Heparin and ddH2O to a final volume of 1 L. Stir until thoroughly mixed. Add sodium azide to a final concentration of 0.02% (w/v). Store at room temperature.
2.7. Remove the spinal cord from the straightener and transfer 5 mL of the prepared PTwH into a 5 mL conicals. Wash the sample in PTwH for 30 min. Repeat.
Checkpoint: Possible stopping point – tissue may be stored in 1x PBS with 0.01% (w/v) sodium azide at 4°C for several weeks.
Immunofluorescent labeling
Purpose: Immunofluorescent labeling allows for the visualization of endogenous or transgenic epitopes within the cleared tissue.
1. Primary antibody staining.
1.1. Prepare 1 mL of the primary antibody solution in a 1.7 mL microcentrifuge tube.
1.1.1. Combine: PTwH, 5% DMSO, 3% Normal Donkey Serum, and primary antibodies. If samples are too large for 1.5 mL microcentrifuge tubes, use a 5 mL conical and adjust volume of primary antibody solution accordingly.
1. 1.2. Incubate the spinal cord sample in the primary antibody solution for 2 days at 37°C with gentle rocking.
2. PTwH washes.
2.1. Wash the spinal cord sample 5 times for 30 min each with 12.5 mL of PTwH in a 15 mL conical while rocking with a nutating rocker at room temperature.
2.2. Leave the sample in the 5th wash overnight (12–24 h) with gentle rocking at room temperature.
3. Secondary antibody staining.
3.1. Prepare 1.2 mL of the secondary antibody solution in a 1.7 mL microcentrifuge tube.
3.1.1. Combine PTwH, 5% NDS, and secondary antibodies.
3.1.2. Once prepared, filter the secondary antibody solution using a 3 mL syringe attached to a 0.22 μm syringe filter. Filter into a new 1.7 mL microcentrifuge. Some loss of the secondary antibody solution will occur, but the final volume should be ~ 1 mL.
3.2. Incubate spinal cord samples for 2 days at 37°C in the secondary antibody solution with gentle rocking.
4. Wash samples in PTwH.
4.1. Repeat PTwH Washes (Step 2), leaving the sample in the 5th PTwH wash overnight (12–24 h) on the nutating rocker at room temperature.
Agarose mounting and refractive index matching
Purpose: Mounting spinal cord samples in agarose can help with positioning and manipulating the sample safely within the Zeiss lightsheet microscope. Refractive index matching helps homogenize the refractive indices of the tissue and agarose to minimize light diffraction during imaging.
1. Mounting spinal cord in agarose (optional).
1.1. Prepare 0.1 M phosphate buffer (PB), pH 7.4.
1.1.1. Dissolve 3.1 g of sodium phosphate (NaH2PO4, monobasic, monohydrate) and 10.9 g of sodium phosphate (Na2HPO4, dibasic, anhydrous) in 900 mL of ddH2O. Add ddH2O to a final volume of 1 L.
1.2. Prepare 0.02 M phosphate buffer by combining 200 mL of 0.1 M PB with 800 mL of ddH2O.
1.3. Prepare sufficient volume of 2% agarose to fully immerse the spinal cord and any microscope attachments necessary for moving the sample.
1.3.1. To make 20 mL of 2% agarose, combine 400 mg of low-melt agarose in 20 mL of 0.02 M phosphate buffer.
1.3.2. Microwave in increments to avoid boiling over until all the agarose is fully dissolved.
1.3.3. Aliquot 1 mL into 1.5 mL microcentrifuge tubes. Aliquots can be stored at 4°C.
1.3.4. To re-melt, place microcentrifuge tubes in the heat block at 95°C. Cool the agarose to ~60°C prior to use.
1.4. Position the spinal cord and any microscope handling components in a mold for the agarose. Slowly pipette in melted agarose until the sample and components are fully submerged. Allow agarose to solidify in a light protected environment for at least 30 min. Do not let agarose set for more than 2 h as it can dry out and crack. Once sample is embedded, proceed to the next step.
2. Dehydration of the spinal cord.
2.1. Prepare the dehydration series (MeOH/H2O) in 5 mL conicals with the following concentrations of methanol: 20, 40, 60, 80, and 100%.
2.2. Dehydrate the sample through the prepared series in increasing order of methanol concentration.
2.2.1. Lay the conicals horizontally on a nutating rocker to prevent the spinal cords and agarose from warping along the bottom of the conicals during the dehydration steps. Gently rock for 1 h each at room temperature.
2.3. Incubate the spinal cord sample in 100% MeOH overnight (12–24 h) in a 5 mL conical with gentle rocking at room temperature.
3. Refractive index matching.
3.1. Prepare 5 mL of 66% DCM/ 33% MeOH (v/v) solution in a 5 mL conical. BTF can be used instead of DCM.
3.2. Incubate sample in the DCM/MeOH solution for 3 h on a nutating rocker at room temperature.
3.3. Wash sample twice in 5 mL of 100% DCM in 5 mL conicals for 15 min each on the nutating rocker at room temperature. BTF can be used instead of DCM.
3.4. Prepare 5 mL of ethyl cinnamate in the 5 mL conical.
3.5. Transfer the spinal cord sample to the ethyl cinnamate solution and equilibrate for at room temperature for at least 1 full day prior to imaging. For best results, gently invert the conical every 1–2 h before leaving overnight (12–24 h).
3.5.1. The following day, replace with fresh ethyl cinnamate and let the sample rest for at least 2 h prior to imaging. Gently invert the conical every 1–2 h to ensure mixing of the ethyl cinnamate throughout the sample.
Lightsheet microscopy
After equilibration in ethyl cinnamate for at least 1 day, spinal cord samples were imaged using a Zeiss Z1 Lightsheet microscope with a 5x acquisition objective. Virtual image z-stacks were captured using both right and left sided lightsheet illumination (5x illumination objectives) and fused together in Zen software.
Image analysis
Lightsheet images were analyzed in Imaris 3D software (Bitplane), Matlab, Python, or ImageJ (Schindelin et al., 2012). To calculate the mean fluorescent intensity of NeuN labeling, an image mask was created for NeuN+ pixels for each z-plane in the 3D images in MATLAB. The NeuN+ image mask was used to generate a binary image file removing any background fluorescence. The masked image file was then imported into a Python script, which was used to calculate the average fluorescent intensity of each image. To calculate the signal-to-background ratio of NeuN labeling, a signal mask for the NeuN+ pixels was created using a manual threshold, and a background mask was generated by thresholding the entire tissue and subtracting the signal mask. For quantitative comparisons, all samples were cleared, immunolabeling, and imaged in batch and the same signal and background threshold levels were used for every image. The signal-to-background ratio for each image was calculated by dividing the mean intensity within the signal mask by the mean intensity within the background mask. MRP8+ cell quantification was performed by using the Imaris spot function. The number of MRP8+ cells within 500 μm of the lesion center were counted in each image.
Statistical analysis
Statistical analysis was performed using Prism software (Graphpad). Sample sizes for replicates are presented in figure legends. Data is presented as mean ± SEM. Normality of data was confirmed using the Shapiro–Wilk test. p values were calculated by two-sided Student’s t test, one-way ANOVA with Tukey’s post hoc test, or two-way ANOVA with Tukey’s post hoc test. p < 0.05 was considered significant.
Results
sciDISCO clears the full intact murine spinal cord in a time efficient manner
To develop a streamlined tissue clearing protocol, we identified and combined the essential and most effective components of multiple tissue clearing procedures including iDISCO+ and ethanol-ECi (Becker et al., 2012; Renier et al., 2016; Klingberg et al., 2017). The resulting sciDISCO optical clearing protocol is comprised of three main steps: delipidation, tissue labeling, and refractive index matching (Figure 1A). The first step involves delipidation, in which methanol and dichloromethane (DCM) or benzotrifluoride (BTF) are used to remove the majority of light scattering lipids from the tissue. Next in the immunolabeling step, primary and secondary antibodies are used to label target epitopes in order visualize biomolecules with cellular and subcellular resolution. Finally in the refractive index matching step, the spinal cord is encased in agarose using our 3D printed agarose mounting chamber (Figure 1B), dehydrated again, and then incubated in ethyl cinnamate for refractive index matching for at least 24 h prior to imaging (Figure 1D). Samples can be kept light protected at room temperature in ethyl cinnamate until imaging.
