 Vinicius Tavares Kütter1*†
Vinicius Tavares Kütter1*† Ingrid Tavares Costa2†
Ingrid Tavares Costa2† Alina Criane de Oliveira Pires1†
Alina Criane de Oliveira Pires1† Maria Montes-Bayón3,4†Alfredo Sanz-Medel3,4†
Maria Montes-Bayón3,4†Alfredo Sanz-Medel3,4† Wilson Machado5†
Wilson Machado5† Emmanoel Vieira Silva Filho5†
Emmanoel Vieira Silva Filho5†- 1Geology and Geochemistry Program, Federal University of Pará, Belém, Pará, Brazil
- 2Faculty of Oceanography, Federal University of Pará, Belém, Pará, Brazil
- 3Department of Physical and Analytical Chemistry, Faculty of Chemistry, University of Oviedo, Oviedo, Spain
- 4Instituto de Investigación Sanitaria Del Principado de Asturias (ISPA), Hospital Universitario, Oviedo, Spain
- 5Geochemistry Program, University Federal Fluminense, Niterói, Rio de Janeiro, Brazil
Introduction: Mangrove forests play a crucial role in the biogeochemical cycling of trace elements in intertropical coastal regions. This study evaluates concentrations of metals and arsenic in mangrove plants and ferns (leaves, flowers, branches, and roots) along a tidal creek chronically affected by residues from a zinc smelter and along another creek influenced by iron ore storage in Sepetiba Bay, Rio de Janeiro, Brazil.
Methods: To investigate potential physiological responses to metal exposure, selected plant samples were analyzed for the occurrence of proteins like phytochelatins. Total element contents (As, Zn, Cd, Cr, Cu, Mn, Fe, Ni, and Pb) were determined using ICP-MS, and the proteins were analyzed through SEC-HPLC-ICP-MS.
Results: Elevated concentrations of As (average of 34.81 mg kg−1) were detected in fern leaves, and proteins similar to phytochelatins indicated the presence of two distinct chemical species of As in these plants. In mangrove trees, three chemical species were identified in leaves and two in flowers and branches. Statistical comparisons showed significant differences in Cr, Cu, and Pb in mangrove leaves and Pb in branches from the Arapucaia Channel relative to Saco do Engenho Creek. These higher concentrations were associated with untreated domestic effluent from the Mazomba River and effluent from ore and coal activities in Itaguaí Port.
Discussion: Results indicate a dilution gradient for all metals (except Fe) from the former Cia Mercantil Ingá waste site toward the bay, whereas the Arapucaia Channel does not show a comparable dilution pattern. These outcomes highlight consideration contaminant sources, hydrodynamics, and plant uptake pathways in managing mangroves in higher polluted area.
1 Introduction
Mangrove ecosystems are a special realm due to the efficient trapping effect they have on elements transported in the intertropical coastal zones mitigate pollutant transfer through estuaries (Sultana et al., 2023; Lacerda et al., 2022; Lacerda, 2025). Mangrove contamination by chemical pollutants has a cascading effect on nutrient cycling throughout the ecosystem. Studies indicate that mangrove leaves contaminated by metals and hydrocarbons have a greater production of phenolic compounds and tannins, a defense response mechanism against stressors (Yan et al., 2017; Jiang et al., 2017; Numbere and Camilo, 2016). Consequently, the cycling and isotopic fractionation of carbon and nitrogen is altered, because mangrove crabs and other herbivores find the leaves more difficult to digest, the subsequent processing of the carbon and nitrogen is impaired (Nordhaus and Wolff, 2007), leading to alterations in their biogeochemical cycling and isotopic fractionation (Almeida et al., 2022). Furthermore, the exudates released by the roots alter the rhizosphere sediment with an increase in pH and consequently inhibit functional bacteria-mediated Fe and S cycles and promote the immobilization of metals (e.g., Cd) in the sediments (Leng et al., 2022).
Mangrove plants have evolved biochemical defense mechanisms to regulate internal metal concentrations and maintain physiologically tolerable limits, minimizing possible harmful effects. This mechanism results in a change in the plant and sediment chemical composition when exposed to a high concentration of metals. One such mechanism involves the chelation of trace elements by phytochelatins (or proteins like phytochelatins), metal binding peptides synthesized to avoid toxicological effects on plant tissues (Faizan et al., 2024; Inouhe, 2005). Phytochelatins are biomarkers and constitute a family of metal/metalloid-complexing peptides consisting of repetitive γ-glutamylcysteine units with a carboxyl-terminal glycine and range from 5 to 17 amino acids in length (2–10 kDa) (Hall, 2002). They have been identified in a wide variety of plant species, as well as in some microorganisms (Bhat et al., 2025; Cobbett and Goldsbrough, 2002). Furthermore, phytochelatins play an important role in metal detoxification by transporting the metal to the vacuoles (Cobbett and Goldsbrough, 2002). Phytochelatins are biomarkers that have been successfully employed in biomonitoring, allowing the evaluation of metal exposure effects (Sanhita et al., 2024; Lara-Almazán et al., 2021; Gawel and Hemond, 2004). However, a limited number of studies have investigated the production of phytochelatins and/or phytochelatin genes expression in mangrove species, with a predominant focus on laboratory-based analyses in Indo-Pacific taxa (Nualla-Ong et al., 2020; Huang and Wang, 2010; Gonzalez-Mendoza et al., 2007), resulting in a significant paucity of data regarding mangrove species from the Americas.
In southeastern Brazil, due to the growth of large urban and industrial centers over the last few decades, mangroves have been massively suppressed with an area reduction of about 30% (Ferreira and Lacerda, 2016; Diniz et al., 2019). In addition, they have been exposed to high levels of domestic and industrial pollution (Machado et al., 2002; Luiz-Silva et al., 2008) and major large-scale technological accidents (tailing dam rupture, oil spill, sewage rupture) (Kütter et al., 2023; Queiroz et al., 2018; Farias et al., 2008). This region houses Sepetiba Bay, Rio de Janeiro, which is the eighth largest bay (447 km2) in Brazil, and which for 40 years received effluents from the zinc electroplating industry and suffered repeated occurrences of tailing dike rupture (1996, 2002, and 2003), spreading metal contamination throughout the bay causing chronic local contamination (Rodrigues et al., 2020; Tonhá et al., 2020).
The aim of the present study is to present new data on the concentration of metals (Fe, Mn, Cr, Cu, Zn, Ni, Cd, As, and Pb) and expression of proteins phytochelatins-like in the mangrove and mangrove-associated vegetation affected by disposals from the zinc electroplating industry (currently decommissioned) and iron ore and coal wastes (before the decommissioning process of the electroplating metallurgy) in Sepetiba Bay. These data will also serve as a starting point for future assessments of the efficiency of the metallurgical decommissioning process, as the samples were collected and analyzed 2 years before this process.
Chronic geochemical anomalies persist in sediments at Sepetiba Bay despite numerous remediation actions, including dredging and subaquatic confinement of contaminated sediments and complete closure and confinement of the 390,000 m3 zinc electroplating industry residues (Rodrigues et al., 2020). Baseline data are strategic to area management and evaluated the best remediation practices applied in this tropical environment.
Considering the worldwide ecological and economic relevance of coastal bays, which host a great aquatic biodiversity (e.g., coastal vegetation, fishes, invertebrates from rocky shores, muddy banks, and sandy beaches, as well as sea birds and mammals), the evaluation of organism contamination and a better comprehension of the physiological mechanisms adopted as defenses against adverse effects need to be addressed. The starting point of expression of proteins similar to phytochelatin in mangroves and ferns impacted by zinc electroplating industry residues can help to understand the toxicological impact on the biota of Sepetiba Bay.
