 Luis Armando Valenzuela-Ahumada1
Luis Armando Valenzuela-Ahumada1 Octavio Fabián Mercado-Gómez2
Octavio Fabián Mercado-Gómez2 Rubi Viveros-Contreras3
Rubi Viveros-Contreras3 Rosalinda Guevara-Guzmán2*
Rosalinda Guevara-Guzmán2* Alberto Camacho-Morales1*
Alberto Camacho-Morales1*- 1Department of Biochemistry, College of Medicine, Universidad Autónoma de Nuevo León, Monterrey, Mexico
- 2Department of Physiology, College of Medicine, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 3Centro de Investigaciones Biomédicas, Universidad Veracruzana, Xalapa, Mexico
Neurodegenerative diseases affect up to 349.2 million individuals worldwide. Preclinical and clinical advances have documented that altered energy homeostasis and mitochondria dysfunction is a hallmark of neurological disorders. Diet-derived ceramides species might target and disrupt mitochondria function leading to defective energy balance and neurodegeneration. Ceramides as bioactive lipid species affect mitochondria function by several mechanism including changes in membrane chemical composition, inhibition of the respiratory chain, ROS overproduction and oxidative stress, and also by activating mitophagy. Promising avenues of intervention has documented that intermittent fasting (IF) is able to benefit and set proper energy metabolism. IF is an eating protocol that involves alternating periods of fasting with periods of eating which modulate ceramide metabolism and mitochondria function in neurons. This review will address the detrimental effect of ceramides on mitochondria membrane composition, respiratory chain, ROS dynamics and mitophagy in brain contributing to neurodegeneration. We will focus on effect of IF on ceramide metabolism as a potential avenue to improve mitochondria function and prevention of neurodegeneration.
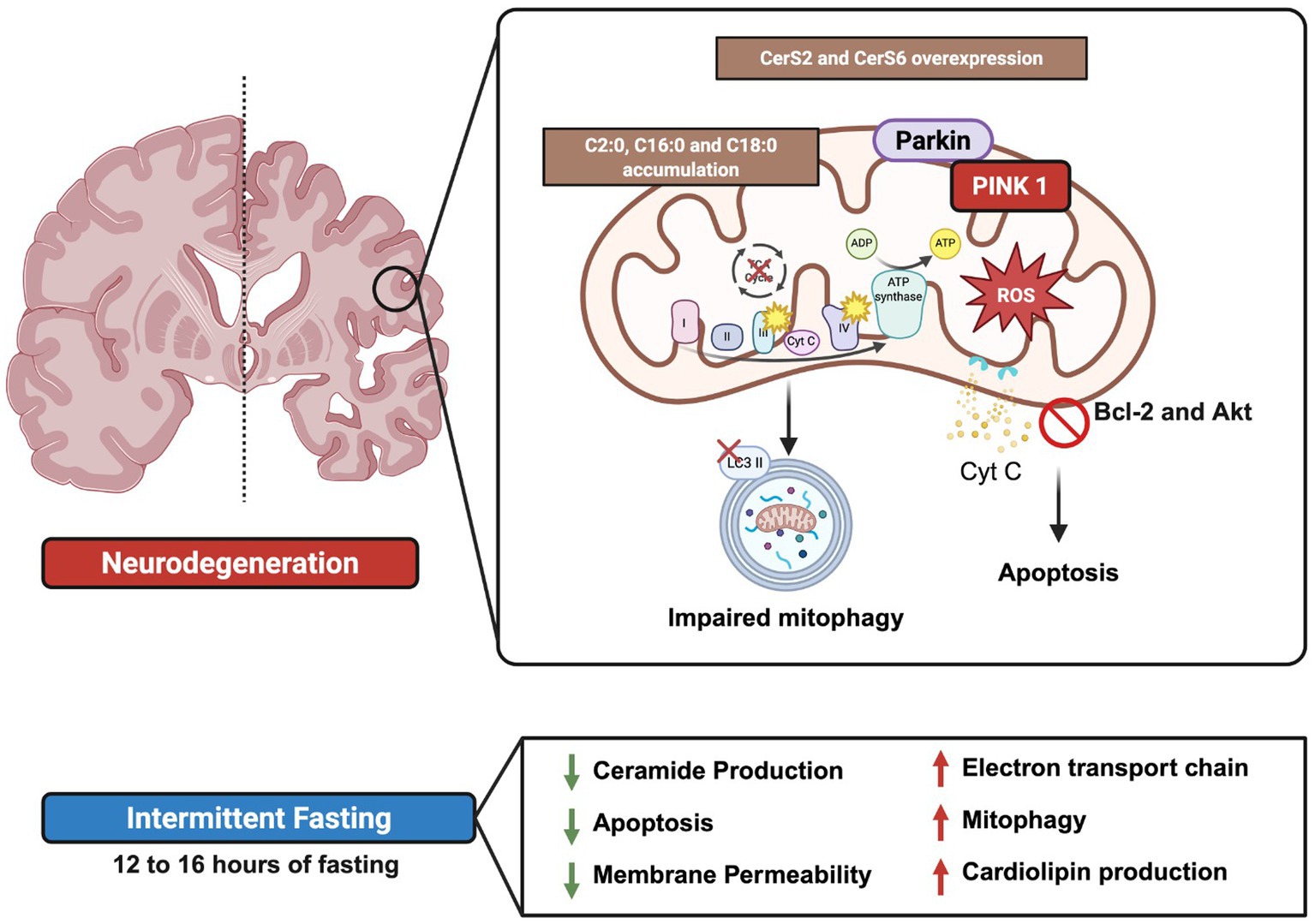
Graphical Abstract.
1 Introduction
Brain is a major body organ that integrate several neurodegenerative diseases including Alzheimer disease (AD), Parkinson disease (PD), primary tauopathies, frontotemporal dementia (FTD), amyotrophic lateral sclerosis (ALS), synucleinopathies (such as Lewy body dementia [LBD] and multisystem atrophy [MSA]), polyglutamine-related diseases (Huntington disease (HD) and spinocerebellar ataxias [SCA]), prion disease (PrD), traumatic brain injury (TBI), chronic traumatic encephalopathy (CTE), spinal cord injury (SCI), and multiple sclerosis (MS) (Wilson et al., 2023). According to the incidence, prevalence, and social impact up to 349.2 million individuals were affected by major neurological disorders worldwide (Ding et al., 2022; Huang et al., 2023). Recent reports documented the eight hallmarks of neurological disorders such as: pathological protein aggregation, synaptic and neuronal network dysfunction, aberrant proteostasis, cytoskeletal abnormalities, DNA and RNA defects, inflammation, neuronal cell death and altered energy homeostasis and mitochondria function (Wilson et al., 2023). While multifactorial triggers contribute to susceptibility to neurological disorders, defective in brain energetic deeply affect brain function. Accordingly, brain demands a high energy supply provided by glucose metabolism and mitochondria supporting synaptic transmission and maintain the plasma resting potential (Camacho and Massieu, 2006; Hamzeh et al., 2023). A decrease in glucose supply or pharmacologic inhibition of glucose or mitochondria metabolism into the brain, result in neurodegeneration (Shichkova et al., 2024; Camacho et al., 2006, 2007; Kazemeini et al., 2024).
Preclinical and clinical models documented that diet-derived ceramides might target and disrupt mitochondria function leading to neurodegeneration (Hamzeh et al., 2023). Ceramides are bioactive lipid species that serve as precursors of all complex sphingolipids (SLs) (Bernal-Vega et al., 2023). Ceramides are structural components in cell and mitochondrial membranes and modulates several cellular signaling events such as cell growth, cycle, death, and senescence, inflammation, immune modulation, cell adhesion, and migration, autophagy and stress response (Bernal-Vega et al., 2023). We and others have reported that ceramides affect mitochondria function favoring changes in membrane chemical composition, inhibition of the respiratory chain, overproduction of ROS reducing intracellular ATP production and causing oxidative stress and mitophagy in brain (Camacho-Morales et al., 2024). Accordingly, subjects diagnosed with neurodegenerative diseases have reported ceramide accumulation. For instance, plasma ratio of very long (C22-24) to long (C16-18) ceramides were found in AD susceptibility (McGrath et al., 2020). Also, elevated Cer species were found in the lesion area at early stages of AD (Han et al., 2002; Chowdhury et al., 2022; Reveglia et al., 2023). This evidence supports the notion that ceramides lipid species promote mitochondria dysfunction in brain which potentially might contribute to neurodegenerative susceptibility in subjects.
Intermittent fasting (IF) is an eating protocol that involves alternating periods of fasting with periods of eating and has recently gained popularity as a strategy for reducing body weight (Erdem et al., 2022; Steger et al., 2021). Furthermore, IF has been reported to decrease circulating levels of inflammatory markers in healthy subject (Faris et al., 2012; Perez-Kast and Camacho-Morales, 2025), as well as the inflammatory state associated with obesity (Zouhal et al., 2020). Also, IF decrease plasma levels of ceramide C17, C22, and C24 sphingomyelin, as well as C20, C22, C24, and C24:1 dihydrosphingomyelin (Madkour et al., 2023). IF also regulate the peroxisome proliferator-activated receptor γ coactivator 1α (PGC-1α), a transcription factor of mitochondria function, in neurons (Wilhelmi de Toledo et al., 2020; Mattson et al., 2018). IF upregulate the brain-derived neurotrophic factor (BDNF), favoring neurogenesis, synaptic plasticity and mitochondrial biogenesis (Mattson et al., 2018). This evidence supports that IF can prevent inflammatory response and enhance mitochondria functionality in individuals with chronic inflammation of elevated risk of poor cognitive function.
In this review, we will address the effect of IF on ceramide metabolism linked to mitochondria dynamics in brain and its potential effects on neurodegeneration. We will focus on the effect of ceramide assisting lipid composition of the mitochondria membrane, mitochondrial respiratory chain activity, ROS overproduction, oxidative stress and changes in mitophagy – apoptosis, and how these are mutually related. We aim this review within the framework of neurodegenerative diseases, which integrate the highest incidence and prevalence of mental illnesses in the population. The manuscript will describe findings in animal models (mainly rats and mice) and contrasting them with what has been reported so far in humans.
2 Ceramide in brain function and neurodegeneration
Ceramides are a family of bioactive lipids composed of a long-chain sphingoid base linked to a fatty acid chain through an amide bond. They vary in structure based on the length, hydroxylation, and saturation levels of both the sphingoid base and the fatty acid residues (Alonso and Goñi, 2018). Ceramides are predominantly found in the liver, adipose tissue, and neural tissue, where they modulate several signaling pathways. Three major metabolic pathways are involved in ceramide biosynthesis: the de novo, the salvage pathway and the sphingomyelinase pathway. The De Novo Pathway is located on the cytoplasmic surface of the endoplasmic reticulum (ER) and integrate the most important source of ceramides. The De Novo Pathway begins with the condensation of L-serine and a fatty-acyl CoA such as palmitoyl-CoA, myristoyl CoA, or stearoyl CoA at the endoplasmic reticulum catalyzed by the enzyme serine palmitoyltransferase (SPT) to form 3-ketosphinganine, which is then reduced to dihydrosphingosine by the NADH-dependent 3-ketosphinganine reductase (KDSR). Ceramide synthases (CerS) catalyze the acylation of dihydrosphingosine to dihydroceramide. According to their carbon chain lengths, six CerS isoforms have been reported in mammals displaying selective specificity for acyl CoAs. CerS1 attaches the C18 fatty acyl CoA (long chain); CerS2 exhibits activity toward the very long acyl CoAs such as C22–C24; CerS3 attaches the ultra-long fatty acyl CoA as C26, CerS4 specifies the C18–C20 CoA; and CerS5 and 6 have specificity for the C14–C16 CoA (Bernal-Vega et al., 2023). Ceramide synthesis is responsive to tissue-dependent distribution of ceramide synthases. For instance, CerS1 is mainly expressed in the brain, CerS2 is highly abundant in the liver and kidney, high ex- pression of CerS3 is found in the testis and skin, and elevated levels of CerS4 mRNA are found in the skin, heart, liver, and leukocytes, CerS5 is mainly expressed in lung epithelia and brain, whereas CerS6 show high expression in the intestine (Brachtendorf et al., 2019; Levy and Futerman, 2010; Bernal-Vega et al., 2023). Interestingly, Canals and Clarke (2022) found that mRNA expression levels of CerS do not directly correlate with the concentrations of ceramides with specific chain lengths. Finally, ceramide is produced by a desaturation reaction assisted by dihydroceramide desaturase (DDase) (Quinville et al., 2021; Bernal-Vega et al., 2023). This evidence suggests that ceramides synthesis is responsive to tissue-dependent distribution of enzymes and which are post-transcriptionally modulated by unknown mechanisms (Mullen et al., 2012).
In the salvage pathway, ceramide is generated from partial degradation of membrane glycolipids by glucosylceramidase (GCase). Also, in the sphingomyelin pathway complex sphingolipids are hydrolyzed by sphingomyelinases (SMase) in lysosomes. Next, ceramide is transported to the Golgi complex through vesicular and non-vesicular protein ATP-dependent coupling allowing the production of complex SLs. Finally, ceramide can follow four major catabolic pathways: (1) phosphorylated by the ceramide kinase (CERK) to form ceramide-1-phosphate (C1P), (2) incorporated into sphingomyelin-by-sphingomyelin synthase (SMS) activity, (3) or into glucosylceramide by the activation of glucosylceramide synthases (GCS), leading to the generation of complex SLs like cerebrosides, gangliosides, sulfatides and globosides; and (4) ceramides are deacylated by the ceramidases (CDase) to produce sphingosine. Sphingosine kinases (SKs) phosphorylate sphingosine to produce sphingosine-1-phosphate (S1P), which is degraded by sphingosine phosphate phosphatase (SPP) or S1P lyase synthesizing. The genes encoding CerS enzymes differ in their spatial and temporal expression patterns, as well as in their ability to produce ceramides with varying chain lengths. Although all identified CerS enzymes exhibit similar Km values (affinity for substrates), they vary in their selectivity for acyl-CoAs depending on the length of the acyl chain (Bernal-Vega et al., 2023).