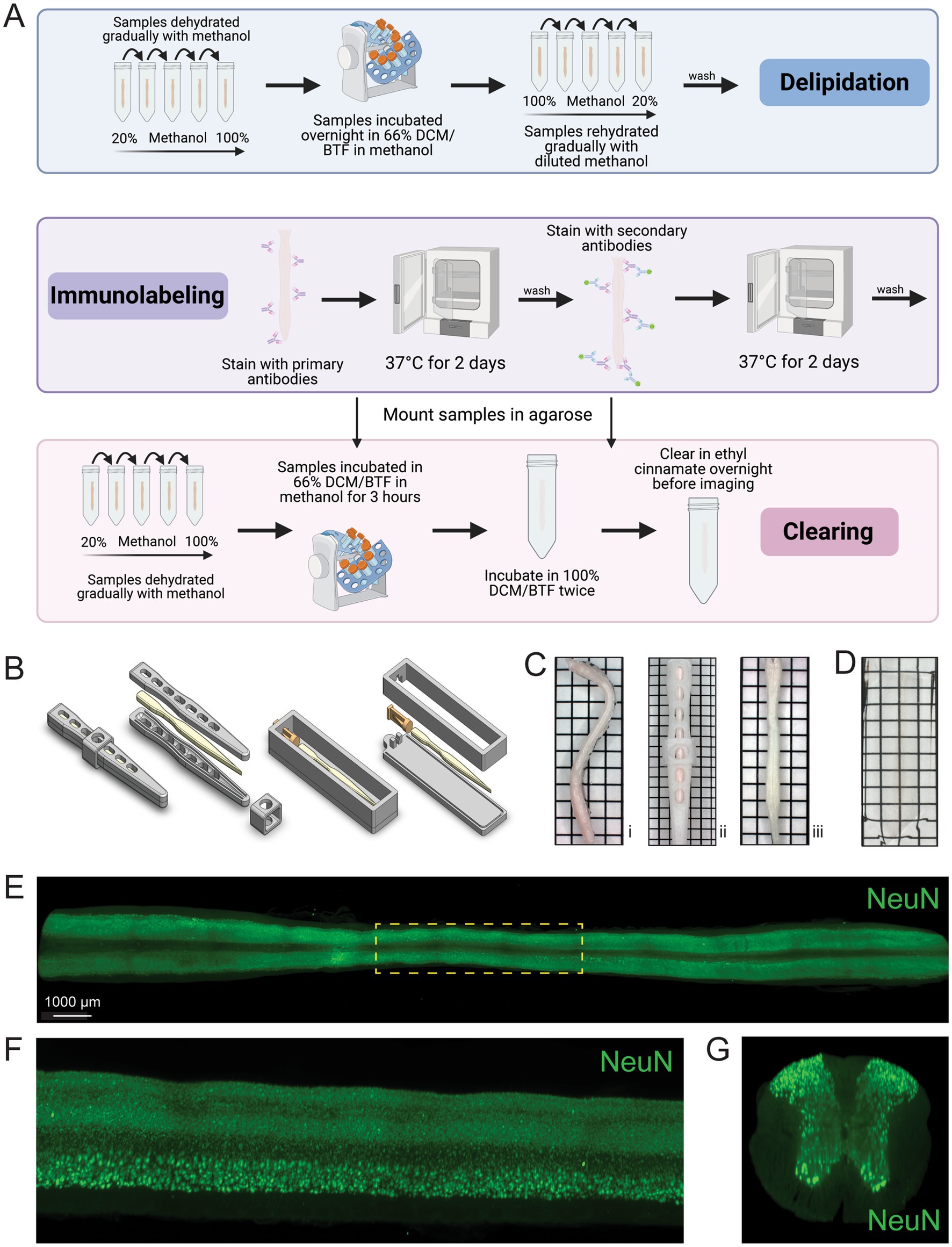
Figure 1. The sciDISCO protocol allows for the 3D imaging of the entire spinal cord. (A) Visual schematic of the sciDISCO protocol involving the delipidation of the spinal cord, immunolabeling, and RI matching in ethyl cinnamate. Created in BioRender (https://BioRender.com/7kqz2km). (B) 3D schematics of the spinal cord straightener and agarose mounting chamber. (C) (i) A curved spinal cord after post-fixation and dissection. (ii) The spinal cord in the 3D printed spinal cord straightener and (iii) the straightened spinal cord after delipidation and removal from the straightener. (D) A cleared spinal cord embedded in agarose and RI matched with ethyl cinnamate. (E) Horizontal view of a 3D lightsheet image of a spinal cord including cervical, thoracic, and lumbar segments labeled with NeuN antibody to visualize neuronal cell bodies. (F) Zoomed in sagittal view of the outline thoracic region (yellow) from (E). (G) Virtual transverse cross section of the thoracic spinal cord. Grids on images are made up of 2.5 mm × 2.5 mm squares.
The murine spinal column is inherently curved and we noticed that the dissected spinal cords reverted to a pronounced curved shape during the dehydration step (Figure 1Ci). Imaging the curved spinal cord is technically challenging and increases imaging time and image data size by requiring imaging of void space below and above the curved spinal cord regions. To address these limitations, we developed a 3D printed straightener device to house the spinal cord during the initial delipidation steps (Figures 1B,Cii). Following delipidation using this cage, the spinal cord remained straight for the duration of the protocol (Figure 1Ciii).
Using the sciDISCO protocol, we were able to capture immunofluorescent staining (NeuN) in the full length, intact murine spinal cord using lightsheet microscopy (Figure 1E). The resulting 3D images enabled visualization of the entire labeled spinal cord tissue (Figure 1E, horizontal view). Multiple view angles of selected regions of the 3D image can be readily obtained including the sagittal (Figure 1F) and transverse views (Figure 1G).
In respect to comparable protocols, iDISCO+ and PACT, sciDISCO is considerably more time efficient, able to clear and immunolabel the whole spinal cord in as little as 9 days, compared to 12 days for iDISCO+ and 16 days for PACT (Figure 2A). Despite the reduced time required for clearing and immunolabeling, sciDISCO delivers successful immunolabeling using intact spinal cords with the meninges removed (Figure 2B).