2 Materials and methods
2.1 Study area
Sepetiba Bay is a highly impacted coastal lagoon system that has an area of 447 km2 during high tide and 419 km2 during low tide, and an average depth of 6 m. It has brackish water (salinity range: 22–32 psu) due to its connection with the Atlantic Ocean. The north shore of this bay is the most industrialized and urbanized area, hosts the largest mangrove area, and receives the main continental drainage, with the São Franscico River providing to 86% of freshwater input into the bay. Close to this area for four decades the zinc smelter effluents discharge in the mangrove adjacent to the defunct Companhia Mercantil and Industrial Ingá, resulting in the increased concentration of various metals (Zn, Cd, Pb, Cu) and arsenic (As) in the mangrove sediments adjacent to the industrial plant (Jeong et al., 2023; Fonseca et al., 2013; Magalhães et al., 2001). According to Gonçalves et al. (2020), metallurgy decommissioning in 2011 had a positive impact, reducing concentrations by 100-fold (Cd) and 70-fold (Zn) in bottom sediments of the bay, although the mangrove sediments directed affected by metallurgy waste are still hotspots of contamination. Metals enrichments in forms weakly bound to sediments have pointed relevant risks for the sediment biota of Sepetiba Bay (de Carvalho Vicente et al., 2024; da Silva et al., 2022; Rodrigues et al., 2017).
Companhia Mercantil and Industrial Ingá, situated at north portion of the bay in Madeira Island, operated for 40 years and disposed steel waste and effluents into the mangroves and the bay. This company became the largest producer of Zn and Cd, representing around 30% of the Brazilian market until 1998. The company discharged 24 tons year−1 of Cd and 3,660 tons year−1 of Zn in Sepetiba Bay until its closure after 40 years of activity (Barcellos and Lacerda, 1994; Molisani et al., 2004). This waste discharge resulted in a nearly 200-fold increase in the deposition of Zn and Cd in the coastal environment (Barcellos and Lacerda, 1994; Barcellos et al., 1997). Additionally, the presence of As contamination in bay sediments is related to the arsenic trioxide (As2O3) used for coal purification (Magalhães et al., 2001). Repeated tailing dike rupture disasters occurred (1996, 2002, and 2003), directly affecting the nearby mangroves. Recent studies indicate changes in the epicuticular wax of Avicennia shaueriana and Laguncularia racemosa leaves in areas with higher metal contamination in sediments in Sepetiba Bay (Victório et al., 2020, 2023). A morphology of the epicuticular wax has been applied as a biomarker of air pollution (Banerjee et al., 2022; Gostin, 2016).
The discharge of effluent into the bay by Companhia Mercantil and Industrial Ingá ceased in 2008 with the start of the zinc smelter decommissioning project, which was concluded in 2015. During this remediation project, sediment from some areas with higher metal levels was removed by dredging, and the material was disposed of in an underwater confined disposal facility. The waste ore pile in the dike was encapsulated and confined in the underground. However, the contaminated mangrove sediments were not remediated. Accord to Gonçalves et al. (2020), the decommissioning project resulted in reductions of bay sediment of Cd and Zn concentrations by 100% and 70%, respectively.
2.2 Sampling
Plant samples were collected in February 2009, before the completion of the decommissioning project (Figure 1). During this period, the mangrove received discharge of treated effluents from the Zn waste pile.

Figure 1. Sampling site at Sepetiba Bay, close to Itaguaí Port. The bay receives drainage from large cities (Itaguaí, Mangaratiba, Japeri, Miguel Pereira, and partially Rio de Janeiro) and an expanding industrial park (Santa Cruz), in addition to housing four large ports (operation mainly containers, ore, and steel).
At the edge of the tailing waste ore pile, the fern Nephrolepis biserrata was sampled and the mangrove Rhizophora mangle was sampled in six points at Saco do Engenho creek (receive tailing waste, buried by repeated tailing dike rupture). Four other sampling points for Rhizophora mangle were also included along the Arapucaia Channel, which receives the coal and ore effluents from Itaguaí Port and drainage from the Mazomba River. Points Arapucaia 2 and Arapucaia 4 are situated in front of the discharge tubes of coal ore piles area drainage. Fern roots, stems, and leaves were collected, while mangrove flowers, green leaves, and branches were sampled. Four neighbor plants with the same size were collected at each sampling point, and a composite sample (10 units of leaves, 5–10 branches, four roots) of different trees or ferns was used for each type of plant tissue. The mangrove leaves were sampled manually in the branches disposed around 2 m of the soil. Only adult Rhizophora mangle was selected.
2.3 Metal analysis
In the laboratory, the plants were washed with distilled water and their structures were separated. They were subsequently dried in an oven at 60 °C, homogenized and pulverized in agate grade, and aliquots of 250 mg (n = 3) of the sample were digested in aqua regia [6.0 mL of HCl (37% Merck) and 2.0 mL of HNO3 (65% Merck)] in a microwave (Anton PAAR MW3000). The extracts obtained were used to determine the total content of elements (As, Zn, Cd, Cr, Cu, Mn, Fe, Ni, and Pb) by inductively coupled plasma mass spectrometry (ICP-MS) (Agilent 7500). The detection limits were 0.5 mg kg−1 (Fe), 0.005 mg kg−1 (Mn, Ni), 0.001 mg kg−1 (Cr, Cu), 0.01 mg kg−1 (Zn), and 0.002 mg kg−1 (As). Analytical quality control was verified through blank samples reagents, triplicate, internal standard (Rh), and the use of certified reference material (BCR 414 plankton; NIST SEM 1515 apple leaf) analyses. The recovery rates for the certified reference material were within the acceptable range at 92% to 100%.
2.4 Analysis of proteins similar to phytochelatin
The determination of biomolecules associated with arsenic was carried out following the methodology of Montes-Bayón et al. (2004). The biomolecules were extracted from 200 mg of plant samples in 50 mM ammonium acetate (pH ~5) (99% Sigma-Aldrich) over 1 h in ultrasound and subsequent filtration through a 0.45μm membrane. The determination of the molecule was carried out by injecting 50 μL of sample into the size-exclusion chromatography—high performance liquid chromatography (LC-10 AD Shimadzu)—ICP-MS (Agilent 7500) (SEC-HPLC-ICP-MS) system with a Superdex peptide column at a flow rate of 0.6 mL/min.
2.5 Statistical analyses
In Statistica® 13, the Shapiro–Wilk test was performed to assess the normality of the data. The results indicated that the data were non-normally distributed. Therefore, a non-parametric Mann–Whitney test could have been used to compare samples from Saco do Engenho inlet and Arapucaia Channel. The Kruskal–Wallis test was applied to investigate differences between metal concentrations in the plant's tissues.
3 Results and discussion
3.1 Total metal concentration
Arsenic, Zn, Cd, Cr, Cu, Fe, Ni, and Pb exhibited higher concentrations in the fern species than in the mangrove species (Figures 2, 3). Supplementary Table 1 presents the mean concentrations (in parts per million, mg Kg−1) of nine metals across various plant tissues, including leaves, stems, roots, and flowers. The data highlight variations in metal accumulation depending on plant type and tissue, providing insights into potential bioaccumulation patterns and environmental exposure.

Figure 2. Metal concentrations in the fern Nephrolepis biserrata close to the waste ore pile.

Figure 3. Metals distribution in mangrove impacted by waste ore in Sepetiba Bay.