The role of ceramides in the brain is essential for maintaining the structure and function of cell membranes, contributing to their integrity, fluidity and stability (Mencarelli and Martinez-Martinez, 2013). For instance, C24:0 ceramide maintains cell membrane integrity by enhancing lipid packing and promoting a more rigid membrane structure, thereby regulating permeability. Conversely, replacing C24:0 ceramide with shorter-chain ceramides like C16:0 increased membrane permeability, underscoring the importance of very long-chain ceramides in barrier function (DiPasquale et al., 2022; Ventura et al., 2020). In fact, ceramides are expressed in oligodendrocytes, where they support myelin integrity. The ceramides that serve as precursors for complex sphingolipids essential to myelin sheath formation. Initial reports documented active sphingolipid metabolism of C2:0 and C6:0 ceramides to Sph and S1P in differentiated oligodendrocytes (Qin et al., 2010). Exogenous S1P is efficiently metabolized back to Sph and predominantly C18-ceramide supporting myelin confirmation. Oligodendrocytes also accumulate sphingomyelin and GalCer and C22:0, C24:0 and C24:1 ceramides are found in oligodendrocytes (Hammerschmidt and Brüning, 2022) (Figure 1).
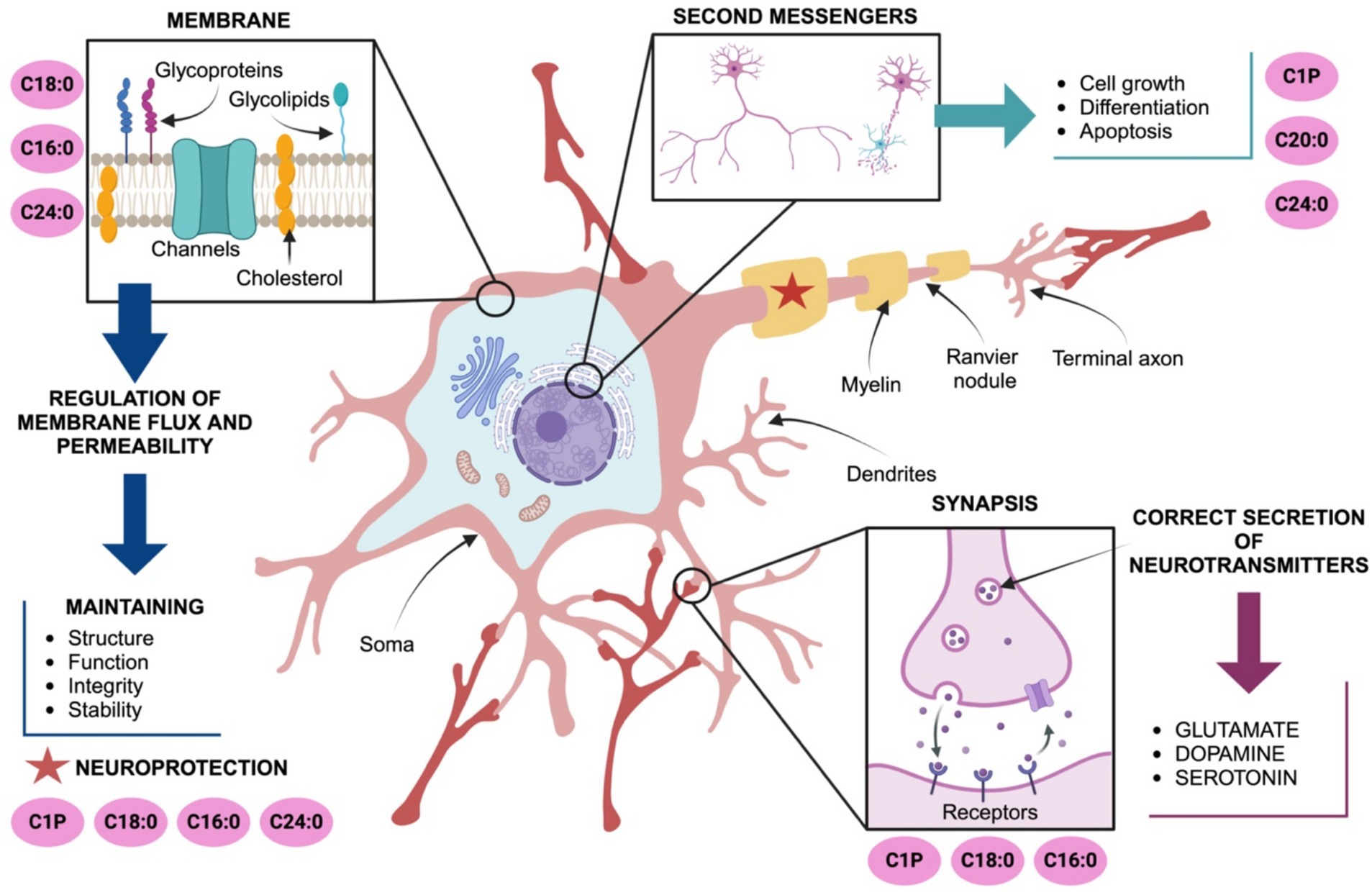
Figure 1. Ceramides modulate neuronal development, function, and protection. C18:0, C16:0 and C24:0 maintain plasma membrane integrity and fluidity. C20:0, C24:0 and ceramide-1-phosphate (C1P) are also involved in the fine regulation of processes including cell growth, differentiation, neuroplasticity and apoptosis. C24:0 ceramide supports myelination and axonal health, indirectly protecting neuronal endings and facilitating signal propagation to synaptic terminals. Ceramides, particularly C16:0, C18:0, and C24:0, play significant roles in modulating neurotransmitter release coordinating excitotoxicity and ensuring proper neuronal communication. Abbreviations: ceramide-1-phosphate (C1P).
Beyond these structural roles, ceramides are also involved in intracellular signaling, acting as second messengers in critical processes such as cell growth, differentiation, apoptosis, and stress responses essential for neuronal survival and synaptic function. C16:0 ceramide maintains synaptic integrity and plasticity, especially under conditions of metabolic stress or excess fatty acids assuring memory and learning (Furuya et al., 1998). Also, C20:0 and C24:0 ceramides are critical during neuronal development maintaining neuronal networks, modulating neurite outgrowth and axon guidance (Olsen and Færgeman, 2017; Chaurasia et al., 2019; Mullen et al., 2012). The ceramide-1-phosphate (C1P), a phosphorylated derivative of ceramide, favors cell survival, proliferation, and anti-apoptotic signaling through the activation of PI3K/Akt, ERK1/2, and NF-κB pathways. C1P also modulates inflammatory responses and inhibits acid sphingomyelinase activity supporting neuronal survival (Furuya et al., 1998; Gómez-Muñoz, 2006). C1P and C2 ceramides also regulate the dynamics of the cytoskeleton and axonal guidance by their effects on G-protein coupled receptors coupled to Rho GTPases signaling. Additionally, C1P modulates the activity of neurotrophic factors, such as BDNF (brain-derived neurotrophic factor), which are vital for axon growth, survival, and synaptic plasticity (Furuya et al., 1998; Gómez-Muñoz, 2006) (Figure 1).
Furthermore, in neurons ceramides regulate neurotransmitter release, particularly modulating glutamate, dopamine, and serotonin, which are vital for neuronal communication and synaptic plasticity (Farsi et al., 2021; López et al., 2025). Under normal conditions, C16:0 ceramide, produced by Ceramide Synthase 6 (CerS6), plays a crucial role in synaptic glutamate release by organizing lipid rafts configuration assuring clustering of the soluble N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) complex and vesicle fusion (Sapoń et al., 2023). In fact, C16:0 acts as a second messenger, activating protein kinase C (PKCζ) and promoting Ca2+-dependent exocytosis of glutamate (Wheeler et al., 2009), supporting short-term plasticity. C18:0 ceramide is enriched in hippocampal and cortical neurons (Qin et al., 2010) where higher basal levels of C18:0 ceramide correlate with efficient dopamine release and signaling (Schoffelmeer et al., 2000). C1P regulates the function of serotonin receptors and influence serotonin-related cellular signaling (Ji et al., 2011). C1P may also influence serotonin production by regulating the activity of tryptophan hydroxylase possibly through changes in cellular stress responses or phosphorylation (Sinenko et al., 2023). These evidences documented the role of C16:0, C18:0, and C1P:0 on neurotransmitter release and synaptic plasticity.
Dysregulation of ceramide metabolism in the brain has been linked to several brain pathologies, including neurodegenerative diseases such as Alzheimer’s and Parkinson’s, and psychiatric disorders like anxiety disorders, depression, and schizophrenia, and neurological injuries (Sambolín-Escobales et al., 2022; Schumacher et al., 2022; Mielke et al., 2013; Liu et al., 2024; Afridi et al., 2022). Ceramides promote neuroprotection by modulating stress, including oxidative stress, inflammation, and excitotoxicity in neurons and glia. In neurons, ceramides support the coating of neuronal endings, safeguarding synaptic connections (Mencarelli and Martinez-Martinez, 2013). Ceramides are also expressed in glia cells. In a recent report McInnis et al. (2024) documented high expression of C24:0 ceramide in astrocytes and C24:1 in microglia when compare with C16:0 and C18:0 ceramides in neurons. Also, S1P modulates the translocation of NF-κB to the nucleus in astrocytes (Rajagopalan et al., 2015) leading to experimental autoimmune encephalomyelitis (Linnerbauer et al., 2020). LacCer activates the IRF-1 and NF-κB expression favoring proinflammatory gene profiles in astrocytes (Giovannoni and Quintana, 2020). Accordingly, exposure microglia to palmitic acid (C16:0/PA) enhanced the expression of proinflammatory markers (Duffy et al., 2015), and a decreased in C22:0–C24:0 ceramides by deletion of CerS2 triggered the activation of microglial cells and progressive myelin atrophy (Teo et al., 2023). Potential molecular mechanisms contributing to demyelination and neurodegeneration suggest that ceramides C18:0, C22:0, C24:0 and C24: activate the neuronal NADPH oxidase 2 (NOX2)1 (Arsenault et al., 2021) or endoplasmic reticulum stress response (García-González et al., 2018). These evidences support the role of ceramides on glia function.
Finally, ceramides might be synthetized by mitochondria itself. For instance, initial reports in murine models identified three ceramide synthase subtypes CerS1/2/6 in the mitochondria of mouse brain (Novgorodov et al., 2011; S. Ding et al., 2024). In fact, CerS2 co-localizes with the translocase of outer mitochondrial membrane 20 (TOM20) on the outer mitochondria membrane and the CerS6 associates with the adenine nucleotide translocase (ANT) on the inner mitochondria membrane (Yabu et al., 2009; Ding et al., 2024). In addition, mitochondria also recruit additional enzymes related to ceramide metabolism including neutral sphingomyelinase (nSMase) (Novgorodov et al., 2011; Ding et al., 2024) and neutral ceramidase (Rajagopalan et al., 2015; Ding et al., 2024). This suggests that mitochondria by itself integrate the enzymatic machinery to properly regulate ceramides and assist its biological activity potentially by its interaction to ER.
Accumulation of selective ceramides species might target and disrupt mitochondria function leading to neurodegeneration (Lima et al., 2023). In an elegant preclinical report authors documented accumulation of long-chain and very long-chain ceramides in brain of 5xFAD mice, which was associated with mitochondrial dysfunction (Crivelli et al., 2024). This evidence suggest that defective ceramide metabolism might target mitochondria favoring neurodegeneration. We next describe the role of ceramides on mitochondria dysfunction assisting neurodegeneration.
3 Ceramides and mitochondrial damage
Mitochondria are dynamic organelles that use oxidative phosphorylation to supply energy by producing adenosine triphosphate, which is indispensable for life and health in most eukaryotic organisms (Luo et al., 2020). Mitochondria display critical roles including metabolic homeostasis, redox signaling, regulation of Ca2+, mitochondrial-nuclear communication and cell death (Chen et al., 2023). When mitochondrial dysfunction occurs in cells, homeostasis is coordinating by mitophagy which balance a physiological response of mitochondrial biogenesis and clearance of damaged mitochondria (Luo et al., 2020). In fact, mitochondrial turnover and dynamics are precisely coordinated by a balance between fission—fusion processes (Chen et al., 2023).
Dysregulation of mitochondrial dynamics is one key pathogenic mechanism of diverse human pathologies including neurodegenerative diseases (Luo et al., 2020). Accordingly, ceramides species might target mitochondria in brain favoring susceptibility to neurodegeneration. Major advances addressing the role of mitochondria on neurodegenerative diseases have been mainly reported in two neurodegenerative diseases, Parkinson and Alzheimer (Alecu and Bennett, 2019). Although the precise molecular mechanisms remain largely elusive, we will describe how ceramides target mitochondria function in brain by several mechanisms: (1) changes in the lipid composition of the mitochondria membrane, (2) alteration in mitochondrial respiratory chain activity and increased in ROS production, and (3) changes in mitophagy—apoptosis process.