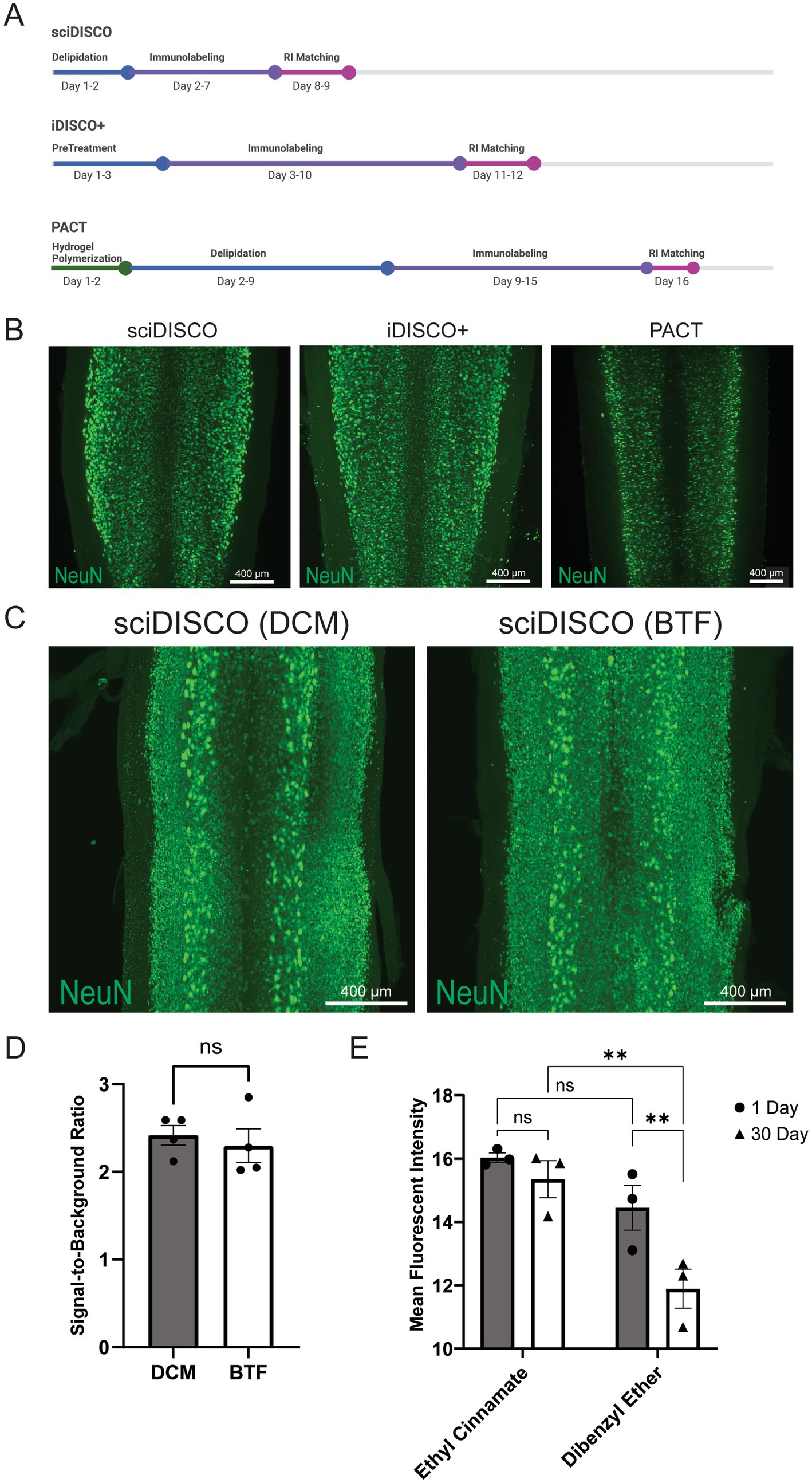
Figure 2. sciDISCO is faster than comparable clearing protocols and delivers successful immunolabeling. (A) Timeline schematic detailing the average times for each main step in sciDISCO, iDISCO+, and the PACT clearing protocols. Created in BioRender (https://BioRender.com/ghn1jij). (B) sciDISCO delivers similar immunolabeling of NeuN when compared to iDISCO+ and PACT clearing protocols. Representative maximum intensity projections of 3D lightsheet images of the lower cervical and upper thoracic regions are shown. (C) Representative maximum intensity projections of NeuN-labeled thoracic spinal cord regions cleared using either DCM or BTF in the sciDISCO protocol. (D) Quantification of the signal-to-background ratio of samples cleared with sciDISCO using either DCM or BTF. Mean ± SEM. Student’s t-test. (E) Quantification of mean fluorescent intensity of samples incubated in ethyl cinnamate (sciDISCO) vs. dibenzyl ether (iDISCO+) after 1 and 30 days of incubation. Mean ± SEM. ** p < 0.01. Two-way ANOVA with Tukey’s multiple comparisons post-hoc test.
DCM was originally used for delipidation in the development of the sciDISCO protocol. However, there are potential health and environmental risks when working with DCM (Schlosser et al., 2015), as well as an upcoming ban in the United States for most uses of DCM. We found that another, more environmentally friendly organic solvent, benzotrifluoride (BTF) (Verebélyi and Iván, 2012), performs comparably when used as a direct alternative to DCM in the sciDISCO protocol (Figure 2C). BTF also maintains the same signal-to-background ratio of NeuN labeling compared to DCM with sciDISCO (Figure 2D). We also wanted to test the long-term storage capability of samples in RI matching solutions for sciDISCO vs. iDISCO+. We allowed samples to incubate in ethyl cinnamate and dibenzyl ether, respectively, in light protected boxes at room temperature for 30 days prior to imaging. Relative to dibenzyl ether, ethyl cinnamate is a less toxic compound that has been shown to provide successful RI matching during tissue clearing while maintaining fluorescence long-term in other clearing protocols (Klingberg et al., 2017). Maximum intensity projections were generated from 3D images for each sample and a mask of all NeuN+ pixels was generated using MATLAB. The NeuN+ areas of each image were then assessed in Python to calculate the mean fluorescent intensity of all NeuN+ voxels for each sample. After 1 day of refractive index matching, there was no significant difference in mean fluorescent intensity of NeuN labeling between the sciDISCO samples stored in ethyl cinnamate and the iDISCO+ samples stored in dibenzyl ether (Figure 2E). However, after 30 days of incubation the iDISCO+ samples had reduced fluorescent intensity relative to sciDISCO samples as well as iDISCO+ samples at 1 day of incubation. No decline in fluorescent intensity was observed with sciDISCO samples stored in ethyl cinnamate.
sciDISCO clears through the lesion in the sub-acutely injured spinal cord
Following contusive SCI in mice, the lesion site undergoes extensive remodeling and becomes filled with ECM proteins such as collagen IV and is surrounded by a layer of reactive astrocytes commonly referred to the as the glial border or scar (Figure 3A) (Cooper et al., 2018; Yang et al., 2020). The ability to clear through the lesion site would allow for the 3D analysis of lesion formation and structure. Previous studies have demonstrated limited clearing of the lesion site after the sub-acute phase (10–14 dpi) with aqueous-based protocols such as PACT or CLARITY (Asboth et al., 2018; McCreedy et al., 2021). We also observed poor clearing of the lesion site with PACT, however, the sciDISCO protocol readily cleared through the ECM-rich lesion at 14 dpi (Figure 3B).