The fern showed the highest concentrations of metals (Cr, Cu, Fe, Ni, Zn, Cd, and Pb) in the roots, while Mn and As presented the highest concentrations in the leaves. The higher metal concentrations in the fern roots are associated to the higher capacity of phytostabilization of the Nephrolepis sp. (Ancheta et al., 2020).
Manganese participates in the biochemical and physiological functions of the plants, such as photosynthesis and serving as a cofactor in the production of superoxide dismutase (Graham et al., 1988). The higher concentrations of Mn in the leaf would then be associated with the detoxification processes of free oxygen radicals produced when the plant is under environmental stress, with high concentrations of metals in the soil. The present Sepetiba Bay sediments, enriched in Mn due to influence of ore dispersion from the port terminal, increase the concentration in the sand beaches (Gueiros et al., 2003; Arruda et al., 2025).
Figure 3 shows a marked increase in As concentration (34.81 mg kg−1) detected in leaves of the fern sample, while the values found in the stems and roots were, respectively, 4.46 mg kg−1 and 3.20 mg kg−1, 10 times lower than detected in leaves. A previous study conducted by Melendez et al. (2011) at Companhia Mercantil and Industrial Ingá area, reported that As concentration in N. biserrata leaves range from 1.2 to 26.4 mg kg−1 in the Saco do Engenho inlet and 5.4–36.7 mg kg−1 close to the metallurgical waste pile.
On other hand, in Cu-Au mine tailing from Philippines in the Nueva Vizcaya area highest metals concentration in N. biserrata:191.41 mg kg−1 Cu and 153.96 mg kg−1 (leaves), 5,756.07 mg kg−1 Cu, < 0.01 mg kg−1 As (roots) (Ancheta et al., 2020). In the same way Manan et al. (2015) demonstrate higher activities of catalase and ascorbate peroxidase and production of phenolics and total flavonoids in N. biserrata from contaminated soil in Pasir Gudang Industrial Park, Malaysia compared to control (Manan et al., 2015). Manan et al. (2015) reported concentration of metals in leaves were: Zn 4,837 mg kg−1 (contaminated area) and 760 mg kg−1 (control area); Pb 133.7 mg kg−1 (contaminated area) and 11.3 mg kg−1 (control area); Cu 82 mg kg−1 (contaminated area) and 15.6 mg kg−1 (control area).
The highest Zn levels in fern in our study were associated with pollution from Companhia Mercantil and Industrial Ingá. The fern showed Zn concentrations of 287.91 mg kg−1 in roots, 70.12 mg kg−1 in stems, and 86.12 mg kg−1 in leaves; these are high values when compared to other works (Supplementary Table 1). Unlike the concentrations obtained for Cd, which did not exceed 3 mg kg−1, being 2.66, 1.63, and 1.68 mg kg−1 for root, stem, and leaf, respectively. These lower concentrations of Zn and Cd in the aerial biomass of N. biserrata may indicate that environments saturated with trace elements may limit translocation and bioaccumulation mechanisms, not passing on nutrients to the rest of the plant, for example. Thus, these elements become toxic at high concentrations (>300 mg Zn/kg and >5 mg Cd/kg dry mass), resulting in serious physiological disorders, such as inhibition of photosynthesis, hormonal changes, and cell death (Marschner, 2012; Benavides et al., 2005).
Chromium (4.09 mg kg−1), Cu (18.23 mg kg−1), Fe (1,264.62 mg kg−1), and Ni (2.77 mg kg−1), however, are essential for the development of plants and, depending on their concentrations, they can have specific functions in physiology. The levels of these elements are higher in fern roots and no excess was observed in our data when compared to other studies (Table 1). The essential elements mentioned are within the tolerable limit for plants, participating as micronutrients with specific functions for these vegetables.
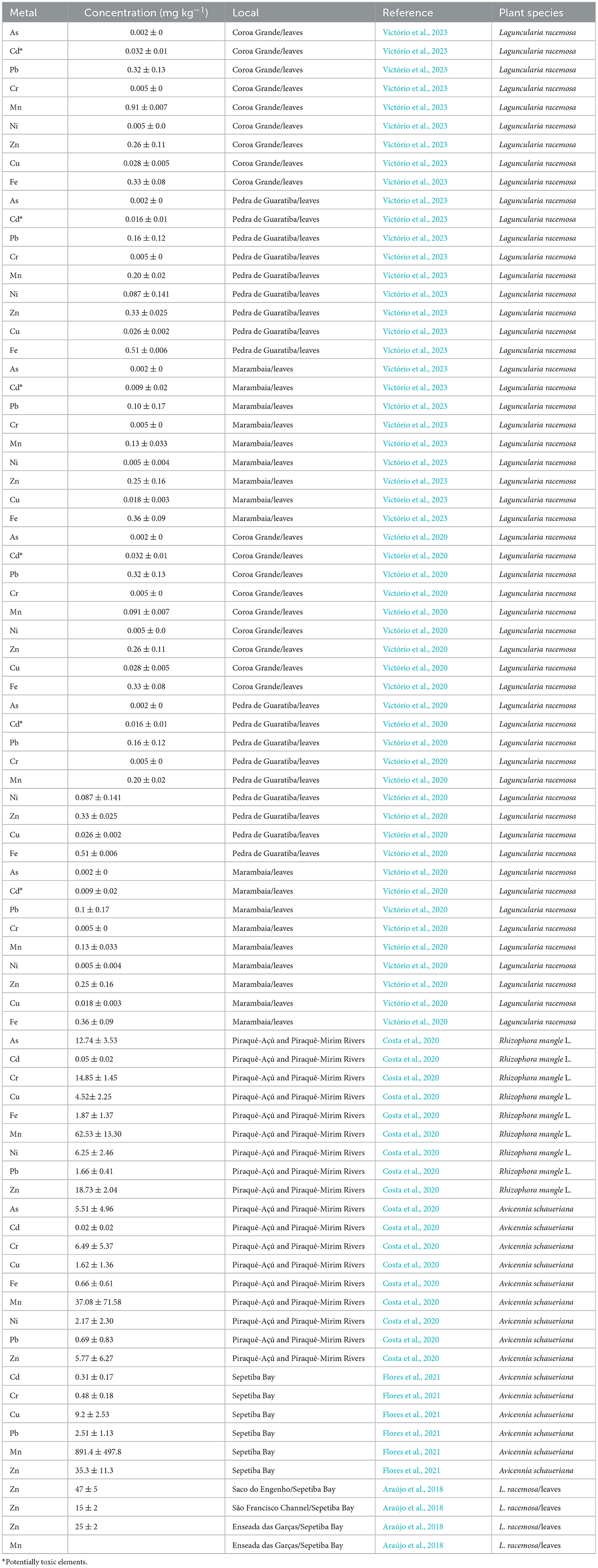
Table 1. Concentration of metal in leaves of mangrove species in different studies in Sepetiba Bay.
Rhizophora mangle did not present high levels of As, Cr, Cu, Ni, Fe, Zn, Cd, or Pb, compared to Nephrolepis biserrata (Figure 4). The mangrove from Arapucaia Channel and Saco do Engenho creek showed, using a Mann–Whitney U-test, significance at p < 0.05 for the metals Cr, Cu, and Pb present in the leaf and Pb in the branches between these areas. The higher values of Cr, Cu, and Pb in the leaf and Pb in the branches of mangroves from Arapucaia Channel can be attributed to domestic untreated effluent discharge from Mazomba River in the channel and the effluents of coal and ore operation in Itaguaí Port. Cunha et al. (2009) demonstrated the Pb input from the river's discharges to Sepetiba Bay.