3.1 Ceramides affects the lipid composition of the mitochondria membranes
Mitochondrial membrane lipids are essential for several mitochondria processes such as membrane architecture, transport of proteins into the matrix, activity of respiratory proteins, mitochondrial fission and fusion, and contribution of lipids to endoplasmic reticulum related through contacts by the mitochondria-associated membranes (MAMs) (Böttinger et al., 2016; Aufschnaiter et al., 2017). The major lipid species in mitochondria included up to 40% phosphatidylcholine (PC), 27% phosphatidylethanolamine (PE), 3% phosphatidylserine (PS), 15% phosphatidylinositol (PI), 2% phosphatidic acid (PA) and 13% cardiolipin (CL) (Aufschnaiter et al., 2017). Lipidomic profiles into the outer mitochondrial membrane, included up to 46% PC, 33% PE, 1% PS, 10% PI, 2% PA and 6% CL; whereas in the inner mitochondrial membrane it is found 38% PC, 24% PE, 4% PS, 16% PI, 2% PA and 16% CL (Aufschnaiter et al., 2017). Notably, ceramides content in mitochondria is located up to three-fold higher in the outer rather than in the inner mitochondrial membrane, which potentially might regulate the formation of protein-permeable channels (Siskind and Colombini, 2000). MAMs also integrates a subcellular compartmentalization of ceramide metabolism between mitochondria and endoplasmic reticulum capable of modulate cellular responses (Bionda et al., 2004). In fact, in an elegant study authors proposed an evolutionary scenario to explain mitochondria lipid metabolism, suggesting that mitochondria are outcomes of a merger between (alphaproteo-) bacteria and archaea showing ancestral combination of two main genes for lipid synthesis: cardiolipin and ceramide (Geiger et al., 2023). This evidence confirms that lipidomic profile and ceramide content in mitochondria membranes might modulate cellular responses and contributes to cellular dysfunction by disrupting mitochondrial integrity, morphology and function (Figure 2).
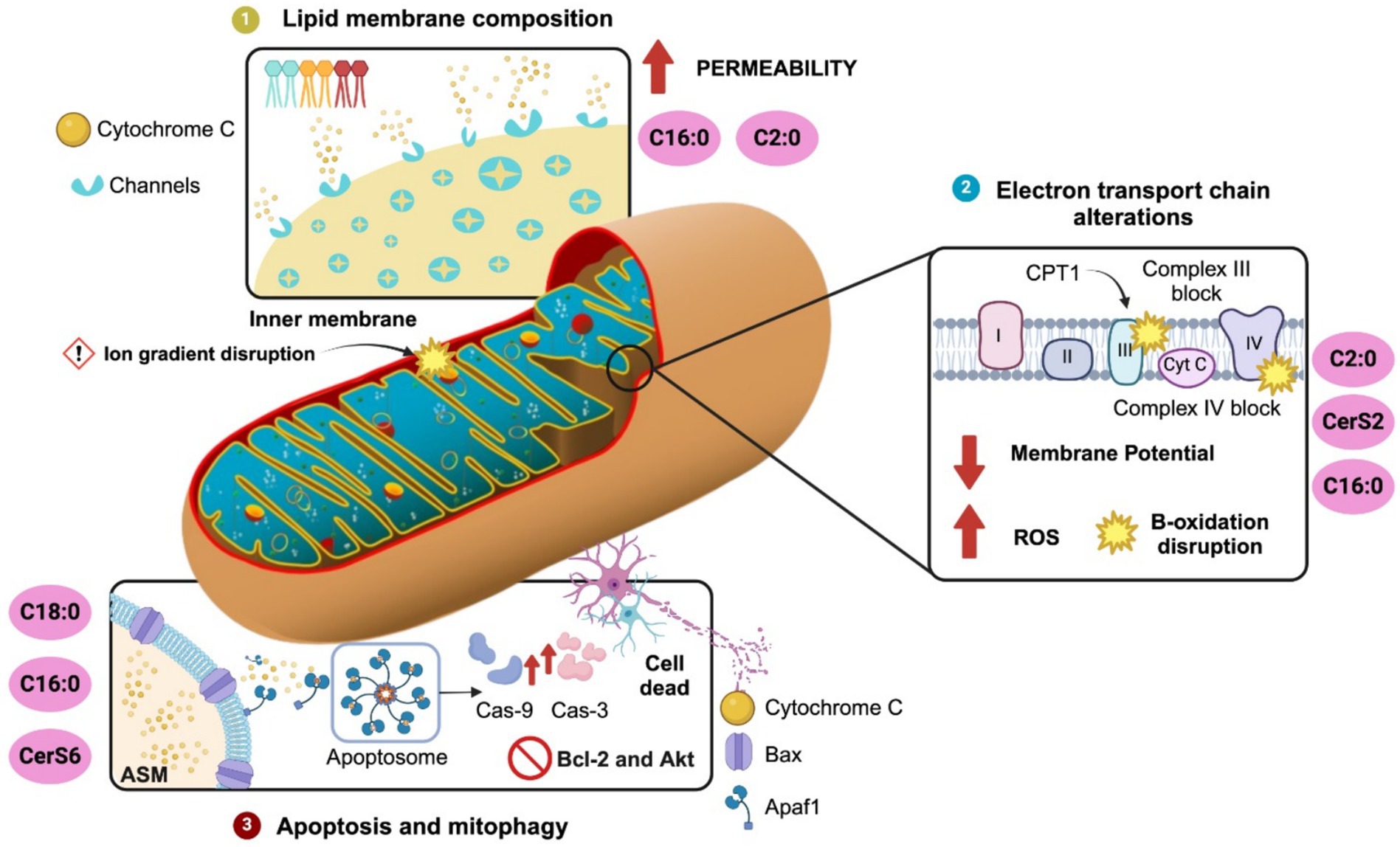
Figure 2. Ceramide-driven mitochondrial regulation. Ceramides modulates mitochondria dynamics by several pathways. (1) Lipid membrane composition: C2:0 and C16:0 ceramides increase mitochondrial membrane permeability by integrating channels into the inner mitochondria membrane assisting cytochrome c release. (2) Disruption of electron transport chain: C2:0 and C16:0 ceramides or CerS2 overexpression promoted ROS production via mPTP interaction and Complex III inhibition (antimycin A site). Also, C16:0 inhibits Complex IV and CPT1, disrupting β-oxidation. (3) Ceramides activates apoptosis and mitophagy: C16:0 and C18:0 promoted Bcl-2 depletion and Akt inhibition and activated apoptosis by MOMP through Bcl-xL-sensitive channel formation and interaction with Bax/VDAC. C18:0 ceramide, produced by CerS1 transported via p17/PERMIT, triggers mitophagy through LC3B-II independently of caspases. ASM and ceramides coordinate mitophagosome formation. Conversely, CerS6 impairs mitophagy via SQSTM1 interaction. Ceramide synthase 2, CerS2; Ceramide synthase 6, CerS6.
Initial reports documented that changes in ceramide species might affect lipid composition in the mitochondrial membrane. For instance, C16:0 and C2:0-ceramides might increase the permeability of mitochondria membranes releasing cytochrome c tentatively by channel formation (Siskind and Colombini, 2000). Mitochondrial permeability disrupts the ion gradients across the inner mitochondrial membrane, resulting in mitochondrial depolarization, a loss of oxidative phosphorylation, and an increase in ROS production (García-Ruiz et al., 1997). Mitochondrial permeability might be also facilitated by external factors such as mitochondrial calcium loading, pH changes, thiol oxidation, and activation of the adenine nucleotide translocator (ANT). In fact, researcher identified that mitochondrial permeability transition pore (mPTP) was sensitive to C2 ceramide incubation by promoting ROS overproduction (García-Ruiz et al., 1997). The accumulation of C16:0 ceramide also might alter the balance between sphingolipids, phospholipids, and other membrane constituents, leading to increased membrane rigidity and permeability (Fucho et al., 2017). In isolated liver mitochondria, C16:0 ceramides formed much higher membrane channels than the C22:0 ceramide (Stiban and Perera, 2015). It is proposed that membrane channels are formed from columns of ceramides that arrange in an anti-parallel fashion making a cylindrical shape spanning the hydrophobic interior of the mitochondrial outer membrane. Each column is composed of six Cer molecules associated to the MOMP and mitochondria permeabilization (Abou-Ghali and Stiban, 2015). The formation of ceramide-induced pores has been associated with interactions between ceramides and proteins such as Bax or the voltage-dependent anion channel (VDAC), which facilitate cytochrome c release and initiate apoptosis (Siskind et al., 2006; Pastorino et al., 2002) (Figure 2).
Finally, changes in the lipidomic profile of mitochondria membranes might compromise mitochondria permeabilization and cellular responses susceptible to neurodegeneration. A recent report documented accumulation of the ceramide precursor sphingosine and C20:0, C22:0, C22:1, C24:0 and C24:1 ceramide species in mitochondria isolated from both 5xFAD mice and AD patient brains (Crivelli et al., 2024). Also, defective CL synthesis, a lipid specie of ceramide metabolism, might alter mitochondrial cristae structure reported in AD and PD’s diseases (Argueti-Ostrovsky et al., 2024). Conversely, pharmacologic inhibition of the ceramide synthesis improved mitochondria morphology in lymphoid cells from patients with HD (Ciarlo et al., 2012) (Figure 2).
Together, changes in ceramide metabolism appears to modulate mitochondria membrane structure in neurodegenerative diseases. It is expected that membrane structure associates to mitochondrial dynamics and metabolism, we will address this topic in the next sections.
3.2 Ceramides affect the electron transport chain (ETC) and promote ROS generation
Mitochondria integrates the ETC to the oxidative phosphorylation to regulate cellular energy metabolism. The protein machinery of the ETC is composed of five enzyme complexes and two electron mobility carriers including the complex I (CI, NADH–ubiquinone oxidoreductase), complex II (CII, succinate dehydrogenase, SxDH), complex III (CIII, cytochrome bc1 oxidoreductase), complex IV (CIV, cytochrome c oxidase), ATP synthase, and the electron mobility carriers ubiquinone and cytochrome c (Kobayashi et al., 2023). Functionally, reducing equivalents from NADH and FADH provide electrons to the ETC which are finally accepted by oxygen to generate ATP. Accordingly, ROS such as the superoxide anion production originates from the Complex I and III, and can damage proteins, lipids or nucleic acids within the mitochondria and cell (Hadrava Vanova et al., 2020). Overexpression of CerS2 has been reported to reduce mitochondrial membrane potential compared to control groups (Qiu et al., 2018). Experimental evidence indicates that ceramide accumulation can reduce mitochondrial membrane potential by 30–50% in specific cell types (Novgorodov et al., 2008). This decline can be detected using fluorescent dyes such as TMRM or JC-1, which show a decrease in fluorescence when mitochondrial potential drops. Notably, C2-ceramide has been shown to induce a rapid reduction in mitochondrial membrane potential. For example, in rat cardiomyocytes, C2-ceramide caused a marked decrease in mitochondrial membrane potential, accompanied by fragmentation of the mitochondrial network (Parra et al., 2013). Further mechanistic insights were provided by Novgorodov et al. (2016), who explored the interaction between SIRT3, a mitochondrial deacetylase, and ceramide synthases (CerS1, CerS2, CerS6) in cerebral mitochondria. Their investigation using SIRT3 knockout mice revealed that the absence of SIRT3 led to increased mitochondrial ROS production and protein carbonylation, markers of oxidative stress and protein damage (Figure 2).
Several reports have documented the effect of ceramides accumulation on ETC and mitochondrial function in peripheral tissues and brain. For example, accumulation of ceramides in the mitochondria of skeletal muscle cells showed loss of the mitochondrial respiratory chain components, resulting in mitochondrial dysfunction and insulin resistance (Diaz-Vegas et al., 2023). Also, C2:0 ceramide block the activity of Complex III favoring ROS production, tentatively by electron transfer to molecular oxygen at or near the same site where antimycin A (AA) acts within the Q-cycle of Complex III (Fato et al., 2009; Gudz et al., 1997). C16:0 ceramide was also reported to inhibit the Complex IV activity in isolated liver mitochondria and induce ROS production (Zigdon et al., 2013). C16:0 ceramide also disrupts mitochondrial β-oxidation by inhibiting carnitine palmitoyltransferase 1 (CPT1), a key enzyme regulating the entry of long-chain fatty acids into mitochondria for oxidation. This inhibition results in a lipid overload state, where non-oxidized fatty acids are diverted into pathways generating more ceramides, perpetuating a self-amplifying cycle of metabolic stress and dysfunction (Serra et al., 2013). A recent report documented that pharmacologic accumulation of ceramide resulted in disruption in ETC and ROS production in human neurons (Kennedy et al., 2016). Crivelli et al. (2024) reported that elevated levels of 18:0, 22:0, and 24:1 ceramide reduced the ETC and increased ROS production in isolated synaptic and non-synaptic mitochondria from 5xFAD mice, a widely used Alzheimer’s disease model. This result resembles what it was found in recent reports documented that ceramide accumulation compromises mitochondria respiration in the cerebral cortex (Carr et al., 2023) (Figure 2).
3.3 Ceramides coordinate apoptosis and mitophagy
Ceramides have been implicated in the induction of apoptosis and mitophagy by several mechanisms. Ceramide accumulation can lead to mitochondrial and membrane permeabilization, triggering the release of cytochrome c into the cytoplasm favoring caspase-3-dependent apoptosis. Ceramides also activate apoptosis by depleting the anti-apoptotic proteins like Bcl-2 in neurons (Jazvinšćak Jembrek et al., 2015), inhibits the Akt/PKB survival pathway and activates the mitogen-activated protein kinases (MAPKs) (Iwayama and Ueda, 2013). As described, ceramides are capable of forming membrane channels in both planar phospholipid membranes and the outer membranes of isolated mitochondria, large enough to allow the passage of proteins released during mitochondrial outer membrane permeabilization (MOMP) (Fisher-Wellman et al., 2021). The anti-apoptotic protein Bcl-xL has been identified as a key regulator of this pathway, as it inhibits both MOMP in vivo and the formation of ceramide channels in vitro (Chang et al., 2015). During apoptosis, ceramides can affect mitochondria by ceramide-formed channels, favoring mitochondrial outer membrane permeability (MOMP), kinase activation, and inhibition of respiration (Stiban and Perera, 2015). Conversely, the ceramide-metabolite sphingosine-1-phosphate (S1P) inhibits apoptosis in endothelial cells (Bonnaud et al., 2007). Accordingly, a recent report documented that brain apoptosis faced an increase in the C16-ceramide levels potentially by the conversion from mitochondrial sphinganine and sphingomyelin (Mignard et al., 2020). Notably, C16-ceramide is enhanced in mitochondria when sphingomyelin levels are decreased in the MAMs microdomains (Mignard et al., 2020). These results propose that ceramide induces apoptosis by increasing mitochondrial Bax expression which supports the notion that ceramide assist channels formation. Also, ceramides seem to integrate a mutual dependent crosstalk between mitochondria – ER by MAMs microdomains during apoptosis regulating by C16-species (Figure 2).