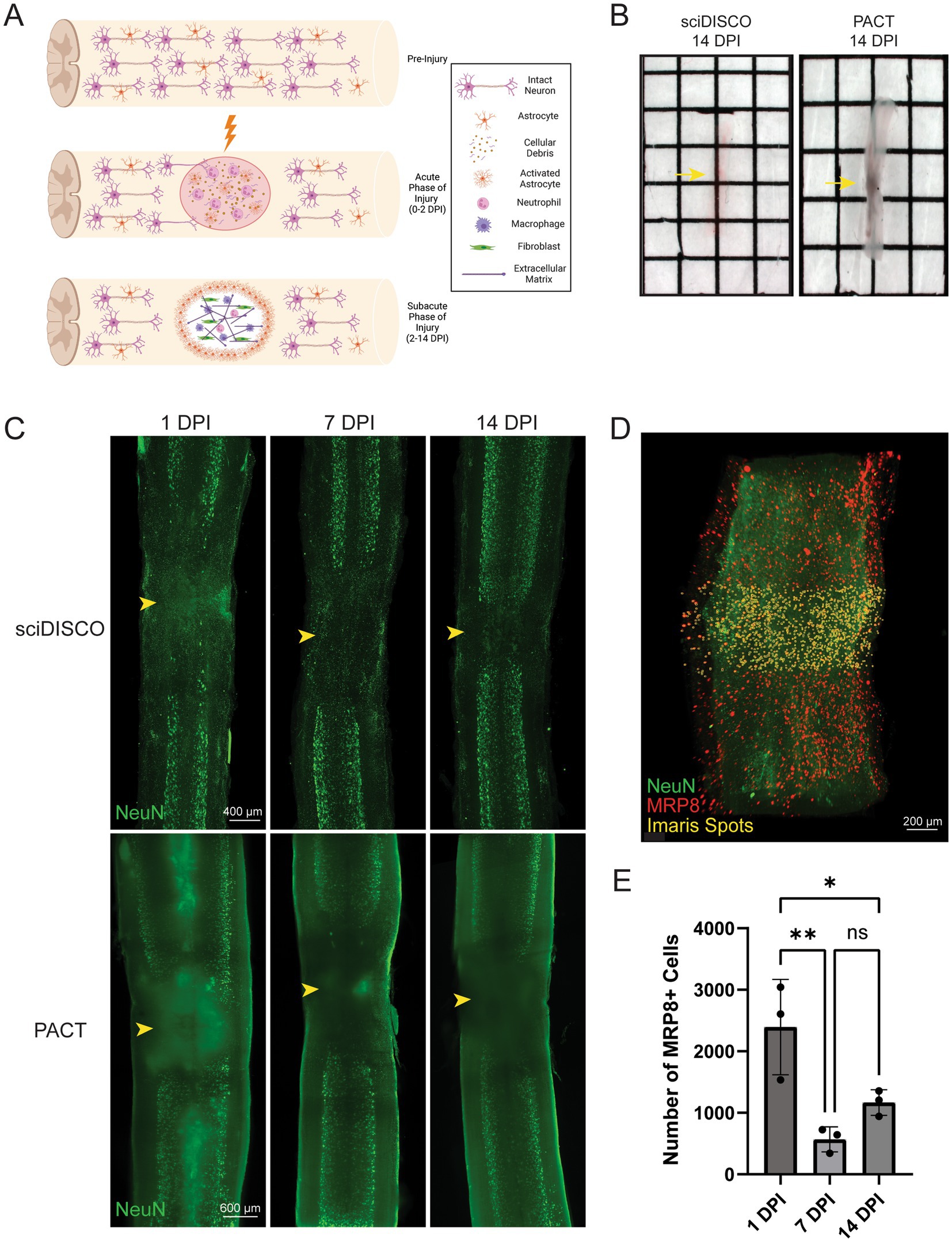
Figure 3. sciDISCO clears through the subacute lesion after contusive spinal cord injury. (A) Graphical representation of the lesion formation and extracellular matrix deposition following contusive spinal cord injury in mice. By the subacute phase of injury, the lesion site is filled with extracellular matrix proteins, macrophages, and fibroblasts. Created in BioRender (https://BioRender.com/yzaetus). (B) Macroscopic images of thoracic spinal cords at 14 dpi. Samples are RI matched in ethyl cinnamate (sciDISCO) or RIMS (PACT) with the lesion site indicated by the yellow arrow. The lesion site remains opaque in the PACT sample. (C) Maximum intensity projections of sciDISCO and PACT cleared thoracic spinal cords at 1, 7, and 14 days post contusive injury. The lesion center is indicated by arrowhead. (D) Example image of quantification of MRP8+ cells within 500 μm of the lesion center using the Imaris spots function. (E) Quantification of MRP8+ cell counts in sciDISCO cleared cords at 1, 7, and 14 dpi. * p < 0.05, ** p < 0.01. One-way ANOVA with Tukey’s post-hoc test.
The increased capacity to clear through the sub-acute lesion site led to improved lightsheet imaging of the injured spinal cord cleared with sciDISCO relative to PACT (Figure 3C). Beginning at as early as 1 dpi, the SCI lesion site failed to fully clear with PACT leading to opaque regions in the lightsheet images due to light scattering. Acutely injured spinal cord tissues readily cleared with sciDISCO, leading to virtually no opaque regions in the resulting lightsheet images. With PACT clearing, the light scattering effect of the poorly cleared lesion site is more pronounced at 7 and 14 dpi timepoint as the lesion site is remodeled and the deposited ECM is resistant to the aqueous clearing technique (Azaripour et al., 2016). Despite the increased presence of ECM, sciDISCO readily cleared through the SCI lesion site of the subacutely injured spinal cord.
To test the ability to quantify cells within the lesion after clearing, we labeled neutrophils in the injured spinal cord with an anti-MRP8 antibody and quantified the number of neutrophils within 500 μm of the lesion center using Imaris (Figures 3D,E). Neutrophils were used as our target cell as they are among the first peripheral immune cells to infiltrate in large numbers following SCI and have been shown to peak in number at 1 dpi before gradually decreasing over time in the injury site (Donnelly and Popovich, 2008; Neirinckx et al., 2014). Using the Imaris spots function, MRP8-positive cells were quantified within the lesion site for 1, 7, and 14 dpi spinal cords. Quantitative analysis showed that there is an initial influx of neutrophils at the 1 dpi timepoint that decreases at the 7 and 14 dpi timepoints (Figure 3E). Overall, these results indicate that our sciDISCO protocol is efficient in clearing through the acutely and sub-acutely injured spinal cord and allows for the quantification and study of inflammation and lesion formation after injury.
sciDISCO allows for immunolabeling and clearing of the meningeal layers around the spinal cord
The meninges comprise a set of protective membranes that surround the central nervous system, including the spinal cord, and contain lymphatic vessels that play a crucial role in immune surveillance and the transport of immune cells (Jacob et al., 2019). The meningeal layers of the spinal cord have also been recently identified as a critical location for immune cell trafficking in mouse models of SCI or experimental autoimmune encephalitis (Cugurra et al., 2021; Mazzitelli et al., 2023). However, many protocols require the removal of meninges to improve optical transparency and allow for swelling of the spinal cord tissue parenchyma during the tissue clearing procedures. We have previously found that the meningeal layer had to be removed from the fixed spinal cord prior to PACT clearing (McCreedy et al., 2021). Without removal, the meninges frequently constricted around the swelling tissue and prevented adequate tissue clearing and antibody penetration (Figures 4A,B). While removal of the meninges improved PACT clearing and immunolabeling, imaging of Lyve1+ lymphatic vessels in the meninges was severely compromised (Figure 4B). After clearing with sciDISCO, the meninges and spinal cord cleared completely (Figure 4C) and robust labeling of both neurons in the parenchyma and the lymphatic vessels within the meninges was readily observed (Figure 4D). Collectively, our findings demonstrate the flexibility of sciDISCO for tissue clearing and immunolabeling with and without the meningeal layers surrounding the spinal cord.