Figure 4. SEC-UV-Vis-ICP-MS (Superdex peptide) in fern.
This difference in the distribution and concentration of metals between the two plant species analyzed is related to evolutionary and physiological differences between the species. While ferns are arsenic hyperaccumulators (Gonzaga et al., 2006), mangroves are tolerant to metals and have several metal homeostasis systems, such as the formation of iron plaques in the roots, limiting the translocation of toxic metals to the aerial parts (Yan et al., 2017). Laboratory experiments demonstrate that Avicennia marina is As-tolerant, and iron plaque reduces As translocation in the root, especially when there is a supply of and Fe2+ (Lin et al., 2018). In addition, Avicennia marina can excrete Zn through leaf glandular trichomes (MacFarlane and Burchett, 1999). Many mangrove species showed high resistance to metals (MacFarlane et al., 2007; Lu et al., 2014). Soil retention and root ultrafiltration would exclude most toxic metals, and only those absorbed and translocated to the aerial part would interfere with plant performance. The relatively higher tolerances to metals in the Rhizophoraceae mangrove species are attributed to their thick lignified/suberized exodermis, which could directly delay the entry of metals into the roots (Cheng et al., 2014). The concentrations of metals and metalloids showed distribution patterns in different compartments in Rhizophora mangle (Figure 4).
δ66ZnJMC in contaminated mangroves exhibits significant isotopic fractionation during its assimilation by vegetation. Studies on Laguncularia racemosa in Sepetiba Bay revealed that leaves present Zn isotopic signatures (δ66ZnJMC between +0.08‰ and +0.23‰) consistently lighter than those of the sediments where the trees are rooted (δ66ZnJMC: +0.36‰ to +0.84‰). This difference highlights critical biogeochemical processes: (1) selective bioavailability in the rhizosphere, where sulfides and organic complexes enrich the dissolved phase in light isotopes; (2) preferential translocation of 64Zn via xylem sap; and (3) tolerance mechanisms, such as active excretion through glandular leaf trichomes and the formation of root iron plaques, which immobilize heavy isotopes. The isotopic homogeneity in leaves (variation ≤ 0.15‰), even under sediment contamination gradients, suggests a uniformly accessible bioavailable pool to roots, reinforcing the role of mangroves as efficient biogeochemical filters. Such patterns allow the use of δ66Zn as a proxy for physiological stress in impacted coastal ecosystems (Araújo et al., 2018). The highest As concentrations in mangrove vegetation were reported only in flowers (Figure 4). This high concentration of As in the flowers of Saco do Engenho may be related to the higher concentration of this element in the local sediment (Magalhães et al., 2001) and also to the fact that these flowers have a different biochemical composition in comparison to another tissues, with sugars and anthocyanin to attract pollinating insects. Studies report the presence of anthocyanin in mangrove flowers (Paul et al., 2024), this pigment may increase arsenic tolerance (Ahammed and Yang, 2022). In relation to Zn, it can be observed that the levels showed in Arapucaia Channel and Saco do Engenho creek shoe similar values, probably due to the proximity of these points to Ingá Mercantil tailing waste, which it has a high concentration of this metal (Flores et al., 2021).
Cadmium did not present high concentrations (< 1 mg kg−1 in leaves), as well as Ni and Cr, whose content did not exceed 1.8 mg kg−1 in leaves and 1.6 mg kg−1 in branches, respectively. Manganese accumulated in mangrove branches and leaves, reaching 67.6 mg kg−1 in branches and 63 mg kg−1 in leaves, with similar concentrations in both areas. This accumulation pattern suggests a tendency for Mn to translocate to aerial tissues, which may be related to its high mobility in the xylem and its essential physiological role, especially as an enzymatic cofactor in photosynthesis and in the activation of cellular metabolic enzymes (Marschner, 2012). The higher concentration in branches and leaves may also reflect the demand for Mn by these tissues in photosynthetic and antioxidant processes, and may also indicate that, under the reducing conditions common in mangrove soils, some Mn remains immobilized in the roots in insoluble forms, while the available fraction is readily transported to the upper parts of the plant (Kabata-Pendias, 2011; Fernando and Lynch, 2015). These results point to an efficient physiological control of Mn homeostasis in Rhizophora mangle, favoring its accumulation in tissues with greater metabolic activity.
Lead, Cu, and Cr exhibited significant high levels in different compartments of Rhizophora mangle from Arapucaia Channel compared to Saco do Engenho creek. Jeong et al. (2023) identified Cu (167 mg kg−1) and Pb (257 mg kg−1) sediment hotspots in the Saco do Engenho inlet. Sepetiba Bay has multiple sources of Pb contamination (discharge of domestic and industrial effluents, gasoline, particulate matter transported by rivers, seawater, etc.) resulting in local biota contamination (Jeong et al., 2023; Kütter et al., 2021; Morales et al., 2019; Cunha et al., 2009).
Manganese accumulated in the branches and leaves of the mangrove, reaching 67.6 mg kg−1 in the branches and 63 mg kg−1 in the leaves with similar concentration in the two areas.
Among the elements studied here, Fe presented the highest concentration per sampling point, reaching 752.6 mg kg−1 leaves in S. Engenho 6. Iron is characterized as an essential micronutrient present mainly in the process of respiration and photosynthesis in plants. Likewise, Mn has a structural role and also acts in the photosynthetic process. Furthermore, these concentrations of Fe and Mn may be related to anthropogenic discharges from various industrial and port activities close to the inlet and channel around Sepetiba Bay. The high concentrations of Fe and Mn observed in the sediments may be related to local anthropogenic sources associated with industrial and logistical activities in Sepetiba Bay, particularly near Arapucaia and Saco do Engenho. Notable sources include steel and metallurgical complexes, such as Companhia Mercantil and Industrial Ingá and the private port terminal of TKCSA (ThyssenKrupp CSA), whose implementation from the 1960s to the 2000s has led to mangrove removals. Dredging and significant water alteration in iron ore logistics, with rail transport and port unloading in the Mangaratiba, Sepetiba region, resulted in the dispersion of dust rich in Fe, Mn, Cd, and other metals, near Sepetiba Bay (Arruda et al., 2025) and the Sepetiba Tecon/Porto de Itaguaí container port terminal, which handles large volumes of bulk cargo and operates on bottoms subject to dredging, contributing to the remobilization of metals in the sediments. The concentrations reported in this work indicate that the metals present in the Saco do Engenho inlet tend to follow a decreasing pattern, that is, with a higher content in the innermost area close to the courtyard of the former Cia Mercantil Ingá, diluting toward the mouth, except for Fe, which follows the opposite pattern.
In relation to the elements of the Arapucaia Channel, the influence of anthropogenic activities was observed, which tended to the strong assimilation of metals along the entire length of this channel. Silva et al. (1990) presented lower values for Zn and Fe in Rhizophora mangle, since the first Cia Ingá disaster occurred only in 1996, after their study.
The metal concentration values are below those reported by Kehrig et al. (2003), for mangrove sediment environments in southeastern Brazil, in addition to being below the mangrove sediment concentrations of some studies (Guzman and Jiménez, 1992; Che, 1999; Zan et al., 2002) in other parts of the world. This difference may reflect attenuation processes over time, such as the burial of contaminated layers by more recent, less impacted sediments. The adoption of environmental controls since the 2000s, including effluent treatment, CONAMA regulations, and increased oversight, may have contributed to the reduction in the direct input of metals to Sepetiba Bay.