Mitophagy and apoptosis are mutually regulated to secure cell integrity during neuronal death and survival. Maintenance of the mitochondrion pool is sensitive to both fusion and fission molecular machinery in a way to selectively identify and remove damaged mitochondria (Scheibye-Knudsen et al., 2015). Mitophagy, a form of autophagy, is responsible for the basal mitochondrial turnover that eliminates dysfunctional mitochondria. Notably, new evidence documented that mitochondria split into two separate types, one of which concentrates on energy production, the other on producing essential cellular building blocks (Ryu et al., 2024). Accordingly, mitochondrial fusion preserves the mitochondria for greater energy production during conditions of high metabolic activity, whereas fission facilitates the removal of damaged mitochondria via mitophagy (Scheibye-Knudsen et al., 2015). C18:0 ceramide specie is a stress-mediated mitophagy response which is mediated by the subcellular localization of CerS1 in damaged mitochondria vs. endoplasmic reticulum in the brain (Oleinik et al., 2023; Ryu et al., 2024). Recently, it was reported that mitophagy requires the subcellular interaction of CerS1 by the p17/PERMIT protein (17 kDa transporter) to secure proper mitochondrial quality control in the brain by mitophagy (Oleinik et al., 2023). Based on this model, CerS1 is transported to the outer mitochondrial membrane by the p17/PERMIT to recognize and induce C18:0 ceramide-mediated mitophagy in brain (Oleinik et al., 2023). In fact, CerS1 and its metabolic product, C18-ceramide (Dany and Ogretmen, 2015), induce non-apoptotic lethal mitophagy independent of Bax, Bal, or caspase activity in carcinoma cells (Sentelle et al., 2012). Interestingly, ectopic expression of CerS1 or treatment with C18-pyridinium-ceramide resulted in LC3-II formation and promoted its direct binding to ceramide on the mitochondrial membranes. This lipid-protein binding then allowed the mitochondria to be targeted by the LC3-II-containing autophagosome (Sentelle et al., 2012). Conversely, endogenous C16-ceramide generated by ceramide synthase 6 did not activate mitophagy-promoting function (Sentelle et al., 2012). These results support the role of ceramide signaling in mediating mitophagy through ceramide-LC3-II binding (Figure 2).
Mitophagy might be also regulated by acid sphingomyelinase (ASM), a ceramide metabolite enzyme. The ASM is activated by several cellular stresses implicated in both apoptosis and autophagy by modulating the balance between ceramide and its metabolic derivatives, sphingosine (Sph) and sphingosine-1-phosphate (S1P) (Lee et al., 2014). Precisely, ASM modulates the lysosomal nutrient-sensing complex (LYNUS), which integrate a mTOR and transcription factor EB (TFEB) mutual crosstalk. Justice et al. (2018) reported that ASM activity supports physiological mTOR signaling by preventing TFEB activation. Pharmacologic or genomic ASM inhibition enhanced nuclear translocation of TFEB and autophagic activity. Also, Oleinik et al. (2023) demonstrated that C18-ceramide promoted mitophagy through LC3B lipidation to form LC3B-II. In a recent paper, authors documented that oxidative stress targets ceramide to mitochondria by extracellular vesicles from the plasma membrane. Authors reported that elevation of ceramide by sphingomyelinase 2 and acid sphingomyelinase activity were target to mitochondria by ceramide-rich extracellular vesicles favoring cell death (Quadri et al., 2024). These results suggest that ceramides and ASM coordinate mitochondria and autophagosome membrane formation assisting mitophagy (Figure 2).
Defective mitophagy sensitive to ceramides has also been reported in PD (Vos et al., 2021). PINK (PTEN-induced putative kinase1)/Parkin pathway is the most well-characterized signaling that orchestrates mitochondrial degradation in PD (Doxaki and Palikaras, 2020). Under physiological conditions, the cytosolic PINK1 enters the mitochondria through the translocases of the outer and inner membranes complexes (Markaki et al., 2023). PINK1 is cleaved by mitochondrial processing peptidase and the presenilin-associated rhomboid-like protease and is finally completely degraded by an MG132-sensitive protease (Greene et al., 2012). Then, PINK1 is stabilized on the outer membrane and is activated by autophosphorylation, leading to the recruitment of cytosolic E3 ubiquitin ligase Parkin, which in turn, ubiquitinates several outer mitochondrial membrane proteins promoting mitochondrial (Sekine and Youle, 2018; Doxaki and Palikaras, 2020). In fact, homozygous loss of PINK1 or Parkin results in PD (Vos and Klein, 2022), supporting the role of PINK1 on mitochondria quality control. Of note, authors reported interaction between ceramides and the PINK1 protein affecting mitochondria function and mitophagy. Notably, defective PINK1 leads to decreasing beta-oxidation, increasing ceramide accumulation in mitochondria and mitophagy (Vos et al., 2021). Moreover, lower ceramide levels improved mitochondrial phenotypes in PINK1-mutant flies and PINK1-deficient patient-derived fibroblasts (Vos and Klein, 2022). Moreover, lower ceramide levels improved mitochondrial phenotypes in PINK1-mutant flies and PINK1-deficient patient-derived fibroblasts (Vos et al., 2021). Also, using other brain damage models, it has been described that intracerebral hemorrhage by hemin treatment increases the upregulation of ceramide synthase 6 (CerS6) and CerS6-derived C16 ceramide biosynthesis (Xu et al., 2024). Furthermore, CerS6 was found to have a detrimental effect on mitochondrial dynamics and function, while increasing apoptosis in neuronal cell culture. Additionally, CerS6 interacts with the sequestosome 1 protein, impairing mitophagy and contributing to mitochondrial dysfunction (Xu et al., 2024).
Finally, ceramide synthesis and accumulation in brain might be sensitive to metabolic triggers. A recent report documented that insulin significantly increased C16:1, C20, C24, and C24:1 synthesis in the cerebral cortex of ApoE4 mice model (Carr et al., 2023). Also, C18-ceramide production seems to modulate the argininosuccinate/fumarate/malate axis and mitophagy in neurodegenerative diseases. Authors reported that exogenous fumarate or malate inhibited mitophagy, leading to mitochondrial damage and sensorimotor abnormalities associated with aging in p17/PERMIT−/− mice (van Eijk et al., 2017). Conversely, C18-ceramide activates mitophagy and improved sensorimotor deficits in aged p17/PERMIT−/− mice (van Eijk et al., 2017).
These findings reinforce the link between ceramides species positively or negatively modulating mitophagy and mitochondrial dysfunction in neurodegenerative diseases.
4 Intermittent fasting (IF)
By definition, fasting is a voluntary deprivation of food and caloric beverages, for different purposes, ranging from religious (Galassi et al., 2018) or political reasons to obtaining health benefits. Alternating between periods of food restriction and periods of food intake is known as IF. IF has currently been proposed as a strategy for reducing and controlling body weight (Chair et al., 2022; Jospe et al., 2020; Templeman et al., 2021), as well as improving metabolic and cardiovascular health (Liu et al., 2022; Sutton et al., 2018).
There are various IF protocols, distinguished by the duration and frequency of fastin periods. In this review, we focus on the following protocols: time-restricted eating, alternate-day fasting and twice-per-week fasting (Koppold et al., 2024). During the time-restricted eating (TRE) (or time restricted feeding -TRF- in animal models) daily eating hours are reduced (Liu et al., 2022), allowing food intake ad libitum in windows ranging from 4 up to 10 h (14–20 h of fasting per day) (Lowe et al., 2020; Mayra et al., 2022; Wilkinson et al., 2020). Several authors do not properly consider TRE as a type of IF due to time of eating to certain hours each day, and it may not involve significant food deprivation compared to other types of fasting (Attinà et al., 2021; Fanti et al., 2021). Accordingly, alternate day fasting (ADF), also called every other day fasting (Alzoubi et al., 2021), involves 1 day of ad libitum feeding followed by a full day of fasting (Stekovic et al., 2019; Hoddy et al., 2014). In twice-per-week fasting (such as the so-called “5:2 diet”), fasting is performed on 2 non-consecutive days of the week, and the remaining 5 days are for habitual food intake (Scholtens et al., 2020). In both ADF and twice-per-week fasting regimen, caloric intake can be very low during fasting periods (less than 25% of daily caloric intake), which is referred to as modified (Hoddy et al., 2014; Koppold et al., 2024).
Despite variations in fasting protocols, alternating between fasting and regular eating leads to significant metabolic and hormonal adaptations of body physiology. In IF, the use of the body’s energy reserves, such as liver glycogen and adipose tissue triacylglycerols, is increased. After about 12 h of fasting, a metabolic switch occurs and the use of triacylglycerols increases (Mattson et al., 2018), leading to changes in brain metabolic activity.
Next, we will address the effect of IF on ceramides profiles assisting mitochondria function and dynamics.
5 IF regulates ceramides profiles and mitochondrial function and dynamics
5.1 Effect of IF on ceramides levels and lipid composition in the mitochondria membranes
As mentioned above, IF and CR have been demonstrated to modify mitochondrial function, however, little is known regarding the effects of these dietary regimens on mitochondrial membrane composition. Cardiolipins are specific phospholipids of the mitochondrial inner membrane that participates in many aspects of its organization and function, hence promoting proper mitochondrial ATP production (Prola and Pilot-Storck, 2022). It is known that oxidative phosphorylation generates the electrochemical gradients to produce ATP synthesis, which occurs in the mitochondrial inner membrane and requires specific membrane morphology. Cardiolipins is an anionic molecule, with a dimeric molecular structure comprising three glycerol groups, two phosphate moieties, and four esterified fatty acyl chains, which are all bound to a compact polar head (Ikon and Ryan, 2017). Cardiolipin represents around 10–15% of mitochondrial membrane phospholipids and promotes the formation of mitochondrial cristae (Joubert and Puff, 2021), Cardiolipin also interacts with numerous other proteins involved in protein import, calcium transport, and cell death (Ghosh et al., 2020) (Figure 3).
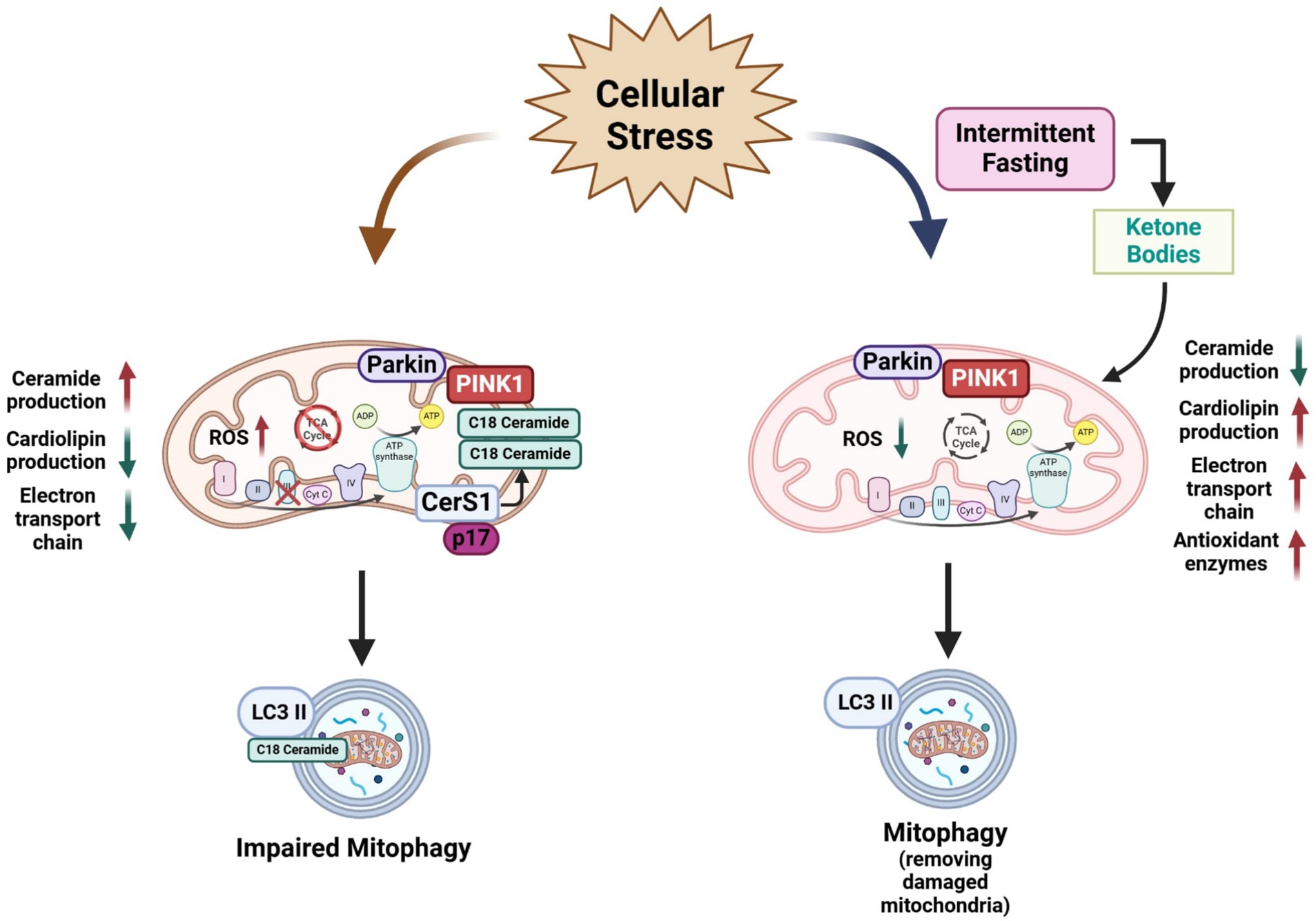
Figure 3. Schematic representation of the negative effects of ceramide in mitochondria physiology and mitophagy and a possible beneficial effect of intermittent fasting on mitochondria. Ceramide overproduction produces negative effects on mitochondria physiology such as an alteration in the mitochondrial respiratory chain function and ROS accumulation leading to oxidative stress. Furthermore, increasing levels of ceramide, especially C18:0 specie activates mitophagy. Conversely, IF improves mitochondrial function in several cell types by reducing ceramide production, reducing the ROS production by increasing antioxidant enzyme levels, production of cardiolipin levels in the mitochondria membrane, and improving the electron transport chain by enhancing mitochondrial oxidative metabolism and mitochondrial respiratory control rate. IF also can induce mitophagy to remove damaged mitochondria by several cellular insults. Scheme created in https://BioRender.com.