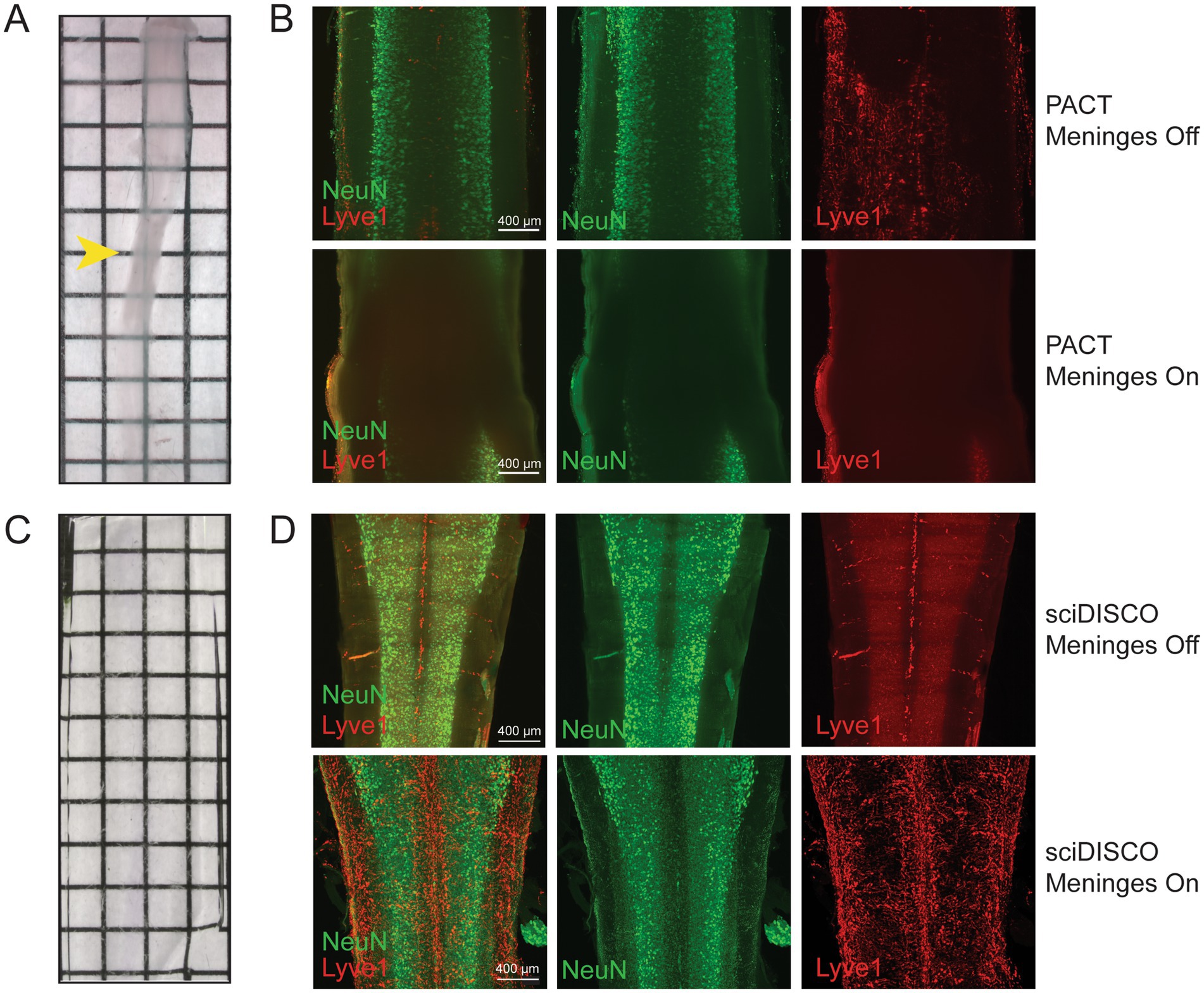
Figure 4. sciDISCO allows for the immunolabeling and imaging of the meningeal layers around the spinal cord. (A) Macroscopic image of a mouse spinal cord cleared and RI matched in RIMS (PACT) with the meninges still intact. The PACT clearing process causes the tissue parenchyma to swell and the meninges to constrict around the spinal cord (arrowhead). (B) NeuN and Lyve1 immunostaining of a PACT cleared spinal cord with meninges removed (top) and intact (bottom). (C) Macroscopic image of a mouse spinal cord cleared and RI matched with ethyl cinnamate (sciDISCO) with the meninges still intact. (D) NeuN and Lyve1 immunostaining of a sciDISCO cleared spinal cord with the meninges removed (top) and with the meninges intact (bottom). All images in (B,D) are maximum intensity projections of 3D lightsheet images. Grids on macroscopic images are made up of 2.5 mm × 2.5 mm squares.
sciDISCO is compatible with transgenic sparse labeling techniques to identify morphological characteristics of neuronal populations
Finally, to test that our protocol is compatible with reporter mouse lines and multiple types of transgenic labels, we tested two different sparse labeling techniques to assess neuronal morphology. We first bred Chx10-Cre mice with the Confetti (Gt(ROSA)26Sortm1(CAG-Brainbow2.1)Cle) reporter mouse line resulting in offspring (referred to as Chx10-Confetti mice) with Chx10+ neurons (putatively V2a interneurons) stochastically expressing either red fluorescent protein (RFP), cyan fluorescent protein (CFP), green fluorescent protein (GFP), or yellow fluorescent protein (YFP; Figure 5A). V2a interneurons were selected as they have recently been shown to play a vital role in recovery of locomotor function after injury (Courtine et al., 2008; Zholudeva et al., 2017; Zholudeva et al., 2021; Zhong et al., 2010; Zavvarian et al., 2020). As with other organic solvent-based clearing protocols, endogenous fluorescence is quenched, therefore an antibody against RFP was used to label RFP-expressing V2a interneurons (Figure 5B). This labeling strategy allowed for the visualization of V2a interneuron soma and tracts.
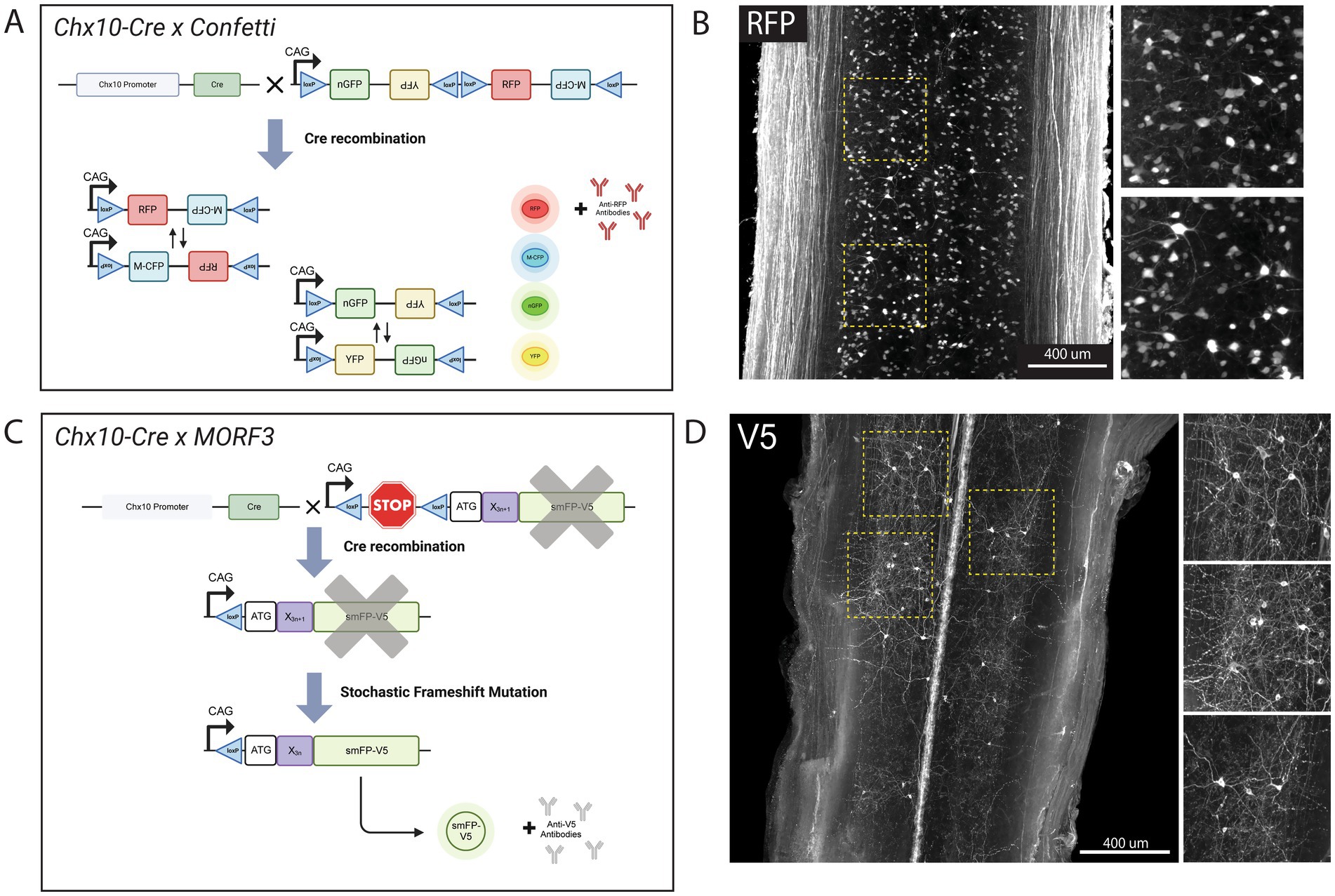
Figure 5. sciDISCO is compatible with transgenic mouse lines for sparse labeling of neuronal populations. (A) Graphical schematic of the stochastic Confetti fluorescent reporter expression upon crossing with Chx10-Cre mice. Created in BioRender (https://BioRender.com/5cwn62g). (B) Lightsheet imaging of RFP+ V2a interneurons in the thoracic spinal cord following clearing with sciDISCO. (C) Graphical schematic of the MORF3 sparse labeling system upon crossing with Chx10-Cre mice. Created in BioRender (https://BioRender.com/fh674nv). (D) Lightsheet images of very sparse labeling of V2a interneurons in the thoracic region of the spinal cord following sciDISCO clearing. Both images in (B,D) are partial maximum intensity projections of the intermediate laminae of the spinal cord from the 3D lightsheet images.