Dilution by less contaminated terrigenous sediments from rivers such as the Guarda and Guandu may also be involved, especially in areas with greater hydrodynamics. From a geochemical perspective, processes such as the precipitation of Fe and Mn oxides, complexation with organic matter, and incorporation into less bioavailable phases (such as the residual or sulfide-associated fraction) favor the stabilization of metals in surface sediments.
Cheng et al. (2010) revealed that Zn significantly reduced the barriers to radial oxygen loss in three mangrove species (Aegiceras corniculatum, Bruguiera gymnorrhiza, and Rhizophora stylosa), thus inhibiting seedling growth due to changes in the anatomical structure of the root, such as thickening of the root, source, and cortex, and increased lignification in the exodermis and endodermis. Victório et al. (2020) also detected the presence of Al, Mn, Sr, and Zn in the leaves of Avicennia schaueriana, reducing the leaf area, directly affecting the photosynthetic process, and reducing epicuticular wax. Furthermore, leaf necrosis can be observed in some species, being induced by the hyperaccumulation of metals (Campos et al., 2018). These processes would end up occurring in the plants investigated in this study due to the higher concentrations of Zn and Cd in sediments and even in plant organisms.
Exposure of plants to high concentrations of As in the soil can have a deleterious effect. Campos et al. (2018), when demonstrating the hyperaccumulation of As in Pityrogramma calomelans, observed necrosis in the leaves, morphological changes in the root, and progressive darkening, which intensified with increasing concentration and exposure time to 30 mM As in the solution, reaching a total concentration of 6,289 mg kg−1 from root to leaves.
3.2 Binding of to proteins similar to phytochelatin
In fern species, biomolecule analyzes demonstrate the existence of two chemical species of As in the plant structure (Figure 4), while for mangroves, three chemical species were found in the leaves and two in flowers and branches, all in the low molecular weight range. In R. mangle, greater speciation diversity was observed, particularly in the leaves, where three distinct peaks were recorded (18, 26, and 31 min), while flowers and branches exhibited two main peaks (26 and 30 min) (Figure 5). The most intense and recurrent peak, located at 26 min, coincides with the sulfur absorption spectral region, which strongly suggests the involvement of compounds containing thiol (–SH) groups, such as GSH and PCs, in arsenic complexation. This association is consistent with studies indicating that phytochelatins are essential for the sequestration and intracellular transport of metals and metalloids in plants, functioning as chelating agents that reduce the toxicity of elements such as As(III) (Raab et al., 2004; Abbas et al., 2018).

Figure 5. SEC-UV-Vis-ICP-MS (Superdex peptide) in mangrove.
The greater species diversity in R. mangle, especially in the leaves, the most metabolically active tissues, may reflect greater complexity in the defense mechanisms and compartmentalization of As in this woody species (Mishra et al., 2021), typical of estuarine wetlands subject to various environmental stressors. These data indicate not only the presence of sulfur-complexed species but also possible differences in the metabolic pathway or the availability of intracellular ligands between species and tissues. Thus the results demonstrate that both N. biserrata and R. mangle have the ability to form As complexes with thiolated ligands, suggesting a conserved detoxification mechanism, although with different levels of speciative complexity (Navaza et al., 2006; Mishra et al., 2021).
4 Conclusion
Environments saturated with Zn and Cd may limit the translocation and bioaccumulation mechanisms of Nephrolepis biserrata. The high concentrations of these metals in the roots suggest an initial defense barrier against their toxicity. In contrast, arsenic is more mobile in the plant, being translocated to the aboveground biomass, classifying the species as a potential phytoextractor of this element. The persistent influence of industrial and port discharges continues to shape the distribution of metals in the region, especially near the Saco do Engenho mangrove swamp. The presence of high concentrations of Fe and Mn indicates ongoing impacts from port tailings and industrial emissions, with an increasing gradient of contamination toward the metallurgical tailings pile. In this context, mangroves act as natural filters, functioning as phytostabilizers of metallic pollutants. Therefore, it is essential to investigate the arsenic chemical species present in the mangrove forest to understand their ecotoxicological risks and inform remediation strategies. The results obtained contribute to future studies evaluating the effects of climate change on the dynamics of metals and the biogeochemical cycle of sediments impacted by industrial waste.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material; further inquiries can be directed to the corresponding author.
Author contributions
VK: Writing – review & editing, Formal analysis, Supervision. IC: Writing – original draft. AP: Writing – review & editing. MM-B: Methodology, Formal analysis, Writing – review & editing, Supervision. AS-M: Writing – review & editing, Supervision. WM: Writing – review & editing. ES: Writing – review & editing, Supervision.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by CAPES (001).
Acknowledgments
We thank Professor Anderson de Araújo Rocha for valuable support in the first analysis during pre-project experiment.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/focsu.2025.1602115/full#supplementary-material
References
Abbas, G., Murtaza, B., Bibi, I., Shahid, M., Niazi, N. K., Khan, M. I., et al. (2018). Arsenic uptake, toxicity, detoxification, and speciation in plants: physiological, biochemical, and molecular aspects. Int. J. Environ. Res. Public Health 15:59. doi: 10.3390/ijerph15010059
Ahammed, G. J., and Yang, Y. (2022). Anthocyanin-mediated arsenic tolerance in plants. Environ. Pollut. 292:118475. doi: 10.1016/j.envpol.2021.118475
Almeida, E. V. D., Kütter, V. T., and Silva-Filho, E. V. (2022). Trophic analysis of female mangrove crabs at two sites from Southeastern Brazil (Rio de Janeiro). Ocean Coast. Res. 70:e22002. doi: 10.1590/2675-2824070.21032evda
Ancheta, M. H., Quimado, M. O., Tiburan, C. L. Jr., Doronila, A., and Fernando, E. S. (2020). Copper and arsenic accumulation of Pityrogramma calomelanos, Nephrolepis biserrata, and Cynodon dactylon in Cu- and As-mine tailings. J. Degrad. Mining Land Management, 7, 2201–2208. doi: 10.15243/jdmlm.2020.073.2201
Araújo, D. F., Machado, W., Weiss, D., Mulholland, D. S., Garnier, J., Souto-Oliveira, C. E., et al. (2018). Zinc isotopes as tracers of anthropogenic sources and biogeochemical processes in contaminated mangroves. Appl. Geochem. 95, 25–32. doi: 10.1016/j.apgeochem.2018.05.008