A few reports have shown that CR or fasting could modulate cardiolipin biosynthesis or promote changes in mitochondria membranes. Amigo et al. (2017) demonstrated that isolated phospholipids from rat liver mitochondria from animals on ad libitum diet present an increased content of lipoperoxides and higher polarization. Conversely, CR diminished the lipoperoxides content and expression levels of the enzymes involved in cardiolipin biosynthesis and remodeling were found to be upregulated in CR-subjected animals. When mitochondrial membranes were fractionated, the outer membrane presented higher cardiolipin content, indicating a redistribution of this phospholipid mediated by phospholipid scramblase in CR animals. These changes were associated with the upregulation of DRP1-mediated mitochondrial fragmentation and autophagy proteins (Amigo et al., 2017) (Figure 3).
Menezes-Filho et al. (2019) reported that an overnight fasting (15 h) leads to a shift in the liver mitochondrial bioenergetics profile, with a reduction in ADP-stimulated and maximal respiration, lower membrane potential, and lower resistance to Ca2 + −induced mitochondrial permeability transition. Interestingly, 15 h fasting was able to increase cardiolipin quantities in isolated mitochondria from liver (Menezes-Filho et al., 2019). These findings show that dietary regimens could promote the remodeling of cardiolipin content in mitochondrial membranes.
5.2 Effect of IF on ceramides levels, ETC and ROS generation
Excessive glucose flux has been described as increasing the activity of the mitochondrial ETC, leading to ROS overproduction (Hallan and Sharma, 2016; Joaquim et al., 2022). IF protocols such as the ADF for one month was shown to reduce mitochondrial respiratory control in liver but not in the brain of young mice, showing a tissue-specific effect toward energy saving (Sorochynska et al., 2019). In normal aging, a decline in brain mitochondrial respiration and antioxidant enzymes has also been observed. Notably, aged mice exposed to ADF reduced oxidative stress and increased antioxidant enzyme levels, while having a modest effect in improving respiratory control (Bayliak et al., 2021). Furthermore, both ADF and time-restricted feeding reduced malondialdehyde accumulation in the mitochondria and increased complex I and III activity of the ETC in young and aged mice (Chen et al., 2024). Otherwise, IF is also beneficial to mitochondrial and peroxisomal lipid proteins homeostasis. For instance, IF restored ETC proteins and normalized the oxygen consumption rates of Complex I and II in a rat model of pulmonary arterial hypertension (PAH) (Almuraikhy et al., 2024). Mechanistically, it is documented that mitochondria electron transport chain genes are progressively sensitive to fasting. At 12 h of fasting, 14 genes were downregulated, whereas 6 genes, scattered over all 5 complexes were upregulated. At 24 h, 3 genes remained upregulated among 20 downregulated species. At 72 h of fasting, no fewer than 63% of the respiratory chain genes, including cytochrome C, were downregulated (Sokolović et al., 2007). This suggest that IF might modulate the mitochondrial electron transport chain at transcriptional level. (Figure 3).
IF has been demonstrated to confer survival benefits in several disease models (de Cabo and Mattson, 2019). Also, the role of IF has been studied in clinical trials. In a recent study in overweight and obese women with rheumatoid arthritis, IF decreased the oxidative stress marker as malondialdehyde levels in serum and, neutrophil-to-lymphocyte ratio together with an increase in catalase levels. Also, liver enzymes such as aspartate transaminase and alanine transaminase had a significant decrease in the IF group (Tavakoli et al., 2025). Such evidence suggests that IF might be integrated as an interventional approach to tackle oxidative stress and inflammatory profiles (Figure 3).
5.3 Effect of IF on apoptosis and mitophagy
IF positively modulates mitochondrial dynamics resulting in preserved mitochondrial health and cellular energetics (Diab et al., 2024). Authors reported that preservation of mitochondrial integrity could be attributed to suppressing excessive mitochondrial fission proteins by the upregulation of adipose triglyceride lipase Atg1, the rate limiting enzyme of triglyceride hydrolysis. The liberation of fatty acids by Atg1 during fasting contributes to enhanced mitochondrial oxidative capacity (Lettieri-Barbato et al., 2018). On the other hand, Real-Hohn et al. (2018) showed that IF increases mitochondrial mass compared with ad libitum animals. Furthermore, these authors found that IF group had a lower level of oxidative stress and greater oxidative capacity and promoting a balanced fusion-fission equilibrium and inducing mitophagy (Real-Hohn et al., 2018). McWilliams et al. (2018) also reported the effect of fasting on basal mitophagy in mouse tissues with high metabolic demand such as the brain, heart, eye, liver, kidney, and spleen. According, a high degree of mitophagy occurred in neuronal cells such as dopaminergic neurons and microglia (McWilliams et al., 2018). In high fat-fructose diet-induced liver and skeletal muscle injury model, Bastawy et al. (2024) showed that IF and metformin upregulated mitophagy-related genes such as PINK1, Parkin, and LAMP, attenuated the hepatic triglyceride content and decreased oxidative stress markers (Bastawy et al., 2024). During low energetic supply, it is expected that activation of the AMP-activated protein kinase (AMPK), which activates mitophagy. Conversely, mice lacking skeletal muscle AMPK subjected to fasting conditions showed that both mRNA and protein level of autophagy adaptor protein p62 was increased in fasted conditions. Moreover, Parkin protein level increased by more than two-fold in AMPK-MKO compared with wild-type mice (Bujak et al., 2015) (Figure 3).
Finally, calorie restriction (CR) has also probed to modulates mitophagy. Long-term CR reduces mitochondrial biogenesis and mitophagy in liver, as measured by quantitative proteomics (Price et al., 2015). Also, CR was able to become susceptibility to mitophagy, together with an increase of both Pink1 mRNA and an increase of Drp1 protein (Zhao et al., 2013).
This evidence suggests that intermittent fasting, calorie restriction or fasting induces mitophagy and mitophagy-related markers, whereby it could be concluded that restrictive interventions have a role in the regulation of health by inducing mitochondria autophagy in different organs of the body (Mehrabani et al., 2020).
6 Conclusion
The current review provided experimental evidence supporting that dysregulation of ceramides metabolism assists neurodegenerative disorders through mitochondria dysfunction. Here, we propose that ceramides susceptibilizes to neurodegeneration by affecting mitochondria membrane composition, respiratory chain, ROS dynamics, ATP production and mitophagy-apoptosis in brain. We conceive that dysregulation of ceramide metabolism is sensitive to ultra-processed foods intake normally exposed during the current day life. Elucidating how ceramides exert their effects can enhance pathophysiological understanding of neurodegenerative disorders and constitute potential biological targets for novel drugs and diagnostic and prognostic biomarkers.
A major potential conclusion of our review is the effect of IF on neurodegeneration. IF has been shown to be an effective strategy to control body weight and improve several metabolic parameters. However, its application in the context of neurodegenerative disorders may be limited by some contrasting or inconclusive results, representing a constantly evolving area of research. We provide evidence documented that IF positively targets mitochondria at three levels: membrane lipid composition, respiratory chain and ROS generation and mitophagy-apoptosis. In this context, IF might be integrated as a future non-pharmacological strategy supporting integral treatment of neurodegenerative disorders. Additionally, a potential combination of IF with pharmacological treatments may enhance therapeutic intervention and efficacy by targeting metabolic and inflammatory pathways which normally accounts during physiological brain aging.
By now, research on IF and its role on neurodegeneration faces significant challenges, including a limited number of studies addressing IF protocols on neurodegeneration. This heterogeneity complicates the ability to draw definitive conclusions or recommend a standardized protocol mainly in preclinical models. Future research should prioritize the development of consistent IF protocols in animal models and characterize selective molecular targets. Addressing these challenges will identify selective cellular and molecular pathways linking ceramides metabolism to neurodegenerative disorders, which are essential for translating IF into sustainable and broadly applicable strategies in clinic for improving brain health potentially focusing on neurodegeneration.
Author contributions
LV-A: Writing – review & editing, Writing – original draft, Funding acquisition, Supervision, Formal analysis, Investigation, Conceptualization. OM-G: Writing – original draft, Software, Funding acquisition, Formal analysis, Conceptualization, Supervision, Writing – review & editing, Investigation. RV-C: Funding acquisition, Visualization, Formal analysis, Investigation, Writing – review & editing, Conceptualization, Writing – original draft. RG-G: Conceptualization, Supervision, Writing – review & editing, Writing – original draft, Software, Investigation, Funding acquisition, Formal analysis. AC-M: Supervision, Formal analysis, Conceptualization, Funding acquisition, Writing – review & editing, Writing – original draft, Investigation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Grant No. CBF2023-2024-1804 from the Secretary of Science, Humanities, Technology and Innovation (SECIHTI).
Acknowledgments
We thank M. S. Alejandra Arreola-Triana for her support in editing this manuscript, and Victor Mendoza for his assisting to images design.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abou-Ghali, M., and Stiban, J. (2015). Regulation of ceramide channel formation and disassembly: insights on the initiation of apoptosis. Saudi J. Biol. Sci. 22, 760–772. doi: 10.1016/j.sjbs.2015.03.005
Afridi, R., Rahman, M. H., and Suk, K. (2022). Implications of glial metabolic dysregulation in the pathophysiology of neurodegenerative diseases. Neurobiol. Dis. 174:105874. doi: 10.1016/j.nbd.2022.105874
Alecu, I., and Bennett, S. A. L. (2019). Dysregulated lipid metabolism and its role in α-Synucleinopathy in Parkinson’s disease. Front. Neurosci. 13:328. doi: 10.3389/fnins.2019.00328
Almuraikhy, S., Naja, K., Anwardeen, N., Sellami, M., Al-Amri, H. S., Al-Sulaiti, H., et al. (2024). Metabolic signatures of combined exercise and fasting: an expanded perspective on previous telomere length findings. Front. Aging 5:1494095. doi: 10.3389/fragi.2024.1494095
Alonso, A., and Goñi, F. M. (2018). The physical properties of ceramides in membranes. Annu. Rev. Biophys. 47, 633–654. doi: 10.1146/annurev-biophys-070317-033309
Alzoubi, K. H., Khabour, O. F., Al-Awad, R. M., and Aburashed, Z. O. (2021). Every-other day fasting prevents memory impairment induced by high fat-diet: role of oxidative stress. Physiol. Behav. 229:113263. doi: 10.1016/j.physbeh.2020.113263
Amigo, I., Menezes-Filho, S. L., Luévano-Martínez, L. A., Chausse, B., and Kowaltowski, A. J. (2017). Caloric restriction increases brain mitochondrial calcium retention capacity and protects against excitotoxicity. Aging Cell 16, 73–81. doi: 10.1111/acel.12527
Argueti-Ostrovsky, S., Barel, S., Kahn, J., and Israelson, A. (2024). VDAC1: a key player in the mitochondrial landscape of neurodegeneration. Biomol. Ther. 15:33. doi: 10.3390/biom15010033