While visualization of individual V2a interneuron soma and proximal neurites was readily accomplished with tissue clearing and lightsheet imaging of the spinal cords from Chx10-Confetti mice, long-distance tracing of axons could not be performed due to the substantial density of RFP+ axons in the white matter. To achieve sparser labeling of V2a interneurons, we crossed Chx10-Cre mice with MORF3 (Gt(ROSA)26Sortm3(CAG-smfp-V5*)Xwy) reporter mice. The MORF3 mouse line contains a stop codon under control of the Cre-Lox system, and then a spaghetti monster fluorescent protein V5 (smFP-V5-F) behind a MORF (Mononucleotide Repeat Frameshift) sequence (Veldman et al., 2020). During neural development, as V2a interneurons are generated, the stop codon will be removed by Cre and a stochastic frameshift of the mononucleotide repeat region will occur in a small subset of V2a interneurons bringing the smFP-V5-F reporter protein coding sequence into frame (Figure 5C). Immunolabeling with anti-V5 antibodies after sciDISCO clearing enabled visualization of sparsely-labeled V2a interneurons, including their cell bodies and neurites in the spinal cord (Figure 5D). Combining sciDISCO and sparse-labeling reporter lines allows for detailed morphological analysis of spinal interneuron populations, which can be utilized for future connectomic studies.
Discussion
Optical tissue clearing has emerged as an important method for the study of 3D structures within tissues without the need for mechanical sectioning of the tissue or the requirement to rebuild a tissue from hundreds of individual tissue sections. In addition to allowing novel insights into the structure and function of normal healthy tissues, tissue clearing and 3D imaging can be highly valuable for assessing changes associated with injury or disease states. While tissue clearing techniques have been described since 1914 (Spalteholz, 1914) and come in a wide range of varieties, the need for optimizations for different disease states or tissues is still warranted. By combining the most effective components of PACT, iDISCO+ and Eci-based tissue clearing procedures, we have developed sciDISCO as an optimized and streamlined protocol for optical clearing of the spinal cord, including the lesion site after SCI.
An issue that arises with respect to optical tissue clearing and lightsheet microscopy is the immense amount of image data that needs to be stored. Large-scale 3D imaging of cleared samples can generate many gigabytes to terabytes of images, particularly when dealing with larger tissues such as the spinal cord (Andreev and Koo, 2020). Curves or bends within the tissue can lead to excess data acquisition and storage as more void space can be required to image the entire tissue. Specifically, when dehydrating the murine spinal cord, the curvature of the spinal cord is exacerbated, which can lead to an increase in image data when imaging the entire spinal cord. The use of our 3D printed spinal cord straightener keeps the spinal cord straight during dehydration, allowing for images to be captured with fewer imaging planes (optical sections) to image the entire cord and reducing the amount of data and time for each image, as well as lowering the required storage space. One limitation of straightening the spinal cord may be the potential distortion of the tissue from its native shape.
One major area of SCI research involves the lesion and glial border that forms following traumatic SCI. In mice, the glial border forms around the lesion center in the sub-acute phase of injury. Within the lesion, a dense network of collagen fibers and other ECM proteins fills the lesion (Cooper et al., 2018), and is surrounded by border-forming astrocytes (Alizadeh et al., 2019). sciDISCO, with the ability to clear through this dense lesion and visualize components within the lesion and within the glial border, opens up areas of research to further understand lesion formation and glial border development over time, and understanding how the lesion and glial border interact with spared neurons around the injury site.
Following the disruption of neural circuitry caused by SCI, studies have shown that some damaged neurons display a degree of plasticity and form detour circuits, circumventing the injury and lesion area in an attempt to contribute to functional recovery (Filli et al., 2014). Detour circuits can form from damaged neurons sprouting collaterals that establish new connections with neurons spared after the injury (Granier et al., 2020; Meehan et al., 2020). Previous research has shown long-distance propriospinal interneurons contribute to detour circuit formation and locomotor functional recovery following cervical injury (Zholudeva et al., 2017). sciDISCO, with the ability to clear through the lesion site, and combined with sparse neuronal labeling technologies such as the MORF3 mouse line, can allow for the visualization and mapping of detour circuit formation following SCI. In addition to detour circuit mapping, identification of morphological changes in spared neurons following SCI can lead to a better understanding of endogenous recovery mechanisms and spontaneous functional recovery within the spinal cord following injury.
The detailed protocol for sciDISCO described in this report will provide a powerful tool for studying the spinal cord, including SCI and recovery. By streamlining the procedures, we aim to make tissue clearing of the spinal cord more accessible and efficient. While lightsheet microscopy was used in this study to generate 3D images, the optically cleared spinal cord could be imaged with other modalities including confocal microscopy. The advances provided by the sciDISCO protocol have the potential to rapidly advance our understanding of the cellular mechanisms of normal spinal cord function and dysfunction caused by injury or disease, thereby leading to the discovery of new therapies to improve long-term outcomes for afflicted individuals.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Texas A&M University Institutional Animal Care and Use Committee. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
AB: Formal analysis, Methodology, Data curation, Writing – original draft, Software, Writing – review & editing, Investigation, Conceptualization. FJ: Writing – original draft, Data curation, Formal analysis, Writing – review & editing, Conceptualization, Investigation, Methodology. MH: Investigation, Writing – original draft, Data curation, Methodology. JK: Investigation, Writing – original draft, Methodology, Data curation. DM: Methodology, Resources, Validation, Conceptualization, Writing – review & editing, Funding acquisition, Supervision, Writing – original draft, Project administration, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Funding was provided by Mission Connect, a program of TIRR Foundation, (grants 019-117 and 021-103) and NIH NINDS R01NS122961.