Arruda, S. R., Martins, M. V. A., Reis, G. A., Terroso, D. L., Telles, R. C. M., Lima, D. S. L., et al. (2025). Impact caused by the transportation of iron ore on the beaches of a tourist region in SE Brazil: Mangaratiba (Rio de Janeiro). J. Sediment. Environ. 1:227. doi: 10.1007/s43217-025-00227-8
Banerjee, S., Banerjee, A., and Palit, D. (2022). Morphological and biochemical study of plant species—a quick tool for assessing the impact of air pollution. J. Clean. Prod. 339:130647 doi: 10.1016/j.jclepro.2022.130647
Barcellos, C., and Lacerda, L. D. (1994). Cadmium and zinc source assessment in the Sepetiba Bay and basin region. Environ. Monit. Assess. 29, 183–199. doi: 10.1007/BF00546874
Barcellos, C., Rezende, C. E., and Pfeiffer, W. C. (1997). Zn and Cd production and pollution in a Brazilian coastal region. Mar. Pollut. Bull. 22, 558–561. doi: 10.1016/0025-326X(91)90896-Z
Benavides, M. P., Gallego, S. M., and Tomaro, M. L. (2005). Cadmium toxicity in plants. Braz. J. Plant Physiol. 17, 21–34. doi: 10.1590/S1677-042020050001000
Bhat, B. A., Rather, M. A. R. A., Pirzadah, T. B., Nazir, R., Mir, R. A., Qadir, R. U., et al. (2025). Plant hyperaccumulators: a state-of-the-art review on mechanism of heavy metal transport and sequestration. Front. Plant Sci. Sec. Plant Abiotic Stress 16:1631378. doi: 10.3389/fpls.2025.1631378
Campos, N. V., Arcanjo-Silva, S., Freitas-Silva, L., Araújo, T. O., Souza-Fernandes, D. P., Azevedo, A. A., et al. (2018). Arsenic hyperaccumulation in Pityrogramma calomelanos L. (Link): Adaptive traits to deal with high metalloid concentrations. Environ. Sci. Pollut. Res. 25, 10720–10729. doi: 10.1007/s11356-017-1085-9
Che, R. G. O. (1999). Concentration of 7 heavy metals in sediments and mangrove root samples from Mai Po, Hong Kong. Mar. Pollut. Bull. 39, 269–279. doi: 10.1016/S0025-326X(99)00056-9
Cheng, H., Jiang, Z. Y., Liu, Y., Ye, Z. H., Wu, M. L., Sun, C. C., et al. (2014). Metal (Pb, Zn and Cu) uptake and tolerance by mangroves in relation to root anatomy and lignification/suberization. Tree Physiol. 34, 646–656. doi: 10.1093/treephys/tpu042
Cheng, H., Liu, Y., Tam, N. F. Y., Wang, X., Li, S. Y., Chen, G. Z., et al. (2010). The role of radial oxygen loss and root anatomy on zinc uptake and tolerance in mangrove seedlings. Environ. Pollut. 158, 1189–1196. doi: 10.1016/j.envpol.2010.01.025
Cobbett, C., and Goldsbrough, P. (2002). Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annu. Rev. Plant Biol. 53, 159–182. doi: 10.1146/annurev.arplant.53.100301.135154
Costa, E. S., Sá, F., Gomes, L. E. O., Silva, C. A., Lima, A. T., Lehrback, B. D., et al. (2020). Can severe drought periods increase metal concentrations in mangrove sediments? A case study in eastern Brazil. Sci. Tot. Environ. 748:142443. doi: 10.1016/j.scitotenv.2020.142443
Cunha, B. C. A., Rocha, D., Geraldes, M. C., Pereira, S. D., and Almeida, A. C. (2009). Pb isotopic signatures in sediments of a sub-tropical coastal lagoon: anthropogenic sources for metal contamination in the Sepetiba Bay (SE—Brazil). J. Coastal Research, 797–801. Available online at: http://www.jstor.org/stable/25737688
da Silva, L. C., Martins, M. V. A., Castelo, W. F. L., Saibro, M. B., Rangel, D., Pereira, E., et al. (2022). Trace metals enrichment and potential ecological risk in sediments of the Sepetiba Bay (Rio de Janeiro, SE Brazil). Mar. Pollut. Bull. 177:113485. doi: 10.1016/j.marpolbul.2022.113485
de Carvalho Vicente, M., Trevisan, C. L., de Carvalho, A. C. B., de Oliveira, B. C. V., de Rezende, C. E., Machado, W. V., et al. (2024). Geochemical fractionation of trace metals and ecological risk assessment of surface sediments in Sepetiba Bay, Brazil. Environ. Sci. Pollut. Res., 31, 14254–14269. doi: 10.1007/s11356-024-32095-6
Diniz, C., Cortinhas, L., Nerino, G., Rodrigues, J., Sadeck, L., Adami, M., et al. (2019). Brazilian mangrove status: Three decades of satellite data analysis. Remote Sensing, 11, 808. doi: 10.3390/rs11070808
Faizan, M., Alam, P., Hussain, A., Karabulut, F., Tonny, S. H., Cheng, S. H., et al. (2024). Phytochelatins: key regulator against heavy metal toxicity in plants. Plant Stress 11:100355. doi: 10.1016/j.stress.2024.100355
Farias, C. O., Hamacher, C., Wagener, A. de L. R., and Scofield, A. de L. (2008). Origin and degradation of hydrocarbons in mangrove sediments (Rio de Janeiro, Brazil) contaminated by an oil spill. Org. Geochem. 39, 289–307. doi: 10.1016/j.orggeochem.2007.12.008
Fernando, D. R., and Lynch, J. P. (2015). Manganese phytotoxicity: new light on an old problem. Ann. Bot. 116, 313–319. doi: 10.1093/aob/mcv111
Ferreira, A. C., and Lacerda, L. D. (2016). Degradation and conservation of Brazilian mangroves: status and perspectives. Ocean Coast. Manage. 125, 38–46. doi: 10.1016/j.ocecoaman.2016.03.011
Flores, V. R., Berbert, L. C., Succar, J. B., Victório, C. P., Direito, I. C. N., Cardoso, A. M., et al. (2021). Avicennia schaueriana as a highly efficient accumulator for Manganese in Sepetiba Bay, Brazil. Int. J. Environ. Sci. Technol. 10, 364–366. doi: 10.1007/s13762-021-03482-0
Fonseca, E. F., Baptista Neto, J. B., and Silva, C. G. (2013). Heavy metal accumulation in mangrove sediments surrounding a large waste reservoir of a local metallurgical plant, Sepetiba Bay, SE, Brazil. Environ. Earth Sci. 70, 643–650. doi: 10.1007/s12665-012-2148-3
Gawel, J. E., and Hemond, H. F. (2004). Biomonitoring for metal contamination near two Superfund sites in Woburn, Massachusetts, using phytochelatins. Environ. Pollut. 131, 125–135. doi: 10.1016/j.envpol.2004.01.012
Gonçalves, R. A., Oliveira, D. F., Rezende, C. E., Almeida, P., Lacerda, L. D., Gama, B. A., et al. (2020). Spatial and temporal effects of decommissioning a zinc smelter on the sediment quality of an estuary system: Sepetiba Bay, Rio de Janeiro, Brazil. J. Braz. Chem. Soc. 31, 683–693. doi: 10.21577/0103-5053.20190232
Gonzaga, M. I. S., Santos, J. A. G., and Ma, L. Q. (2006). Arsenic phytoextraction and hyperaccumulation by fern species. Sci. Agric. 63, 90–101. doi: 10.1590/S0103-90162006000100015
Gonzalez-Mendoza, D., Moreno, A. Q., and Zapata-Perez, O. (2007). Coordinated responses of phytochelatin synthase and metallothionein genes in black mangrove, Avicennia germinans, exposed to cadmium and copper. Aquat. Toxicol. 83, 306–314. doi: 10.1016/j.aquatox.2007.05.005
Gostin, I. (2016). “Air pollution stress and plant response,” in Plant Responses to Air Pollution, eds. U. Kulshrestha and P. Saxena (Singapore: Springer).
Graham, R. D., Hannam, R. J., and Uren, N. C. (Eds.) (1988). Manganese in Soils and Plants. Dordrecht: Kluwer Academic Publishers. doi: 10.1007/978-94-009-2817-6
Gueiros, B. B., Machado, W., Lisboa Filho, S. D., and Lacerda, L. D. D. (2003). Manganese behavior at the sediment-water interface in a mangrove dominated area in Sepetiba Bay, SE Brazil. J. Coast Res. 19, 550–559.