Arsenault, E. J., McGill, C. M., and Barth, B. M. (2021). Sphingolipids as regulators of neuro-inflammation and NADPH oxidase 2. NeuroMolecular Med. 23, 25–46. doi: 10.1007/s12017-021-08646-2
Attinà, A., Leggeri, C., Paroni, R., Pivari, F., Dei Cas, M., Mingione, A., et al. (2021). Fasting: how to guide. Nutrients 13:1570. doi: 10.3390/nu13051570
Aufschnaiter, A., Kohler, V., Diessl, J., Peselj, C., Carmona-Gutierrez, D., Keller, W., et al. (2017). Mitochondrial lipids in neurodegeneration. Cell Tissue Res. 367, 125–140. doi: 10.1007/s00441-016-2463-1
Bastawy, N., Soliman, G. F., Sadek, N. B., Gharib, D. M., Gouda, M. A., Rashed, L. A., et al. (2024). Metformin and intermittent fasting mitigate high fat-fructose diet-induced liver and skeletal muscle injury through upregulation of mitophagy genes in rats. Beni Suef Univ. J. Basic Appl. Sci. 13:88. doi: 10.1186/s43088-024-00548-z
Bayliak, M. M., Dmytriv, T. R., Melnychuk, A. V., Strilets, N. V., Storey, K. B., and Lushchak, V. I. (2021). Chamomile as a potential remedy for obesity and metabolic syndrome. EXCLI J. 20, 1261–1286. doi: 10.17179/excli2021-4013
Bernal-Vega, S., García-Juárez, M., and Camacho-Morales, A. (2023). Contribution of ceramides metabolism in psychiatric disorders. J. Neurochem. 164, 708–724. doi: 10.1111/jnc.15759
Bionda, C., Portoukalian, J., Schmitt, D., Rodriguez-Lafrasse, C., and Ardail, D. (2004). Subcellular compartmentalization of ceramide metabolism: MAM (mitochondria-associated membrane) and/or mitochondria? Biochem. J. 382, 527–533. doi: 10.1042/BJ20031819
Bonnaud, S., Niaudet, C., Pottier, G., Gaugler, M.-H., Millour, J., Barbet, J., et al. (2007). Sphingosine-1-phosphate protects proliferating endothelial cells from ceramide-induced apoptosis but not from DNA damage-induced mitotic death. Cancer Res. 67, 1803–1811. doi: 10.1158/0008-5472.CAN-06-2802
Böttinger, L., Ellenrieder, L., and Becker, T. (2016). How lipids modulate mitochondrial protein import. J. Bioenerg. Biomembr. 48, 125–135. doi: 10.1007/s10863-015-9599-7
Brachtendorf, S., El-Hindi, K., and Grösch, S. (2019). Ceramide synthases in cancer therapy and chemoresistance. Prog. Lipid Res. 74, 160–185. doi: 10.1016/j.plipres.2019.04.002
Bujak, A. L., Crane, J. D., Lally, J. S., Ford, R. J., Kang, S. J., Rebalka, I. A., et al. (2015). AMPK activation of muscle autophagy prevents fasting-induced hypoglycemia and myopathy during aging. Cell Metab. 21, 883–890. doi: 10.1016/j.cmet.2015.05.016
Camacho, A., and Massieu, L. (2006). Role of glutamate transporters in the clearance and release of glutamate during ischemia and its relation to neuronal death. Arch. Med. Res. 37, 11–18. doi: 10.1016/j.arcmed.2005.05.014
Camacho, A., Montiel, T., and Massieu, L. (2006). The anion channel blocker, 4,4′-dinitrostilbene-2,2′-disulfonic acid prevents neuronal death and excitatory amino acid release during glycolysis inhibition in the hippocampus in vivo. Neuroscience 142, 1005–1017. doi: 10.1016/j.neuroscience.2006.07.004
Camacho, A., Montiel, T., and Massieu, L. (2007). Sustained metabolic inhibition induces an increase in the content and phosphorylation of the NR2B subunit of N-methyl-D-aspartate receptors and a decrease in glutamate transport in the rat hippocampus in vivo. Neuroscience 145, 873–886. doi: 10.1016/j.neuroscience.2006.12.069
Camacho-Morales, A., Noriega, L. G., Sánchez-García, A., Torre-Villalvazo, I., Vázquez-Manjarrez, N., Maldonado-Ruiz, R., et al. (2024). Plasma C24:0 ceramide impairs adipose tissue remodeling and promotes liver steatosis and glucose imbalance in offspring of rats. Heliyon 10:e39206. doi: 10.1016/j.heliyon.2024.e39206
Canals, D., and Clarke, C. J. (2022). Compartmentalization of sphingolipid metabolism: implications for signaling and therapy. Pharmacol. Ther. 232:108005. doi: 10.1016/j.pharmthera.2021.108005
Carr, S. T., Saito, E. R., Walton, C. M., Saito, J. Y., Hanegan, C. M., Warren, C. E., et al. (2023). Ceramides mediate insulin-induced impairments in cerebral mitochondrial bioenergetics in ApoE4 mice. Int. J. Mol. Sci. 24:16635. doi: 10.3390/ijms242316635
Chair, S. Y., Cai, H., Cao, X., Qin, Y., Cheng, H. Y., and Ng, M. T. (2022). Intermittent fasting in weight loss and Cardiometabolic risk reduction: a randomized controlled trial. J. Nurs. Res. 30:e185. doi: 10.1097/jnr.0000000000000469
Chang, K.-T., Anishkin, A., Patwardhan, G. A., Beverly, L. J., Siskind, L. J., and Colombini, M. (2015). Ceramide channels: destabilization by Bcl-xL and role in apoptosis. Biochim. Biophys. Acta 1848, 2374–2384. doi: 10.1016/j.bbamem.2015.07.013
Chaurasia, B., Tippetts, T. S., Mayoral Monibas, R., Liu, J., Li, Y., Wang, L., et al. (2019). Targeting a ceramide double bond improves insulin resistance and hepatic steatosis. Science 365, 386–392. doi: 10.1126/science.aav3722
Chen, T.-H., Wang, H.-C., Chang, C.-J., and Lee, S.-Y. (2024). Mitochondrial glutathione in cellular redox homeostasis and disease manifestation. Int. J. Mol. Sci. 25:1314. doi: 10.3390/ijms25021314
Chen, Y.-Z., Zimyanin, V., and Redemann, S. (2023). Loss of the mitochondrial protein SPD-3 elevates PLK-1 levels and dysregulates mitotic events. Life Sci. Alliance 6:e202302011. doi: 10.26508/lsa.202302011
Chowdhury, M. R., Jin, H. K., and Bae, J. (2022). Diverse roles of ceramide in the progression and pathogenesis of Alzheimer’s disease. Biomedicines 10:1956. doi: 10.3390/biomedicines10081956
Ciarlo, L., Manganelli, V., Matarrese, P., Garofalo, T., Tinari, A., Gambardella, L., et al. (2012). Raft-like microdomains play a key role in mitochondrial impairment in lymphoid cells from patients with Huntington’s disease. J. Lipid Res. 53, 2057–2068. doi: 10.1194/jlr.M026062
Crivelli, S. M., Quadri, Z., Elsherbini, A., Vekaria, H. J., Sullivan, P. G., Zhi, W., et al. (2024). Abnormal regulation of mitochondrial sphingolipids during aging and Alzheimer’s disease. ASN Neuro 16:2404367. doi: 10.1080/17590914.2024.2404367
Dany, M., and Ogretmen, B. (2015). Ceramide induced mitophagy and tumor suppression. Biochim. Biophys. Acta 1853, 2834–2845. doi: 10.1016/j.bbamcr.2014.12.039
de Cabo, R., and Mattson, M. P. (2019). Effects of intermittent fasting on health, aging, and disease. N. Engl. J. Med. 381, 2541–2551. doi: 10.1056/NEJMra1905136
Diab, R., Dimachkie, L., Zein, O., Dakroub, A., and Eid, A. H. (2024). Intermittent fasting regulates metabolic homeostasis and improves cardiovascular health. Cell Biochem. Biophys. 82, 1583–1597. doi: 10.1007/s12013-024-01314-9
Diaz-Vegas, A., Madsen, S., Cooke, K. C., Carroll, L., Khor, J. X. Y., Turner, N., et al. (2023). Mitochondrial electron transport chain, ceramide and coenzyme Q are linked in a pathway that drives insulin resistance in skeletal muscle. Elife. 27:12:RP87340. doi: 10.7554/eLife.87340
Ding, S., Li, G., Fu, T., Zhang, T., Lu, X., Li, N., et al. (2024). Ceramides and mitochondrial homeostasis. Cell. Signal. 117:111099. doi: 10.1016/j.cellsig.2024.111099
Ding, C., Wu, Y., Chen, X., Chen, Y., Wu, Z., Lin, Z., et al. (2022). Global, regional, and national burden and attributable risk factors of neurological disorders: the global burden of disease study 1990-2019. Front. Public Health 10:952161. doi: 10.3389/fpubh.2022.952161
DiPasquale, M., Deering, T. G., Desai, D., Sharma, A. K., Amin, S., Fox, T. E., et al. (2022). Influence of ceramide on lipid domain stability studied with small-angle neutron scattering: the role of acyl chain length and unsaturation. Chem. Phys. Lipids 245:105205. doi: 10.1016/j.chemphyslip.2022.105205
Doxaki, C., and Palikaras, K. (2020). Neuronal Mitophagy: friend or foe? Front. Cell Develop. Biol. 8:611938. doi: 10.3389/fcell.2020.611938
Duffy, C. M., Yuan, C., Wisdorf, L. E., Billington, C. J., Kotz, C. M., Nixon, J. P., et al. (2015). Role of orexin a signaling in dietary palmitic acid-activated microglial cells. Neurosci. Lett. 606, 140–144. doi: 10.1016/j.neulet.2015.08.033
Erdem, N. Z., Bayraktaroğlu, E., Samancı, R. A., Geçgil-Demir, E., Tarakçı, N. G., and Mert-Biberoğlu, F. (2022). The effect of intermittent fasting diets on body weight and composition. Clinic. Nutr. ESPEN 51, 207–214. doi: 10.1016/j.clnesp.2022.08.030
Fanti, M., Mishra, A., Longo, V. D., and Brandhorst, S. (2021). Time-restricted eating, intermittent fasting, and fasting-mimicking diets in weight loss. Curr. Obes. Rep. 10, 70–80. doi: 10.1007/s13679-021-00424-2
Faris, M. A.-I. E., Kacimi, S., Al-Kurd, R. A., Fararjeh, M. A., Bustanji, Y. K., Mohammad, M. K., et al. (2012). Intermittent fasting during Ramadan attenuates proinflammatory cytokines and immune cells in healthy subjects. Nutr. Res. 32, 947–955. doi: 10.1016/j.nutres.2012.06.021
Farsi, Z., Walde, M., Klementowicz, A. E., Paraskevopoulou, F., and Woehler, A. (2021). Single synapse glutamate imaging reveals multiple levels of release mode regulation in mammalian synapses. iScience 24:101909. doi: 10.1016/j.isci.2020.101909
Fato, R., Bergamini, C., Bortolus, M., Maniero, A. L., Leoni, S., Ohnishi, T., et al. (2009). Differential effects of mitochondrial complex I inhibitors on production of reactive oxygen species. Biochim. Biophys. Acta 1787, 384–392. doi: 10.1016/j.bbabio.2008.11.003
Fisher-Wellman, K. H., Hagen, J. T., Neufer, P. D., Kassai, M., and Cabot, M. C. (2021). On the nature of ceramide-mitochondria interactions—dissection using comprehensive mitochondrial phenotyping. Cell. Signal. 78:109838. doi: 10.1016/j.cellsig.2020.109838
Fucho, R., Casals, N., Serra, D., and Herrero, L. (2017). Ceramides and mitochondrial fatty acid oxidation in obesity. FASEB J. 31, 1263–1272. doi: 10.1096/fj.201601156R
Furuya, S., Mitoma, J., Makino, A., and Hirabayashi, Y. (1998). Ceramide and its interconvertible metabolite sphingosine function as indispensable lipid factors involved in survival and dendritic differentiation of cerebellar Purkinje cells. J. Neurochem. 71, 366–377. doi: 10.1046/j.1471-4159.1998.71010366.x
Galassi, F. M., Bender, N., Habicht, M. E., Armocida, E., Toscano, F., Menassa, D. A., et al. (2018). St. Catherine of Siena (1347-1380 AD): one of the earliest historic cases of altered gustatory perception in anorexia mirabilis. Neurol. Sci. 39, 939–940. doi: 10.1007/s10072-018-3285-6
García-González, V., Díaz-Villanueva, J. F., Galindo-Hernández, O., Martínez-Navarro, I., Hurtado-Ureta, G., and Pérez-Arias, A. A. (2018). Ceramide metabolism balance, a multifaceted factor in critical steps of breast Cancer development. Int. J. Mol. Sci. 19:2527. doi: 10.3390/ijms19092527
García-Ruiz, C., Colell, A., Marí, M., Morales, A., and Fernández-Checa, J. C. (1997). Direct effect of ceramide on the mitochondrial electron transport chain leads to generation of reactive oxygen species. Role of mitochondrial glutathione. J. Biol. Chem. 272, 11369–11377. doi: 10.1074/jbc.272.17.11369
Geiger, O., Sanchez-Flores, A., Padilla-Gomez, J., and Degli Esposti, M. (2023). Multiple approaches of cellular metabolism define the bacterial ancestry of mitochondria. Sci. Adv. 9:eadh0066. doi: 10.1126/sciadv.adh0066
Ghosh, S., Basu Ball, W., Madaris, T. R., Srikantan, S., Madesh, M., Mootha, V. K., et al. (2020). An essential role for cardiolipin in the stability and function of the mitochondrial calcium uniporter. Proc. Natl. Acad. Sci. USA 117, 16383–16390. doi: 10.1073/pnas.2000640117
Giovannoni, F., and Quintana, F. J. (2020). The role of astrocytes in CNS inflammation. Trends Immunol. 41, 805–819. doi: 10.1016/j.it.2020.07.007
Gómez-Muñoz, A. (2006). Ceramide 1-phosphate/ceramide, a switch between life and death. Biochim. Biophys. Acta 1758, 2049–2056. doi: 10.1016/j.bbamem.2006.05.011
Greene, A. W., Grenier, K., Aguileta, M. A., Muise, S., Farazifard, R., Haque, M. E., et al. (2012). Mitochondrial processing peptidase regulates PINK1 processing, import and Parkin recruitment. EMBO Rep. 13, 378–385. doi: 10.1038/embor.2012.14