Acknowledgments
We would like to acknowledge the Texas A&M University Microscopy and Imaging Center for the use of the lightsheet microscope as well as the Imaris software, particularly Dr. Holly Gibbs for her expertise and knowledge of lightsheet microscopy.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alizadeh, A., Dyck, S. M., and Karimi-Abdolrezaee, S. (2019). Traumatic spinal cord injury: an overview of pathophysiology, models and acute injury mechanisms. Front. Neurol. 10:282. doi: 10.3389/fneur.2019.00282
Andreev, A., and Koo, D. E. (2020). Practical guide to storage of large amounts of microscopy data. Microsc. Today 28, 42–45. doi: 10.1017/S1551929520001091
Ariel, P. (2017). A beginner’s guide to tissue clearing. Int. J. Biochem. Cell Biol. 84, 35–39. doi: 10.1016/j.biocel.2016.12.009
Asboth, L., Friedli, L., Beauparlant, J., Martinez-Gonzalez, C., Anil, S., Rey, E., et al. (2018). Cortico–reticulo–spinal circuit reorganization enables functional recovery after severe spinal cord contusion. Nat. Neurosci. 21, 576–588. doi: 10.1038/s41593-018-0093-5
Azaripour, A., Lagerweij, T., Scharfbillig, C., Jadczak, A. E., Willershausen, B., and van Noorden, C. J. F. (2016). A survey of clearing techniques for 3D imaging of tissues with special reference to connective tissue. Prog. Histochem. Cytochem. 51, 9–23. doi: 10.1016/j.proghi.2016.04.001
Azim, E., Jiang, J., Alstermark, B., and Jessell, T. M. (2014). Skilled reaching relies on a V2a propriospinal internal copy circuit. Nature 508, 357–363. doi: 10.1038/nature13021
Becker, K., Jährling, N., Saghafi, S., Weiler, R., and Dodt, H. U. (2012). Chemical clearing and dehydration of GFP expressing mouse brains. PLoS One 7:e33916. doi: 10.1371/journal.pone.0033916
Berke, I. M., Miola, J. P., David, M. A., Smith, M. K., and Price, C. (2016). Seeing through musculoskeletal tissues: improving in situ imaging of bone and the lacunar Canalicular system through optical clearing. PLoS One 11:e0150268. doi: 10.1371/journal.pone.0150268
Chung, K., Wallace, J., Kim, S. Y., Kalyanasundaram, S., Andalman, A. S., Davidson, T. J., et al. (2013). Structural and molecular interrogation of intact biological systems. Nature 497, 332–337. doi: 10.1038/nature12107
Cooper, J. G., Jeong, S. J., McGuire, T. L., Sharma, S., Wang, W., Bhattacharyya, S., et al. (2018). Fibronectin EDA forms the chronic fibrotic scar after contusive spinal cord injury. Neurobiol. Dis. 116, 60–68. doi: 10.1016/j.nbd.2018.04.014
Courtine, G., Song, B., Roy, R. R., Zhong, H., Herrmann, J. E., Ao, Y., et al. (2008). Recovery of supraspinal control of stepping via indirect propriospinal relay connections after spinal cord injury. Nat. Med. 14, 69–74. doi: 10.1038/nm1682
Cugurra, A., Mamuladze, T., Rustenhoven, J., Dykstra, T., Beroshvili, G., Greenberg, Z. J., et al. (2021). Skull and vertebral bone marrow are myeloid cell reservoirs for the meninges and CNS parenchyma. Science 373:eabf7844. doi: 10.1126/science.abf7844
Donnelly, D. J., and Popovich, P. G. (2008). Inflammation and its role in neuroprotection, axonal regeneration and functional recovery after spinal cord injury. Exp. Neurol. 209, 378–388. doi: 10.1016/j.expneurol.2007.06.009
Ertürk, A., Becker, K., Jährling, N., Mauch, C. P., Hojer, C. D., Egen, J. G., et al. (2012). Three-dimensional imaging of solvent-cleared organs using 3DISCO. Nat. Protoc. 7, 1983–1995. doi: 10.1038/nprot.2012.119
Fiederling, F., Hammond, L. A., Ng, D., Mason, C., and Dodd, J. (2021). Tools for efficient analysis of neurons in a 3D reference atlas of whole mouse spinal cord. Cell Rep. Methods 1:100074. doi: 10.1016/j.crmeth.2021.100074
Filli, L., Engmann, A. K., Zörner, B., Weinmann, O., Moraitis, T., Gullo, M., et al. (2014). Bridging the gap: a reticulo-propriospinal detour bypassing an incomplete spinal cord injury. J. Neurosci. 34, 13399–13410. doi: 10.1523/jneurosci.0701-14.2014
Frenkel, N., Poghosyan, S., van Wijnbergen, J. W., van den Bent, L., Wijler, L., Verheem, A., et al. (2023). Tissue clearing and immunostaining to visualize the spatial organization of vasculature and tumor cells in mouse liver. Front. Oncol. 13:1062926. doi: 10.3389/fonc.2023.1062926
Garcia-Ramirez, D. L., Ha, N. T. B., Bibu, S., Stachowski, N. J., and Dougherty, K. J. (2021). Spinal cord injury alters spinal Shox2 interneurons by enhancing excitatory synaptic input and serotonergic modulation while maintaining intrinsic properties in mouse. J. Neurosci. 41, 5833–5848. doi: 10.1523/jneurosci.1576-20.2021
Granier, C., Schwarting, J., Fourli, E., Laage-Gaupp, F., Hennrich, A. A., Schmalz, A., et al. (2020). Formation of somatosensory detour circuits mediates functional recovery following dorsal column injury. Sci. Rep. 10:10953. doi: 10.1038/s41598-020-67866-x
Hayashi, M., Hinckley, C. A., Driscoll, S. P., Moore, N. J., Levine, A. J., Hilde, K. L., et al. (2018). Graded arrays of spinal and supraspinal V2a interneuron subtypes underlie forelimb and hindlimb motor control. Neuron 97:e865, 869–884. doi: 10.1016/j.neuron.2018.01.023
Izmailov, A. A., Povysheva, T. V., Bashirov, F. V., Sokolov, M. E., Fadeev, F. O., Garifulin, R. R., et al. (2017). Spinal cord molecular and cellular changes induced by adenoviral vector- and cell-mediated triple gene therapy after severe contusion. Front. Pharmacol. 8:813. doi: 10.3389/fphar.2017.00813
Jacob, L., Boisserand, L. S. B., Geraldo, L. H. M., de Brito Neto, J., Mathivet, T., Antila, S., et al. (2019). Anatomy and function of the vertebral column lymphatic network in mice. Nat. Commun. 10:4594. doi: 10.1038/s41467-019-12568-w
Jalufka, F. L., Min, S. W., Platt, M. E., Pritchard, A. L., Margo, T. E., Vernino, A. O., et al. (2022). Hydrophobic and hydrogel-based methods for passive tissue clearing. Methods Mol. Biol. 2440, 197–209. doi: 10.1007/978-1-0716-2051-9_12
Jing, D., Zhang, S., Luo, W., Gao, X., Men, Y., Ma, C., et al. (2018). Tissue clearing of both hard and soft tissue organs with the PEGASOS method. Cell Res. 28, 803–818. doi: 10.1038/s41422-018-0049-z
Klingberg, A., Hasenberg, A., Ludwig-Portugall, I., Medyukhina, A., Männ, L., Brenzel, A., et al. (2017). Fully automated evaluation of total glomerular number and capillary tuft size in nephritic kidneys using lightsheet microscopy. J. Am. Soc. Nephrol. 28, 452–459. doi: 10.1681/ASN.2016020232
Livet, J., Weissman, T. A., Kang, H., Draft, R. W., Lu, J., Bennis, R. A., et al. (2007). Transgenic strategies for combinatorial expression of fluorescent proteins in the nervous system. Nature 450, 56–62. doi: 10.1038/nature06293
Masselink, W., Reumann, D., Murawala, P., Pasierbek, P., Taniguchi, Y., Bonnay, F., et al. (2019). Broad applicability of a streamlined ethyl cinnamate-based clearing procedure. Development 146:dev.166884. doi: 10.1242/dev.166884
Matryba, P., Kaczmarek, L., and Golab, J. (2019). Advances in ex situ tissue optical clearing. Laser Photonics Rev. 13:1800292. doi: 10.1002/lpor.201800292
Mazzitelli, J. A., Pulous, F. E., Smyth, L. C. D., Kaya, Z., Rustenhoven, J., Moskowitz, M. A., et al. (2023). Skull bone marrow channels as immune gateways to the central nervous system. Nat. Neurosci. 26, 2052–2062. doi: 10.1038/s41593-023-01487-1