Guzman, H. M., and Jiménez, C. E. (1992). Contamination of coral reefs by heavy metals along the Caribbean coast of Central America (Costa Rica and Panama). Mar. Pollut. Bull. 24, 554–561. doi: 10.1016/0025-326X(92)90708-E
Hall, J. L. (2002). Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 53, 1–11. doi: 10.1093/jexbot/53.366.1
Huang, G. Y., and Wang, Y. S. (2010). Expression and characterization analysis of type 2 metallothionein from grey mangrove species (Avicennia marina) in response to metal stress. Aquat. Toxicol. 99, 86–92. doi: 10.1016/j.aquatox.2010.04.004
Inouhe, M. (2005). Fitoquelatinas. Brazil. J. Plant Physiol. 17, 65–78. doi: 10.1590/S1677-04202005000100006
Jeong, H., Araújo, D. F., Garnier, J., Mulholland, D., Machado, W., Cunha, B., et al. (2023). Copper and lead isotope records from an electroplating activity in sediments and biota from Sepetiba Bay (southeastern Brazil). Mar. Pollut. Bull. 190:114848. doi: 10.1016/j.marpolbul.2023.114848
Jiang, S., Weng, B., Liu, T., Su, Y., Liu, J., Lu, H., et al. (2017). Response of phenolic metabolism to cadmium and phenanthrene and its influence on pollutant translocations in the mangrove plant Aegiceras corniculatum (L.) Blanco (Ac). Ecotoxicol. Environ. Saf. 141, 290–297. doi: 10.1016/j.ecoenv.2017.03.041
Kabata-Pendias, A. (2011). Trace Elements in Soils and Plants (4th Edn.). Boca Raton, FL: CRC Press.
Kehrig, H. A., Pinto, F. N., Moreira, I., and Malm, O. (2003). Heavy metals and methylmercury in a tropical coastal estuary and a mangrove in Brazil. Org. Geochem. 34, 661–669. doi: 10.1016/S0146-6380(03)00021-4
Kütter, V. T., Martins, G. S., Brandini, N., Cordeiro, R. C., Almeida, J. P. A., Marques, E. D., et al. (2023). Impacts of a tailings dam failure on water quality in the Doce river: the largest environmental disaster in Brazil. J. Trace Elem. Miner. 5:100084. doi: 10.1016/j.jtemin.2023.100084
Kütter, V. T., Moreira, V. A., Kütter, M. T., Silva-Filho, E. V., Marques, E. D., Garnier, J., et al. (2021). Metal bioavailability and distribution in the fish community in a tropical estuary, Sepetiba Bay, Rio de Janeiro, Brazil. Geochimica Brasiliensis 35, 17–38. doi: 10.21715/GB2358-2812.2021351017
Lacerda, L. D. (2025). “Mangroves mitigate pollutant transfer through estuaries,” in Mangroves in the Anthropocene: Climate Change and Conservation Challenges. ISME Mangrove Ecosystems Technical Reports, Vol. 7, eds. L. D. Lacerda, H. E. Evans, S. Baba and H. T. Chan, p. 114.
Lacerda, L. D., Ward, R. D., Borges, R., and Ferreira, A. C. (2022). Mangrove trace metal biogeochemistry response to global climate change. Front. For. Glob. Change 5:817992. doi: 10.3389/ffgc.2022.817992
Lara-Almazán, N., Zarazúa-Ortega, G., Ávila-Pérez, P., Carreño-De León, C., and Barrera-Díaz, C. E. (2021). Multielemental analysis by total reflection X-ray fluorescence spectrometry and phytochelatins determination in aquatic plants. X-Ray Spectrom. 50, 414–424. doi: 10.1002/xrs.3220
Leng, Z., Wu, Y., Li, J., Nie, Z., Jia, H., Yan, C., et al. (2022). Phenolic root exudates enhance Avicennia marina tolerance to cadmium under the mediation of functional bacteria in mangrove sediments. Mar. Pollut. Bull. 185:114227. doi: 10.1016/j.marpolbul.2022.114227
Lin, Y., Fan, J., Yu, J., Jiang, S., Yan, C., Liu, J., et al. (2018). Root activities and arsenic translocation of Avicennia marina (Forsk.) Vierh seedlings influenced by sulfur and iron amendments. Mar. Pollut. Bull. 135, 1174–1182. doi: 10.1016/j.marpolbul.2018.08.040
Lu, H., Liu, B., Zhang, Y., Ye, J., and Yan, C. (2014). Comparing analysis of elements sub-cellular distribution in Kandelia obovata between SEM-EDX and chemical extraction. Aquat. Bot. 112, 10–15. doi: 10.1016/j.aquabot.2013.07.007
Luiz-Silva, W., Machado, W., and Matos, R. H. (2008). Multi-elemental contamination and historic record in sediments from the Santos-Cubatão Estuarine System, Brazil. J. Braz. Chem. Soc. 19, 1490–1500. doi: 10.1590/S0103-50532008000800008
MacFarlane, G. R., and Burchett, M. D. (1999). Zinc distribution and excretion in the leaves of the grey mangrove, Avicennia marina (Forsk.) Vierh. Environ. Exp. Bot. 41, 167–175. doi: 10.1016/S0098-8472(99)00002-7
MacFarlane, G. R., Koller, C. E., and Blomberg, S. P. (2007). Accumulation and partitioning of heavy metals in mangroves: a synthesis of field-based studies. Chemosphere 69, 1454–1464. doi: 10.1016/j.chemosphere.2007.04.059
Machado, W., Moscatelli, M., Rezende, L. G., and Lacerda, L. D. D. (2002). Mercury, zinc, and copper accumulation in mangrove sediments surrounding a large landfill in southeast Brazil. Environ. Pollut. 120, 455–461. doi: 10.1016/S0269-7491(02)00108-2
Magalhães, V. F., Carvalho, C. E. V., and Pfeiffer, W. C. (2001). Arsenic contamination and dispersion in the Engenho Inlet, Sepetiba Bay, SE, Brazil. Water Air Soil Pollut. 129, 83–90. doi: 10.1023/A:1010381902874
Manan, F. A., Mamat, D. D., Samad, A. A., Ong, Y. S., Ooh, K. F., Chai, T. T., et al. (2015). Heavy metal accumulation and antioxidant properties of Nephrolepis biserrata growing in heavy metal-contaminated soil. Global NEST J. 17, 544–554. doi: 10.30955/gnj.001463
Marschner, P. (2012). Marschner's Mineral Nutrition of Higher Plants (3rd Edn.). Cambridge, MA: Academic Press.