Gudz, T. I., Tserng, K. Y., and Hoppel, C. L. (1997). Direct inhibition of mitochondrial respiratory chain complex III by cell-permeable ceramide. J. Biol. Chem. 272, 24154–24158. doi: 10.1074/jbc.272.39.24154
Hadrava Vanova, K., Kraus, M., Neuzil, J., and Rohlena, J. (2020). Mitochondrial complex II and reactive oxygen species in disease and therapy. Redox Rep. 25, 26–32. doi: 10.1080/13510002.2020.1752002
Hallan, S., and Sharma, K. (2016). The role of mitochondria in diabetic kidney disease. Curr. Diab. Rep. 16:61. doi: 10.1007/s11892-016-0748-0
Hammerschmidt, P., and Brüning, J. C. (2022). Contribution of specific ceramides to obesity-associated metabolic diseases. Cell. Mol. Life Sci. 79:395. doi: 10.1007/s00018-022-04401-3
Hamzeh, O., Rabiei, F., Shakeri, M., Parsian, H., Saadat, P., and Rostami-Mansoor, S. (2023). Mitochondrial dysfunction and inflammasome activation in neurodegenerative diseases: mechanisms and therapeutic implications. Mitochondrion 73, 72–83. doi: 10.1016/j.mito.2023.10.003
Han, X., Holtzman, M., McKeel, D. W., Kelley, J., and Morris, J. C. (2002). Substantial sulfatide deficiency and ceramide elevation in very early Alzheimer’s disease: potential role in disease pathogenesis. J. Neurochem. 82, 809–818. doi: 10.1046/j.1471-4159.2002.00997.x
Hoddy, K. K., Kroeger, C. M., Trepanowski, J. F., Barnosky, A., Bhutani, S., and Varady, K. A. (2014). Meal timing during alternate day fasting: impact on body weight and cardiovascular disease risk in obese adults. Obesity 22, 2524–2531. doi: 10.1002/oby.20909
Huang, Y., Li, Y., Pan, H., and Han, L. (2023). Global, regional, and national burden of neurological disorders in 204 countries and territories worldwide. J. Glob. Health 13:04160. doi: 10.7189/jogh.13.04160
Ikon, N., and Ryan, R. O. (2017). Cardiolipin and mitochondrial cristae organization. Biochim. Biophys. Acta Biomem. 1859, 1156–1163. doi: 10.1016/j.bbamem.2017.03.013
Iwayama, H., and Ueda, N. (2013). Role of mitochondrial Bax, caspases, and MAPKs for ceramide-induced apoptosis in renal proximal tubular cells. Mol. Cell. Biochem. 379, 37–42. doi: 10.1007/s11010-013-1624-8
Jazvinšćak Jembrek, M., Hof, P. R., and Šimić, G. (2015). Ceramides in Alzheimer’s disease: key mediators of neuronal apoptosis induced by oxidative stress and aβ accumulation. Oxidative Med. Cell. Longev. 2015:346783. doi: 10.1155/2015/346783
Ji, J. E., Kim, S. K., Ahn, K. H., Choi, J. M., Jung, S. Y., Jung, K. M., et al. (2011). Ceramide induces serotonin release from RBL-2H3 mast cells through calcium mediated activation of phospholipase A2. Prostaglandins Other Lipid Mediat. 94, 88–95. doi: 10.1016/j.prostaglandins.2011.01.001
Joaquim, L., Faria, A., Loureiro, H., and Matafome, P. (2022). Benefits, mechanisms, and risks of intermittent fasting in metabolic syndrome and type 2 diabetes. J. Physiol. Biochem. 78, 295–305. doi: 10.1007/s13105-021-00839-4
Jospe, M. R., Roy, M., Brown, R. C., Haszard, J. J., Meredith-Jones, K., Fangupo, L. J., et al. (2020). Intermittent fasting, Paleolithic, or Mediterranean diets in the real world: exploratory secondary analyses of a weight-loss trial that included choice of diet and exercise. Am. J. Clin. Nutr. 111, 503–514. doi: 10.1093/ajcn/nqz330
Joubert, F., and Puff, N. (2021). Mitochondrial cristae architecture and functions: lessons from minimal model systems. Membranes 11:465. doi: 10.3390/membranes11070465
Justice, M. J., Bronova, I., Schweitzer, K. S., Poirier, C., Blum, J. S., Berdyshev, E. V., et al. (2018). Inhibition of acid sphingomyelinase disrupts LYNUS signaling and triggers autophagy. J. Lipid Res. 59, 596–606. doi: 10.1194/jlr.M080242
Kazemeini, S., Nadeem-Tariq, A., Shih, R., Rafanan, J., Ghani, N., and Vida, T. A. (2024). From plaques to pathways in Alzheimer’s disease: the mitochondrial-neurovascular-metabolic hypothesis. Int. J. Mol. Sci. 25:11720. doi: 10.3390/ijms252111720
Kennedy, M. A., Moffat, T. C., Gable, K., Ganesan, S., Niewola-Staszkowska, K., Johnston, A., et al. (2016). A signaling lipid associated with Alzheimer’s disease promotes mitochondrial dysfunction. Sci. Rep. 6:19332. doi: 10.1038/srep19332
Kobayashi, A., Takeiwa, T., Ikeda, K., and Inoue, S. (2023). Roles of noncoding RNAs in regulation of mitochondrial Electron transport chain and oxidative phosphorylation. Int. J. Mol. Sci. 24:9414. doi: 10.3390/ijms24119414
Koppold, D. A., Breinlinger, C., Hanslian, E., Kessler, C., Cramer, H., Khokhar, A. R., et al. (2024). International consensus on fasting terminology. Cell Metab. 36:e4, 1779–1794. doi: 10.1016/j.cmet.2024.06.013
Lee, J. K., Jin, H. K., Park, M. H., Kim, B., Lee, P. H., Nakauchi, H., et al. (2014). Acid sphingomyelinase modulates the autophagic process by controlling lysosomal biogenesis in Alzheimer’s disease. J. Exp. Med. 211, 1551–1570. doi: 10.1084/jem.20132451
Lettieri-Barbato, D., Cannata, S. M., Casagrande, V., Ciriolo, M. R., and Aquilano, K. (2018). Time-controlled fasting prevents aging-like mitochondrial changes induced by persistent dietary fat overload in skeletal muscle. PLoS One 13:e0195912. doi: 10.1371/journal.pone.0195912
Levy, M., and Futerman, A. H. (2010). Mammalian ceramide synthases. IUBMB Life 62, 347–356. doi: 10.1002/iub.319
Lima, T. I., Laurila, P.-P., Wohlwend, M., Morel, J. D., Goeminne, L. J. E., Li, H., et al. (2023). Inhibiting de novo ceramide synthesis restores mitochondrial and protein homeostasis in muscle aging. Sci. Transl. Med. 15:eade6509. doi: 10.1126/scitranslmed.ade6509
Linnerbauer, M., Wheeler, M. A., and Quintana, F. J. (2020). Astrocyte crosstalk in CNS inflammation. Neuron 108, 608–622. doi: 10.1016/j.neuron.2020.08.012
Liu, C., Dhindsa, D., Almuwaqqat, Z., Ko, Y.-A., Mehta, A., Alkhoder, A. A., et al. (2022). Association between high-density lipoprotein cholesterol levels and adverse cardiovascular outcomes in high-risk populations. JAMA Cardiol. 7, 672–680. doi: 10.1001/jamacardio.2022.0912
Liu, X., Jin, Y., Cheng, X., Song, Q., Wang, Y., He, L., et al. (2024). The relevance between abnormally elevated serum ceramide and cognitive impairment in Alzheimer’s disease model mice and its mechanism. Psychopharmacology 241, 525–542. doi: 10.1007/s00213-024-06530-y
López, M., Diéguez, C., Tena-Sempere, M., and González-García, I. (2025). Ceramides in the central control of metabolism. Trends Endocrinol Metab 36, 11–14. doi: 10.1016/j.tem.2024.06.007
Lowe, D. A., Wu, N., Rohdin-Bibby, L., Moore, A. H., Kelly, N., Liu, Y. E., et al. (2020). Effects of time-restricted eating on weight loss and other metabolic parameters in women and men with overweight and obesity: the TREAT randomized clinical trial. JAMA Intern. Med. 180, 1491–1499. doi: 10.1001/jamainternmed.2020.4153
Luo, Y., Ma, J., and Lu, W. (2020). The significance of mitochondrial dysfunction in Cancer. Int. J. Mol. Sci. 21:5598. doi: 10.3390/ijms21165598
Madkour, M. I., Islam, M. T., Tippetts, T. S., Chowdhury, K. H., Lesniewski, L. A., Summers, S. A., et al. (2023). Ramadan intermittent fasting is associated with ameliorated inflammatory markers and improved plasma sphingolipids/ceramides in subjects with obesity: Lipidomics analysis. Sci. Rep. 13:17322. doi: 10.1038/s41598-023-43862-9
Markaki, M., Tsagkari, D., and Tavernarakis, N. (2023). Mitophagy and long-term neuronal homeostasis. J. Cell Sci. 136:jcs260638. doi: 10.1242/jcs.260638
Mattson, M. P., Moehl, K., Ghena, N., Schmaedick, M., and Cheng, A. (2018). Intermittent metabolic switching, neuroplasticity and brain health. Nat. Rev. Neurosci. 19, 81–94. doi: 10.1038/nrn.2017.156
Mayra, S. T., Kravat, N., Chondropoulos, K., De Leon, A., and Johnston, C. S. (2022). Early time-restricted eating may favorably impact cognitive acuity in university students: a randomized pilot study. Nutrit. Res. 108, 1–8. doi: 10.1016/j.nutres.2022.10.002
McGrath, E. R., Himali, J. J., Xanthakis, V., Duncan, M. S., Schaffer, J. E., Ory, D. S., et al. (2020). Circulating ceramide ratios and risk of vascular brain aging and dementia. Ann. Clinic. Trans. Neurol. 7, 160–168. doi: 10.1002/acn3.50973
McInnis, J. J., Sood, D., Guo, L., Dufault, M. R., Garcia, M., Passaro, R., et al. (2024). Unravelling neuronal and glial differences in ceramide composition, synthesis, and sensitivity to toxicity. Commun. Biol. 7, 1597–1517. doi: 10.1038/s42003-024-07231-0
McWilliams, T. G., Prescott, A. R., Montava-Garriga, L., Ball, G., Singh, F., Barini, E., et al. (2018). Basal Mitophagy occurs independently of PINK1 in mouse tissues of high metabolic demand. Cell Metab. 27, 439–449.e5. doi: 10.1016/j.cmet.2017.12.008
Mehrabani, S., Bagherniya, M., Askari, G., Read, M. I., and Sahebkar, A. (2020). The effect of fasting or calorie restriction on mitophagy induction: a literature review. J. Cachexia. Sarcopenia Muscle 11, 1447–1458. doi: 10.1002/jcsm.12611
Mencarelli, C., and Martinez-Martinez, P. (2013). Ceramide function in the brain: when a slight tilt is enough. Cell. Mol. Life Sci. 70, 181–203. doi: 10.1007/s00018-012-1038-x
Menezes-Filho, S. L., Amigo, I., Luévano-Martínez, L. A., and Kowaltowski, A. J. (2019). Fasting promotes functional changes in liver mitochondria. Biochim. Biophy. Acta 1860, 129–135. doi: 10.1016/j.bbabio.2018.11.017
Mielke, M. M., Maetzler, W., Haughey, N. J., Bandaru, V. V. R., Savica, R., Deuschle, C., et al. (2013). Plasma ceramide and glucosylceramide metabolism is altered in sporadic Parkinson’s disease and associated with cognitive impairment: a pilot study. PLoS One 8:e73094. doi: 10.1371/journal.pone.0073094
Mignard, V., Dubois, N., Lanoé, D., Joalland, M.-P., Oliver, L., Pecqueur, C., et al. (2020). Sphingolipid distribution at mitochondria-associated membranes (MAMs) upon induction of apoptosis. J. Lipid Res. 61, 1025–1037. doi: 10.1194/jlr.RA120000628
Mullen, T. D., Hannun, Y. A., and Obeid, L. M. (2012). Ceramide synthases at the Centre of sphingolipid metabolism and biology. Biochem. J. 441, 789–802. doi: 10.1042/BJ20111626
Novgorodov, S. A., Chudakova, D. A., Wheeler, B. W., Bielawski, J., Kindy, M. S., Obeid, L. M., et al. (2011). Developmentally regulated ceramide synthase 6 increases mitochondrial Ca2+ loading capacity and promotes apoptosis. J. Biol. Chem. 286, 4644–4658. doi: 10.1074/jbc.M110.164392
Novgorodov, S. A., Gudz, T. I., and Obeid, L. M. (2008). Long-chain ceramide is a potent inhibitor of the mitochondrial permeability transition pore. J. Biol. Chem. 283, 24707–24717. doi: 10.1074/jbc.M801810200
Novgorodov, S. A., Riley, C. L., Keffler, J. A., Yu, J., Kindy, M. S., Macklin, W. B., et al. (2016). SIRT3 deacetylates ceramide synthases: implications for mitochondrial dysfunction and brain injury. J. Biol. Chem. 291, 1957–1973. doi: 10.1074/jbc.M115.668228
Oleinik, N., Albayram, O., Kassir, M. F., Atilgan, F. C., Walton, C., Karakaya, E., et al. (2023). Alterations of lipid-mediated mitophagy result in aging-dependent sensorimotor defects. Aging Cell 22:e13954. doi: 10.1111/acel.13954
Olsen, A. S. B., and Færgeman, N. J. (2017). Sphingolipids: membrane microdomains in brain development, function and neurological diseases. Open Biol. 7:170069. doi: 10.1098/rsob.170069
Parra, V., Moraga, F., Kuzmicic, J., López-Crisosto, C., Troncoso, R., Torrealba, N., et al. (2013). Calcium and mitochondrial metabolism in ceramide-induced cardiomyocyte death. Biochim. Biophys. Acta 1832, 1334–1344. doi: 10.1016/j.bbadis.2013.04.009
Pastorino, J. G., Shulga, N., and Hoek, J. B. (2002). Mitochondrial binding of hexokinase II inhibits Bax-induced cytochrome c release and apoptosis. J. Biol. Chem. 277, 7610–7618. doi: 10.1074/jbc.M109950200
Perez-Kast, R. C., and Camacho-Morales, A. (2025). Fasting the brain for mental health. J. Psychiatr. Res. 181, 215–224. doi: 10.1016/j.jpsychires.2024.11.041