McCreedy, D. A., Jalufka, F. L., Platt, M. E., Min, S. W., Kirchhoff, M. A., Pritchard, A. L., et al. (2021). Passive clearing and 3D Lightsheet imaging of the intact and injured spinal cord in mice. Front. Cell. Neurosci. 15:684792. doi: 10.3389/fncel.2021.684792
Meehan, C. F., Ford, T. W., and Kirkwood, P. A. (2020). Plasticity of thoracic interneurones rostral to a lateral spinal cord lesion. Exp. Neurol. 331:113361. doi: 10.1016/j.expneurol.2020.113361
Naqvi, S., Panghal, A., and Flora, S. J. S. (2020). Nanotechnology: a promising approach for delivery of neuroprotective drugs. Front. Neurosci. 14:494. doi: 10.3389/fnins.2020.00494
Neirinckx, V., Coste, C., Franzen, R., Gothot, A., Rogister, B., and Wislet, S. (2014). Neutrophil contribution to spinal cord injury and repair. J. Neuroinflammation 11:150. doi: 10.1186/s12974-014-0150-2
Oh, S. W., Harris, J. A., Ng, L., Winslow, B., Cain, N., Mihalas, S., et al. (2014). A mesoscale connectome of the mouse brain. Nature 508, 207–214. doi: 10.1038/nature13186
Pan, C., Cai, R., Quacquarelli, F. P., Ghasemigharagoz, A., Lourbopoulos, A., Matryba, P., et al. (2016). Shrinkage-mediated imaging of entire organs and organisms using uDISCO. Nat. Methods 13, 859–867. doi: 10.1038/nmeth.3964
Qi, Y., Yu, T., Xu, J., Wan, P., Ma, Y., Zhu, J., et al. (2019). FDISCO: advanced solvent-based clearing method for imaging whole organs. Sci. Adv. 5:eaau8355. doi: 10.1126/sciadv.aau8355
Ragan, T., Kadiri, L. R., Venkataraju, K. U., Bahlmann, K., Sutin, J., Taranda, J., et al. (2012). Serial two-photon tomography for automated ex vivo mouse brain imaging. Nat. Methods 9, 255–258. doi: 10.1038/nmeth.1854
Renier, N., Adams, E. L., Kirst, C., Wu, Z., Azevedo, R., Kohl, J., et al. (2016). Mapping of brain activity by automated volume analysis of immediate early genes. Cell 165, 1789–1802. doi: 10.1016/j.cell.2016.05.007
Renier, N., Wu, Z., Simon, D. J., Yang, J., Ariel, P., and Tessier-Lavigne, M. (2014). iDISCO: a simple, rapid method to immunolabel large tissue samples for volume imaging. Cell 159, 896–910. doi: 10.1016/j.cell.2014.10.010
Richardson, D. S., Guan, W., Matsumoto, K., Pan, C., Chung, K., Ertürk, A., et al. (2021). Tissue clearing. Nat. Rev. Methods Primers 1:84. doi: 10.1038/s43586-021-00080-9
Rocha, M. D., Düring, D. N., Bethge, P., Voigt, F. F., Hildebrand, S., Helmchen, F., et al. (2019). Tissue clearing and light sheet microscopy: imaging the Unsectioned adult Zebra finch brain at cellular resolution. Front. Neuroanat. 13:13. doi: 10.3389/fnana.2019.00013
Romer, S. H., Seedle, K., Turner, S. M., Li, J., Baccei, M. L., and Crone, S. A. (2017). Accessory respiratory muscles enhance ventilation in ALS model mice and are activated by excitatory V2a neurons. Exp. Neurol. 287, 192–204. doi: 10.1016/j.expneurol.2016.05.033
Saritas, T., Puelles, V. G., Su, X. T., Ellison, D. H., and Kramann, R. (2019). Optical clearing and imaging of immunolabeled kidney tissue. J. Vis. Exp. 149:e60002. doi: 10.3791/60002
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: an open-source platform for biological-image analysis. Nat. Methods 9, 676–682. doi: 10.1038/nmeth.2019
Schlosser, P. M., Bale, A. S., Gibbons, C. F., Wilkins, A., and Cooper, G. S. (2015). Human health effects of dichloromethane: key findings and scientific issues. Environ. Health Perspect. 123, 114–119. doi: 10.1289/ehp.1308030
Shen, X. Y., Tao, C. L., Ma, L., Shen, J. H., Li, Z. L., Wang, Z. G., et al. (2021). Influence of spinal cord injury on core regions of motor function. Neural Regen. Res. 16, 567–572. doi: 10.4103/1673-5374.293158
Spalteholz, W. Über das durchsichtigmachen von menschlichen und tierischen präparaten und seine theoretischen bedingungen. Leipzig. (1914).
Susaki, E. A., Tainaka, K., Perrin, D., Kishino, F., Tawara, T., Watanabe, T. M., et al. (2014). Whole-brain imaging with single-cell resolution using chemical cocktails and computational analysis. Cell 157, 726–739. doi: 10.1016/j.cell.2014.03.042
Susaki, E. A., Tainaka, K., Perrin, D., Yukinaga, H., Kuno, A., and Ueda, H. R. (2015). Advanced CUBIC protocols for whole-brain and whole-body clearing and imaging. Nat. Protoc. 10, 1709–1727. doi: 10.1038/nprot.2015.085
Treweek, J. B., Chan, K. Y., Flytzanis, N. C., Yang, B., Deverman, B. E., Greenbaum, A., et al. (2015). Whole-body tissue stabilization and selective extractions via tissue-hydrogel hybrids for high-resolution intact circuit mapping and phenotyping. Nat. Protoc. 10, 1860–1896. doi: 10.1038/nprot.2015.122
Veldman, M. B., Park, C. S., Eyermann, C. M., Zhang, J. Y., Zuniga-Sanchez, E., Hirano, A. A., et al. (2020). Brainwide genetic sparse cell labeling to illuminate the morphology of neurons and glia with Cre-dependent MORF mice. Neuron 108, 111–127.e6. doi: 10.1016/j.neuron.2020.07.019
Verebélyi, K., and Iván, B. (2012). Cationic polymerization of styrene by the TiCl /N,N,N′,N′-tetramethylethylenediamine (TMEDA) catalyst system in benzotrifluoride, an environmentally benign solvent, at room temperature. Polymer 53, 3426–3431. doi: 10.1016/j.polymer.2012.05.055
Viswanathan, S., Williams, M. E., Bloss, E. B., Stasevich, T. J., Speer, C. M., Nern, A., et al. (2015). High-performance probes for light and electron microscopy. Nat. Methods 12, 568–576. doi: 10.1038/nmeth.3365
Wang, Z., Maunze, B., Wang, Y., Tsoulfas, P., and Blackmore, M. G. (2018). Global connectivity and function of descending spinal input revealed by 3D microscopy and retrograde transduction. J. Neurosci. 38, 10566–10581. doi: 10.1523/jneurosci.1196-18.2018
Yang, T., Dai, Y., Chen, G., and Cui, S. (2020). Dissecting the dual role of the glial scar and scar-forming astrocytes in spinal cord injury. Front. Cell. Neurosci. 14:78. doi: 10.3389/fncel.2020.00078
Yang, B., Treweek, J. B., Kulkarni, R. P., Deverman, B. E., Chen, C. K., Lubeck, E., et al. (2014). Single-cell phenotyping within transparent intact tissue through whole-body clearing. Cell 158, 945–958. doi: 10.1016/j.cell.2014.07.017
Zavvarian, M.-M., Hong, J., and Fehlings, M. G. (2020). The functional role of spinal interneurons following traumatic spinal cord injury. Front. Cell. Neurosci. 14:127. doi: 10.3389/fncel.2020.00127
Zholudeva, L. V., Abraira, V. E., Satkunendrarajah, K., McDevitt, T. C., Goulding, M. D., Magnuson, D. S. K., et al. (2021). Spinal interneurons as gatekeepers to neuroplasticity after injury or disease. J. Neurosci. 41, 845–854. doi: 10.1523/JNEUROSCI.1654-20.2020
Zholudeva, L. V., Karliner, J. S., Dougherty, K. J., and Lane, M. A. (2017). Anatomical recruitment of spinal V2a interneurons into phrenic motor circuitry after high cervical spinal cord injury. J. Neurotrauma 34, 3058–3065. doi: 10.1089/neu.2017.5045
Keywords: optical tissue clearing, spinal cord injury, lightsheet imaging, neutrophils, interneurons
Citation: Buxton AW, Jalufka FL, Hruska ME, Kubaney JR and McCreedy DA (2025) Rapid and efficient optical tissue clearing for volumetric imaging of the intact and injured spinal cord in mice. Front. Neurosci. 19:1601360. doi: 10.3389/fnins.2025.1601360
Edited by:
Munehisa Shinozaki, Keio University, JapanReviewed by:
Qi Zhang, Florida Atlantic University, United StatesCaleb Padgett, Augusta University, United States
Copyright © 2025 Buxton, Jalufka, Hruska, Kubaney and McCreedy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dylan A. McCreedy, ZG1jY3JlZWR5QGJpby50YW11LmVkdQ==
†These authors have contributed equally to this work