Melendez, L. B., Silva-Filho, E. V., Miekeley, N., Vieira, F. A., and Sella, S. M. (2011). Determination of arsenic species in P. calomelanos and N. biserrata. J. Braz. Chem. Soc. 22, 1961–1967. doi: 10.1590/S0103-50532011001000018
Mishra, R. K., Tiwari, S., Patel, A., and Prasad, S. H. (2021). Arsenic contamination, speciation, toxicity and defense strategies in plants. Braz. J. Bot 44, 1–10. doi: 10.1007/s40415-020-00694-5
Molisani, M. M., Marins, R. V., Machado, W., Paraquetti, H. H. M., Bidone, E. D., Lacerda, L. D., et al. (2004). Environmental changes in Sepetiba Bay, SE Brazil. Reg. Environ. Change 4, 17–27. doi: 10.1007/s10113-003-0060-9
Montes-Bayón, M., Meija, J., LeDuc, D. L., Terry, N., Caruso, J. A., and Sanz-Medel, A. (2004). HPLC-ICP-MS and ESI-Q-TOF analysis of biomolecules induced in Brassica juncea during arsenic accumulation. J. Anal. At. Spectrom. 19, 153–158. doi: 10.1039/B308986J
Morales, S. J. D., Guerra, J. V., Nunes, M. A. S., Martins, M. V. A., Souza, A. M., Geraldes, M. C., et al. (2019). Anthropogenic impacts on the Western sector of Sepetiba bay (Brazil) assessed by the Pb isotopic composition of surface sediments. J. Sediment. Environ. 4, 291–311. doi: 10.12957/jse.2019.45269
Navaza, A. P., Montes-Bayón, M., LeDuc, D. L., Terry, N., and Sanz-Medel, A. (2006). Study of phytochelatins and other related thiols as complexing biomolecules of As and Cd in wild type and genetically modified Brassica juncea plants. J. Mass Spectrom. 41, 323–331. doi: 10.1002/jms.992
Nordhaus, I., and Wolff, M. (2007). Feeding ecology of the mangrove crab Ucides cordatus (Ocypodidae): food choice, food quality, and assimilation efficiency. Mar. Biol. 151, 1665–1681. doi: 10.1007/s00227-006-0597-5
Nualla-Ong, A., Phongdara, A., and Buapet, P. (2020). Copper and zinc differentially affect root glutathione accumulation and phytochelatin synthase gene expression of Rhizophora mucronata seedlings: implications for mechanisms underlying trace metal tolerance. Ecotoxicol. Environ. Saf. 205:111175. doi: 10.1016/j.ecoenv.2020.111175
Numbere, A. O., and Camilo, G. R. (2016). Mangrove leaf litter decomposition under mangrove forest stands with different levels of pollution in the Niger River Delta, Nigeria. Afr. J. Ecol. 55, 162–167. doi: 10.1111/aje.12335
Paul, I., Manna, S., Bera, R., Paine, A. K., Mridha, D., Gorain, P. C., et al. (2024). Floral scents, specialized metabolites and stress-response activities in Heritiera fomes and Bruguiera gymnorrhiza from Sundarban mangrove ecosystem. J. Plant Res. 137, 463–484. doi: 10.1007/s10265-024-01527-2
Queiroz, H. M., Nóbrega, G. N., Ferreira, T. O., Almeida, L. S., Romero, T. B., Santaella, S. T., et al. (2018). The Samarco mine tailing disaster: a possible time-bomb for heavy metals contamination? Sci. Total Environ. 637, 498–506. doi: 10.1016/j.scitotenv.2018.04.370
Raab, A., Feldmann, J., and Meharg, A. A. (2004). The nature of arsenic-phytochelatin complexes in Holcus lanatus and Pteris cretica. Plant Physiol. 134, 1113–1122. doi: 10.1104/pp.103.033506
Rodrigues, S. K., Abessa, D. M. S., Rodrigues, A. P. C., Soares-Gomes, A., Freitas, C. B., Santelli, R. E., et al. (2017). Sediment quality in a metal-contaminated tropical bay assessed with a multiple lines of evidence approach. Environ. Pollut. 228, 265–276. doi: 10.1016/j.envpol.2017.05.045
Rodrigues, S. K., Machado, W., Guerra, J. V., Geraldes, M., Morales, S., Vinzón, S. B., et al. (2020). Changes in Cd and Zn distribution in sediments after closure of an electroplating industry, Sepetiba bay, Brazil. Mar. Pollut. Bull. 161:111758. doi: 10.1016/j.marpolbul.2020.111758
Sanhita, Sikri, N., Dutta, D., Behera, B., Dhaka, S., and Mehta, J. (2024). “Chapter 10—Plant biomarkers for environmental monitoring,” in Biomarkers in Environmental and Human Health Biomonitoring, eds. R. Mishra, S. Madhav, R. K. Dhaka and P. Garg (Cambridge, MA: Academic Press), 185–203.
Silva, C. A. R., Lacerda, L. D., and Rezende, C. E. (1990). Metals Reservoir in a Red Mangrove Forest. Biotropica 22:339. doi: 10.2307/2388551
Sultana, S., Huang, R., and Van Zwieten, L. (2023). Trapping effect of mangrove and saltmarsh habitats on geochemical elements: a case study in Ximen Island, Zhejiang, China. J. Soils Sediments 23, 2327–2343. doi: 10.1007/s11368-023-03503-0
Tonhá, M. S., Garnier, J., Araújo, D. F., Cunha, B. C., Machado, W., Dantas, E., et al. (2020). Behavior of metallurgical zinc contamination in coastal environments: a survey of Zn from electroplating wastes and partitioning in sediments. Sci. Total Env. 743:140610. doi: 10.1016/j.scitotenv.2020.140610
Victório, C. P., Dos Santos, M. S., Mello, M. C., Bento, J. P. S. P., da Costa Souza, M., Simas, N. K., et al. (2020). The presence of heavy metals in Avicennia schaueriana Stapf and Leechman ex Moldenke leaf and epicuticular wax from different mangroves around Sepetiba Bay, Rio de Janeiro, Brazil. Environ. Sci. Pollut. Res. 27, 23714–23729. doi: 10.1007/s11356-020-08606-6
Victório, C. P., Silva dos Santos, M., Cordeiro Dias, A., Bento, J. P. S. P., Ferreira, B. H. S., Souza, M. C., et al. (2023). Laguncularia racemosa leaves indicate the presence of potentially toxic elements in mangroves. Sci. Rep. 13:4845. doi: 10.1038/s41598-023-31986-x
Yan, Z., Sun, X., Xu, Y., Zhang, Q., and Li, X. (2017). Accumulation and tolerance of mangroves to heavy metals: a review. Curr. Pollut. Rep. 3, 302–317. doi: 10.1007/s40726-017-0066-4
Zan, Q. J., Wang, Y. J., and Wang, B. S. (2002). Accumulation and cycle of heavy metal in Sonneratia apetala and S. caseolaris mangrove community at Futian of Shenzhen, China. Chin. J. Environ. Sci. 23, 81–88. Available online at: https://europepmc.org/article/med/12371110
Keywords: Sepetiba Bay, metallurgical waste, trace metals, arsenic speciation, Nephrolepis biserrata, Rhizophora mangle
Citation: Kütter VT, Costa IT, Pires ACdO, Montes-Bayón M, Sanz-Medel A, Machado W and Silva Filho EV (2025) Mangroves and associated vegetation under metals and arsenic lingering impact: the legacies of zinc smelter industry residues and iron ore in southeast Brazil. Front. Ocean Sustain. 3:1602115. doi: 10.3389/focsu.2025.1602115
Received: 28 March 2025; Accepted: 26 September 2025;
Published: 03 December 2025.
Edited by:
Natalia Soares Quinete, Florida International University, United StatesReviewed by:
Leila S. Lemos, Florida International University, United StatesAjaya Bhattarai, Tribhuvan University, Nepal
Copyright © 2025 Kütter, Costa, Pires, Montes-Bayón, Sanz-Medel, Machado and Silva Filho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vinicius Tavares Kütter, a3V0dGVyQHVmcGEuYnI=
†ORCID: Vinicius Tavares Kütter orcid.org//0000-0001-7295-6800
Ingrid Tavares Costa orcid.org/0009-0000-1577-9609
Alina Criane de Oliveira Pires orcid.org/0000-0002-5132-0368
Maria Montes-Bayón orcid.org/0000-0001-6114-9405
Alfredo Sanz-Medel orcid.org/0000-0003-2899-1347
Wilson Machado orcid.org/0000-0003-3117-8584
Emmanoel Vieira Silva Filho orcid.org/0000-0001-6444-6851