Price, M., Higgs, S., and Lee, M. (2015). Self-reported eating traits: underlying components of food responsivity and dietary restriction are positively related to BMI. Appetite 95, 203–210. doi: 10.1016/j.appet.2015.07.006
Prola, A., and Pilot-Storck, F. (2022). Cardiolipin alterations during obesity: exploring therapeutic opportunities. Biology 11:1638. doi: 10.3390/biology11111638
Qin, J., Berdyshev, E., Goya, J., Natarajan, V., and Dawson, G. (2010). Neurons and oligodendrocytes recycle sphingosine 1-phosphate to ceramide. J. Biol. Chem. 285, 14134–14143. doi: 10.1074/jbc.M109.076810
Qiu, W., Fu, Z., Xu, G. G., Grassucci, R. A., Zhang, Y., Frank, J., et al. (2018). Structure and activity of lipid bilayer within a membrane-protein transporter. Proc. Natl. Acad. Sci. USA 115, 12985–12990. doi: 10.1073/pnas.1812526115
Quadri, Z., Elsherbini, A., Crivelli, S. M., El-Amouri, S. S., Tripathi, P., Zhu, Z., et al. (2024). Ceramide-mediated orchestration of oxidative stress response through filopodia-derived small extracellular vesicles. J. Extracell. Vesicles 13:e12477. doi: 10.1002/jev2.12477
Quinville, B. M., Deschenes, N. M., Ryckman, A. E., and Walia, J. S. (2021). A comprehensive review: sphingolipid metabolism and implications of disruption in sphingolipid homeostasis. Int. J. Mol. Sci. 22:5793. doi: 10.3390/ijms22115793
Rajagopalan, V., Canals, D., Luberto, C., Snider, J., Voelkel-Johnson, C., Obeid, L. M., et al. (2015). Critical determinants of mitochondria-associated neutral sphingomyelinase (MA-nSMase) for mitochondrial localization. Biochim. Biophys. Acta 1850, 628–639. doi: 10.1016/j.bbagen.2014.11.019
Real-Hohn, A., Navegantes, C., Ramos, K., Ramos-Filho, D., Cahuê, F., Galina, A., et al. (2018). The synergism of high-intensity intermittent exercise and every-other-day intermittent fasting regimen on energy metabolism adaptations includes hexokinase activity and mitochondrial efficiency. PLoS One 13:e0202784. doi: 10.1371/journal.pone.0202784
Reveglia, P., Paolillo, C., Angiolillo, A., Ferretti, G., Angelico, R., Sirabella, R., et al. (2023). A targeted mass spectrometry approach to identify peripheral changes in metabolic pathways of patients with Alzheimer’s disease. Int. J. Mol. Sci. 24:9736. doi: 10.3390/ijms24119736
Ryu, K. W., Fung, T. S., Baker, D. C., Saoi, M., Park, J., Febres-Aldana, C. A., et al. (2024). Cellular ATP demand creates metabolically distinct subpopulations of mitochondria. Nature 635, 746–754. doi: 10.1038/s41586-024-08146-w
Sambolín-Escobales, L., Tirado-Castro, L., Suarez, C., Pacheco-Cruz, D., Fonseca-Ferrer, W., Deme, P., et al. (2022). High-fat diet and short-term unpredictable stress increase long-chain ceramides without enhancing behavioral despair. Front. Mol. Biosci. 9:859760. doi: 10.3389/fmolb.2022.859760
Sapoń, K., Mańka, R., Janas, T., and Janas, T. (2023). The role of lipid rafts in vesicle formation. J. Cell Sci. 136:jcs260887. doi: 10.1242/jcs.260887
Scheibye-Knudsen, M., Fang, E. F., Croteau, D. L., Wilson, D. M., and Bohr, V. A. (2015). Protecting the mitochondrial powerhouse. Trends Cell Biol. 25, 158–170. doi: 10.1016/j.tcb.2014.11.002
Schoffelmeer, A. N., Vanderschuren, L. J., De Vries, T. J., Hogenboom, F., Wardeh, G., and Mulder, A. H. (2000). Synergistically interacting dopamine D1 and NMDA receptors mediate nonvesicular transporter-dependent GABA release from rat striatal medium spiny neurons. J. Neurosci. 20, 3496–3503. doi: 10.1523/JNEUROSCI.20-09-03496.2000
Scholtens, E. L., Krebs, J. D., Corley, B. T., and Hall, R. M. (2020). Intermittent fasting 5:2 diet: what is the macronutrient and micronutrient intake and composition? Clinic. Nutrit. 39, 3354–3360. doi: 10.1016/j.clnu.2020.02.022
Schumacher, F., Edwards, M. J., Mühle, C., Carpinteiro, A., Wilson, G. C., Wilker, B., et al. (2022). Ceramide levels in blood plasma correlate with major depressive disorder severity and its neutralization abrogates depressive behavior in mice. J. Biol. Chem. 298:102185. doi: 10.1016/j.jbc.2022.102185
Sekine, S., and Youle, R. J. (2018). PINK1 import regulation; a fine system to convey mitochondrial stress to the cytosol. BMC Biol. 16:2. doi: 10.1186/s12915-017-0470-7
Sentelle, R. D., Senkal, C. E., Jiang, W., Ponnusamy, S., Gencer, S., Selvam, S. P., et al. (2012). Ceramide targets autophagosomes to mitochondria and induces lethal mitophagy. Nat. Chem. Biol. 8, 831–838. doi: 10.1038/nchembio.1059
Serra, D., Mera, P., Malandrino, M. I., Mir, J. F., and Herrero, L. (2013). Mitochondrial fatty acid oxidation in obesity. Antioxid. Redox Signal. 19, 269–284. doi: 10.1089/ars.2012.4875
Shichkova, P., Coggan, J. S., Markram, H., and Keller, D. (2024). Brain metabolism in health and neurodegeneration: the interplay among neurons and astrocytes. Cells 13:1714. doi: 10.3390/cells13201714
Sinenko, S. A., Kuzmin, A. A., Skvortsova, E. V., Ponomartsev, S. V., Efimova, E. V., Bader, M., et al. (2023). Tryptophan Hydroxylase-2-mediated serotonin biosynthesis suppresses cell reprogramming into pluripotent state. Int. J. Mol. Sci. 24:4862. doi: 10.3390/ijms24054862
Siskind, L. J., and Colombini, M. (2000). The lipids C2- and C16-ceramide form large stable channels. Implications for apoptosis. J. Biol. Chem. 275, 38640–38644. doi: 10.1074/jbc.C000587200
Siskind, L. J., Kolesnick, R. N., and Colombini, M. (2006). Ceramide forms channels in mitochondrial outer membranes at physiologically relevant concentrations. Mitochondrion 6, 118–125. doi: 10.1016/j.mito.2006.03.002
Sokolović, M., Wehkamp, D., Sokolović, A., Vermeulen, J., Gilhuijs-Pederson, L. A., van Haaften, R. I., et al. (2007). Fasting induces a biphasic adaptive metabolic response in murine small intestine. BMC Genomics 8:361. doi: 10.1186/1471-2164-8-361
Sorochynska, O. M., Bayliak, M. M., Gospodaryov, D. V., Vasylyk, Y. V., Kuzniak, O. V., Pankiv, T. M., et al. (2019). Every-other-day feeding decreases glycolytic and mitochondrial energy-producing potentials in the brain and liver of Young mice. Front. Physiol. 10:1432. doi: 10.3389/fphys.2019.01432
Steger, F. L., Donnelly, J. E., Hull, H. R., Li, X., Hu, J., and Sullivan, D. K. (2021). Intermittent and continuous energy restriction result in similar weight loss, weight loss maintenance, and body composition changes in a 6 month randomized pilot study. Clinic. Obes. 11:e12430. doi: 10.1111/cob.12430
Stekovic, S., Hofer, S. J., Tripolt, N., Aon, M. A., Royer, P., Pein, L., et al. (2019). Alternate day fasting improves physiological and molecular markers of aging in healthy, non-obese humans. Cell Metab. 30:e6, 462–476. doi: 10.1016/j.cmet.2019.07.016
Stiban, J., and Perera, M. (2015). Very long chain ceramides interfere with C16-ceramide-induced channel formation: a plausible mechanism for regulating the initiation of intrinsic apoptosis. Biochim. Biophys. Acta 1848, 561–567. doi: 10.1016/j.bbamem.2014.11.018
Sutton, E. F., Beyl, R., Early, K. S., Cefalu, W. T., Ravussin, E., and Peterson, C. M. (2018). Early time-restricted feeding improves insulin sensitivity, blood pressure, and oxidative stress even without weight loss in men with prediabetes. Cell Metab. 27, 1212–1221.e3. doi: 10.1016/j.cmet.2018.04.010
Tavakoli, A., Akhgarjand, C., Ansar, H., Houjaghani, H., Khormani, A., Djafarian, K., et al. (2025). The effects of intermittent fasting on antioxidant and inflammatory markers and liver enzymes in postmenopausal, overweight and obese women with rheumatoid arthritis: a randomized controlled trial. Sci. Rep. 15:2357. doi: 10.1038/s41598-025-86734-0
Templeman, I., Smith, H. A., Chowdhury, E., Chen, Y.-C., Carroll, H., Johnson-Bonson, D., et al. (2021). A randomized controlled trial to isolate the effects of fasting and energy restriction on weight loss and metabolic health in lean adults. Sci. Transl. Med. 13:eabd8034. doi: 10.1126/scitranslmed.abd8034
Teo, J. D., Marian, O. C., Spiteri, A. G., Nicholson, M., Song, H., Khor, J. X. Y., et al. (2023). Early microglial response, myelin deterioration and lethality in mice deficient for very long chain ceramide synthesis in oligodendrocytes. Glia 71, 1120–1141. doi: 10.1002/glia.24329
van Eijk, R. P. A., Jones, A. R., Sproviero, W., Shatunov, A., Shaw, P. J., Leigh, P. N., et al. (2017). Meta-analysis of pharmacogenetic interactions in amyotrophic lateral sclerosis clinical trials. Neurology 89, 1915–1922. doi: 10.1212/WNL.0000000000004606
Ventura, A. E., Varela, A. R. P., Dingjan, T., Santos, T. C. B., Fedorov, A., Futerman, A. H., et al. (2020). Lipid domain formation and membrane shaping by C24-ceramide. Biochim. Biophys. Acta Biomembr. 1862:183400. doi: 10.1016/j.bbamem.2020.183400
Vos, M., Dulovic-Mahlow, M., Mandik, F., Frese, L., Kanana, Y., Haissatou Diaw, S., et al. (2021). Ceramide accumulation induces mitophagy and impairs β-oxidation in PINK1 deficiency. Proc. Natl. Acad. Sci. USA 118:e2025347118. doi: 10.1073/pnas.2025347118
Vos, M., and Klein, C. (2022). Ceramide-induced mitophagy impairs ß-oxidation-linked energy production in PINK1 deficiency. Autophagy 18, 703–704. doi: 10.1080/15548627.2022.2027193
Wheeler, D., Knapp, E., Bandaru, V. V. R., Wang, Y., Knorr, D., Poirier, C., et al. (2009). Tumor necrosis factor-alpha-induced neutral sphingomyelinase-2 modulates synaptic plasticity by controlling the membrane insertion of NMDA receptors. J. Neurochem. 109, 1237–1249. doi: 10.1111/j.1471-4159.2009.06038.x
Wilhelmi de Toledo, F., Grundler, F., Sirtori, C. R., and Ruscica, M. (2020). Unravelling the health effects of fasting: a long road from obesity treatment to healthy life span increase and improved cognition. Ann. Med. 52, 147–161. doi: 10.1080/07853890.2020.1770849
Wilkinson, M. J., Manoogian, E. N. C., Zadourian, A., Lo, H., Fakhouri, S., Shoghi, A., et al. (2020). Ten-hour time-restricted eating reduces weight, blood pressure, and Atherogenic lipids in patients with metabolic syndrome. Cell Metab. 31, 92–104.e5. doi: 10.1016/j.cmet.2019.11.004
Wilson, D. M., Cookson, M. R., Van Den Bosch, L., Zetterberg, H., Holtzman, D. M., and Dewachter, I. (2023). Hallmarks of neurodegenerative diseases. Cell 186, 693–714. doi: 10.1016/j.cell.2022.12.032
Xu, W., Zhang, D., Ma, Y., Gaspar, R. C., Kahn, M., Nasiri, A., et al. (2024). Ceramide synthesis inhibitors prevent lipid-induced insulin resistance through the DAG-PKCε-insulin receptorT1150 phosphorylation pathway. Cell Rep. 43:114746. doi: 10.1016/j.celrep.2024.114746
Yabu, T., Shimuzu, A., and Yamashita, M. (2009). A novel mitochondrial sphingomyelinase in zebrafish cells. J. Biol. Chem. 284, 20349–20363. doi: 10.1074/jbc.M109.004580
Zhao, Y., Chen, F., Chen, S., Liu, X., Cui, M., and Dong, Q. (2013). The Parkinson’s disease-associated gene PINK1 protects neurons from ischemic damage by decreasing mitochondrial translocation of the fission promoter Drp1. J. Neurochem. 127, 711–722. doi: 10.1111/jnc.12340
Zigdon, H., Kogot-Levin, A., Park, J.-W., Goldschmidt, R., Kelly, S., Merrill, A. H., et al. (2013). Ablation of ceramide synthase 2 causes chronic oxidative stress due to disruption of the mitochondrial respiratory chain. J. Biol. Chem. 288, 4947–4956. doi: 10.1074/jbc.M112.402719
Keywords: neurodegeneration, microglia, intermittent fasting, ceramides, mitophagy
Citation: Valenzuela-Ahumada LA, Mercado-Gómez OF, Viveros-Contreras R, Guevara-Guzmán R and Camacho-Morales A (2025) Fasting the mitochondria to prevent neurodegeneration: the role of ceramides. Front. Neurosci. 19:1602149. doi: 10.3389/fnins.2025.1602149
Edited by:
Xiao-Yuan Mao, Central South University, ChinaReviewed by:
Cai Qi, Yale University, United StatesFiorella Colasuonno, University of Rome Tor Vergata, Italy
Copyright © 2025 Valenzuela-Ahumada, Mercado-Gómez, Viveros-Contreras, Guevara-Guzmán and Camacho-Morales. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alberto Camacho-Morales, YWNtNTkwQGhvdG1haWwuY29t; YWxiZXJ0by5jYW1hY2hvbXJAdWFubC5lZHUubXg=; Rosalinda Guevara-Guzmán, cmd1ZXZhcmFAdW5hbS5teA==